Embed Size (px)
Citation preview

118
Limnol. Oceanogr., 48(1), 2003, 118–128q 2003, by the American Society of Limnology and Oceanography, Inc.
Abundance of amino sugars and peptidoglycan in marine particulate and dissolvedorganic matter
Ronald Benner1 and Karl KaiserDepartment of Biological Sciences and Marine Science Program, University of South Carolina, Columbia,South Carolina 29208
Abstract
Several recent studies indicated that amino sugars were likely to be major components of marine carbon andnitrogen cycles, but there has been insufficient data to investigate this hypothesis. In the present study, hydrolyzableamino sugars were measured in a variety of marine organisms and particulate organic matter (POM) and in ultraf-iltered dissolved organic matter (UDOM) from various ocean basins and depths. Glucosamine (GlcN) and galac-tosamine (GalN) were abundant and common to all samples. Concentrations of these two amino sugars were similarto their neutral sugar counterparts, glucose and galactose, and in surface water they accounted for up to ;1.5% ofC and N in POM and 2.5% of C and 7.1% of N in UDOM. Chitin, a polymer of glucosamine, is produced bymany marine organisms, but based on the low GlcN : GalN ratios (1.0–2.5) in POM and UDOM, it does not appearto be a primary source of glucosamine in these samples. Muramic acid, an amino sugar found only in the bacterialcell wall polymer peptidoglycan, was relatively abundant in POM, indicating that bacterial detritus is a principalcomponent of submicron particles in seawater. Muramic acid was measured in all UDOM samples, but its concen-trations were low (,1 nmol L21), indicating that intact fragments of peptidoglycan were a relatively minor com-ponent of UDOM. The abundance of GlcN and GalN and their similar concentrations in POM and UDOM providenovel information suggesting a major prokaryotic source.
Amino sugars are often considered to be abundant com-ponents of marine organic matter, yet there are surprisinglyfew measurements of amino sugars in the marine environ-ment. Chitin, a structural polymer of the amino sugar N-acetyl-glucosamine, is produced by a large variety of marineorganisms (Muzzarelli 1977), and the production of chitin inthe oceans is undoubtedly quite large. Kirchman and White(1999) estimated that about 10% of marine bacterial pro-duction could be supported by chitin. In addition to chitinand its derivatives, amino sugars are found in other poly-saccharides, glycoproteins, and glycolipids that are commonto many organisms (Sharon 1965). Amino sugars are im-portant structural components of prokaryotic cell walls,where they occur in peptidoglycan, lipopolysaccharides, andpseudopeptidoglycan (Brock et al. 1994). Several studiesprovide measurements of amino sugars in marine particlesand sediments (Ittekot et al. 1984; Muller et al. 1986; Lie-bezeit 1993; Dauwe and Middelburg 1998), but existing in-
1 Corresponding author (benner @biol.sc.edu).
AcknowledgmentsWe dedicate this paper to the memory of John I. Hedges. John
provided valuable comments on an earlier draft of this manuscriptand many hours of stimulating discussion on the chemical compo-sition of DOM. We thank Rumi Fukuda for providing samples ofnatural populations of marine bacteria collected from seawater, W.Whitman for providing cultures of methanogens, and Jose Bersanofor assistance with the collection and sorting of copepods. We thankscientists and crew aboard the USS Hawkbill and RV Weatherbirdand Vickers for assistance with sampling, David Kirchman, the bio-geochemistry group at the University of South Carolina, and twoanonymous reviewers for comments on an earlier draft of this man-uscript. This research was funded by National Science Foundationgrants OCE-0080782 and OPP-9996438.
formation is insufficient to provide more than a glimpse ofthe significance of amino sugars in the ocean.
It is now well recognized that carbohydrates are amongthe most abundant and reactive components of the marine Ccycle (Romankevich 1984; Benner et al. 1992; Pakulski andBenner 1994). The development of sensitive methods for themeasurement of neutral sugars in seawater has greatly ad-vanced understanding of the abundance, distribution, and cy-cling of these common carbohydrates in the ocean (Mopperet al. 1995; Borch and Kirchman 1997; Skoog and Benner1997). Similar methods were recently developed for themeasurement of amino sugars in seawater (Kaiser and Ben-ner 2000), and this report provides a first look at the bio-geochemistry of amino sugars in particulate and dissolvedorganic matter. Amino sugars are unusual carbohydrates inthat they contain nitrogen. Most N in marine dissolved or-ganic matter (DOM) resides in amide functional groups,which are common to combined amino acids and acetylatedamino sugars (McCarthy et al. 1997). Hydrolyzable aminoacids account for ,15% of dissolved organic nitrogen inseawater, suggesting that amino sugars could be importantin the marine N cycle (McCarthy et al. 1997).
Chemical characterization of a variety of biomolecules inmarine organic matter indicates that bacteria are an impor-tant source of marine DOM (Tanoue et al. 1995; Boon et al.1998; McCarthy et al. 1998). Most of the characterized bio-molecules are commonly or uniquely found in bacterial cellwalls. Peptidoglycan is the dominant cell wall polymerfound in bacteria, and existing data suggest this bacterialpolymer could be a major component of refractory DOM inthe ocean (Boon et al. 1998; McCarthy et al. 1998). Aminosugars form the glycan ‘‘backbone’’ of peptidoglycan, andthe characterization of specific amino sugars in marine par-ticulate and dissolved organic matter provides new insights

119Amino sugars in marine organic matter
Table 1. Recoveries of amino sugars from purified polymers.
Polymer Source Amino sugar Recovery (%)
ChitinChondroitin sulfate APeptidoglycan
(from M. luteus)
Sigma C-8908Sigma C-8529Wako 162-18101
GlucosamineGalactosamineGlucosamineMuramic acid
78787463
about the occurrence and refractory nature of peptidoglycanin marine organic matter.
Materials and methods
Sampling—Seawater samples were collected from the Pa-cific (February–March 1992, RV John Vickers), Atlantic(May 1992, RV Weatherbird), and Arctic Oceans (August–September 1998, USS Hawkbill ) using a high-volume pump(Pacific and Atlantic surface samples) or Niskin bottles.Samples collected in the Pacific and Atlantic were processedaboard ship immediately following collection. Samples col-lected from the Arctic were preserved with mercuric chloride(0.15 mmol L21) and processed in the home laboratory with-in 3 months of collection. Pacific samples were passedthrough a Nitex sieve (60 mm pore size) and processed bytangential-flow filtration to recover two size classes of ma-rine organic matter: particulate organic matter (POM; 0.1–60 mm pore size) and ultrafiltered dissolved organic matter(UDOM; 1–100 nm). Atlantic and Arctic samples werepassed through Nuclepore cartridge filters (0.2 mm poresize), and the UDOM size fraction (1–200 nm) was recov-ered using tangential-flow ultrafiltration. Concentrates fromtangential-flow filtration were diafiltered with Milli-Q waterto remove sea salts, and the desalted concentrates were driedunder vacuum. Sample handling and ultrafiltration proce-dures followed the methods of Benner et al. (1997).
Cultures—Cultures and natural populations of a wide di-versity of organisms were analyzed to determine primarysources and yields of amino sugars. Axenic cultures of ma-rine algae (Heterocapsa niei CCMP 447, Nannochloris sp.CCMP 518, Emiliania huxleyi CCMP 373, Thalassiosiraoceanica CCMP 1005) and the marine bacterium Synecho-coccus sp. CCMP 1334 were obtained from the culture col-lection at Bigelow. The marine bacterium Synechococcus ba-cillaris CCMP 1333 also was obtained from Bigelow andcultured as described in Biddanda and Benner (1997).Freeze-dried cultures of two soil bacteria, Bacillus subtilisATCC 6633 and Pseudomonas fluorescens ATCC 13430,were obtained from Sigma Chemical. A wild population ofTrichodesmium sp. was collected in August 1995 from sur-face waters of the Gulf of Mexico (Biddanda 1995). Naturalpopulations of marine bacteria (prokaryotes) were collectedfrom coastal and oceanic waters by size-selective filtrationmethods (Fukuda et al. 1998). Natural populations of thecopepod Acartia tonsa were collected from coastal waters ofthe Gulf of Mexico and sorted (male and female) using adissecting microscope. Two cultures of methanogenic Ar-chaea, Methanobrevibacter smithii and Methanococcus mar-
ipaludis, were obtained from W. Whitman at the Universityof Georgia.
Measurements—Concentrations of dissolved organic car-bon (DOC) were determined using a high-temperature com-bustion method and a Shimadzu TOC 5000 analyzer (Bennerand Strom 1993). The organic carbon and nitrogen contentof dried POM and UDOM samples were measured after va-por phase acidification using a Carlo Erba 1108 CHN ana-lyzer.
Amino sugars (glucosamine, galactosamine, mannosami-ne, muramic acid) were separated and measured using high-performance anion exchange chromatography (HPAEC) withpulsed amperometric detection (PAD). Sample preparationand chromatography conditions followed the procedures ofKaiser and Benner (2000) and are briefly presented here. ADionex 500 ion chromatography system with a CarboPacPA1 column and PAD detector were used for all analyses.Dried POM and UDOM samples were hydrolyzed with 3 MHCL at 1008C for 5 h. Hydrolyzed samples were neutralizedand split into two aliquots. One aliquot was used for analysisof glucosamine, galactosamine, and mannosamine, and theother aliquot was used for analysis of muramic acid. Afterdesalting using a cation exchanger, glucosamine, galactos-amine, and mannosamine were separated using isocratic con-ditions and a mobile phase with 12 mmol L21 NaOH and 1mmol L21 barium acetate. Muramic acid was measured in aseparate chromatographic analysis using isocratic conditionsand a mobile phase with 100 mmol L21 NaOH and 100mmol L21 sodium acetate.
During strong acid hydrolysis, acetyl groups are removedfrom amino sugars. Acetylated forms of amino sugars arecommon in nature (e.g., chitin, peptidoglycan), but we donot know whether the amino sugars measured in this studywere acetylated prior to acid hydrolysis. The amino sugarsmeasured in this study have six carbon atoms each, and thisis the value we used for calculations of amino sugar carbonin samples. Therefore, the values presented herein underes-timate amino sugar carbon by as much as 25% because theacetylated forms of these amino sugars have eight carbonatoms each.
Results
The recoveries of total hydrolyzable amino sugars fromseveral common polymers ranged from 63 to 78% (Table 1).Recoveries of glucosamine and galactosamine from poly-mers linked by glycosidic bonds were uniformly high (74–78%). The recovery of muramic acid from peptidoglycanwas somewhat lower (63%) and likely reflects the occur-

120 Benner and Kaiser
Table 2. Yields of amino sugars in pure cultures and natural assemblages of organisms collected from various marine environments.
Culture SourceClass orfamily
C/Natom
Amino sugar (nmol [mg C]21)
Ga1N ManN GlcN MA
Phototrophic algaeHeterocapsa nieiNannochloris sp.Emiliania huxleyiThalassiosira oceanica
CCMP 447CCMP 518CCMP 373CCMP 1005
DinophyceaeChlorophyceaePrymnesiophyceaeCoscinodiscophyceae
7.67.08.67.4
13.5bd5.13.1
3.04.70.23.1
5.914.6
9.163.7
bdbdbdbd
Phototrophic bacteriaSynechococcus bacillarisSynechococcus sp.Trichodesmium sp.
CCMP 1333CCMP 1334GOM wild
CyanophyceaeCyanophyceaeCyanophyceae
7.44.86.0
14.14.33.1
7.324.9
3.3
113.0243.2
25.0
15.133.911.0
Heterotrophic bacteriaBacillus subtilisPseudomonas fluorescensNatural populationNatural populationNatural populationNatural population
ATCC 6633ATCC 13430Southern OceanEquatorial PacificTokyo BayOtsuchi Bay
BacillaceaePseudomonadaceaeMixedMixedMixedMixed
4.54.16.15.87.75.2
30.530.910.016.935.810.2
4.7bd2.62.43.34.3
251.297.423.142.958.834.4
166.744.64.63.0nd7.7
CopepodAcartia tonsaAcartia tonsa
GOM femaleGOM male
CrustaceaCrustacea
4.54.4
32.358.0
bdbd
457.41,210.4
ndnd
Abbreviations: GalN, galactosamine; ManN, mannosamine; GlcN, glucosamine; MA, muramic acid; GOM, Gulf of Mexico; bd, below detection; nd, notdetermined.
rence of peptide as well as glycosidic bonds to this subunit.Overall, these recoveries indicate the hydrolysis is well suit-ed for the analysis of polymers that are likely to be abundantin seawater. No corrections for hydrolysis efficiency wereused in this study.
Analyses of total hydrolyzable amino sugars in a varietyof marine organisms indicated that galactosamine and glu-cosamine are common and abundant components of all cells(Table 2), including the cultures of methanogenic Archaea(data not shown). Mannosamine is also widely distributedbut is usually less abundant. The diatom, T. oceanica, pro-duces chitin (chitan—partially acetylated) fibrils, which ac-count for the relatively high yield of glucosamine in thismarine alga (McLachlan et al. 1965). Amino sugars account-ed for 0.1–0.5% and 0.2–0.6% of the carbon and nitrogen,respectively, in microalgae. The yields of galactosamine andglucosamine were generally several-fold higher in bacteriathan algae. Amino sugars accounted for 0.3–3.9% and 0.3–2.5% of the carbon and nitrogen, respectively, in phototroph-ic and heterotrophic bacteria. Glucosamine was very abun-dant in the copepod A. tonsa because of its chitinousexoskeleton. Amino sugar yields were lower in female co-pepods, which had threefold greater mass than males. Aminosugars accounted for ;12% and ;7% of male copepod car-bon and nitrogen, respectively.
Muramic acid was not present in any of the axenicallygrown algal cultures, but it was present in all phototrophicand heterotrophic bacterial cultures and samples (Table 2).Yields of muramic acid were much higher in the Gram-pos-itive bacterium B. subtilis than in the Gram-negative bacteria(Synechococcus, Trichodesmium, Pseudomonas). The cellwall of Gram-positive bacteria is much thicker and accountsfor a larger percentage of cellular carbon than in Gram-neg-
ative bacteria (Brock et al. 1994). Higher yields of muramicacid were found in cultured Gram-negative bacteria com-pared with natural populations collected from seawater. It isunclear whether this reflects variability in the contributionsof peptidoglycan to cellular carbon or the inclusion of Ar-chaea or detritus during the collection of natural populationsof bacteria from seawater. Muramic acid yields varied two-fold between the two cultured Synechococcus sp., indicatinga considerable variability in the peptidoglycan content ofclosely related marine bacteria. Muramic acid was not de-tected in the methanogen cultures (data not shown), whichis consistent with previous studies of the molecular com-position of cell walls in Archaea (Brock et al. 1994). Yieldsof muramic acid were typically less than half those of glu-cosamine in the same bacterium. These amino sugars occurin a 1 : 1 ratio in peptidoglycan, indicating the majority ofthe glucosamine in Gram-negative bacteria is not associatedwith this cell wall polymer.
Galactosamine and glucosamine were the most abundantamino sugars in POM collected from the Pacific Ocean (Ta-ble 3). Seawater concentrations of galactosamine and glu-cosamine in POM ranged from 0.07 to 1.09 nmol L21, withhighest concentrations in surface waters. The ratio of glu-cosamine : galactosamine (GlcN : GalN) ranged from 1.6 to2.5 and averaged 1.9. Mannosamine was detected in POMsamples, but it was in low concentration and could not bereliably quantified in the presence of high galactosamineconcentrations. Muramic acid was also present in low con-centration, but it was quantified in a separate chromatograph-ic analysis (see Methods). The carbon-normalized yields ofgalactosamine, glucosamine, and muramic acid in POM (Ta-ble 3) were similar to yields in natural populations of marinebacteria (Table 2). Glucosamine was much more abundant

121Amino sugars in marine organic matter
Table 3. Concentrations and yields of amino sugars in particulate organic matter (POM) from the Pacific Ocean.
Depth(m)
POC(mmol L21)
C/Natom
GalN(nmol L21)
GalN(nmol [mg C]21)
GlcN(nmol L21)
GlcN(nmol [mg C]21)
MA(nmol L21)
MA(nmol [mg C]21)
28S, 1408W2
100400
4,000
1.020.460.710.35
7.77.08.07.2
0.520.100.250.07
42.317.029.716.6
1.050.240.440.11
85.242.352.026.4
0.0710.0330.0270.012
5.815.923.212.88
128S, 1358W2
100200375
4,000
0.830.820.320.430.32
7.77.47.68.67.0
0.610.310.090.140.07
55.431.823.727.117.3
1.090.650.170.250.12
99.566.743.049.632.1
0.0650.0350.0170.0100.024
5.933.544.441.916.18
Table 4. Concentrations and yields of amino sugars in ultrafiltered dissolved organic matter (UDOM).
Depth(m)
DOC(mmol L21)
UDOC(mmol L21)
C/Natom
GalN(nmol L21)
GalN(nmol [mg C]21)
GlcN(nmol L21)
GlcN(nmol [mg C]21)
MA(nmol L21)
MA(nmol [mg C]21)
Pacific Ocean58359N, 1408009W
2 76 18.8 15.9 14.6 64.7 27.9 123.7 0.31 1.3728S, 1408W
2100400
4,000
72685144
21.714.411.7
8.0
16.116.616.718.4
23.111.7
4.82.9
88.767.734.230.1
37.217.5
7.14.4
142.9101.3
50.645.7
0.650.120.130.12
2.500.690.931.25
128S, 1358W2
100200375
4,000
8285575345
22.222.212.610.6
8.1
16.815.616.216.918.4
35.823.4
8.15.52.3
134.287.853.843.123.7
56.336.913.1
9.33.8
211.5138.4
86.373.039.0
0.780.440.430.260.10
2.931.652.842.041.03
Atlantic Ocean318509N, 648109W
1900
2,400
724746
16.48.39.2
17.115.618.1
18.34.43.8
92.844.534.3
30.18.35.6
152.883.350.5
0.270.090.10
1.370.910.91
Arctic Ocean848N, 1758W
1055
100132180
137104907776
32.724.918.416.416.5
21.019.318.416.116.4
14.310.5
7.16.97.9
36.535.132.135.240.0
19.618.2
8.06.7
10.6
49.861.136.234.153.8
0.440.220.070.100.10
1.130.750.330.510.53
226300600
1,0001,600
6562595854
13.414.412.813.013.5
14.215.215.216.515.9
6.86.25.25.84.9
42.136.033.636.930.0
10.09.67.15.84.8
62.455.846.136.929.4
0.110.110.130.130.27
0.700.640.830.831.67
(5–25-fold) than muramic acid, indicating most glucosaminewas not associated with peptidoglycan.
Total hydrolyzable amino sugars were analyzed in UDOMsamples from various locations and depths in three oceanbasins (Table 4). Surface water concentrations of galactos-amine and glucosamine in the UDOM size fraction rangedfrom 10 to 36 nmol L21 and 17 to 56 nmol L21, respectively.Deep water concentrations were less variable and ranged
from 2.3 to 5.8 nmol L21 and 3.8 to 5.8 nmol L21, respec-tively. Surface water concentrations of galactosamine andglucosamine were typically higher in the Pacific and Atlanticcompared with the Arctic. As with POM samples, manno-samine was detected in UDOM samples, but it was in lowconcentration and could not be reliably quantified in thepresence of high galactosamine concentrations. Muramicacid concentrations were low and ranged from 0.12 to 0.78

122 Benner and Kaiser
Fig. 1. Concentrations (nmol L21) of galactosamine (GalN),glucosamine (GlcN), and muramic acid in ultrafiltered dissolvedorganic matter (UDOM) from the Pacific, Atlantic, and ArcticOceans.
Fig. 2. Molar ratios of glucosamine : galactosamine (GlcN :GalN) and glucosamine : muramic acid (GlcN : MA) in UDOM fromthe Pacific, Atlantic, and Arctic Oceans. Vertical lines representmean values.
nmol L21 in surface waters and 0.10 to 0.27 nmol L21 indeep waters.
Composite depth profiles of total hydrolyzable galactos-amine, glucosamine, and muramic acid concentrations in theUDOM size fraction from the Pacific, Atlantic, and ArcticOceans are presented in Fig. 1. The profiles indicate similarconcentrations and depth trends among ocean basins. Sur-face water concentrations were 5–10-fold higher than deepwater concentrations, indicating a major surface water sourcefor these amino sugars. Glucosamine and galactosaminewere found in similar relative proportions in all UDOM sam-ples, and a surprisingly narrow range of GlcN : GalN values(1–2) was observed (Fig. 2). The average GlcN : GalN valuein UDOM was 1.5 and is similar to the average value (1.9)in POM. The ratio of glucosamine : muramic acid (GlcN :MA) in UDOM exhibited a wide range of values (18–146)with an average value of 70 (Fig. 2). These high valuesindicate most of the glucosamine in UDOM is not associatedwith the bacterial cell wall polymer peptidoglycan.
Carbon-normalized yields of galactosamine, glucosamine,and muramic acid in POM and UDOM from the same watersamples are compared at two stations (128S, 1358W and 28S,1408W) in the Pacific Ocean (Figs. 3, 4). Galactosamine andglucosamine yields are consistently lower in POM comparedto UDOM. Yields of galactosamine and glucosamine inPOM and UDOM decrease with increasing depth, althoughthe change is greater in UDOM. Unlike galactosamine andglucosamine yields, muramic acid yields were higher inPOM than UDOM. Muramic acid yields in POM andUDOM were variable and did not exhibit a consistent depth-related trend. Based on these overall patterns and distribu-tions, it appears that galactosamine and glucosamine areclosely associated in marine organic matter.
Total hydrolyzable amino sugars accounted for 0.7–2.5%of the carbon and 2.2–7.1% of the nitrogen in UDOM fromsurface waters (Fig. 5). About 0.6 and 1.5% of the carbonand nitrogen in UDOM from deep water was accounted foras amino sugars. Similar depth-related trends were observed
with POM. Amino sugars accounted for 0.34–1.30% of or-ganic carbon and 0.38–1.61% of organic nitrogen in POM.
The average carbon-normalized yields of galactosamine,glucosamine, and muramic acid in marine phytoplankton (al-gae), bacterioplankton, POM, and UDOM from surface anddeep waters are presented in Fig. 6. The yields of galactos-amine and glucosamine in surface water POM are greaterthan those in marine microorganisms. This could indicate anenrichment of these amino sugars during the decompositionof particulate material, or it could indicate other sources ofamino sugar–rich particles, such as zooplankton. An evengreater enrichment of amino sugars is observed in surfacewater UDOM. Again, it is not clear whether this trend isindicative of variable sources or diagenesis. The galactos-amine and glucosamine yields in deep water POM andUDOM are considerably lower than surface water counter-parts, and we interpret this as a diagenetic trend.
Muramic acid yields follow a very different pattern fromthat of galactosamine and glucosamine (Fig. 6). Bacteria arethe only known source of this amino sugar, and yields ofmuramic acid are highest in marine bacteria. The POM sam-ples include marine bacteria as well as other organisms andnonliving particles, and these samples have the next highestyields of muramic acid followed by UDOM. Muramic acidyields in POM and UDOM decrease with depth, as was ob-served with the other amino sugars.
Discussion
Abundance and distribution of amino sugars—Glucos-amine and galactosamine, the most common amino sugars,were measured in all marine phytoplankton, bacterioplank-ton, and zooplankton analyzed. Glucosamine was particular-ly abundant in marine copepods, accounting for as much as12% of biomass carbon. These two amino sugars were mea-sured in all samples of marine particulate and dissolved or-ganic matter, demonstrating their widespread distribution inthe ocean. Glucosamine and galactosamine occurred in fairlyconstant proportion (GlcN : GalN; 1.0–2.5) in all POM and
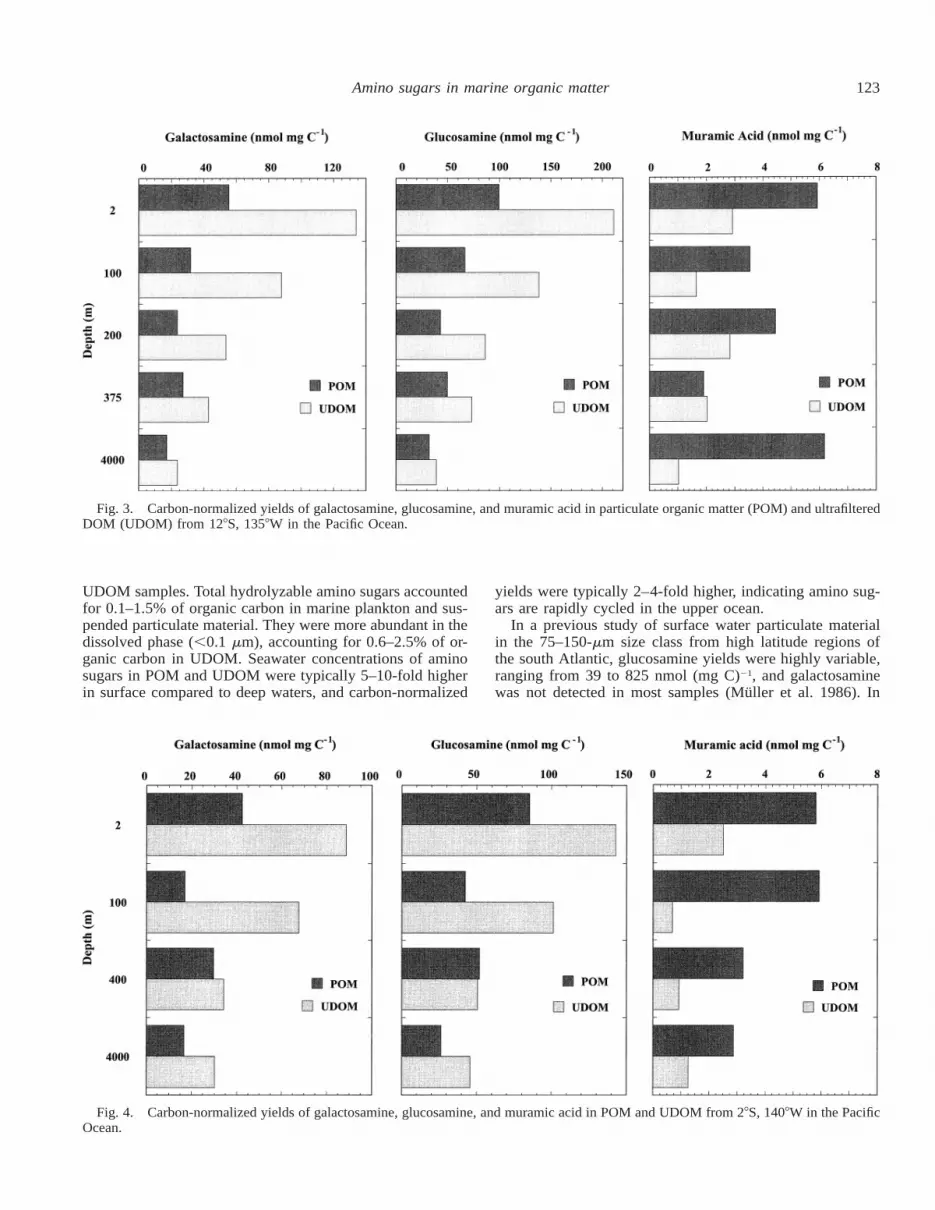
123Amino sugars in marine organic matter
Fig. 3. Carbon-normalized yields of galactosamine, glucosamine, and muramic acid in particulate organic matter (POM) and ultrafilteredDOM (UDOM) from 128S, 1358W in the Pacific Ocean.
Fig. 4. Carbon-normalized yields of galactosamine, glucosamine, and muramic acid in POM and UDOM from 28S, 1408W in the PacificOcean.
UDOM samples. Total hydrolyzable amino sugars accountedfor 0.1–1.5% of organic carbon in marine plankton and sus-pended particulate material. They were more abundant in thedissolved phase (,0.1 mm), accounting for 0.6–2.5% of or-ganic carbon in UDOM. Seawater concentrations of aminosugars in POM and UDOM were typically 5–10-fold higherin surface compared to deep waters, and carbon-normalized
yields were typically 2–4-fold higher, indicating amino sug-ars are rapidly cycled in the upper ocean.
In a previous study of surface water particulate materialin the 75–150-mm size class from high latitude regions ofthe south Atlantic, glucosamine yields were highly variable,ranging from 39 to 825 nmol (mg C)21, and galactosaminewas not detected in most samples (Muller et al. 1986). In
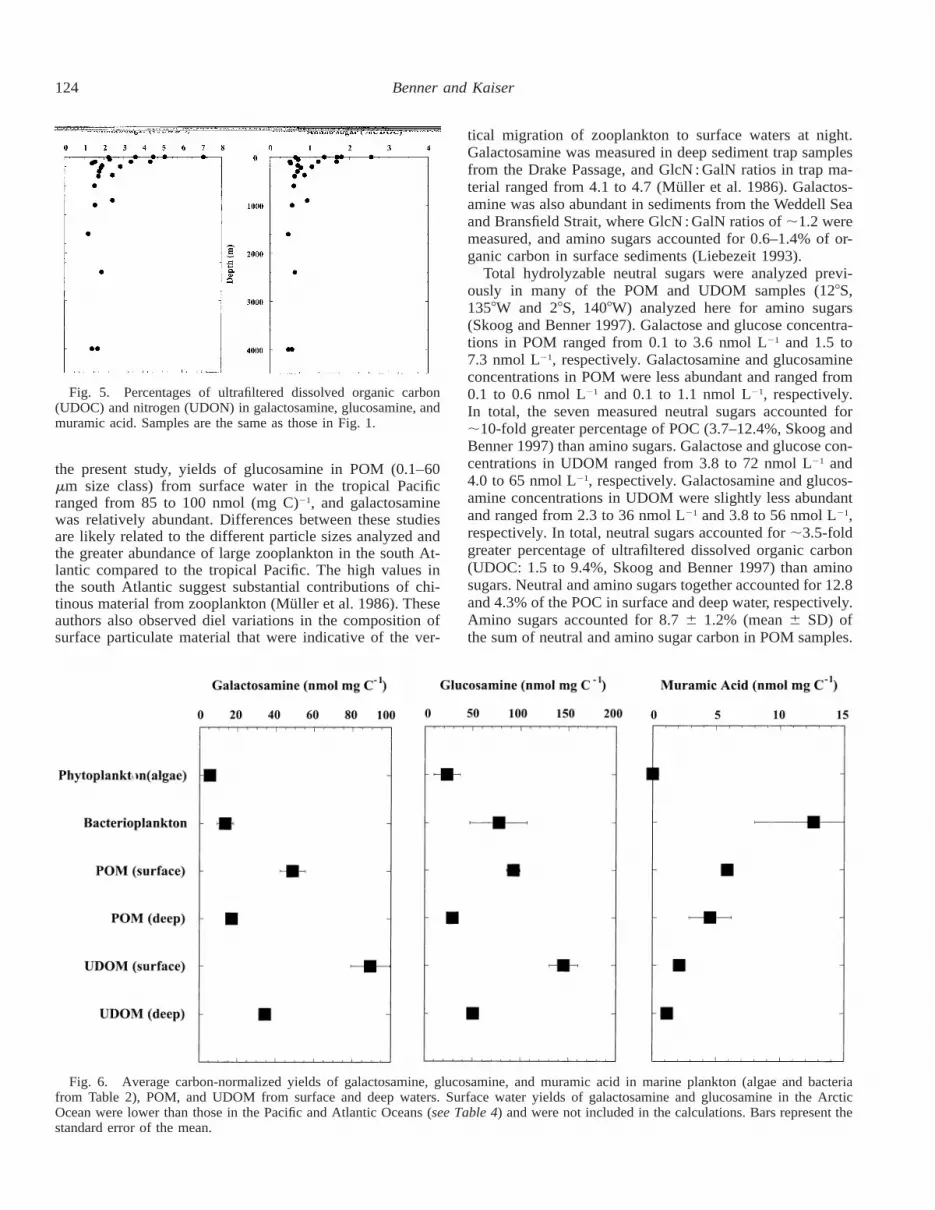
124 Benner and Kaiser
Fig. 5. Percentages of ultrafiltered dissolved organic carbon(UDOC) and nitrogen (UDON) in galactosamine, glucosamine, andmuramic acid. Samples are the same as those in Fig. 1.
Fig. 6. Average carbon-normalized yields of galactosamine, glucosamine, and muramic acid in marine plankton (algae and bacteriafrom Table 2), POM, and UDOM from surface and deep waters. Surface water yields of galactosamine and glucosamine in the ArcticOcean were lower than those in the Pacific and Atlantic Oceans (see Table 4) and were not included in the calculations. Bars represent thestandard error of the mean.
the present study, yields of glucosamine in POM (0.1–60mm size class) from surface water in the tropical Pacificranged from 85 to 100 nmol (mg C)21, and galactosaminewas relatively abundant. Differences between these studiesare likely related to the different particle sizes analyzed andthe greater abundance of large zooplankton in the south At-lantic compared to the tropical Pacific. The high values inthe south Atlantic suggest substantial contributions of chi-tinous material from zooplankton (Muller et al. 1986). Theseauthors also observed diel variations in the composition ofsurface particulate material that were indicative of the ver-
tical migration of zooplankton to surface waters at night.Galactosamine was measured in deep sediment trap samplesfrom the Drake Passage, and GlcN : GalN ratios in trap ma-terial ranged from 4.1 to 4.7 (Muller et al. 1986). Galactos-amine was also abundant in sediments from the Weddell Seaand Bransfield Strait, where GlcN : GalN ratios of ;1.2 weremeasured, and amino sugars accounted for 0.6–1.4% of or-ganic carbon in surface sediments (Liebezeit 1993).
Total hydrolyzable neutral sugars were analyzed previ-ously in many of the POM and UDOM samples (128S,1358W and 28S, 1408W) analyzed here for amino sugars(Skoog and Benner 1997). Galactose and glucose concentra-tions in POM ranged from 0.1 to 3.6 nmol L21 and 1.5 to7.3 nmol L21, respectively. Galactosamine and glucosamineconcentrations in POM were less abundant and ranged from0.1 to 0.6 nmol L21 and 0.1 to 1.1 nmol L21, respectively.In total, the seven measured neutral sugars accounted for;10-fold greater percentage of POC (3.7–12.4%, Skoog andBenner 1997) than amino sugars. Galactose and glucose con-centrations in UDOM ranged from 3.8 to 72 nmol L21 and4.0 to 65 nmol L21, respectively. Galactosamine and glucos-amine concentrations in UDOM were slightly less abundantand ranged from 2.3 to 36 nmol L21 and 3.8 to 56 nmol L21,respectively. In total, neutral sugars accounted for ;3.5-foldgreater percentage of ultrafiltered dissolved organic carbon(UDOC: 1.5 to 9.4%, Skoog and Benner 1997) than aminosugars. Neutral and amino sugars together accounted for 12.8and 4.3% of the POC in surface and deep water, respectively.Amino sugars accounted for 8.7 6 1.2% (mean 6 SD) ofthe sum of neutral and amino sugar carbon in POM samples.

125Amino sugars in marine organic matter
Neutral and amino sugars together accounted for 9.7 and2.3% of the UDOC in surface and deep water, respectively.Amino sugars accounted for 22.9 6 4.6% of the sum ofneutral and amino sugar carbon in UDOM samples.
Amino sugars are unique among the carbohydrates as ni-trogen-bearing molecules. They account for a substantialpercentage (2–7%) of ultrafiltered dissolved organic nitrogen(UDON) and are therefore likely important components ofthe marine N cycle. For comparison, total hydrolyzable ami-no acids accounted for 14–28% of UDON in samples thatwere similarly collected from the Pacific Ocean (McCarthyet al. 1996). Amino acids clearly account for a much greaterpercentage of marine organic nitrogen than amino sugars,but dissolved amino sugar concentrations and carbon-nor-malized yields decrease more rapidly with depth than aminoacid concentrations and yields, suggesting amino sugars aremore efficiently cycled in the upper water column (Benner2002).
Analyses by nuclear magnetic resonance (NMR) spec-troscopy of similar UDOM samples from the Pacific Oceanindicate that carbohydrates account for ;50% and ;25% ofUDOC in surface and deep water, respectively (Benner et al.1992). Total hydrolyzable neutral and amino sugars togetheraccount for only ;20% and ;10% of the carbohydrate car-bon indicated by NMR in UDOM from surface and deepwater, respectively. Acidic sugars (e.g., uronic acids) are an-other important group of carbohydrates not characterized inthis study, but recent spectrophotometric analyses of uronicacids indicate they are no more abundant than neutral oramino sugars (Hung et al. 2001). It is now apparent thatmost of the carbohydrate-type structures in UDOM indicatedby NMR are not characterized at the molecular level usingexisting hydrolytic and chromatographic methods. The hy-drolytic conditions chosen for carbohydrate analyses need tobe sufficient to cleave hydrolysis-resistant bonds without al-tering the structures of the carbohydrates released. It is pos-sible that our current methods fail to meet these criteria withnatural organic matter, resulting in low carbohydrate recov-eries after acid hydrolysis. Another possibility is that car-bohydrates are modified by biological and photochemicalprocesses in seawater (Benner and Biddanda 1998; Ober-nosterer et al. 1999; Ogawa et al. 2001), resulting in struc-tures that retain biochemical functional groups (e.g., C-Oand O-C-O) characteristic of carbohydrates and detected byNMR but lose their molecular identity. It is also conceivablethat unusual carbohydrates not characterized using currentanalytical procedures are the dominant carbohydrates in sea-water. Unraveling this mystery is critical for gaining a betterunderstanding of the source and reactivity of a large fractionof marine DOM.
Reactivity of amino sugars—Much can be learned aboutthe reactivity of organic matter by comparing depth-relatedtrends in the concentrations of carbon and specific biomol-ecules (Benner 2002). Such comparisons led Liebezeit(1993) to suggest that chitinous particulate material is largelyproduced and consumed in the upper ocean. Comparison ofthe concentrations of POC and total hydrolyzable amino sug-ars in surface and oxygen-minimum (375–400 m) waters ofthe tropical Pacific indicates that decreases in concentrations
of amino sugars were ;2-fold greater than decreases inPOC. Based on the same comparison for UDOM, decreasesin concentrations of amino sugars were ;2.5-fold greaterthan decreases in UDOC. These comparisons indicate thatamino sugars are considerably more reactive than bulk car-bon in upper ocean particulate and dissolved organic matter.A similar analysis of total hydrolyzable neutral sugars inthese samples (Skoog and Benner 1997) indicates neutralsugars are of similar reactivity as amino sugars. Overall,carbohydrates are among the most abundant and reactivecomponents of the upper ocean carbon and nitrogen cycles.
Glucosamine and galactosamine occur in relatively con-stant proportion in POM (;1.9) and UDOM (;1.5) fromdiverse geographic regions, suggesting these amino sugarsare closely associated and have similar reactivities. SimilarGlcN : GalN ratios of ;1.2 and ;1.9 were observed in ma-rine sediments and DOM derived from bacteria, respectively(Liebezeit 1993; Ogawa et al. 2001). Previous studies ofmarine POM and DOM noted fairly constant neutral sugarcompositions among samples from different ocean basins(McCarthy et al. 1996; Aluwihare et al. 1997; Borch andKirchman 1997; Skoog and Benner 1997; Kirchman et al.2001). This narrow range of neutral and amino sugar com-positions in marine POM and DOM is surprising. The bio-logical sources of these carbohydrates have quite variablecompositions. Moreover, these compositional similarities arecommon to very reactive organic matter that cycles rapidlyin the upper ocean, as well as refractory organic matter thatpersists in the deep ocean. These observations indicate thatcompositional uniformity is rapidly imprinted in marine or-ganic matter during early diagenesis. Results from a recentstudy monitoring neutral sugar compositions during experi-mental phytoplankton blooms (Meon and Kirchman 2001)also suggest that decomposition processes play an importantrole in shaping the carbohydrate composition of organic mat-ter in the ocean.
Muramic acid, peptidoglycan, and bacterial contributionsto marine organic matter—Peptidoglycan is a unique cellwall polymer of bacteria that consists of glycan strands of arepeating disaccharide (N-acetyl-glucosamine and N-acetyl-muramic acid) covalently cross-linked by peptide side chainscomposed of both L- and D-amino acids. Muramic acid isonly found in peptidoglycan, where it is linked to both theglycan strand and the peptide side chain. The peptide sidechains vary in amino acid composition, length, and positionof cross-linking, whereas muramic acid occurs in the con-stant stoichiometry of one residue per subunit of peptido-glycan (Schleifer and Kandler 1972). These characteristicsmake muramic acid an ideal biomarker for peptidoglycan.
Yields of muramic acid were highly variable among thecultured bacteria analyzed in this study. Previous analysesof muramic acid in terrestrial and marine bacteria have notedsimilar yields and variability (Moriarty 1977; Mimura andRomano 1985). The greatest difference in muramic acidyields is between Gram-positive and Gram-negative cells,with Gram-positive cells having 5- to 10-fold greater yieldsof muramic acid (Moriarty 1977; this study). Most bacteriain seawater and marine sediments are Gram-negative (Mor-iarty and Hayward 1982; Giovannoni 2000), and based on

126 Benner and Kaiser
analyses of cultured Gram-negative marine bacteria, we ex-pected muramic acid yields ranging from 15 to 35 nmol (mgC)21 (Moriarty 1977; Mimura and Romano 1985; this study).The yield (11 nmol muramic acid (mg C)21) in a populationof Trichodesmium sp. collected during a bloom in Gulf ofMexico surface water is similar to the lower values observedin cultures. However, muramic acid yields (3–5 nmol mur-amic acid (mg C)21) in natural populations of prokaryoticmicroorganisms collected from the open ocean were partic-ularly low in the present study. These low muramic acidyields in size-fractionated samples could result from marinebacteria with unusually thin cell walls or the presence ofdetrital material. Members of the domain Archaea are re-ported to be abundant in the deep ocean (Karner et al. 2001),but it is unlikely they were included with bacteria in thesurface water samples we analyzed. Archaea do not containmuramic acid in their cell walls (Brock et al. 1994), so in-clusion of Archaea in samples would result in lower yieldsof muramic acid.
The yields of muramic acid in POM were relatively highand similar to those in the natural populations of marinebacteria analyzed in this study. Based on direct counts ofbacteria (prokaryotes) in the initial water samples collectedfor POM isolation (Benner et al. 1997), we estimated theamount of muramic acid in POM associated with bacterialbiomass. We assumed that all cells counted were bacteria(i.e., no Archaea) and therefore had muramic acid in theircell walls. Karner et al. (2001) recently found that Archaeacomprise about half of the total direct count of cells in deepwater, so our calculations provide a maximal estimate ofmuramic acid associated with bacterial cells in deep water.For these calculations, we used 12.4 fg C cell21 for the car-bon content of marine bacteria (Fukuda et al. 1998) and 12.6nmol (mg C)21 for the muramic acid yield of marine bac-teria. This muramic acid yield is the average of all culturedand natural populations of marine bacteria analyzed in thisstudy. Based on this estimate, most of the muramic acidrecovered in the POM size class (0.1–60 mm) was associatedwith nonliving particles. On average, 3.4-fold more muramicacid was recovered in POM than could be accounted for inbacterial cells. Only 10–15% of the muramic acid in the twoPOM samples from 4,000 m was attributable to intact bac-terial cells. These data provide strong evidence of a sub-stantial reservoir of detrital particulate material of bacterialorigin throughout the ocean water column. This bacterial de-tritus includes cell wall material with fragments of pepti-doglycan, as indicated by the occurrence of muramic acid.Various processes, including viral lysis and protozoan graz-ing, could lead to the formation of bacterial detritus (Fuhr-man 2000; Nagata 2000; Strom 2000). These results indicatebacterial detritus is abundant and likely accounts for a sub-stantial fraction of the submicron and colloidal particles ob-served in seawater (Koike et al. 1990; Wells and Goldberg1991).
Several recent studies have identified specific biochemi-cals of bacterial origin in marine DOM, including a mem-brane protein (Tanoue et al. 1995), D-amino acids (McCarthyet al. 1998), and components of lipopolysaccharides (Wake-ham et al. in press). All of these biomarkers are componentsof bacterial cell wall complexes. Based on the abundance of
specific D-amino acids in marine UDOM, McCarthy et al.(1998) suggested that peptidoglycan was an abundant andimportant component of marine DON. In the present study,muramic acid was measured in all UDOM samples, confirm-ing the ubiquitous occurrence of peptidoglycan in marineDOM. However, yields of muramic acid in UDOM are morethan 10-fold lower than those of D-Ala, a common compo-nent of the peptide side chain in peptidoglycan. Muramicacid and D-Ala are typically found in equimolar yields inpeptidoglycan (Schleifer and Kandler 1972). Every subunitof peptidoglycan includes muramic acid, so there is muchmore D-Ala in UDOM than can be accounted for in pepti-doglycan. It appears the D-amino acids in UDOM are rem-nants derived from bacterial cell walls rather than compo-nents of intact subunits of peptidoglycan. Recent studies ofthe production and decomposition of DOM derived frommarine bacteria indicate that the glycan component of pep-tidoglycan is relatively rapidly degraded (Ogawa et al.2001). It is unclear why the peptide side chains of peptido-glycan would be more resistant to decomposition than theglycan strand. Further studies of the decomposition of bac-terial cell walls in seawater and the sources of D-amino acidsare needed to explain the prevalence of D-amino acids inmarine DOM.
Galactosamine and glucosamine were abundant in POMand UDOM and could be of bacterial origin, but these com-pounds have many potential sources. These amino sugarswere present in all organisms analyzed in this study, includ-ing members of the Bacteria, Archaea and Eukarya domains.As shown in the present study, marine phytoplankton andzooplankton that produce chitin are rich in glucosamine andhave high (.14) GlcN : GalN ratios. Muller et al. (1986)found high concentrations of glucosamine and trace concen-trations of galactosamine in large (.75 mm) surface waterparticles, indicating they were largely composed of chitinousmaterial and derived from zooplankton. In contrast, deep ma-rine sediments have GlcN : GalN ratios of 1–2, indicatingthey are largely derived from heterotrophic microorganisms(Liebezeit 1993). We observed GlcN : GalN ratios of 1.0–2.5in marine POM and UDOM from various depths and geo-graphic regions, and this could be indicative of a microbialorigin. Recent analyses of marine UDOM by direct temper-ature-resolved mass spectrometry noted the presence of N-acetyl amino sugars (Boon et al. 1998). These authors sug-gested a bacterial origin for most polysaccharides in UDOMbased on characteristic spectra produced under various ion-ization conditions and the occurrence of methylated neutralsugars, which are biomarkers of bacterial origin. Glucos-amine and galactosamine were important components of re-fractory DOM derived from bacteria, and they remained infairly constant proportion (;1.9, respectively) during a year-long decomposition study (Ogawa et al. 2001). The macro-molecular identity of this polymeric material with relativelylow and constant GlcN : GalN ratios remains unknown. Itsbiological origin is also uncertain, but there is growing ev-idence of a major prokaryotic source in the ocean.
References
ALUWIHARE, L. I., D. J. REPETA, AND R. F. CHEN. 1997. A majorbiopolymeric component to dissolved organic carbon in surfaceseawater. Nature 387: 166–169.

127Amino sugars in marine organic matter
BENNER, R. 2002. Chemical composition and reactivity, p. 59–90.In D. Hansell and C. Carlson [eds.], Biogeochemistry of ma-rine dissolved organic matter. Academic.
, AND B. BIDDANDA. 1998. Photochemical transformationsof surface and deep marine dissolved organic matter: Effectson bacterial growth. Limnol. Oceanogr. 43: 1373–1378.
, AND M. STROM. 1993. A critical evaluation of the analyt-ical blank associated with DOC measurements by high-tem-perature catalytic oxidation. Mar. Chem. 41: 153–160.
, J. D. PAKULSKI, M. MCCARTHY, J. I. HEDGES, AND P. G.HATCHER. 1992. Bulk chemical characterization of dissolvedorganic matter in the ocean. Science 255: 1561–1564.
, B. BIDDANDA, B. BLACK, AND M. MCCARTHY. 1997.Abundance, distribution, and stable carbon and nitrogen iso-topic compositions of marine organic matter isolated by tan-gential-flow ultrafiltration. Mar. Chem. 57: 243–263.
BIDDANDA, B. A. 1995. Trichodesmium bloom in Gulf of Mexico,summer 1995. IOC/UNESCO Harmful Algae News 12/13.
, AND R. BENNER. 1997. Carbon, nitrogen and carbohydratefluxes during the production of particulate and dissolved or-ganic matter by marine phytoplankton. Limnol. Oceanogr. 42:506–518.
BOON, J. J., V. A. KLAP, AND T. I. EGLINTON. 1998. Molecularcharacterization of microgram amounts of oceanic colloidal or-ganic matter by direct temperature-resolved ammonia chemicalionization mass spectrometry. Org. Geochem. 29: 1051–1061.
BORCH, N. H., AND D. L. KIRCHMAN. 1997. Concentration and com-position of dissolved combined neutral sugars (polysaccha-rides) in seawater determined by HPLC-PAD. Mar. Chem. 57:85–95.
BROCK, T. D., M. T. MADIGAN, J. M. MARTINKO, AND J. PARKER.1994. Biology of microorganisms, 7th ed. Prentice Hall.
DAUWE, B., AND J. J. MIDDELBURG. 1998. Amino acids and hex-osamines as indicators of organic matter degradation state inNorth Sea sediments. Limnol. Oceanogr. 43: 782–798.
FUHRMAN, J. 2000. Impact of viruses on bacterial processes, p. 327–350. In D. L. Kirchman [ed.], Microbial ecology of the oceans.Wiley.
FUKUDA, R., H. OGAWA, T. NAGATA, AND I. KOIKE. 1998. Directdetermination of carbon and nitrogen contents of natural bac-terial assemblages in marine environments. Appl. Environ. Mi-crobiol. 64: 3352–3358.
GIOVANNONI, S., AND M. RAPPE. 2000. Evolution, diversity, andmolecular ecology of marine prokaryotes, p. 47–84. In D. L.Kirchman [ed.], Microbal ecology of the oceans. Wiley.
HUNG, C.-C., D. TANG, K. W. WARNKEN, AND P. H. SANTSCHI.2001. Distributions of carbohydrates, including uronic acids,in estuarine waters of Galveston Bay. Mar. Chem. 73: 305–318.
ITTEKOT, V., W. G. DEUSER, AND E. T. DEGENS. 1984. Seasonalityin the fluxes of sugars, amino acids, and amino sugars to thedeep ocean: Sargasso Sea. Deep-Sea Res. 31: 1057–1069.
KAISER, K., AND R. BENNER. 2000. Determination of amino sugarsin environmental samples with high salt content by high-per-formance anion-exchange chromatography and pulsed amper-ometric detection. Anal. Chem. 72: 2566–2572.
KARNER, M. B., E. F. DELONG, AND D. M. KARL. 2001. Archaealdominance in the mesopelagic zone of the Pacific Ocean. Na-ture 409: 507–510.
KIRCHMAN, D. L., AND J. WHITE. 1999. Hydrolysis and minerali-zation of chitin in the Delaware estuary. Aquat. Microb. Ecol.18: 187–196.
, B. MEON, H. W. DUCKLOW, C. A. CARLSON, D. A. HAN-SELL, AND G. F. STEWARD. 2001. Glucose fluxes and concen-trations of dissolved combined neutral sugars (polysaccharides)
in Ross Sea and Polar Front Zone, Antarctica. Deep-Sea Res.II 48: 4179–4197.
KOIKE, I., S. HARA, K. TERAUCHI, AND K. KOGURE. 1990. Role ofsub-micrometer particles in the ocean. Nature 345: 242–244.
LIEBEZEIT, G. 1993. Amino sugars in Bransfield Strait and WeddellSea sediments. Senckenb. Marit. 23: 29–35.
MCCARTHY, M., J. I. HEDGES, AND R. BENNER. 1996. Major bio-chemical composition of dissolved high molecular weight or-ganic matter in seawater. Mar. Chem. 55: 281–298.
, T. PRATUM, J. I. HEDGES, AND R. BENNER. 1997. Chemicalcomposition of dissolved organic nitrogen in the ocean. Nature390: 150–154.
, J. I. HEDGES, AND R. BENNER. 1998. Major bacterial con-tribution to marine dissolved organic nitrogen. Science 281:231–234.
MCLACHLAN, J., A. G. MCINNES, AND M. FALK. 1965. Studies onthe chitan (chitin: poly-N-acetyl-glucosamine) fibers of the di-atom Thalassiosira fluviatilis Hustedt. Production and isolationof chitan fibers. Can. J. Bot. 43: 707–713.
MEON, B., AND D. L. KIRCHMAN. 2001. Dynamics and molecularcomposition of dissolved organic material during experimentalphytoplankton blooms. Mar. Chem. 75: 185–199.
MIMURA, T., AND J.-C. ROMANO. 1985. Muramic acid measurementsfor bacterial investigations in marine environments by high-pressure liquid chromatography. Appl. Environ. Microbiol. 50:229–237.
MOPPER, K., J. ZHOU, K. S. RAMANA, U. PASSOW, H. G. DAM, AND
D. T. DRAPEAU. 1995. The role of surface-active carbohydratesin the flocculation of a diatom bloom in a mesocosm. Deep-Sea Res. 42: 47–73.
MORIARTY, D. J. W. 1977. Improved method using muramic acidto estimate biomass of bacteria in sediments. Oecologia 26:317–323.
, AND A. C. HAYWARD. 1982. Ultrastructure of bacteria andthe proportion of Gram-negative bacteria in marine sediments.Microb. Ecol. 8: 1–14.
MULLER, P. J., E. SUESS, AND C. A. UNGERER. 1986. Amino acidsand amino sugars of surface particulate and sediment trap ma-terial from waters of the Scotia Sea. Deep-Sea Res. 33: 819–838.
MUZZARELLI, R. A. A. 1977. Chitin. Pergamon.NAGATA, T. 2000. Production mechanisms of dissolved organic mat-
ter, p. 121–152. In D. L. Kirchman [ed.], Microbial ecology ofthe oceans. Wiley.
OBERNOSTERER, I., B. REITNER, AND G. J. HERNDL. 1999. Contrast-ing effects of solar radiation on dissolved organic matter andits bioavailability to marine bacterioplankton. Limnol. Ocean-ogr. 44: 1645–1654.
OGAWA, H., Y. AMAGAI, I. KOIKE, K. KAISER, AND R. BENNER.2001. Production of refractory dissolved organic matter bybacteria. Science 292: 917–920.
PAKULSKI, J. D., AND R. BENNER. 1994. Abundance and distributionof dissolved carbohydrates in the ocean. Limnol. Oceanogr. 39:930–940.
ROMANKEVICH, E. A. 1984. Geochemistry of organic matter in theocean. Springer.
SCHLEIFER, K. H., AND O. KANDLER. 1972. Peptidoglycan types ofbacterial cell walls and their taxonomic implications. Bacteriol.Rev. 36: 407–477.
SHARON, N. 1965. Microorganisms, plants, and invertebrates, p. 1–45. In E. A. Balazs and R. W. Jeanloz [eds.], The amino sugars:the chemistry and biology of compounds containing aminosugars. Academic.
SKOOG, A., AND R. BENNER. 1997. Aldoses in various size fractionsof marine organic matter: Implications for carbon cycling.Limnol. Oceanogr. 42: 1803–1813.

128 Benner and Kaiser
STROM, S. L. 2000. Bacterivory: Interactions between bacteria andtheir grazers, p. 351–386. In D. L. Kirchman [ed.], Microbialecology of the oceans. Wiley.
TANOUE, E., S. NISHIYAMA, M. KAMO, AND A. TSUGITA. 1995. Bac-terial membranes: Possible source of a major dissolved proteinin seawater. Geochim. Cosmochim. Acta 59: 2643–2648.
WAKEHAM, S. G., T. K. PEASE, AND R. BENNER. In press. Hydroxy
fatty acids in marine dissolved organic matter as indicators ofbacterial membrane material. Org. Geochem.
WELLS, M. L., AND E. D. GOLDBERG. 1991. Occurrence of smallcolloids in seawater. Nature 353: 342–344.
Received: 15 March 2002Accepted: 21 August 2002
Amended: 17 September 2002





























