Embed Size (px)
Citation preview

Schizophrenia Research 143 (2013) 269–276
Contents lists available at SciVerse ScienceDirect
Schizophrenia Research
j ourna l homepage: www.e lsev ie r .com/ locate /schres
Altered cortico-basal ganglia motor pathways reflect reduced volitional motoractivity in schizophrenia
Tobias Bracht a, Susanne Schnell b,c, Andrea Federspiel d, Nadja Razavi d, Helge Horn a, Werner Strik a,Roland Wiest e, Thomas Dierks d, Thomas J. Müller a, Sebastian Walther a,⁎a University Hospital of Psychiatry, University of Bern, Bolligenstrasse 111, 3000 Bern 60, Switzerlandb Department of Diagnostic Radiology, Medical Physics, Freiburg, University Medical Centre Freiburg, Breisacherstrasse 60a, 79106, Germanyc Departments of Radiology and Biomedical Engineering, Northwestern University, Feinberg School of Medicine, 737N. Michigan Ave Suite 1600, Chicago, IL 60611, USAd University Hospital of Psychiatry, Department of Psychiatric Neurophysiology, University of Bern, Bolligenstrasse 111, 3000 Bern 60, Switzerlande Institute of Diagnostic and Interventional Neuroradiology, Inselspital, University of Bern, Freiburgstrasse 3010, Bern, Switzerland
⁎ Corresponding author. Tel.: +41 31 930 9111; fax:E-mail addresses: [email protected] (T. Bracht),
[email protected] (S. Schnell), Federsp(A. Federspiel), [email protected] (N. Razavi), horn@[email protected] (W. Strik), [email protected] (R(T. Dierks), [email protected] (T.J. Müller), walther
0920-9964/$ – see front matter © 2012 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.schres.2012.12.004
a b s t r a c t
a r t i c l e i n f oArticle history:Received 7 September 2012Received in revised form 9 November 2012Accepted 4 December 2012Available online 28 December 2012
Keywords:Diffusion tensor imagingWhite matterTractographyMovement disordersSchizophreniaActigraphy
Little is known about the neurobiology of hypokinesia in schizophrenia. Therefore, the aim of this study was toinvestigate alterations of white matter motor pathways in schizophrenia and to relate our findings to objectivelymeasuredmotor activity. We examined 21 schizophrenia patients and 21 healthy controls using diffusion tensorimaging and actigraphy. We applied a probabilistic fibre tracking approach to investigate pathways connectingthe dorsolateral prefrontal cortex (dlPFC), the rostral anterior cingulate cortex (rACC), the pre-supplementarymotor area (pre-SMA), the supplementary motor area proper (SMA-proper), the primary motor cortex (M1),the caudate nucleus, the striatum, the pallidumand the thalamus. Schizophrenia patients had lower activity levelsthan controls. In schizophrenia we found higher probability indices forming part of a bundle of interest (PIBI) inpathways connecting rACC, pre-SMA and SMA-proper as well as in pathways connecting M1 and pre-SMA withcaudate nucleus, putamen, pallidum and thalamus and a reduced spatial extension of motor pathways in schizo-phrenia. There was a positive correlation between PIBI and activity level in the right pre-SMA–pallidum and theleftM1–thalamus connection in healthy controls, and in the left pre-SMA–SMA-proper pathway in schizophrenia.Our results point to reduced volitional motor activity and altered motor pathway organisation in schizophrenia.The identified associations between the amount of movement and structural connectivity of motor pathwayssuggest dysfunction of cortico-basal ganglia pathways in the pathophysiology of hypokinesia in schizophrenia.Schizophrenia patients may use cortical pathways involving the supplementary motor area to compensate forbasal ganglia dysfunction.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Motor behaviour abnormalities such as Parkinsonism, dyskinesia,neurological soft signs or catatonic phenomena are an essential partof schizophrenia psychopathology (Koning et al., 2010; Walther andStrik, 2012). Motor abnormalities can be observed in children wholater develop schizophrenia (Walker et al., 1994). Furthermore,motor symptoms are prevalent in first episode, never-medicatedpatients (Peralta et al., 2010) and in medication-naïve chronicpatients (McCreadie et al., 2002). Despite the high prevalence of
+41 31 930 9404.
[email protected] (H. Horn),. Wiest), [email protected]@puk.unibe.ch (S. Walther).
rights reserved.
motor symptoms in schizophrenia, the neurobiological backgroundof motor symptoms is an unresolved issue (Walther and Strik, 2012).
Motor behaviour is a result of a complex network of brain areasincluding the dorsolateral prefrontal cortex (dlPFC), the rostral ante-rior cingulate cortex (rACC), the supplementary motor area (SMA),the primary motor cortex (M1), caudate nucleus, putamen, pallidumand thalamus (Graybiel, 1997; Groenewegen, 2003). Therefore, theconnection pathways of this functional network are of particular in-terest for understanding motor behaviour.
Abnormal connectivity of neuronal systemsmay underlie schizophre-nia psychopathology (Friston, 1998). This has led to an increasing focuson diffusion tensor imaging (DTI) studies in schizophrenia which pointtowhitemattermicrostructure alterations ofmotor-relatedwhitematterbundles, such as the corpus callosum, the corticospinal tract, the internalcapsule or the superior longitudinal fasciculus (Douaud et al., 2007;Cheung et al., 2008; Kyriakopoulos et al., 2008; Wang et al., 2011).
DTI-based fibre tracking enables the reconstruction of specificpathways based on the directionality of the diffusion tensor. It has

270 T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
been used to investigate cortico-basal ganglia pathways in healthy sub-jects (Behrens et al., 2003; Draganski et al., 2008; Cohen et al., 2009) aswell as in patientswith neuropsychiatric disorders such asHuntington'sdisease or schizophrenia (Kim et al., 2008; Klöppel et al., 2008).Where-as deterministic fibre trackingdelineates a single best path, probabilisticfibre tracking quantifies the probability that two brain regions are struc-turally interconnected thereby taking uncertainties of the diffusion ten-sor into account (Jones et al., 2012). Therefore, several groups appliedprobabilistic fibre tracking approaches (Behrens et al., 2007; Priceet al., 2007; Kubicki et al., 2011). Despite advantages of probabilisticfibre tracking in regions with crossing fibres, inaccuracies of the diffu-sion tensor might cumulate, in particular in long distance pathways.Therefore, we used a bilateralfibre tracking approach from both regionsof interest in conjunction with a combination of the resulting probabil-ity maps; a method specifically designed to extract long distance path-ways between two regions (Kreher et al., 2008).
White matter abnormalities of motor pathways have been reportedbefore it remains difficult to relate thosefindings to relevant behaviour-al measures. Actigraphy is a valuable tool for studyingmotor behaviourin schizophrenia (Farrow et al., 2005; Walther et al., 2009a,b;Wichniaket al., 2011; Bracht et al., 2012). Combined brain imaging and actigraphystudies indicate reduced quantitative motor behaviour and altered as-sociations of motor activity and brain structure and function of regionsof the motor system including the ACC, the SMA and the thalamus inschizophrenia (Farrow et al., 2005; Walther et al., 2011a,b). However,to date no fibre tracking study specifically investigated the relationshipbetween cortico-basal ganglia motor pathway organisation and motorbehaviour in schizophrenia.
The aim of the study was to investigate associations between quan-titative motor behaviour and structural connectivity of cortico-corticaland cortico-basal ganglia motor pathways in schizophrenia. We hy-pothesized that (i) quantitative volitional motor activity is reducedin schizophrenia as compared to controls (Walther and Strik, 2012).(ii) Structural connectivity between key areas of the motor systemdiffers between schizophrenia patients and controls. Based on previousDTI-basedfibre tracking studies in schizophrenia demonstrating reduced
Table 1Demographics of schizophrenia patients and healthy controls.
Variable SZ patients (n=21)
Gender Male 13Female 8
Age (years) 34.2±8.2Handedness Right 21
Left 0Caucasian 18Married 1Schizophrenia typeParanoid type 12Disorganized type 5Catatonic type 2Residual type 2Years of education 13.7±4.8Age at onset 26.4±6.9Duration of illness 8.07±9.6Number of episodes 4.80±6.4Medication dosage (CPZ equivalent)c 698±644Type of medication Atypical 20(Atypical/typical)d Both 1PANSS total 55.5±17.03PANSS positive 12.0±5.3PANSS negative 17.4±8.2Actigraphy 13,542±7272
Data are means±standard deviations.Abbreviations: SZ, schizophrenia; CPZ, chlorpromazine equivalents; PANSS, Positive and Ne
a Mann–Whitney-U-tests were calculated.b Chi2 test were calculated.c Chlorpromazine equivalents were calculated based on (Woods, 2003).d 20 patients received atypical antipsychotics (clozapine, risperidone, olanzapine, aripipre Two-sample t-tests were calculated.
pathway volume and length (Buchsbaum et al., 2006; Kubicki et al.,2011), we assumed a reduced spatial extension ofmotor pathways lead-ing to an increase of probability of connection within these pathwaysand (iii) the alterations of structural connectivity in the motor systemin schizophrenia are associated with the reduced motor activity.
2. Material and methods
2.1. Participants
We recruited 21 inpatients with schizophrenia and 21 healthycontrols from the University Hospital of Psychiatry, Bern. Diagnoseswere made according to DSM-IV criteria after clinical examinationsand based on review of all available case files. Psychopathology wasassessed using the Positive and Negative Syndrome Scale (PANSS)(Kay et al., 1987). Exclusion criteria to all participants were historyof substance abuse other than nicotine, neurological disorders or ahistory of severe head trauma. All participants provided written in-formed consent. The study was approved by the local ethics commit-tee and adhered to the Declaration of Helsinki. This study includessubjects who participated in previous studies (Walther et al., 2010,2011a). See Table 1 for demographic information.
2.2. Data acquisition
2.2.1. ActigraphyFollowing the MRI scans in the morning, participants wore an
actigraph (Actiwatch®, Cambridge Neurotechnology, Inc., UK) on thewrist of the left (non-dominant) arm for the 24 consecutive hours.Actigraphy on the non-dominant arm reflects total motor activity(Middelkoop et al., 1997). Activity counts were stored in 2-second in-tervals. Participants provided information about the timing and dura-tion of sleep. Data were extracted using Sleep analysis 5® software(Cambridge Neurotechnology, Inc., UK). We calculated the activitylevel (AL), defined as the cumulative activity counts while participants
Controls (n=21) SZ vs. controls
Male 13Female 8
34.6±8.2 Z=0.56, P=0.58a
Right 21Left 0
21 Chi2=3.23, P=0.07b
6 Chi2=4.29, P=0.04b
17.2±4.0 Z=2.69, P=0.007a
– –
– –
– –
– –
–
–
– –
– –
– –
21,211±6462 T=3.612, P=0.001e
gative Syndrome Scale.
azole, amisulpride), 1 patient received haloperidol and olanzapine.

271T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
were awake divided by the net recording time in hours for each partic-ipant (Walther et al., 2010, 2011a).
2.2.2. MRI acquisitionAll images were acquired with a 12-channel signal reception head
coil on a 3-TMR scanner (SiemensMagnetomTrio, Erlangen, Germany).High-resolution T1-weighted MR images were obtained using a 3DModified Driven Equilibrium Fourier Transform (MDEFT) sequence(Deichmann et al., 2004) with 176 sagittal slices, 256×224 matrixwith an isotropic resolution of 1 mm3, acquired in 12 min. Identicalprescription of MR images was achieved using the Siemens Autoalignsequence, which automatically sets up consistent slice orientationbased on a standard MRI atlas.
2.2.3. Diffusion Tensor Imaging (DTI)For DTI measurements, we used a spin-echo echo-planar-imaging
(EPI) sequence (55 slices, FOV=256×256 mm2, sampled on a 128×128matrix resulting in 2 mm3 voxel size, TR/TE=6000/78 ms) coveringthe whole brain (40 mT/m gradient, 5/8 partial Fourier, no accelera-tion factor, bandwidth 1346 Hz/Px). Forty-two diffusion-weightedvolumes were measured in the axial plane parallel to the AC-PCline with a b-value=1300 s/mm2. Four T2-weighted images weremeasured after the first and every 12th diffusion-weighted image(i.e. b-value=0, B0 image). We used a balanced and rotationally in-variant diffusion-encoding scheme over the unit sphere to generatethe DTI data (Hasan et al., 2001).
2.3. Data analysis
Imaging data were analysed using Statistical Parametric Mapping(SPM8) (www.fil.ion.ucl.ac.uk/spm), implemented in Matlab 7.6.0(R2008a; Mathworks, Natick, MA, USA).
2.3.1. Regions of interestGrey matter regions of interest (ROIs) localised within the Montreal
Neurological Institute (MNI) reference brain were selected using theSPM8 WFU-Pick-Atlas (Maldjian et al., 2003). The dlPFC includesBrodmann area (BA) 9 and 46 (Nieuwenhuys et al., 2008). The rACC iscomposed of parts of BA24 rostral to a vertical section through thegenu (Nieuwenhuys et al., 2008; Vogt, 2009). The border of pre-SMAand SMA-proper coincides with the vertical anterior commissural line(AC) of BA6 (Habas, 2010). Definitions of the primary motor cortex(M1), thalamus, putamen, caudate nucleus and pallidum have beentaken from the WFU-Pick-Atlas.
2.3.2. Pre-processingThe 4 B0 images were motion corrected using the SPM8 diffusion
toolbox (http://sourceforge.net/projects/spmtools). DTI-images werethen realigned on the B0 images. The T1-images were segmentedinto white and grey matter masks. This procedure provided thenormalisation parameters for forward and backward transforma-tions between individual native spaces and the MNI space. Becausefibre tracking is done in the native space of each subject, allROIs were transferred into the native space using the inversenormalisation parameters of each subject. The T1-image and theinverse normalised ROIs of each subject were co-registered to thecorresponding B0-image.
2.3.3. Probabilistic DTI-based fibre trackingThe fibre tracking and combination of probabilistic maps were
performed using a method of pathway extraction (Kreher et al., 2008)from the Matlab-based ‘DTI & Fibre Toolbox’ (www.uniklinik-freiburg,de/mr/live/arbeitsgruppen/diffusion_en.html). The same approach hasbeen used in previous studies e.g. (Saur et al., 2008; Bracht et al.,2009). First, the diffusion tensor was computed (Basser et al., 1994)and second a Monte Carlo simulation of random walks similar to the
probabilistic index of connectivity (Pico) method (Parker et al., 2003)was used to calculate probabilistic maps separately for each ROI. Inthese maps the visiting frequency of a voxel represents a degree ofconnectivity to the seed region. The number of random walks wasset to 25,000 and the maximum fibre length to 150 voxels. The track-ing area was restricted to the white matter mask obtained from thesegmentation of the T1-image to avoid tracking across anatomicalborders. A rim of grey matter (10% of the grey matter mask) wasincluded in the mask to ensure contact with the grey matter ROIs(Saur et al., 2008).
2.3.4. Connection between two seed regionsRegion to region anatomical connectivity between two ROIs was
computed by combining their respective probability maps (Kreheret al., 2008). This combination implies amultiplication that takes the di-rectional information of the random walk into account. Random walksstarting from ROI A and B may face in opposing directions (connectingfibres) or in the same direction (merging fibres). Within the pathwayconnecting A and B the proportion of connecting fibres should exceedthe proportion of merging fibres. Merging fibres are suppressed andconnecting fibre is preserved (Kreher et al., 2008). This method enablesthe extraction of the most probable direct pathway between two re-gions of interest. The resulting values represent a voxel-wise estimationof the probability index that a voxel is part of the connecting fibre bun-dle of interest (PIBI).
In our study, we bilaterally assessed structural connectivity be-tween cortico-cortical connections (dlPFC–pre-SMA, rACC–pre-SMA,pre-SMA–SMA-proper and SMA-proper–M1), between the pre-SMAand basal ganglia (pre-SMA–caudate nucleus, pre-SMA–putamen,pre-SMA–pallidum and pre-SMA–thalamus), and between the M1and basal ganglia (M1–caudate nucleus, M1–putamen, M1–pallidumand M1–thalamus).
The combined probability maps of each subject were scaled be-tween 0 and 1, spatially normalised into the standard MNI space,and smoothed using an isotropic 3-mm Gaussian kernel.
2.3.5. Statistical analysisIn order to compare the structural connectivity between groups,
PIBI-values of normalisedprobabilitymaps for each putative connectionwere compared between groups using independent t-tests in SPM8.Weset the statistical threshold at pb0.05 applying a family-wise error(FWE) correction for multiple comparisons.
To further investigate the nature of the group differences in PIBI-values, we were interested in the characteristics of the probabilitymaps. Therefore, mean-fractional anisotropy (FA), mean–mean dif-fusivity (MD) and the number of voxels were extracted from thosenative space probability maps which showed differences in PIBIbetween groups. To remove random artefacts only voxels withPIBI>0.0148 were included representing the upper 5% of PIBIs(Saur et al., 2008). Mean-FA, mean-MD and number of voxels wereeach entered in separate MANOVAs with the dependent variablegroup and the respective values of the extracted maps as indepen-dent variables.
To explore the differences between groups regarding the relation-ship between motor behaviour and fibre tract organisation of the 24combined probabilitymaps a general linearmodel (GLM)was calculatedin SPM8. Independent variable was group (controls vs. schizophrenia)and dependent variables were PIBI and AL. We employed a significancethreshold of pb0.05, FWE corrected.
We used Shapiro–Wilk tests to check for normal distribution of vari-ables. The demographic characteristics of patients and controls werecompared with independent t-tests, Mann–Whitney U-tests, Pearsoncorrelations, Spearman correlations and chi-square tests. Statisticalanalyses were performed using SPSS 18 (Chicago, IL, USA).

Table 3Post-hoc independent t-tests for the number of voxels within the probabilistic pathways.
Connection Controls SZ T df p
rACC–pre-SMA (L) 1542±787 961±558 2.785 40 0.009⁎⁎
pre-SMA–SMA-proper (R) 2762±765 1613±960 4.223 40 b0.001⁎⁎⁎
Pre-SMA–Caudate (R) 3633±1521 2483±1388 2.558 40 0.014⁎
Pre-SMA–pallidum (R) 3616±1253 2759±1354 2.128 40 0.040⁎
M1–Caudate (L) 1866±877 2238±1689 −0.894 40 0.377M1–putamen (L) 2292±772 1677±736 2.641 40 0.012⁎
M1–pallidum (L) 2493±856 1816±824 2.612 40 0.013⁎
M1–thalamus (L) 2396±494 2184±1256 0.721 40 0.475M1–thalamus (R) 2225±880 2203±1623 0.055 40 0.957
Abbreviations: SZ, schizophrenia patients; L, left; R, right; rACC, rostral anterior cingulatecortex; SMA, supplementary motor area; M1, primary motor cortex.
⁎ pb0.05.⁎⁎ pb0.01.
⁎⁎⁎ pb0.001.
272 T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
3. Results
3.1. Group comparisons of activity level
ALwas higher in controls than in patients (see Table 1). AL in healthycontrols was not correlated with age (r=0.07, p=0.78) or years ofeducation (r=0.34, p=0.13). AL in patients was not significantlycorrelated with age (r=−0.20, p=0.38), years of education (r=0.053,p=0.82), duration of illness (r=−0.10, p=0.66), chlorpromazineequivalents (r=−0.098, p=0.67), PANSS score [total syndrome score(r=−0.11, p=0.65), positive syndrome score (r=0.42, p=0.062),negative syndrome score (r=−0.12, p=0.59)], number of episodes(r=−0.11, p=0.64) or age at onset (r=−0.14, p=0.56).
3.2. Group comparisons of probability indices forming part of a bundle ofinterest
In 9 of 24 maps, independent t-tests with FWE correction revealedsignificantly higher PIBI in schizophrenia patients than in controls(see Table 2). None of the map controls had higher PIBI values.MANOVAs of the 9mapswith higher PIBI in schizophrenia revealed sig-nificant differences between groups regarding the number of voxels(F=5.266, df=9, pb0.001) and mean-MD (F=2.416, df=9, p=0.032) but not regarding mean-FA (F=2.112, df=9, p=0.058).Post-hoc independent t-tests comparingmean-MDof the ninemaps be-tween groups did not reveal significant results. Post-hoc independentt-tests for the number of voxels are given in Table 3.
3.3. Associations of activity level and probability indices forming part of abundle of interest
The GLM revealed significant different associations between PIBIand AL between groups in the left pre-SMA–SMA-proper connection(F=22.45, pb0.001, number of voxels=2, MNI-coordinates, −32,2, 48, controls: r=−0.044, p=0.850; patients r=0.479, p=0.028,see Fig. 2A), in the right pre-SMA-pallidum connection (F=20.44,pb0.001, number of voxels=5, MNI-coordinates 26,−4, 2, controls:r=0.577, p=0.006; patients: r=0.310, p=0.172, Fig. 2B), and inthe left M1-thalamus connection (F=16.8, pb0.001, number ofvoxels=1, MNI-coordinates −22, −24, 10, controls: r=0.551,p=0.010; patients r=0.218, p=0.343, Fig. 2C).
4. Discussion
The investigation had three main findings. First, we found reducedactivity levels in schizophrenia. Second, pathways of schizophreniapatients involved in motor control showed higher PIBI values. Third,
Table 2Comparisons between PIBI in schizophrenia patients and healthy controls.
Connections Left side
T-value p-Value MNI-coordinates
rACC–pre-SMA 2.7 0.026 −10, 14, 24dlPFC–pre-SMA n.s.Pre-SMA–SMA-proper 4.8 0.001 −32, 2, 48SMA-proper–BA4 n.sPre-SMA–Caudate n.s.Pre-SMA–putamen n.s.Pre-SMA–pallidum n.s.Pre-SMA–thalamus n.s.M1–Caudate 4.1 0.012 −20, −6, 22M1–putamen 4.2 0.011 −26, −30, 40M1–pallidum 3.9 0.028 −30, −16 18M1–thalamus 3.9 0.033 −24, −30, 32
Maximum T-values, p-values, corresponding MNI coordinates and the number of voxels (k)higher PIBI values in all clusters. For all analyses, pb0.05 after FWE correction for multiple ccortex; SMA, supplementary motor area; M1, primary motor cortex; dlPFC, dorsolateral pre
there were positive correlations between PIBI values and motor activ-ity. In healthy controls compared to schizophrenia patients, these cor-relations were found in different pathways of the motor system.
We emphasize that higher PIBI does not reflect higher connectionstrength or pathway integrity. Rather, it indicates that a candidatepathway was reconstructed more frequently in the probabilisticfibre tracking calculations (Jones et al., 2012). Possible causesinclude variations of pathway width, length, branching, curvature,axon density or myelination of the candidate pathways (Beaulieu,2002). The higher PIBI values in schizophrenia may result from thereduced spatial extension of statistically defined motor pathways.In other words, like the increased flow in a bottleneck, the identicalnumber of random walks is condensed in smaller pathway volumesin schizophrenia compared to controls (see supplementary figure).Our finding is consistent with a recent probabilistic fibre trackingstudy identifying elevated connectivity of the thalamus with thesomato-sensory cortex in schizophrenia (Marenco et al., 2012). Thereduced spatial distribution of probabilistic maps is in line with re-ports of disrupted fibre connectivity (Zalesky et al., 2011) as wellas volume decreases and shortening of pathways in schizophrenia(Buchsbaum et al., 2006; Kubicki et al., 2011). Furthermore, previousprobabilistic fibre tracking studies investigating pre-symptomatic andsymptomatic Huntington's disease revealed reductions in cortico-caudate streamlines, which were associated with motor impairment(Klöppel et al., 2008; Bohanna et al., 2011). In contrast, diffusion prop-erties but not PIBI values of motor pathways differed in major depres-sive disorder compared to healthy controls (Bracht et al., 2012). Onemight speculate whether the reduced spatial extension of motor path-ways is caused by neuronal loss in schizophrenia which has beensuggested in some histological studies in schizophrenia (Harrison,
Right side
k T-value p-Value MNI-coordinates k
4 n.s.n.s.
4 n.s.n.s.
3.5 0.048 20, 8, 16 1n.s.
3.9 0.033 22, 0, 0 4n.s.
2 n.s.2 n.s.4 n.s.2 4.05 0.011 26, −22, 16 9
are the result of independent t-tests. Controls and patients are contrasted. Patients hadomparisons was considered significant. Abbreviations: rACC, rostral anterior cingulatefrontal cortex; n.s., not significant.
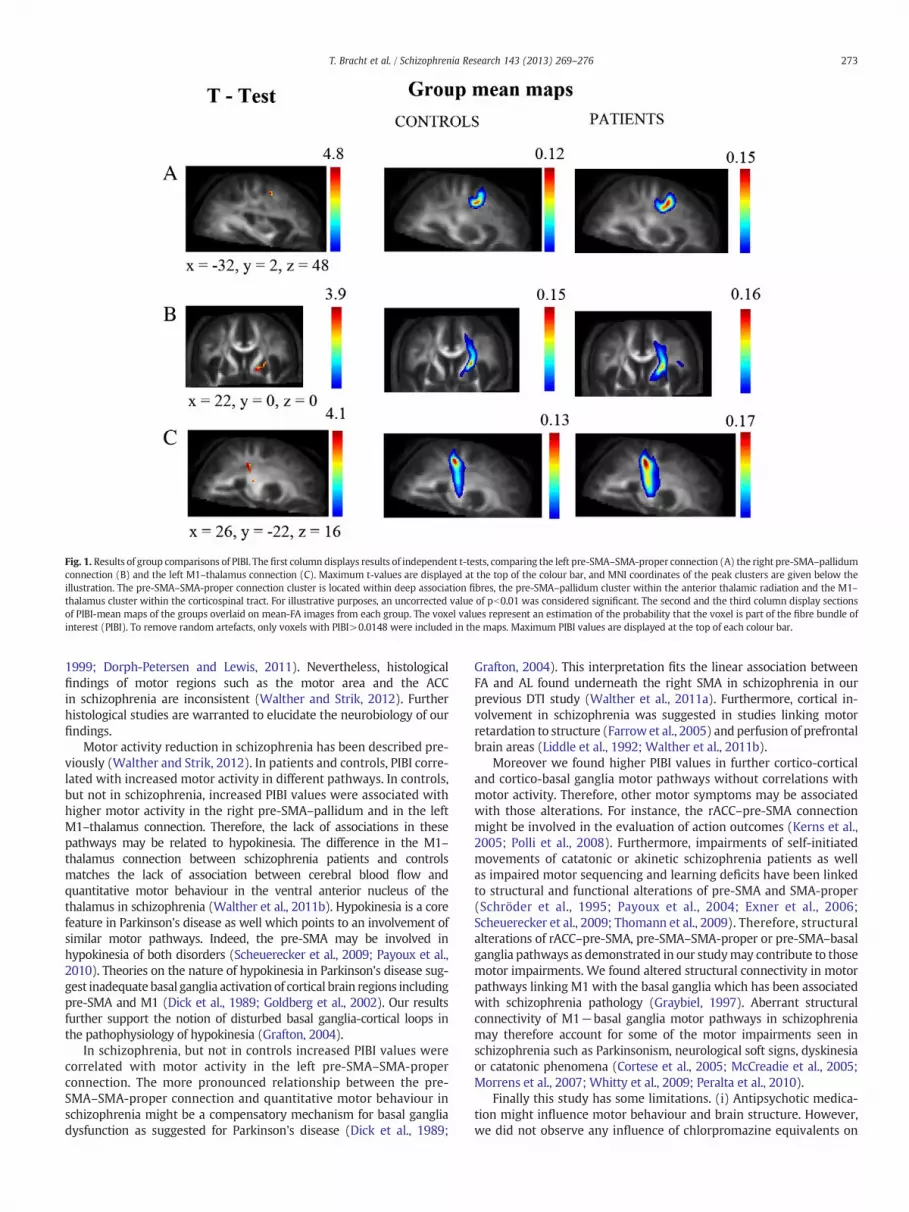
Fig. 1. Results of group comparisons of PIBI. The first column displays results of independent t-tests, comparing the left pre-SMA–SMA-proper connection (A) the right pre-SMA–pallidumconnection (B) and the left M1–thalamus connection (C). Maximum t-values are displayed at the top of the colour bar, and MNI coordinates of the peak clusters are given below theillustration. The pre-SMA–SMA-proper connection cluster is located within deep association fibres, the pre-SMA–pallidum cluster within the anterior thalamic radiation and the M1–thalamus cluster within the corticospinal tract. For illustrative purposes, an uncorrected value of pb0.01 was considered significant. The second and the third column display sectionsof PIBI-mean maps of the groups overlaid on mean-FA images from each group. The voxel values represent an estimation of the probability that the voxel is part of the fibre bundle ofinterest (PIBI). To remove random artefacts, only voxels with PIBI>0.0148 were included in the maps. Maximum PIBI values are displayed at the top of each colour bar.
273T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
1999; Dorph-Petersen and Lewis, 2011). Nevertheless, histologicalfindings of motor regions such as the motor area and the ACCin schizophrenia are inconsistent (Walther and Strik, 2012). Furtherhistological studies are warranted to elucidate the neurobiology of ourfindings.
Motor activity reduction in schizophrenia has been described pre-viously (Walther and Strik, 2012). In patients and controls, PIBI corre-lated with increased motor activity in different pathways. In controls,but not in schizophrenia, increased PIBI values were associated withhigher motor activity in the right pre-SMA–pallidum and in the leftM1–thalamus connection. Therefore, the lack of associations in thesepathways may be related to hypokinesia. The difference in the M1–thalamus connection between schizophrenia patients and controlsmatches the lack of association between cerebral blood flow andquantitative motor behaviour in the ventral anterior nucleus of thethalamus in schizophrenia (Walther et al., 2011b). Hypokinesia is a corefeature in Parkinson's disease as well which points to an involvement ofsimilar motor pathways. Indeed, the pre-SMA may be involved inhypokinesia of both disorders (Scheuerecker et al., 2009; Payoux et al.,2010). Theories on the nature of hypokinesia in Parkinson's disease sug-gest inadequate basal ganglia activation of cortical brain regions includingpre-SMA and M1 (Dick et al., 1989; Goldberg et al., 2002). Our resultsfurther support the notion of disturbed basal ganglia-cortical loops inthe pathophysiology of hypokinesia (Grafton, 2004).
In schizophrenia, but not in controls increased PIBI values werecorrelated with motor activity in the left pre-SMA–SMA-properconnection. The more pronounced relationship between the pre-SMA–SMA-proper connection and quantitative motor behaviour inschizophrenia might be a compensatory mechanism for basal gangliadysfunction as suggested for Parkinson's disease (Dick et al., 1989;
Grafton, 2004). This interpretation fits the linear association betweenFA and AL found underneath the right SMA in schizophrenia in ourprevious DTI study (Walther et al., 2011a). Furthermore, cortical in-volvement in schizophrenia was suggested in studies linking motorretardation to structure (Farrowet al., 2005) and perfusion of prefrontalbrain areas (Liddle et al., 1992; Walther et al., 2011b).
Moreover we found higher PIBI values in further cortico-corticaland cortico-basal ganglia motor pathways without correlations withmotor activity. Therefore, other motor symptoms may be associatedwith those alterations. For instance, the rACC–pre-SMA connectionmight be involved in the evaluation of action outcomes (Kerns et al.,2005; Polli et al., 2008). Furthermore, impairments of self-initiatedmovements of catatonic or akinetic schizophrenia patients as wellas impaired motor sequencing and learning deficits have been linkedto structural and functional alterations of pre-SMA and SMA-proper(Schröder et al., 1995; Payoux et al., 2004; Exner et al., 2006;Scheuerecker et al., 2009; Thomann et al., 2009). Therefore, structuralalterations of rACC–pre-SMA, pre-SMA–SMA-proper or pre-SMA–basalganglia pathways as demonstrated in our studymay contribute to thosemotor impairments. We found altered structural connectivity in motorpathways linking M1 with the basal ganglia which has been associatedwith schizophrenia pathology (Graybiel, 1997). Aberrant structuralconnectivity of M1−basal ganglia motor pathways in schizophreniamay therefore account for some of the motor impairments seen inschizophrenia such as Parkinsonism, neurological soft signs, dyskinesiaor catatonic phenomena (Cortese et al., 2005; McCreadie et al., 2005;Morrens et al., 2007; Whitty et al., 2009; Peralta et al., 2010).
Finally this study has some limitations. (i) Antipsychotic medica-tion might influence motor behaviour and brain structure. However,we did not observe any influence of chlorpromazine equivalents on

Fig. 2. Associations between probability of connection and activity level in a general linear model. The first column displays results of the general linear model for the left pre-SMA–SMAproper connection (A), for the right pre-SMA–pallidum connection (B) and for the left M1–thalamus connection (C). The pre-SMA–SMA-proper cluster is located within deepcortico-cortical association fibres and localised identical to where we found the differences of PIBI (Fig. 1A). The pre-SMA–pallidum cluster is located in close proximity to globuspallidus externus and putamen and slightly more lateral, posterior and dorsal to the cluster that shows a difference of PIBI (Fig. 1B). The M1–thalamus cluster is located within thecorticospinal tract andmedial, anterior and ventral to the cluster that shows a difference of PIBI (Fig. 1C). MNI-coordinates of peak values of the clusters are given below the illustrations.F-values are given on top of the colour bar. For illustrative purposes, results are displayed at a level of significance of pb0.01, uncorrected. The second columndisplays PIBI values extractedfrom the identified clusters and the corresponding activity level values of the respective subjects. Controls are displays as blue circles, patients as red diamonds. Horizontal dotted linesindicate mean PIBI values, vertical dotted lines indicate mean activity levels. Abbreviations: SMA, supplementarymotor area; M1, primarymotor cortex; PIBI, probability indices formingpart of a bundle of interest.
274 T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
AL in schizophrenia patients, which is in line with previous studies(Walther et al., 2009a,b). Moreover, larger studies were unable toconfirm an effect of medication on white matter microstructure
(Kanaan et al., 2009). (ii) DTI-based measures such as FA or PIBI areunspecific regarding themicrostructural changes. Future studies shouldapply novel white matter mapping techniques aiming at quantify axon

275T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
density, diameter and myelination (Jones et al., 2012). (iii) Identifiedclusters are relatively small in size. However, one should bear in mindthat we applied a FWE correction and that we restricted group compar-isons to the limited region of the identified fibre tracts (unlike wholebrain approaches). Comparing PIBI values of controls and patients insurrounding voxels it becomes evident that surrounded voxels displaythe same trend but do not reach significance with FWE correction(seemean probabilitymaps Fig. 1). Therefore, identified clusters shouldrather be seen as peaks of structural alterations than as isolated spots.
In conclusion, identified correlations between the amount of move-ment and structural connectivity suggest dysfunction of cortico-basalganglia pathways in the pathophysiology of hypokinesia in schizophre-nia. Schizophrenia patients may use cortical brain regions to compen-sate for basal ganglia dysfunction.
Role of the funding sourceThere was no external funding.
ContributorsTB analysed the data and wrote the first draft of the paper. SS, AF and RW provided
methodological support. NR recruited the patients and discussed findings. HH designedthe study and discussed findings. TD and TM interpreted the data. WS edited the paperand interpreted the data. SW designed the study, analysed data, supervised the studyand edited the paper. All authors contributed to and have approved the final manuscript.
Conflict of interestDr. Horn has received speaker's fees from Eli-Lilly, Bristol-Myers-Squibb, Lundbeck,
Boehringer-Ingelheim and Vifor. Prof. Strik received speaker's fees from/is advisoryboardmember of Eli-Lilly, Pfizer and Lundbeck. Prof. Müller has received research supportfrom Eli-Lilly, speaker's fees from AstraZeneca, Bristol-Myers-Squibb, Janssen-Cilag,Eli-Lilly, Pfizer, Sanofi-Aventis, Glaxo-Smith-Kline, Desitin-Pharma and Vifor. Dr. Mülleris advisory board member of Eli-Lilly, Janssen-Cilag, Sandoz and Lundbeck; he has beena consultant to Janssen-Cilag and Glaxo-Smith-Kline. Dr. Walther has received speaker'sfees from Janssen-Cilag and Sandoz. All other authors declare that they have no conflictsof interest.
AcknowledgementNone.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.schres.2012.12.004.
References
Basser, P.J., Mattiello, J., LeBihan, D., 1994. Estimation of the effective self-diffusiontensor from the NMR spin echo. J. Magn. Reson. B 103 (3), 247–254.
Beaulieu, C., 2002. The basis of anisotropic water diffusion in the nervous system — atechnical review. NMR Biomed. 15 (7–8), 435–455.
Behrens, T.E., Johansen-Berg, H., Woolrich, M.W., Smith, S.M., Wheeler-Kingshott, C.A.,Boulby, P.A., Barker, G.J., Sillery, E.L., Sheehan, K., Ciccarelli, O., Thompson, A.J.,Brady, J.M., Matthews, P.M., 2003. Non-invasive mapping of connections betweenhuman thalamus and cortex using diffusion imaging. Nat. Neurosci. 6 (7), 750–757.
Behrens, T.E., Berg, H.J., Jbabdi, S., Rushworth,M.F., Woolrich, M.W., 2007. Probabilistic dif-fusion tractography with multiple fibre orientations: what can we gain? Neuroimage34 (1), 144–155.
Bohanna, I., Georgiou-Karistianis, N., Egan, G.F., 2011. Connectivity-based segmentationof the striatum in Huntington's disease: vulnerability of motor pathways. Neurobiol.Dis. 42 (3), 475–481.
Bracht, T., Tüscher, O., Schnell, S., Kreher, B., Rüsch, N., Glauche, V., Lieb, K., Ebert, D.,Il'yasov, K.A., Hennig, J., Weiller, C., van Elst, L.T., Saur, D., 2009. Extraction ofprefronto-amygdalar pathways by combining probability maps. Psychiatry Res. 174(3), 217–222.
Bracht, T., Heidemeyer, K., Koschorke, P., Horn, H., Razavi, N., Wopfner, A., Strik, W.,Walther, S., 2012. Comparison of objectively measured motor behavior with ratingsof the motor behavior domain of the Bern Psychopathology Scale (BPS) in schizo-phrenia. Psychiatry Res. 198 (2), 224–229.
Bracht, T., Federspiel, A., Schnell, S., Horn, H., Höfle, O., Wiest, R., Dierks, T., Strik, W.,Müller, T.J., Walther, S., 2012. Cortico-cortical white matter motor pathway micro-structure is related to psychomotor retardation in major depressive disorder. PLoSONE 7 (12), e52238 http://dx.doi.org/10.1371/journal.pone.0052238.
Buchsbaum, M.S., Schoenknecht, P., Torosjan, Y., Newmark, R., Chu, K.W., Mitelman, S.,Brickman, A.M., Shihabuddin, L., Haznedar, M.M., Hazlett, E.A., Ahmed, S., Tang, C.,2006. Diffusion tensor imaging of frontal lobe white matter tracts in schizophrenia.Ann. Gen. Psychiatry 5, 19.
Cheung, V., Cheung, C., McAlonan, G.M., Deng, Y., Wong, J.G., Yip, L., Tai, K.S., Khong,P.L., Sham, P., Chua, S.E., 2008. A diffusion tensor imaging study of structuraldysconnectivity in never-medicated, first-episode schizophrenia. Psychol. Med.38 (6), 877–885.
Cohen, M.X., Schoene-Bake, J.C., Elger, C.E., Weber, B., 2009. Connectivity-based segre-gation of the human striatum predicts personality characteristics. Nat. Neurosci. 12(1), 32–34.
Cortese, L., Caligiuri, M.P., Malla, A.K., Manchanda, R., Takhar, J., Haricharan, R.,2005. Relationship of neuromotor disturbances to psychosis symptoms infirst-episode neuroleptic-naive schizophrenia patients. Schizophr. Res. 75(1), 65–75.
Deichmann, R., Schwarzbauer, C., Turner, R., 2004. Optimisation of the 3D MDEFTsequence for anatomical brain imaging: technical implications at 1.5 and 3 T.Neuroimage 21 (2), 757–767.
Dick, J.P., Rothwell, J.C., Day, B.L., Cantello, R., Buruma, O., Gioux, M., Benecke, R.,Berardelli, A., Thompson, P.D., Marsden, C.D., 1989. The Bereitschaftspotential isabnormal in Parkinson's disease. Brain 112 (Pt 1), 233–244.
Dorph-Petersen, K.A., Lewis, D.A., 2011. Stereological approaches to identifying neuro-pathology in psychosis. Biol. Psychiatry 69 (2), 113–126.
Douaud, G., Smith, S., Jenkinson, M., Behrens, T., Johansen-Berg, H., Vickers, J., James, S.,Voets, N., Watkins, K., Matthews, P.M., James, A., 2007. Anatomically related greyand white matter abnormalities in adolescent-onset schizophrenia. Brain 130(Pt 9), 2375–2386.
Draganski, B., Kherif, F., Klöppel, S., Cook, P.A., Alexander, D.C., Parker, G.J., Deichmann,R., Ashburner, J., Frackowiak, R.S., 2008. Evidence for segregated and integrativeconnectivity patterns in the human Basal Ganglia. J. Neurosci. 28 (28), 7143–7152.
Exner, C., Weniger, G., Schmidt-Samoa, C., Irle, E., 2006. Reduced size of the pre-supplementary motor cortex and impaired motor sequence learning in first-episode schizophrenia. Schizophr. Res. 84 (2–3), 386–396.
Farrow, T.F., Hunter, M.D., Wilkinson, I.D., Green, R.D., Spence, S.A., 2005. Structuralbrain correlates of unconstrained motor activity in people with schizophrenia.Br. J. Psychiatry 187, 481–482.
Friston, K.J., 1998. The disconnection hypothesis. Schizophr. Res. 30 (2), 115–125.Goldberg, J.A., Boraud, T., Maraton, S., Haber, S.N., Vaadia, E., Bergman, H., 2002. Enhanced
synchrony among primary motor cortex neurons in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine primate model of Parkinson's disease. J. Neurosci. 22(11), 4639–4653.
Grafton, S.T., 2004. Contributions of functional imaging to understanding Parkinsoniansymptoms. Curr. Opin. Neurobiol. 14 (6), 715–719.
Graybiel, A.M., 1997. The basal ganglia and cognitive pattern generators. Schizophr.Bull. 23 (3), 459–469.
Groenewegen, H.J., 2003. The basal ganglia and motor control. Neural Plast. 10 (1–2),107–120.
Habas, C., 2010. Functional connectivity of the human rostral and caudal cingulatemotor areas in the brain resting state at 3 T. Neuroradiology 52 (1), 47–59.
Harrison, P.J., 1999. The neuropathology of schizophrenia. A critical review of the dataand their interpretation. Brain 122 (Pt 4), 593–624.
Hasan, K.M., Parker, D.L., Alexander, A.L., 2001. Comparison of gradient encodingschemes for diffusion-tensor MRI. J. Magn. Reson. Imaging 13 (5), 769–780.
Jones, D.K., Knosche, T.R., Turner, R., 2012. White matter integrity, fiber count, andother fallacies: the do's and don'ts of diffusion MRI. Neuroimage. http://dx.doi.org/10.1016/j.neuroimage.2012.06.081.
Kanaan, R., Barker, G., Brammer, M., Giampietro, V., Shergill, S., Woolley, J., Picchioni, M.,Toulopoulou, T., McGuire, P., 2009. White matter microstructure in schizophrenia:effects of disorder, duration and medication. Br. J. Psychiatry 194 (3), 236–242.
Kay, S.R., Fiszbein, A., Opler, L.A., 1987. The positive and negative syndrome scale(PANSS) for schizophrenia. Schizophr. Bull. 13 (2), 261–276.
Kerns, J.G., Cohen, J.D., MacDonald III, A.W., Johnson, M.K., Stenger, V.A., Aizenstein, H.,Carter, C.S., 2005. Decreased conflict- and error-related activity in the anterior cingu-late cortex in subjects with schizophrenia. Am. J. Psychiatry 162 (10), 1833–1839.
Kim, D.J., Kim, J.J., Park, J.Y., Lee, S.Y., Kim, J., Kim, I.Y., Kim, S.I., Park, H.J., 2008. Quantifi-cation of thalamocortical tracts in schizophrenia on probabilistic maps. NeuroReport19 (4), 399–403.
Klöppel, S., Draganski, B., Golding, C.V., Chu, C., Nagy, Z., Cook, P.A., Hicks, S.L., Kennard,C., Alexander, D.C., Parker, G.J., Tabrizi, S.J., Frackowiak, R.S., 2008. White matterconnections reflect changes in voluntary-guided saccades in pre-symptomaticHuntington's disease. Brain 131 (Pt 1), 196–204.
Koning, J.P., Tenback, D.E., vanOs, J., Aleman, A., Kahn, R.S., vanHarten, P.N., 2010. Dyskinesiaand parkinsonism in antipsychotic-naive patients with schizophrenia, first-degree rela-tives and healthy controls: a meta-analysis. Schizophr. Bull. 36 (4), 723–731.
Kreher, B.W., Schnell, S., Mader, I., Il'yasov, K.A., Hennig, J., Kiselev, V.G., Saur, D.,2008. Connecting and merging fibres: pathway extraction by combining proba-bility maps. Neuroimage 43 (1), 81–89.
Kubicki, M., Alvarado, J.L., Westin, C.F., Tate, D.F., Markant, D., Terry, D.P., Whitford, T.J.,De Siebenthal, J., Bouix, S., McCarley, R.W., Kikinis, R., Shenton, M.E., 2011. Stochas-tic tractography study of Inferior Frontal Gyrus anatomical connectivity in schizo-phrenia. Neuroimage 55 (4), 1657–1664.
Kyriakopoulos, M., Bargiotas, T., Barker, G.J., Frangou, S., 2008. Diffusion tensor imagingin schizophrenia. Eur. Psychiatry 23 (4), 255–273.
Liddle, P.F., Friston, K.J., Frith, C.D., Hirsch, S.R., Jones, T., Frackowiak, R.S., 1992. Patternsof cerebral blood flow in schizophrenia. Br. J. Psychiatry 160, 179–186.
Maldjian, J.A., Laurienti, P.J., Kraft, R.A., Burdette, J.H., 2003. An automated method forneuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets.Neuroimage 19 (3), 1233–1239.
Marenco, S., Stein, J.L., Savostyanova, A.A., Sambataro, F., Tan, H.Y., Goldman, A.L., Verchinski,B.A., Barnett, A.S., Dickinson, D., Apud, J.A., Callicott, J.H., Meyer-Lindenberg, A.,

276 T. Bracht et al. / Schizophrenia Research 143 (2013) 269–276
Weinberger, D.R., 2012. Investigation of anatomical thalamo-cortical connectivity andFMRI activation in schizophrenia. Neuropsychopharmacology 37 (2), 499–507.
McCreadie, R.G., Padmavati, R., Thara, R., Srinivasan, T.N., 2002. Spontaneous dyskinesiaand parkinsonism in never-medicated, chronically ill patients with schizophrenia:18-month follow-up. Br. J. Psychiatry 181, 135–137.
McCreadie, R.G., Srinivasan, T.N., Padmavati, R., Thara, R., 2005. Extrapyramidal symp-toms in unmedicated schizophrenia. J. Psychiatr. Res. 39 (3), 261–266.
Middelkoop, H.A., van Dam, E.M., Smilde-van den Doel, D.A., Van Dijk, G., 1997. 45-hourcontinuous quintuple-site actimetry: relations between trunk and limb movementsand effects of circadian sleep-wake rhythmicity. Psychophysiology 34 (2), 199–203.
Morrens, M., Hulstijn, W., Sabbe, B., 2007. Psychomotor slowing in schizophrenia.Schizophr. Bull. 33 (4), 1038–1053.
Nieuwenhuys, R., Voogd, J., van Huizen, C., 2008. The Human Central Nervous System,Fourth edition. Springer.
Parker, G.J., Haroon, H.A., Wheeler-Kingshott, C.A., 2003. A framework for a streamline-based probabilistic index of connectivity (PICo) using a structural interpretation ofMRI diffusion measurements. J. Magn. Reson. Imaging 18 (2), 242–254.
Payoux, P., Boulanouar, K., Sarramon, C., Fabre, N., Descombes, S., Galitsky, M., Thalamas,C., Brefel-Courbon, C., Sabatini, U., Manelfe, C., Chollet, F., Schmitt, L., Rascol, O., 2004.Cortical motor activation in akinetic schizophrenic patients: a pilot functional MRIstudy. Mov. Disord. 19 (1), 83–90.
Payoux, P., Brefel-Courbon, C., Ory-Magne, F., Regragui, W., Thalamas, C., Balduyck, S.,Durif, F., Azulay, J.P., Tison, F., Blin, O., Esquerre, J.P., Rascol, O., 2010. Motor activa-tion in multiple system atrophy and Parkinson disease: a PET study. Neurology 75(13), 1174–1180.
Peralta, V., Campos, M.S., De Jalon, E.G., Cuesta, M.J., 2010. Motor behavior abnormali-ties in drug-naive patients with schizophrenia spectrum disorders. Mov. Disord. 25(8), 1068–1076.
Polli, F.E., Barton, J.J., Thakkar, K.N., Greve, D.N., Goff, D.C., Rauch, S.L., Manoach, D.S.,2008. Reduced error-related activation in two anterior cingulate circuits is relatedto impaired performance in schizophrenia. Brain 131 (Pt 4), 971–986.
Price, G., Cercignani, M., Parker, G.J., Altmann, D.R., Barnes, T.R., Barker, G.J., Joyce, E.M.,Ron, M.A., 2007. Abnormal brain connectivity in first-episode psychosis: a diffusionMRI tractography study of the corpus callosum. Neuroimage 35 (2), 458–466.
Saur, D., Kreher, B.W., Schnell, S., Kummerer, D., Kellmeyer, P., Vry, M.S., Umarova, R.,Musso, M., Glauche, V., Abel, S., Huber, W., Rijntjes, M., Hennig, J., Weiller, C.,2008. Ventral and dorsal pathways for language. Proc. Natl. Acad. Sci. U. S. A.105 (46), 18035–18040.
Scheuerecker, J., Ufer, S., Kapernick, M., Wiesmann, M., Bruckmann, H., Kraft, E., Seifert,D., Koutsouleris, N., Moller, H.J., Meisenzahl, E.M., 2009. Cerebral network deficitsin post-acute catatonic schizophrenic patients measured by fMRI. J. Psychiatr.Res. 43 (6), 607–614.
Schröder, J., Wenz, F., Schad, L.R., Baudendistel, K., Knopp, M.V., 1995. Sensorimotorcortex and supplementary motor area changes in schizophrenia. A study withfunctional magnetic resonance imaging. Br. J. Psychiatry 167 (2), 197–201.
Thomann, P.A., Wustenberg, T., Santos, V.D., Bachmann, S., Essig, M., Schröder, J., 2009.Neurological soft signs and brain morphology in first-episode schizophrenia.Psychol. Med. 39 (3), 371–379.
Vogt, B., 2009. Cingulate Neurobiology and Disease. Oxford University Press.Walker, E.F., Savoie, T., Davis, D., 1994. Neuromotor precursors of schizophrenia. Schizophr.
Bull. 20 (3), 441–451.Walther, S., Strik, W., 2012. Motor symptoms and schizophrenia. Neuropsychobiology
66 (2), 77–92.Walther, S., Horn, H., Razavi, N., Koschorke, P., Müller, T.J., Strik, W., 2009a. Quantitative
motor activity differentiates schizophrenia subtypes. Neuropsychobiology 60 (2),80–86.
Walther, S., Koschorke, P., Horn, H., Strik, W., 2009b. Objectively measured motor activ-ity in schizophrenia challenges the validity of expert ratings. Psychiatry Res. 169(3), 187–190.
Walther, S., Federspiel, A., Horn, H., Wiest, R., Dierks, T., Strik, W., Müller, T.J., 2010.White matter integrity associated with volitional motor activity. NeuroReport 21(5), 381–385.
Walther, S., Federspiel, A., Horn, H., Razavi, N., Wiest, R., Dierks, T., Strik, W., Müller, T.J.,2011a. Alterations of white matter integrity related to motor activity in schizo-phrenia. Neurobiol. Dis. 42 (3), 276–283.
Walther, S., Federspiel, A., Horn, H., Razavi, N., Wiest, R., Dierks, T., Strik, W., Müller, T.J.,2011b. Resting state cerebral blood flow and objective motor activity reveal basalganglia dysfunction in schizophrenia. Psychiatry Res. 192 (2), 117–124.
Wang, Q., Deng, W., Huang, C., Li, M., Ma, X., Wang, Y., Jiang, L., Lui, S., Huang, X., Chua,S.E., Cheung, C., McAlonan, G.M., Sham, P.C., Murray, R.M., Collier, D.A., Gong, Q., Li,T., 2011. Abnormalities in connectivity of white-matter tracts in patients withfamilial and non-familial schizophrenia. Psychol. Med. 41 (8), 1691–1700.
Whitty, P.F., Owoeye, O., Waddington, J.L., 2009. Neurological signs and involuntarymovements in schizophrenia: intrinsic to and informative on systems pathobiology.Schizophr. Bull. 35 (2), 415–424.
Wichniak, A., Skowerska, A., Chojnacka-Wojtowicz, J., Taflinski, T., Wierzbicka, A.,Jernajczyk, W., Jarema, M., 2011. Actigraphic monitoring of activity and rest inschizophrenic patients treated with olanzapine or risperidone. J. Psychiatr. Res.45 (10), 1381–1386.
Woods, S.W., 2003. Chlorpromazine equivalent doses for the newer atypical antipsy-chotics. J. Clin. Psychiatry 64 (6), 663–667.
Zalesky, A., Fornito, A., Seal, M.L., Cocchi, L., Westin, C.F., Bullmore, E.T., Egan, G.F., Pantelis,C., 2011. Disrupted axonal fiber connectivity in schizophrenia. Biol. Psychiatry 69 (1),80–89.





























