Embed Size (px)
Citation preview

Arch Dermatol Res (2008) 300:495–504
DOI 10.1007/s00403-008-0836-3ORIGINAL PAPER
Acute stretch promotes endothelial cell proliferation in wounded healing mouse skin
Carl D. Shrader · Holly G. Ressetar · Jia Luo · Eugene V. Cilento · Frank D. Reilly
Received: 21 May 2007 / Revised: 4 January 2008 / Accepted: 28 January 2008 / Published online: 11 March 2008© Springer-Verlag 2008
Abstract We have developed a novel in vivo model utiliz-ing acute stretch to investigate endothelial cell proliferationas a marker of vascular growth in healing mouse skin. Thisstudy is a follow-up to ones revealing immediate stretchimproves blood Xow, decreases total tissue necrosis, andinduces tissue insulin transcription. Dorsal distally basedXaps of skin were stretched for 3 min using linear (skinhook) plus hemispherical load cycling (inXated subcutane-ous silicone catheter). Unstretched, wounded skin along theback and sternum served as postoperative controls. LaserDoppler Xowmetry demonstrated a threefold increase in Xapperfusion at postoperative day 7. A stretch-induced sixfoldincrease in endothelial cell mitogenesis accompaniedenhancements in blood Xow and extracorporal wound heal-ing over the sternum. Western blots revealed up-regulation/activation of insulin and mitogenic signaling intermediatesin stretched skin. Activated insulin and insulin growth factorreceptors (pIR/pIGFR), protein kinase B (Akt, pAkt),
vascular endothelial growth factor (VEGF) and vascularendothelial growth factor receptor 2 (Xk-1) were among theidentiWed stretch-responsive intermediates. These resultsindicate the beneWts of acute stretch are mediated throughenhanced vascularity as evidenced by endothelial cell mito-genesis and up-regulation/activation of insulin and keyangiogenic eVectors in dorsal distally based skin Xaps.
Keywords Acute stretch · Skin · Endothelium · Proliferation · Insulin
Introduction
We have reported long-term beneWts and improved skinXap viability arising from the application of rapid, acutestretch in a SKH1 mouse model [43]. These beneWts weredemonstrated with Doppler Xowmetry and by measuringnecrosis along the margin of a surgically created dorsal dis-tally based skin Xap. Increases up to 50% in nutritive bloodXow and decreases of 75% in tissue necrosis were observedat post-operative day 5. Despite numerous studies by othersattesting to the beneWts of tissue stretch, no thorough inves-tigation into the underlying cell type(s) and molecularmechanism(s) that promote viability and improved healinghave been reported [29, 35].
The microvasculature bears the enormous task of main-taining systemic homeostasis following tissue wounding. Itmust initiate clot formation, chemokine signaling, cellularrecruitment, vasoreactivity, and immune response whilemaintaining normal functional and/or nutritive blood Xow[3, 7, 23]. These processes are critical to survival and heal-ing, and occur in three phases: an inXammatory phase, aproliferative phase, and a maturational phase. The endothe-lial cell serves a pivotal role in each phase.
C. D. Shrader · H. G. Ressetar · E. V. Cilento · F. D. Reilly (&)Department of Neurobiology and Anatomy, School of Medicine, West Virginia University, Morgantown, WV 26506-9128, USAe-mail: [email protected]
C. D. Shradere-mail: [email protected]
J. LuoDepartment of Microbiology, Immunology and Cell Biology, School of Medicine, West Virginia University, Morgantown, WV 26506-9177, USA
E. V. CilentoDepartment of Chemical Engineering, College of Engineering and Mineral Resources, West Virginia University, Morgantown, WV 26506-6070, USA
123

496 Arch Dermatol Res (2008) 300:495–504
Despite many advances in understanding the science ofendothelium, there is a gap in knowledge of mitogenicstimuli, i.e., stretch and molecular mechanisms that triggerendothelial cell proliferation. Mitogenic activity of endo-thelium promotes new vascular growth as seen from exist-ing capillary beds during angiogenesis. Angiogenesis isinvolved in a variety of physiological and pathologicalroles, and has become a central focus for many investiga-tors [4, 8, 14, 21, 39]. Angiogenesis is imperative to theproliferative phase of healing as it sustains the process.Chronic unhealed wounds result if angiogenesis becomesimpaired [19, 20, 36].
Characterization of the cellular and molecular mecha-nisms underlying improved viability and healing in stretchedtissue carries enormous clinical signiWcance, warranting fur-ther investigation. Using diVerential display reverse tran-scription polymerase chain reaction (DDRT-PCR) andNorthern blot analysis, we reported stretch-induced expres-sion of tissue insulin mRNA [44]. Expression of insulinmRNA was coincident with improved healing as increasedblood Xow and decreased necrosis accompanied up-regula-tion. Despite the traditional view of insulin as a metabolicregulatory hormone, recent evidence suggests insulin canserve as a key mitogenic regulator [2, 9, 22, 34, 40, 41].
In the current study, we demonstrate stretch-inducedgene-speciWc tissue insulin expression and enhanced up-regulation/activation of multiple angiogenic signaling inter-mediates using Western blot analysis. Immunohistochemis-try revealed enhanced mitogenesis in endothelium ofstretched tissue and conWrmed activation of insulin receptorand insulin growth factor receptor (pIR/pIGFR). These dataincrease our understanding of the beneWcial eVects of acutestretch on skin Xap viability and healing in local and remote(extracorporal) sites.
Materials and methods
Animal care and anesthesia
A total of 70 outbred Crl: SKH1-hrBR albino (hairless)male mice (20–30 g) of 20–42 days of age were purchasedfrom Charles River Laboratories (Wilmington, MA). Theywere kept in a light and temperature regulated environmentand were permitted free access to food and water whilemaintained on Tek fresh bedding. The mice were anesthe-tized intraperitoneally with Nembutal (0.05 mg/g bodyweight) and 20 min later with Ketamine (0.03 mg/g). Theinstitutional Animal Care and Use Committee approved theproject and key personnel were trained in Core AnimalWelfare (protocol #: 03-0401). NIH Publication 85-23(revised in 1985) “Principles of laboratory animal care”was followed. The animal care facilities and program met
the requirements of the Laboratory Animal Welfare Act P2-89-541, 91-579 and 94-279, NIH policies and ONM Circu-lar A-21, the Public Health Service Act, and the AmericanVeterinary Medical Association on euthanasia.
Treatment groups and surgical protocol
The experimental design incorporated two groups of four tosix anesthetized mice. A dorsally based random Xap mea-suring 30 mm in width by 35 mm in length was delineatedon the animals using established methods [11–13, 43]. SkinXaps from the control group were undermined, following acranial apex incision, without the application of acutestretch. Flaps from the experimental group were under-mined and then a constant 5 g of linear tension plus a sub-cutaneous 3 min hemispherical load cycle (acute stretch)applied using a Foley catheter (REF 0165L 12, C.R. Bard,Inc., Covington, KY). Constant initial linear tension wasaccomplished by securing anesthetized mice with surgicaltape to a custom built Plexiglas apparatus housing a tensi-ometer (Chatillon Tensiometer DGGRS-gram gauge,Greensboro, NC) so that, movements of this instrument inrelation to the Xap were precisely controlled. The Xap wassecured to the tensiometer with a suture hook. Experimentalanimals, similar to control animals, received an apical inci-sion allowing for undermining and stretching of the Xap.The skin hook was attached and an initial 5 g of linear ten-sion applied. The hemispherical load cycle was achieved byinserting the inXation tube of the catheter under the Xapthrough the cranial incision. The catheter was inXated with6 ml of sterile saline.
Both stretched (experimental) and unstretched (control)Xaps were raised by cutting the two lateral sides. They werethen re-approximated to the original donor site and closedwith 5–0 nylon simple sutures. A total of 15 sutures, dis-tributed approximately 5 mm, were used for closure.
Flaps were excised and hemi-sected at multiple post-operative time points. One half was stored at ¡80°C andthe other half was placed in 10% neutral buVered formalinprior to paraYn embedding. After Xap excision, mice wereeuthanized with an excess of pentobarbital.
Another four to six anesthetized mice in the stretchedand unstretched groups had an extracorporeal skin incisionof 15 mm in length applied along the sternum. Four sutures,3 mm apart were used to close these surgically createdwounds. Total wound length was measured using amicrometer between 6 h and postoperative day 7.
Laser Doppler Xowmetry
Blood Xow measurements were repeated three times (tripli-cate) in each animal using a laser Doppler Xowmeter ModelALF 21 (Transonic Systems, Ithaca, NY). Means of triplicate
123

Arch Dermatol Res (2008) 300:495–504 497
measurements § SEM were graphed. The measurementsite was 10 mm from the center of the caudal Xap borders,and its exact position was determined from previous studies[13, 43]. Total Xap perfusion was estimated by recordingblood Xow before surgery (baseline) and then again at post-operative time points.
Immunohistochemistry
Skin Xaps were collected, Wxed in 10% neutral buVered for-malin and paraYn-embedded. The samples were Wxed1 day and processed the following day to preserve the anti-genicity and morphology of the tissue. Five micrometersections were cut and mounted on poly-lysine coated slidesand air-dried overnight at room temperature. Double-label-ing of factor VIII (demonstrated with alkaline phospha-tase—fast red chromagen) with pIR/pIGFR or proliferatingcellular nuclear antigen (PCNA) (both demonstrated withDAB—brown chromagen) in vascular endothelial cells wasassessed using established immunohistochemical methods[32, 33].
The basic immunostaining protocol was as follows: Xapswere processed immunohistochemically for the presence ofspeciWc proteins using an avidin-biotin method [32, 33].The proteins of interest (PCNA; Ventana, Tucson, AZ, pIR/pIGFR; Chemicon International, Temecula, CA) were dou-ble-labeled with factor VIII (Ventana, Tucson, AZ) to iden-tify endothelial cells. Flaps were sectioned (5 �m) with amicrotome. Sections were washed with 3.0% bovine serumalbumin (BSA) and 2% normal goat serum (NGS) in0.10 M phosphate buVer for 10 min each to block non-spe-ciWc staining. Antigen retrieval was performed to removeany cross-linking damage done during Wxation and process-ing. The samples were incubated for 30 min at 37°C withprimary antibodies (PCNA and/or pIR/pIGFR) diluted at1:50 in a solution of 3% BSA in 0.15 M phosphate buVer.Next, samples were washed in 1£ tris buVered saline(TBS) and placed in horse radish peroxidase (HRP) poly-mer for 30 min. Sections were washed in 1£ TBS and DABplaced on them for 5–10 min. Slides were then placed in arunning water wash for 5 min. Double stain blocker wasadded for 10 min, followed by another 5 min rinse in 1£TBS. A second primary antibody was added at this point(factor VIII) for 30 min, rinsed in 1£ TBS for 5 min andplaced in alkaline phosphatase (ALP) for 55 min. Sampleswere placed in 1£ TBS for 5 min and fast red was added toslides for 5–10 min. Triple labeled samples were rinsedagain in 1£ TBS and placed in the third antibody (PCNA),then rinsed and placed in ALP for 55 min. Finally, thesesamples were rinsed in 1£ TBS and placed in 5-bromo-4-chloro-3-indoxyphosphate/nitroblue tetrazolium chloride(BCIP/NBT) (Ventana, Tucson, AZ) substrate for 5 min.Slides were then washed well in a running water rinse for
10 min. Hematoxylin (30–45 s) was used as a counterstainon double labeled sections. Slides were again washed welland coverslipped as an aqueous media. Due to light sensi-tivity, samples were stored in a dark area for up to 1 week.QuantiWed antigens were labeled with DAB and densitome-try performed using Optimas 6.2. Incubation in NGS andomission of primary antibodies served as control reactionsfor non-speciWc binding. The results of these controls werenegative.
Western blot analysis
The quantitative immunoblotting protocol was similar tothat used previously [27]. All analyses were repeated threetimes (triplicate) with new animal groups. Means of tripli-cate measurements § SEM were graphed. Time points pre-ceding those graphed were not signiWcant from baseline,and therefore not displayed in Wgures.
Flaps were placed in liquid nitrogen for 1 min, pulver-ized, homogenized and lysed with ice cold RIPA buVer(150 mM NaCl, 50 mM Tris pH 8.0, 1.0% Polydet N-40,0.10% sodium dodecylsulfate (SDS), 0.50% sodium deoxy-cholic acid, 0.10 mg/ml phenylmethylsulfonyl Xuoride,1.0 mM sodium orthovanadate, and 3.0% aprotinin, Sigma)for 10 min. The homogenates were centrifuged at 10,000 gfor 10 min, and the supernatant containing cellular proteinswas collected and protein concentration determined with aBio-Tek microplate reader. Protein samples (50 �g/lane)were loaded on 10% SDS-polyacrylamide gels (PAGE),separated by electrophoresis and then transferred to nitro-cellulose membranes. Non-speciWc activity was blocked bywashing the membrane with 5.0% nonfat dry milk and0.10% Tween-20 in 0.01 M PBS (TPBS) for 1 h at roomtemperature. The membranes were incubated overnight at4°C with primary antibodies against PCNA, insulin, Akt,VEGF, Xk-1 (Santa Cruz Biotechnology, Santa Cruz, CA),pAkt (S473, Zymed Laboratories, San Francisco, CA) andpIR/pIGFR (Cell Signaling Technology, Danvers, MA).After two washes in TPBS, the membranes were incubatedwith a peroxidase-conjugated secondary antibody (Amer-sham, Piscataway, NJ; 1:2,000 in TPBS) for 1 h. Theimmune complexes were detected by the enhanced chemi-luminescence (ECL) method (Amersham, Piscataway, NJ).As with the immunohistochemical studies, non-speciWcbinding was assessed on immunoblots processed withoutthe primary or secondary antibodies. The results of suchcontrols were consistently negative.
In order to control for inter-sample variation of the totalamount of protein loaded onto the gel, the membranes werestripped of antibodies directed against target proteins andre-probed with anti-tubulin Ab as an internal standard. Den-sitometric readings of target protein contents were normal-ized against those of the internal standard using Optimas
123

498 Arch Dermatol Res (2008) 300:495–504
6.2. The membranes were stripped by incubating in a solu-tion containing 100 mM �-mercaptoethanol, 2.0% SDS,and 62.5 mM Tris–HCl (pH 6.7) for 30 min at 50°C orovernight at 4°C. The membranes were then washed twicewith TPBS for 15 min. Non-speciWc binding was blockedwith either 5.0% milk or 2.5% BSA in PBS, and the mem-brane was re-blotted as described above. With this method,we were able to detect small diVerences in protein expres-sion [28]. The relative amounts of stretch-induced proteinexpression in the Wlms prepared from the immunoblotswere quantiWed using an LKB Ultrascan XL Laser Densi-tometer. For detection of low abundance proteins, such asphosphorylated kinases, the homogenates were concen-trated using the Microcon Centrifugal Filter Device (Milli-pore Inc., Billercia, MA) prior to electrophoresis. We havesuccessfully employed this method to detect low levels ofspeciWc proteins.
Statistical analysis
DiVerences among treatment groups were tested using aone-way analysis of variance (ANOVA) and Tukey’s (posthoc) test. DiVerences with P-values · 0.05 were consid-ered statistically signiWcant. Data are presented in Wgures asmean § error of the mean (SEM).
Results
Doppler Xowmetry demonstrated, that all Xaps were ade-quately perfused. Total estimated perfusion at the Xap basewas signiWcantly higher (P · 0.05) at postoperative day 7(Fig. 1) in acutely stretched wounded skin. This increase wasaccompanied by a signiWcant reduction (P · 0.05) in woundlength along the sternum between post-operative days 1 and5 (Fig. 2). These Wndings were consistence with thosereported previously where marginal tissue necrosis along thecut edge of stretched Xaps with elevated blood Xow was sig-niWcantly decreased at post-operative day 5 [43].
PCNA antigen (brown chromagen) was isolated on fac-tor VIII (red chromagen) positive endothelial cells ofwounded skin (identiWed by arrows in Fig. 3a). QuantiWca-tion of brown chromagen as a percentage (%) of the totalcross-sectional area of a blood vessel’s lining revealed sig-niWcant stretch-enhanced labeling (P · 0.05). This increasein expression was conWrmed by Western blots that werestripped of PCNA antigen and re-blotted with tubulin as aninternal standard (Fig. 3b).
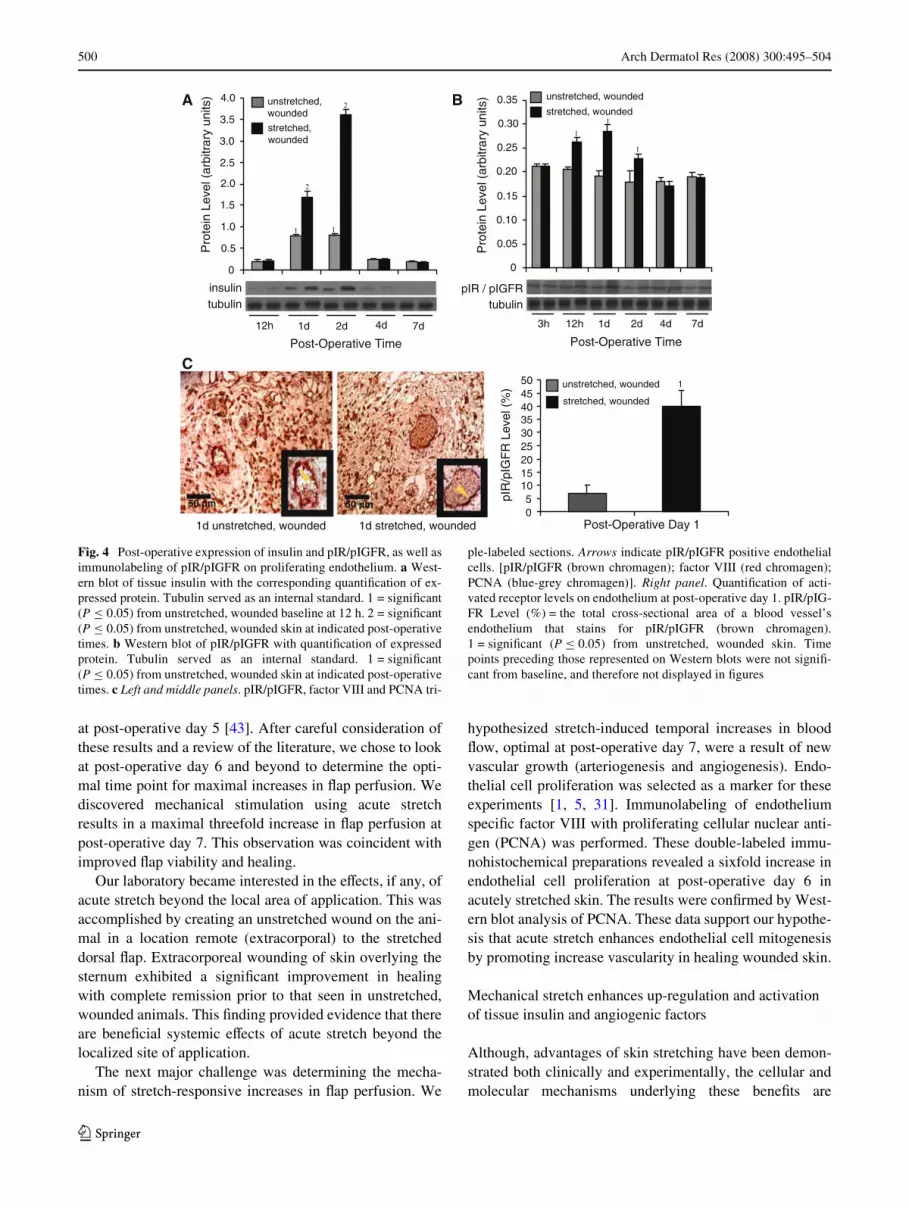
Insulin expression was signiWcantly elevated (P · 0.05)at day 1 and optimal at day 2 (Fig. 4a), while pIR/pIGFRactivity peaked at day 1 (Fig. 4b) in stretched Xaps. Triple-immunolabeling of tissue sections with pIR/pIGFR (brownchromagen), factor VIII (red chromagen) and PCNA
(blue-grey chromagen) conWrmed the presence of pIR/pIGFR on proliferating endothelium (Fig. 4c left and mid-dle panels with arrows indicating pIR/pIGFR positiveendothelial cells). QuantiWcation of brown chromagen as apercentage of the total cross-sectional area of a blood ves-sel’s lining demonstrated signiWcant pIR/pIGFR activation(P · 0.05) on post-operative day 1 (Fig. 4c right panel).
Acute stretching signiWcantly upregulated (P · 0.05)expression of VEGF, VEGF receptor 2 (Xk-1) (Fig. 5), andAkt (Fig. 6). It also activated (pAkt) Akt above the levels
Fig. 1 Temporal changes in total Xap perfusion at post-operative days2, 5, 7, 10 and 15. Estimated total perfusion (%) = blood Xow as a per-centage of that measured at the Xap base prior to surgery.1 = signiWcant (P · 0.05) from unstretched, wounded skin
0
50
100
150
200
250
300
350
2d 5d 7d 10d 15d
unstretched, wounded
stretched,wounded 1
Post-Operative Time
Tot
al F
lap
Per
fusi
on (
%)
Fig. 2 Extracorporeal wound length at post-operative times 6 h, 1, 2,5 and 7 days. 1 = signiWcant (P · 0.05) from unstretched, woundedskin at indicated post-operative times
0
2
4
6
8
10
12
14
16
6h 1d 2d 5d 7d
unstretched, wounded
stretched, wounded
1
1
1
Post-Operative Time
Wou
nd L
engt
h (m
m)
123

Arch Dermatol Res (2008) 300:495–504 499
provoked by wounding alone (Fig. 6). Protein levels forupregulated/activated angiogenic factors correlated posi-tively with the timing of insulin and PCNA expression instretched, wounded Xaps (Fig. 7).
Discussion
One of the major challenges that currently faces micro-vas-cular research is vital knowledge of mechanisms that pro-mote vascular growth. Recent data demonstrate theimportance of mechanical–chemical signaling that is trans-mitted throughout the endothelial cell and through extracel-lular matrix (ECM) adhesions on the surface ofendothelium [6, 10, 15, 16, 37]. Most of these experimentalmodels have elucidated mechanisms of endothelial cellmigration and tubulogenesis as physiologically relevantmodels to study angiogenesis [16, 25].
We have recently developed a novel in vivo approachutilizing acute mechanical stretch to investigate endothelial
cell proliferation as a marker vascular growth in healingmouse skin [1, 5, 31]. This model has the following fea-tures: (1) the modulus of elasticity in the animal approxi-mates the skin of the human face, (2) Xowmetrydemonstrates surgically created Xaps are well perfused, (3)mechanical manipulation (acute stretch) drasticallyimproves Xap viability as assessed by decreases up to 75%in marginal tissue necrosis, (4) wounding alone(unstretched skin) enhances proliferation and protein kinaseB expression, and (5) the dorsal distally based skin Xap is areliable model for studying wound closure.
Mechanical stimulation enhances local Xap perfusion and remote extracorporeal wound healing that is coincident with endothelial cell proliferation
Acute stretching of a surgically created skin Xap in ourSKH1 hairless mouse model improves viability and heal-ing. As previously reported, increases up to 50% in bloodXow and decreases of 75% in tissue necrosis were detected
Fig. 3 Post-operative expression and endothelial immunolabeling ofPCNA. a Left and middle panels. PCNA antigen (brown chromagen)is located on endothelium (factor VIII antigen—red chromagen) ofwounded skin (identiWed by arrows). Right panel. QuantiWcation ofPCNA reveals signiWcant stretch enhanced labeling in endothelium.PCNA Level (%) = the total cross-sectional area of a blood vessel’sendothelium that stains for PCNA (brown chromagen). 1 = signiWcant
(P · 0.05) from unstretched, wounded skin. b Western blot of PCNAwith the corresponding quantiWcation of expressed protein. Tubulinserved as an internal standard. 1 = signiWcant (P · 0.05) from un-stretched, wounded baseline at 12 h. 2 = signiWcant (P · 0.05) fromunstretched, wounded skin at indicated post-operative times. Timepoints preceding those represented on Western blots were not signiW-cant from baseline, and therefore not displayed in Wgures
PCNA
tubulin
7d4d2d
0
0.5
1.0
1.5
2.0
2.5
3.0
22
22
1 11 1
Post-OperativeTime
unstretched, wounded
stretched, wounded
12h 1d
Pro
tein
Lev
el (
arbi
trar
y un
its)
50 µm 0
5
10
15
20
25
30
35
401
Post-Operative Day 6
PC
NA
Lev
el (
%)
6d unstretched, wounded 6d stretched, wounded
stretched, wounded
unstretched, wounded
50 µm
A
B
123
500 Arch Dermatol Res (2008) 300:495–504
at post-operative day 5 [43]. After careful consideration ofthese results and a review of the literature, we chose to lookat post-operative day 6 and beyond to determine the opti-mal time point for maximal increases in Xap perfusion. Wediscovered mechanical stimulation using acute stretchresults in a maximal threefold increase in Xap perfusion atpost-operative day 7. This observation was coincident withimproved Xap viability and healing.
Our laboratory became interested in the eVects, if any, ofacute stretch beyond the local area of application. This wasaccomplished by creating an unstretched wound on the ani-mal in a location remote (extracorporal) to the stretcheddorsal Xap. Extracorporeal wounding of skin overlying thesternum exhibited a signiWcant improvement in healingwith complete remission prior to that seen in unstretched,wounded animals. This Wnding provided evidence that thereare beneWcial systemic eVects of acute stretch beyond thelocalized site of application.
The next major challenge was determining the mecha-nism of stretch-responsive increases in Xap perfusion. We
hypothesized stretch-induced temporal increases in bloodXow, optimal at post-operative day 7, were a result of newvascular growth (arteriogenesis and angiogenesis). Endo-thelial cell proliferation was selected as a marker for theseexperiments [1, 5, 31]. Immunolabeling of endotheliumspeciWc factor VIII with proliferating cellular nuclear anti-gen (PCNA) was performed. These double-labeled immu-nohistochemical preparations revealed a sixfold increase inendothelial cell proliferation at post-operative day 6 inacutely stretched skin. The results were conWrmed by West-ern blot analysis of PCNA. These data support our hypothe-sis that acute stretch enhances endothelial cell mitogenesisby promoting increase vascularity in healing wounded skin.
Mechanical stretch enhances up-regulation and activation of tissue insulin and angiogenic factors
Although, advantages of skin stretching have been demon-strated both clinically and experimentally, the cellular andmolecular mechanisms underlying these beneWts are
Fig. 4 Post-operative expression of insulin and pIR/pIGFR, as well asimmunolabeling of pIR/pIGFR on proliferating endothelium. a West-ern blot of tissue insulin with the corresponding quantiWcation of ex-pressed protein. Tubulin served as an internal standard. 1 = signiWcant(P · 0.05) from unstretched, wounded baseline at 12 h. 2 = signiWcant(P · 0.05) from unstretched, wounded skin at indicated post-operativetimes. b Western blot of pIR/pIGFR with quantiWcation of expressedprotein. Tubulin served as an internal standard. 1 = signiWcant(P · 0.05) from unstretched, wounded skin at indicated post-operativetimes. c Left and middle panels. pIR/pIGFR, factor VIII and PCNA tri-
ple-labeled sections. Arrows indicate pIR/pIGFR positive endothelialcells. [pIR/pIGFR (brown chromagen); factor VIII (red chromagen);PCNA (blue-grey chromagen)]. Right panel. QuantiWcation of acti-vated receptor levels on endothelium at post-operative day 1. pIR/pIG-FR Level (%) = the total cross-sectional area of a blood vessel’sendothelium that stains for pIR/pIGFR (brown chromagen).1 = signiWcant (P · 0.05) from unstretched, wounded skin. Timepoints preceding those represented on Western blots were not signiW-cant from baseline, and therefore not displayed in Wgures
0
0.05
0.10
0.15
0.20
0.25
0.30
0.35
1
1
1
0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
2
2
1 1
insulintubulin
unstretched, wounded
stretched, wounded
unstretched, wounded
stretched, wounded
Pro
tein
Lev
el (
arbi
trar
y un
its)
Pro
tein
Lev
el (
arbi
trar
y un
its)
Post-Operative Time
12h 1d 2d 7d4d
Post-Operative Time
3h 12h 1d 2d 4d 7d
tubulinpIR / pIGFR
1d stretched, wounded
1d unstretched, wounded
pIR
/pIG
FR
Lev
el (
%)
05
101520253035404550 1unstretched, wounded
stretched, wounded
Post-Operative Day 1
50 µm 50 µm
A B
C
123

Arch Dermatol Res (2008) 300:495–504 501
unknown [33, 35, 38]. We have previously reported the up-regulation of tissue insulin mRNA in stretched skin [44]. Itwas unclear if this increased transcription was biologicallysigniWcant. We hypothesized that acute stretch-inducedendothelial cell proliferation was tissue insulin mediatedthrough phosphoinositide-3 kinase (PI3-K) eVectors, i.e.,protein kinase B (Akt) and vascular endothelial growth fac-tor (VEGF).
Using Western blot analysis, we conWrmed the up-regu-lation of tissue insulin in response to acute stretch. Stretchalso promoted activation of insulin receptor (pIR) and insu-
lin growth factor receptor (pIGFR). Despite only a 40%homology in insulin and insulin growth factor, both recep-tor types respond to insulin activation and are associatedwith mitogenic insulin signaling [24, 30]. Our preliminarystudies (unpublished observation) demonstrated, thatstretch does not alter endothelial expression of these recep-tors (P ¸ 0.05). However, stretch did signiWcantly impactthe activation of IR/IGFR. Immunolabeling with factor VIIIand PCNA conWrmed the presence and activation of thesereceptors on proliferating endothelium. The relatively largeincreases in receptor activity in immunolabeled tissue
Fig. 5 Post-operative expression of VEGF and Xk-1. a Western blotof VEGF with the corresponding quantiWcation of expressed protein.Tubulin served as an internal standard. 1 = signiWcant (P · 0.05) fromunstretched, wounded skin at indicated post-operative times. b West-ern blot of Xk-1 with the corresponding quantiWcation of expressed
protein. Tubulin served as an internal standard. 1 = signiWcant(P · 0.05) from unstretched, wounded skin at indicated post-operativetimes. Time points preceding those represented on Western blots werenot signiWcant from baseline, and therefore not displayed in Wgures
VEGF
tubulin
0
0.5
1.0
1.5
2.0
2.5
1
1 1
1
11
1
unstretched, wounded
stretched, woundedunstretched, wounded
stretched, wounded
Pro
tein
Lev
el (
arbi
trar
y un
its)
0
0.5
1.0
1.5
2.0
2.5
Pro
tein
Lev
el (
arbi
trar
y un
its)
Post-Operative Time 12h 1d 2d 4d 7d
Post-Operative Time 12h 1d 2d 4d 7d
tubulin
flk-1
A B
Fig. 6 Post-operative expression of Akt and pAkt. a Western blot ofAkt with the corresponding quantiWcation of expressed protein. Tubu-lin served as an internal standard. 1 = signiWcant (P · 0.05) from un-stretched, wounded skin at indicated post-operative times. b Westernblot of pAkt with the corresponding quantiWcation of expressed pro-
tein. Tubulin served as an internal standard. 1 = signiWcant (P · 0.05)from unstretched, wounded baseline at 3 h. 2 = signiWcant (P · 0.05)from unstretched, wounded skin at indicated post-operative times.Time points preceding those represented on Western blots were notsigniWcant from baseline, and therefore not displayed in Wgures
Post-Operative Time
0
0.51.0
1.52.0
2.5
3.03.5
4.04.5
5.0
Pro
tein
Lev
el (
arbi
trar
y un
its) unstretched, wounded
stretched, wounded1
1
1
Akt
tubulin tubulinpAkt
Pro
tein
Lev
el (
arbi
trar
y un
its)
2
1
0
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.45unstretched, woundedstretched, wounded
2
2
1
1
Post-Operative Time
12 h 1d 7d4d2d 3h 12h 1d 2d 4d 7d
1
2
A B
123

502 Arch Dermatol Res (2008) 300:495–504
sections compared to Western blots indicate that endothe-lial cells are the predominant cell type expressing the acti-vated insulin receptors. These data demonstrate that tissueinsulin and its receptors are stretch-responsive and arelocated on mitotic endothelial cells. Moreover, these resultssupport recent evidence demonstrating insulin’s role as akey mitogenic regulator and indicate a biologically signiW-cant role for insulin in the observed beneWts of acute stretch[9, 22, 26, 34, 40, 41].
Angiogenesis has been shown to be mediated throughPI3-K and downstream eVectors Akt and VEGF [6, 10, 15,17, 18, 42]. We demonstrated that acute stretch up-regu-lates VEGF, VEGF receptor 2 (Xk-1), and Akt, whileenhancing activation (pAkt) of Akt. Therefore, acutestretch, unlike wounding alone, works through both VEGFand Akt. This could be the result of acute stretch-inducedmechanical–chemical signaling that is transmitted through-out the endothelial cell and through extracellular matrix(ECM) adhesions [6, 10, 15, 16, 37]. These results are coin-cident with the up-regulation of tissue insulin and receptoractivation. These intracellular signaling events may serveas the link between stretch-induced insulin release andendothelial cell proliferation.
Mechanical induction of protein expression and activation provides temporal support for insulin-mediated endothelial cell proliferation
We hypothesized that acute stretch-induced endothelial cellproliferation is tissue insulin mediated through PI3-K eVec-tors, i.e., Akt (pAkt) and VEGF. The temporal relationshipsamong proteins up-regulated/activated by acute stretch pro-vides further evidence that insulin activates it’s receptors(pIR/pIGFR) and stimulates endothelial cell proliferation
(indicated by PCNA) through PI3-K by Akt (pAkt), VEGFand Xk-1 signaling in our in vivo mouse model.
Conclusions
We have presented a novel in vivo approach to study thebeneWts of a mechanical stimulus (i.e., acute stretch).Although, acute stretch plays a signiWcant role in regulatingendothelial cell proliferation and up-regulation/activationof insulin and key angiogenic factors, the exact mechanismthat results in angiogenesis remains elusive. The Westernblot data presented here are representative of all cell typesfound in murine skin. Future in vitro studies will focus onconXuent endothelial cell (EC) monolayers. This will allowfor a conclusive characterization of the implicated angio-genic pathway(s) in ECs using blockade studies lethal inour in vivo model.
It is important to note that our observations of improvedviability, as assessed by increased blood Xow and decreasednecrosis, can be attributed to several variables workingindividually or in conjunction. Among these are increasedvascular density, vasodilation, and physiological shunting.Consequences of interactions among these variables remainunclear. However, temporal observations of a steadyincrease in blood Xow, becoming optimal at post-operativeday 7 and regressing by day 10, are indicative of vascularremodeling and support a potential role for arteriogenesisand angiogenesis in the initial stages of this process.
Finally, these experiments demonstrate that acute stretchenhances the up-regulation/activation of proteins (e.g.PCNA, pAkt) that respond to wounding alone. Therefore,we conclude that tissue stretch not only promotes up-regu-lation/activation, but enhances underlying angiogenicmechanisms that are wound-responsive. Interestingly, wedid not demonstrate increases in perfusion from woundingalone in unstretched (control) Xaps (Fig. 1). This is attrib-uted to vascular growth-related blood Xow being below thedetection sensitivity of the laser Doppler Xowmeter [43].
Although, the exact mechanism(s) underlying stretch-responsive angiogenesis has yet to be determined, thisstudy provides the framework for future in vitro studies thatwill explicate the underlying signaling cascade(s).
Acknowledgments This research was supported by interim fundsfrom the WVU Department of Neurobiology and Anatomy (account #:DA491030020 fund #: 12301430) and WVU College of Engineeringand Mineral Resources (account #: DA310880010 fund #: 11100124).
References
1. Badr I, Brown MD, Egginton S, Hudlicka O, Milkiewicz M,Verhaeg J (2003) DiVerences in local environment determine the
Fig. 7 The temporal relationship of protein expression in stretched,wounded Xaps. Peak responses for pAkt, pIR/pIGFR, and PCNA ap-pear small, because baseline values used for normalization were sub-stantially elevated in Western blots
insulin
pcna
VEGF
flk-1
Akt
pAkt
pIR/pIGFR
0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
3h 1d 2d 4d 7d
Pro
tein
Lev
el (
arbi
trar
y un
its)
Post-Operative Time
123

Arch Dermatol Res (2008) 300:495–504 503
site of physiological angiogenesis in rat skeletal muscle. ExpPhysiol 88:565–568
2. Beeson M, Sajan MP, Dizon M, Grebenev D, Gomez-Daspet J,Miura A, Kanoh Y, Powe J, Bandyopadhyay G, Standaert ML, Fa-rese RV (2003) Activation of protein kinase C-zeta by insulin andphosphatidylinositol-3,4,5-(PO4)3 is defective in muscle in type 2diabetes and impaired glucose tolerance: amelioration by rosiglit-azone and exercise. Diabetes 52:1926–1934
3. Brewster L, Brey EM, Addis M, Xue L, Husak V, Ellinger J, Hau-denschild CC, Greisler HP (2006) Improving endothelial healingwith novel chimeric mitogens. Am J Surg 192:589–593
4. Brey EM, Uriel S, Greisler HP, McIntire LV (2005) Therapeuticneovascularization: contributions from bioengineering. TissueEng 11:567–584
5. Brown MD, Kent J, Kelsall CJ, Milkiewicz M, Hudlicka O (2003)Remodeling in the microcirculation of rat skeletal muscle duringchronic ischemia. Microcirculation 10:179–191
6. Chau CH, Chen KY, Deng HT, Kim KJ, Hosoya K, Terasaki T,Shih HM, Ann DK (2002) Coordinating Etk/Bmx activation andVEGF upregulation to promote cell survival and proliferation.Oncogene 21:8817–8829
7. Davis GE, Saunders WB (2006) Molecular balance of capillarytube formation versus regression in wound repair: role of matrixmetalloproteinases and their inhibitors. J Investig Dermatol SympProc 11:44–56
8. Folkman J (2003) Fundamental concepts of the angiogenic pro-cess. Curr Mol Med 3:643–651
9. Gordon SR (2002) MicroWlament disruption in a noncycling orga-nized tissue, the corneal endothelium, initiates mitosis. Exp CellRes 272:127–134
10. Haas TL (2002) Molecular control of capillary growth in skeletalmuscle. Can J Appl Physiol 27:491–515
11. Hochberg J, Raman M, Cilento E, Kemp K, Barrett M, Thomas R,Reilly F (1994) Development and evaluation of an in vivo mousemodel for studying myocutaneous Xap microcirculation and via-bility before and after suturing or stapling. Int J Microcirc Clin Exp14:67–72
12. Hochberg J, Beasley M, Jennings T, Zhu X, Boo S, Cilento EV,Reilly FD (1998) EVects of hold time, cycling, and catheter Wll-volume on the reported beneWts of rapid intraoperative tissueexpansion over simple undermining. Plast Surg Forum 286–289
13. Hochberg J, Zhu X, Beasley M, Cilento E, Reilly F (2000) Thebest estimates of total perfusion are from the base of myocutane-ous Xaps. In: Plast Surg Forum 452
14. Hudlicka O (1998) Is physiological angiogenesis in skeletal mus-cle regulated by changes in microcirculation? Microcirculation5:5–23
15. Ingber DE (2002) Mechanical signaling and the cellular responseto extracellular matrix in angiogenesis and cardiovascular physiol-ogy. Circ Res 91:877–887
16. Joung IS, Iwamoto MN, Shiu YT, Quam CT (2006) Cyclic strainmodulates tubulogenesis of endothelial cells in a 3D tissue culturemodel. Microvasc Res 71:1–11
17. Kanda S, Mochizuki Y, Kanetake H (2003) Stromal cell-derivedfactor-1alpha induces tube-like structure formation of endothelialcells through phosphoinositide 3-kinase. J Biol Chem 278:257–262
18. Kawaguchi M, Koshimura K, Sohmiya M, Murakami Y, Gonda T,Kato Y (2001) EVect of insulin on nitric oxide synthase-likeimmunostaining of arteries in various organs in Zucker diabeticfatty rats. Eur J Endocrinol 145:343–349
19. Kawamura A, Horie T, Tsuda I, Abe Y, Yamada M, Egawa H, IidaJ, Sakata H, Onodera K, Tamaki T, Furui H, Kukita K, Meguro J,Yonekawa M, Tanaka S (2006) Clinical study of therapeuticangiogenesis by autologous peripheral blood stem cell (PBSC)
transplantation in 92 patients with critically ischemic limbs. J ArtifOrgans 9:226–233
20. Kelkar BR (2003) Induced angiogenesis for limb ischemia. ClinOrthop Relat Res 234–240
21. Kim KE, Cho CH, Kim HZ, Baluk P, McDonald DM, Koh GY(2006) In vivo actions of angiopoietins on quiescent and remodel-ing blood and lymphatic vessels in mouse airways and skin. Arte-rioscler Thromb Vasc Biol
22. Kobayashi T, Kamata K (2002) Short-term insulin treatment andaortic expressions of IGF-1 receptor and VEGF mRNA in diabeticrats. Am J Physiol Heart Circ Physiol 283:H1761–H1768
23. Laing AJ, Dillon JP, Condon ET, Street JT, Wang JH, McGuin-ness AJ, Redmond HP (2007) Mobilization of endothelial precur-sor cells: systemic vascular response to musculoskeletal trauma. JOrthop Res 25:44–50
24. Layton CJ, Becker S, Osborne NN (2006) The eVect of insulin andglucose levels on retinal glial cell activation and pigment epithe-lium-derived Wbroblast growth factor-2. Mol Vis 12:43–54
25. Li S, Huang NF, Hsu S (2005) Mechanotransduction in endothelialcell migration. J Cell Biochem 96:1110–1126
26. Linn T, Erb D, Schneider D, Kidszun A, Elcin AE, Bretzel RG, El-cin YM (2003) Polymers for induction of revascularization in therat fascial Xap: application of vascular endothelial growth factorand pancreatic islet cells. Cell Transplant 12:769–778
27. Luo J, Miller MW (1996) Ethanol inhibits basic Wbroblast growthfactor-mediated proliferation of C6 astrocytoma cells. J Neuro-chem 67:1448–1456
28. Luo J, Lang JA, Miller MW (1998) Transforming growth factorbeta1 regulates the expression of cyclooxygenase in cultured cor-tical astrocytes and neurons. J Neurochem 71:526–534
29. Machida BK, Liu-Shindo M, Sasaki GH, Rice DH, ChandrasomaP (1991) Immediate versus chronic tissue expansion. Ann PlastSurg 26:227–231
30. Marino-Buslje C, Martin-Martinez M, Mizuguchi K, Siddle K,Blundell TL (1999) The insulin receptor: from protein sequence tostructure. Biochem Soc Trans 27:715–726
31. Milkiewicz M, Brown MD, Egginton S, Hudlicka O (2001) Asso-ciation between shear stress, angiogenesis, and VEGF in skeletalmuscles in vivo. Microcirculation 8:229–241
32. Munoz-Chapuli R, Gonzalez-Iriarte M, Carmona R, Atencia G,Macias D, Perez-Pomares JM (2002) Cellular precursors of thecoronary arteries. Tex Heart Inst J 29:243–249
33. Neuman CA (1957) The expansion of an area of skin by progres-sive distension of subcutaneous balloon. Plast Reconstr Surg 124–130
34. Pelegrinelli FF, Thirone AC, Gasparetti AL, Araujo EP, VellosoLA, Saad MJ (2001) Early steps of insulin action in the skin of in-tact rats. J Invest Dermatol 117:971–976
35. Sasaki GH (1987) Intraoperative sustained limited expansion(ISLE) as an immediate reconstructive technique. Clin Plast Surg14:563–573
36. Schramm JC, Dinh T, Veves A (2006) Microvascular changes inthe diabetic foot. Int J Low Extrem Wounds 5:149–159
37. Shiu YT, Weiss JA, Hoying JB, Iwamoto MN, Joung IS, Quam CT(2005) The role of mechanical stresses in angiogenesis. Crit RevBiomed Eng 33:431–510
38. Siegert R, Weerda H, HoVmann S, Mohadjer C (1993) Clinicaland experimental evaluation of intermittent intraoperative short-term expansion. Plast Reconstr Surg 92:248–254
39. Sieminski AL, Gooch KJ (2000) Biomaterial-microvasculatureinteractions. Biomaterials 21:2232–2241
40. Sundell J, Knuuti J (2003) Insulin and myocardial blood Xow. Car-diovasc Res 57:312–319
41. Wertheimer E, Trebicz M, Eldar T, Gartsbein M, Nofeh-Moses S,Tennenbaum T (2000) DiVerential roles of insulin receptor and
123

504 Arch Dermatol Res (2008) 300:495–504
insulin-like growth factor-1 receptor in diVerentiation of murineskin keratinocytes. J Invest Dermatol 115:24–29
42. Yu X, Rajala RV, McGinnis JF, Anderson RE, Yan X, Li S,Elias RV, Knapp RR, Zhou X, Cao W (2004) Involvement ofinsulin/phosphoinositide 3-kinase/akt signal pathway in 17-estradiol-mediated neuroprotection. J Biol Chem 279:13086–13094
43. Zhu X, Hall D, Ridenour G, Boo S, Jennings T, Hochberg J,Cilento E, Reilly F (2003) A mouse model for studying rapic intra-operative methods of skin closure and wound healing. Med SciMonit 9:109–115
44. Zhu Y, Luo J, Barker J, Hochberg J, Cilento E, Reilly F (2002)IdentiWcation of genes induced by rapid intraoperative tissueexpansion in mouse skin. Arch Dermatol Res 293:560–568
123





























