Embed Size (px)
Citation preview

Biology of Human Tumors
Activation of IL6/IGFIR Confers Poor Prognosis ofHBV-Related Hepatocellular Carcinoma throughInduction of OCT4/NANOG ExpressionTe-Sheng Chang1,2,3, Yu-Chih Wu1, Ching-Chi Chi4,5,Wei-Chi Su1,6, Pey-Jium Chang2,Kam-Fai Lee7,Tao-HsinTung8, JuiWang9, Jun-JenLiu10, Shui-YiTung3,4, Liang-MouKuo2,11,Hong-Nerng Ho12,13, Thai-Yen Ling14, and Yen-Hua Huang1,6,15,16,17
Abstract
Purpose: To unravel the role of interleukin (IL)-6 and insulin-like growth factor (IGF)-I receptor (IGFIR) in expressing stemness-related properties and to evaluate the prognostic values of plu-ripotent transcription factor OCT4/NANOG, and IGFIR in hepa-tocellular carcinoma (HCC).
Experimental Design: Serum levels of IL6 were detectedusing ELISA assays (n ¼ 120). The effects of IL6/IGFI onstemness expression in HCC were examined using OCT4/NANOG promoter luciferase reporter, RNA interference, sec-ondary sphere formation, side population, and xenograftanimal models. The OCT4/NANOG protein and phospho-IGFI receptor (p-IGFIR) in tissues were detected by Westernblotting (n ¼ 8) and immunohistochemical staining (n ¼ 85).OCT4, NANOG, and IGFIR expression levels in tissues (n ¼191) were analyzed by real-time qRT-PCR and was correlatedwith early tumor recurrence using the Kaplan–Meier survivalanalysis.
Results: A high positive correlation between the expressionlevels of OCT4/NANOG and IGFIR/p-IGFIR in human HCCtissues was observed. The concurrent expression of OCT4/NANOG/IGFIR was mostly confined to hepatitis B virus(HBV)–related HCC (HBV-HCC) and was significantly correlatedwith early tumor recurrence. High serum levels of IL6 weresignificantly correlated with high OCT4/NANOG expression. IL6stimulated an autocrine IGFI/IGFIR expression STAT3dependent-ly, which stimulated stemness-related properties in both the celllines and the xenografted mouse tumors. The inhibition of IGFIRactivation by either RNA interference or by treatment with theinhibitor picropodophyllin (PPP) significantly suppressed theIL6-induced stemness-related properties both in vitro and in vivo.
Conclusions: The expression of pluripotency-related genes isassociated with early tumor recurrence and is regulated by IL6-induced IGF/IGFIR activation, particularly in HBV-HCC. ClinCancer Res; 21(1); 201–10. �2014 AACR.
IntroductionHepatocellular carcinoma (HCC) is the sixth most common
cancer, and the third leading cause of cancer-related mortalityworldwide (1). Surgical resection can be curative in the earlystages of HCC, but the risk of recurrence remains high. As tumorstemness properties are related to HCC recurrence (2), investi-gating the mechanisms responsible for the expression of tumor
stemness could lead to strategies to prevent tumor recurrence andto improvements in the efficacy of HCC treatment.
Clinical and epidemiologic studies have identified a clearrelationship between chronic hepatic inflammation and HCCpathogenesis (3). The inflammation associated with chronichepatitis B virus (HBV) infection contributes to HCC (4). Fol-lowing HBV infection, Kupffer cells activate nuclear factor-kB
1Department of Biochemistry and Molecular Cell Biology, Collegeof Medicine, Taipei Medical University, Taipei, Taiwan. 2GraduateInstitute of Clinical Medical Sciences, College of Medicine, ChangGung University, Taoyuan, Taiwan. 3Department of Gastroenter-ology and Hepatology, Chang Gung Memorial Hospital, Chiayi,Taiwan. 4College of Medicine, Chang Gung University, Taoyuan,Taiwan. 5Department of Dermatology and Centre for Evidence-Based Medicine, Chang Gung Memorial Hospital, Chiayi, Taiwan.6Graduate Institute of Medical Sciences, College of Medicine,Taipei Medical University, Taipei, Taiwan. 7Department of Pathol-ogy, Chang Gung Memorial Hospital, Chiayi, Taiwan. 8Departmentof Medical Research and Education, Cheng-Hsin General Hospital,and Faculty of Public Health, School of Medicine, Fu-Jen CatholicUniversity, Taipei, Taiwan. 9School of Public Health, College ofPublic Health and Nutrition, Taipei Medical University, Taipei, Tai-wan. 10School of Medical Laboratory Sciences and Biotechnology,Taipei Medical University, Taipei, Taiwan. 11Department of GeneralSurgery, Chang Gung Memorial Hospital, Chiayi, Taiwan. 12Grad-uate Institute of Clinical Genomics, College of Medicine, NationalTaiwan University, Taipei, Taiwan. 13Department of Obstetrics andGynecology, Division of Reproductive Endocrinology and Infer-
tility, National Taiwan University and Hospital, Taipei, Taiwan.14Department of Pharmacology, College of Medicine, NationalTaiwan University, Taipei, Taiwan. 15Center for Reproductive Med-icine,Taipei Medical University Hospital, Taipei, Taiwan. 16Compre-hensive Cancer Center of Taipei Medical University,Taipei,Taiwan.17The Ph.D. Program for Translational Medicine, Taipei MedicalUniversity, Taipei, Taiwan.
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
T.-S. Chang and Y.-C. Wu contributed equally to this article.
Corresponding Authors: Yen-Hua Huang, Taipei Medical University, No. 250,Wuxing Street, Taipei 110, Taiwan. Phone: 886-2-27361661, ext. 3150; Fax:886-2-27356689; E-mail: [email protected]; or Thai-Yen Ling, NationalTaiwan University, No. 1, Section I, Je-Ai Road, Taipei 100, Taiwan. Phone:886-2-33936987; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-13-3274
�2014 American Association for Cancer Research.
ClinicalCancerResearch
www.aacrjournals.org 201
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

and secrete inflammatory cytokines, such as tumornecrosis factor-a, interleukin (IL)-8, IL1b, and IL6 (5). Meanwhile, the HBV Xprotein (HBx) can stimulate IL6 expression in hepatocytes andHCCs (6). A clinical association between IL6 and HBV-relatedHCC (HBV-HCC) has been previously reported. It has beenshown that IL6 controls early gene expression in HBV infection(5), and that a high serum level of IL6 can predict future HCCdevelopment in patients with chronic hepatitis B (7). It wasrecently suggested that aberrant IL6-mediated signaling cantransform hepatic progenitor cells in HBx transgenic mice(8), and that aberrant IL6-mediated signaling is responsiblefor liver progenitor/stem cells giving rise to cancer (9). Inaddition, it has been demonstrated that CD24þ liver tumor-initiating cells can drive self-renewal and tumor initiationthrough STAT3-mediated NANOG regulation (10). Theseobservations highlight the association between IL6 and stem-ness, which may include octamer 4 (OCT4)/NANOG expres-sion, in HBV-induced liver tumorigenesis.
In addition to stimulating IL6 secretion (6), HBV is highlyassociated with the expression of both insulin-like growth factor(IGF) and the IGFI receptor (IGFIR) in HCC (11–15). Activationof IGFIR has been shown to promote HCCdevelopment (16, 17);interestingly, the upregulation of IGFIR and its downstream Aktsignaling is predominantly confined to HBV-HCC (13).
HBV apparently links the endocrine signals of both the IL6 andIGFIR in HCC formation. However, the relationships among IL6,IGFIR-mediated signaling, and pluripotency-related gene expres-sion (OCT4 andNANOG) in HBV-HCC are unclear. The purposeof this study was to investigate the interplay among IL6, IGFIR,and OCT4/NANOG expression and the clinical significance ofthese relationships. Here, we show that the upregulation of IGFIR,OCT4, andNANOG expression is associated withHCC recurrenceand occurs predominately in patients with HBV-HCC. We foundthat patients with HBV-HCC have a higher serum level of IL6, andthe IL6 increases the expression of autocrine IGFI and IGFIR,which then promote expression of the transcription factors OCT4andNANOGina STAT3-dependentmanner. This studyhighlightsthe significance of IL6-IGFI/IGFIR–mediated signaling as a poten-tial therapeutic target for individualized adjuvant therapy forpatients with HBV-related HCC.
Materials and MethodsCell lines and HCC tissues
The Hep3B (HBVþHBsAgþ humanHCC, HB-8064) andHepG2(HBV� human hepatoblastoma, HB-8065) cells were purchasedfrom the American Type Culture Collection. The Huh7 (HBV�
human HCC) cells were obtained from the Japanese Collection ofResearch Bioresources. The HepG2.2.15 (HBVþHBsAgþ humanhepatoblastoma) cells were kindly provided by Dr. Jun-Jen Liu(Institute of Medical Biotechnology, Taipei Medical University,Taipei, Taiwan). The PLC5 (HBVþHBsAgþ humanHCC) cells werekindly provided by Dr. Ching-Tai Lin (Department of MedicalResearch, Chang Gung Memorial Hospital, Chiayi, Taiwan).
Cells were maintained in Dulbecco's Modified Eagle Medium(DMEM;Gibco-BRL) supplemented with 10% fetal bovine serum(FBS) and 3.7 g/L sodium bicarbonate. For the IGFI treatment,cells were cultured in serum-free media with IGFI (0–50 ng/mL;R&D Systems); for IL6 treatments, cells in serum-free media weretreated with IL6 (50 ng/mL; Sigma-Aldrich) with or without theJAK/STAT3 inhibitor AG490 (Cayman Chemical) or S3I-201(Merck Millipore). The solutions of 50 mmol/L AG490 and1 mmol/L picropodophyllin (PPP; Calbiochem), an IGFIR-phos-phorylation-inhibitor, were prepared in dimethyl sulfoxide(DMSO).
Frozen HCC tissue samples were obtained from 191 patients(146men)of ages 21 to 80 yearswith amean ageof 60.57�11.24years who had received curative hepatectomy between 2004 and2012 at Chang Gung Memorial Hospital (Chiayi, Taiwan).Among the 191patients, formalin-fixed, paraffin-embeddedHCCtissue samples were available for 85 patients, and serum sampleswere available for 120 patients (Supplementary Fig. S1). Themean follow-up duration was 32.43 months (range, 1.2–106months). Our study was approved by the Institutional ReviewBoard of Chang Gung Medical Foundation. The study was con-ducted according to the Reporting Recommendations for TumorMarker Prognostic Studies (REMARK) guideline (18) and a ret-rospectively analysis, pathologic evaluation, and statistical anal-ysis plan. The REMARK guideline was shown in SupplementaryTable S1.
RNA isolation and real-time quantitative reverse-transcriptionPCR
Cell lines and frozen HCC tissues were subjected to total RNAisolation followed by real-time quantitative reverse transcription-PCR (qRT-PCR). Detailed descriptions are provided in the Sup-plementary Materials and Methods.
Western blot analysisProteins from the human HCC tissues and the cell lines
were extracted with lysis buffer containing 0.1% SDS, 0.1%Nonidet P-40, 0.5% Sodium deoxycholate, 150 mmol/L NaCl,1 mmol/L EDTA, and 10 mmol/L Tris–HCl (pH 7.5) with aprotease inhibitor cocktail (Roche Diagnostics). Detailed descrip-tions are provided in the Supplementary Materials and Methods.
Detailed descriptions of the RNA isolation, real-time qRT-PCRanalysis, Western blotting, immunohistochemical staining, dualluciferase assay, shRNA and lentivirus production, flow cytometryfor side population cells, tumor sphere formation assay, xenografttumormodel, enzyme-linked immunosorbent assay (ELISA), andstatistical analysis are provided in the Supplementary Materialsand Methods.
Translational Relevance
This study shows that patients with hepatitis B virus (HBV)–related hepatocellular carcinoma (HBV-HCC) exhibit a highrate of concurrent expression of interleukin (IL)-6 in sera, andOCT4, NANOG, and insulin-like growth factor (IGF) I recep-tor-(IGFIR) in primary tumors.We found the IL6–IGFIR signalmediates the expression of stemness-related properties that areassociatedwith early recurrence andpoor prognosis inpatientswith HCC. The IL6 stimulates the expression of IGFIR andautocrinal IGFI STAT3 dependently inHBV-HCCs. Blockage ofIGFIR signal by IGFIR-specific short hairpin RNA (shRNA) orIGFIR phosphorylation inhibitor PPP suppresses the IL6-induced stemness-related properties in vitro and in a xenograftmouse model. These results suggest that IL6–IGFIR signal is apotential target for individualized adjuvant therapy and pre-venting early tumor relapse of HBV-HCCs.
Chang et al.
Clin Cancer Res; 21(1) January 1, 2015 Clinical Cancer Research202
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

ResultsPositive correlation between OCT4 and NANOG mRNAexpression and IGFIRmRNA expression in human HCC tissues
To examine the associations between OCT4, NANOG,and IGFIR mRNA expression in human HCC, the tumor tissue(T) and adjacent peritumor tissue (PT) from 191 frozen tissuesamples were analyzed using real-time qRT-PCR. As shownin Supplementary Fig. S2, mRNA levels of IGFIR were signif-icantly correlated with mRNA levels of both OCT4 (R ¼ 0.7281;P < 0.0001) and NANOG (R ¼ 0.7144; P < 0.0001).
The expression levels of OCT4, NANOG, and IGFIR were alsocompared among patients with HCC infected with hepatitis Cvirus (HCV), HBV, HBV and HCV (BC), and those with neitherHBV nor HCV (NBNC). The patients with HBV-HCC hadsignificantly higher levels of OCT4, NANOG, and IGFIR mRNAcompared with patients with HCV-HCC (Fig. 1A; P < 0.001 forall comparisons). When the 191 patients with HCC weredivided into high (gene expression level of T/PT � 2) and lowexpression groups (gene expression level of T/PT < 2), thepatients with HBV-HCC showed significantly higher rates of
synchronous high expression of OCT4, NANOG, and IGFIRmRNA (Table 1; P ¼ 0.038).
Association ofOCT4,NANOG, and IGFIR expression with earlytumor recurrence in human HCC
The clinical relevance ofOCT4,NANOG, and IGFIR expressionon the early tumor recurrence (within 2 years) of HCC wasexamined on the basis of the gene expression levels (either high[T/PT � 2] or low [T/PT < 2]) of these three genes in the 191patients with HCC (Table 2, Fig. 1B, and Supplementary Fig. S3).Univariate analysis revealed that expression ofOCT4 (P < 0.001),NANOG (P < 0.001), IGFIR (P < 0.001), and high expression of allthree genes (P < 0.001) were significantly associated with earlytumor recurrence (Table 2). HBV (P ¼ 0.005), HCV (P ¼ 0.029),tumor—node–metastasis (TNM) stage, microvascular/macrovas-cular invasion, satellite nodules, and tumor size (P¼ 0.001) werealso significantly associatedwith early tumor recurrence (Table 2).Multivariate analysis also demonstrated that expression of OCT4(P¼ 0.012),NANOG (P¼ 0.013), IGFIR (P < 0.001) and all threegenes (P ¼ 0.001) were significantly associated with early tumor
Figure 1.Correlation of the expressions of OCT4, NANOG, and IGFIR/p-IGFIR with the recurrence of human HCC. A, correlations of transcriptional levels (�DDCt of tumor/peritumor) of IGFIR with OCT4 or NANOG in HCC tumor samples (T) and adjacent peritumor liver tissues (PT) from patients without HCV or HBV (NBNC, n ¼ 13);patientswith HBV-HCC (HBV, n¼ 73); patientswith HCV-HCC (HCV, n¼ 81); and dually infected patients (BC, n¼ 24). Relationships between the viral etiologies andthe transcriptional levels of OCT4, NANOG, and IGFIR are shown. ��� , P < 0.001 by the Mann–Whitney U test. B, the Kaplan–Meier curves of time-to-early tumorrecurrence after HCC resection for the transcriptional levels ofOCT4,NANOG, and IGFIR in human HCC tissues (n¼ 191). After normalization with the correspondingperitumor (PT) tissue sample, the tumor (T) tissues that expressed low levels (T/PT < 2-fold) ofOCT4 (n¼ 143),NANOG (n¼ 150), or IGFIR (n¼ 137) were comparedwith tumor tissues that expressed high levels (T/PT� 2-fold) ofOCT4 (n¼ 48), NANOG (n¼ 41), or IGFIR (n¼ 54). The Kaplan–Meier curves of time-to-early tumorrecurrence for the patients with HBV-HCC and HCV-HCC following hepatectomy was also shown based on the Peto-Peto-Prentice test. C, top, theimmunohistochemical staining of OCT4, NANOG, and p-IGFIR in one representative tissue sample from patient with HBV-HCC (bar, 50 mm). Bottom, the significantpositive correlations between OCT4 and NANOGprotein levels with p-IGFIR in humanHCC tissues by immunohistochemical staining (n¼ 85). Spearman correlationanalysis. D, Western blot analysis of the expression of p-IGFIR, IGFIR, OCT4, and NANOG protein in the tumor (T) and peri-tumor (PT) cells of eight representativesamples.
IL6/IGFIR in Stemness Expression and HCC Early Recurrence
www.aacrjournals.org Clin Cancer Res; 21(1) January 1, 2015 203
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

recurrence. The Kaplan–Meier curves of early tumor recurrencewere significant for high versus low expression of the individualgenes (OCT4, NANOG, or IGFIR; Fig. 1B; P < 0.0001). TheKaplan–Meier curves of early tumor recurrence for concurrenthigh versus concurrent low expression of OCT4 and IGFIR orNANOG and IGFIR (Supplementary Fig. S3A; P < 0.0001), forconcurrent high expression of all three genes versus two genes orless (Supplementary Fig. S3B, left; P < 0.0001) and for highexpression of at least one of the three genes versus none of thethree genes (Supplementary Fig. S3B, right; P < 0.0001) wereall significant. In addition, concurrent high expression ofall three genes (OCT4, NANOG, and IGFIR) occurred 71% inhepatitis B–positive HCC (P ¼ 0.014), but only 35% in hepatitis
C–positive HCC (P ¼ 0.017; Supplementary Table S2). Concur-rent high expression of all three genes was also correlated with thepresence of microvascular invasion (P ¼ 0.037) and satellitenodules (P ¼ 0.008; Supplementary Table S2). There wasalso a significant difference between patients with HBV-HCCand patients with HCV-HCC in early tumor recurrence (Fig. 1B;P ¼ 0.005). The Kaplan–Meier curves of overall survival (OS;Supplementary Fig. S4) and the time-to-recurrence (Supplemen-tary Fig. S5) for the gene expression were shown. The training/validation set analysis was further examined (Supplementary Fig.S6). Because of the lack of an external database of pluripotenttranscriptional factor OCT4 and NANOG expression in HCC, werandomly divided our samples into two equal groups, the trainedsamples and the validation samples, to examine whether theassociations between biologic measures and mortality were validin our proposedmodel. As shown in Supplementary Fig. S6, therewere no significant differences between the observed and pre-dicted survival curves of IGFIR, OCT4, and NANOG (all P > 0.05by the log-rank test). These results confirmed that our proposedmodel of mortality assessment through the use of biologic mea-sures is valid.
Positive correlation between OCT4 and NANOG proteinexpression and IGFIR phosphorylation in human HCCtissues
OCT4 andNANOGprotein expression levels and the activationof IGFIR by phosphorylation (p-IGFIR) in human HCC tumortissues were examined by immunohistochemical staining (Fig.1C). Figure 1C shows the immunohistochemical staining ofOCT4, NANOG, and p-IGFIR in tumor and peri-tumor samplesfrom one representative patient with HBV-HCC. The level of p-IGFIR positively correlatedwith the levels ofOCT4 (R¼ 0.2377; P¼ 0.0285) and NANOG (R ¼ 0.2151; P ¼ 0.048) in HCC tumortissues (Fig. 1C; n¼ 85). The expression of p-IGFIR/IGFIR, OCT4,and NANOG proteins in the tumor tissues was verified usingWestern blotting (Fig. 1D). These results highlight the associationbetween IGFIR activation and expression of the pluripotenttranscription factors OCT4 and NANOG in human HCC.
IGFI and IGFIR regulate expression of OCT4 and NANOG inHBV-related human HCC cells
As the patient with HBV-HCC population displayed a greaterproportion of patients with concurrent high expression ofOCT4/NANOG and IGFIR (Table 1), and there is a positive correlationbetween the level of p-IGFIR (IGFIR activation) and OCT4/NANOG expression in HCC (Fig. 1C), we examined the effect ofIGFI, a high-affinity ligand of IGFIR, on OCT4 and NANOG
Table 2. Variables associated with early tumor recurrence after hepatectomyfor HCC (N ¼ 191)
Recurrenceb
Multivariate analysis
VariableUnivariateanalysis P P HR (95% CI)
Gender (male vs. female) 0.571 NS —
Age, y (<55 vs. �55) 0.469 NS —
HBV (yes vs. no) 0.005a 0.216 1.49 (0.79–2.80)HCV (yes vs. no) 0.029a 0.984 0.99 (0.53–1.83)Bilirubin, mg/dL (<1.2 vs. �1.2) 0.232 NS —
Albumin, g/dL (<3.5 vs. �3.5) 0.078 0.276 1.34 (0.79–2.26)ALT, U/L (<35 vs. �35) 0.585 NS —
PT, INR (<1.2 vs. �1.2) 0.141 NS —
AFP, ng/mL (<400 vs. �400) 0.246 NS —
ICG, retention rate (<15% vs. �15%) 0.135 NS —
TNM stage (T1/2 vs. T3/4) <0.001a 0.803 1.10 (0.53–2.24)Multiple tumors (yes vs. no) 0.749 NS —
Child–Pugh class (A vs. B) 0.505 NS —
Complete tumor capsule (yes vs. no) 0.362 NS —
Microvascular invasion (yes vs. no) <0.001a 0.005a 2.22 (1.28–3.84)Macrovascular invasion (yes vs. no) <0.001a 0.226 1.97 (0.66–5.87)Cutoff margin free (yes vs. no) 0.079 0.282 0.60 (0.24–1.52)Differentiation grade (1/2 vs. 3/4) 0.027a 0.511 1.20 (0.69–2.10)Tumor size, cm (< 3 vs. �3) 0.001a 0.222 1.47 (0.79–2.74)Satellite nodules (yes vs. no) <0.001a 0.053 1.87 (0.99–3.52)OCT4 expression (<2X vs. �2X) <0.001a 0.012a 1.97 (1.16–3.33)NANOG expression (<2X vs. �2X) <0.001a 0.013a 1.98 (1.15–3.39)IGFIR expression (<2X vs. �2X) <0.001a <0.001a 2.86 (1.70–1.81)High expression of all three genes of
OCT4, NANOG, and IGFIR (<2X vs.�2X)
<0.001a 0.001a 2.65 (1.51–4.63)
Abbreviations: AFP, serum a-fetoprotein; ALT, alanine aminotransferase; ICG,indocyanine green; PT, prothrombin time; INR, international normalized ratio;Tumor size, the largest one if multiple.aP < 0.05.bTime to early recurrence (less than 2 years).
Table 1. Relationship between viral etiology and concurrent gene expression
Number of high Etiologyexpression genes NBNC (n ¼ 13) HBV (n ¼ 73) HCV (n ¼ 81) BC (n ¼ 24)
0 69.2% (9/13) 46.6% (34/73) 74.1% (60/81) 75.0% (18/24)1 23.1% (3/13) 16.4% (12/73) 13.6% (11/81) 8.3% (2/24)2 0% (0/13) 11.0% (8/73) 2.5% (2/81) 4.2% (1/24)3 7.7% (1/13) 26.0% (19/73) 9.9% (8/81) 12.5% (3/24)0/1 12 (92.3%) 46 (63.0%) 71 (87.7%) 20 (83.3%)2/3 1 (7.7%) 27 (37.0%) 10 (12.3%) 4 (16.7%)Pa Referent 0.038b 0.628 0.446
NOTE: Expression genes include OCT4, NANOG, and IGFIR, n ¼ 191.ax2 test compared with the NBNC group.bP < 0.05.
Chang et al.
Clin Cancer Res; 21(1) January 1, 2015 Clinical Cancer Research204
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

expression inHBV-HCC cells. We used humanHCC cell lines thatcontain (Hep3B and HepG2.2.15) or do not contain (Huh7) theHBV genome. mRNA levels ofOCT4 andNANOGweremeasuredusing qRT-PCR. As shown in Supplementary Fig. S7, IGFI treat-ment significantly increasedOCT4 and/orNANOGmRNAexpres-sion inHBVþHBsAgþHep3B andHepG2.2.15 cells, but not in theHBV�HBsAg� Huh7 cell line. Because of the hepatoblastomacharacter of HepG2.2.15 cells, we used Hep3B and Huh 7 as thedifferentiation cell model for further study. We found that IGFIhad a dose-dependent effect on OCT4 and NANOGmRNA levelsin Hep3B cells, although the effect on NANOG mRNA levels wasnot as pronounced. Huh7 showed a negative response to IGFItreatment (Fig. 2A). In addition, we performed experiments usingshRNAs to knockdown the expression of the endogenous IGFIRprotein (shIGFIR#1 and shIGFIR#2) and found that shIGFIRsignificantly suppressed the IGFI-induced OCT4 and NANOGexpression in Hep3B (Fig. 2B) and HepG2.2.15 cells (Supple-mentary Fig. S8A).We also found that IGFI significantly increasedthe number of secondary tumor spheres of Hep3B andHepG2.2.15 cells in a serum-free medium, and that this effect
was suppressed by either PPP (an IGFIR phosphorylation inhib-itor) or shIGFIR#2 (Fig. 2C and Supplementary Figs. S8B andS8C). The IGFI effect on stemness-related properties was furthersupported by a side population assay, in which IGFI increased thepercentage of Hep3B cells with the side population phenotype,and shIGFIR#2 significantly suppressed this effect of IGFI (Fig. 2Dand Supplementary Fig. S9). These results suggest that IGFI andIGFIR may regulate the expression of OCT4 and/or NANOG inhuman HBV-HCC cells.
IL6 activates the expression of IGFI, IGFIR, and OCT4/NANOGin HBV-related human HCC cells
IL6 has been associated with the expression of genes thatcontribute to cancer stemness in HCC (7–9). We detectedsignificantly higher levels of IL6 in the serum from patientswith HBV-HCC compared with serum from patients with HCV-HCC (Fig. 3A, top). Patients with HCC who displayed highlevels of OCT4 and NANOG expression also had significantlyhigher serum levels of IL6 (Fig. 3A, bottom), and the patientswith high serum IL6 levels exhibited early tumor recurrence
Figure 2.IGFI/IGFIR signaling regulatesstemness expression in human HBV-HCC cell lines. A, the dose effect of IGFI(0–50 ng/mL) on mRNA levels ofOCT4 and NANOG in Hep3B and Huh7cells. B, OCT4 and NANOG proteinlevels and relative luciferase activity ofOCT4/NANOG promoter-luciferaseshCtrl.- or shIGFIR Hep3B cells withIGFI treatment (50 ng/mL). C,secondary sphere formation of Hep3Bcells in serum free medium control,IGFI (50 ng/mL), IGFI and DMSO, IGFIþ PPP (1 mmol/L), IGFI þ shIGFIR#2,and the quantitative analysis. Bar, 100mm. D, the percentage of Hep3B cellswith side population phenotypecultivated in medium only (Mock),IGFI (50 ng/mL), IGFIþ shCtrl., or IGFIþ shIGFIR#2. Note that shCtrl. is thecontrol vector, and shIGFIR refers tothe shRNA of IGFIR of shIGFIR#1 andshIGFIR#2. � , #, P <0.05; �� , ##, P <0.01;and ��� , ###, P < 0.001 by the Studentt test.
IL6/IGFIR in Stemness Expression and HCC Early Recurrence
www.aacrjournals.org Clin Cancer Res; 21(1) January 1, 2015 205
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
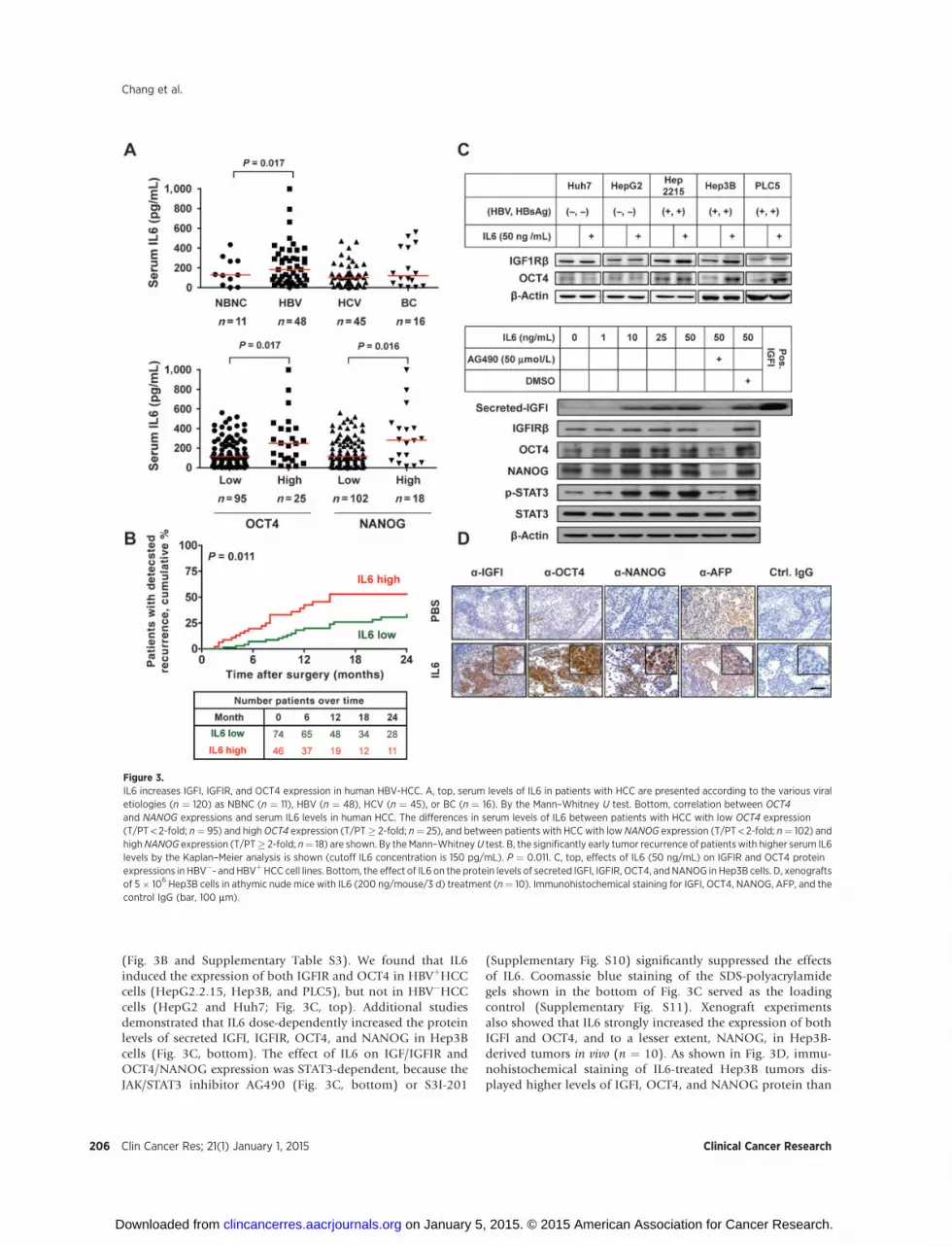
(Fig. 3B and Supplementary Table S3). We found that IL6induced the expression of both IGFIR and OCT4 in HBVþHCCcells (HepG2.2.15, Hep3B, and PLC5), but not in HBV�HCCcells (HepG2 and Huh7; Fig. 3C, top). Additional studiesdemonstrated that IL6 dose-dependently increased the proteinlevels of secreted IGFI, IGFIR, OCT4, and NANOG in Hep3Bcells (Fig. 3C, bottom). The effect of IL6 on IGF/IGFIR andOCT4/NANOG expression was STAT3-dependent, because theJAK/STAT3 inhibitor AG490 (Fig. 3C, bottom) or S3I-201
(Supplementary Fig. S10) significantly suppressed the effectsof IL6. Coomassie blue staining of the SDS-polyacrylamidegels shown in the bottom of Fig. 3C served as the loadingcontrol (Supplementary Fig. S11). Xenograft experimentsalso showed that IL6 strongly increased the expression of bothIGFI and OCT4, and to a lesser extent, NANOG, in Hep3B-derived tumors in vivo (n ¼ 10). As shown in Fig. 3D, immu-nohistochemical staining of IL6-treated Hep3B tumors dis-played higher levels of IGFI, OCT4, and NANOG protein than
Figure 3.IL6 increases IGFI, IGFIR, and OCT4 expression in human HBV-HCC. A, top, serum levels of IL6 in patients with HCC are presented according to the various viraletiologies (n ¼ 120) as NBNC (n ¼ 11), HBV (n ¼ 48), HCV (n ¼ 45), or BC (n ¼ 16). By the Mann–Whitney U test. Bottom, correlation between OCT4and NANOG expressions and serum IL6 levels in human HCC. The differences in serum levels of IL6 between patients with HCC with low OCT4 expression(T/PT < 2-fold; n¼ 95) and highOCT4 expression (T/PT� 2-fold; n¼ 25), and between patients with HCC with low NANOG expression (T/PT < 2-fold; n¼ 102) andhighNANOG expression (T/PT� 2-fold; n¼ 18) are shown. By the Mann–WhitneyU test. B, the significantly early tumor recurrence of patientswith higher serum IL6levels by the Kaplan–Meier analysis is shown (cutoff IL6 concentration is 150 pg/mL). P ¼ 0.011. C, top, effects of IL6 (50 ng/mL) on IGFIR and OCT4 proteinexpressions in HBV�- and HBVþHCC cell lines. Bottom, the effect of IL6 on the protein levels of secreted IGFI, IGFIR, OCT4, andNANOG in Hep3B cells. D, xenograftsof 5� 106 Hep3B cells in athymic nude mice with IL6 (200 ng/mouse/3 d) treatment (n¼ 10). Immunohistochemical staining for IGFI, OCT4, NANOG, AFP, and thecontrol IgG (bar, 100 mm).
Chang et al.
Clin Cancer Res; 21(1) January 1, 2015 Clinical Cancer Research206
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

PBS-treated tissue samples. These results demonstrate that IL6activates expression of IGFI, OCT4, and/or NANOG in HBV-HCC cells both in vitro and in vivo.
IL6 increases stemness-related properties through IGFIRactivation in HBV-related human HCC cells
To examine whether IL6 regulates the stemness properties ofHBV-HCC cells through the activation of IGFIR, RNA interferenceof endogenous IGFIR expression was performed. We found thatthe shIGFIRs suppressed the IL6-induced expression of OCT4 andNANOG in HBVþ Hep3B (Fig. 4A) and HepG2.2.15 cells (Sup-plementary Fig. S13A). IL6 treatment also increased the numberof secondary tumor spheres (Fig. 4B, top and Supplementary Figs.S12 and S13B) and side population cells (Fig. 4B, bottom and
Supplementary Fig. S14) of HBV-HCC cells; this effect of IL6 wassuppressed by the addition of either PPP or shIGFIR. The xeno-graft experiments (Figs. 4C and Supplementary Fig. S15) provideadditional support that the downstream effect of IL6 inducesstemness via activation of IGFI/IGFIR signaling.
In the xenograft experiments using 1 � 106 Hep3B cells forimplantation, more tumors formed in IL6-treated mice than inPBS-treated mice (Supplementary Table S4). The risk of tumorformation significantly increased when �5 �106 cells wereimplanted (Supplementary Table S5; HR, 5.53; P < 0.001). Inaddition, immunohistochemical analysis showed strong OCT4and NANOG expression in IL6-treated tumor cells (Fig. 4C; IL6panel). Importantly, PPP markedly suppressed the IL6 effect ontumor growth (Supplementary Fig. S15; IL6þPPP vs. IL6) as well
Figure 4.IL6 increases stemness properties through IGFIR activation in human HBV-HCC cells. A, OCT4 and NANOG protein levels and the relative luciferase activityof the OCT4/NANOG promoter-luciferase shCtrl.- or shIGFIR Hep3B cells with IL6 treatment (50 ng/mL). Student t test. B, top, the effect of IL6 and IGFIR on theformation of secondary spheres of Hep3B cells. Bottom, the percentage of Hep3B cells with the side population phenotype cultivated in medium only (Mock),IL6 (50 ng/mL), IL6 þ shCtrl. or IL6 þ shIGFIR#2. Student t test. C, immunohistochemical analysis of OCT4/NANOG expression following PBS, IL6, and IL6plus PPP treatment (bar, 100mm). D, serum levels of IL6 in patientswithHCC stratifiedby expression level of all three genes (low¼ T/PT< 1X, n¼ 51; high¼T/PT� 1X,n ¼ 23). By the Mann–Whitney U test. E, model of how IL6 increases the expression of pluripotent gene OCT4 and NANOG through IGFIR activation in humanHBV-HCC cells. � , #, P < 0.05; �� , P < 0.01; and ��� , P < 0.001.
IL6/IGFIR in Stemness Expression and HCC Early Recurrence
www.aacrjournals.org Clin Cancer Res; 21(1) January 1, 2015 207
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

as OCT4/NANOG expression in Hep3B-derived tumors (Fig. 4C;panel of IL6þPPP vs. IL6). The strong correlation between IL6 andIGFIR/OCT4/NANOG was further supported by clinical observa-tion. When compared with patients exhibiting low expression ofall three genes, patients exhibiting high expression of the threegenes had significantly higher serum levels of IL6 (Fig. 4D). Theseresults suggest that IL6 increases pluripotency-related gene expres-sion, particularly for OCT4, in HBV-HCC cells through the acti-vation of IGFI/IGFIR signaling pathway.
DiscussionThe expression of pluripotency-related genes in tumors, such as
OCT4 and NANOG, has been linked to poor prognosis and drugresistance (19, 20). However, the mechanisms regulating cancerstemness expression and tumor recurrence in HCC remainunclear. There is a definite causal relationship between inflam-mation and HCC, and the upregulation of certain inflammatorycytokines induced by HBV or HCV may contribute to hepatocar-cinogenesis (21). However, the pathogenesis of HCC may varyamong different etiologies. For example, a previous reportshowed that HCV infection can activate WNT signaling (13). Thecurrent work demonstrates that IL6 induces the IGFIR-mediatedupregulation of OCT4 and NANOG expression, which is associ-ated with early tumor recurrence in patients with HBV-HCC. Thisheterogeneity in the molecular pathogenesis and signaling path-ways may contribute to the suboptimal efficacy of current ther-apies for HCC (22, 23). A better understanding of the differentetiologies and their diversemechanisms that drive tumorigenicityand/or tumor recurrence is crucial for the optimization of indi-vidualized therapy for HCC.
The mechanism for HBV-induced expression of pluripotency-related genes in cancer is still to be determined. Studies using a cellmodel of hepatoblastoma suggest that it occurs through activa-tion of the HBx protein (24), while other studies suggest it occursvia hepatic progenitor cell activation (8, 25). How HBV inducespluripotency gene expression in differentiated HBV-infectedhepatocytes still remains largely unknown. The clinical observa-tion shows that patients with HCV-HCC and BC-HCC presentlower expression levels of OCT4, NANOG, and IGFIR comparedwith patients with HBV-HCC (Fig. 1A). As HBV is frequentlysuppressed by HCV in dual HBV and HCV hepatitis (26), the factthat patients with BC-HCC expressed lower OCT4, NANOG, andIGFIR than did patients with HBV-HCC indirectly supports a rolefor HBV in OCT4, NANOG, and IGFIR expression, and empha-sizes the etiologic differences that contribute to the prognosis ofpatients with HCC (12).
Niche environment might play a critical role in cell fate deter-mination. Our current findings support an upstream role of HBV-induced inflammation in the development of stemness-relatedproperties in HBV-HCC through IL6-induced IGFI/IGFIR expres-sion. The direct transactivation of IGFIR by IL6 was recentlyreported in prostate cancer (27). However, unlike prostate tumor-igenesis, we found that IL6 did not directly transactivate thephosphorylation of IGFIR in Hep3B cells in either our IGFIRphosphorylation experiments (Supplementary Fig. S16) or inpull-down assays (data not shown). In contrast, our resultsshowed that IL6 activates IGFIR through STAT3-dependentincreases in IGFI and IGFIR expression, and that IGFIR activationmediates the OCT4/NANOG expression in Hep3B cells. In sup-port, studies of human embryonic stem cells and pluripotent
mouse germ line stem cells demonstrated the role of IGFIRsignaling in maintaining stemness (28–30). However, in contrastto the previous report that showed that NANOG-positive HCCcells may be IGFIR-mediated (31), we found that p-IGFIR levelswere significantly correlatedwithOCT4 levels inHBV-HCC tumortissues (Supplementary Fig. S17A); but, were even more signifi-cantly correlated with NANOG levels in HCV-HCC tumor tissues(Supplementary Fig. S17B). This result was further supported byour experiments using a xenograft tumor model that demonstrat-ed the role of IGFIR activation as a dominant signal for IL6-induced stemness, particularly for OCT4, in HBV-HCC tumors(Fig. 4C). These results highlight the association of IGFIR activa-tion with the expression of the different pluripotent transcriptionfactors OCT4 and NANOG in etiologically different cancers.
Despite the large CD133-positive population in Hep3B cells,the tumor-formation efficiency was relatively low inmice that didnot receive treatment of IL6 (Supplementary Table S4) or IGFI(data not shown). This result suggests a role for IL6 in stimulatingHCC cell proliferation during the early stages of HCC. Further-more, as IL6 is the principle cytokine in liver inflammation, theeffects of IL6 may explain the lower incidence of HCC in patientswith inactive carrier status compared with those with chronicactive HBV hepatitis (3). When a larger number (1 � 107) ofHep3B cells were implanted, the IL6 treatment increased cellproliferation only slightly while the expression ofOCT4 increasedsignificantly (Fig. 4C and Supplementary Fig. S15). These findingssuggest that the IL6-induced pluripotency-related gene expressionmay have less effect on tumor proliferation with increasing tumorburden in late-stage HCC. The downstream IGFIR activation inIL6-induced OCT4/NANOG expression in HBV-HCC cells wasalso examined in the xenograft model using a low-dose PPP (1mg/mL) treatment (16) to prevent cell apoptosis in the IL6-induced tumors. The low-dose PPP treatment not only reducedtumor size, but alsomarkedly suppressed the IL6-induced expres-sion ofOCT4/NANOG in the tumors (Fig. 4C and SupplementaryFig. S15). These findings support the fact that activation of IGF/IGFIR mediated the downstream signaling in IL6-induced OCT4/NANOG expression in vivo.
The mechanisms regulating HCC recurrence are still largelyunknown. The patients with HBV-HCC in our study displayedhigher expression levels of pluripotency-related genes, especiallyof OCT4 and IGFIR, and tended to have early tumor recurrence(Fig. 1B) and worse OS (Supplementary Fig. S4) following hep-atectomy. The significant impact of OCT4 and/or NANOG over-expression on tumor recurrence suggests that upregulated OCT4/NANOG confers the ability to micrometastasize. This associationbetween pluripotency-related gene expression and micrometas-tasis of HBV-HCC is supported by the observation that NANOG-positive HCC cells expressed higher metastatic ability (31).
Although the disappointing preliminary results from clinicaltrials using IGFIR-specific monoclonal antibodies have raiseddoubts regarding the future of IGFIR inhibition in various cancers(32–34), several alternate approaches deserve attention. Theblockage of IGFIR in HCC was shown to induce adoptiveresponses, such as upregulation of the epidermal growth factorreceptor, thus emphasizing the rationale for cotargeting morethan one signaling (35). Another approach is to facilitate HCCpersonalized therapy for treating IGFIR by incorporating bio-marker data that have been under intense investigation (36).Although no optimal biomarkers have been identified for HCCthus far, our results suggested that IGFIR signaling inhibition
Chang et al.
Clin Cancer Res; 21(1) January 1, 2015 Clinical Cancer Research208
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

mightworkmore effectively in the subset ofHCCs associatedwithHBV infection.
In summary, HCC has a high rate of recurrence followingtreatment. Effective strategies for the prevention of HCC recur-rence are needed to improve the prognosis of patients with HCC(37). In this study, we demonstrated that the inflammatorycytokine IL6 stimulates the autocrine IGF loop to activate IGFIR,and initiates OCT4 (and/or NANOG) expression in HBV-HCCcells (Fig. 4E). The IL6-induced expression of OCT4 and/orNANOG is primarily limited to patients with HBV-HCC, and isassociated with early tumor recurrence. Our findings suggest thattargeting both the IL6 and IGFIR signaling pathways would be thefuture strategies to improve individualized adjuvant therapy andprevent early tumor recurrence for patients with HBV-HCC.
Disclosure of Potential Conflict of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: T.-S. Chang, Y.-C. Wu, T.-Y. Ling, Y.-H. HuangDevelopment of methodology: T.-S. Chang, Y.-C. Wu, T.-Y. Ling, Y.-H. HuangAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): T.-S. Chang, Y.-C. Wu, S.-Y. Tung, L.-M. Kuo,Y.-H. HuangAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): Y.-C. Wu, C.-C. Chi, K.-F. Lee, T.-H. Tung, J. Wang,S.-Y. Tung, H.-N. Ho, Y.-H. HuangWriting, review, and/or revision of the manuscript: T.-S. Chang, C.-C. Chi,T.-Y. Ling, Y.-H. Huang
Administrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): T.-S. Chang, W.-C. Su, P.-J. Chang, K.-F. Lee,S.-Y. Tung, H.-N. Ho, T.-Y. Ling, Y.-H. HuangStudy supervision: T.-S. Chang, T.-Y. Ling, Y.-H. HuangOther (discussion and suggestion of study goal): J.-J. Liu
AcknowledgmentsThe authors thank Profs. Muh-Hwa Yang and Jaw-ChingWu (National Yang
Ming University, Taipei, Taiwan) and Chia-Ning Shen (Genomics ResearchCenter, Academia Sinica, Taipei, Taiwan) for their critical review of this article.The authors also thank Hsu-Liang Chiang, Ya-Ting Yang, and Jui-Hung Changfor the excellent technical support;Drs. Ching-ChuanHsieh andChia-SiuWang,and Tissue Bank (Chang Gung Memorial Hospital, Chiayi, Taiwan) for pro-viding the HCC tissues; and the Cell-Bio Biotechnology Co., Ltd. (Taipei,Taiwan) for the qualification of immunohistochemical staining.
Grant SupportThis work was supported by research grants from the National Science
Council, Taiwan (NSC99-2628-B-038-009-MY3, NSC(100-102)-2321-B-038-003, NSC102-2628-B-038-008-MY3, and NSC101-2314-B-182A-028), Minis-try of Science and Technology, Taiwan (MOST 103-2321-B-038-011), HealthandWelfare Surcharge of Tobacco Products (MOHW103-TD-B-111-01), ChangGung Memorial Hospital (CMRPG680311, CMRPG690411, CMRPG6A0471,and CMRPG6B0381), Taipei Medical University (TMUTOP103002-9).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received December 2, 2013; revised September 3, 2014; accepted September17, 2014; published online January 6, 2015.
References1. Ferenci P, Fried M, Labrecque D, Bruix J, Sherman M, Omata M, et al.
Hepatocellular carcinoma (HCC): a global perspective. J Clin Gastroen-terol 2010;44:239–45.
2. Yang XR, Xu Y, Yu B, Zhou J, Qiu SJ, Shi GM, et al. High expression levels ofputative hepatic stem/progenitor cell biomarkers related to tumour angio-genesis and poor prognosis of hepatocellular carcinoma. Gut 2010;59:953–62.
3. Berasain C, Castillo J, Perugorria MJ, Latasa MU, Prieto J, Avila MA.Inflammation and liver cancer: new molecular links. Ann NY Acad Sci2009;1155:206–21.
4. Nakamoto Y, Guidotti LG, Kuhlen CV, Fowler P, Chisari FV. Immunepathogenesis of hepatocellular carcinoma. J Exp Med 1998;188:341–50.
5. H€osel M, Quasdorff M, Wiegmann K, Webb D, Zedler U, Broxtermann M,et al. Not interferon, but interleukin-6 controls early gene expression inhepatitis B virus infection. Hepatology 2009;50:1773–82.
6. XiangWQ, FengWF, KeW, SunZ, ChenZ, LiuW.Hepatitis B virus Xproteinstimulates IL-6 expression in hepatocytes via a MyD88-dependent path-way. J Hepatol 2011;54:26–33.
7. Wong VW, Yu J, Cheng AS, Wong GL, Chan HY, Chu ES, et al. High seruminterleukin-6 level predicts future hepatocellular carcinoma developmentin patients with chronic hepatitis B. Int J Cancer 2009;124:2766–70.
8. Wang C, Yang W, Yan HX, Luo T, Zhang J, Tang L, et al. HBx inducestumorigenicity of hepatic progenitor cells in 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) treated HBx transgenic mice. Hepatology 2012;55:108–20.
9. Tang Y, Kitisin K, JogunooriW, Li C,DengCX,Mueller SC, et al. Progenitor/stem cells give rise to liver cancer due to aberrant TGF-beta and IL-6signaling. Proc Natl Acad Sci U S A 2008;105:2445–50.
10. Lee TK, Castilho A, Cheung VC, Tang KH,Ma S, Ng IO. CD24þ liver tumor-initiating cells drive self-renewal and tumor initiation through STAT3-mediated NANOG regulation. Cell Stem Cell 2011;9:50–63.
11. Park BC, Huh MH, Seo JH. Differential expression of transforming growthfactor alpha and insulin-like growth factor II in chronic active hepatitis B,cirrhosis and hepatocellular carcinoma. J Hepatol 1995;22:286–94.
12. Iizuka N, Oka M, Yamada-Okabe H, Mori N, Tamesa T, Okada T, et al.Comparison of gene expression profiles between hepatitis B virus- andhepatitis C virus-infected hepatocellular carcinoma by oligonucleotidemicroarray data on the basis of a supervised learning method. Cancer Res2002;62:3939–44.
13. Boyault S, RickmanDS, deReyni�esA, BalabaudC,Rebouissou S, Jeannot E ,et al. Transcriptome classification of HCC is related to gene alterations andto new therapeutic targets. Hepatology 2007;45:42–52.
14. Kim SO, Park JG, Lee YI. Increased expression of the insulin-like growthfactor I (IGF-I) receptor gene in hepatocellular carcinoma cell lines:implications of IGF-I receptor gene activation by hepatitis B virus X geneproduct. Cancer Res 1996;56:3831–6.
15. Qian J, Yao D, Dong Z, WuW, Qiu L, Yao N, et al. Characteristics of hepaticIGF-II expression and monitored levels of circulating IGF-II mRNA inmetastasisofhepatocellular carcinoma.AmJClinPathol2010;134:799–806.
16. Nussbaum T, Samarin J, Ehemann V, Bissinger M, Ryschich E, Khamidja-nov A, et al. Autocrine insulin-like growth factor-II stimulation of tumorcell migration is a progression step in human hepatocarcinogenesis.Hepatology 2008;48:146–56.
17. Tovar V, Alsinet C, Villanueva A, Hoshida Y, Chiang DY, Sol�e M, et al. IGFactivation in a molecular subclass of hepatocellular carcinoma and pre-clinical efficacy of IGF-IR blockage. J Hepatol 2010;52:550–9.
18. McShane LM, Altman DG, Sauerbrei W, Taube SE, Gion M, Clark GM.Statistics subcommittee of the NCI-EORTC working group on cancerdiagnostics. Reporting recommendations for tumor marker prognosticstudies. J Clin Oncol 2005;23:9067–72.
19. Wang XQ,OngkekoWM, Chen L, Yang ZF, Lu P, Chen KK , et al. Octamer 4(Oct4) mediates chemotherapeutic drug resistance in liver cancer cellsthrough a potential Oct4 AKT–ATP-binding cassette G2 pathway. Hepa-tology 2010;52:528–39.
20. Wu YC, Ling TY, Lu SH, Kuo HC, Ho HN, Yeh SD, et al. Chemotherapeuticsensitivity of testicular germ cell tumors under hypoxic conditions isnegatively regulated by SENP1-controlled sumoylation of OCT4. CancerRes 2012;72:4963–73.
IL6/IGFIR in Stemness Expression and HCC Early Recurrence
www.aacrjournals.org Clin Cancer Res; 21(1) January 1, 2015 209
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

21. Haybaeck J, Zeller N, Wolf MJ, Weber A, Wagner U, Kurrer MO, et al. Alymphotoxin-driven pathway to hepatocellular carcinoma. Cancer Cell2009;16:295–308.
22. Fransvea E, Paradiso A, Antonaci S, Giannelli G. HCC heterogeneity:molecular pathogenesis and clinical implications. Cell Oncol 2009;31:227–33.
23. Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, et al.Sorafenib in advanced hepatocellular carcinoma. N Engl J Med 2008;359:378–90.
24. Arzumanyan A, Friedman T, Ng IO, Clayton MM, Lian Z, Feitelson MA.Does the hepatitis B antigen HBx promote the appearance of liver cancerstem cells? Cancer Res 2011;71:3701–8.
25. Hsia CC, Thorgeirsson SS, Tabor E. Expression of hepatitis B surface andcore antigens and transforming growth factor-alpha in "oval cells" of theliver in patients with hepatocellular carcinoma. J Med Virol 1994;43:216–21.
26. Sheen IS, Liaw YF, Chu CM, Pao CC. Role of hepatitis C virus infection inspontaneous hepatitis B surface antigen clearance during chronic hepatitisB virus infection. J Infect Dis 1992;165:831–4.
27. Rojas A, LiuG,Coleman I,NelsonPS, ZhangM,DashR, et al. IL-6 promotesprostate tumorigenesis and progression through autocrine cross-activationof IGF-IR. Oncogene 2011;30:2345–55
28. Bendall SC, Stewart MH, Menendez P, George D, Vijayaragavan K, Wer-bowetski-Ogilvie T, et al. IGF and FGF cooperatively establish the regula-tory stem cell niche of pluripotent human cells in vitro. Nature 2007;448:1015–21.
29. Wang L, Schulz TC, Sherrer ES, Dauphin DS, Shin S, Nelson AM, et al. Self-renewal of human embryonic stem cells requires insulin-like growth factor-1 receptor and ERBB2 receptor signaling. Blood 2007;110:4111–9.
30. Huang YH, Chin CC, Ho HN, Chou CK, Shen CN, Kuo HC, et al.Pluripotency of mouse spermatogonial stem cells maintained by IGF-I-dependent pathway. FASEB J 2009;23:2076–87.
31. Shan J, Shen J, Liu L, Xia F, XuC,DuanG, et al.Nanog regulates self-renewalof cancer stem cell through IGF pathway in human hepatocellular carci-noma. Hepatology 2012;56:1004–14.
32. Pollak M. The insulin and insulin-like growth factor receptor family inneoplasia: an update. Nat Rev Cancer 2012;12:159–69.
33. AllisonM. Clinical setbacks reduce IGF-1 inhibitors to cocktail mixers. NatBiotechnol 2012;30:906–7.
34. Baserga R. The decline and fall of the IGF-I receptor. J Cell Physiol2013;228:675–9.
35. Desbois-Mouthon C, Baron A, Eggelpo€el MB, Fartoux L, Venot C, Bladt F,et al. Insulin-like growth factor-1 receptor inhibition induces a resistancemechanism via the epidermal growth factor receptor/HER3/AKT signalingpathway: rational basis for cotargeting insulin-like growth factor-1 receptorand epidermal growth factor receptor in hepatocellular carcinoma. ClinCancer Res 2009;15:5445–56.
36. Singhal A, JayaramanM,DhanasekaranDN, Kohli V.Molecular and serummarkers in hepatocellular carcinoma: predictive tools for prognosis andrecurrence. Crit Rev Oncol Hemat 2012;82:116–40.
37. Kudo M. Adjuvant therapy after curative treatment for hepatocellularcarcinoma. Oncology 2011;81(Suppl 1):50–5.
Clin Cancer Res; 21(1) January 1, 2015 Clinical Cancer Research210
Chang et al.
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

2015;21:201-210. Clin Cancer Res Te-Sheng Chang, Yu-Chih Wu, Ching-Chi Chi, et al. ExpressionHepatocellular Carcinoma through Induction of OCT4/NANOG Activation of IL6/IGFIR Confers Poor Prognosis of HBV-Related
Updated version
http://clincancerres.aacrjournals.org/content/21/1/201
Access the most recent version of this article at:
Cited Articles
http://clincancerres.aacrjournals.org/content/21/1/201.full.html#ref-list-1
This article cites by 37 articles, 12 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications Department at
on January 5, 2015. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from





























