Embed Size (px)
Citation preview

BIOLBIOLBIOLBIOLBIOLOGIAOGIAOGIAOGIAOGIAAMBIENTAMBIENTAMBIENTAMBIENTAMBIENTALEALEALEALEALE
CentroItalianoStudi diBiologiaAmbientale
ISSN 1129-504X
Volume 18
Numero 2
Dicembre 2004

Tipografia NUOVA FUTURGRAF, Via Soglia 1, REGGIO EMILIA
BIOLBIOLBIOLBIOLBIOLOGIAOGIAOGIAOGIAOGIAAMBIENTAMBIENTAMBIENTAMBIENTAMBIENTALEALEALEALEALE
PROPRIETÀ: Rossella Azzoni, Presidente del C.I.S.B.A.
DIRETTORE RESPONSABILE: Rossella AzzoniREDAZIONE:Giuseppe Sansoni [email protected] resp. di redazioneRoberto Spaggiari [email protected] resp. di segreteriaGilberto N. Baldaccini [email protected] redattorePietro Genoni [email protected] redattore
Pubblicazione del C.I.S.B.A., vol. 18, n. 2/2004Autorizzazione del Tribunale di Reggio Emilia n. 837 del 14 maggio 1993
Biologia Ambientale, viene inviata ai soci del Centro Italiano Studi di Biologia Ambientale (C.I.S.B.A).Per iscriversi o per informazioni: Centro Italiano Studi di Biologia Ambientale, via Amendola 2, 42100 Reggio EmiliaSegretario: Roberto Spaggiari, tel. 0522 336060 – 0335 7712847; fax 0522 330546; e-mail: [email protected]
www.cisba.it [email protected] annuali di iscrizione al Centro Italiano Studi di Biologia Ambientale: socio ordinario: E 40,00; socio collaboratoreE 30,00; socio sostenitore E 310,00. conto corrente postale n. 10833424 intestato a: CISBA, RE
Biologia Ambientale raccoglie e diffonde in-formazioni sulle tematiche ambientali, con parti-colare attenzione ai seguenti campi di interesse:
Bioindicatori e biomonitoraggioEcotossicologiaDepurazione delle acque reflueEcologia delle acque interne e dell’ambientemarinoGestione dell’ambienteIgiene ambientaleEcologia urbanaImpatto ambientaleIngegneria naturalisticaRinaturazione e riqualificazione ambientaleConservazione della naturaEcologia del paesaggio
Biologia Ambientale è articolata in due sezioni:Lavori Originali, in cui vengono pubblicati arti-coli e rassegne bibliografiche originali;Informazione & Documentazione –sezione vol-ta a favorire la circolazione di informazioni e diidee tra i soci– in cui vengono riportate recensio-ni di libri, riviste e altre pubblicazioni nonché no-tizie e lavori già pubblicati ritenuti di particolareinteresse o attualità.
Comitato ScientificoRoberto ANTONIETTI
Dip. Scienze Ambientali, Univ. di ParmaNatale Emilio BALDACCINI
Dip. di Etologia, Ecologia, Evoluzione, Univ. di PisaRoberto BARGAGLI
Dip. Scienze Ambientali, Univ. di SienaAntonio DELL’UOMO
Dip. di Botanica ed Ecologia, Univ. di CamerinoSilvana GALASSI
Università dell’Insubria, ComoPier Francesco GHETTI
Dip. Scienze Ambientali, Univ. Cà Foscari, VeneziaStefano LOPPI
Dip. Scienze Ambientali, Univ. di SienaSergio MALCEVSCHI
Ist. Ecologia del territorio e degli ambienti terrestri,Univ. di Pavia
Maurizio G. PAOLETTIDip. di Biologia, Univ. di Padova
Luciano SANTINIDip. C.D.S.L. Sez. Entomologia agraria, Univ. di Pisa
Paolo Emilio TOMEIDip. Agronomia e gestione agroecosistema, Univ. di Pisa
Mariagrazia VALCUVIA PASSADOREDip. Ecologia del territorio e degli ambienti terrestri,Univ. di Pavia
Pierluigi VIAROLIDip. Scienze Ambientali, Univ. di Parma
Luigi VIGANÓIRSA - CNR, Brugherio MI
Sergio ZERUNIANParco Nazionale del Circeo, Sabaudia (LT)
Aldo ZULLINIDip. di Biotecnologie e Bioscienze, Univ. Milano Bicocca

BIOLBIOLBIOLBIOLBIOLOGIAOGIAOGIAOGIAOGIAAMBIENTAMBIENTAMBIENTAMBIENTAMBIENTALEALEALEALEALE Volume 18
Numero 2Dicembre 2004
SOMMARIO
3
11
19
25
LAVORI ORIGINALI
INFORMAZIONE & DOCUMENTAZIONE
FERRARINI A., ROSSI P., MIGLIAZZI M., ROSSI O., GIAVELLI G. - L’analisi da remoto delpaesaggio mediante finestre mobili: il caso di studio della Val Baganza(Parma)
SFRISO A., FACCA C., CEOLDO S., GHETTI P.F. - Variazioni delle concentrazioni dinutrienti e dei flussi di sedimentazione nella parte centrale della laguna diVenezia
FACCA C., SFRISO A., GHETTI P.F. - Abbondanza e diversità del fitoplancton e dellediatomee bentoniche in laguna di Venezia
ZERUNIAN S. - Proposta di un Indice dello Stato Ecologico delle Comunità Itticheviventi nelle acque interne italiane
GAITER S., BODON M., ROCCA D. - Applicazione della metodica di indagine biologi-ca, basata sulla fauna bentonica e sulla componente organica del sedimen-to, in acque di sorgente ed emergenze assimilabili
PICCININI A. - Il Siluro tra mito e realtà
Rassegna scientifica (a cura di P. Genoni)
Recensioni
31
48
53
72


Biologia Ambientale, 18 (2): 3-10, 2004
L’analisi da remoto del paesaggio mediante finestremobili: il caso di studio della Val Baganza (Parma)Alessandro Ferrarini*, Pierfrancesca Rossi, Mauro Migliazzi, Orazio Rossi, Giovanni Giavelli
Dipartimento di Scienze Ambientali, Università degli Studi di Parma, Viale delle Scienze 11/A - 43100 Parma
* Autore referente per la corrispondenza: [email protected]; [email protected]
Pervenuto il 24.11.2003; accettato il 3.3.2004
RiassuntoLa geometria non euclidea amplia la capacità di descrizione della natura, permettendo l’analisi anche di oggetti e strutture irregolari. Per unpaesaggio dalla forte impronta antropica, dove linee diritte e figure regolari sono state create trasformando aree naturali in zoneurbanizzate o rurali è sufficiente l’utilizzo degli strumenti euclidei per descriverne i semplici motivi spaziali. Al contrario, se si consideraun paesaggio più naturale, prevale l’irregolarità delle forme degli habitat e l’approccio frattale risulta il più idoneo e soddisfacente per ladescrizione e lo studio del territorio. I frattali sono oggetti di dimensione non intera, la cui struttura geometrica è così complicata da nonpermettere di attribuire loro una dimensione geometrica di tipo euclideo.In questo lavoro vengono analizzate le forme di alcune tipologie di habitat presenti nel bacino idrografico del torrente Baganza mediantel’utilizzo della metodologia di Krummel delle finestre mobili applicata ad immagini telerilevate Landsat. I risultati vengono discussi intermini di gestione e conservazione del territorio.
PAROLE CHIAVE: paesaggio / telerilevamento / analisi a finestre mobili / forma degli habitat
Remotely sensed landscape moving window analysis: the case study of the Baganza stream watershed (Parma, Italy)The not euclidean geometry widens the ability of nature description, allowing the analysis both of regular and irregular structures. Letconsider a landscape with a strong anthropic imprint, where right lines and regular figures have been created transforming natural areas indeveloped regions; in this case it will be sufficient the use of euclidean tools to describe the spatial patterns through the perimeter-arearelationship, the area of an habitat or its distance from other habitats. On the contrary, if a more natural landscape is considered, then theirregularity of the shapes largely prevails. Fractals are objects whose geometric shape is so complex and distant from the simplicity ofeuclidean features, that don’t allow to attribute them a “whole” dimension. The fractal approach results the most satisfactory for thedescription and the study of the landcover.In this paper, the shapes of some kinds of habitats inside the Baganza stream watershed have been analyzed by means of Krummel’smoving window method applied to Landsat satellite images. The results have been inserted into a GIS and then discuss in terms oflandscape management and conservation.
KEY WORDS: landscape / remote sensing / moving window analysis / habitat shape
INTRODUZIONEL’ecologia del paesaggio ha ormai dimostrato nu-
merose proprietà ecologico-ambientali della forma de-gli habitat presenti sul territorio. Alcune di queste sono:a) le forme degli habitat rappresentano il risultato di
processi che operano su tempi lunghi (FORMAN eGODRON , 1986; FORMAN, 1995, HAMMETT, 1992);
b) le forme degli habitat rappresentano la risultante di un
bilancio tra forze interne all’habitat e forze esternedella matrice adiacente (FORMAN, 1995, FOSTER, 1983);
c) la forma incide sui processi tra gli habitat, condizio-nando la migrazione di piccoli mammiferi (BUECHNER,1989), la colonizzazione delle piante (HARDT e FORMAN
1989), le strategie di foraggiamento degli animali(FORMAN e GODRON,1986);

FERRARINI et al. - Analisi a finestre mobili del paesaggio4
d) ci sono tre principi, cosiddetti di “forma e funzione”(THOMPSON, 1961; FORMAN, 1995), che risultano, ingenere, rispettati:– le forme compatte sono utili per conservare le
risorse in quanto viene minimizzato il perimetroesposto all’esterno
– le forme perimetralmente complesse sono utili nelfavorire le interazioni con l’intorno poiché vienemassimizzata la probabilità di attraversamento delconfine dell’habitat
– le forme disposte come un labirinto tendono arappresentare un sistema di trasporto trasmetten-do l’informazione da una porzione all’altra delmosaico degli habitat;
e) le forme degli habitat variano soprattutto con latopografia e con l’intensità dell’attività umana. Ingenere, in pianura gli habitat hanno forme compattee poco variabili e confini regolari. In collina gli habitathanno forme intermedie e confini meno regolari. Inmontagna, nelle zone più naturali e a maggiorependenza, le forme sono più complesse perimetral-mente e lunghe e la variabilità di forme è moltoelevata. Esistono anche molte forme miste che com-binano gli effetti di meccanismi umani e naturali;
f) gli habitat grandi sono caratterizzati da condizioniproprie della zona interna (interior) e quindi anche daspecie dell’habitat interno che non sono presenti neglihabitat piccoli (FORMAN, 1981);
g) processi ad alta entropia e casualità danno luogo aforme casuali caratterizzate soprattutto da convolu-zione e da perimetri aspri. Forze direzionali, qualivento, acqua, ghiaccio tendono ad allungare gradual-mente gli habitat. L’effetto netto di queste forze èquello di attendersi forme allungate, con molti lobi eperimetri aspri, poiché è necessaria una grandequantità di energia per mantenere forme circolari conperimetri regolari (FORMAN, 1995);
h) la forma di un habitat può essere descritta mediantequattro attributi– area– perimetro– elongazione (allungamento in una direzione prefe-
renziale)– complessità dei bordi con presenza di molti lobi
i) gli habitat il cui perimetro presenta molti lobi hanno,in generale, ampi scambi con la matrice esterna e unasuddivisione di popolazioni in sottopopolazioni cheproduce una maggiore variazione genetica rispetto adun habitat circolare dove il flusso genico è piùdiffuso; infine producono l’incanalamento degli ani-mali da un’area ampia (matrice) in un’altra strettacome un lobo.Il presente contributo si focalizza sulla complessità
perimetrale degli habitat, espressione di processi natu-
rali e/o antropici, mediante l’applicazione del metododella finestra mobile proposto da Krummel (KRUMMELet al., 1987).
AREA DI STUDIO, MATERIALI E METODIL’area di studio coincide con il bacino idrografico
del Torrente Baganza (Fig. 1) posto nel versante emi-liano dell’Appennino settentrionale nella provincia diParma. Nella zona pianeggiante, la vegetazione natura-le è limitata alle aree soggette a minor pressione antro-pica in prossimità del greto del torrente, con piopponero e robinia. Nell’area collinare è presente la vegeta-zione submediterranea con boschi misti a roverella suiversanti caldi e secchi e a carpino nero in condizionipiù mesofile. Nell’area submontana, che si estendefino alla quota di circa 1000 m s.l.m., sono frequentiprati da sfalcio, cerrete sui versanti meno acclivi edostrieti su quelli a prevalente esposizione nord. Infine,alle quote più elevate, dominano pressoché incontra-stati i boschi neutrofili a faggio, con zone a castagno,oggi non più curato. Le formazioni erbacee sono per lopiù rappresentate da praterie a Brachypodium genuen-se (ROSSI, 1999). L’attività umana si concentra neinuclei urbani limitrofi al capoluogo di provincia. L’areaplaniziale e di prima collina è dominata da un’agricoltu-ra intensiva legata alle necessità produttive della filieraagro-alimentare del parmense. La zona montana risen-te, al contrario, di un ventennale processo di spopola-
Fig. 1. Posizione del bacino idrografico del Torrente Baganza(18000 ettari, prov. Parma).
Val Baganza

FERRARINI et al. - Analisi a finestre mobili del paesaggio 5
mento ed invecchiamento, con un progressivo abban-dono della presenza umana sul territorio. La conserva-zione di prati a sfalcio per la produzione a foraggiera haun qualche impatto estensivo, mentre scarso è losfruttamento delle risorse forestali nel settore dellalavorazione del legno.
Per indagare l’area di studio, è stata utilizzata unascena Landsat TM5 del 24-7-95. Integrando il datosatellitare con ricognizioni in situ (ROSSI, 1999) è statarealizzata entro il Progetto Carta della Natura (ROSSI,2001; ROSSI et al., 2002) la mappatura degli habitat delbacino idrografico del Torrente Baganza su base CO-RINE Biotopes dell’UE (C.E.C., 1991) e relativa al-l’anno 1999. Tale mappatura è stata successivamenteaggiornata nell’anno 2000 (TOMASELLI, 2000; nonpubblicato). L’analisi frattale è stata applicata a seitipologie di habitat CORINE caratteristiche della ValBaganza:a) prati da sfalcio submontani e montani (codice CORI-
NE: 38.2). Si tratta di prati da sfalcio mesofili,fertilizzati e ben drenati, con Arrenatherum elatius,Trisetum flavescens, Bromus hordeaceus, Poa pra-tensis, Galium album;
b) boschi di cerro (codice CORINE: 41.74). Si tratta diboschi a dominanza di cerro (Quercus cerris), conOstrya carpinifolia, Fraxinus ornus, Quercus pube-scens, maggiociondolo (Laburnum anagyroides) epioppo tremolo (Populus tremula). Lo strato arbusti-vo è caratterizzato dalla predominanza di Corylusavellana; un’altra specie arbustiva relativamentefrequente è Rosa arvensis. Nello strato erbaceopredominano specie mesofile quali Primula vulgaris,Hepatica nobilis, Listera ovata e Geranium nodo-sum. Sono diffusi nella fascia submontana in stazionifresche con suoli da basici a subacidi;
c) boschi di carpino nero e supramediterranei (codiceCORINE: 41.812). Sono boschi a dominanza dicarpino nero, con orniello, querce, aceri (Acer opu-lifolim, Acer campestre), e sottobosco di essenzemesofile. Sono diffusi prevalentemente sui versantifreschi della fascia supramediterranea. Le speciediagnostiche principali sono: Acer opulifolium, He-patica nobilis, Lilium croceum;
d) cespuglieti di tipo medio-europeo (codice CORINE:31.81). Si tratta di mantelli di bosco o cespuglietisecondari composti da Prunus spinosa, Crataegusmonogyna, Rosa canina, Cornus sanguinea. Sonodiffusi dalla fascia supramediterranea calda a quellasubmontana;
e) praterie semiaride appenniniche (codice CORINE:34.3266). Sono praterie chiuse da moderatamentearide a semimesofile, di suoli relativamente profondie per lo più carbonatici, caratterizzate dalla dominan-za di erbe perenni, come Bromus erectus, Brachypo-
dium rupestre, Ranunculus bulbosus, Sanguisorbaminor, diverse leguminose (Lotus corniculatus, Me-dicago lupulina, Hippocrepis comosa) e orchidee(Orchis morio, Anacamptis pyramidalis, Gymnade-nia conopsea, Ophrys apifera). Sono diffuse nellefasce supramediterranea e submontana;
f) praterie a Brachypodium genuense (codice CORINE:36.334). Sono praterie chiuse a dominanza di Bra-chypodium genuense limitate alla fascia montana. Lespecie accompagnatrici più frequenti sono: Luzulamultiflora, Anthoxanthum alpinum, Avenella flexuo-sa.Rispetto all’approccio classico euclideo (ZURLINI et
al., 2001a), la geometria frattale permette una descri-zione più realistica della forma degli oggetti naturali.Questi ultimi, poiché non interessati dalle attività uma-ne, mostrano forme molto irregolari. Il grado di com-plessità di queste forme può essere espresso in terminidimensione frattale perimetrale (D). Per i singoli habi-tat di un mosaico D è data da (NIKORA et al., 1999):(1) D = α · (1+H)dove α è l’esponente nella relazione perimetro-area(MANDELBROT, 1983), P è il perimetro, A l’area e c unacostante. H è l’esponente di Hurst detto «shape expo-nent» nella relazione area-lunghezza (MANDELBROT,1983):(2) A = b · L(1+H)dove b è una costante. In particolare, H è uguale a 1quando gli habitat a scale differenti sono geometrica-menti molto simili (self-similar). Qualora si verifichiquesta condizione (MILNE, 1991; HASTINGS e SUGIHA-RA, 1993), si ha che:(3) α = D/2con α che fornisce una stima diretta di D dalle relazio-ni empiriche perimetro-area (LOVEJOY, 1982) entroogni dominio scalare. Dati l’area ed il perimetro di nhabitat irregolari, la relazione perimetro-area è:(4) Pi = c · Ai
α
dove Pi e Ai sono rispettivamente il perimetro e l’areadell’habitat i-esimo. Passando alla trasformazione lo-garitmica si ottiene:(5) ln(Pi) = ln(c) + α · ln(Ai)dove D è il doppio del coefficiente angolare α delmodello di regressione lineare. D è vincolato ad unpiano ed assume valori prossimi od uguali ad 1 performe euclidee semplici (cerchi e rettangoli) ed au-menta fino a 2 per quelle più complesse. Un salto nelladimensione frattale perimetrale, o nei parametri frattaliconnessi, potrebbe essere indice di un cambiamentosostanziale nei processi che generano e mantengonogli habitat del paesaggio alle differenti scale (KRUMMELet al., 1987; MILNE, 1991).
Il metodo di Krummel (KRUMMEL et al., 1987),studia la relazione tra area e perimetro degli habitat

FERRARINI et al. - Analisi a finestre mobili del paesaggio6
mediante una regressione a finestra mobile che eviden-zia l’eventuale esistenza di punti di rottura nei parame-tri di forma. Il metodo può essere così schematizzato:1) siano dati n habitat ognuno con perimetro Pi e area
Ai, 1 ≤ i ≤ n2) si attua la trasformata logaritmica dell’area e del
perimetro degli n habitat3) sia data una finestra iniziale di dimensioni m4) si calcola la regressione tra P ed A per i primi m
habitat5) la finestra si sposta di una osservazione6) si calcola la regressione tra P ed A per gli habitat
da 2 ad m+17) la finestra si sposta di una osservazione8) si calcola la regressione tra P ed A per gli habitat
da 3 ad m+29) la finestra mobile si ferma quando i + m = n, dove
i è l’i-esimo habitat da cui parte la regressione10) la dimensione della finestra viene aumentata da m
ad m2= m + k, con k a piacere11) si riprende dal punto 4) con la sola differenza che
m è sostituito da m212) l’iterazione del processo avviene un numero di
volte pari a g = [(n – m) / k] + 1I break point individuati dividono le osservazioni
analizzate in più intervalli; ciascun intervallo è chiama-to dominio frattale ed è caratterizzato da un coefficien-te di regressione D. Individuati i punti di rottura nellaregressione area-perimetro, vengono calcolati i valoridi ln(Ai) corrispondenti. La differenza tra i coefficientidi regressione dei diversi domini funzionali tra P ed A èindice dell’efficacia del metodo, una variazione anchese lieve è sufficiente a dimostrare la validità del metodo.
L’intero processo è piuttosto oneroso dal punto divista computazionale. Per tale motivo abbiamo realiz-zato un applicativo in linguaggio Visual Basic chiama-to “Rolling Regression Software” (MIGLIAZZI et al.,2002). Il programma (che può essere richiesto alprimo autore del presente lavoro) realizza tutti i pas-saggi sopra descritti e restituisce i risultati in un forma-to immediatamente importabile entro un GIS per unarappresentazione cartografica dei domini frattali indivi-duati. Il software richiede in input tre parametri:a) number of observations (n): indica il numero di
osservazioni (habitat) su cui operareb) first lag (m): indica la dimensione del primo intervallo
Fig. 2 Algoritmo utilizzato nell’individuazione dei domini frattali per le sei tipologie di habitat CORINE analizzati. Sulla sinistra,l’immagine satellitare Landsat TM5 dell’area di studio.
un

FERRARINI et al. - Analisi a finestre mobili del paesaggio 7
per il calcolo della regressione.c) increment (k): indica l’incremento cioè il valore che
sarà sommato ad m ad ogni regressioneL’algoritmo utilizzato per la determinazione dei do-
mini frattali viene, in sintesi, illustrato nella figura 2.
RISULTATINell’area di studio le tipologie CORINE considerate
sono così quantificate:a) codice 31.81: 302 habitat;b) codice 34.3266: 84 habitat;c) codice 36.334: 129 habitat;d) codice 38.2: 375 habitat;e) codice 41.74: 32 habitat;f) codice 41.812: 65 habitat.Relativamente al codice CORINE 31.81, il punto di
rottura della relazione area-perimetro si ha per ln(A)=9,13 ovvero A=9228 mq. Si possono, quindi, calcolarei coefficienti di regressione per i 2 intervalli scalari:
D1 = 1,15D2 = 1,36
La differenza tra i due coefficienti di regressione è0,21. Questo significa che, relativamente al codiceCORINE 31.81, gli habitat con un valore areale infe-riore a 9228 mq presentano una relazione area-perime-tro molto più semplice rispetto a quelli con superficiemaggiore di quel valore. È quindi possibile suddividerei 302 habitat di questo codice in due cluster, il primocaratterizzato da forme mediamente semplificate e com-patte, il secondo da forme mediamente complesse(Fig. 3).
Relativamente al codice CORINE 34.3266, il puntodi rottura della relazione area-perimetro si ha per ln(A)=9,05 cioè A=8518 mq. Si possono calcolare i coeffi-cienti:
D1 = 1,14D2 = 1,31
La differenza tra i due coefficienti di regressione è0,17. Per il codice 36.334, il punto di rottura si ha perln(A)= 9,21 (Fig. 4 e Fig. 5) cioè A= 9996 mq. Icoefficienti di regressione per i 2 intervalli scalari sonorispettivamente:
D1 = 1,23D2 = 1,58
La differenza tra i due coefficienti di regressione è0,35. Per il codice 38.2, il punto di rottura si ha perln(A)= 9,32 cioè A= 11158 mq. Si possono calcolare icoefficienti di regressione:
D1 = 1,11D2 = 1,39
La differenza tra i due coefficienti di regressione è0,28. Per il codice CORINE 41.74 non è stato possibi-le rilevare alcun punto di rottura nella relazione area-perimetro. Il codice 41.812 ha un punto di rottura per
Fig. 3 Esempio di un habitat appartenente al primo dominiofrattale (a) del Codice 31.81 e di un habitat del secondo dominiosempre del Codice 31.81 (b). Si nota con nettezza la differenzanella complessità della forma (convoluzione).
Fig. 4 Rappresentazione per il codice CORINE 36.334 dellaregressione spezzata, risultante dall’applicazione del metodo diKrummel. Il punto di rottura (evidenziato dal circolo) èposizionato in ln(A)= 9,21. La dimensione frattale D1 del primogruppo di habitat è 1,23 (D1= 2 · 0,6158), mentre D2 è pari a 1,58(D2= 2 · 0,7927).
Fig. 5 Rappresentazione GIS dei 2 domini frattali del codiceCORINE 36.334: in grigio chiaro il primo dominio, in grigio scuroil secondo. Il primo dominio si trova esclusivamente a destra deltorrente Baganza dove le pendenze dolci permettono la presenzadell’uomo, mentre il secondo si trova quasi esclusivamente sulversante sinistro dove, per l’alta acclività, l’uomo è quasitotalmente assente.
7,4 7,6 7,8 8,0 8,2 8,4 8,6 8,8 9,0 9,2 9,4 9,6 9,8 10 10,2 10,4 10,6 10,8 11 11,2 11,4 11,6 11,8 12
ln (P)
Codice CORINE 36.334
ln (A)
0,6158 · x + 0,5339 x < 9,21y=0,7927 · x - 1,1566 x > 9,21
8,88,68,48,28,07,87,67,47,27,06,86,66,46,26,05,85,65,45,25,04,8

FERRARINI et al. - Analisi a finestre mobili del paesaggio8
ln(A)= 10,7 cioè A= 44355 mq. I coefficienti di re-gressione per i 2 intervalli scalari sono rispettivamente:
D1 = 1,23D2 = 1,66
La differenza tra i due coefficienti di regressione è0,43. I risultati ottenuti sono riportati nella tabella I.
DISCUSSIONEIl metodo di Krummel presenta alcuni evidenti punti
di forza. Si tratta di una metodologia concettualmentesemplice e relativamente intuitiva che prevede che illegame tra una variabile dipendente ed una indipenden-te possa non essere costante lungo tutto il dominiodella funzione. Essa permette inoltre di individuare glieventuali punti di rottura. Il metodo consente inoltre ditrovare punti di rottura anche su un campione minimodi osservazioni: nel caso del codice 41.812, è statotrovato un punto di rottura nonostante fossero presentisolo 65 osservazioni. Avvalendosi di esperienze empi-riche, si può porre uguale a 40 il numero di osservazio-ni sotto il quale il metodo non è in grado di trovarebreakpoint.
Il metodo presenta tuttavia alcuni limiti. Innanzituttoè un metodo semi-quantitativo e, a differenza di altremetodologie con finalità simili (GROSSI et al., 2001;ZURLINI et al., 2001b; ZURLINI et al., 2002), non esistealcun test che restituisca con esattezza il punto dirottura della relazione area-perimetro. I punti di shiftdella funzione vengono individuati per ispezione visua-le del biplot avente ln(A) sull’asse delle ascisse e D suquello delle ordinate. I punti di rottura possono essereindividuati solo per una certa dimensione della finestrache scorre sulle osservazioni: le altre finestre general-mente non sono in grado di rivelare i breakpointpresenti. Questo è il motivo per cui è necessario usareun numero elevato di finestre di dimensione variabile.La scelta della dimensione ottimale della finestra derivadal compromesso fra due opposte esigenze. Da unlato, la dimensione frattale deve essere calcolata su uninsieme di habitat relativamente omogenei: ovviamen-te, più piccola è la finestra, maggiormente omogeneisono gli habitat che ricadono al suo interno. Dall’altrolato, per ottenere stime statisticamente significativedella dimensione frattale, la finestra non deve esseretroppo piccola. La dimensione ottimale della finestraviene quindi ottenuta iterativamente partendo dalla di-mensione più piccola possibile ed incrementandolafinché non vengono prodotte stime significative delladimensione frattale. Per esempio, il ritrovamento deidomini frattali per le sei tipologie studiate ha richiestole seguenti dimensioni della finestra mobile: 245 osser-vazioni per il codice 31.81, 65 per il codice 34.3266,110 per il codice 36.334, 305 per il codice 38.2, 60 peril codice 41.812. Non esiste una regola, né teorica né
empirica, per determinare la dimensione ottimale di talefinestra, se non che essa deve essere maggiore di 30osservazioni affinché la regressione calcolata sia signi-ficativa. Inoltre, l’incremento k deve essere scelto conla sola accortezza che m+k sia minore del numerototale n delle osservazioni.
L’ispezione visuale degli habitat appartenenti a di-versi domini frattali evidenzia che effettivamente cam-bia in modo netto il grado di convoluzione (Fig. 3 eFig. 5). Da qui il significato ecologico delle analisirealizzate: suddividere l’intero range degli habitat ap-partenenti ad una certa tipologia CORINE in due o piùcluster a cui, a parità di complessità morfologica (do-minio frattale), corrispondano processi naturali/antro-pici simili che creano e mantengono quel gruppo dihabitat. Un salto di dominio frattale lascia, viceversa,supporre che cambino i processi dominanti entro o alconfine degli habitat.
Sotto l’ipotesi che a gradi di complessità differenticorrisponda una differente embriologia e diversi pro-cessi dominanti negli ecosistemi, sono in corso ulterio-ri analisi per saggiare questa ipotesi. Ciò permetterebbeanche l’individuazione delle variabili ecologiche e/oantropogeniche che possono effettivamente spiegare isalti di dominio scalare rinvenuti. Particolarmente utilerisulta l’uso congiunto di questa metodologia e delletecniche di telerilevamento come strumento d’indaginedel territorio (FERRARINI et al., 2002a; FERRARINI et al.,2002b; FERRARINI et al., 2002c).
CONCLUSIONIIl metodo di Krummel individua gli eventuali punti di
rottura della funzione di regressione della relazionearea-perimetro degli habitat e, quindi, la presenza didue o più funzioni, in dipendenza dalla forma degli
Tab. I. Risultati dell’applicazione del metodo della finestramobile di Krummel ad alcuni codici CORINE presenti nel bacinoidrografico del torrente Baganza. La colonna D1 rappresenta ilcoefficiente di regressione del primo dominio frattale, D2 delsecondo. La colonna D2-D1 rappresenta il salto tra il primo e ilsecondo dominio frattale. Tanto maggiore è questo valore, tantomaggiore è la differenza nella complessità media delle formedegli habitat appartenenti ai due domini. È stato tralasciato ilcodice 41.74 per il quale non sono stati rinvenuti punti di rotturanella regressione perimetro-area.
Codice CORINE D1 D2 D2-D1
31.81 1,15 1,36 0,2134.3266 1,14 1,31 0,1736.334 1,23 1,58 0,3538.2 1,11 1,39 0,2841.812 1,23 1,66 0,43

FERRARINI et al. - Analisi a finestre mobili del paesaggio 9
habitat stessi.La suddivisione degli habitat in due o più gruppi
(domini) permette di avanzare alcune ipotesi pianifica-torie plausibili sulla loro conservazione. Per esempio,forme maggiormente regolari e compatte sono menosensibili rispetto alla matrice esterna, risentono menodell’influenza degli habitat adiacenti, i loro confini nettiagiscono maggiormente da barriere o da filtri. La loroconformazione minimizza l’esposizione dei confini equesto è molto utile al fine di una maggior conserva-zione delle risorse interne. Confini diritti facilitano,inoltre, il movimento dei predatori lungo gli stessi equesto rende l’habitat ancor più difficilmente penetra-bile da parte di specie estranee. Habitat circolari, ocomunque regolari, sono caratterizzati, in genere, daun flusso genico ristretto rispetto ad habitat convoluti,dove le popolazioni si suddividono in sottopopolazionilocalizzate nei diversi lobi dei lobi e dei loro microhabi-tat, e questo comporta una minor varietà di specie.
Gli habitat frastagliati e con molti lobi sono utili nelfavorire le interazioni con l’intorno, vi è una massimiz-zazione della probabilità di attraversamento del confinepoiché più lungo. I lobi originano un effetto imbuto,una concentrazione di specie in un piccolo spazio, chefacilita l’entrata e/o l’uscita dall’habitat. Molte speciepossono attraversare un habitat solo attraverso i lobi.Questi spazi hanno, inoltre condizioni microclimatiche
proprie e generano microambienti dove aumentano leprobabilità di nascondersi ed evitare predatori, mante-nendo in tal modo una diversità genetica elevata nel-l’habitat. Maggior interesse conservazionistico dovràin generale essere posto nei riguardi degli habitat con-voluti piuttosto che ad habitat compatti, difficilmenteperturbabili.
Nel presente lavoro, il metodo di Krummel è statoapplicato a sei tipologie di habitat CORINE la cuimappatura proviene da immagini telerilevate Landsat.Mediante l’utilizzo di un software realizzato apposita-mente per questo lavoro, è stato possibile individuarediversi domini frattali per ogni singola tipologia CORI-NE e riportare il tutto all’interno di un GIS. Ulterioriapprofondimenti dovranno spiegare l’appartenenza diun habitat ad un certo dominio frattale sulla base divariabili naturali e/o antropogeniche che, con riferi-mento alla letteratura scientifica disponibile, si ritienesiano in grado di influenzare e spiegare i punti dirottura della relazione area-perimetro. Tali breakpointsono determinanti ecologico-ambientali di fondamen-tale importanza per un’analisi sinottica del mosaicodegli habitat naturali mediante immagini telerilevate.
RingraziamentiSi ringraziano i referees anonimi per i consigli forniti.
BIBLIOGRAFIA
BUECHNER M., 1989. Are small-scale landscape features importantfactors for field studies of small mammal dispersal sinks?Landscape Ecology, 2: 191-199.
C.E.C. (COMMISSION OF EUROPEAN COMMUNITY), 1991. CORINEBiotopes Manual, habitats of the European Community. Amethod to identify and describe consistently sites of majorimportance for nature conservation. EUR 12587/3, Bruxelles.
FERRARINI A., ROSSI P., ZACCARELLI N., 2002a. Un classificatoreefficiente di dati iperspettrali MIVIS per la mappatura terri-toriale: CART (Classification And Regression Trees). RivistaItaliana di Telerilevamento, 22: 3-12.
FERRARINI A., ROSSI P., ROSSI O., SOLIANI L., 2002b. Metodiavanzati di analisi e progettazione territoriale. Notiziario dellaSocietà Italiana di Ecologia (S.It.E.), pp. 7-9.
FERRARINI A., ROSSI P., ZACCARELLI N., 2002c. Mappature vegeta-zionali ad elevata accuratezza: la metodologia delle reti neuraliapplicata ai dati iperspettrali MIVIS. Rivista Italiana di Tele-rilevamento, 22: 13-22.
FORMAN R.T.T., 1981. Interaction among landscape elements: acore of landscape ecology. In S. P., Tjallingii e A.A. de Veer,
eds. Perspectives in Landscapes, pp. 35-48, PUDOC, Wage-nigen, Netherlands.
FORMAN R.T.T., GODRON M., 1986. Landscape ecology. Whiley& Sons, New York.
FORMAN R.T.T., 1995. Land Mosaics. The Ecology of Ladscapesand Regions, Cambridge University Press.
FOSTER D.R., 1983. The history and pattern of fire in the borealfoster of southeastern Labrador. Canadian Journal of Botany,61: 2549-71.
GROSSI L., ZURLINI G., ROSSI O., 2001. Statistical detection ofmultiscale landscape patterns. Journal of Ecological andEnvironmetal Statistics, 8: 253-267.
HAMMETT J.E., 1992. The shapes of adaptation: historical ecologyof anthropogenic landscapes in the southeastern United States.Landscape Ecology, 7: 121-135.
HARDT, R.A., FORMAN, R.T.T., 1989. Boundary form effects onwoody colonization of reclaimed surface mines. Ecology, 70:1252-1260.
HASTINGS, H.M., SUGIHARA, G., 1993. Fractals - a user’s guide forthe natural sciences, Oxford University Press, New York.

FERRARINI et al. - Analisi a finestre mobili del paesaggio10
KRUMMEL J.R., GARDNER R.H., SUGIHARA G., O’NEILL R.V., COLE-MAN P.R., 1987. Landscape patterns in a disturbed environ-ment. Oikos, 48: 321-324.
LOVEJOY S., 1982. Area-perimeter relation for rain and cloud areas.Science, 216: 185-187.
MANDELBROT B.B., 1983. The Fractal Geometry of Nature. W. H.Freeman (ed.), San Francisco.
MIGLIAZZI M., ROSSI O. (relatore), FERRARINI A. (correlatore), ROSSI
P. (correlatore), ALLESINA S. (correlatore), 2002. Metodi avan-zati di analisi dell’ecomosaico ambientale mediante tecnichequantitative applicate alla base di dati di un Sistema Informa-tivo Territoriale. Tesi di laurea - Facoltà di Scienze Matema-tiche Fisiche e Naturali, Università di Parma.
MILNE B.T., 1991. Lessons from applying fractal models tolandscape patterns. In: Turner M.G. and.Gardner R.H (eds.)Quantitative Methods in Landscape Ecology. Springer-Verlag:199-235.
NIKORA V.I., PEARSON C.P., SHANKAR, U., 1999. Scaling propertiesin landscape patterns: New Zealand experience. LandscapeEcology, 14: 17-33.
ROSSI O., 2001. La Carta della Natura del Paese: aspetti generalie prospettive. In: Cartografia multiscalare della natura. S.It.E.- Atti XXIII, 11-20.
ROSSI O., ROSSI P., FERRARINI A., 2002. Principii e metodologie
generali per la Carta della Natura del Paese. In: Le Ofioliti: isolesulla terraferma. Per una rete di Aree Protette, Regione Emilia-Romagna, 235-242
ROSSI P., 1999. Analisi della diversità vegetazionale in Val Bagan-za mediante la classificazione CORINE Biotopes. Tesi diLaurea in Scienze Biologiche, Facoltà di Scienze MatematicheFisiche e Naturali, Università di Parma.
THOMPSON D.W., 1961. On Growth and Form. Cambridge Univer-sity Press, Cambridge.
ZURLINI G., ROSSI O., FERRARINI A., ROSSI P., ZACCARELLI N., 2001a.Risultati preliminari sulla relazione tra pattern spaziali dimosaici di habitat CORINE e tassi di disturbo ambientale.S.It.E. - Atti XXV.
ZURLINI G., ROSSI O., FERRARINI A., ROSSI P., ZACCARELLI N., 2001b.Assessing multi-scale fragility of landscapes: concepts, metho-ds and recent results of the Map of Italian Nature. In BelwardA., Binaghi E., Brivio P.A., Lanzarone G.A. e Tosi G. (eds.):Geospatial knowledge processing for natural resources ma-nagement, European Commission, JRC ISPRA, NationalResearch Council, Artestampa, Baveno (Varese), 153-159.
ZURLINI G., ROSSI O., ZACCARELLI N., PETROSILLO I., FERRARINI A.,ROSSI P., 2002. Spatial patterns of ecological change in water-shed. in: Acts of the annual meeting of International Associationfor Landscape Ecology (IALE), Lincoln, NE (USA), pp.73.

Biologia Ambientale, 18 (2): 11-18, 2004
Variazioni delle concentrazioni di nutrienti e dei flussi disedimentazione nella parte centrale della laguna di Venezia
Adriano Sfriso*, Chiara Facca, Sonia Ceoldo, Pier Francesco Ghetti
Dipartimento di Scienze Ambientali, Università di Venezia, Calle Larga 2137 - 30123 Venezia* Referente per la corrispondenza: Fax +39 041 2348584; [email protected]
Pervenuto il 7.8.2003; accettato il 29.1.2004
RiassuntoIn questo lavoro sono presentate le variazioni dello stato trofico e dei flussi di sedimentazione avvenute negli ultimi quindici anni in alcunestazioni della parte centrale della laguna di Venezia. Da giugno 2000 a giugno 2001 sono state analizzate le concentrazioni dei nutrientidisciolti nella colonna d’acqua, dei nutrienti totali presenti nei sedimenti superficiali, nel particellato e nei tessuti di alcune macrofite e iflussi di sedimentazione. I dati ottenuti sono stati confrontati con quelli rilevati dal 1984 al 1998.Nella colonna d’acqua le concentrazioni dei nutrienti, soprattutto dell’azoto inorganico disciolto, presentano una marcata variabilitàinterannuale. Nel sedimento superficiale (orizzonte 0-5 cm), dal 1984 al 2001, le concentrazioni di azoto e fosforo sono diminuite, spessopiù che dimezzate. Dall’analisi delle concentrazioni tissutali dei nutrienti, Ulva rigida C. Ag. e Gracilaria armata (C. Ag.) J. Ag. risultanopotenzialmente limitate da azoto, mentre la rizofita Cymodocea nodosa (Ucria) Asher è limitata prevalentemente da fosforo.Nell’ultimo decennio i flussi di particellato, valutati con trappole di sedimentazione poste nei fondali delle aree di studio, sono aumentatidi 4-5 volte. Nella laguna centrale l’incremento è stato di circa un ordine di grandezza, con valori medi intorno a 700 kg m-2 y-1, su basesecca. Questo incremento, inizialmente associato alla riduzione della copertura algale, è stato fortemente accentuato dall’impatto dellapesca delle vongole (Tapes philippinarum Adams & Reeve) svolta nel bacino che, una volta, era occupato dalle macroalghe.Parallellamente, la biomassa macroalgale si è ridotta a circa 1-2% dei valori misurati negli anni ’80.
PAROLE CHIAVE: nutrienti / flussi di sedimentazione / pesca delle vongole / macrofite.
Changes of nutrient concentrations and sediment fluxes in the central part of the Venice lagoonIn the last 15 years, trophic conditions and sediment fluxes in the central part of the Venice lagoon changed remarkably. Macroalgalbiomass decreased to 1-2% of the values recorded in the 1980s. Since 1983, the Manila clam, Tapes philippinarum Adams & Reeve, wasintroduced in the lagoon and colonised the bare sediments. Clam harvesting by hydraulic and mechanical dredging affected severely thebenthic system.A general decrease of sedimentary nutrients was detected, especially in areas such as San Giuliano where C, N and P were about halvedfrom 1989-90 to 2000-01. In this area macroalgae disappeared in the early 1990s. In parallel, high sediment resuspension and settlementwere caused by clam fishing. As a consequence, sediment fluxes where ca. 4-5 times (up to 12 times in other areas) higher than thosemeasured in the 1990s and favoured the loss of the finest material and the dispersal in the lagoon of high amounts of particulate nutrients.The reactive phosphorus (RP) concentrations in the water column decreased remarkably, as a result of the increased oxidation state of thelagoon and the polyphosphate banning from the commercial detergent formulations. The dissolved inorganic nitrogen (DIN) increasedweakly or remained similar to the concentrations monitored in the past. Both RP and DIN concentrations were higher than the imperativevalues fixed for total N and P in the water column by the 23rd April 1998 Ronchi-Costa decree. The tissue concentrations of nutrients in Ulva rigida C. Ag. and Gracilaria armata (C. Ag.) J. Ag., in areas with high macroalgalbiomass, showed a potential nitrogen limitation. The nutrient concentrations in the shoots of Cymodocea nodosa (Ucria) Asher.evidenced a potential phosphorus limitation.
KEY WORDS: nutrients / sediment fluxes / clam fishing / macrophytes

SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia12
INTRODUZIONEFin dagli anni ’60 la laguna di Venezia è stata
soggetta a profonde modificazioni morfologiche, chi-miche e biologiche con frequenti e repentini cambia-menti che hanno reso necessario un continuo aggior-namento delle conoscenze sia dello stato trofico chedel livello di inquinamento di acque e sedimenti.
Tra gli anni ’60 e ’70, l’espansione della produzioneindustriale e l’avvento di fertilizzanti e monocolture inagricoltura hanno causato lo sversamento in laguna dirilevanti quantità di sostanze eutrofizzanti (PAVONI etal., 1992; MARCOMINI et al., 1995). La disponibilità dinutrienti in combinazione con gli sconvolgimenti idro-dinamici causati dallo scavo di larghi e profondi canalicommerciali (Canale Malamocco-Marghera detto “Ca-nale dei Petroli”) ha creato le condizioni per una cre-scita incontrollata di macroalghe nitrofile e galenofilecome Ulva rigida C. Ag. (SFRISO et al., 1993; SFRISO eMARCOMINI, 1996a). Nel giro di pochi anni imponentibloom macroalgali hanno completamente soppiantatole praterie di fanerogame marine preesistenti in lagunacentrale e in parte della laguna nord.
La dominanza macroalgale ha perdurato fino all’ini-zio degli anni ’90, quando le macroalghe hanno comin-ciato un rapido e progressivo declino, inizialmentecausato da variazioni climatiche e, poi, rafforzato daaltre importanti concause come l’incremento dei flussidi risospensione e sedimentazione dei sedimenti super-ficiali e l’incremento del pascolo degli organismi erbi-vori, non più eliminati dalle crisi distrofiche (SFRISO eMARCOMINI, 1996b; BALDUCCI et al., 2001). Contem-poraneamente, la raccolta delle macroalghe, in presen-za di biomasse oramai limitate ad areali circoscritti, hacominciato a dare risultati positivi. Da ultimo sonodiminuite anche le immissioni di fosforo e azoto inlaguna ma, soprattutto, sono diminuiti gli apporti sta-gionali di nutrienti che si accumulavano temporanea-mente nei sedimenti superficiali.
Grazie alla sua elevata capacità di accrescimento,Ulva rigida assimilava i nutrienti dall’acqua e li trasfe-riva ai sedimenti durante la degradazione (SFRISO et al.,1987). Pertanto le macroalghe erano responsabili del-l’incremento di nutrienti sia nei sedimenti superficialiche nel particellato. Poiché la produzione lorda di Ulvaera intorno a 200-220 kg m-2 y-1 (peso umido) e nellasola laguna centrale la produzione annua era di circa 10milioni di tonnellate (SFRISO e MARCOMINI, 1994, SFRI-SO et al., 2003a) si può ben comprendere l’importanzadelle macroalghe nei cicli dei nutrienti in laguna.Similmente si possono immaginare i cambiamenti veri-ficatisi quando le biomasse macroalgali sono regredite.Attualmente in laguna centrale la produzione lorda dellemacroalghe è di ca. 230000 tonnellate per anno (SFRI-SO et al., 2003a).
Successivamente alla scomparsa delle macroalghe,la vongola filippina, Tapes philippinarum Adams &Reeve, introdotta in laguna solo nel 1983, ha coloniz-zato i fondali liberi dalle biomasse algali creando lepremesse per nuovi e drastici cambiamenti dell’am-biente lagunare. La pesca delle vongole determina unforte impatto sul comparto bentonico, dovuto all’im-piego di draghe turbosoffianti, draghe vibranti e ru-sche. Il raccolto ha raggiunto il suo apice nel 1996 conca. 50000 tonnellate di pescato e continua tuttora conuna media di ca. 30000-40000 tonnellate per anno(OREL et al., 2000). In quegli anni erano operanti ca.120 turbosoffianti ed almeno 600 barchini, apposita-mente attrezzati per questo tipo di pesca. Gli effettisono stati devastanti. In attesa di poter inquadrarequesto tipo di pesca in un’attività di pesca-colturagestita in aree concessionarie, lo scempio della lagunacontinua ancora cambiando sia la morfologia dei fon-dali che la composizione ed abbondanza delle comunitàfito e zoobentoniche.
Per effetto della pesca delle vongole, non solo sonostate spazzate via le fanerogame marine che ricomin-ciavano a popolare le aree liberate dalle macroalghe,ma anche il fitoplancton attualmente appare fortemen-te limitato. I bloom fitoplantonici rilevabili in passatonon si verificano più, a causa della distruzione dellecomunità microbentoniche e delle enormi quantità disedimenti risospesi (FACCA et al., 2002). I fondalisono in erosione per la perdita della componente sedi-mentaria più fine e gli inquinanti, accumulatisi neisedimenti durante il boom industriale post-bellico edattualmente seppelliti a 15-20 cm di profondità, sonorimessi in circolazione e ridistribuiti (SFRISO et al.,2003b).
Attualmente, nonostante la laguna sia soggetta acontinui cambiamenti in risposta alle nuove ingerenzeumane, la scomparsa dei fenomeni distrofici e dellecrisi anossiche, che puntualmente si verificavano ognianno in presenza delle imponenti biomasse di Ulva, hapermesso una miglior ossigenazione dell’ambiente e ilripopolamento da parte di molte specie algali nuove opresenti in passato prima della diffusione dell’Ulva.
Questo lavoro si propone di completare il quadrodelle conoscenze acquisite in questi ultimi 15 anni sullostato trofico della parte centrale della laguna di Vene-zia. In particolare, sono considerate le variazioni spa-zio-temporali: a) dei nutrienti nella colonna d’acqua,nei sedimenti superficiali, nei solidi sospesi e nellabiomassa delle macrofite dominanti e b) dei flussi dirisospensione e sedimentazione dei sedimenti superfi-ciali, facendo un confronto con serie di dati storicirilevati in passato dagli stessi ricercatori nelle stessearee di studio utilizzando le stesse metodiche di cam-pionamento e di analisi.
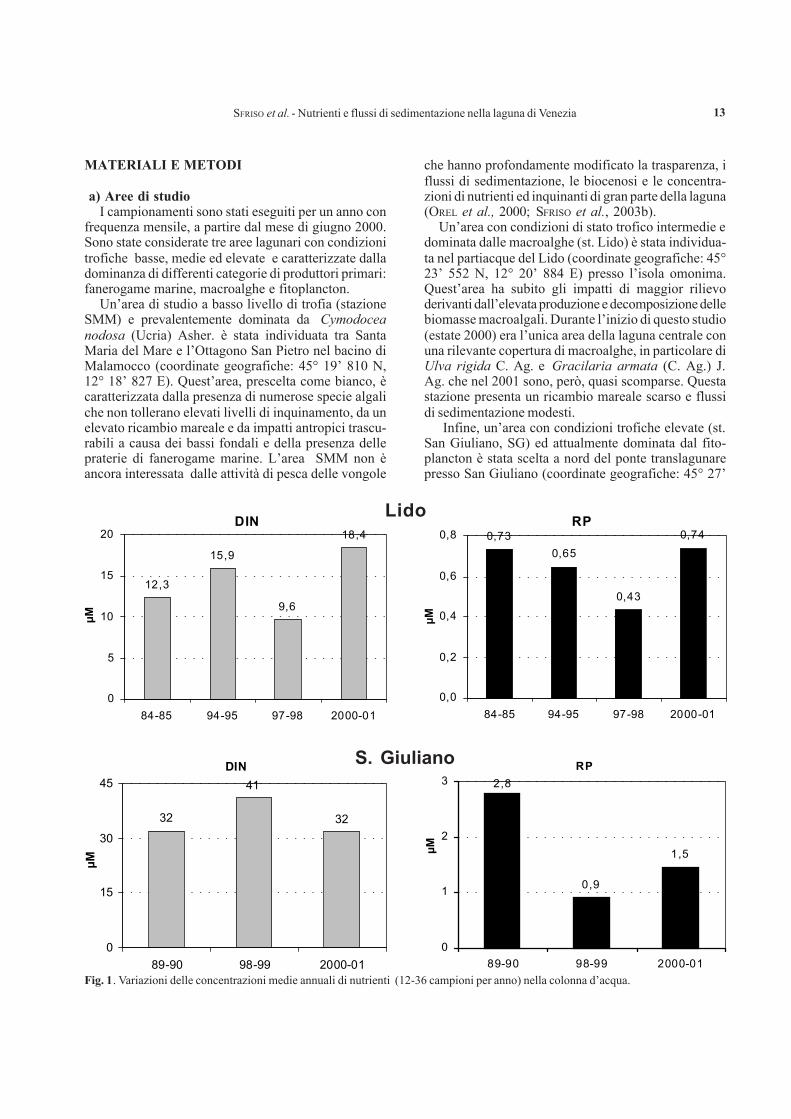
SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia 13
RP 0,73
0,65
0,43
0,74
0,0
0,2
0,4
0,6
0,8
84-85 94-95 97-98 2000-01
µM
RP
0,9
1,5
2,8
0
1
2
3
89-90 98-99 2000-01
µM
MATERIALI E METODI
a) Aree di studioI campionamenti sono stati eseguiti per un anno con
frequenza mensile, a partire dal mese di giugno 2000.Sono state considerate tre aree lagunari con condizionitrofiche basse, medie ed elevate e caratterizzate dalladominanza di differenti categorie di produttori primari:fanerogame marine, macroalghe e fitoplancton.
Un’area di studio a basso livello di trofia (stazioneSMM) e prevalentemente dominata da Cymodoceanodosa (Ucria) Asher. è stata individuata tra SantaMaria del Mare e l’Ottagono San Pietro nel bacino diMalamocco (coordinate geografiche: 45° 19’ 810 N,12° 18’ 827 E). Quest’area, prescelta come bianco, ècaratterizzata dalla presenza di numerose specie algaliche non tollerano elevati livelli di inquinamento, da unelevato ricambio mareale e da impatti antropici trascu-rabili a causa dei bassi fondali e della presenza dellepraterie di fanerogame marine. L’area SMM non èancora interessata dalle attività di pesca delle vongole
che hanno profondamente modificato la trasparenza, iflussi di sedimentazione, le biocenosi e le concentra-zioni di nutrienti ed inquinanti di gran parte della laguna(OREL et al., 2000; SFRISO et al., 2003b).
Un’area con condizioni di stato trofico intermedie edominata dalle macroalghe (st. Lido) è stata individua-ta nel partiacque del Lido (coordinate geografiche: 45°23’ 552 N, 12° 20’ 884 E) presso l’isola omonima.Quest’area ha subito gli impatti di maggior rilievoderivanti dall’elevata produzione e decomposizione dellebiomasse macroalgali. Durante l’inizio di questo studio(estate 2000) era l’unica area della laguna centrale conuna rilevante copertura di macroalghe, in particolare diUlva rigida C. Ag. e Gracilaria armata (C. Ag.) J.Ag. che nel 2001 sono, però, quasi scomparse. Questastazione presenta un ricambio mareale scarso e flussidi sedimentazione modesti.
Infine, un’area con condizioni trofiche elevate (st.San Giuliano, SG) ed attualmente dominata dal fito-plancton è stata scelta a nord del ponte translagunarepresso San Giuliano (coordinate geografiche: 45° 27’
Lido
S. Giuliano
Fig. 1. Variazioni delle concentrazioni medie annuali di nutrienti (12-36 campioni per anno) nella colonna d’acqua.
DIN
32
41
32
0
15
30
45
89-90 98-99 2000-01
µM
DIN
9,6
18,4
12,3
15,9
0
5
10
15
20
84-85 94-95 97-98 2000-01
µM
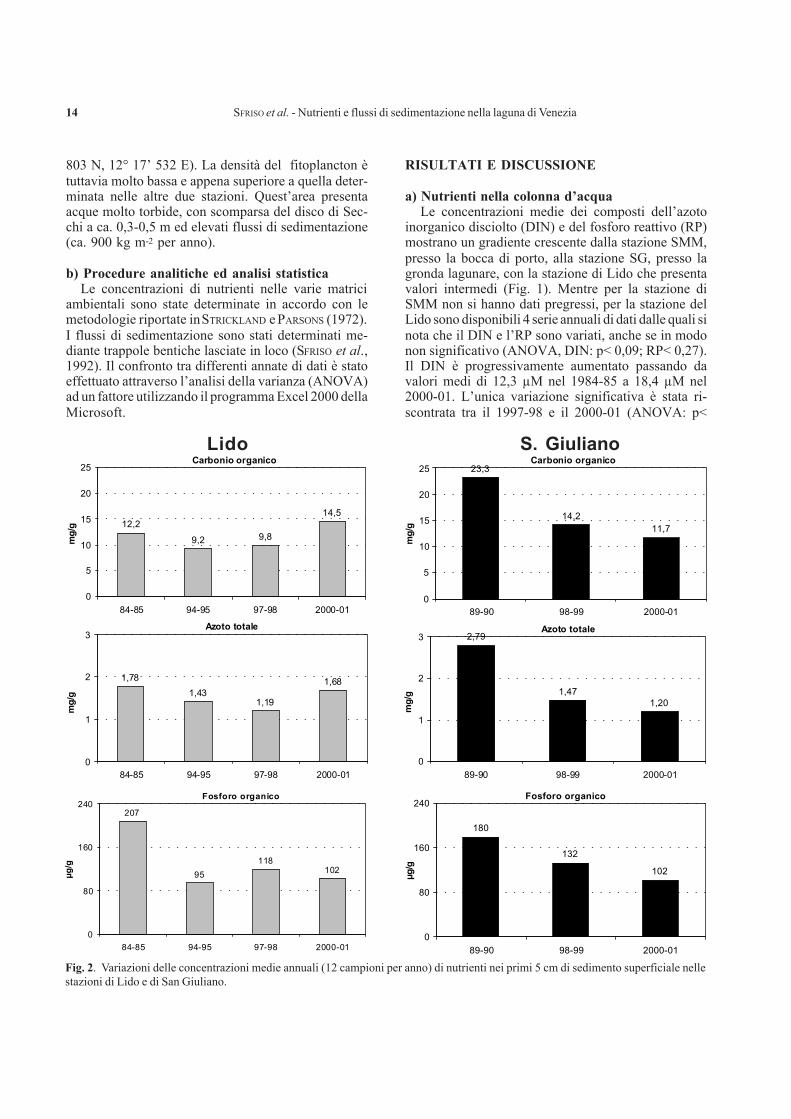
SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia14
803 N, 12° 17’ 532 E). La densità del fitoplancton ètuttavia molto bassa e appena superiore a quella deter-minata nelle altre due stazioni. Quest’area presentaacque molto torbide, con scomparsa del disco di Sec-chi a ca. 0,3-0,5 m ed elevati flussi di sedimentazione(ca. 900 kg m-2 per anno).
b) Procedure analitiche ed analisi statisticaLe concentrazioni di nutrienti nelle varie matrici
ambientali sono state determinate in accordo con lemetodologie riportate in STRICKLAND e PARSONS (1972).I flussi di sedimentazione sono stati determinati me-diante trappole bentiche lasciate in loco (SFRISO et al.,1992). Il confronto tra differenti annate di dati è statoeffettuato attraverso l’analisi della varianza (ANOVA)ad un fattore utilizzando il programma Excel 2000 dellaMicrosoft.
RISULTATI E DISCUSSIONE
a) Nutrienti nella colonna d’acquaLe concentrazioni medie dei composti dell’azoto
inorganico disciolto (DIN) e del fosforo reattivo (RP)mostrano un gradiente crescente dalla stazione SMM,presso la bocca di porto, alla stazione SG, presso lagronda lagunare, con la stazione di Lido che presentavalori intermedi (Fig. 1). Mentre per la stazione diSMM non si hanno dati pregressi, per la stazione delLido sono disponibili 4 serie annuali di dati dalle quali sinota che il DIN e l’RP sono variati, anche se in modonon significativo (ANOVA, DIN: p< 0,09; RP< 0,27).Il DIN è progressivamente aumentato passando davalori medi di 12,3 µM nel 1984-85 a 18,4 µM nel2000-01. L’unica variazione significativa è stata ri-scontrata tra il 1997-98 e il 2000-01 (ANOVA: p<
Fig. 2. Variazioni delle concentrazioni medie annuali (12 campioni per anno) di nutrienti nei primi 5 cm di sedimento superficiale nellestazioni di Lido e di San Giuliano.
Lido S. Giuliano
Fosforo organico
207
95118
102
0
80
160
240
84-85 94-95 97-98 2000-01
µg/g
Carbonio organico
12,2
9,2 9,8
14,5
0
5
10
15
20
25
84-85 94-95 97-98 2000-01
mg/
g
Azoto totale
1,781,43
1,19
1,68
0
1
2
3
84-85 94-95 97-98 2000-01
mg/
g
Fosforo organico
180
132
102
0
80
160
240
89-90 98-99 2000-01
µg/g
Carbonio organico
11,714,2
23,3
0
5
10
15
20
25
89-90 98-99 2000-01
mg/
g
Azoto totale2,79
1,471,20
0
1
2
3
89-90 98-99 2000-01
mg/
g
SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia 15
0,04). Viceversa, le concentrazioni di RP sono dimi-nuite passando da 0,73 µM nel 1984-85 a 0,43 µM nel1997-98 per aumentare nuovamente a 0,74 µM nel2000-01. Nella stazione di SG, dove le macroalghesono scomparse all’inizio degli anni ’90, si nota che ilDIN non ha presentato variazioni di rilievo, oscillandotra 32 e 41µM (ANOVA: p< 0,39). L’RP è invecediminuito significativamente (ANOVA: p< 5,0E-04)passando da 2,8 µM nel 1989-90 a 0,9 µM nell’ultimoperiodo. Nella stazione di SG, sia il DIN che l’RPraggiungono concentrazioni notevolmente superiori aivalori del decreto Ronchi-Costa del 23 aprile 1998 chefissava come imperative le concentrazioni di 0,8 µMper il fosforo totale e 25 µM per l’azoto totale.
b) Nutrienti nei sedimenti superficialiNelle stazioni di Lido, ancora popolata da Ulva
rigida nel 2000-2001, e di SG, priva di macroalghe findal 1993, le concentrazioni di azoto e fosforo nelsedimento superficiale seguono andamenti temporalidiversi (Fig. 2).
Nella stazione di SG, le concentrazioni medie deinutrienti tra il 1989 e il 2001 si sono significativamenteridotte passando da 23,3±1,6 a 11,7±3,5 mg g-1 per ilCorg (carbonio organico) (ANOVA: p< 1,3E-12), da2,8±0,2 a 1,2±0,2 mg g-1 per l’Ntot (azoto totale)(ANOVA: p< 9,0E-20) e da 180±23 a 102±21 µg g-1
per il Porg (fosforo organico) (ANOVA: p< 6,6E-06).Nella stazione di Lido sia l’Ntot (ANOVA: p< 1,3E-
Carbonio Totale
270
320
370
420
giu
lug
ago
set
ott
nov
dic
gen
feb
mar ap
r
mag
mg/
g
Ge rmogli (372±12)Rizomi (370±17)Parti morte (333±26)Cymodocea nodosa
Azoto Totale
010203040
giu
lug
ago
set
ott
nov
dic
gen
feb
mar ap
r
mag
mg/
g
Germogli (34.5±3.9)Rizomi (17.1±4.6)Parti morte (22.1±5.2)
Valorecritico
18 mg/g
Cymodocea nodosaAzoto Totale
10
20
30
40
Giu
Lug
Ago
Set
Ott
Nov
Dic
Gen Feb
Mar Apr
Mag
mg/
g
Ulva (24.7±4.3)Gracilaria (25.5±6.9)
Va lore cri tico
20 mg/g
Fosforo Totale
0
1
2
3
giu
lug
ago
set
ott
nov
dic
gen
feb
mar ap
r
mag
mg/
g
Ge rmogli (2.16±0.32)Rizomi (1.63±0.52)Parti morte (0.84±0.15)
Va lore cri tico
2.0 mg/g
Cymodocea nodosaFosforo Totale
0.0
0.5
1.0
1.5
2.0
Giu
Lug
Ago
Set
Ott
Nov
Dic
Gen Feb
Mar Apr
Mag
mg/
g
Ulva (1.07± 0.4)Gracilaria (1.23±0.4)
Va lorecri tico
0.25 mg/g
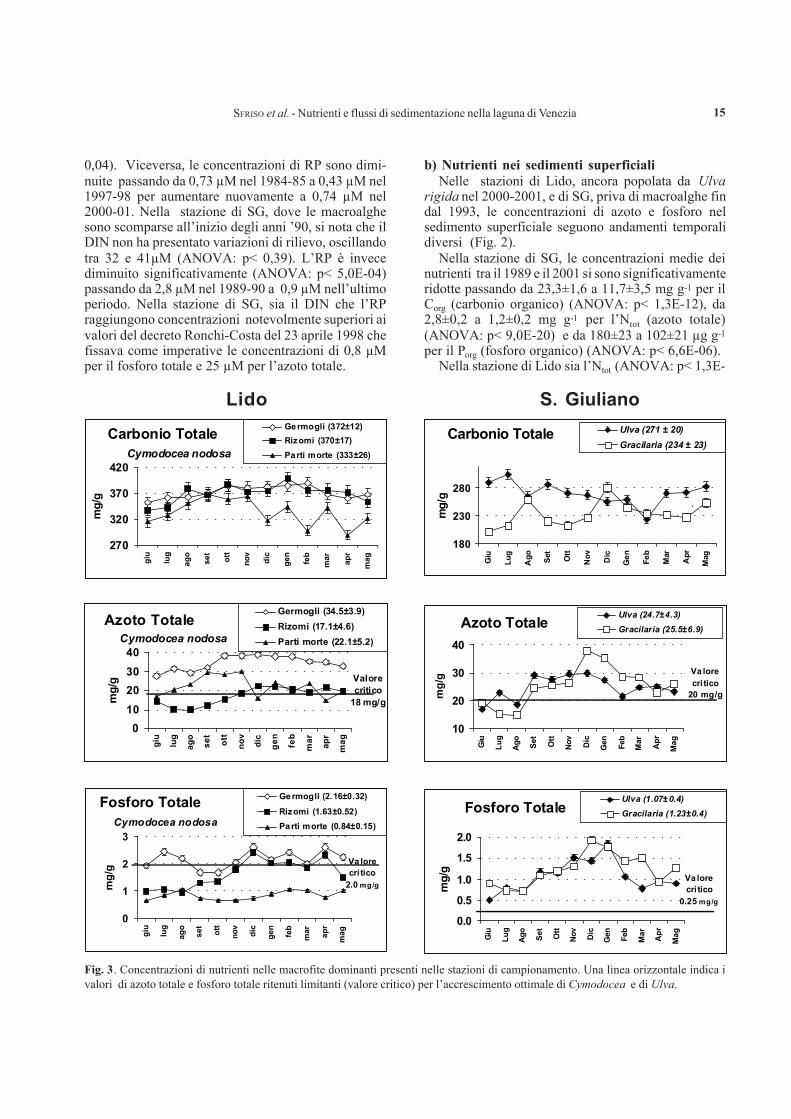
Fig. 3. Concentrazioni di nutrienti nelle macrofite dominanti presenti nelle stazioni di campionamento. Una linea orizzontale indica ivalori di azoto totale e fosforo totale ritenuti limitanti (valore critico) per l’accrescimento ottimale di Cymodocea e di Ulva.
Lido S. Giuliano
Carbonio Totale
180
230
280G
iu
Lug
Ago Se
t
Ott
Nov Dic
Gen Fe
b
Mar
Apr
Mag
mg/
gUlva (271 ± 20)Gracilaria (234 ± 23)
.
SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia16
04) che il Corg (ANOVA: p< 1,1E-10) hanno presenta-to variazioni altamente significative durante tutto ilperiodo considerato. Tuttavia, dopo un significativodecremento monitorato fra il 1984-85 e il 1997-98(ANOVA, Ntot: p< 1,2E-05; Corg: p< 2,0E-09), leconcentrazioni di questi nutrienti sono nuovamenteaumentate nel 2000-01 (ANOVA, Ntot: p< 1,3E-03;Corg: p< 8,7E-07), probabilmente a causa degli elevatiflussi di sedimentazione e del trasporto di sedimenticausati dalla pesca delle vongole. Il Porg mostra inveceuna continua tendenza alla diminuzione passando da207± 64 a 102±21 µg g-1 (ANOVA: 5,0E-09).
c) Nutrienti nei tessuti delle macrofite dominantiAttualmente le macroalghe presentano una bio-
massa e una produzione primaria pari a circa il 2% deivalori trovati negli anni ’80 (SFRISO et al. 2003a).Tuttavia il numero di specie presenti in laguna è piùche doppio rispetto a una decina di anni fa (SFRISO etal., 2002). La diffusione delle fanerogame marine èinvece in espansione poiché queste piante hanno oc-cupato molti areali, un tempo popolati dalle macroal-ghe, soprattutto in aree marginali lungo i canali e neibassi fondali dove l’attività di pesca delle vongole èmodesta.
Nella stazione di SMM, le concentrazioni di Ctot
(carbonio totale) nei germogli e nei rizomi di Cymodo-cea nodosa, fanerogama marina che raggiunge bio-masse medie elevate (3048±1134 g m-2, su base umi-da), sono risultate simili (372±12 e 370±17 mg g-1),mentre le concentrazioni di Ntot e Ptot (azoto e fosforototali) sono risultate sempre maggiori nei germogli chenegli altri tessuti. Le concentrazioni di Ntot nei germogli(34,5±33,9 mg g-1) non sono mai state al di sotto delleconcentrazioni ritenute critiche (18 mg g-1) per unaccrescimento ottimale di questa specie, mentre quelledel Ptot, variando tra ca. 1,8 e ca. 2,3 mg g-1, sonorisultate potenzialmente limitanti (<2,0 mg g-1) trasettembre ed ottobre, durante il periodo del massimoincremento di biomassa (Fig. 3).
In Ulva rigida e Gracilaria armata, macroalghepresenti con una discreta biomassa nella stazione Lido,le concentrazioni di Ntot (24,7±4.3 e 25,6±6,9 mg g-1,rispettivamente) hanno raggiunto concentrazioni inter-medie rispetto a quelle rilevate nei germogli e nei rizomidi Cymodocea. Le concentrazioni di Ptot (1,1±0,4 e1,2±0,4 mg g-1, rispettivamente) erano, invece, sem-pre inferiori. Mentre per il fosforo le concentrazionierano sempre notevolmente più elevate della concen-trazione considerata critica per l’accrescimento (0,25mg g-1), per l’azoto sono stati trovati valori potenzial-mente limitanti (<20 mg g-1) tra giugno ed agosto.
Flussi SPM
0
1000
2000
3000
4000
5000
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu
g/m
² gio
rno,
DW
89-90 (387)98-99 (1963)2000-01 (1741)
Flussi medi
141
636721
0
200
400
600
800
Kg/
m² a
nno,
DW
89-90 98-99 2000-01
Flussi medi
39
142
38
0
50
100
150
Kg/
m² a
nno,
DW
97-98 2000-0194-95
Flussi SPM
0
250
500
750
1000
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu
g/m
² gio
rno,
DW
94-95 (108)97-98 (102)2000-01 (391)
2050
Lido
S. Giuliano
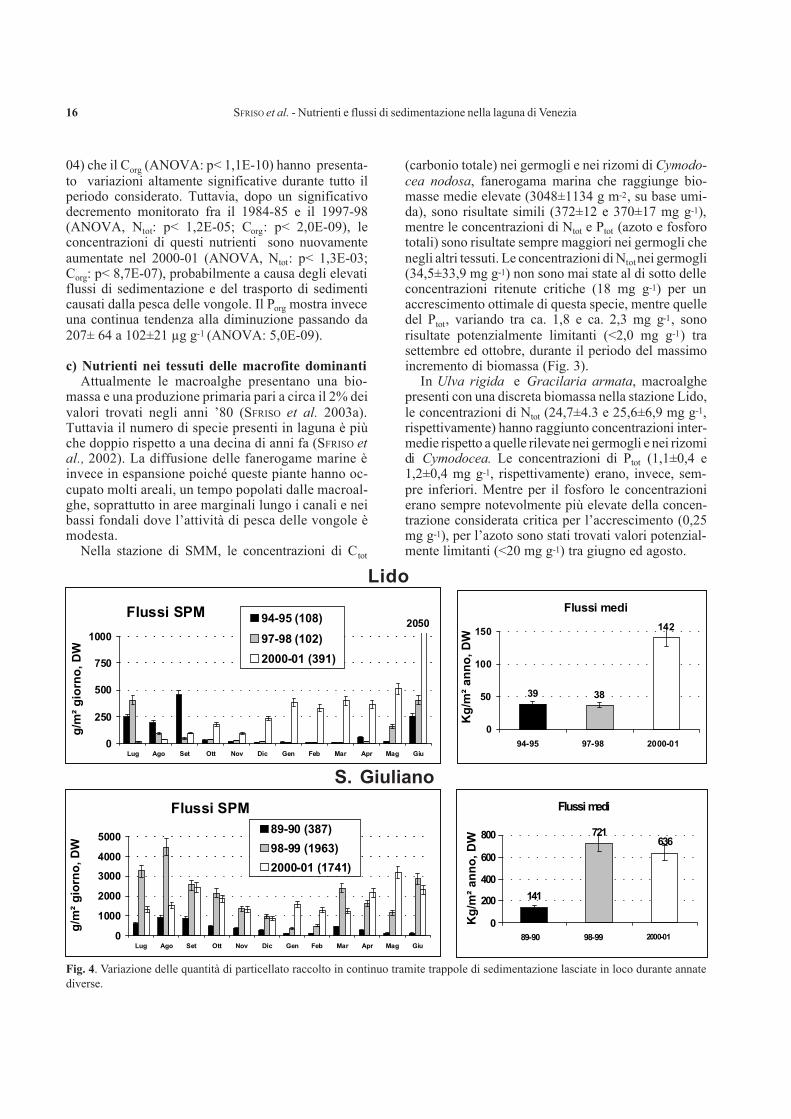
Fig. 4. Variazione delle quantità di particellato raccolto in continuo tramite trappole di sedimentazione lasciate in loco durante annatediverse.

SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia 17
d) Flussi di sedimentazione e di nutrientiCon la scomparsa delle macroalghe e la diffusione e
pesca delle vongole i flussi di sedimentazione, tra il1989 e il 2000-01, sono notevolmente aumentati pas-sando da 141 a 600-700 kg m-2 y-1, come sostanzasecca, nella stazione di SG (ANOVA: p< 5,9E-05),mentre al Lido l’incremento è stato più contenuto(ANOVA: p<0,07), variando tra 38-39 e 142 kg m-2
y-1 (Fig. 4). In quest’ultima stazione, dopo la scom-parsa della biomassa macroalgale avvenuta alla fine del2000, i flussi di particellato sono passati da valoriinferiori a 250 g m-2 d-1 a valori fino a 2050 g m-2 d-1.I flussi sono stati marcatamente più elevati nella sta-gione calda presentando picchi di sedimentazione traaprile e giugno e tra settembre e novembre, in conco-mitanza con il maggior sforzo di pesca delle vongolefilippine.
Nel complesso, anche se le concentrazioni di nu-trienti nel particellato sono risultate inferiori a quellerilevate in passato, i flussi totali di nutrienti veicolati colparticellato nel 2000-01 sono risultati più elevati chenegli anni precedenti. Infatti, i flussi di nutrienti legatial particellato dipendono dai flussi di sedimentazione edalla concentrazione dei nutrienti nel materiale sedi-mentato. Nella stazione di Lido, caratterizzata dallapresenza delle macroalghe, il particellato sedimentatoaveva un contenuto medio mensile di C, N e P piùelevato che nelle altre stazioni, raggiungendo concen-trazioni di Corg di 32±7 mg g-1, Ntot di 4,2±1,2 mg g-1 edi Porg di 279±67 µg g-1. Le concentrazioni medie piùbasse sono state determinate nella stazione di SMM.
Sulla base dei flussi di particellato nella stazione diSG, la ricaduta dei nutrienti ha raggiunto ca. 12,0 kgm-2 y-1 per il Corg, ca. 1,3 kg m-2 y-1 per Ntot e ca. 0,1kg m-2 y-1 per il Porg. Nella stazione di SMM i flussi dinutrienti sono stati di circa un ordine di grandezza piùbassi: ca. 1,7 kg m-2 y-1 per il Corg, ca. 0,2 kg m-2 y-1
per Ntot e ca. 0,01 kg m-2 y-1 per il Porg. La stazione delLido presentava valori intermedi, ma più prossimi aquelli della stazione di SMM.
CONCLUSIONI Nelle aree lagunari considerate, dalla fine degli anni
’80 al 2001, le concentrazioni dei nutrienti nelle acquee nei sedimenti superficiali sono variate in concomitan-za con la riduzione e la scomparsa quasi totale dellemacroalghe e con la diffusione e la pesca delle vongolefilippine. Le variazioni più significative si sono osser-vate soprattutto nei sedimenti superficiali nelle areeprive di macroalghe, dove, in alcuni casi, le concentra-zioni di Corg, Ntot e Porg sono addirittura dimezzate.
Nelle acque si nota, invece, una tendenza alla dimi-nuzione del RP e all’incremento del DIN. La riduzionedel RP può essere attribuita all’eliminazione dei fosfatinelle formulazioni commerciali dei detergenti e allecondizioni ossidative del sedimento superficiale. L’in-cremento del DIN può essere invece indotto dallaridotta assimilazione dei produttori primari.
Le concentrazioni tissutali di azoto e fosforo neigermogli della fanerogama Cymodocea nodosa, checresce in acque ben ossigenate, mostrano una sualimitazione potenziale da fosforo. Nelle macroalghe,che prediligono le aree lagunari più eutrofizzate, èinvece l’azoto che appare ancora l’elemento critico,ma limitatamente al periodo di maggior accrescimento,in giugno-agosto.
Contemporaneamente alla riduzione delle biomassemacroalgali e alla diffusione e alla pesca delle vongolesi è registrato anche un significativo incremento dellarisospensione, del trasporto e dei flussi di sedimenta-zione. Attualmente (2003) in laguna centrale i flussimedi di sedimentazione sono circa un ordine di gran-dezza più elevati rispetto agli anni ’80. Tuttavia larecente regolamentazione delle attività di pesca allevongole e l’assegnazione da parte della Provincia diVenezia di aree per la venericoltura (ca. 3000 ha)distribuite in tutta la laguna (OREL et al., 2000), hannodimostrato che la produzione e la raccolta di vongole inaree confinate hanno un impatto limitato sui flussi disedimentazione considerati su base annuale (PESSA eSFRISO, 2002).
Le attività di raccolta delle vongole e di sistemazionedei fondali, compresa l’estirpazione di eventuali fane-rogame marine, hanno infatti un impatto ambientaleparagonabile a quello di un normale evento meteorolo-gico avverso.
BIBLIOGRAFIA
BALDUCCI C., SFRISO A., PAVONI B., 2001. Macrofauna impacton Ulva rigida C.Ag. production and relationship withenvironmental variables in the lagoon of Venice. Mar. Env.Res. 52: 27-49.
FACCA C., SFRISO A., SOCAL G., 2002. Changes in abundance and
composition of phytoplankton and microphytobenthosdue to increased sediment fluxes in the Venice lagoon,Italy. Estuarine, Coastal and Shelf Science, 54: 773-792.
MARCOMINI A., SFRISO A., PAVONI B., ORIO A.A., 1995. Eutrophi-cation of the lagoon of Venice: nutrient loads and exchanges.

SFRISO et al. - Nutrienti e flussi di sedimentazione nella laguna di Venezia18
In: (A.J.Mc Comb ed.), Eutrophic Shallow Estuaries andLagoons, CRC Press, Boca Raton, Fl., U.S.A. 59-80 pp.
OREL G., BOATTO V., SFRISO A., PELLIZZATO M., 2000. Piano perla gestione delle risorse alieutiche delle lagune della Provin-cia di Venezia. In: (Provincia di Venezia ed.), Sannioprint,Benevento. 102 pp.
PAVONI B., MARCOMINI A., SFRISO A., DONAZZOLO R., ORIO A.A.,1992. Changes in an estuarine ecosystem. The Lagoon ofVenice as a case study. In: (D.A. Dunnette and R.J. O’Brieneds.), The Science of Global Change, American ChemicalSociety, Washington, D.C., U.S.A. 287-305 pp.
PESSA G., SFRISO A., 2002. Monitoraggio degli effetti della pescadi Tapes philippinarum sui flussi di sedimentazione, sui pro-cessi di erosione/sedimentazione e sulla distribuzione di ma-croalghe e fanerogame marine in aree bersaglio e prossimealle zone date in concessione per la pesca allevamento diquesti molluschi. Provincia di Venezia. Rapporto Finale.
SFRISO A., MARCOMINI A., PAVONI B., 1987. Relationship betwe-en macroalgal biomass and nutrient concentrations in a hyper-trophic area of the Venice lagoon. Marine EnvironmentalResearch, 22: 297-312.
SFRISO A., PAVONI B., MARCOMINI A., RACCANELLI S., ORIO A.A.,1992. Particulate matter deposition and nutrient fluxes ontothe sediments of the Venice lagoon. Environmental Technolo-gy, 13: 473-483.
SFRISO A., MARCOMINI A., PAVONI B., ORIO A.A., 1993. Species
composition, biomass and net primary production in coastalshallow waters: the Venice lagoon. Bioresource Technology,44: 235-250.
SFRISO A., MARCOMINI A., 1994. Gross primary production andnutrient behaviours in shallow lagoon waters. BioresourceTechnology, 45: 59-66.
SFRISO A., MARCOMINI A., 1996a. Chap.15 Italy - The Lagoon ofVenice. In: (W.Schramm and P.N. Nienhuis eds.), MarineBenthic Vegetation, Ecological Studies, Springer Verlag, Ber-lin Heidelberg, Vol. 123: 339-368.
SFRISO A., MARCOMINI A., 1996b. Decline of Ulva growth in thelagoon of Venice. Bioresource Technology, 58: 299-307.
SFRISO A., LA ROCCA B., GODINI E., 2002. Inventario di taxamacroalgali in tre aree della laguna di Venezia a differentelivello di trofia. Lavori Soc. Ven. Sci. Nat., 27: 85-99.
SFRISO A., FACCA, C., GHETTI P.F. 2003a. Temporal and spatialchanges of macroalgae and phytoplankton in a Mediterraneancoastal area: the Venice lagoon as a case study. MarineEnvironmental Research, 56: 617-636.
SFRISO A., FACCA C., CEOLDO S., SILVESTRI S., GHETTI P.F. (2003b).Role of macroalgal biomass and clam fishing on spatial andtemporal changes in N and P sedimentary pools in the centralpart of the Venice lagoon. Oceanologica Acta, 26: 3-12
STRICKLAND J.D.H., PARSONS T.R., 1972. A Practical Handbookof Seawater Analyses. Fisheries Research Board of Canada,Ottawa. 309 pp.

Biologia Ambientale, 18 (2): 19-24, 2004
RiassuntoGli Autori presentano i risultati relativi allo studio delle comunità planctoniche e bentoniche, effettuato concampionamenti mensili, da giugno 2000 a maggio 2001, in tre stazioni della laguna di Venezia e con osservazioni al microscopio otticoinvertito. Gli andamenti del fitoplancton seguono le variazioni stagionali con minimi in inverno (ca. 0,15 x 106 cells dm-3) e massimi inprimavera-estate (da 1 a 19 x 106 cells dm-3). Le abbondanze maggiori sono state individuate nell’area più fortemente impattata dalleattività antropiche, a ridosso della terraferma. Le diatomee bentoniche, invece, hanno presentato andamenti più stabili con variazioni, adun primo esame, indipendenti dalle condizioni meteorologiche e densità mediamente più elevate (2,7 x 106 cells cm-3) nelle aree popolateda macrofite. Nella colonna d’acqua le specie dominanti sono state Chaetoceros socialis Lauder e Skeletonema costatum Cleve, mentrenel sedimento hanno prevalso Cocconeis scutellum Ehrenberg, nelle aree popolate da macrofite, e Thalassiosira sp., in prossimità dellaterraferma.
PAROLE CHIAVE: fitoplancton / diatomee bentoniche / biodiversità / laguna di Venezia
Phytoplankton and benthic diatom abundance and diversity in the lagoon of VeniceWater and surface sediment samples were collected monthly from May 2000 to June 2001 in three stations of Venice lagoon to investigatephytoplankton and benthic diatom distribution. Phytoplankton abundance was minimum in December at St. B and C and in January atSt. A (ca. 0.15 x 106 cells dm-3). The first increase was observed in February with Skeletonema costatum Cleve bloom whereas themaximum value occurred in April with Chaetoceros socialis Lauder. On average the highest abundance was recorded at St. C (4.4 ± 5.6x 106 cells dm-3), where macrophytes lack and human activities are intense, and the lowest one at St. A (0.6 ± 0.3 x 106 cells dm-3),populated by seagrasses. Trends of benthic diatoms are more complex to explain. In fact, at all stations maximum values were recordedin June and July and then abundances progressively decreased and no more blooms were observed except for a light increase in Februaryat St. C. The highest mean abundance was recorded at St. B (2.7 ± 0.8 x 106 cells cm-3), populated by macroalgae, whereas at St. A andC the mean values were rather similar (1.4 ± 0.9 and 1.8 ± 0.6 x 106 cells cm-3, respectively). At a first exam the presence/absence ofmacrophyte and the fluxes of sediment appear to be the factors which mainly affected benthic diatoms distribution. Althoughmacrophytes can compete with microalgae for resources they are, in general, rich of epiphytes and favour cells settlement. In surfacesediment Cocconeis scutellum Ehrenberg determined, in fact, the high abundance recorded at St. A and B.No direct correspondence was observed between the population found in the water column and in the sediment samples. Also therelationship between diversity and abundance was different: in sediment, dominance didn’t correspond to bloom phenomena.
KEY WORDS: phytoplankton / benthic diatoms / biodiversity / Venice lagoon
Abbondanza e diversità del fitoplancton edelle diatomee bentoniche in laguna di VeneziaChiara Facca, Adriano Sfriso*, Pier Francesco GhettiDipartimento di Scienze Ambientali, Università di Venezia, Calle Larga Santa Marta 2137, 30123 Venezia
* Referente per la corrispondenza. Fax: +39 (0)41 2348584; e-mail: [email protected]
Pervenuto il 7.8.2003; accettato il 8.10.2003
INTRODUZIONENelle aree costiere, in cui la colonna d’acqua ha
scarsa profondità e la radiazione luminosa penetra finoal fondo, le macrofite sono, in genere, i maggiori
produttori primari rispetto al fitoplancton, la cui pro-duzione è importante soprattutto in mare aperto, e almicrofitobenthos, che prevale nelle zone intertidali e

FACCA et al. - Fitoplancton e diatomee bentoniche in laguna di Venezia20
nei bassifondi privi di macrofite. Questa è stata lasituazione, per diversi decenni, anche della laguna diVenezia, in cui le massicce fioriture macroalgali hannospesso provocato gravi crisi anossiche. All’inizio deglianni Novanta, tuttavia, l’elevata produzione macroal-gale è andata diminuendo, soprattutto per effetto dicondizioni meteorologiche sfavorevoli alla crescita diUlva rigida C. Ag., ripetutesi per più anni consecutivi(SFRISO e MARCOMINI, 1996). Uno studio condottorecentemente (1998-99: SFRISO, 2000) ha evidenziatoche la biomassa macroalgale è quasi completamentescomparsa dall’area centrale della laguna. In questecondizioni si è supposto che il fitoplancton e il microfi-tobenthos fossero in grado di sostituirsi alle macrofiteed assumerne il ruolo. A questo scopo lo studio diqueste due comunità e delle loro interazioni si è fattopiù attento, anche alla luce delle condizioni ambientaliche si sono stabilite nell’area a seguito della scomparsadelle macroalghe e della raccolta incontrollata di Tapesphilippinarum Adams e Reeve. I flussi di risospensio-ne/sedimentazione sono diventati fino a 12 volte mag-
giori rispetto a quelli registrati durante le fioriture diUlva, prima della diffusione del bivalve (SFRISO, 2000).Questo fenomeno ha aspetti molto preoccupanti sul-l’ecosistema perché riduce la trasmissione della radia-zione luminosa nella colonna d’acqua e, quindi, lacapacità fotosintetica degli organismi autotrofi, alteral’habitat bentonico impedendo l’attecchimento di qua-lunque forma di vita e favorisce l’allontanamento deisedimenti fini verso il mare.
Studi condotti alla fine degli anni Novanta in quattrostazioni dell’area centrale (di cui tre prive di macrofite)hanno dato esiti inaspettati: si è notata, infatti, unaflessione dell’abbondanza fitoplanctonica (FACCA etal., 2001), probabilmente indotta dai frequenti feno-meni di risospensione dei sedimenti superficiali.
Lo scopo del presente lavoro è quello di tracciare unquadro della distribuzione non solo temporale, vistoche comunque la campagna si è protratta per un anno,ma anche spaziale delle comunità fitoplanctonica edelle diatomee bentoniche in relazione al tipo di macro-fite presenti e al grado di trofia della zona.
Fig. 1. Stazioni di campionamento
ITALY SLOVENIA
CROATIA
ADRIATICSEA
VENICE
A: Santa Mariadel Mare
B: Lido
C: SanGiuliano
BURANO
TORCELLO
LIDOINLET
DRY LANDS
TIDAL LANDS
MALAMOCCOINLET
AIRPORT
PORTOMARGHERA
LIDO
VENICE

FACCA et al. - Fitoplancton e diatomee bentoniche in laguna di Venezia 21
MATERIALI E METODII campioni sono stati raccolti con cadenza mensile
da giugno 2000 a maggio 2001 in tre stazioni dellalaguna di Venezia (Fig. 1). Una stazione (St. A, SantaMaria del Mare), posta a sud della bocca di porto diMalamocco, risente direttamente degli scambi con ilmare, ha sedimenti prevalentemente sabbiosi ed è po-polata da fanerogame (soprattutto Cymodocea nodosaAsch). La seconda stazione (St. B, Lido), in prossimitàdel Lido di Venezia, presso lo spartiacque, risente degliscarichi urbani dell’isola, è popolata soprattutto damacroalghe (Ulva rigida C. Ag. e Gracilaria armata(C. Agardh) J. Agardh) e presenta sedimenti costituitiper il 60% da silt ed argilla (frazione <63 μm). La terzastazione (St. C, San Giuliano), priva di macrofite, èposta a nord del ponte translagunare che collega Vene-zia alla terraferma. Quest’area è caratterizzata da in-tenso traffico marittimo, scarichi urbani ed agricoliprovenienti dagli insediamenti dell’entroterra, bassoriciclo mareale, sedimenti prevalentemente fini e flussidi sedimentazione che risultano cinque e dieci voltemaggiori che nelle St. B ed A, rispettivamente.
Il fitoplancton è stato stimato su campioni rappre-sentativi di tutta la colonna d’acqua (raccolti medianteuna bottiglia di Plexiglas di ca. 1,5 m d’altezza e 4 cmdi diametro). Gli strati superficiali di sedimento, dipochi millimetri di spessore, sono stati prelevati dabarca mediante carotatore e sono stati trattati e con-servati secondo i suggerimenti proposti da SMYTH(1955) e TOLOMIO et al. (2000) e la metodologia de-scritta in FACCA et al. (2002).
L’analisi quantitativa e qualitativa è stata condotta almicroscopio ottico invertito (Axiovert, ZEISS) secon-do la procedura proposta da UTHERMÖL (1958). L’iden-tificazione tassonomica e l’aggiornamento della no-menclatura sono stati eseguiti sulla base dei seguentitesti: PERAGALLO e PERAGALLO (1897-1908), VANLAN-DINGHAM (1967-1979), ROUND et al. (1990) e TOMAS(1993, 1996). I valori dell’indice di diversità di Shan-non sono stati calcolati mediante il software PRIMERv. 5 (PRIMER-E Ltd., Plymouth, UK).
RISULTATI
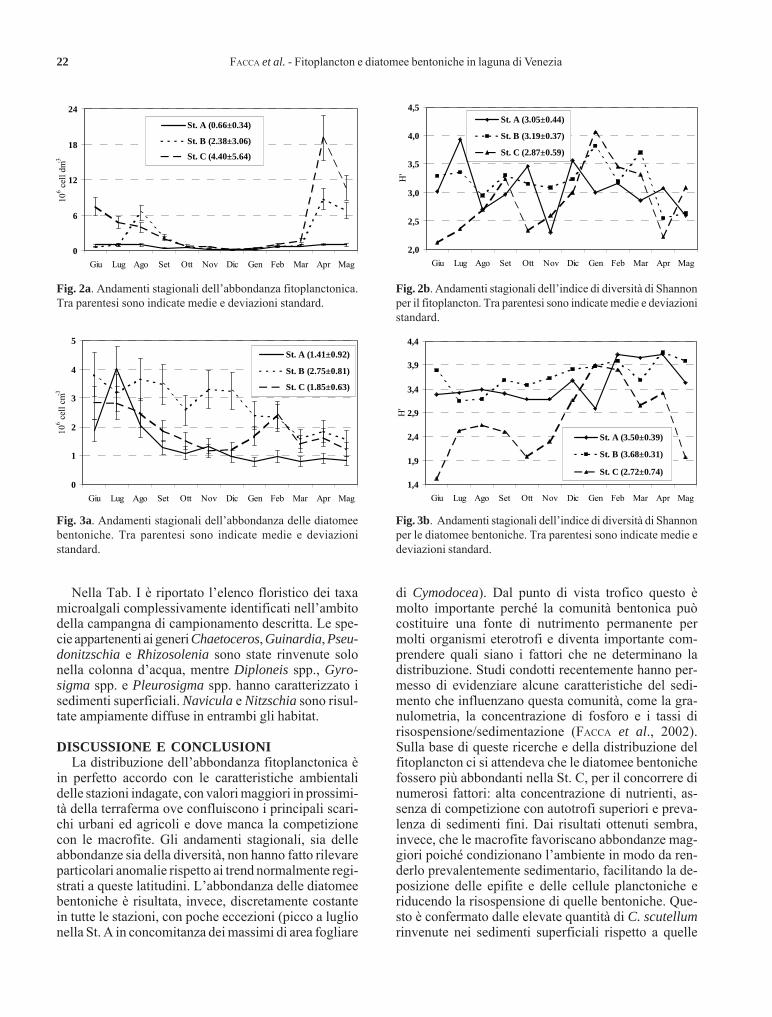
FitoplanctonI valori più bassi (ca. 0,15 x 106 cells dm-3; Fig. 2a)
sono stati registrati tra dicembre e gennaio mentre, giàin febbraio, si è osservato un bloom di Skeletonemacostatum Cleve. Specie appartenenti al genere Chaeto-ceros hanno determinato la fioritura più abbondante (1,8,7 e 19 x 106 cells dm-3, rispettivamente nelle St. A, Be C) in aprile. La St. A è stata quella a più bassa densitàfitoplanctonica (media 0,6 ± 0,3 x 106 cells dm-3) conuna percentuale di diatomee pari al 50% e di nanofla-
gellati (termine con cui si individuano cellule flagellatesferiche di meno di 5 μm di diametro) pari al 30% dellecellule complessivamente stimate. Le diatomee sonoprevalse (94%) a febbraio, a seguito del già citatobloom di S. costatum e di una discreta quantità diPseudonitzschia seriata Peragallo. Le diatomee sonostate, mediamente, più abbondanti (73%) nella St. B, incui sono state registrate due fioriture simili, una adagosto ed una ad aprile, sempre di specie appartenential genere Chaetoceros (in particolare C. socialis Lau-der e C. tenuissimus Meunier). Dinophyceae sonosporadicamente apparse solo nelle St. A e B, mentreCryptophyceae sono risultate discretamente abbon-danti in tutte le stazioni. L’abbondanza fitoplanctonicaè stata, mediamente, maggiore (4,4 ± 5,6 x 106 cellsdm-3) a ridosso della terraferma, dove le diatomeehanno rappresentato il 43% delle cellule complessiva-mente identificate. Oltre alle sensibili abbondanze diChaetoceros, nel mese di aprile Micromonas sp. harappresentato ca. il 50% della comunità.
Gli andamenti di diversità (Fig. 2b) hanno evidenzia-to fluttuazioni repentine con massimi tra dicembre egennaio e minimi in concomitanza del bloom di Chae-toceros in aprile.
Diatomee bentonicheNella St. A, sebbene si sia registrato un picco a
luglio (4 x 106 cells cm-3; Fig. 3a), a novembre si èraggiunta un’abbondanza maggiore (1,3 x 106 cellscm-3) che in tutta la primavera successiva. Nella St. Bsi può notare un trend progressivamente decrescenteda giugno 2000 (3,8 x 106 cells cm-3) a maggio 2001(1,5 x 106 cells cm-3). Proprio in questa stazione èstato, comunque, registrato il valore medio massimo(2,7 ± 0,8 x 106 cells cm-3). Gli andamenti di questedue stazioni sono stati condizionati soprattutto dall’ab-bondanza di Cocconeis scutellum Ehrenberg, diatomeapennata tipicamente epifita, e da cellule di piccoledimensioni (<10 μm), in prevalenza appartenenti algenere Navicula. Sostanziali differenze si sono notatenella St. C, a partire dalla scarsa abbondanza di C.scutellum (fino a cinque/sei volte inferiore alle altrestazioni) e dalla dominanza di Thalassiosira sp., princi-pale responsabile dell’andamento registrato fino a no-vembre. Il picco di febbraio è in buona misura attri-buibile ad un aumento di Gyrosigma distortum Griffithe Henfrey e a diatomee pennate (principalmente Navi-cula di ca. 20-30 μm) non facilmente identificabili.
Le variazioni dell’indice di Shannon (Fig. 3b) sonorisultate piuttosto contenute e graduali; non ci sonostate flessioni significative in corrispondenza degli au-menti delle abbondanze, anzi spesso gli andamentierano simili. Questo sembra, una volta in più, confer-mare la teoria sull’elevata stabilità della comunità.
FACCA et al. - Fitoplancton e diatomee bentoniche in laguna di Venezia22
Nella Tab. I è riportato l’elenco floristico dei taxamicroalgali complessivamente identificati nell’ambitodella campangna di campionamento descritta. Le spe-cie appartenenti ai generi Chaetoceros, Guinardia, Pseu-donitzschia e Rhizosolenia sono state rinvenute solonella colonna d’acqua, mentre Diploneis spp., Gyro-sigma spp. e Pleurosigma spp. hanno caratterizzato isedimenti superficiali. Navicula e Nitzschia sono risul-tate ampiamente diffuse in entrambi gli habitat.
DISCUSSIONE E CONCLUSIONILa distribuzione dell’abbondanza fitoplanctonica è
in perfetto accordo con le caratteristiche ambientalidelle stazioni indagate, con valori maggiori in prossimi-tà della terraferma ove confluiscono i principali scari-chi urbani ed agricoli e dove manca la competizionecon le macrofite. Gli andamenti stagionali, sia delleabbondanze sia della diversità, non hanno fatto rilevareparticolari anomalie rispetto ai trend normalmente regi-strati a queste latitudini. L’abbondanza delle diatomeebentoniche è risultata, invece, discretamente costantein tutte le stazioni, con poche eccezioni (picco a luglionella St. A in concomitanza dei massimi di area fogliare
di Cymodocea). Dal punto di vista trofico questo èmolto importante perché la comunità bentonica puòcostituire una fonte di nutrimento permanente permolti organismi eterotrofi e diventa importante com-prendere quali siano i fattori che ne determinano ladistribuzione. Studi condotti recentemente hanno per-messo di evidenziare alcune caratteristiche del sedi-mento che influenzano questa comunità, come la gra-nulometria, la concentrazione di fosforo e i tassi dirisospensione/sedimentazione (FACCA et al., 2002).Sulla base di queste ricerche e della distribuzione delfitoplancton ci si attendeva che le diatomee bentonichefossero più abbondanti nella St. C, per il concorrere dinumerosi fattori: alta concentrazione di nutrienti, as-senza di competizione con autotrofi superiori e preva-lenza di sedimenti fini. Dai risultati ottenuti sembra,invece, che le macrofite favoriscano abbondanze mag-giori poiché condizionano l’ambiente in modo da ren-derlo prevalentemente sedimentario, facilitando la de-posizione delle epifite e delle cellule planctoniche eriducendo la risospensione di quelle bentoniche. Que-sto è confermato dalle elevate quantità di C. scutellumrinvenute nei sedimenti superficiali rispetto a quelle
0
6
12
18
24
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
106 c
ell d
m-3
St. A (0.66±0.34)
St. B (2.38±3.06)
St. C (4.40±5.64)
Fig. 2a. Andamenti stagionali dell’abbondanza fitoplanctonica.Tra parentesi sono indicate medie e deviazioni standard.
2,0
2,5
3,0
3,5
4,0
4,5
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
H'
St. A (3.05±0.44)
St. B (3.19±0.37)
St. C (2.87±0.59)
Fig. 2b. Andamenti stagionali dell’indice di diversità di Shannonper il fitoplancton. Tra parentesi sono indicate medie e deviazionistandard.
0
1
2
3
4
5
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
106 c
ell c
m-3
St. A (1.41±0.92)
St. B (2.75±0.81)
St. C (1.85±0.63)
Fig. 3a. Andamenti stagionali dell’abbondanza delle diatomeebentoniche. Tra parentesi sono indicate medie e deviazionistandard.
1,4
1,9
2,4
2,9
3,4
3,9
4,4
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
H'
St. A (3.50±0.39)
St. B (3.68±0.31)
St. C (2.72±0.74)
Fig. 3b. Andamenti stagionali dell’indice di diversità di Shannonper le diatomee bentoniche. Tra parentesi sono indicate medie edeviazioni standard.

FACCA et al. - Fitoplancton e diatomee bentoniche in laguna di Venezia 23
Tab. I. Elenco floristico delle specie identificate nella colonna d’acqua e nel sedimento superficiale.
BACILLARIOPHYCEAE Nitzschia constricta RalfAchnanthes sp. Nitzschia cursoria GrunowAmphipleura micans Cleve Nitzschia dissipata GrunowAmphiprora paludosa Smith Nitzschia dissipata var. media GrunowAmphiprora sp. Nitzschia lanceolata SmithAmphora coffeaeformis Kützing Nitzschia lanceolata var. minor GrunowAmphora cymbelloides Grunow Nitzschia longa GrunowAmphora dubia Gregory Nitzschia longissima GrunowAmphora exigua Gregory Nitzschia lorenziana GrunowAmphora fluminensis Grunow Nitzschia lorenziana var. incurva GrunowAmphora cfr. macilenta Gregory Nitzschia macilenta GregoryAmphora truncata Cleve Nitzschia marina GrunowAmphora turgida Gregory Nitzschia microcephala GrunowAmphora veneta Kützing Nitzschia recta HantzschAmphora sp. Nitzschia sigma SmithAsterionella bleakeleyi Smith Nitzschia sigma var. rigida GrunowAsterionella sp. Nitzschia socialis GregoryAsterionellopsis glacialis Round Nitzschia tenuissima PeragalloBacillaria paradoxa Gmelin Nitzschia sp.Bacteriastrum delicatulum Cleve Pleurosigma australe GrunowCaloneis blanda Cleve Pleurosigma acuminatum GrunowCampyloneis grevillei Grunow Pleurosigma angulatum SmithCerataulina pelagica Hendey Pleurosigma decorum SmithChaetoceros affinis Lauder Pleurosigma ibericum PeragalloChaetoceros atlanticus Cleve Pleurosigma naviculaceum BrebissonChaetoceros decipiens Cleve Pleurosigma obscurum SmithChaetoceros lorenzianus Grunow Pleurosigma salinarum Cleve & GrunowChaetoceros mitra Cleve Pleurosigma speciosum SmithChaetoceros socialis Lauder Pleurosigma sp.Chaetoceros tenuissimus Meunier Proboscia alata SundströmChaetoceros sp. Psammodictyon panduriforme MannCocconeis molesta Kützing Pseudonitzschia fraudulenta HasleCocconeis scutellum Ehrenberg Pseudonitzschia heimii ManguinCocconeis sp. Pseudonitzschia lineola HasleCylindrotheca closterium Reimann & Lewin Pseudonitzschia seriata PeragalloDactyliosolen fragilissimus Hasle Pseudonitzschia subpacifica HasleDiploneis bombus Ehrenberg Pseudonitzschia sp.Diploneis chersonensis Cleve Rhopalodia musculus Otto MüllerDiploneis crabro Ehrenberg Rhizosolenia decipiens SundströmDiploneis hyalina Cleve Rhizosolenia striata GrevilleDiploneis sp. Rhizosolenia styliformis BrightwellEucampia groenlandica Cleve Rhizosolenia sp.Gomphonema olivaceum Kützing Skeletonema costatum CleveGrammatophora angulosa Ehrenberg Striatella delicatula GrunowGrammatophora oceanica Ehrenberg Striatella unipunctata AgardhGrammatophora sp. Surirella gemma EhrenbergGuinardia cylindrus Hasle Surirella ovata KützingGuinardia delicatula Hasle Surirella robusta EhrenbergGuinardia flaccida Peragallo Surirella sp.Guinardia striata Hasle Synedra formosa HantzschGyrosigma attenuatum Cleve Synedra pulchella KützingGyrosigma balticum Rabenhorst Synedra tabulata KützingGyrosigma distortum Griffith & Henfrey Synendra sp.Gyrosigma fasciola Griffith & Henfrey Thalassionema nitzschioides Van HeurckGyrosigma fasciola var. sulcata Cleve Thalassiosira sp.Gyrosigma spenceri Griffith & Henfrey Tropidoneis lepidoptera CleveHantzschia amphioxys Grunow Centriche indeterminateHantzschia hyalina Grunow Pennate indeterminateHemiaulus hauckii Grunow Bacillariophyceae indeterminate
(segue)

FACCA et al. - Fitoplancton e diatomee bentoniche in laguna di Venezia24
Licmophora abbreviata Agardh CHLOROPHYCEAELicmophora debilis Grunow Chlorophyceae indeterminateLicmophora flabellata Agardh CHRYSOPHYCEAELicmophora gracilis Grunow Chrysophyceae indeterminateLicmophora jurgensii Agardh CRYPTOPHYCEAELicmophora tenuis Grunow Cryptophyceae indeterminateLicmophora sp. DINOPHYCEAEMelosira nummuloides Agardh Alexandrium insuetum BalechMelosira sulcata Kützing Alexandrium sp.Melosira sp. Gyrodinium estuariale HulburtNavicula ammophila Grunow Gyrodinium sp.Navicula arenaria Donkin Prorocentrum arcuatum IsselNavicula arenicola Grunow Prorocentrum gracile SchüttNavicula cancellata Donkin Prorocentrum micans EhrenbergNavicula cincta Ralf Prorocentrum sp.Navicula clavata Gregory Dinoflagellati indeterminateNavicula cryptocephala Kützing EUGLENOPHYCEAENavicula inflexa Ralf Eutreptiella gymnasticaNavicula lanceolata Kützing NANOFLAGELLATINavicula latissima Gregory PRASINOPHYCEAENavicula sp. Micromonas sp.Nitzschia bilobata Smith Prasinophyceae indetrminateNitzschia clausii Hantzsch PRYMNESIOPHYCEAENitzschia commutata Grunow Prymnesiophyceae indeterminateNitzschia compressa Boyer
(continua)
incontrate nella colonna d’acqua delle St A e B. NellaSt. C, invece, oltre a mancare l’apporto delle epifite siè osservato che è prevalsa la risospensione visto che,ad es., nella colonna d’acqua sono state più abbondantile specie del genere Navicula con dimensioni di ca. 10μm, che una volta risospese hanno difficoltà maggioria sedimentare, rispetto a quelle di taglia di ca. 20-30μm, abbondanti nel sedimento. Il prevalere della riso-
spensione sulla sedimentazione è testimoniato anchedall’assenza, tra le diatomee bentoniche, delle formetipicamente planctoniche come Chaetoceros spp. eGuinardia spp. Al contrario, Amphora exigua Gre-gory, A. veneta Kützing, Psammodictyon pandurifor-me Mann e Surirella ovata Kützing, normalmentediffuse nel sedimento, sono state spesso trovate neicampioni d’acqua.
Bibliografia
FACCA C., SFRISO A., SOCAL G., 2001. Distribuzione spazio-temporale del fitoplancton nell’area centrale della laguna diVenezia. XXXII° Congresso SIBM, 4-9 giugno 2001, Nu-mana (AN).
FACCA C., SFRISO A., SOCAL G., 2002. Temporal and spatialdistribution of diatoms in the surface sediment of theVenice lagoon. Botanica Marina, 45: 170-183.
PERAGALLO M.M.H., PERAGALLO M., 1897-1908. DiatoméesMarines de France et des Districts Maritimes Voisins. In:(M.J. Tempere ed.), Grez-sur-Loing. 491 pp.
ROUND F.E., CRAWFORD R.M., MANN D.G., 1990. The Diato-ms. Biology & Morphology of the genera. Cambridge Uni-versity Press, Cambridge, 747 pp.
SFRISO A., 2000. Eutrofizzazione e inquinamento delle acque edei sedimenti nella parte centrale della laguna di Venezia.In: (CVN, MAV, LLPP, eds), Nuovi Interventi per laSalvaguardia di Venezia, Venezia. 3 vol., 258 pp.
SFRISO A., MARCOMINI A., 1996. Decline of Ulva growth in the
lagoon of Venice. Bioresource Technology 58: 299-307.SMYTH J.C., 1955. A study of the benthic diatoms of Loch
Sween (Argyll). Journal of Ecology 43: 149-171.TOLOMIO C., MOSCHIN E., MORO I., VALANDRO A., 2000. Ap-
proccio allo studio delle diatomee bentoniche nei sedimen-ti della laguna di Venezia. Ist. Ven. Sci., Lett. ed Arti, AttiConvegno SLV 2 (2): 1145-1153.
TOMAS C.R., 1993. Marine Phytoplankton. A Guide to NakedFlagellates and Coccolithophorids. Academic Press, SanDiego, 263 pp.
TOMAS C.R., 1996. Identifying Marine Diatoms and Dinoflagel-lates. Academic Press, San Diego, 598 pp.
UTHERMÖHL H., 1958. Zur Vervollkomnung der quantitati-ven Phytoplankton-Methodik. Mitt int Verein Limnol 9:1-38.
VANLANDINGHAM S.L., 1967-79. Catalogue of the fossil andrecent genera and species of diatoms and their synonyms.Vaduz: J. Cramer. 8 vol., 4654 pp.

Biologia Ambientale, 18 (2): 25-30, 2004
Proposta di un Indice dello Stato Ecologico delleComunità Ittiche viventi nelle acque interne italiane
Sergio ZerunianLaboratorio di Ittiologia delle Acque Dolci – Contrada Ponte la Vetrina, 04010 Maenza (LT). [email protected]
RiassuntoViene proposto un Indice dello Stato Ecologico delle Comunità Ittiche (ISECI) applicabile agli ecosistemi delle acque interne italiane, inprimo luogo per soddisfare quanto previsto dalla Direttiva 2000/60/CE “Quadro per l’azione comunitaria in materia di acque”. L’ISECItiene conto di due aspetti principali: 1) la naturalità della comunità, intesa come la normale ricchezza determinata dalla presenza di tuttele specie indigene attese in relazione al quadro zoogeografico ed ecologico e dall’assenza di specie aliene; 2) la buona condizione dellepopolazioni indigene, intesa come la capacità di autoriprodursi ed avere normali dinamiche ecologico-evolutive. Prima di applicareestesamente l’ISECI si propone una fase sperimentale al fine di saggiare l’indice e mettere a punto adeguate metodologie standardizzatedi campionamento.
PAROLE CHIAVE: Comunità ittiche / acque interne / indici biologici / stato ecologico / ISECI
Proposal of an Index of the Ecological State of the Fish Communities living in the Italian inland watersIt is proposed an Index of the Ecological State of the Fish Communities (ISECI) applicable to the ecosystems of the Italian inland waters,in the first place to fulfil the points of the Directive 2000/60/EC “Outline for the Community action on the subject of waters”. The indextakes into account two main aspects: 1) the natural condition of the community, understood as the normal richness determined by absenceof the alien species and the presence of all the native ones expected with regard to the zoogeographical and ecological framework; 2) thegood condition of the native populations, understood as the skill to self-reproducing and having the normal ecological-evolutionarydynamics. Before to apply extensively the index it is proposed an experimental stage in order to test it and to adjust standardized andadequate methodologies of sampling.
KEY WORDS: Fish communities / inland waters / biological index / ecological state / ISECI
INTRODUZIONELa Direttiva 2000/60/CE istituisce un Quadro per
l’azione comunitaria in materia di acque tale da “impe-dire un ulteriore deterioramento, proteggere e miglio-rare lo stato degli ecosistemi acquatici e degli ecosiste-mi terrestri e delle zone umide direttamente dipendentidagli ecosistemi acquatici sotto il profilo del fabbiso-gno idrico” (art. 1.a). Per la classificazione dello statoecologico delle acque superficiali (fiumi, laghi, acquedi transizione, acque costiere) individua tre tipologie dielementi qualitativi (allegato V): elementi biotici; ele-
menti idromorfologici, a sostegno degli elementi bioti-ci; elementi chimico-fisici, a sostegno degli elementibiotici. Risulta evidente il ruolo primario assegnatodalla Direttiva agli elementi biotici. Per questi ultimivengono anche individuati i gruppi tassonomico-eco-logici da prendere in considerazione: composizione,abbondanza e biomassa del fitoplancton (non per ifiumi); composizione e abbondanza della flora acqua-tica, macrofite e fitobentos; composizione e abbon-danza dei macroinvertebrati bentonici; composizione,
Nota della Redazione:in piena sintonia con lo spirito del CISBA, si pubblica questa “proposta di indice” nell’auspicio che possa stimolarne una fase di sperimentazionee di taratura, allo scopo di arrivare nel giro di qualche anno a formulare un “indice” standardizzato e largamente applicabile.

ZERUNIAN S. - Indice dello Stato Ecologico Comunità Ittiche26
abbondanza e struttura di età della fauna ittica (nonper le acque costiere).
Alcuni dei gruppi tassonomico-ecologici individuatidalla Direttiva 2000/60/CE vengono utilizzati da anniper valutare la qualità biologica di determinate tipologiedi ecosistemi delle acque interne, quali ad esempio icorsi d’acqua. Il gruppo usato con maggiore frequen-za e con i migliori risultati è sicuramente quello deimacroinvertebrati bentonici, per il quale da oltre ventianni è stata avviata nel nostro paese una approfonditadiscussione che ha portato alla messa a punto di unindice applicabile alle acque correnti italiane: l’IndiceBiotico Esteso (IBE); per l’applicazione dell’IBE esisteuna serie di manuali con finalità tassonomiche e meto-dologiche che rendono l’indice concretamente utilizza-bile nelle diverse realtà del nostro paese (per una sintesivedi GHETTI, 1997).
Per quanto riguarda la fauna ittica delle acque inter-ne, invece, non esiste in Italia alcun indice sintetico ingrado di permettere una valutazione dello stato bio-ecologico delle varie comunità; alcune ipotesi di lavorosono state formulate recentemente (SCARDI et al.,2002; FORNERIS et al., 2004), ma sono ancora moltolontane dal rappresentare strumenti operativi concreta-mente applicabili. Questa lacuna si riflette negativa-mente sulle attività di gestione delle acque interne, permolti aspetti assegnate per legge alle AmministrazioniProvinciali, e rende inapplicabili alcuni punti significa-tivi della Direttiva 2000/60/CE. Da queste motivazioninasce la presente proposta di un Indice dello StatoEcologico delle Comunità Ittiche (ISECI) applicabileagli ecosistemi delle acque interne italiane: fiumi (defi-niti, secondo l’art. 2 della Direttiva, “corpi idrici interniche scorrono prevalentemente in superficie ma chepossono essere parzialmente sotterranei”), laghi (“cor-pi idrici superficiali fermi”), e acque di transizione(“corpi idrici superficiali in prossimità delle foci deifiumi, che sono parzialmente di natura salina a causadella loro vicinanza alle acque costiere, ma sostanzial-mente influenzati dai flussi di acqua dolce”).
L’indice proposto vuole in primo luogo fornire unostrumento operativo per gli addetti al monitoraggioprevisto dalla Direttiva 2000/60/CE. È inoltre comple-mentare agli altri indicatori della qualità biologica degliecosistemi acquatici, contribuendo così a produrre“immagini” sempre più rappresentative della realtà.Infine, può risultare utile alle Amministrazioni deputatealla gestione delle acque interne nell’ottica del lorobuon governo e, più in generale, di una razionalepolitica ambientale.
MATERIALI E METODIScopo principale dell’ISECI è la valutazione dello
stato di una determinata comunità di pesci delle acque
interne (per una definizione di questa categoria tasso-nomico-ecologica vedi ZERUNIAN, 2002), secondo quan-to previsto dalla Direttiva 2000/60/CE che individuaper la fauna ittica tre diversi livelli di “stato ecologico”:elevato, buono, sufficiente. Nell’indice proposto talevalutazione viene effettuata tenendo conto di dueaspetti principali: 1) la naturalità delle comunità, intesacome la normale ricchezza determinata dalla presenzadi tutte le specie indigene attese in relazione al quadrozoogeografico ed ecologico e dall’assenza di speciealiene; 2) la buona condizione delle popolazioni indige-ne, intesa come la capacità di autoriprodursi ed averenormali dinamiche ecologico-evolutive. La situazioneottimale, rappresentata dal valore massimo dell’ISECI(20), individua la condizione naturale di riferimentonella stima/misurazione della qualità ambientale.
L’ISECI propone di valutare lo stato ecologico diuna comunità ittica procedendo nel modo seguente.1. Mediante analisi cartografica ed idonei sopralluoghi
sul campo si individua una serie di stazioni di campio-namento, rappresentativa delle diverse situazioni eco-
Tab. I. Metodi di campionamento
ELETTROSTORDITORE
a) con gli operatori in acqua se la profondità dell’area in cui sieffettua il campionamento è inferiore a 1,20 m (piccoli corsid’acqua, aree spondali di medi corsi d’acqua, risorgive, areecircumlacuali con bassa profondità di bacini d’acqua dolce, ecc.);b) con gli operatori in barca se la profondità dell’area in cui sieffettua il campionamento è compreso tra 1,20 e 2,50 m (areespondali di medi e grandi corsi d’acqua, aree circumlacuali conmedia profondità di bacini d’acqua dolce, ecc.).
Il campionamento può essere effettuato sia con strumento acorrente continua pulsata sia con strumento ad impulsi, mediantepassaggi ripetuti in settori preventivamente individuati e per untempo non inferiore a 60 minuti per ciascuna stazione
RETI (barracuda, sciabica, tramaglio, bilancia, martavello, ecc.)
Da utilizzare nelle seguenti tipologie ambientali: aree profonde dimedi e grandi corsi d’acqua, comprese le acque di transizione;aree di laghi d’acqua dolce dove la profondità, superiore a 2,50 m,non consente l’uso dell’elettrostorditore e dove la comunità itticapotenzialmente presente comprenda specie pelagiche di acquelibere (Alburnus alburnus alborella, Salmo carpio, Coregonusspp., ecc.) e/o specie bentoniche di acque profonde (Salvelinusalpinus, Lota lota, ecc.); laghi costieri.
Il campionamento con barracuda, sciabiche e bilance va effettuatoper un tempo non inferiore a 60 minuti per ciascuna stazione; lereti da posta (tramagli e martavelli) devono restare in opera per8-10 ore, dalla sera al mattino successivo. Se nel fiume o lago inesame esiste attività di pesca professionale, è preferibile fareseguire il campionamento da pescatori locali.

ZERUNIAN S. - Indice dello Stato Ecologico Comunità Ittiche 27
logiche del corpo idrico in esame. Il numero dellestazioni sarà in funzione delle dimensioni del corpoidrico e della scala a cui viene condotta l’indagine;all’interno di un sistema sensibile o di particolarevalore conservazionistico, come ad esempio un’areaprotetta, è necessario un numero di stazioni propor-zionalmente più alto.
2. Per ciascuna stazione di campionamento si individuain via teorica la comunità ittica attesa, con riferimen-to alle sole specie indigene e tenendo conto deiseguenti elementi: a) distribuzione delle specie (inrelazione al quadro zoogeografico nazionale di tutti itaxa presenti nelle acque interne italiane); b) ecologiadelle specie; c) periodo del campionamento (in rela-zione alla possibile presenza degli stadi adulti di speciemigratrici o, in alternativa, dei relativi stadi larvali ogiovanili). Tutti i dati necessari all’individuazionedella comunità ittica attesa sono reperibili in lavori disintesi come ad esempio il volume allegato all’ “Ico-nografia dei Pesci delle acque interne d’Italia” (ZERU-NIAN e DE RUOSI, 2002) e il volume “Pesci delle acqueinterne d’Italia” (ZERUNIAN, 2004a).
3. Si eseguono i campionamenti, che devono avere: (1)frequenza di almeno due volte l’anno, di cui una nelperiodo tardo primaverile (da metà aprile a metàgiugno) per valutare l’eventuale presenza di speciemigratrici in fase riproduttiva oppure dei relativi stadilarvali o giovanili, e (2) metodologia basata, dovepossibile, sull’uso dell’elettrostorditore; in alcunetipologie ambientali è necessario l’uso di strumenti dicattura (reti) propri della pesca professionale (Tab.I). Per le specie catturate si ritiene sufficiente l’appli-cazione di un indice di abbondanza semiquantitativo,come quello di Moyle per le acque correnti, medianteil quale viene attribuita una delle seguenti categorie:scarso (1-3 individui in 50 m lineari), presente (4-10individui …), frequente (11-20 individui …), abbon-dante (21-50 individui …), dominante (più di 50individui …); esempi di applicazione di questo indicealle acque interne italiane sono riportati da TURIN et al.(1999)
4. Si procede alla determinazione del materiale campio-nato, con il fine di individuare la comunità itticareale. Per la classificazione delle specie indigenepossono essere utilizzate le chiavi dicotomiche con-tenute nel volume “Pesci delle acque interne d’Italia”(ZERUNIAN, 2004a); per la maggior parte delle speciealiene si può fare riferimento al volume “I Pesci delleacque interne italiane” (GANDOLFI et al., 1991).
5. Limitatamente ai taxa indigeni, su un sub-campionerappresentativo dei due sessi e delle diverse taglie, sirileva una serie di parametri biologici qualitativi (fe-notipo) e quantitativi (lunghezza totale, lunghezzastandard, peso totale, peso delle gonadi, età) neces-
sari per effettuare una valutazione delle condizionibiologiche delle popolazioni indigene; per questavalutazione si tiene anche conto della consistenzademografica di ciascuna popolazione, stimata con unindice di abbondanza semiquantitativo (si veda ilpunto 3).
6. Si calcola il valore dell’ISECI utilizzando una tabellaa doppia entrata che tiene conto della composizionedella comunità e della condizione biologica dellepopolazioni indigene (Tab. II).
7. Si procede alla conversione dei valori dell’ISECI inlivelli di stato ecologico delle comunità ittiche, chevanno da I a V (Tab. III). I primi tre livelli (elevato,buono, sufficiente) corrispondono a quelli individuatidalla Direttiva 2000/60/CE. L’ISECI individua altridue livelli (scadente, pessimo) fornendo così unmaggior dettaglio per il livello “insufficiente” implici-tamente previsto dalla Direttiva.
DISCUSSIONEIl primo elemento considerato dall’ISECI nella valu-
tazione dello stato ecologico di una comunità ittica è lacomposizione della comunità (primo ingresso, oriz-zontale, in Tab. II); ciò è in accordo con la Direttiva2000/60/CE, che attribuisce il migliore stato ecologicoa un corpo d’acqua dove la fauna ittica abbia “compo-sizione e abbondanza delle specie che corrispondonototalmente o quasi alle condizioni inalterate; presenzadi tutte le specie sensibili alle alterazioni tipiche specifi-che”. Nella proposta sintetizzata in Tabella II si prefi-gurano 5 condizioni riferibili al gruppo delle specieindigene e 3 riferibili a quello delle specie aliene. L’indi-ce assegna quindi valori tanto più alti quanto più lacomunità risulta naturale. La condizione ottimale diriferimento (presenza di tutte le specie indigene attesee assenza di specie aliene) risulta ormai rara in Italia,così come nella gran parte d’Europa: estinzioni locali eintroduzioni di specie aliene hanno infatti modificato inmisura più o meno profonda le comunità ittiche delleacque interne del continente (CRIVELLI e MAITLAND,1995; LELEK, 1996). In considerazione dell’alto nume-ro di taxa endemici nel nostro paese e della necessità diconcrete misure finalizzate alla loro conservazione, ènecessario puntare mediante opportune azioni a con-servare o ripristinare la normale ricchezza biologicadelle comunità ittiche (per una discussione più appro-fondita vedi ZERUNIAN, 2003). Azioni di miglioramentodelle condizioni alterate, come può essere consideratala ricostituzione mediante reintroduzioni di una comu-nità ittica depauperata da estinzioni locali, sono previ-ste dalla Direttiva 2000/60/CE. L’attribuzione di valoriprogressivamente più bassi dell’ISECI in presenza dispecie aliene, soprattuto se ad alto impatto sulle comu-nità indigene come nel caso del Siluro (Silurus glanis),

ZERUNIAN S. - Indice dello Stato Ecologico Comunità Ittiche28
vuole evidenziare la negatività di queste presenze estimolare l’adozione di tutte le misure possibili perevitare il dilagare del fenomeno delle introduzioni fau-nistiche e delle immissioni accidentali (vedi ZERUNIAN,2002).
Il secondo elemento considerato dall’ISECI nellavalutazione dello stato ecologico di una comunità itticaè la condizione biologica delle popolazioni indigene(secondo ingresso, verticale, in Tab. II); anche questoè in accordo con la Direttiva 2000/60/CE, che attribu-
isce il migliore stato ecologico a un corpo d’acquadove la fauna ittica abbia “strutture di età delle comuni-tà ittiche che presentano segni minimi di alterazioniantropiche e non indicano l’incapacità a riprodursi o asvilupparsi di specie particolari”. Nella proposta sinte-tizzata nella tabella II si prefigurano 4 condizioni (A-D)che tengono conto dei seguenti elementi: struttura inclassi di età delle popolazioni indigene, loro consisten-za demografica, possibile ibridazione delle popolazioniindigene appartenenti ai generi Salmo, Thymallus, Esox
Tab. II. Calcolo del valore dell’ISECI.
1) Silurus glanis2) Petromyzon marinus, Lampetra fluviatilis, Acipenser sturio, Huso huso3) Lampetra zanandreai, Acipenser naccarii, Rutilus rubilio, R. erythrophthalmus, Leuciscus souffia muticellus, Alburnus
alburnus alborella, A. albidus, Chondrostoma soetta, Ch. genei, Barbus plebejus, B. meridionalis caninus, Cobitistaenia bilineata, Sabanejewia larvata, Salmo (trutta) marmoratus, S. (trutta) macrostigma, S. fibreni, S. carpio,Pomatoschistus canestrini, Knipowitschia panizzae, K. punctatissima, Padogobius martensii, Gobius nigricans
A) Tutte le popolazioni ben strutturate in classi di età e mostranti una sufficiente o buona consistenza demografica(presente, frequente, abbondante o dominante). Popolazioni dei generi Salmo, Thymallus, Esox e Barbus nonibride con popolazioni alloctone.
B) Tutte le popolazioni ben strutturate in classi di età e mostranti una sufficiente o buona consistenza demografica.Una o più popolazioni dei generi Salmo, Thymallus, Esox e Barbus ibride con popolazioni alloctone.
C) La maggior parte (più del 50%) delle popolazioni ben strutturate in classi di età e mostranti una sufficiente o buonaconsistenza demografica.
D) Il 50% o meno delle popolazioni ben strutturate in classi di età e mostranti una sufficiente o buona consistenzademografica.
COMPOSIZIONE DELLA COMUNITÀ (primo ingresso, orizzontale)
CONDIZIONE BIOLOGICA delle popolazioni indigene (secondo ingresso, verticale)
Specie indigene Specie aliene A B C D Assenti 20 18 16 14 Presenti una-due; Siluro¹ assente 19 17 15 13 Presenti tutte quelle attese
Presenti più di due o il Siluro 18 16 14 12 Assenti 18 16 14 12 Presenti una-due; Siluro¹ assente 17 15 13 11
Presenti tutte quelle attese, tranne i Ciclostomi e gli Acipenseridi migratori² Presenti più di due o il Siluro 16 14 12 10
Assenti 16 14 12 10
Presenti una-due; Siluro¹ assente 15 13 11 9
Presenti la maggior parte (più del 50%) di quelle attese, tranne i Ciclostomi e gli Acipenseridi migratori e taxa non endemici in Italia Presenti più di due o il Siluro 14 12 10 8
Assenti 14 12 10 8 Presenti una-due; Siluro¹ assente 13 11 9 7
Presenti la maggior parte (più del 50%) di quelle attese, tranne i Ciclostomi e gli Acipenseridi migratori e taxa endemici in Italia³
Presenti più di due o il Siluro 12 10 8 6
Assenti 12 10 8 6 Presenti una-due; Siluro¹ assente 11 9 7 5 Presenti il 50% o meno di
quelle attese Presenti più di due o il Siluro 10 8 6 4
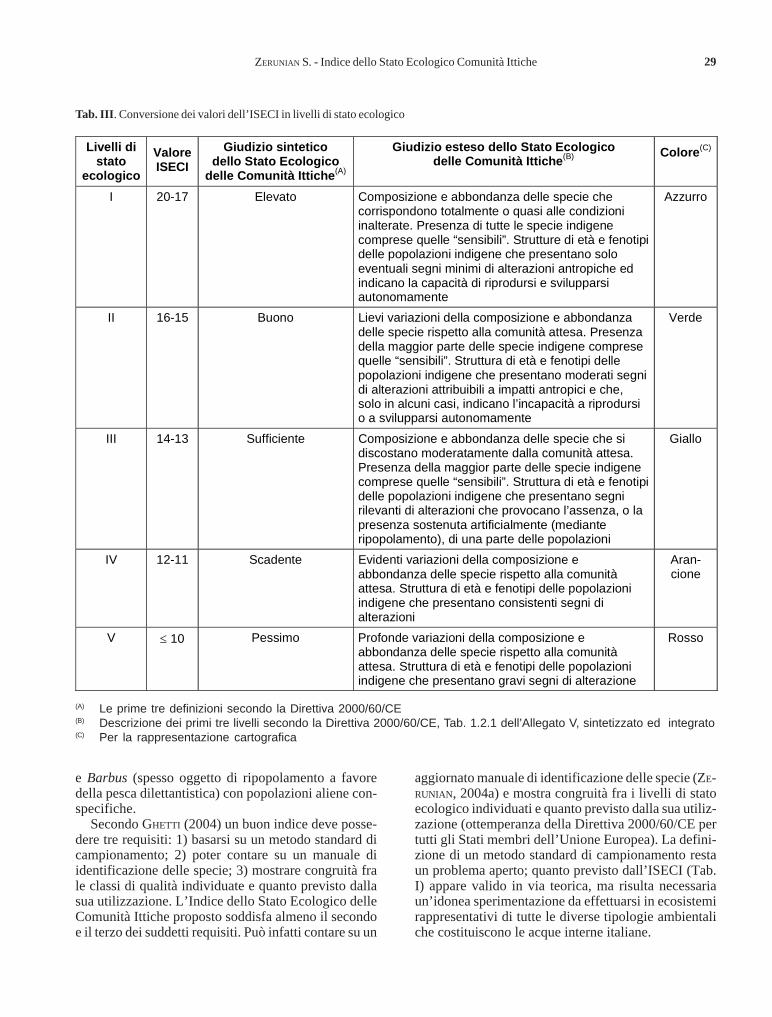
ZERUNIAN S. - Indice dello Stato Ecologico Comunità Ittiche 29
e Barbus (spesso oggetto di ripopolamento a favoredella pesca dilettantistica) con popolazioni aliene con-specifiche.
Secondo GHETTI (2004) un buon indice deve posse-dere tre requisiti: 1) basarsi su un metodo standard dicampionamento; 2) poter contare su un manuale diidentificazione delle specie; 3) mostrare congruità frale classi di qualità individuate e quanto previsto dallasua utilizzazione. L’Indice dello Stato Ecologico delleComunità Ittiche proposto soddisfa almeno il secondoe il terzo dei suddetti requisiti. Può infatti contare su un
aggiornato manuale di identificazione delle specie (ZE-RUNIAN, 2004a) e mostra congruità fra i livelli di statoecologico individuati e quanto previsto dalla sua utiliz-zazione (ottemperanza della Direttiva 2000/60/CE pertutti gli Stati membri dell’Unione Europea). La defini-zione di un metodo standard di campionamento restaun problema aperto; quanto previsto dall’ISECI (Tab.I) appare valido in via teorica, ma risulta necessariaun’idonea sperimentazione da effettuarsi in ecosistemirappresentativi di tutte le diverse tipologie ambientaliche costituiscono le acque interne italiane.
Tab. III. Conversione dei valori dell’ISECI in livelli di stato ecologico
(A) Le prime tre definizioni secondo la Direttiva 2000/60/CE(B) Descrizione dei primi tre livelli secondo la Direttiva 2000/60/CE, Tab. 1.2.1 dell’Allegato V, sintetizzato ed integrato(C) Per la rappresentazione cartografica
Livelli di stato
ecologico
Valore ISECI
Giudizio sintetico dello Stato Ecologico
delle Comunità Ittiche(A)
Giudizio esteso dello Stato Ecologico delle Comunità Ittiche(B)
Colore(C)
I 20-17 Elevato Composizione e abbondanza delle specie che corrispondono totalmente o quasi alle condizioni inalterate. Presenza di tutte le specie indigene comprese quelle “sensibili”. Strutture di età e fenotipi delle popolazioni indigene che presentano solo eventuali segni minimi di alterazioni antropiche ed indicano la capacità di riprodursi e svilupparsi autonomamente
Azzurro
II 16-15 Buono Lievi variazioni della composizione e abbondanza delle specie rispetto alla comunità attesa. Presenza della maggior parte delle specie indigene comprese quelle “sensibili”. Struttura di età e fenotipi delle popolazioni indigene che presentano moderati segni di alterazioni attribuibili a impatti antropici e che, solo in alcuni casi, indicano l’incapacità a riprodursi o a svilupparsi autonomamente
Verde
III 14-13 Sufficiente Composizione e abbondanza delle specie che si discostano moderatamente dalla comunità attesa. Presenza della maggior parte delle specie indigene comprese quelle “sensibili”. Struttura di età e fenotipi delle popolazioni indigene che presentano segni rilevanti di alterazioni che provocano l’assenza, o la presenza sostenuta artificialmente (mediante ripopolamento), di una parte delle popolazioni
Giallo
IV 12-11 Scadente Evidenti variazioni della composizione e abbondanza delle specie rispetto alla comunità attesa. Struttura di età e fenotipi delle popolazioni indigene che presentano consistenti segni di alterazioni
Aran-cione
V ≤ 10 Pessimo Profonde variazioni della composizione e abbondanza delle specie rispetto alla comunità attesa. Struttura di età e fenotipi delle popolazioni indigene che presentano gravi segni di alterazione
Rosso

ZERUNIAN S. - Indice dello Stato Ecologico Comunità Ittiche30
CONCLUSIONIL’Indice dello Stato Ecologico delle Comunità Itti-
che colma una evidente lacuna fra gli indicatori biolo-gici utilizzati negli ecosistemi delle acque interne italia-ne. Rappresenta così uno strumento che permette direalizzare pienamente il monitoraggio previsto dallaDirettiva 2000/60/CE. Prima di poter applicare con-cretamente l’ISECI è però necessaria una fase speri-mentale volta a saggiare e validare l’indice e mettere apunto adeguate metodologie standardizzate di campio-namento. È auspicabile che tale fase sperimentale iniziquanto prima, possibilmente con la supervisione di unEnte Nazionale (quale l’Agenzia per la Protezione del-l’Ambiente ed i servizi Tecnici).
In considerazione del ritardo già accumulato dalnostro paese nell’applicazione della Direttiva 2000/60/CE, è necessario affrontare con chiarezza di intentiogni problematica legata all’uso delle comunità ittichenel monitoraggio delle acque interne; bisogna quindiporsi e rispettare precise scadenze che, tenendo contodi quanto previsto dalla Direttiva, potrebbero essere leseguenti (ZERUNIAN, 2004b, modificato ed integrato):- entro il 2004 individuazione delle comunità ittiche
originarie in ogni distretto idrografico e delle speciedi interesse comunitario che ne fanno parte (AllegatoII della Direttiva 92/43/CEE);
- entro il 2006 completamento della fase sperimentalein cui saggiare e validare l’ISECI in un campione
rappresentativo di ecosistemi delle acque interneitaliane; redazione di un manuale di applicazionedell’ISECI;
- entro il 2008 redazione di piani d’azione finalizzatialla conservazione dell’ittiofauna, a livello di distrettiidrografici (con particolare riferimento alle speciedell’Allegato II della Direttiva 92/43/CEE); individua-zione della rete di monitoraggio e prima raccolta didati sulle comunità ittiche (momento zero);
- entro il 2010 avviamento della rimozione delle causeche hanno determinato eventuali estinzioni locali dispecie ittiche; reintroduzioni faunistiche delle speciesensibili (stenoecie) eventualmente estintesi nei sin-goli distretti idrografici; redazione di piani di gestionedelle comunità ittiche; proseguimento del monitorag-gio;
- entro il 2012 proseguimento della rimozione dellecause che hanno determinato eventuali estinzionilocali di specie ittiche; verifica del successo dellereintroduzioni faunistiche e nuovi tentativi in caso diinsuccesso; proseguimento del monitoraggio.
RINGRAZIAMENTIL’Autore desidera ringraziare Giuseppe Sansoni e Paolo Turinper le critiche costruttive formulate a una prima stesura dell’In-dice, che hanno permesso di migliorare e rendere più chiari alcuniaspetti.
BIBLIOGRAFIA
mediante reti neurali artificiali. In: “Atti IX Conv. Naz.AIIAD”, pubbl. in Biologia Ambientale, 18 (1): 25-31.
TURIN P., MAIO G., ZANETTI M., BILÒ M.F., ROSSI V., SALVIATI
S., 1999. Carta ittica delle acque dolci interne. Prov. diRovigo, 326 pp.
ZERUNIAN S., 2002. Condannati all’estinzione? Biodiversità,biologia, minacce e strategie di conservazione dei Pescid’acqua dolce indigeni in Italia. Edagricole, Bologna, X +220 pp.
ZERUNIAN S., DE RUOSI T., 2002. Iconografia dei Pesci delleacque interne d’Italia / Iconography of Italian Inland WaterFishes. Min. Ambiente - Ist. Naz. Fauna Selvatica, 263 pp.+ 33 tavv.
ZERUNIAN S., 2003. Piano d’azione generale per la conserva-zione dei Pesci d’acqua dolce italiani. Quad. Cons. Natura,17, Min. Ambiente - Ist. Naz. Fauna Selvatica, 123 pp.
ZERUNIAN S., 2004a. Pesci delle acque interne d’Italia. Min.Ambiente - Ist. Naz. Fauna Selvatica, 258 pp + CD-Rom.
ZERUNIAN S., 2004b. Ruolo della comunità ittica nell’applica-zione della Direttiva Quadro. Atti Seminario “Classifica-zione Ecologica delle Acque interne” (Trento, 12-13.02.2004). Prov. Aut. Trento - CISBA, in stampa.
CRIVELLI A.J., MAITLAND P.S. (eds.), 1995. Endemic Freshwa-ter Fishes of the Northern Mediterranean Region. Biol.Conserv., 72: 121-337.
FORNERIS G., MERATI F., PASCALE M., PEROSINO G.C., 2004.Proposta di un Indice Ittico per il bacino occidentale del Po.Abstract X Congr. Naz. AIIAD: 31.
GANDOLFI G., ZERUNIAN S., TORRICELLI P., MARCONATO E., 1991.I Pesci delle acque interne italiane. Ist. Poligr. e Zecca delloStato, Roma, XVI + 617 pp.
GHETTI P.F., 1997. Indice Biotico Esteso (IBE), i macroinverte-brati nel controllo della qualità degli ambienti di acquecorrenti. Prov. Aut. di Trento, 222 pp.
GHETTI P.F., 2004. Conclusioni. Atti Seminario “Classificazio-ne Ecologica delle Acque interne” (Trento, 12-13. 02.2004).Prov. Aut. Trento - CISBA, in stampa.
LELEK A., 1996. General consideration regarding the decline ofspecies. In: Conservation of Endengered Freshwater Fishin Europe (Kirchhofer A., Hefti D. eds.), Birkhäuser Verlag,Basel / Switzerland: 1-7.
SCARDI M., CATAUDELLA S., CICCOTTI E., DI DATO P., MAIO G.,MARCONATO E., SALVIATI S., TANCIONI L., TURIN P., ZANETTI
M., 2002. Previsione della composizione della fauna ittica

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 31
Informazione&Documentazione
Applicazione della metodica di indagine biologica,basata sulla fauna bentonica e sulla componenteorganica del sedimento, in acque di sorgente edemergenze assimilabili
Silvio Gaiter, Marco Bodon, Daniela Rocca*
Agenzia Regionale per la Protezione dell’Ambiente Ligure, Dipartimento di Genova, Via Gropallo 5 - 16121 Genova
* Autore referente per la corrispondenza ([email protected])
INTRODUZIONEDalla fine degli anni ’80 sono
state intraprese dal Presidio Multi-zonale di Prevenzione di Genova(PMP), poi Agenzia Regionale perla Protezione dell’Ambiente Ligu-re (ARPAL), indagini e studi sulmondo biologico sotterraneo e sor-givo dell’Appennino Ligure, svoltianche in collaborazione con l’Isti-tuto di Zoologia dell’Università diGenova, attualmente Dipartimen-to per lo Studio del Territorio edelle sue Risorse (DIP.TE.RIS.). Leesperienze si sono subito orientateverso la conoscenza delle biocenosie la lettura delle stesse in relazioneagli aspetti di alterazione ambien-tale e di rischio igienico delle ac-que. Nel 1989 è stata stesa unaprima descrizione delle metodichedi campionamento e analisi (BO-DON e GAITER, 1989), nella quale sidelineano le informazioni che pos-sono essere tratte dallo studio diqueste biocenosi; successivamente,una aumentata esperienza appli-cativa ha permesso di definire me-glio il quadro conoscitivo del bioto-po sorgente. Sono stati quindi pre-sentati i risultati di differenti situa-zioni faunistiche e alcuni criteriinterpretativi, applicati ad alcune
realtà territoriali (BODON e GAITER,1995; GAITER et al., 1995). In questiultimi anni sono state sviluppatealtre iniziative parallele, fra le qualila realizzazione di un database su-gli ambienti sorgivi (con dati bioce-notici, ambientali, fisico-chimici emicrobiologici), la ricerca e sche-datura dei lavori scientifici italianied europei e la sintesi delle infor-mazioni ecologiche e biogeografi-che sui singoli taxa. Gli sforzi sisono concentrati sul significato diindicatore degli organismi del ben-thos in relazione ai differenti aspettidella risorsa idrica e hanno porta-to alla stesura di un set di descritto-ri che permettono di formulare ipo-tesi sulla tipologia delle acque (ori-gine dell’acquifero alimentante),grado di vulnerabilità del sito d’in-dagine, in particolare per risorsecaptate, e grado di inquinamento odi alterazione delle acque.
L’elaborazione degli indica-tori è basata sul documento pro-dotto, nel 1999, per il Centro Te-matico Nazionale Acque Interne eMarino Costiere - CTN_AIM (BO-DON e RAFFETTO, 1999), proposto,con opportune modifiche e inte-grazioni, in occasione del corso di
formazione “Metazoi delle acquesorgive e sotterranee”, tenutosi aGenova nel settembre 2000 (ROCCA
et al., 2003). Il metodo è in fasesperimentale ed è stato applicatosu situazioni e realtà geografichelimitate (oltre 500 sorgenti e am-bienti assimilabili per un totale dicirca 900 siti di campionamentodell’Appennino Ligure). Il conte-nuto della tabella applicativa è solouna prima proposta, da sperimen-tare e perfezionare, destinata a su-bire certamente modifiche con l’ac-quisizione di migliori conoscenzeecologiche e biogeografiche sui sin-goli taxa, ancora molto limitate eristrette a piccole aree del territo-rio italiano. Un buon contributopotrebbe giungere dalle iniziativedi studio che si svilupperanno inItalia a seguito del progetto PA-SCALIS (Protocols for the ASsess-ment and Conservation of AquaticLIfe in the Subsurface) nell’ambi-to del programma quadro per laricerca e lo sviluppo tecnologicodella Commissione Europea (GA-LASSI, 2002; MALARD et al., 2003).
La metodica proposta nelpresente lavoro è stata impostatasecondo la traccia descritta nelle

Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)32
Informazione&Documentazione
procedure previste per i metodi diprova di laboratorio dal ServizioAssicurazione Qualità di ARPAL(ARPAL, 2000). La procedura ap-pare, quindi, piuttosto schematicae rigida, ma la standardizzazioneed omologazione del metodo puòrappresentare una buona spintaatta a favorire la diffusione dellasperimentazione dell’indagine bio-logica.
GENERALITÀLo scopo che si prefigge l’in-
dagine biologica in acque sorgive,cioè in ambienti di passaggio fraacque sotterranee e acque superfi-ciali (ecotoni acqua-acqua a dina-miche forti; GILBERT et al., 1990), èquello di formulare diagnosi sul-l’origine e la qualità delle acque diquesti siti peculiari. Questo tipo diapproccio si basa sull’esame siadella componente faunistica –co-stituita da organismi invertebratiappartenenti quasi esclusivamenteal benthos di acque dolci (che vivo-no a livello del fondo e nello stratopiù superficiale del sedimento)– siadella componente organica del se-dimento, che comprende resti ve-getali e animali. La lettura attentadella biocenosi e della tanatoceno-si, può fornire utili elementi percompletare il quadro conoscitivodella risorsa idrica rilevato da altriesami e analisi.
Le attività previste per l’ap-plicazione della metodica possonoessere raggruppate in due fasi prin-cipali: la fase di campo, in cui sieffettuano il campionamento bio-logico e il rilevamento dei parame-tri di campagna accessori, e la fasedi laboratorio, in cui si procedealla preparazione e separazione delmateriale raccolto, al riconoscimen-to dei taxa, all’elaborazione dei datie alla sintesi dei risultati.
CAMPO DI APPLICAZIONELa metodologia è applicabi-
le a diverse tipologie di sorgente e,in generale, ad ambienti di acquesotterranee. È idonea per emergen-ze naturali, ovvero siti che non han-no subito interventi di modifica-zione, sorgenti parzialmente modi-ficate ma che conservano ancoracaratteristiche naturali e ancheemergenze captate, in parte o to-talmente, con manufatto chiuso oaperto, parzialmente o completa-mente. L’ampia gamma dei siti diinvestigazione è quindi riconduci-bile a due tipologie fondamentali:emergenze idriche in siti naturalied emergenze idriche in siti artifi-cializzati. A questi si aggiungono,per le loro caratteristiche fisico-chi-miche e biologiche, alcuni ambien-ti idrici sotterranei assimilabili, suiquali è possibile applicare, con cau-tela, la metodica, come cavità car-siche naturali e cavità artificiali,originate in seguito ad interventiindotti nei complessi rocciosi con-tenenti serbatoi acquiferi (gallerieferroviarie, autostradali, miniere,cave, etc.). La metodica proposta èapplicabile, con riserva, anche agliambienti di falda alluvionale (poz-zi), solo quando la conformazionedel lume e la modesta profonditàlo consentono. Il presente elabora-to e la relativa modellistica si riferi-scono comunque ai soli ambientisorgivi o assimilabili. Le tipologieambientali e i relativi siti di possi-bile applicazione vengono schema-tizzate nell’All. 1.
Questa metodologia d’inda-gine non è invece applicabile adalcune tipologie particolari di sor-gente, come acque che provengonodalla fusione di ghiacciai, sorgentitermali, sorgenti minerali con chi-mismo particolare, sorgenti sulfu-ree, sorgenti temporanee.
PRINCIPIO DEL METODOQuesto tipo di indagine vie-
ne svolta prevalentemente sul cam-po, come per altre discipline profes-
sionali (geologia ambientale, fisicaambientale, etc.), dove è necessarioeffettuare una lettura attenta del-l’ecosistema, o di una parte di esso.Il metodo è applicabile –da partedi operatori aventi una adeguataformazione in campo ecologico,idrobiologico e tassonomico– dopoun periodo di tirocinio sotto la gui-da di personale esperto o mediantespecifici corsi di formazione. Lateoria che sta alla base di questotipo di metodologia presuppone chele condizioni di un ecosistema pos-sano essere valutate mediante lamisura di parametri propri dellacomunità biotica e delle sue com-ponenti, da parte di personale qua-lificato.
PRECAUZIONIIl pericolo a cui l’operatore
va incontro è principalmente lega-to all’attività svolta sul campo, poi-ché in laboratorio l’utilizzo di agen-ti chimici che possano arrecare dan-no è modesto e si riduce al soloutilizzo di piccole quantità di al-cool per la fissazione e la conserva-zione dei campioni. I principali ri-schi, riconducibili all’attività sulcampo, sono:– fenomeni climatici;– morfologia del territorio;– sollevamento e trasporto della
strumentazione durante il per-corso a piedi;
– agibilità all’interno del manu-fatto di captazione o delle cavi-tà naturali e artificiali.
In generale, l’operatore devepoter disporre dell’idonea attrez-zatura individuale (casco protetti-vo o da minatore, guanti, giacconee copri pantaloni impermeabili, cal-zature quali scarpe da trekking estivali) e di una o più torce elettri-che. I dispositivi di protezione col-lettiva (corde e moschettoni, scalet-ta portatile) sono indispensabili alnucleo di lavoro in alcune specifi-che situazioni. Sono inoltre oppor-

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 33
Informazione&Documentazione
tune la presenza di un numero suf-ficiente di persone, in relazione allaquantità di materiale da trasporta-re, e una conoscenza specifica delterritorio da investigare. Nel casodi indagini in cavità artificiali onaturali è necessario disporre diuna guida abilitata.
INTERFERENZECiò che può influenzare il
risultato della prova, è legato pre-valentemente alla fase di campo edipende soprattutto dall’alterazio-ne dell’habitat. Un’alterazione di-retta sui siti di colonizzazione de-termina una variazione quali-quan-titativa della biocenosi fino allacompleta distruzione della stessa. Ifattori d’interferenza possono esse-re ricondotti a tre tipologie princi-pali:– eventi alluvionali che interessa-
no il sito sorgivo o parte di esso;– impiego di sostanze disinfettan-
ti (es. a base di cloro o calceviva) all’interno del sito di cam-pionamento, nel caso di emer-genze captate;
– interventi di asportazione deisedimenti da parte degli addettialla manutenzione delle operedi captazione delle emergenzeidriche.
Sono ovviamente esclusi ifenomeni di inquinamento indiret-to delle acque che il metodo si pro-pone di valutare.
APPARECCHIATURE
Attrezzature di campo (Fig. 1):– retino triangolare da benthos per
raccolta diretta, con breve im-pugnatura, di piccole dimensio-ni (20-25 cm di lato), e con retefiltrante a sacco in nylon, con-nessa all’anello metallico (adat-tabile al substrato) mediantetelo robusto; a maglie opportu-ne:– 500 μm per macrobenthos;
– 100 μm per meio- e micro-benthos;
– retino da drift, con rete filtrantein nylon a maglie opportune(500 μm o 100 μm), dotato diun collettore terminale svitabi-le, utilizzabile in alcune parti-colari situazioni;
– manico allungabile da adattareai retini, per prelievi in acqueprofonde (fino a 3 m ca.);
– paletta metallica con impugna-tura in plastica per smuovere ilsedimento o per raccoglierlo;
– pinzette da entomologo per laraccolta a vista degli organismidi maggiori dimensioni;
– sacchetti e contenitori per laconservazione temporanea delcampione;
– matita ed etichette;– torcia elettrica con illuminazio-
ne potente;– borsa termica per il trasporto
dei campioni (temperatura diconservazione 3-5 °C);
– piastre frigorifere.
Attrezzature di laboratorio:– frigorifero per la conservazione
dei campioni;– secchio in plastica di capacità 5
litri;– contenitori graduati di varie ca-
pacità;– setacci in acciaio di 12-13 cm di
diametro, a maglia opportuna:– 4 mm (UNI n° 1);– 2 mm (UNI n° 7);– 1 mm (UNI n° 13);– 0,5 mm (UNI n° 20);
– ritaglio di rete in nylon da 100μm (20-30 cm ca. di lato).
– cucchiaio;– capsule Petri di 5-10 cm di dia-
metro;– microscopio stereoscopico, a bas-
si e medi ingrandimenti (6-50x), dotato di lente aggiuntiva perulteriori ingrandimenti (100 x);
– microscopio ottico a 100-1000ingrandimenti.
– pinzette da orologiaio;– aghi montati;– pipette in plastica graduate da 3
mL;– vetrini e coprioggetti per micro-
scopia;– alcool etilico a 80°;– glicerina;
Fig. 1. Attrezzature di campo per il campionamento di sorgenti: retino triangola-re da benthos per la raccolta diretta (a sinistra) e retino da drift, dotato di uncollettore terminale svitabile (a destra).

Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)34
Informazione&Documentazione
– provette e contenitori per la con-servazione in alcool degli orga-nismi.
Supporti strutturali:– banconi da lavoro predisposti
per il collocamento dei micro-scopi;
– lavandino idoneo alla prepara-zione del campione.
Modulistica:– modello SORG: per l’annotazio-
ne dei dati ispettivo-ambientali,mirato ad evidenziare le carat-teristiche specifiche del puntostazione oggetto d’indagine (All.2).
– modello BIOL: per l’annotazio-ne dei dati descrittivi del sito dicampionamento e per la compi-lazione dei risultati (All. 3).
REAGENTIL’unico prodotto normal-
mente utilizzato durante la fase dilaboratorio è l’alcool a 80°. La pre-parazione avviene utilizzando al-cool etilico a 94-96° (o alcool dena-turato), diluendo 80 mL di alcoolcon 14-16 mL di acqua. Occasio-nalmente, per i preparati, può es-sere impiegata anche qualche goc-cia di glicerina.
PROCEDURA
1 Fase di campo1.1 Rilevamento dati ambientali· Compilazione del modello
SORG (vedi istruzioni operati-ve, All. 2):- dati identificativi e di inqua-
dramento geografico;- valutazione ambientale del-
la stazione;- scelta dei siti per l’esame bio-
logico (Fig. 2): possono esse-re campionati più siti perciascuna sorgente; ad ognisito campionato deve corri-spondere un singolo model-
lo BIOL.– Compilazione del modello BIOL
(vedi istruzioni operative, All. 3):denominazione della sorgente,data e riferimento al sito cam-pionato, corrispondente a quan-to indicato nel modello SORG(“identificazione del punto d’ac-qua” e “siti dell’esame biologi-co”); compilazione dei campidella sezione “caratteristiche del
sito” e del primo campo dellasezione “caratteristiche del cam-pionamento”.
1.2Campionamento biologico– Raccolta del macrobenthos in
modo diretto: a vista si cercanogli organismi di maggiori dimen-sioni; il campionamento a vistaè integrativo ma non sostitutivodi quello tradizionale riportato
Fig. 2. Rappresentazione dei siti investigabili all’interno di una captazione (A, B)e all’esterno (C, D). A: ambiente interno di una captazione con scaturigine acces-sibile (SC), scorrimento interno (SI), scaturigine da tubo (ST), vasca di arrivo (VA),vasca successiva (VS); B: spaccato di una manufatto di captazione con scaturigineinglobata nella vasca di arrivo (SV) e vasca successiva (VS); C: ambiente esterno diuna captazione con scorrimento esterno (SE), permeazioni adiacenti (PA), rivoadiacente (RA) e vasca lungo la rete (VR); D: sorgente captata dopo un tratto discorrimento superficiale, scaturigine (SC), scorrimento esterno (SE) e captazione avalle della scaturigine (CV).
A B
C D

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 35
Informazione&Documentazione
di seguito;– raccolta di benthos e sedimen-
to mediante retino: raccolta nel-le diverse tipologie di substratopresenti nella sorgente, comemassi e ciottoli, ghiaia, sabbiao limo, muschi e fogliame mar-cescente, lavando direttamenteil materiale controcorrente da-vanti all’apertura del retino.
Anche per scorrimenti inter-ni posti in stretti cunicoli si puòapplicare la stessa metodologia dicampionamento. Eventualmente,se l’opera captativa lo permette,effettuare un ulteriore campiona-mento dalle vasche di raccolta (va-sca d’arrivo o vasche successive),disponendo il retino allo sboccodel tubo di scarico di fondo, racco-gliendo così il sedimento presentesul fondo dei manufatti. Se il depo-sito di sedimento accumulato è ri-levante, asportare solo lo strato su-perficiale.
La metodica può essere ap-plicata effettuando il campiona-mento di tutte le classi dimensio-nali di organismi (macro-, micro- emeiobenthos, con rete filtrante di100 μm), ma, ai fini dell’applica-zione degli indicatori su acque sor-give, può essere sufficiente consi-derare solo la componente macro-bentonica; in questo caso sarannoutilizzati i retini con maglie da 500μm.
1.3 CompletamentoCompilazione della tabella
relativa alla lista faunistica (mo-dello BIOL - sezione “Esame delbenthos” – “in situ”) solo per quan-to riguarda eventuali organismi ri-levati presso il sito di campiona-mento durante il sopralluogo. Se-gnalare l’esecuzione o il rilevamen-to sull’entità idrica in oggetto dialtre indagini non biologiche. Laregistrazione è prevista nella sche-da SORG (sezione “Altri rilievi ri-guardanti l’emergenza idrica” e
“campionamenti per esami di la-boratorio”).
Generalmente, unitamente alcampionamento, si rilevano i para-metri di campo fisico-chimici e idro-logici (temperatura aria e acqua,pH, ossigeno disciolto espresso inmg/L e come percentuale di satu-razione, conducibilità e portata) ele note meteo-climatiche (condizio-ni metereologiche) e idrologiche (re-gime idrico locale); vengono inoltreeffettuati i prelievi delle acque de-stinati alle analisi chimiche e bat-teriologiche di laboratorio. Posso-no essere condotte sul territorio leindagini ispettive sulle aree di sal-vaguardia dell’emergenza idrica ealtri rilevamenti (geologico-struttu-rali, geomorfologici, geomeccanici,etc.).
Il sedimento viene riposto inuno o più sacchetti che verrannomantenuti in borsa termica, unita-mente alle piastre frigorifere, du-rante il trasporto al laboratorio.
Giunti al laboratorio, i sac-chetti saranno conservati, aperti,in frigorifero a 3-5 °C. L’esame delbenthos e del sedimento dovrà av-venire entro le 24 ore successive almomento del campionamento.
2 Fase di laboratorio2.1 Preparazione del campione– Misurazione del volume: ripor-
re il sedimento in un contenito-re graduato e lasciarlo decanta-re per alcuni minuti prima diprocedere alla lettura del volu-me di sedimento.
– Lavaggio e separazione delle fra-zioni: per separare dal campio-ne la parte grossolana e ottene-re frazioni più omogenee, si pro-cede mediante l’utilizzo dei se-tacci. Con l’ausilio della retedel retino stesso (maglia 500 μmper macrobenthos, 100 μm permeio- e microbenthos), lavandoil campione in acqua, si elimi-na la parte più fine. Questa fase
può essere evitata quando ilmateriale raccolto è scarso e digranulometria intermedia; inquesto caso si elimina il mate-riale grossolano manualmentee si sciacqua velocemente il re-siduo dentro la rete. Le variefrazioni separate sono suddivi-se, in piccole quantità, in cap-sule Petri.
– Arricchimento: per campionivoluminosi e costituiti prevalen-temente da frazioni minerali siprocede alla concentrazione delmateriale organico prima del-l’esame. Il materiale organico,meno pesante del sedimento,può essere estratto mediante agi-tazione del campione, imprimen-do movimenti rotatori, rimesco-lando il substrato in un reci-piente pieno d’acqua e versan-do e filtrando nel retino rapida-mente l’acqua con le frazionipiù leggere in sospensione.
– Compilazione del modello BIOLnella sezione “Caratteristiche delcampione”.
2.2 Esame del benthos– Separazione degli organismi: le
frazioni vengono esaminate conl’ausilio di un microscopio ste-reoscopico, a bassi ingrandi-menti (6-10 x), per individuaregli organismi presenti che ven-gono prelevati con pinzette etrasferiti nel fissativo. Un fon-do nero facilita la visualizza-zione degli organismi depig-mentati e ialini.
– Fissazione: è preferibile utiliz-zare alcool a 80° in quanto nonindurisce i tessuti e il materialecosì trattato si presta meglio perlo studio anatomico (la formali-na neutralizzata al 4 % è spessousata, ma sconsigliabile, ancheperché cancerogena).
– Classificazione degli organismi.Per l’osservazione si utilizza unmicroscopio stereoscopico, con

Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)36
Informazione&Documentazione
luce diretta, a medi ingrandi-menti (25-50 x), dotato di lenteaggiuntiva quando sono neces-sari ulteriori ingrandimenti (100x). I caratteri minuti di impor-tanza diagnostica si osservanomediante preparati su vetrino,tramite un microscopio ottico aluce trasmessa. La classifica-zione va approfondita sino alivello sistematico in uso per ilmacrobenthos di acque super-ficiali (GHETTI, 1997), ad ecce-zione di alcuni gruppi aventitaxa con esigenze ecologichedifferenti, come Molluschi(Hydrobiidae) e Crostacei (An-fipodi e Isopodi), per i quali ènecessario identificare il gene-re o i gruppi di specie. Le chiavitassonomiche sono contenute inmolti articoli e manuali, comele “Guide per il riconoscimentodelle specie animali di acqueinterne italiane del CNR” (RUF-FO ed., 1977-1985), l’atlante diSANSONI (1992), i volumi di CAM-PAIOLI et al. (1994, 1999) e altrilavori specialistici su singoligruppi. Questa fase è molto cri-tica per la buona esecuzionedel metodo, in quanto l’utilizzodelle guide opportune, la corret-ta osservazione del materiale,ma soprattutto le approfonditeconoscenze tassonomiche daparte dell’operatore, sono di pri-maria importanza.
- Compilazione del modelloBIOL. La sezione “Esame delbenthos”, è predisposta apposi-tamente per annotare la listadei taxa rinvenuti, indicando ilnumero di esemplari riscontra-ti (approssimato se superiorealle dieci unità) e il giudizio diabbondanza, articolato in quat-tro classi di frequenza. Quest’ul-timo viene valutato in relazio-ne allo sforzo di campionamen-to ed al volume di sedimentoraccolto.
- Conservazione: si usa general-mente alcool a 80° (preferibil-mente con l’aggiunta di qual-che goccia di glicerina per gliartropodi). I campioni smistatia livello dei singoli gruppi van-no conservati in provette ade-guate, dotate di chiusura ido-nea (contenitori o provette invetro o materiale plastico) e ri-posti entro barattoli a chiusuraermetica anch’essi provvisti dialcool sul fondo.
- Etichettatura: il materiale smi-stato deve essere contrassegnatoda etichette indelebili collocateentro le singole provette; gene-ralmente si riportano le seguen-ti voci: gruppo tassonomico, ta-xon, corpo idrico, località e fra-zione, comune, provincia, rac-coglitori e data, coordinate UTMe quota.
2.3Esame del sedimento– Identificazione e valutazione
semi-quantitativa della parte or-ganica presente nel sedimento:le frazioni vengono esaminatecon l’ausilio di un microscopiostereoscopico, a bassi ingrandi-menti (6-10 x), per identificare,ed eventualmente separare, lecomponenti vegetali e animali.I resti di invertebrati acquatici,quando identificabili, devonoessere classificati allo stesso li-vello del benthos.
– Compilazione del modello BIOLnella sezione “Esame del sedi-mento”, predisposto apposita-mente per annotare la presenzadelle diverse componenti del se-dimento, suddivise in resti diorigine animale e resti di origi-ne vegetale. Sulla scheda sonoelencate le principali componen-ti che solitamente si riscontra-no; sono però previsti ulteriorispazi per approfondimenti, inparticolare per i resti di origineanimale. Anche in questo caso
il giudizio di abbondanza, arti-colato in quattro classi di fre-quenza, viene valutato in rela-zione al volume di substrato rac-colto.
3 Calcolo ed espressionedei risultati
3.1 Inquadramento biologico del sitoL’operatore, dalla scheda
SORG e dalla specifica schedaBIOL, procede alla valutazione deirisultati fornendo un primo qua-dro interpretativo. Questo si ottie-ne esaminando la lista faunistica equella relativa alla tanatocenosi,valutando le diverse voci con lafrequenza relativa delle diversecomponenti e assegnando a ciascu-na di esse la caratterizzazione eco-logica nel contesto ambientale diriferimento.
3.2 Applicazione degli indicatoriPer una valutazione più obiet-
tiva, l’applicazione della metodicaalle sorgenti captate prende in con-siderazione tre aspetti diversi a cuicorrispondono tre indicatori:– descrittore di inquinamento;– descrittore di vulnerabilità cap-
tativa;– descrittore idrologico.
Ciascun indicatore biologicoha come riferimento una schedaesplicativa specifica, riportata nel-l’All. 4, e strutturata come segue.La prima parte, riferita all’inqua-dramento generale, riporta gli at-tributi e le referenze dell’indicato-re (descrizione dell’indicatore, me-todi di misura, scopo, unità di mi-sura, copertura geografica dei dati,periodo di riferimento dei dati, do-cumenti di riferimento, osservazio-ni e commenti). La tabella prendein considerazione, nelle diverse si-tuazioni, la presenza e la frequen-za della componente faunistica(biocenosi) e di quella organicadel sedimento (tanatocenosi). L’in-terpretazione e la formulazione del

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 37
Informazione&Documentazione
giudizio si basa sul confronto tra irisultati della scheda BIOL con laserie di situazioni codificate (“ta-bella relativa alla biocenosi e ta-natocenosi di riferimento”).
Se le situazioni sono tipiche,l’ambiente e l’area geografica omo-genei, la biocenosi e la tanatoceno-si caratteristiche, il giudizio biolo-gico può dare un contributo valido
sugli aspetti presi in considerazio-ne o una conferma di quanto l’esa-me ispettivo o altre analisi forni-scono. Talvolta, tuttavia, specifichecondizioni locali, indipendenti dafattori antropici, come vicissitudi-ni paleogeografiche, substrato o chi-mismo particolare delle acque, pos-sono portare a difficoltà interpre-tative.
ALLEGATI– Allegato 1: tabella ambienti in-
vestigabili;– Allegato 2: modello SORG e
istruzione operativa;– Allegato 3: modello BIOL e istru-
zione operativa;– Allegato 4: tabelle relative ai tre
indicatori biologici.
ARPAL, 2000. Manuale AssicurazioneQualità. ARPAL, Laboratorio delDipartimento di Genova, ed. 00 del11/09/00, documento interno.
BODON M., GAITER S., 1989. Conside-razioni sul popolamento macro-bentonico delle acque sorgive. Bio-logia Ambientale, 3 (2): 5-12.
BODON M., GAITER S., 1995. Nuovi crite-ri di valutazione basati sulla com-ponente biologica per le captazionidi acque destinate al consumo uma-no. Biologia Ambientale, 9 (1): 5-17.
BODON M., RAFFETTO G., 1999. Appro-fondimento dello stato dell’arte degliindici biologici e trofici per la valu-tazione della qualità delle acque sot-terranee. ARPAL, Centro TematicoNazionale Acque Interne e Marinocostiere CTN_AIM, documento in-terno, 89 pp.
CAMPAIOLI S., GHETTI P.F., MINELLI A.,RUFFO S. 1994. Manuale per il ricono-scimento dei macroinvertebrati delleacque dolci italiane. Provincia Auto-noma di Trento, Vol. I, 357 pp.
CAMPAIOLI S., GHETTI P.F., MINELLI A. eRUFFO S. 1999. Manuale per il ricono-scimento dei macroinvertebrati delleacque dolci italiane. Agenzia Provin-ciale per la Protezione dell’Ambien-te, Trento, Vol. II: 358-484.
GAITER S., CHIAPPARA G.M., BALDINI I.,BERTOLOTTO R.M., TRAVERSO P., 1995.Metodi investigativi e analitici attia valutare lo stato di protezione e irischi igienici di piccoli acquedottiapprovvigionati da acquiferi mon-tani: applicazione alla situazioneconcreta di un piccolo comune(Maissana, La Spezia). In: Atti 2°Conv. Naz. “Protezione e gestione del-le acque sotterranee: metodologie, tec-
nologie e obiettivi”, Nonantola (MO),17-19 maggio 1995. Quaderni diGeologia Applicata, Pitagora Ed.,Bologna: 3.385-395.
GALASSI D., 2002. Il progetto PASCALIS.Biologia Ambientale, 16 (1): 60-61.
GHETTI P.F., 1997. Manuale di applica-zione. Indice Biotico Esteso (I.B.E.). Imacroinvertebrati nel controllo di qua-lità degli ambienti di acque correnti.Provincia Autonoma di Trento, 222pp.
GILBERT J., DOLE-OLIVIER M.J., MARMONIER
P., VERVIER P.,1990. Surface water-groundwater ecotones. In: R.J.Naiman et H. Decamps (eds.), Eco-logy and Management of acquatic-terrestrial ecotones. Partenon Publ.,London: 199-225.
MALARD. F., DOLE-OLIVIER M.-J., MATHIEU
J., STOCH F., 2003. Sampling ma-nual for the assessment of regionalgroundwater biodiversity. PASCA-LIS, European Project: Protocols forthe Assessment and Conservation ofAquatic Life in the Subsurface,http//www.pascalis-project.org.
PESCE G.L., 1996. Biocenosi a copepodidel bacino dell’alto corso del fiumeVomano e alcune sorgenti del GranSasso. In “Monitoraggio biologico delGran Sasso”, Consorzio di RicercaGran Sasso, Università degli Studidell’Aquila, Andromeda Editrice:292-296.
PEZZOLI E., 1984. Fenomeni geomor-fologici e faunistici di Valle Imagna(Carso, Acque, Fauna, Uomo). C.A.I.Sezione di Bovisio Masciago, 48 pp.
PEZZOLI E., 1985. I molluschi creno-bionti e stigobionti presenti in Val-le Imagna (Bergamo): carta genera-le delle stazioni ad oggi segnalate.
Bollettino Malacologico, 21: 1-11.PEZZOLI E., 1988. I molluschi creno-
bionti e stigobionti presenti in Ita-lia. Censimento delle stazioni, se-condo aggiornamento per il setten-trione e proseguimento per l’Italiaappenninica. Quad. Civ. Staz. Idro-biol., 15: 65-103.
PEZZOLI E., 1989. I molluschi creno-bionti e stigobionti presenti in Ita-lia. Censimento delle stazioni, ter-zo aggiornamento. Quad. Civ. Staz.Idrobiol., 16: 43-69.
RIPERT M., 1998. Contribution a l’ecolo-gie d’Hydrobiidae (Mollusca Gastro-poda Prosobranchia) souterrainsdans le Sud de la France. Univer-sité de Lyon I-Marseille II et Saint-Etienne: 28 pp.
ROCCA D., BODON M., GAITER S., CASARINO
E., 2003. Resoconto sul Corso diFormazione “Metazoi delle acquesorgive e sotterranee. Biologia,campionamento e riconoscimento,aspetti ambientali ed applicativi”.Genova, 18-22 settembre 2000. Bio-logia Ambientale, 17 (1): 77-84.
RUFFO S., 1977-1985 (Ed.). Guide peril riconoscimento delle specie ani-mali delle acque interne italiane.Collana del progetto finalizzato “Pro-mozione della qualità dell’ambiente”,CNR.
SANSONI G., 1992. Atlante per il riconosci-mento dei macroinvertebrati dei corsid’acqua italiani. Provincia Autonomadi Trento, Stazione SperimentaleAgraria Forestale, Servizio Protezio-ne Ambiente, 191 pp.
STOCH F., 2001. Primi risultati sull’usodegli organismi stigobi come indi-catori di qualità ambientale. Speleo-logia Veneta, 9: 157-163.
BIBLIOGRAFIA
Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)38
Informazione&Documentazione
Defin
izion
eCa
ratte
rizza
zione
Siti
Cara
tteriz
zazio
neCo
mun
itàsin
tetic
aam
bien
tale
oin
vest
igab
iliPe
rcor
so de
ll’acq
uife
roec
olog
icabi
olog
icade
ll’am
bien
teim
pian
tistic
aSo
rgen
te ca
rsica
Stato
natur
aleSC
Falda
in ro
cce c
arsic
he ⇒
Sco
rrime
nto su
perfic
iale
Ecoto
no: C
rena
lCr
enon
Oper
a di c
aptaz
ione
SC, S
T, VA
,Fa
lda in
rocc
e car
siche
⇒ O
pera
di pr
esa
(⇒ S
corri
mento
supe
rficial
e)Ec
otono
: Styg
al-Da
Styg
on, c
on el
emen
ti cre
nobi
di so
rgen
teSV
, VS
Cren
ala C
reno
n, co
n elem
enti s
tigob
iRi
sorg
ente
carsi
caSt
ato na
turale
SCSc
orrim
ento
supe
rficial
e ⇒ S
corri
mento
sotte
rrane
o in r
occe
carsi
che ⇒
Sco
rrime
ntoEc
otono
: Rhit
hral-
Rhith
ron,
con p
ossib
ili ele
menti
cren
obi
supe
rficial
eCr
enal
Oper
a di c
aptaz
ione
SC, S
T, VA
,Sc
orrim
ento
supe
rficial
e ⇒ S
corri
mento
sotte
rrane
o in r
occe
carsi
che ⇒
Ope
ra di
pres
aEc
otono
: Rhit
hral-
Rhith
ron,
con p
ossib
ili ele
menti
cren
obi
di so
rgen
teSV
, VS,
SI
(⇒ S
corri
mento
supe
rficial
e)Cr
enal
Sorg
ente
carsi
ca co
nSt
ato na
turale
SCSc
orrim
ento
supe
rficial
e ⇒ S
corri
mento
sotte
rrane
o + Fa
lda in
rocc
e car
siche
⇒ S
corri
mento
Ecoto
no: S
tygal-
Cren
on, c
on el
emen
ti rhit
hrali
alime
ntazio
ne m
ista
supe
rficial
eCr
enal-
Rhith
ral
Oper
a di c
aptaz
ione
SC, S
T, VA
,Sc
orrim
ento
supe
rficial
e ⇒ S
corri
mento
sotte
rrane
o + Fa
lda in
rocc
e car
siche
⇒ O
pera
diEc
otono
: Styg
al-Da
Styg
on, c
on el
emen
ti cre
nobi
e/o rh
ithra
lidi
sorg
ente
SV, V
S, S
Ipr
esa (
⇒ S
corri
mento
supe
rficial
e)Cr
enal-
Rhith
ral
a Cre
non,
con e
lemen
ti stig
obi e
/o rh
ithra
liSo
rgen
te in
rocc
e fes
sura
teSt
ato na
turale
SCFa
lda in
rocc
e fes
sura
te ⇒
Sco
rrime
nto su
perfic
iale
Ecoto
no: C
rena
lCr
enon
Oper
a di c
aptaz
ione
SC, S
T, VA
,Fa
lda in
rocc
e fes
sura
te ⇒
Ope
ra di
pres
a (⇒
Sco
rrime
nto su
perfic
iale)
Ecoto
no: S
tygal-
Da S
tygon
, con
elem
enti c
reno
bi a C
reno
n,di
sorg
ente
SV, V
S, S
ICr
enal
con e
lemen
ti stig
obi
Sorg
ente
di ve
rsante
Stato
natur
aleSC
Falda
di ve
rsante
in ro
cce p
erme
abili
per p
oros
ità ⇒
Sco
rrime
nto su
perfic
iale
Ecoto
no: C
rena
lCr
enon
(detr
ito di
falda
o mo
rena
)Op
era d
i cap
tazion
eSC
, ST,
VA,
Falda
di ve
rsante
in ro
cce p
erme
abili
per p
oros
ità ⇒
Ope
ra di
pres
aEc
otono
: Styg
al-Da
Styg
on, c
on el
emen
ti cre
nobi
a Cre
non,
di so
rgen
teSV
, VS,
SI
(⇒ S
corri
mento
supe
rficial
e)Cr
enal
con e
lemen
ti stig
obi e
freato
biRi
sorg
iva o
Fonta
nile
Stato
natur
ale o
con l
ievi
SCFa
lda fre
atica
in pi
ana a
lluvio
nale
⇒ S
corri
mento
supe
rficial
eEc
otono
: Cre
nal
Cren
onint
erve
nti pe
r aum
entar
eil d
eflus
soSc
orrim
enti i
n cav
ità ca
rsica
Stato
natur
aleSC
, SI
Falda
in ro
cce c
arsic
he: z
ona e
picar
sica o
zona
di pe
rcolaz
ione;
zona
di ci
rcolaz
ione,
Styg
alSt
ygon
anfib
ia o e
pifre
atica
; zon
a satu
ra, s
otto i
l live
llo di
base
Con o
pera
di ca
ptazio
neSC
, ST,
VA,
Falda
in ro
cce c
arsic
he ⇒
Ope
ra di
pres
aSt
ygal
Styg
onint
erna
SV, V
S, S
ISc
orrim
enti i
n cav
ità ar
tificia
leSe
nza o
pera
di ca
ptazio
neSC
, ST,
SIFa
lda in
rocc
e fes
sura
te o c
arsic
heSt
ygal
Styg
on(g
aller
ie, m
inier
e, ca
ve)
Con o
pera
di ca
ptazio
neSC
, ST,
VA,
Falda
in ro
cce f
essu
rate
o car
siche
⇒ O
pera
di pr
esa
Styg
alSt
ygon
inter
naSV
, VS,
SI
Perfo
razio
neOp
era d
i cap
tazion
eST
, VA,
VS
Falda
in ro
cce f
essu
rate
o car
siche
⇒ O
pera
di pr
esa i
ndott
aSt
ygal
Styg
onda
perfo
razio
niGa
lleria
dren
ante
Oper
a di c
aptaz
ione:
SC, S
T, VA
,Fa
lda di
versa
nte in
rocc
e per
meab
ili pe
r por
osità
⇒ O
pera
di pr
esa i
ndott
aSt
ygal
Styg
on, c
on el
emen
ti fre
atobi
galle
ria dr
enan
teSV
, VS,
SI
Pozz
o di v
ersa
nteOp
era d
i cap
tazion
e: po
zzo
FP, T
P, VA
, VS
Falda
di ve
rsante
in ro
cce p
erme
abili
per p
oros
ità ⇒
Ope
ra di
pres
a ind
otta
Styg
alSt
ygon
, con
elem
enti f
reato
biPo
zzo f
reati
coOp
era d
i cap
tazion
e: po
zzo
FP, T
P, VA
, VS
Falda
freati
ca in
pian
a allu
viona
le ⇒
Ope
ra di
pres
a ind
otta
Styg
alSt
ygon
, con
soli e
lemen
ti fre
atobi
Pozz
o fre
atico
alim
entat
oOp
era d
i cap
tazion
e: po
zzo
FP, T
P, VA
, VS
Scor
rimen
to su
perfic
iale ⇒
Falda
freati
ca in
pian
a allu
viona
le ⇒
Ope
ra di
pres
a ind
otta
Styg
alSt
ygon
, con
elem
enti i
porre
icida
un co
rso d’
acqu
aPo
zzo a
rtesia
noOp
era d
i cap
tazion
e: po
zzo
FP, T
P, VA
, VS
Falda
artes
iana i
n pian
a allu
viona
le ⇒
Ope
ra di
pres
a ind
otta
Styg
alSt
ygon
, con
soli e
lemen
ti fre
atobi
spec
ializz
ati
All
. 1 -
Tab
ella
am
bie
nti
in
vest
igab
ili
Nota.
Per
ogni
ambie
nte so
no rip
ortat
i i pos
sibili
“siti i
nves
tigab
ili” sp
ecific
i dell
’emer
genz
a idr
ica; p
ertan
to no
n son
o ripo
rtati q
uelli
adiac
enti o
situa
ti a va
lle de
lle op
ere d
i pre
sa. S
C: sc
aturig
ine ac
cess
ibile;
ST:
scatu
rigine
da tu
bo; V
A: va
sca d
i arri
vo; S
V:sc
aturig
ine +
vasc
a arri
vo; V
S: va
sche
succ
essiv
e; SI
: sco
rrime
nti in
terni;
VR:
vasc
a lun
go la
rete;
FP:
fond
o del
pozz
o, TP
: tubo
di po
mpag
gio (f
iltraz
ione d
ell’ac
qua a
ll’arri
vo);
una d
efiniz
ione d
ei div
ersi
siti è
ripor
tata s
otto l
a voc
e “sit
o cam
piona
to”ne
ll’istr
uzion
e ope
rativ
a al m
odell
o sor
g. (A
ll. 2)
. Nell
a des
crizio
ne de
l “per
corso
dell’a
cquif
ero”
, ven
gono
indic
ati tra
pare
ntesi
i per
corsi
che p
osso
no va
riare
a se
cond
a dell
a cap
tazion
e. Ne
lla “c
aratt
erizz
azion
e eco
logica
”, si in
tendo
no: e
coton
o: am
biente
di pa
ssag
gio tra
due s
istem
i idric
i dive
rsi; c
rena
l: amb
iente
di so
rgen
te (d
i con
tatto
tra ac
que s
otter
rane
e e su
perfic
iali);
styga
l: amb
iente
di ac
que s
otter
rane
e; rh
ithra
l: amb
iente
di ru
scell
o (ac
que s
uper
ficial
i). La
“com
unità
biolo
gica”
ripor
tata s
i rifer
isce
alla b
iocen
osi a
ttesa
.

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 39
Informazione&Documentazione
All. 2 - modello SORG e istruzione operativaCORPI IDRICI SOTTERRANEI (modello SORG) - parte generale
IDENTIFICAZIONE DEL PUNTO D'ACQUA
Codice: | | | | | - | | | Sopralluogo del | | Denominazione punto di rilevamento: __________________________________________________
INQUADRAMENTO GEOGRAFICO
Provincia di _____________ | | | Comune di ___________ | | | | | | | Fraz. __________________ Loc. __________________ Bacino _____________ | | | | | Sottobacino/i ___________________________ | | | | | Quota __________ m s.l.m. Coordinate UTM (IGM) | | | | | | | | Coordinate UTM (CTR) X | | | | | | | | Y | | | | | | | |
VALUTAZIONE AMBIENTALE DELLA STAZIONE
Ambiente: | | SO: sorgente; PV: pozzo di versante; PE: perforazione; EN: emergenza in cavità naturali; EA: emergenza in cavità artificiali; GD: galleria drenante. Tipologia della sorgente: | | RE: reocrena; LI: limnocrena; EL: eleocrena; AA: ambiente esterno assente. Posizione della sorgente rispetto a rivi attivi a monte: | | 1: nell'alveo; 2: in prossimità (2-5 m); 3: a 5-20 m; 4: a 20-100 m; 5: >100 m o rivi assenti. Posizione della sorgente rispetto a impluvi con scorrimenti a monte non perenni: | | 1: nell'impluvio; 2: in prossimità (2-5 m); 3: a 5-20 m; 4: a 20-100 m; 5: >100 m o impluvi assenti. Manufatto: | | TC: totalmente chiuso; CI: a chiusura imperfetta; PC: parzialmente chiuso o aperto; MR: rudimentale, aperto; MA: manufatto assente. Captazione: | | CT: captazione completa con derivazione totale; CD: captazione completa con dispersione intorno al manufatto; CZ: captazione completa con derivazione parziale; CP: captazione parziale; NC: captazione assente. Litologia: | | | | AC: arenarie calcaree; AG: argille; AS: argilloscisti; AN: andesiti; AR: arenarie; BA: basalti; CA: calcari; CD: calcari dolomitici; CL: calcescisti; CM: calcari metamorfici; CO: conglomerati; DS: diaspri; DR: dioriti; DO: dolomie; FL: flysh; GA: gabbri; GE: gessi; GN: gneiss; GR: graniti; LI: lipariti; MA: marne; ME: metagabbri; MI: micascisti; OC: ofiocalci; OL: ofioliti; PO: porfidi; PR: prasiniti; QU: quarziti; SC: scisti; SI: sieniti; TR: trachiti. Alterazioni del substrato: | | RC: roccia compatta; RA: roccia alterata; DF: detrito di falda; AL: alluvioni.
CORPI IDRICI SOTTERRANEI (modello SORG) - sintesi rilievi
Codice: | | | | | - | | | Campionamenti del | | Denominazione punto di rilevamento: _________________________________________________________
SITI DELL’ESAME BIOLOGICO
Siti campionati totali N. | | |A| Sito campionato: | | Seguono: - modello BIOL - Benthos | | Sedimento | | |B| Sito campionato: | | Seguono: - modello BIOL - Benthos | | Sedimento | | |C| Sito campionato: | | Seguono: - modello BIOL - Benthos | | Sedimento | | Siti della sorgente: SC: scaturigine accessibile; ST: scaturigine da tubo; VA: vasca di arrivo; SV: scaturigine + vasca arrivo; VS: vasche successive; CV: captazione a valle della scaturigine (impropria); SI: scorrimenti interni. Siti adiacenti o a valle della sorgente: SE: scorrimenti esterni; PA: permeazioni adiacenti; RA: rivi adiacenti; VR: vasca lungo la rete.
ALTRI RILIEVI RIGUARDANTI L'EMERGENZA IDRICA
Parametri fisico-chimici Si ; No modello __________ Sito | | Parametri idrologici Si ; No modello __________ Sito | | Infrastruttura Si ; No modello ____________________ Attività antropiche ed opere Si ; No modello ____________________ Geologici Si ; No modello ____________________ Dati anagrafici Si ; No modello ____________________
CAMPIONAMENTI PER ESAMI DI LABORATORIO
Analisi chimiche Si ; No Referto di prova N° | | Sito | | Profilo analitico | | Analisi batteriologiche Si ; No Referto di prova N° | | Sito | | Profilo analitico | | Note: ___________________________________________________________________________________________________________________
I Rilevatori ______________ ______________

Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)40
Informazione&Documentazione
Il modello “SORG” è composto di dueparti, la prima (parte generale) va com-pilata una sola volta per ogni emer-genza idrica (va rivista solo in caso dicambiamenti significativi), la seconda(sintesi rilievi), ogni volta che si effet-tua il sopralluogo o i rilevamenti. Per-tanto, per ogni successivo sopralluogoche comporta un campionamento bio-logico, è necessario compilare un altromodello SORG limitatamente alla par-te “sintesi rilievi”. Ciascuna parte èsuddivisa in sezioni, comprendenti di-versi campi. Ogni campo va compilatointegralmente in base alle spiegazionisotto indicate.
PARTE GENERALE
Prima sezioneIdentificazione del punto d’acqua
Codice: lo spazio riservato a questacampo rappresenta il numero identifi-cativo per ciascuna sorgente, indispen-sabile per l’archiviazione computeriz-zata dei dati. Il criterio di numerazio-ne può essere stabilito come sempliceprogressione cronologica o essere piùarticolato, con numeri e sigle in rela-zione al tipo di fonte o alla localizza-zione territoriale. Ad esempio, può es-sere formato da 4 cifre che identifica-no il numero progressivo che contrad-distingue il gruppo principale dellasorgente, seguito da altre due cifre chespecificano l’emergenza secondaria al-l’interno della scaturigine.
Denominazione punto di rilevamen-to: riportare il nome della sorgente.
Sopralluogo del: inserire la data incui è stata svolta la prima indaginepresso la sorgente ed è stato compilatoil modello SORG.
Seconda sezioneInquadramento geografico
Provincia: riportare la denominazio-ne della provincia e la sigla automobi-listica.
Comune: riportare la denominazionedel comune e il codice ISTAT (3 cifreper la provincia più tre per il comune).
Frazione: è riferita alla denominazio-ne della frazione più prossima allasorgente, dedotta dalle carte tecnicheregionali (o dalle tavolette dell’IGM), eriportata nell’Annuario del TCI deiComuni e Frazioni d’Italia o ricavatada documentazioni ufficiali degli entipubblici.
Località: è riferita alla denominazio-ne della località in cui ricade l’ambiente d’indagine; riportare la denomina-zione dalle carte tecniche regionali (odalle tavolette dell’IGM) e, solo se nonsegnalata, la toponomastica nota a li-vello locale.
Bacino: indicare la denominazione delcorpo idrico principale (con foce a mareo, nel caso di bacini interregionali,quelli principali che defluiscono fuorila regione d’indagine) sotto il qualericade il punto di rilevamento, prece-duto dall’indicazione di fiume (F.), tor-rente (T.) o rio (R.), in base alla carto-grafia regionale (o alle tavolette del-l’IGM).
Sottobacino/i: indicare la denomina-zione della serie di corpi idrici su cuiinsiste il punto di rilevamento, in basealla cartografia regionale (o alle tavo-lette dell’IGM), con le abbreviazioniriportate al campo precedente, ordina-ti da valle verso monte.
Quota: deve essere riportata la quotadella sorgente sul livello del mare,espressa in metri, ricavata dalle cartetecniche regionali e verificata sul po-sto sulla base dei punti quotati (i rile-vamenti della quota tramite GPS sonospesso imprecisi).
Coordinate UTM: a questo campo cor-rispondono due diverse espressioni inbase alla cartografia utilizzata. Nellecaselle relative alle coordinate UTM(IGM) indicare, nelle prime due casel-le le lettere che identificano il quadra-to di 100 X 100 km di lato, nelle casel-
le successive riportare i numeri cheidentificano il quadrato di 1 x 1 km dilato (le prime due cifre per la latitudi-ne, le altre due per la longitudine.Nelle caselle relative alle coordinateUTM (CTR) “X” ed “Y”, corrisponden-ti rispettivamente a latitudine e longi-tudine, riportare i valori delle coordi-nate UTM dedotti dalla cartografiaCTR in cifre (o trasformarle dalle coor-dinate Gauss-Boaga, se la cartografia èimpostata su questo reticolo), localiz-zando esattamente il punto con l’ap-prossimazione di almeno 10 m (in que-sto caso arrotondando a zero l’ultimacifra delle coordinate). La lettura del-la longitudine e della latitudine va ese-guita da sinistra verso destra (per lalongitudine) e dal basso verso l’alto(per la latitudine). Se le coordinate ven-gono registrate tramite un GPS impo-stato in UTM (sistema Europeo), ripor-tare il valore esatto rilevato dallo stru-mento.
Terza sezioneValutazione ambientale della stazione
Per poter effettuare una valutazioneambientale della sorgente da investi-gare sono stati opportunamente sceltialcuni campi, evidenziati in grassetto,ritenuti di fondamentale importanzaper una descrizione accurata dell’am-biente in esame. Ad ognuno di questisegue un apposito spazio che dovràessere compilato con le relative codifi-che (identificate da un numero o dauna sigla letterale, rispettivamente seriferite ad una stima quantitativa o adun’indicazione tipologica), elencatespecificatamente per ogni singolo cam-po.
Ambiente: indicare la tipologia am-bientale dell’emergenza: sorgente (SO),se, almeno in origine, sgorgava natu-ralmente all’esterno; perforazione (PE)se si tratta di una falda in acquiferopermeabile per fratturazione, intercet-tata da un’opera orizzontale; emergen-za in cavità naturale (EN) o emergenzain galleria artificiale (EA), se sgorgaall’interno rispettivamente di una ca-vità naturale o artificiale; pozzo di ver-
ISTRUZIONE OPERATIVA AL MODELLO SORG

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 41
Informazione&Documentazione
sante (PV) o galleria drenante (GD),rispettivamente se l’opera intercettauna falda in acquifero permeabile perporosità mediante un’opera verticaleo uno scavo orizzontale.
Tipologia della sorgente: indicare sela sorgente (o ambiente assimilabile)allo stato naturale, o comunque al difuori del manufatto captativo, originasubito un corso d’acqua, ed è quindidenominata reocrena (RE); se formauna polla sorgiva piuttosto ampia, lim-nocrena (LI); se la vena d’acqua chefuoriesce si espande in un’area palu-dosa, eleocrena (EL). Nel caso di sor-genti totalmente captate, le cui acquenon defluiscono mai all’esterno, indi-care ambiente esterno assente (AA).
Posizione della sorgente rispetto arivi attivi a monte: rilevare la presen-za di eventuali corsi d’acqua superfi-ciali presenti a monte, al fine di valu-tare la possibile connessione tra que-sti e la stessa sorgente. Indicare nel-l’apposito spazio il numero codificatocorrispondente all’intervallo di distan-za che separa la sorgente dallo scorri-mento superficiale, a partire da unasorgente localizzata all’interno dell’al-veo del corso d’acqua (1), fino ad unadistanza superiore ai 100 metri o as-senza di rivi (5). Con “rivi attivi” s’in-tendono i corsi d’acqua con scorrimen-to perenne, e quelli semiperenni chedurante l’arco dell’anno possono ave-re alcuni brevi periodi di secca, peresempio durante la stagione estiva.
Posizione della sorgente rispetto aimpluvi con scorrimenti a montenon perenni: rilevare la presenza dieventuali impluvi a monte in cui loscorrimento sia anche temporaneo, alfine di valutare il percorso e il deflus-so delle acque in caso di precipitazionimeteoriche o nell’eventualità più estre-ma di piogge torrenziali, con conse-guente dilavamento dei terreni circo-stanti e potenziale passaggio dellalama d’acqua entro il manufatto ol’emergenza idrica non captata. Indi-care nell’apposito spazio il numero co-dificato corrispondente alla distanzache separa la sorgente dall’impluvio, apartire da una sorgente localizzata al-l’interno dell’impluvio (1), fino ad una
distanza superiore ai 100 metri o as-senza di impluvi (5).
Manufatto: indicare quando l’opera ètotalmente chiusa e dotata di sistemidi comunicazione con l’ambiente ester-no a protezione efficace (presenza diguarnizioni, griglie a maglie fini, etc.)(TC); quando l’isolamento è imperfet-to, ovvero l’opera di comunicazione(porta, sportello, botola) è sprovvistadi guarnizioni, la chiusura è difettosao sono presenti modeste fessure (es.griglie a maglie superiori a 0,5 mm)(CI); quando il manufatto è aperto o ilsistema di chiusura non svolge alcunaopera protettiva e/o sono presenti am-pie aperture (PC); quando il manufattoè accessorio ed esterno, non ha funzio-ni protettive dell’emergenza idrica mainteressa la zona circostante alla scatu-rigine, come nel caso di trogoli, lava-toi, fontane, vasche aperte (MR); assen-za di manufatto, ovvero scaturigine incondizioni naturali o seminaturali(come modesti interventi in pietra)(MA).
Captazione: campo relativo all’effet-tivo prelievo idrico. Se è presente unmanufatto artificiale con derivazionedel flusso, indicare captazione totale(CT) nel caso in cui venga captata l’in-tera portata della sorgente e il flussosia completamente derivato; captazio-ne totale con dispersione (CD) quandol’acqua si disperde, in parte, intornoal manufatto; (CZ) quando la captazio-ne del flusso è totale ma l’acqua è deri-vata solo parzialmente e la parte re-stante è scaricata in prossimità del-l’emergenza; (CP) quando la captazio-ne è parziale, quindi solo una partedel flusso è captata. Infine, (NC) se ilflusso idrico non è captato, nel caso disorgenti naturali o provviste di operadi presa non attiva.
Litologia: indicare, in ordine di pre-valenza (separate dal simbolo “-”) o dirapporto stratigrafico (separate dalsimbolo “/”), le due principali tipolo-gie litologiche che interessano l’areadi alimentazione e di venuta a giornodella sorgente, secondo le voci codifi-cate.
Alterazioni del substrato: indicare se
lo sgorgo idrico nell’ultimo tratto discorrimento, prima della venuta a gior-no, attraversa: roccia compatta (RC);roccia alterata, ossia roccia soggetta afenomeni di sgretolamento/disgrega-zione ma di spessore contenuto e co-munque immediatamente a contattocon roccia compatta (RA); detrito di fal-da, spessore più rilevante di terrenodetritico derivante dall’accumulo diroccia che ha già compiuto un processodi alterazione, generalmente segnala-to nelle carte geologiche come morene,paleofrane, coltri eluvio-colluviali(DF); alluvioni, terreno alluvionalederivante dall’accumulo di sedimentiincoerenti (AL).
SINTESI RILIEVI
Riportare nell’intestazione il codicedella sorgente e la denominazione delpunto di rilevamento già indicato nelmodello SORG - parte generale -, e ladata in cui vengono effettuati i cam-pionamenti.
Prima sezioneSiti dell’esame biologico
Siti campionati totali N.: per sito cam-pionato s’intende il luogo, all’internoo all’esterno della sorgente, in cui siesegue il campionamento; nello spazioa disposizione riportare il numero to-tale di siti in cui è stato condotto ilcampionamento.
Sito campionato (Fig. 2): i possibilisiti che si possono incontrare sono elen-cati di seguito e ad ognuno corrispon-de una sigla che dovrà essere riportatanella casella relativa al “Sito campio-nato”: SC: scaturigine accessibile, iltratto più a monte accessibile sullo scor-rimento idrico (punto di emergenza na-turale, rinaturalizzato o anche artifi-cializzato dopo un intervento per in-tercettare la vena idrica, ma ancorafacilmente praticabile e, quindi, cam-pionabile); ST: scaturigine da tubo,quando la scaturigine viene intera-mente occlusa e l’acqua è convogliataal manufatto attraverso una tubazionedi piccolo diametro, dove il tubo d’ar-rivo rappresenta il punto più a montecampionabile con idonea strumenta-

Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)42
Informazione&Documentazione
zione; VA: vasca di arrivo, prima vascadi raccolta all’interno o all’esterno del-la captazione; SV: scaturigine con va-sca arrivo, quando la vena sorgiva af-fiora sul fondo o entro la vasca (scaturi-gine sommersa, non campionabile se-paratamente dalla vasca); VS: vaschesuccessive, vasche, nel manufatto cap-tativo, che seguono la prima vasca didecantazione; CV: captazione a valledella scaturigine, quando, prima dellacaptazione, si ha uno scorrimento su-perficiale che segue la scaturigine; SI:scorrimenti interni, scorrimenti suc-cessivi alla scaturigine, all’interno del-l’opera di presa o di una cavità artifi-ciale o naturale; SE: scorrimenti ester-ni, deflussi della sorgente all’esternodell’opera di presa (troppopieni o co-piose perdite); PA: permeazioni adia-centi, piccole perdite o infiltrazionidalla captazione o adiacenti alla sor-gente; RA: rivi adiacenti, acque super-ficiali prossime alla sorgente; VR: va-sca lungo la rete, vasca di raccolta inun manufatto a valle di quello captati-vo (lungo la rete dell’acquedotto).Nel tratto iniziale di una singola emer-genza idrica, i siti SC e ST sono alter-nativi, così come la presenza del sitoSV esclude entrambi i siti SC (o ST) eVA; si potranno quindi avere le se-guenti situazioni: SC + VA o ST + VA o
SV.Sono previsti fino a tre punti diversi,che vanno elencati da monte verso val-le; se necessario aggiungere altri siti.In corrispondenza di ciascun sito cam-pionato sono riportati i campi delledue analisi del modello BIOL, che de-vono essere contrassegnati nel momen-to in cui si effettua il campionamentodel benthos e/o del sedimento.
Seconda sezioneAltri rilievi riguardantil’emergenza idrica
In questa sezione i campi elencati siriferiscono a ulteriori rilevamenti sulterritorio, da riportare su modelli se-parati, che vengono svolti come corol-lario al campionamento o che possonoessere eseguiti durante il sopralluogo.A differenza delle altre sezioni, perciascun campo si deve barrare con unacrocetta la casella corrispondente allerisposte “Si” e “No”, in base all’avve-nuto rilevamento o meno. Inoltre, èriportato il riferimento al relativo mo-dello in cui sono registrati tali dati eche deve essere compilato; per i para-mentri fisico-chimici e idrologici è ne-cessario specificare anche il sito di in-dagine (vedi sezione “siti dell’esamebiologico”).
La titolarità della concessione, l’usodelle acque (potabile, industriale, agri-colo), etc., sono riportati unitamentead altri dati anagrafici su appositomodello.
Terza sezioneCampionamenti peresami di laboratorio
Anche in questa sezione in corrispon-denza dei campi riferiti ai prelievi perle analisi chimiche e batteriologichevi sono degli appositi spazi, da barrarecon una crocetta in base all’avvenutoprelievo del campione d’acqua (“Si”) omeno (“No”). Indicare, inoltre, il nu-mero del referto di prova del laborato-rio, il sito presso il quale il prelievoidrico è stato condotto e la sigla delprofilo analitico che il laboratorio se-gue per condurre le analisi.
Note: spazio relativo ad eventuali os-servazioni; segnalare ad esempio laparticolare tipologia della sorgente nelcaso di emergenze atipiche come lesorgenti artesiane o altre note specifi-che legate all’ambiente.
I Rilevatori: annotare i cognomi e nomidegli operatori che hanno effettuato ilsopralluogo e/o il campionamento.

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 43
Informazione&Documentazione
Allegato 3 - Modello BIOL e istruzione operativaCORPI IDRICI SOTTERRANEI (modello BIOL)
IDENTIFICAZIONE DEL SITO
Codice: |_|_|_|_|- |_|_| ______________________ Campionamento del | | |__| Sito campionato: |__ |
CARATTERISTICHE DEL SITO
Velocità della corrente: | | 1: impercettibile; 2: lenta; 3: media; 4: notevole. Luminosità: | | 1: assente; 2: molto debole; 3: sensibile; 4: normale. Substrati prevalenti: | | | | | | P: pareti artificiali; M: massi o roccia; C: ciottoli; G: ghiaia; S: sabbia; L: limo; D: detrito vegetale; A: detrito animale; R: radici; F: foglie; B: briofite; H: alghe.
CARATTERISTICHE DEL CAMPIONAMENTO
Modalità di raccolta: | | | | | | VI: vista; RA: retino da macrobenthos; RI: retino da microbenthos; DA: retino da drift per macrobenthos; DI: retino da drift per microbenthos. Significatività - BENTHOS: | | - SEDIMENTO: | | 1: insufficiente; 2: incerto; 3: sufficiente; 4: rilevante.
CARATTERISTICHE DEL CAMPIONE
Volume sedimento campionato: | | mL Granulometria prevalente (fraz. minerale): |__| G: ghiaia; S: sabbia; L: limo. Frazione organica: |__| 1: assente; 2: rara; 3: scarsa; 4: frequente; 5: abbondante; 6: dominante. Anaerobiosi: |__| 1: assente; 2: scarsa; 3: media; 4: notevole. Note: _______________________________________________________________________________________________________________
ESAME DEL BENTHOS
Taxa In situ Subcampione 100 μm
Subcampione 500 μm
Tot. N° Freq. (*)
ESAME DEL SEDIMENTO
Resti di origine vegetale Subcampione 100 μm
Subcampione 500 μm
Freq. (*)
Alghe Muschi Semi Detrito di foglie Detrito di radici Detrito legnoso Detrito decomposto Detrito carbonizzato Altro
Resti di origine animale Taxa Subcampione 100 μm
Subcampione 500 μm
Tot. N° Freq. (*)
Nicchi moll. acq. stigob. Nicchi moll. acq. crenob.
Nicchi moll. acq. superf. Nicchi moll. terr. endogei
Nicchi moll. terr. epigei
Gusci ostracodi
Foderi tricotteri
Foderi chironomidi Resti artropodi terrestri Resti artropodi acquatici Resti vertebrati
Altro
(*) R = raro ; S = scarso ; F = frequente ; A = abbondante. L'Analista ______________________

Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)44
Informazione&Documentazione
Il modello “BIOL” è composto di dueparti: la prima suddivisa in 3 sezioni,comprendenti diversi campi, che ca-ratterizzano il campionamento e il re-lativo campione raccolto, la seconda(da “esame del benthos” in poi) predi-sposta per i risultati dell’analisi delcampione. Ogni campo va compilatointegralmente in base alle spiegazionisotto indicate.N.B.: una sorgente in cui si effettua, adesempio, un campionamento in più sitidistinti, presenterà un solo modelloSORG - sintesi rilievi - (un punto d’ac-qua) e tanti modelli BIOL quanti sonoi siti campionati; quindi ad ogni mo-dello BIOL corrisponde un solo sito dicampionamento indicato nel modelloSORG.
PRIMA PARTE
Prima sezioneIdentificazione del sito
Le prime due righe della scheda iden-tificano i dati relativi alla sorgente(come dalla scheda SORG) ed al cam-pionamento (data e sito di campiona-mento).
Codice: riportare lo stesso codice indi-cato nel modello SORG corrisponden-te, seguito dalla denominazione dellasorgente.
Campionamento del: riportare la datain cui è stato effettuato il campiona-mento.
Sito campionato: questo campo è ri-preso dalla prima sezione del modelloSORG - sintesi rilievi, pertanto va com-pilato nello stesso modo: nella primacasella indicare la lettera prestampatacorrispondente al sito campionato ri-portato nel modello SORG; nella ca-sella seguente deve essere indicata lacodifica del sito, come riportato nellostesso modello.
Seconda sezioneCaratteristiche del sito
Per ognuno dei campi che seguono, in
questa e nella successiva sezione, lospazio previsto va compilato con la re-lativa codifica (numero o sigla lettera-le).
Velocità della corrente: valutare lavelocità della corrente nel sito del cam-pionamento, facendo riferimento alleseguenti codifiche: quasi assente, se siha scorrimento o rimescolamento qua-si impercettibile (1); debole, se il mo-vimento dell’acqua risulta essere len-to (2); media, quando la velocità è sen-sibile ma non elevata, con scorrimentolaminare o poco turbolento (3); notevo-le, se la corrente è elevata e il flussoturbolento (4).
Luminosità: valutare l’intensità inbase alle seguenti codifiche: assente,quando non è visibile alcuna presenzadi luce (1); molto debole, se è appenavisibile l’ambiente campionato (2); sen-sibile, quando l’intensità luminosa èridotta rispetto all’esterno (3); norma-le, se la differenza rispetto all’esternoè irrilevante (4). Nella valutazione diquesto parametro, se il sito investigatoè racchiuso da un manufatto o è unambiente ipogeo (grotta, miniera, etc.),occorre verificare la presenza di luceall’interno del sito in condizioni indi-sturbate (con porta o botola chiusa),osservando eventuali infiltrazioni deiraggi luminosi attraverso tutte le aper-ture. Nel caso in cui non sia presentealcuna opera di captazione la lumino-sità sarà sempre normale (4).
Substrati prevalenti: riportare le tretipologie prevalenti, in ordine di do-minanza. Fra le tipologie di substratosono elencate sia le pareti artificiali(P) se è presente un manufatto, sia letipologie granulometriche elementaridel substrato litoide, quali massi o roc-cia (oltre 265 mm, M); ciottoli (compre-si tra 64 e 265 mm, C); ghiaia (tra 2 e64 mm, G); sabbia (tra 0,06 e 2 mm etessitura granulosa, se sfregata tra ledita, S); limo (inferiore a 0,06 mm, L),sia la componente organica, detritodecomposto vegetale (D) o animale (A).Tra i substrati organici costituiti inprevalenza da vegetali, distinguere,
quando possibile: radici (R), foglie (F),briofite (B) e alghe (H).
Terza sezioneCaratteristiche del campionamento
Modalità di raccolta: indicare nell’ap-posito spazio la o le modalità con cui èstato eseguito il campionamento, se avista (VI), con retino da macrobenthos(RA) o da microbenthos (RI), con retinoda drift per macrobenthos (DA) o permicrobenthos (DI).
Significatività: per il benthos valuta-re, in base allo sforzo di campiona-mento (accessibilità e ampiezza delsito campionato, durata del campio-namento), alla quantità di sedimentoesaminato per l’estrazione degli orga-nismi, e ad eventuali altri fattori dialterazione naturali o antropici, la si-gnificatività del risultato, distinguen-do tra: insufficiente, quando le condi-zioni di campionamento e i risultatinon permettono di avere neppure ungiudizio indicativo sul popolamento(1); incerto, se i risultati appaiono in-completi nel valutare il popolamento(2); sufficiente, quando i risultati pos-sono essere considerati rappresenta-tivi della biocenosi (3); rilevante, selo sforzo di campionamento e l’analisifaunistica ottenuta appaiono moltocompleti (4). Per il sedimento valuta-re, con gli stessi criteri, la rappresen-tatività dei resti di origine animale evegetale, rapportando il risultato so-prattutto al volume di sedimento cam-pionato e alla rappresentatività diquest’ultimo (ovviamente è più ido-neo il substrato a granulometria finedepositatosi di recente, in zone a cor-rente debole).
Quarta sezioneCaratteristiche del campione
Volume sedimento campionato: in-dicare nell’apposito spazio la quanti-tà, espressa in mL, di sedimento rac-colto, non ancora filtrato, lasciandolosedimentare per alcuni minuti in uncontenitore graduato per allontanarela presenza dell’acqua in eccesso.
ISTRUZIONE OPERATIVA AL MODELLO BIOL

Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 45
Informazione&Documentazione
Granulometria prevalente (frazioneminerale): annotare nell’apposito spa-zio la codifica corrispondente alla ti-pologia granulometrica dominante sul-la frazione inorganica presente sul to-tale del campione, facendo riferimen-to a quelle di seguito elencate: ghiaia(G), sabbia (S), limo (L).
Frazione organica: valutare ed indi-care nell’apposito spazio la quantitàdi frazione organica (detrito vegetale eanimale) rispetto alla frazione minera-le, sul totale del campione, facendoriferimento alle seguenti codifichenumeriche: assente (1); rara, se infe-riore al 2 % (2); scarsa, se compresa trail 2 e 5 % (3); frequente, se compresatra 5 e 20 % (4); abbondante, se tra 20e 50 % (5); dominante, se maggiore del50 % (6).
Anaerobiosi: indicare la presenza disubstrato anaerobico (annerimentodella faccia inferiore dei massi e deiciottoli, nei punti a debole corrente opresenza di fango e limo nerastri emaleodoranti, con odore di uova mar-ce, dovuti all’azione di solfobatteri),con riferimento alla seguente scala:assente (1), scarsa (2), media (3), note-vole (4).
Note: indicare eventuali osservazionirelative al campionamento, come ladurata, l’area campionata, etc.
SECONDA PARTE
La seconda parte del modello BIOL èpredisposta per riportare i risultatidell’esame del benthos (prima sezio-ne) e del sedimento (seconda sezione).Si compone di tre tabelle: la prima
relativa agli organismi campionati, laseconda e la terza relative all’esamedel sedimento, separatamente per lacomponente organica di origine vege-tale e per quella di origine animale.
Prima sezioneEsame del benthos
La tabella è divisa in sei colonne. Nel-la prima colonna vanno elencati, unoper ogni riga, i taxa reperiti; nelle trecolonne successive va annotato il nu-mero di esemplari ritrovati (se nume-rosi, approssimare alla decina di uni-tà), separatamente per quelli indivi-duati in situ (seconda colonna), nel sub-campione da 100 μm (terza colonna),in quello da 500 μm (quarta colonna).Nella quinta colonna, indicare il nu-mero totale di individui ottenuto dallasomma di quelli riportati nelle colon-ne precedenti. L’ultima colonna si ri-ferisce alla frequenza del taxon in esa-me in relazione allo sforzo di prelievo.Ad esempio, per un campionamento dimedia significatività (sufficiente): raro(R), in genere 1 o 2 individui; scarso(S), mediamente da 3 fino a 10 esem-plari; frequente (F), da 10 a 25 indivi-dui; abbondante (A) se maggiore di 25.Per campionamenti meno significativio molto approfonditi ridurre o amplia-re la scala in proporzione allo sforzo diprelievo.
Seconda sezioneEsame del sedimento
La prima tabella, composta da quattrocolonne, riporta i resti di origine vege-tale presenti nel sedimento. Nella pri-ma colonna sono elencate le principalicomponenti che solitamente si riscon-trano. Per ogni voce riportare una sti-
ma quantitativa, articolata nelle quat-tro classi di frequenza in base alla per-centuale presente sul volume totaledel sedimento: raro, se inferiore al 2 %(1); scarso, se compreso tra 2 e 5 % (2);frequente, se tra 5 e 20 % (3); abbon-dante, se maggiore del 20 % (4). Il giu-dizio va indicato separatamente per idiversi subcampioni, mentre nell’ulti-ma colonna va riportata la stima riferi-ta al campione globale.
La seconda tabella, composta da seicolonne, è predisposta per valutare lacomponente di origine animale pre-sente nel sedimento, raggruppata nel-le categorie principali. Queste com-prendono organismi acquatici che almomento del campionamento nonsono più presenti ma lasciano traccedella loro colonizzazione (conchigliedi molluschi acquatici, gusci di ostra-codi, foderi di tricotteri o di chirono-midi, etc.) e altri non tipici dell’am-biente acquatico (molluschi terrestriendogei o epigei, artropodi terrestri,etc.). Nella seconda colonna specifica-re, quando identificato, il taxon ri-scontrato. Nelle colonne successive,come per la prima tabella sopra de-scritta, riportare a seconda del sub-campione esaminato, il numero di in-dividui raccolti, e successivamente(quinta colonna) il numero totale ditali individui. Nell’ultima colonna ri-portare la frequenza del taxon in esa-me, articolata in quattro classi (raro,scarso, frequente, abbondante), facen-do riferimento a quanto già espressoper la spiegazione alla tabella “esa-me del benthos”.
L’Analista: annotare il cognome enome dell’operatore che ha eseguito laprova analitica.
Metodica di indagini biologiche in acque sorgive Biol. Amb., 18 (n. 2, 2004)46
Informazione&Documentazione
Allegato 4 - Tabelle relative ai tre indicatori biologici
(continua)
STRUTTURA DELLA BIOCENOSI IN RELAZIONE ALL'INQUINAMENTO PER ACQUE SORGIVE DESCRIZIONE DELL’INDICATORE Analisi della struttura della comunità bentonica dell'ambiente di sorgente. METODI DI MISURA Riconoscimento a livello di taxa (specie o genere). Per il giudizio vedi tabella relativa alla
biocenosi di riferimento. SCOPO DELL’INDICATORE Valutazione della qualità dell'acqua di sorgente in relazione al livello di inquinamento,
sulla base dell'esame della comunità bentonica. UNITÀ DI MISURA Analisi qualitativa della composizione della biocenosi. COPERTURA GEOGRAFICA DEI DATI Nord Italia, Abruzzo (Gran Sasso e F. Vomano). PERIODO DI RIFERIMENTO DEI DATI 1970-2001. DOCUMENTI DI RIFERIMENTO PEZZOLI, 1984, 1985, 1988, 1989. PESCE, 1996. RIPERT, 1998.
BODON e Gaiter, 1989, 1995. STOCH, 2001. OSSERVAZIONI E COMMENTI I lavori sulla fauna di acque sorgive italiane sono piuttosto carenti riguardo gli aspetti
relativi ai rapporti tra gli organismi e l'inquinamento delle acque. Pertanto le informazioni citate, desunte dalla letteratura disponibile, sono solo generiche e indicative.
Tabella relativa alla biocenosi di riferimento
• Acque di buona qualità o poco inquinate
Presenza di specie stigobie e/o stigofile, Molluschi Hydrobiidae (Iglica, Graziana, Bythinella); elevato numero di taxa di Copepodi e presenza di Parastenocaris e Elaphoidella.
• Acque inquinate Rarefazione di Molluschi Hydrobiidae (Graziana); presenza di Molluschi Polmonati (Lymnaea peregra, L. truncatula, Ancylus); presenza di Anfipodi (Niphargus), rarefazione e scomparsa di specie esigenti, aumento delle specie tolleranti, comparsa di specie epigee di maggior livello saprobico.
• Acque fortemente inquinate Scomparsa della fauna creno- e stigobionte; scomparsa di Molluschi Hydrobiidae (Bythinella e Graziana); presenza di Oligocheti Enchytraeidae, Bivalvi Pisidium, Ditteri Ptychopteridae.
STRUTTURA DELLA BIOCENOSI E DELLA TANATOCENOSI COME DESCRITTORE IDROLOGICO PER ACQUE SORGIVE
DESCRIZIONE DELL’INDICATORE Analisi della struttura della comunità bentonica e della tanatocenosi dell'ambiente di sorgente.
METODI DI MISURA Riconoscimento a livello di taxa (specie o genere). Per il giudizio vedi tabella relativa alla biocenosi e alla tanatocenosi di riferimento.
SCOPO DELL’INDICATORE Valutazione della provenienza dell'acqua sorgiva in relazione agli apporti idrici (acque di falda o ipogee, acque di scorrimento esterno o superficiale, acque di origine mista).
UNITÀ DI MISURA Analisi qualitativa della composizione della biocenosi. COPERTURA GEOGRAFICA DEI DATI Liguria. PERIODO DI RIFERIMENTO DEI DATI 1988-2000. DOCUMENTI DI RIFERIMENTO BODON e GAITER, 1989, 1995. OSSERVAZIONI E COMMENTI I lavori sulla fauna di acque sorgive italiane sono piuttosto carenti riguardo gli aspetti
relativi ai rapporti tra gli organismi e l'origine delle acque. Pertanto le informazioni citate, desunte dalla letteratura disponibile, sono solo generiche e indicative. Bisogna tuttavia considerare che la comunità che colonizza l'ambiente della sorgente è strettamente dipendente dalle condizioni di illuminazione e dal grado di isolamento con l'ambiente esterno. Pertanto, nelle sorgenti captate, si realizzano condizioni ambientali tali da modificare profondamente la struttura della comunità e, in questi casi, il ritrovamento di componenti epigee assume un significato molto maggiore che permette di avere informazioni sull'origine delle acque.
Tabella relativa alla biocenosi e alla tanatocenosi di riferimento
• Sorgente alimentata solo da acque sotterranee
Biocenosi: presenza di organismi crenobionti o stigobionti (Bythinella, Hydrobiidae, Niphargus, Proasellus)
Tanatocenosi: presenza di nicchi di molluschi Hydrobiidae stigobionti. • Sorgente alimentata sia da
acque sotterranee che da acque superficiali
Biocenosi: presenza di organismi crenobionti o stigobionti (Hydrobiidae: Avenionia, Bythinella; Crostacei: Niphargus, Proasellus) e presenza di organismi di acque superficiali (Leuctra, Dytiscidae, Philopotamidae, Chironomidae, Ceratopogonidae).
Tanatocenosi: abbondante detrito vegetale, soprattutto di foglie. • Sorgente alimentata solo da
acque superficiali Biocenosi: assenza di organismi stigobionti o tipici crenobionti; presenza quasi
esclusiva di larve di insetti. Tanatocenosi: assenza di nicchi di molluschi Hydrobiidae crenobionti o stigobionti.
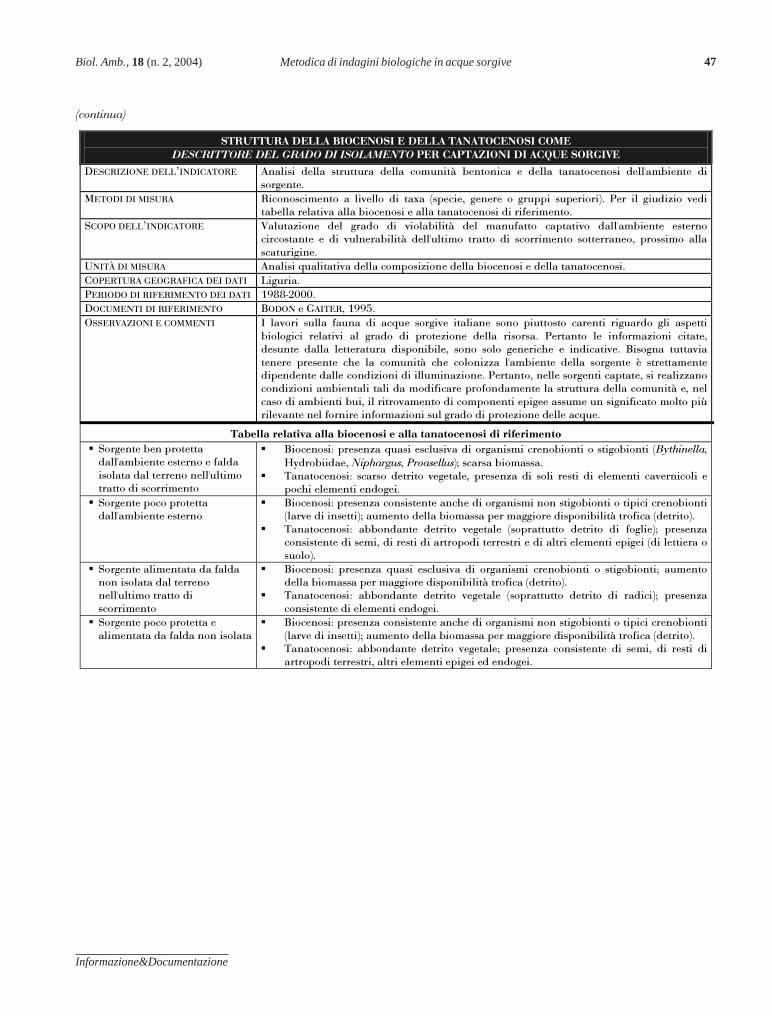
Biol. Amb., 18 (n. 2, 2004) Metodica di indagini biologiche in acque sorgive 47
Informazione&Documentazione
(continua)
STRUTTURA DELLA BIOCENOSI E DELLA TANATOCENOSI COME DESCRITTORE DEL GRADO DI ISOLAMENTO PER CAPTAZIONI DI ACQUE SORGIVE
DESCRIZIONE DELL’INDICATORE Analisi della struttura della comunità bentonica e della tanatocenosi dell'ambiente di sorgente.
METODI DI MISURA Riconoscimento a livello di taxa (specie, genere o gruppi superiori). Per il giudizio vedi tabella relativa alla biocenosi e alla tanatocenosi di riferimento.
SCOPO DELL’INDICATORE Valutazione del grado di violabilità del manufatto captativo dall'ambiente esterno circostante e di vulnerabilità dell'ultimo tratto di scorrimento sotterraneo, prossimo alla scaturigine.
UNITÀ DI MISURA Analisi qualitativa della composizione della biocenosi e della tanatocenosi. COPERTURA GEOGRAFICA DEI DATI Liguria. PERIODO DI RIFERIMENTO DEI DATI 1988-2000. DOCUMENTI DI RIFERIMENTO BODON e GAITER, 1995. OSSERVAZIONI E COMMENTI I lavori sulla fauna di acque sorgive italiane sono piuttosto carenti riguardo gli aspetti
biologici relativi al grado di protezione della risorsa. Pertanto le informazioni citate, desunte dalla letteratura disponibile, sono solo generiche e indicative. Bisogna tuttavia tenere presente che la comunità che colonizza l'ambiente della sorgente è strettamente dipendente dalle condizioni di illuminazione. Pertanto, nelle sorgenti captate, si realizzano condizioni ambientali tali da modificare profondamente la struttura della comunità e, nel caso di ambienti bui, il ritrovamento di componenti epigee assume un significato molto più rilevante nel fornire informazioni sul grado di protezione delle acque.
Tabella relativa alla biocenosi e alla tanatocenosi di riferimento Sorgente ben protetta dall'ambiente esterno e falda isolata dal terreno nell'ultimo tratto di scorrimento
Biocenosi: presenza quasi esclusiva di organismi crenobionti o stigobionti (Bythinella, Hydrobiidae, Niphargus, Proasellus); scarsa biomassa.
Tanatocenosi: scarso detrito vegetale, presenza di soli resti di elementi cavernicoli e pochi elementi endogei.
Sorgente poco protetta dall'ambiente esterno
Biocenosi: presenza consistente anche di organismi non stigobionti o tipici crenobionti (larve di insetti); aumento della biomassa per maggiore disponibilità trofica (detrito).
Tanatocenosi: abbondante detrito vegetale (soprattutto detrito di foglie); presenza consistente di semi, di resti di artropodi terrestri e di altri elementi epigei (di lettiera o suolo).
Sorgente alimentata da falda non isolata dal terreno nell'ultimo tratto di scorrimento
Biocenosi: presenza quasi esclusiva di organismi crenobionti o stigobionti; aumento della biomassa per maggiore disponibilità trofica (detrito).
Tanatocenosi: abbondante detrito vegetale (soprattutto detrito di radici); presenza consistente di elementi endogei.
Sorgente poco protetta e alimentata da falda non isolata
Biocenosi: presenza consistente anche di organismi non stigobionti o tipici crenobionti (larve di insetti); aumento della biomassa per maggiore disponibilità trofica (detrito).
Tanatocenosi: abbondante detrito vegetale; presenza consistente di semi, di resti di artropodi terrestri, altri elementi epigei ed endogei.

Biol. Amb., 18 (n. 2, 2004)48
Informazione&Documentazione
Il Siluro tra mito e realtà*
Da qualche tempo il viscido mo-stro attira l’interesse dei pescatori amosca in funzione della sua più appa-riscente ed importante caratteristica:la dimensione. Fino a qualche decen-nio fa l’Italia ne era esente, ma ora,grazie alla brillante lungimiranza deinostri amministratori e di chi si occu-pa, più o meno legalmente, di gestioneittica, questa specie deleteria conqui-sta sempre più areali contribuendo adegradare quel poco, di fauna più omeno autoctona, che potrebbe soprav-vivere. In ogni modo vediamo almenodi trasformare una brutta storia inuna piccola avventura culturale trascienza antica, mito popolare e, per-che no? Un pizzico di poesia.
Il siluro (Silurus glanis L.) è unpesce straordinario. Un grande pre-datore, molto efficace, in grado diraggiungere dimensioni inusualiper un pesce di acque dolci, a parteovviamente gli storioni. Tantostraordinario che ha convinto mol-tissimi pescatori a dedicarsi allasua pesca, compresi diversi PAM(pescatori a mosca) che hanno ab-bandonato le acque cristalline deitorrenti di montagna per quelle tor-bide e limacciose dei fiumi e deglistagni di pianura.
Armando Piccinini
e-mail: [email protected]
* Considerato l’interesse, si riproduce, con l’au-torizzazione dell’autore e della rivista, l’arti-colo sul siluro comparso su Fly line eco-sistemi fluviali, anno 19, n. 5, sett/ott 2004.
Un bel cambiamento insom-ma, ma che la dice lunga sul fasci-no esercitato da questo pesce.
Il siluro è comparso in Italiaalla fine degli anni ’60 con i primiesemplari rinvenuti nel fiume Ponel 1968. In realtà alcune catture,tra cui una del 1957, erano stategià state fatte nell’Adda ed in alcu-ni fiumi del Friuli-Venezia Giulia,ma erano state considerate come ilfrutto di immissioni accidentali. Il1968 può, in ogni caso, essere con-siderato come l’anno di comparsaufficiale di questa specie in Italia,vale a dire 36 anni fa. Quindi i piùvecchi esemplari di siluro non su-perano probabilmente i 40 annid.età, più o meno come quella ditanti pescatori a mosca.
Il siluro è originario dell’Eu-ropa centrale, dalla catena mon-tuosa dell’Ural fino al Reno; verso
Nord giunge fino in Svezia mentrea Sud il suo areale si estende finoall’Asia Centrale. In alcuni paesi èaddirittura protetto, come in Sve-zia, dove si trova solamente nelSud del paese, ed è considerato unrelitto glaciale. È stato poi introdot-to, per volontà o accidentalmente,in molti altri paesi europei, tra cuila Francia, l’Italia e la Spagna. Inquesti paesi la sua presenza ha datoorigine ad una fiorente attività de-dicata alla sua pesca, con barcheattrezzate, guide, itinerari di pesca,ecc. Insomma un vero e propriobusiness specializzato, a tal puntoche l’atteggiamento dei pescatorinei confronti di questa specie è pas-sato da un iniziale rifiuto e con-danna ad un più moderato assensoe, addirittura, richiesta di tutela neisuoi confronti.
Un’altra curiosità riguardo al
“Piscis pisci praeda, at Siluro omnes”(ogni pesce è preda di un altropesce, ma tutti lo sono del siluro).
Dall’opera di Gesnerus: particolare della stampa rappresentante il siluro.

Biol. Amb., 18 (n. 2, 2004) Il siluro 49
Informazione&Documentazione
siluro è l’esistenza in Europa di unsuo fratellino minore, chiamato Si-luro di Aristotile (Silurus aristotelisL.), presente solamente in alcunifiumi della Grecia. La differenzaprincipale dal siluro europeo è quel-la di possedere solamente 4 barbi-gli invece di 6 e di raggiungereminori dimensioni (alcuni chilo-grammi di peso). Tutti gli altri suoiparenti si trovano invece in Asia.
La presenza del siluro non èmai passata inosservata e già dal-l’antichità gli abitanti di fiumi elaghi conoscevano molto bene que-sta specie. Il primo che ha lasciatotestimonianze riguardanti il suocomportamento è stato il famosofilosofo greco Aristotele (Stagira nel384 A.C. - Calcide nel 322 A.C.).Egli nella sua opera “Historia Ani-malium”, che rimarrà un punto diriferimento per tutti gli studiosi finoal medioevo, descrive la biologiariproduttiva di questa specie, dallascelta del luogo di deposizione (unpunto riparato, ricco di vegetazio-ne), allo sviluppo delle uova, finoalla difesa del nido da parte delmaschio. Tratto da “Historia anima-lium, Ricerche sugli Animali, libro VI,568 a-b” “... I siluri di maggiori dimen-sioni depongono le uova in acque pro-fonde, in certi casi anche alla profon-dità di un braccio, mentre quelli piùpiccoli le depongono in acque basse,in particolare contro le radici di unsalice o di qualche altro albero, pres-so le canne o il muschio. Può capitareche si congiungano tra loro anche uno
molto grande con uno piccolo: avvici-nando insieme quei condotti, che al-cuni chiamano ombelichi, attraverso iquali esce la materia genitale, la fem-mina emette così l’uovo, il maschio illiquido seminale (...) Il maschio difen-de il punto dove le uova sono statedeposte in maggiore quantità, mentrela femmina se ne va dopo la deposizio-ne. Nei siluri, l’accrescimento a parti-re dall’uovo è molto lento, quindi ilmaschio sorveglia la prole anche perquaranta o cinquanta giorni affinchéessa non sia divorata dai pesci chepassano nei dintorni...”
E. probabile che il filosofo gre-co abbia, in realtà, descritto il com-portamento del Silurus aristotelis, cheda lui ha, infatti, preso il nome.
Dopo un greco, a scrivere delsiluro è invece un romano, GaioPlinio Secondo o Plinio il Vecchio(Como 24 D.C. – Pompei 79 D.C.),che morì durante l’eruzione delVesuvio che distrusse Ercolano ePompei, mentre era intento ad os-servare l’attività del vulcano.
Nella sua opera più impor-tante, la “Naturalis Historia”, chesi compone di ben di 32 libri, nellibro 9, che tratta degli animali
acquatici, Plinio paragona i siluriai tonni per le loro dimensioni esegnala questa specie nel Meno, af-fluente di destra del Reno, nel Da-nubio e nel Boristene, attuale Dne-pr che sfocia nel Mar Nero. Pliniolo indica presente anche nel Po,dove lo confonde probabilmentecon lo storione. Famosa è, in ognicaso, la descrizione della pesca alsiluro nel Danubio, con l’uso dicorde e ramponi.
Tratto da “Naturalis Historia,Gli animali acquatici, liber IX, vv. 44-46” “... Di particolare grandezza sonoi tonni (...) Anche in certi fiumi si trova-no dei pesci che non si sviluppanomeno, il siluro nel Nilo, l’isox nel Reno,l’attilo nel Po (...) Il siluro infierisce eattacca, dovunque si trovi, ogni ani-male, spesso affondando i cavalli chenuotano in superficie. Soprattutto nelMeno, fiume della Germania, viene trat-to fuori dall’acqua mediante file dibuoi, e nel Danubio con dei ramponi:è molto simile al porco di mare. Anchenel Boristene si dice che vi sia un pe-sce di particolare grandezza, senzaossa o spine interne, dalla carne mol-to dolce...”
Tuttavia Plinio non si ferma
Il frontespizio dell’opera originale di Gesnerus.
Nel libro III dell’opera di Konrad vonGesner (1550), il capitolo “De siluro”è arricchito da una bella stampa a colo-ri, a tutta pagina.

Il siluro Biol. Amb., 18 (n. 2, 2004)50
Informazione&Documentazione
a questo e nel libro 32, che trattadei rimedi in medicina che si pos-sono ottenere dagli animali acqua-tici scrive che: “... il grasso del siluroliquefatto al sole, misto a miele giovamolto alla vista se spalmato sugli oc-chi (...) i siluri salati o freschi corrobo-rano la voce (...) il siluro col suo brodoe la torpedine come cibo allentanol’intestino (...) il suo grasso cura i gelo-ni (...) il fegato del glanis se applicatosulle verruche le elimina (...)”.
Se qualcuno desidera prova-re, la materia prima non manca inItalia.
Dove ha appreso tutte questenotizie Plinio, riguardo ad un pe-sce che non era allora presente inItalia? Sicuramente durante la suapermanenza in Germania, in qua-lità d’importante funzionario im-periale. Nel momento storico in cuiè ambientato il noto film “Il Gla-diatore” interpretato da Russell eCrowe.
Ben diversa è invece la testi-monianza lasciata dal poeta e lette-rato romano Decimo Magno Auso-nio, nato a Burdigala (l’attuale Bor-deaux) nel 310 D.C. che rivestì l’in-carico di prefetto delle Gallie. Nelsuo poema più importante dal tito-lo “La Mosella” (Mosella, affluentedi sinistra del fiume Reno, nei pres-
si di Coblenza), descrive i pesci pre-senti e le abitudini delle genti chevivono lungo le sue rive. Scrivequindi Decimo Magno Ausonio: trat-to da “La Mosella” “Ora sarai cele-brato tu, potente siluro (Magne silu-ro), animale delle acque, come se l’oliodell’Attica avesse unto il tuo dorso, ioti considero un delfino del fiume, taleè il tuo incedere maestoso, col qualescivoli per le onde e tale è lo sforzo colquale tu spieghi, avanzando, il tuo lun-go corpo, quando piccoli bassifondi oerbe fluviali ti proteggono. Ma quan-do avanzi tra le onde calme, alloradesti l’ammirazione delle rive verdeg-gianti, dell’azzurra moltitudine deipesci, delle acque limpide: i flutti, ri-bollendo, si spandono per il letto delfiume e le ultime onde corrono versole rive. In questo sei simile ad unabalena che nel profondo Atlantico,spinta dal vento o dal suo proprio slan-cio verso le coste della terraferma, fastraripare il mare, solleva grosse on-date e i monti vicini temono di esseremeno elevati. Invece la mite Balenadella Mosella, ben lungi dall’esserecausa di danno, è un grande onoreche si aggiunge al fiume...”
Data la conoscenza che i ro-mani avevano del siluro, forse, pos-siamo dirci fortunati che non ab-biano deciso di introdurlo in Italiafin da epoche così antiche. Infatti,se così fosse stato, adesso potrebbeessere considerato come un’intro-duzione storica (come per la carpa)ed essere considerato come facenteparte dell’ittiofauna italiana, conopportune limitazioni per tutelar-lo. Forse i romani, che non eranocertamente ignoranti, avevano so-spettato che, sotto sotto, era megliolasciarlo in Germania, anche per-ché abituati a mangiare orate espigole nelle loro stupende ville inriva al mare, non avrebbero certofatto cambio con il siluro.
Decimo Magno Ausonio nelsuo poema si sofferma anche, contono rattristato, sui pesci che, cat-
turati dalle reti, cercano di ritorna-re in acqua. Scrive quindi: “L’umi-do bottino si dibatte sull’arida rocciae si spaventa nel sentire i dardi mortalidel giorno che reca la luce. E il pesce,che nel suo fiume mantiene il natiovigore, lentamente consuma la sua vitanel respirare l’aria. Orma, dal corpoindebolito, sono vibrati dei palpiti pi-gri e la coda intorpidita sopporta itremiti estremi. La bocca non si ri-chiude più e le branchie, espirando isoffi letali, cacciano fuori l’aria assor-bita (...) Io ho visto coi miei occhi alcu-ni pesci palpitanti e sul punto di mori-re raccogliere le loro ultime forze ed’un balzo precipitarsi giù nel fiumesottostante, di nuovo impadronendo-si dell’acqua in cui disperavano di nuo-tare...”.
Chissà se Decimo Magno Au-sonio può essere considerato comeun sostenitore dei No-Kill ante lit-teram. La sua è di certo la primatestimonianza, di circa 1700 annifa, a sostegno del Catch and Relea-se.
Non dimentichiamo però chenel 300 d.C. la fame era veramentefame, i pescatori pochi e senza ter-minali al fluorocarbon, i fiumi in-contaminati ed i pesci tanti. Se inostri fiumi fossero meno antropiz-zati (anche solamente un po’ meno)sarebbe possibile un moderato pre-lievo anche da parte dei pescatori
Dall’opera di Konrad von Gesner(1550): lo scazzone (Cottus gobio), aman-te di acque fresche e ben ossigenate econosciuto alla maggior parte dei pe-scatori a mosca.
Un’altra rappresentazione del siluro,questa volta a carattere anatomico. Sipossono distinguere il grande stomacosacciforme, il fegato trilobato e la ve-scica natatoria a forma di cuore, carat-teristica di questa specie.

Biol. Amb., 18 (n. 2, 2004) Il siluro 51
Informazione&Documentazione
senza incidere sulle comunità itti-che.
Per trovare nuove notizie con-cernenti il siluro è ora necessariofare un lungo balzo in avanti neltempo, superare il Medioevo e giun-gere fino al XV e XVI secolo quan-do, grazie all’Umanesimo ed al Ri-nascimento, si assiste ad un rinno-vato interesse per lo studio dellanatura.
È il periodo di Niccolò Coper-nico e Galileo Galilei e, in questianni, accade un fatto importantis-simo che cambierà completamenteil modo di pensare: la nascita dellastampa, grazie a Gutenberg.
La possibilità di stampare ilibri ne permette la loro rapida dif-fusione e così nuove idee e scopertepossono viaggiare rapidamente at-traverso l’intera Europa. È quindinel XVI secolo che nasce la scienzachiamata “Ittiologia” grazie all’ita-liano Ippolito Salviani (nato a Cit-tà di Castello nel 1514 e morto aRoma nel 1572) ed ai francesi Pier-re Belon e Guillaume Rondelet. Edè in questo periodo, grazie alla stam-pa, che ci si può imbattere nell’ope-
ra di Konrad von Gesner o Conra-dus Gesnerus, nato a Zurigo nel1516.
Conradus Gesnerus è oggi, aposteriori, considerato una menteenciclopedica, versato in tutti i cam-pi del sapere: legge quasi tutto, viag-gia, apre un museo, organizza con-vegni medici e pubblica circa 400libri. Alla fine muore nel 1565 perun’epidemia di peste a Zurigo.
La sua opera più importante,dove possiamo scoprire molte inte-ressanti notizie relative al siluro èla “Historiae Animalium”, in 4 vo-lumi, pubblicata tra il 1551 e il1587 (le fotografie allegate risalgo-no ad un’edizione del 1556). Nellibro terzo, dedicato agli animaliacquatici, si incontra quindi unintero capitolo dedicato al siluro:“De Siluro”.
La sua descrizione è dettaglia-ta e ricca di notizie. Dopo averdimostrato che il siluro non deveessere confuso con lo storione (si-mile per dimensioni) ed il luccio(simile per voracità), cita un elencodi località in cui questa specie èpresente, che sono ancora oggi al-l’interno del suo areale di distribu-zione. Gesner continua poi con “Co-rollarium I. De Siluro ex veteri-bus”, dove riporta le testimonianzerelative al siluro, già scritte dagliantichi filosofi e naturalisti. Inte-ressante è la descrizione, tratta daEliano, della pesca ai siluri fattadagli abitanti della città di Tomis(o Tomos, forse l’attuale Costanzavicino alla foce del Danubio) nel-l’Ister, attuale Danubio.
Gesnerus descrive così che ilpescatore dell’Ister conduce pressola riva del fiume una coppia dibuoi, o di cavalli, portando eglistesso un giogo per gli animali. Scel-to così un luogo adatto per la pescae comodo per sè, mette del cibo pergli animali presso la riva e legauno dei due capi di una fune moltorobusta al giogo che ha messo agli
animali e all’altro mette un grossoamo, che innesca con del polmonesecco di bue. Poi dopo aver messoun piombo sulla lenza, la getta inacqua. Quando il siluro mangia ilboccone ed è trafitto dall’amo, ten-ta di liberarsi e con tutte le sueforze tira la fune. Il pescatore allo-ra con uno scudiscio incita i suoianimali e comincia così una lottatra questi ed il grosso pesce e, allafine, questo figlio dell’Ister è vintoed è trascinato a riva.
Sempre in queste località, scri-ve Gesnerus, il fumo del siluro, bru-ciato a fuoco lento, è utilizzato peruccidere le formiche (e forse anchegli umani, possiamo aggiungere, seerano nei paraggi).
Al pari di Plinio, riporta gliutilizzi del siluro nella pratica me-dica, rifacendosi anche al famosomedico dell’antichità Galeno. Il si-luro salato ha così il potere di puli-re le arterie e rendere chiara lavoce, mentre la sua salamoia puòcurare i dolori del ventre e i doloriischiatici. Per quanto riguarda lacapacità di “pulire le arterie” anco-ra oggi, a titolo di prevenzione, sonosomministrati estratti d’olio di pe-sce per il loro alto contenuto diacidi omega-3.
La sua descrizione continuapoi con “Corollarium II De Siluroex nostris observationibus”, vale adire le osservazioni che lo stessoautore ha potuto fare di persona.Interessante è la spiegazione del-l’origine dei nomi del siluro neidifferenti paesi europei.
In Ungheria il siluro è dettoHarcha per la larghezza del dorsoo forse della bocca. Gesnerus sostie-ne con certezza che il pesce, che èchiamato Harcha, è il siluro e unaconferma gli viene da diversi stu-diosi dell’Ungheria. Riporta così ilracconto di uno di questi studiosiche vide su un carro un pesce “bar-buto” lungo 7 o 8 cubiti pescatonel Tibisco (affluente di sinistra del
Non poteva infine mancare la trotafario, chiamata Trutta Fluviatili. Qui èriportata anche la classificazione dellediverse specie che era fatta all’epoca.

Il siluro Biol. Amb., 18 (n. 2, 2004)52
Informazione&Documentazione
Danubio sopra Belgrado). Questopesce sarebbe rimasto nascosto per16 anni sotto la cucina di un nobi-le e alla fine era stato preso con unamo, mentre curava i suoi piccoli,e aveva trascinato i pescatori perdue miglia prima di arrendersi. Ilpesce fu portato nella città di Nad-lac e nel suo ventre fu trovata latesta di un uomo e una mano de-stra con 3 anelli d’oro. Questo fattocolpì tanto gli abitanti di quellacittà che per molti anni non man-giarono più questo genere di pesce,come avevano sempre fatto.
Il pesce del racconto avevabaffi sia sopra che sotto la bocca ela sua apertura era molto grande,la coda si assottigliava verso la fineed i denti erano simili a quei petti-ni, detti cardi, utilizzati per lavora-re la lana ed era privo di squame.
In Dalmazia il siluro è chia-mato invece Sum, nome usato an-che dai Boemi e dai Polacchi, che èlo stesso nome con cui sono chia-mate le balene. Da qui il possibileparagone tra il siluro e la balena.
Tra gli abitanti delle rive del-l’Elba è chiamato Sum: cetaceonemico dei pesci. Il siluro era an-che allora già ben noto per la suagrande voracità, a tal punto chenell’impero Romano Germanicoesisteva un proverbio che diceva:“Piscis pisci praeda, at Siluro om-nes” (ogni pesce è preda di un altropesce, ma tutti lo sono del siluro).Lungo le rive del Danubio, scriveancora Gesnerus, il siluro è chia-mato Schaib o Schaiben, o per ildanno che arreca per la sua voraci-tà (da schab che significa danno) oper la forma simile al fodero di unaspada (che è chiamato schaib oscheib). In altre regioni della Ger-mania è chiamato invece Waller,Weller ,Wale, Walle, Wallern, Wel-line, Walarin, che derivano da wal
o wallfish, nomi con cui i Germanichiamano la balena.
Nel lago Acronio, attuale lagodi Costanza, i pesci più grossi chevi si trovano sono i Walinin con lacarne simile a quella della bottatri-ce. Tuttavia sono catturati di rado,poiché abitano gli abissi più pro-fondi. Talvolta emergono in super-ficie, mai da soli, e ciò sarebbeforiero di grandi mutamenti.
Scrive Gesnerus che nel 1498nel luogo chiamato l’Angolo delReno, dove il Reno esce dal lago diCostanza, furono presi 3 siluri ed ilpiù piccolo, che comunque era piùlungo di un uomo, fu portato aCostanza ed ancora lo si potevavedere in un dipinto. Verso Nord,in Sassonia, è detto Wal o Weller,nome derivato da quello della bale-na, che si chiama wels, e da ciò inlatino è detto Velsum. Infine, scriveGesnerus, seguendo le indicazionidi un altro studioso, che è moltopericoloso immetterlo incautamen-te nelle peschiere e nei vivai per lasua voracità.
Dopo il Gesner, è giusto citareinfine l’opera di Ippolito Salviani,considerato tra i padri dell’ittiolo-gia, che nella sua opera “Aquati-lium animalium historia”, scrittatra il 1554 e il 1557, descrive ilsiluro, riprendendo gran parte del-le notizie del Gesner. Salviani scri-ve, infatti, che “In nostri vero Itali-ae fluviis nullibi (quod sciam) ha-betur” vale a dire “per quanto iosappia, il siluro non è presente neifiumi italiani”. Tra le notizie ripor-tate, vale la pena di menzionare lenotizie che Salviani riceve dal con-te Hieronymus Martinengus, nun-zio pontificio a Vienna, il quale gliriferisce che un fanciullo che nuo-tava nel Danubio presso Possoniamera stato divorato da un grosso silu-ro. Una volta catturato ed esamina-
Nell’opera di Gesnerus sono descrittimolti altri pesci, oltre al siluro, tra cuiil temolo, amato dalla maggior partedei PAM. Qui è rappresentato un indi-viduo adulto ed uno giovane.
to con cura dagli incaricati del ve-scovo, nei suoi intestini furono tro-vati i resti del povero fanciullo. Tut-to quello che segue è storia deinostri giorni. Il siluro è ormai arri-vato in Italia, e per le sue abitudinie dimensioni, continua ancora asuscitare meraviglia e stimolare lanascita di storie e leggende. Chinon ha mai sentito raccontare constupore di un germano o di unanutria che mentre nuotavano sonostate aggrediti da un grande siluro?La storia si ripete dunque. Oggi leconoscenze riguardo a questa spe-cie sono molto maggiori (ad esem-pio si conosce che possiede la capa-cità di individuare le prede anchecon un sistema di elettrolocalizza-zione), eppure continua sempre astimolare il nostro immaginario,compreso quello dei PAM. Infineun’ultima notizia, per chi si reche-rà a pescare i siluri nel fiume Po. Iprimi siluri sono comparsi circa 40anni fa e se alcuni dei primi esem-plari sono ancora vivi (fatto nonimprobabile) dovrebbero ormai averraggiunto il peso di almeno 150-180 kg! Comunque sempre lontanodal più grande mai catturato, chepesava ben 330 kg. Basterà unacanna da salmone?

Biol. Amb., 18 (n. 2, 2004) 53
RASSEGNA SCIENTIFICA
A cura diPietro Genoni
CollaboratoriMichele Arcadipane, Rossana Caroni, Stefano Fenoglio, Alessandra R. Magarelli
Questa rubrica presenta una selezione di articoli di recente pubblicazione su alcune tra le principali riviste scientificheinternazionali. Il suo scopo è quello di offrire ai lettori una panoramica il più possibile aggiornata sullo stato dellaricerca, di base ed applicata, nell’ambito degli argomenti trattati da Biologia Ambientale. I lavori sono raggruppati inbase alla tematica affrontata, indicata mediante parole chiave all’inizio di ciascun gruppo di articoli.Le versioni integrali degli articoli presentati possono essere richieste direttamente all’autore referente indicato inciascuna recensione, oppure alla redazione di Biologia Ambientale ([email protected]).La rubrica è aperta al contributo di tutti i soci, i quali, oltre a segnalare alla redazione articoli ritenuti di interesse, sonoinvitati a collaborare in prima persona alla loro recensione, prendendo preventivi accordi con il curatore([email protected]).
ECOLOGIA DELLE ACQUE INTERNE > MONITORAGGIO BIOLOGICO > MACROINVERTEBRATI BENTONICI
Measuring and controlling data quality in biological assemblage surveys with specialreference to stream benthic macroinvertebratesY. Cao, C.P. Hawkins and M.R. VinsonFreshwater Biology, 48: 1898 (2003)Y. Cao, Department of Aquatic, Watershed and Earth Resources, Utah State University, Logan, UT 84322-5210, USA. E-mail: [email protected]
Le indagini sulle comunitàbiologiche sono finalizzate in pri-mo luogo a definire la composizio-ne in specie e le relative abbondan-ze ad una o più scale spaziali otemporali. L’interpretazione dei datie le conclusioni dipendono da quan-to i campioni sono ben rappresen-tativi della comunità indagata.
Le misure convenzionali diqualità dei dati, come la deviazio-ne standard o il coefficiente di va-riazione, sono stati impiegati per lastima di singole variabili, ma sonoinsufficienti o inidonee per stimarela qualità dei dati che descrivonointere comunità biologiche. Gli in-dici di similarità, invece, tenendoconto della composizione in specie
e della loro abbondanza relativa,possono essere usati per misurare econtrollare efficacemente la quali-tà di questi dati.
Il coefficiente medio di Jac-card (JC) calcolato tra più coppiedi campioni replicati, ossia il coef-ficiente di autosimilarità (AJC), èconcettualmente e numericamentecorrelato al coefficiente medio divariazione della densità di tutte lespecie osservate, una misura di pre-cisione del campionamento, ed allafrazione di ricchezza totale in spe-cie, una misura di accuratezza delcampionamento.
Gli autori esaminano come ilcoefficiente AJC possa essere utiliz-zato per valutare l’effetto di diffe-
renti potenziali cause di errore sul-la qualità dei dati provenienti daindagini sulle comunità biologiche,inclusi lo sforzo di campionamen-to applicato sia all’interno diun’area sia in singoli punti, le pro-cedure di sub-campionamento, laconcordanza nel riconoscimentotassonomico.
Gli autori concludono chel’approccio basato sull’autosimila-rità permette di superare la mag-gior parte dei punti deboli legatialle misure convenzionali di quali-tà dei dati e può essere impiegatoper misurare e controllare efficace-mente i dati delle indagini biologi-che.
[PG]

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)54
Rapid assessment of rivers using macroinvertebrates: the role of experience, and compa-risons with quantitative methodsL. Metzeling, B. Chessman, R. Hardwick, V. WongHydrobiologia, 510: 39-52 (2003)L. Metzeling, Cooperative Research Centre for Freshwater Ecology, EPA Victoria, 40 City Road, Southbank. E-mail:[email protected]
Gli autori hanno confrontatoi risultati ottenuti mediante la va-lutazione biologica rapida (RBA)dei macroinvertebrati rispetto a cam-pioni quantitativi raccolti in 18 cop-pie di siti fluviali dell’Australia su-dorientale. In ciascuna coppia unodei punti di campionamento rap-presentava un sito di riferimento,mentre l’altro presentava diversigradi di disturbo, da lieve a mode-rato, dovuto ad inquinamento pun-tiforme o diffuso (effluenti di alle-vamenti ittici, scarichi di piccolidepuratori urbani, dighe, agricoltu-ra e pascolo).
I campioni sono stati raccoltinelle zone di raschio (riffles), lungole rive e sulle rocce nelle pozze(pools), utilizzando principalmenteretini manuali (RBA) e campiona-tori di Hess (quantitativi).
Dai campioni di macroinver-tebrati raccolti mediante RBA sonosempre stati ottenuti sub-campionimediante lo smistamento degli or-ganismi vivi sul campo per 30 mi-nuti. Sono stati effettuati confrontitra operatori non esperti (studentiuniversitari) e biologi esperti nel-l’applicazione dei protocolli RBA,inclusa l’identificazione degli orga-nismi. I campioni quantitativi era-no raccolti solo da personale esper-to, conservati e smistati al micro-scopio in laboratorio, osservandotutto il materiale od un suo sotto-campione. Gli individui sono staticlassificati a livello di famiglia inentrambi i metodi.
Gli operatori non esperti han-no osservato un numero di fami-glie leggermente inferiore rispettoagli esperti ed hanno commesso al-
cuni errori di identificazione, men-tre i dati degli operatori esperti de-gli stessi siti e degli stessi ambientierano mediamente più consistenti.Tuttavia, un metodo di ordinamen-to dei dati (Procrustes analysis) hapermesso di mostrare che le diffe-renza tra i dati RBA e quelli quan-titativi non dipendevano dall’espe-rienza dell’operatore RBA.
Inoltre, indipendentementedall’operatore, i dati RBA sono ri-sultati migliori rispetto a quelliquantitativi nel discriminare tra isiti di riferimento ed i siti disturbatiper tutti i tre tipi di habitat indaga-ti. Gli autori concludono che ilmonitoraggio RBA è sensibile ed abasso costo e raccomandano di mi-gliorare la consistenza dei dati rac-colti da operatori diversi.
[PG]
Invertebrate traits for the biomonitoring of large European rivers: an initial assessmentof alternative metricsS. Gayraud, B. Statzner, P. Bady, A. Haybachp, F. Schöll, P. Usseglio-Polatera, and Michel BacchiFreshwater Biology, 48: 2045 (2003)B. Statzner, Université de Lyon1, UMR 5023 CNRS, Ecologie des Hydrosystèmes Fluviaux, Boulevard du 11 novembre 1918, 69622Villeurbanne Cedex, France. E-mail: [email protected]
L’applicazione dei regolamen-ti e delle normative che riguardanodiversi ambiti amministrativi su lar-ga scala crea un crescente bisognodi strumenti standardizzati per va-lutare e monitorare lo stato di “sa-lute ecologica” dei fiumi, una ri-chiesta che può essere soddisfattatramite la descrizione di funzioniecologiche delle specie di inverte-brati che costituiscono le comunitàdei fiumi.
Per valutare metriche alter-native, gli autori hanno sperimen-tato in quale misura la strutturafunzionale (descritta da 14 caratte-ristiche biologiche) delle comunitàdi invertebrati, in 190 tratti di gran-di fiumi, differiva rispetto a diversilivelli di identificazione tassonomi-ca (specie, genere, famiglia), di mi-sure di abbondanza (dati grezzi,trasformazione log-normale delleabbondanze, presenza-assenza) e di
considerazione delle specie aliene(incluse od escluse), ed in qualemisura queste differenze influenza-vano la potenzialità dei descrittorifunzionali nel discriminare i trattidi fiume lungo un gradiente di mol-teplici impatti antropici.
I descrittori funzionali rica-vati a livello di specie, genere efamiglia sono risultati molto simili,mentre quelli derivati dai dati grez-zi differivano significativamente da

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 55
quelli derivati dai valori log-nor-male delle abbondanze e dai datidi presenza-assenza. I descrittorifunzionali con l’esclusione dellespecie aliene differivano considere-volmente da quelli con tali specieincluse.
In generale, i descrittori fun-zionali sono riusciti a discrimina-re i tratti di fiume in funzione dellivello di impatto antropico. La ri-soluzione tassonomica ha scarsa-mente influenzato tale discrimina-zione, mentre l’uso dei dati grezziha diminuito la capacità discrimi-natoria rispetto all’uso dei valorilog-normali delle abbondanze e dei
dati di presenza-assenza. Ugual-mente, l’esclusione di specie alie-ne ha diminuito la capacità didistinguere i differenti livelli diimpatto.
Se considerate separatamen-te, le singole caratteristiche biologi-che che descrivono la taglia massi-ma degli individui, il numero dineonati per ciclo riproduttivo, ilnumero di cicli riproduttivi per in-dividuo, la durata della vita degliadulti, il metodo riproduttivo, lecure parentali, la forma del corpo ele abitudini alimentari hanno mo-strato la capacità più elevata didiscriminazione dei livelli di im-
patto antropico.I risultati indicano che l’iden-
tificazione a livello di genere o for-se di famiglia può essere sufficienteper il biomonitoraggio dei grandifiumi quando si usano le caratteri-stiche degli invertebrati. Sebbene idati grezzi di abbondanza possanofornire una migliore discriminazio-ne in presenza di bassi livelli diimpatto, i dati di presenza-assenzadovrebbero essere sufficienti per in-dividuare le modificazioni funzio-nali delle comunità biologiche cau-sate da elevati livelli di disturboantropico nei fiumi europei.
[PG]
Stream macroinvertebrate response to catchment urbanisation (Georgia, U.S.A.)A.H. Roy, A.D. Rosemond, M.J. Paul, D.S. Leigh, J.B. WallaceFreshwater Biology, 48: 329 (2003)A.H. Roy, Institute of Ecology, University of Georgia, Athens, GA 30602, U.S.A. E-mail: [email protected]
Gli effetti dell’urbanizzazionedei bacini idrografici sulla qualitàdelle acque superficiali sono statiesaminati considerando 30 fiumidella Georgia, U.S.A., suddivisi intre classi dimensionali: con bacinidi 15, 50 e 100 km2 (± 25%). Sonostate esaminate le relazioni esisten-ti tra copertura ed uso del suolo(aree urbane, agricole e forestali) ele comunità macrobentoniche. Sonoinoltre state considerate le caratte-ristiche geomorfologiche, idrologi-che e chimiche dei corpi idrici.
Gli autori hanno osservatoevidenti correlazioni tra le caratte-ristiche antropiche del bacino e le
cenosi fluviali. La ricchezza tasso-nomica ed altri indici biotici risul-tano negativamente correlati conl’estensione delle aree urbane e po-sitivamente con quella delle areeforestali. La qualità biologica delleacque peggiora sensibilmente quan-do la presenza urbana è maggioredel 15% sul totale dell’area drena-ta. Una maggior presenza di areeurbane determina inoltre una di-minuzione della granulometriamedia del fondo, un aumento deisolidi sospesi ed una variazione del-la qualità chimica, con un aumen-to delle concentrazioni di azoto efosforo, della conducibilità e della
torbidità. I valori degli indici bioti-ci possono essere meglio previstiquando si utilizzino queste variabi-li a livello di tratto fluviale, piutto-sto che variabili di uso del suolo alivello di bacino.
L’urbanizzazione comportauna riduzione della diversità nellecomunità macrobentoniche, conl’aumento di taxa estremamentetolleranti. Gli indici biotici mag-giormente sensibili sono risultati laricchezza tassonomica, la ricchez-za EPT (Ephemeroptera, Plecopte-ra, Tricoptera) e l’indice ICI (Inver-tebrate Community Index).
[SF]
The effect of an industrial effluent on an urban stream benthic community: water qualityvs. habitat qualityE.J. Nedeau, R.W. Merritt, and M.G. KaufmanEnvironmental Pollution, 123: 1-13 (2003)E.J. Nedeau, Departments of Entomology and Fisheries and Wildlife, Michigan State University, East Lansing, MI 48824, USA
Gli autori hanno studiatol’effetto di un effluente di origineindustriale sulla qualità dell’acqua,
sulla qualità dell’habitat e sui ma-croinvertebrati bentonici di un cor-so d’acqua urbano nel Michigan
sudoccidentale (USA). Lo scaricodeterminava un incremento delletemperature del recettore di 13-18°C

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)56
metodi diversi per raccogliere ma-croinvertebrati bentonici in habi-tat di deposizione e di erosione alfine di comprendere l’importanzarelativa della qualità dell’habitatrispetto alle alterazioni della quali-tà dell’acqua. Le variabili stimatein base ai macroinvertebrati han-no incluso la ricchezza tassonomi-ca, l’abbondanza e l’abbondanzarelativa di gruppi tassonomici sen-sibili.
I risultati indicano che l’ef-fluente produce un effetto positivosulle comunità di macroinvertebra-ti, incrementando la quantità deglihabitat di riffle, ma anche un effet-to negativo a causa del peggiora-mento della qualità dell’acqua. Irisultati hanno dimostrato la ne-cessità di effettuare accurate consi-derazioni sulla qualità degli habi-tat e dell’acqua nei programmi diripristino o recupero fluviale.
[ARM]
durante i mesi più freddi e traspor-tava grandi quantità di ferro (con-centrazioni oltre 20 volte più altedel fondo ambientale) che ricopri-vano il letto fluviale. L’effluentedeterminava un impatto anche sul-le condizioni dell’habitat, incremen-tando la portata del fiume del 50-150% e causando un cambiamentosignificativo del substrato e dellecondizioni di flusso.
Gli autori hanno usato tre
Spatiotemporal variations in species diversity to assess environmental impacts on aqua-tic communities: the use of biomass data versus density dataJ.A. CamargoEnvironmental Monitoring and Assessment, 40: 125-136 (1996)
L’autore esamina l’utilizzo deidati di biomassa rispetto ai dati didensità al fine di stimare le varia-zioni spazio-temporali della diver-sità specifica nelle indagini ecolo-giche volte a valutare gli impattiambientali sulle comunità acquati-che. A tale scopo è stata utilizzatala comunità dei macroinvertebratibentonici. Gli studi in campo sonostati condotti lungo il Rio Duraton(Spagna settentrionale, bacino delDurero), il quale, oltre ad essereregolato da una diga, riceve gli sca-richi di un insediamento industria-le. I valori di diversità sono staticalcolati utilizzando la funzione diShannon-Wiener (H’). Sono statestimate due differenti misure di di-versità, la diversità di biomassa (H’b) e la diversità di densità (H’ d).
I rilasci di acque ipolimnichedal bacino della centrale idroelet-trica determinavano fluttuazioni abreve termine della portata e basseconcentrazioni di ossigeno disciol-to nelle quattro stazioni poste avalle della diga –S2, S3, S4 e S5 (a0,2, 0,4, 2,5 e 7,6 km dalla diga,rispettivamente)– rispetto alla sta-zione di riferimento S1, posta amonte. Lo scarico industriale cau-sava un incremento della concen-trazione di fluoro nelle stazioni S3,S4 e S5. La stazione S1 ha mostratoi valori più elevati di H’ d, S (ric-chezza in specie), B (biomassa tota-le) e N (densità totale), seguita, inordine decrescente, dalle stazioniS5, S4, S2 e S3. Al contrario, i valo-ri di H’ b non hanno mostratoquesta distribuzione spaziale. Di
conseguenza, i valori di S, B e Nsono risultati correlati in misuraminore con H’ b rispetto a H’ d.Inoltre, i coefficienti di variazioneentro e tra i campionamenti ma-crobentonici sono risultati più ele-vati per i valori di H’ b piuttostoche per quelli di H’ d.
In base ai risultati ottenuti, sipuò concludere che l’uso dei datidi densità sembra essere più appro-priato rispetto all’utilizzo dei datidi biomassa per valutare gli impattiambientali sulle comunità acquati-che. Tuttavia queste conclusioni,sebbene significative, non possonoservire come base per delle genera-lizzazioni, dal momento che essesono state ottenute da un unicogruppo di dati raccolti su un brevetratto di un unico fiume. [PG]
Efficiency of a standardised artificial substrate for biological monitoring of river waterqualityA. Pashkevich, T. Pavluk, A. Bij De VaateEnvironmental Monitoring and Assessment, 40: 143-156 (1996)
Prima di utilizzare i macroin-vertebrati nella valutazione dellaqualità del fiume Chusovaya (Rus-
sia), sono stati confrontati i risulta-ti di tre metodi di campionamento:retino manuale, pala circolare e
substrati artificiali standardizzati.Il substrato artificiale era costituitoda un cestello di acciaio riempito

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 57
con biglie di vetro (diametro 20mm). Al fine di confrontare l’effi-cienza di questi metodi di campio-namento il numero totale dei taxatrovati in ciascun sito, con i trecampionatori, è stato consideratopari al 100%.
L’efficienza più elevata è sta-ta ottenuta con il substrato artifi-
ciale, con il quale è stato catturatotra il 75 ed il 100% dei taxa nellediverse stazioni. La pala circolareha permesso di catturare solo il 5-19% ed il 10-20% dei taxa (rispetti-vamente su substrati sabbiosi e ghia-iosi), mostrandosi così il metodomeno efficiente. Risultati interme-di sono stati ottenuti con il retino
manuale, che ha permesso di cattu-rare il 23-38% dei taxa.
In base a questi risultati ed airequisiti generali dei metodi di cam-pionamento, il substrato artificialestandardizzato può essere conside-rato un ottimo metodo di campio-namento dei macroinvertebrati nelmonitoraggio biologico. [PG]
ECOLOGIA DELLE ACQUE INTERNE > MONITORAGGIO BIOLOGICO > AMBIENTE RIPARIO
A simple field method for assessing the ecological quality of riparian habitat in riversand streams: QBR indexA. Munné, N. Prat, C. Solà, N. Bonada, M. RieradevallAquatic Conservation: Marine and Freshwater Ecosystems, 13: 147-163 (2003)N. Prat, Dept. Ecologia, Facultat de Biologia, Universitat de Barcelona, Diagonal, 645, Barcelona E-08028, Spain. E-mail: [email protected]
Lo scopo dell’indice di quali-tà riparia presentato nell’articolo èdi fornire agli enti deputati allagestione di fiumi e torrenti un me-todo semplice per valutare la quali-tà dell’habitat ripario. L’indice èfacile da calcolare e può essere usa-to insieme ad altri indici di qualitàdell’acqua per valutare lo stato eco-logico dei corsi d’acqua. Inoltre,può rappresentare anche un utilestrumento per definire un elevatostato ecologico secondo la Direttivaquadro comunitaria sulle acque.
L’indice, chiamato QBR, èbasato su quattro componenti del-l’habitat ripario: copertura totaledella vegetazione riparia, strutturadella copertura, qualità della co-pertura e alterazioni dell’alveo. Essoprende in considerazione anche le
differenze nella geomorfologia delfiume dalla sorgente ai tratti infe-riori; tali caratteristiche sono valu-tate attraverso semplici misurequantitative.
Il punteggio dell’indice variatra 0 e 100. L’indice QBR vienecalcolato in campo attraverso unascheda (due pagine formato A4) chepuò essere completata in 10 minu-ti.
Lo sviluppo dell’indice QBRha previsto prove in quattro bacinifluviali mediterranei della Catalo-gna (Spagna nordorientale). Sonostati valutati settantadue siti cam-pione ed i risultati sono stati usatiper saggiare l’indice.
Non sono necessari esperti intassonomia per applicare l’indice,sebbene si richiedano alcune cono-
scenze sulla flora locale per distin-guere le specie arboree native daquelle non-native.
I risultati mostrano che l’in-dice QBR può essere usato a pre-scindere dalle differenze regionalinelle comunità di piante. Gli inter-valli di qualità ottenuti attraversol’applicazione dell’indice nello stes-so sito non sono risultati moltoinfluenzati dall’osservatore.
Attualmente l’indice, utilizza-to da diversi gruppi di ricerca, èsaggiato in uno studio comparativoin 12 bacini lungo le coste mediter-ranee della Spagna. È verosimileche l’indice QBR possa essere adat-tato senza significativi cambiamen-ti per l’uso in altre aree geografichein zone temperate e semi-aride.
[ARM]
ECOLOGIA DELLE ACQUE INTERNE > MONITORAGGIO BIOLOGICO > DEMOLIZIONE FOGLIARE
Assessing structural and functional ecosystem condition using leaf breakdown: studieson a polluted riverC. Pascoal, M. Pinho, F. Cássio and P. GomesFreshwater Biology, 48: 2033 (2003)C. Pascoal, Departamento de Biologia, Universidade do Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected]
Gli autori hanno determinatoi tassi di demolizione delle foglie di
Alnus glutinosa ed hanno analizza-to la struttura degli organismi de-
compositori associati alle foglie, alfine di valutare l’effetto dell’inqui-

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)58
acquatici, ma non a significativicambiamenti nella ricchezza tasso-nomica di questi ultimi. L’arricchi-mento in nutrienti nei tratti a valleè stato messo in relazione con unmaggiore tasso di demolizione fo-gliare. Il grado di alterazione fun-zionale –valutato attraverso il rap-porto dei tassi di demolizione fo-gliare in borse a maglia larga e a
maglia stretta– è risultato in accor-do con il gradiente di inquinamen-to definito attraverso due indici bio-tici.
Lo studio supporta l’opinioneche gli esperimenti sulla demolizio-ne fogliare siano uno strumentovalido per valutare l’effetto dell’in-quinamento sulle condizioni eco-logiche dei fiumi.
[ARM]
namento sulle condizioni ecologi-che del fiume Ave (Portogallo nor-doccidentale).
Un incremento in nutrientiorganici ed inorganici è apparsocorrelato ad un aumento della den-sità ed ad una riduzione della ric-chezza di macroinvertebrati, ad unamarcata riduzione della produzio-ne di conidi da parte degli ifomiceti
ECOLOGIA DELLE ACQUE INTERNE > MACROINVERTEBRATI BENTONICI
Vertical distribution of drift in the Erro river (NW Italy)S. Fenoglio, T. Bo, G. Gallina, M. CuccoJournal of Freshwater Ecology 19: 485-492 (2004)S. Fenoglio, Università del Piemonte Orientale, Dipartimento di Scienze e Tecnologie Avanzate., via Cavour 84, 15100 Alessandria. E-mail:[email protected].
In questo studio è stata esa-minata la composizione tassono-mica e quantitativa del drift nelfiume Erro (Provincia di Alessan-dria). Sono stati realizzati 51 cam-pionamenti nel periodo marzo 2002-marzo 2003, utilizzando un retinoda drift in grado di intercettare gliorganismi nella colonna d’acquaseparandoli a tre livelli di profondi-tà: organismi trasportati sul fondo,a metà ed in prossimità della super-ficie. Durante il periodo di studiosono stati realizzati campionamen-
ti quantitativi della fauna macro-bentonica presente nel tratto flu-viale (retino Surber) e sono stateregistrate temperatura dell’acqua evelocità della corrente.
Complessivamente sono statiraccolti e classificati 3.394 organi-smi appartenenti a 28 taxa nel ma-teriale trasportato e 15.351 organi-smi appartenenti a 45 taxa nel sub-strato. Il drift è risultato di maggio-re entità in prossimità del fondo(dal punto di vista sia della ricchez-za tassonomica, sia del numero di
individui/volume d’acqua), anchese è stata rilevata una tendenza didiversi taxa ad essere presenti prefe-renzialmente sempre alla stessa al-tezza nella colonna d’acqua; sonoipotizzate alcune spiegazioni lega-te al comportamento e al ciclo bio-logico dei diversi taxa. È stato inol-tre rilevato come il drift sia inversa-mente legato all’andamento delletemperature e positivamente corre-lato con la densità del popolamen-to naturale presente in quel mo-mento nel tratto fluviale. [SF]
A comparative study of the dispersal of 10 species of stream invertebratesElliott J.M.Freshwater Biology, 48: 1652 (2003)J.M. Elliott, Freshwater Biological Association, Far Sawrey, Ambleside, Cumbria LA22 0LP, U.K. E-mail: [email protected]
Se si eccettuano gli studi ine-renti il drift, pochi dati quantitativisono disponibili per caratterizzarela capacità di dispersione dei ma-croinvertebrati bentonici. Questostudio presenta dati comparativirelativi a dieci specie, utilizzandodue differenti metodi: l’analisi deimovimenti di animali non marcati
(n=20-80) rilasciati in alcuni fiumiartificiali e quelli di animali mar-cati (n=20-60) rilasciati in ambien-te naturale.
I due metodi di indagine han-no fornito risultati simili: la capa-cità dispersiva è risultata indipen-dente dalla densità relativa, mastrettamente proporzionale al nu-
mero di animali rilasciati. I risulta-ti mostrano una certa variabilitàtra le diverse specie e non risultanoinfluenzati da fattori quali la velo-cità media della corrente o la tem-peratura dell’acqua. Tre taxa car-nivori (Perlodes, Rhyacophila, Iso-perla) hanno mostrato la maggiorcapacità di dispersione (70-91% in

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 59
24 h, con una distanza massima di9,5-13,5 m il giorno), seguiti da duetaxa (Protonemura, Rhithrogena) incui circa la metà del numero ini-ziale di individui ha mostrato unabuona capacità di dispersione (50-51% in 24 h, 7,5-8 m/giorno) e,infine, da quattro taxa (Ecdyonu-
rus, Hydropsyche, Gammarus, Bae-tis) in cui meno della metà degliindividui si è spostata (33-40% in24 h, 5,5-7 m/giorno).
Il movimento è stato rivoltocontrocorrente nella maggior partedei casi. Poche larve (20%) di Pota-mophylax si sono spostate, con un
movimento di circa 3,5 m/giornoin entrambe le direzioni. Sono statianche considerati dati provenientida precedenti lavori sul drift. Inconclusione, è stata redatta unalista di taxa con differenti capacitàdi dispersione.
[SF]
ECOLOGIA DELLE ACQUE INTERNE > MACROINVERTEBRATI BENTONICI – FAUNA ITTICA
The influence of scale and geography on relationships between stream communitycomposition and landscape variables: description and predictionC.R. Townsend, S. Dolédec, R. Norris, K. Peacock, C. ArbuckleFreshwater Biology, 48: 768 (2003)C.R. Townsend, Zoology Department, University of Otago, PO Box 56, Dunedin, New Zealand. E-mail: [email protected]
Gli autori hanno analizzato idati relativi alle comunità macro-bentoniche ed ittiche, raffrontan-doli con i dati derivati dall’analisiterritoriale (GIS), con la finalità diesaminare l’esistenza di possibilirelazioni a tre differenti scale (alivello di bacino, di tratto fluviale edi caratteristiche del singolo seg-mento). È stato utilizzato un ap-proccio esplorativo, basato sulladescrizione di eventuali relazioni,ed uno predittivo, basato sul pro-gramma AUSRIVAS.
Le variabili ambientali che
maggiormente influenzano le ce-nosi macrobentoniche sono perlo-più “naturali” se si considera lascala di bacino (geomorfologia,estensione), con alcune influenzeantropiche, quali la presenza agri-cola nelle aree ripariali.
Per quanto riguarda le comu-nità ittiche, le variabili “naturali”mostrano una notevole importan-za specialmente alla scala locale.La posizione geografica influenzadirettamente le cenosi macrobento-niche, in parte perchè è in relazio-ne alle caratteristiche ambientali
ed alla disponibilità di risorse ed inparte perché alcuni taxa non colo-nizzano alcune aree: questa in-fluenza geografica è meno evidenteper i gruppi dotati di stadi immagi-nali alati e con grande capacitàdispersiva.
Ragioni storico-biogeografichespiegano, in parte, anche alcunediversità tra le comunità ittiche.Nelle conclusioni del lavoro è evi-denziata una notevole concordan-za tra i dati esplorativo-osservativie le ipotesi predittive.
[SF]
ECOLOGIA DELLE ACQUE INTERNE > FAUNA ITTICA
Fish zonations and guilds as the basis for assessment of ecological integrity of large riversB.G.W. Aarts, P.H. NienhuisHydrobiologia, 500: 157-178 (2003)B.G.W. Aarts, Department of Environmental Studies, University of Nijmegen, Nijmegen, The Netherlands. E-mail: [email protected]
Il concetto di zonazione lon-gitudinale descrive i cambiamentimonte-valle delle proprietà chimi-co-fisiche e biologiche dei fiumi. Sevengono incluse anche le informa-zioni relative alle associazioni eco-
logiche dei pesci, è possibile incre-mentare l’utilità del concetto di zo-nazione ittica, nel senso che essapuò essere usata come strumentoper la valutazione e la gestione del-l’integrità ecologica dei grandi fiu-
mi.Nell’articolo viene presentata
una caratterizzazione ecologica del-le zone e delle comunità ittichenei grandi fiumi semi-naturali eregolati in Europa (il fiume Doubs

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)60
in Francia ed i fiumi Rhine e Meu-se in Olanda), utilizzando le classi-ficazioni per associazione di moltecaratteristiche dei pesci e delle listerosse nazionali delle specie minac-ciate.
L’insieme dei dati del fiumeDoubs è stato inoltre analizzatousando indici della sensibilità del-le specie ittiche alla alterazione del-l’ambiente e indici di distribuzio-ne.
Nei fiumi studiati, il numerodi associazioni ecologiche aumen-ta verso valle e si osservano chiarevariazioni nella loro struttura. Lepreferenze rispetto alla corrente el’ecologia riproduttiva dei pesci sonostrettamente correlate.
La proporzione delle speciereofile nelle associazioni ittiche di-minuisce verso valle dove aumen-tano le proporzioni delle specie li-mnofile e di quelle che hanno am-pia distribuzione. Le specie che de-
pongono le uova su substrati dipietra (lithophilic spawners) e sullasabbia (psammophilic spawners)sono dominanti nelle zone superio-ri, mentre le zone più basse sonodominate dalle specie che depongo-no le uova sulla vegetazione(phytophilic spawners) e su substratidi ciottoli e vegetazione (phytoli-thophilic spawners).
La proporzione delle specieche si nutrono di zoobenthos e diperiphython diminuisce verso val-le, mentre aumenta quella deglizooplanctivori e degli erbivori. Tut-tavia, essendo la fauna ittica euro-pea costituita principalmente daspecie con alimentazione generali-sta, le discriminanti che si basanosemplicemente sulle associazionialimentari non sono molto alte. Leassociazioni delle specie sensibili econ tolleranze ristrette (stenoecie)aventi strategie fortemente adattatea specifiche condizioni fluviali (re-
ofile o liomnofile), hanno contri-buito alla discriminazione moltodi più delle specie generaliste chepossono sopravvivere in un’ampiavarietà di habitat, anche non carat-teristici degli ecosistemi fluviali na-turali.
A causa della sovrabbondan-za delle specie ad ampia distribu-zione, le zonazioni ittiche longitu-dinali sono appena riconoscibili neigrandi fiumi pesantemente impat-tati come il Rhine. Quindi questifiumi non rispondono ai test diverifica dell’integrità ecologica.
All’interno di una specificaregione ittica, un metodo adattoper analizzare e monitorare l’im-patto del disturbo antropico sullastruttura delle comunità ittiche è ilconfronto tra la struttura delle as-sociazioni dello stato attuale di unazona ittica e quella di una situazio-ne di riferimento.
[MA]
ECOLOGIA DELLE ACQUE INTERNE > CONTINUUM FLUVIALE
Benthic insects and fish of the Doubs River system: typological traits and the develop-ment of a species continuum in a theoretically extrapolated watercourseJ. Verneaux, A. Schmitt, V. Verneaux, C. ProuteauHydrobiologia, 490: 63-74 (2003)J. Verneaux, Laboratoire de Biologie et Ecophysiologie -EA 3184 USC INRA. Pôle Hydrobiologie. Institut des Sciences et Techniques del’Environnement, Université de Franche-Comté, Place Leclerc, 25030 Besançon cedex, France. E-mail: [email protected]
Utilizzando i modelli di di-stribuzione degli insetti bentonici(198 specie) e dei pesci (29 specie)da 11 tributari e dall’asta principa-le del bacino idrografico del fiumeDoubs (Jura francese), gli autorihanno provato a stabilire l’esisten-za di un’organizzazione delle spe-cie in comunità separate e identifi-cabili.
L’analisi delle componentiprincipali (PCA) è stata utilizzata
per identificare un gradiente conti-nuo e per selezionare i 50 siti aminor disturbo, utilizzati per defi-nire un corso d’acqua di riferimen-to teorico. Le classi di densità diogni specie sono state messe in re-lazione con questi gradienti longi-tudinali e la risposta di ogni specieè stata definita da due caratteristi-che tipologiche: le sue preferenzetipologiche (tp) e la sua ampiezzatipologica (ta), generando in questo
modo una sintesi delle caratteristi-che ecologiche.
Nell’indice tipologico (descrit-to nell’appendice dell’articolo) le210 specie, che formano un model-lo biologico, sono elencate in ordi-ne alfabetico con i loro valori di tpe di ta. Queste caratteristiche tipo-logiche delle specie costituisconoutili contributi per una banca datiper il biomonitoraggio delle acquecorrenti. [MA]

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 61
ECOLOGIA DELLE ACQUE INTERNE > SPECIE INVASIVE
Impacts of freshwater invaders at different levels of ecological organisation, with empha-sis on salmonids and ecosystem consequencesK.S. Simon and C.R. TownsendFreshwater Biology, 48: 982 (2003)K.S. Simon, Department of Biology, James Madicon University, Harrisonburg, VA 22807, USA. E-mail: [email protected]
logici multipli. Questi pesci posso-no innescare la cascata trofica do-vuta all’incremento e alla produ-zione della biomassa algale e sonoresponsabili dei cambiamenti deiflussi energetici e dei nutrienti neicorsi d’acqua e nei laghi. I mecca-nismi alla base di questi cambia-menti sono differenti per i due si-stemi e diventano evidenti soltantoquando le informazioni sono con-siderate ai livelli di individuo e dipopolazione.
Nei corsi d’acqua, i salmoni-di possono alterare il comportamen-to degli invertebrati sopprimendo ilpascolo del perifiton. Nei laghi, ilcomportamento alimentare dei sal-monidi può stimolare il fitoplan-cton deviando le sostanze nutrientidal litorale alla zona pelagica.
Lo studio simultaneo a diver-si livelli ecologici dovrebbe fornireuna maggiore comprensione dei
meccanismi che sono alla base del-l’impatto degli animali e delle pian-te invasive, fornendo così una piùsolida base per predire l’impattodelle specie invasive d’acqua dolce.
Le caratteristiche delle specieinvasive che possono essere asso-ciate ad impatti particolarmentegravi includono: un metodo di ac-quisizione delle risorse inizialmen-te assente nel sistema invaso, unalarga nicchia trofica con collega-menti tra comparti dell’ecosistemainizialmente sconnessi, rapporti ali-mentari con conseguenze negativeper le specie native, caratteristichefisiologiche che accrescono la tra-sformazione delle risorse e produ-cono un’alta biomassa, e caratteri-stiche comportamentali o demogra-fiche che forniscono alta resistenzao resilienza nel campo della disper-sione naturale.
[MA]
Le specie esotiche possono in-fluenzare i sistemi d’acqua dolce allivello di individuo, di popolazio-ne, di comunità e di ecosistema.Alcuni di questi effetti possono es-sere semplici o difficili da prevede-re, ma possono essere importantiper comprendere i cambiamenti piùevidenti.
Tuttavia, gli studi sugli effettidelle specie esotiche d’acqua dolceai diversi livelli dell’organizzazio-ne ecologica sono rari. Vengonocomunemente segnalati i cambia-menti della distribuzione o dell’ab-bondanza delle popolazioni dopol’introduzione, mentre è rara la do-cumentazione che riguarda l’im-patto sul funzionamento dell’ecosi-stema e i flussi dell’energia e deinutrienti.
Diversamente dalla maggiorparte delle specie invasive, i salmo-nidi sono stati studiati a livelli eco-
ECOLOGIA DELLE ACQUE INTERNE > ACARI (IDRACNIDI)
Biodiversity and distribution of water mites (Acari, Hydrachnidia) in spring habitatsA. Di Sabatino, B. Cicolani and R. GereckeFreshwater Biology, 48: 2163 (2003)A. Di Sabatino, Dipartimento di Scienze Ambientali, University of L’Aquila, Via Vetoio 20, Coppito, I-67100 L’Aquila, Italy. E-mail:[email protected]
Gli autori hanno studiato ladistribuzione, la ricchezza in spe-cie e l’ecologia degli acari che vivo-no nelle acque sorgive in Italia; loscopo è stato quello di spiegare me-glio il ruolo degli habitat di sorgen-te nel sostenere alti livelli di biodi-versità ed evidenziare il loro contri-buto alla biodiversità delle acque
correnti a scala locale e regionale.Sono state esaminate più di
300 sorgenti in differenti aree geo-grafiche (Alpi, Appennini centrali emeridionali, Sicilia e Sardegna),portando al riconoscimento di untotale di 163 specie di acari acqua-tici. La ricchezza in specie in ogniarea è compresa in un intervallo
che va da 33 specie in Sardegnafino a 77 in Sicilia. La diversità piùalta è stata riscontrata sul GranSasso (Appennini centrali). La fra-zione di crenobionti (specie stretta-mente legate a questo tipo di habi-tat) supera il 50% in quasi tutte learee investigate.
Una cenosi ad acari diversifi-

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)62
cata (oltre 20 specie per sorgente) ealtamente specializzata è stata os-servata in rheocrenes indisturbati ein sorgenti naturali di tipologia in-termedia (rheohelocrenes e rheop-sammocrenes).
In sorgenti sottoposte ad im-patti antropici (pascoli, deforesta-zione, alterazione e trasformazionedelle sorgenti) la ricchezza in spe-cie diminuisce e gli organismi cre-nobionti sono sostituiti da crenoxe-
correnti e possano contribuire peralmeno un terzo alla biodiversitàdelle acque correnti su scala regio-nale. La presenza di crenobiontiendemici e taxa rari mette in lucel’importanza di questi habitat nelmantenimento di alti livelli di bio-diversità, così come il fatto che essicontribuiscono ad una migliorecomprensione dei modelli di biodi-versità delle acque correnti.
[ARM]
ni non specializzati.L’importanza zoogeografica
degli habitat di sorgente è confer-mata dalla presenza di 18 specieendemiche e da gruppi di genericon una interessante distribuzionedisgiunta e relitta.
Un confronto con altre areegeografiche suggerisce che questesorgenti contengano una parte si-gnificativa del numero totale di spe-cie ritrovate negli habitat di acque
ECOLOGIA DELLE ACQUE INTERNE > ANALISI DELL’AMBIENTE FLUVIALE
Habitat change in braided flood plains (Tagliamento, NE-Italy)D. van der Nat, K. Tockner, P.J. Edwards, J.V. Ward and A.M. GurnellFreshwater Biology, 48: 1799 (2003) K. Tockner, Department of Limnology, EAWAG/ETH, überlandstrasse 133, 8600 Dübendorf, Switzerland. E-mail: [email protected]
Sono stati studiati i cambia-menti relativi e la distribuzione dietà di habitat nell’alveo attivo diun tratto del Fiume di Tagliamento(NE-Italia) con presenza di isole ebarre. Tra il settembre 1999 ed ilgennaio 2002, in cinque occasioni,sono stati individuati sei tipi di ha-bitat mediante GPS durante diversecondizioni di portata. È stata uti-lizzata la sovrapposizione di map-pe per calcolare l’età ed il cambia-mento relativo di habitat e sonostati stabiliti i tassi di decadimentoesponenziali (k-valori) per le isole ei più importanti habitat acquatici.
I cambiamenti relativi di tuttigli habitat combinati sono stati dioltre l’82% tra le date di osservazio-
ne nel tratto di alveo con barre,con una tasso cumulativo di 85%su un periodo di 2,5 anni. I cam-biamenti relativi nel tratto di alveocon isole sono stati minori, conuna variazione cumulativa di circail 60% durante il periodo di studio.Nel tratto di alveo con barre sonostate stabilite significative relazioniesponenziali di scomparsa per ca-nali attivi, canali alluvionali, brac-ci d’acque ferme e stagni.
I tempi di dimezzamento era-no particolarmente brevi per i brac-ci d’acque ferme e gli stagni. Neltratto di alveo con isole esistevanorelazioni significative per canaliattivi e canali alluvionali. I tempidi dimezzamento di canali attivi e
canali alluvionali crescevano conla presenza di isole vegetate che,quindi, forniscono stabilità al siste-ma. La composizione relativa del-l’habitat all’interno del corridoioattivo è rimasta quasi costante, so-stenendo l’applicabilità del model-lo dello stato stazionario con spo-stamento a mosaico agli ecosistemidi piana alluvionale meandrizzata.
I risultati indicano che, incondizioni naturali, gli habitat ac-quatici della piana alluvionale pos-sono essere estremamente dinamicisu brevi scale temporali. Anchemodeste piene possono condurre agrossi cambiamenti degli habitat,con importanti conseguenze per lafauna e flora. [ARM]
ECOLOGIA DELLE ACQUE INTERNE > ACIDIFICAZIONE DEI CORSI D’ACQUA
Causes of episodic acidification in Alpine streamsF. Lepori, A. Barbieri and S.J. OrmerodFreshwater Biology, 48: 175 (2003)S.J. Ormerod, Catchment Research Group, Cardiff School of Biosciences, Cardiff University, PO Box 915, Cardiff CF10 3TL, UK. E-mail:[email protected]
Nonostante la loro importan-za complessiva/globale di ambienti
di montagna, gli effetti delle deposi-zioni acide nei corsi d’acqua delle
Alpi non sono ancora stati descritticompletamente. In questo studio è

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 63
relati alla distribuzione delle depo-sizioni atmosferiche di azoto.
Importanti diminuzioni di al-calinità si sono riscontrate nellamaggior parte dei corsi d’acqua adalta portata durante lo scioglimen-to delle nevi (in primavera) o du-rante temporali (in primavera edautunno). Gli episodi autunnali sispiegano con la diluizione dell’al-calinità delle acque dei torrenti daparte dell’acqua piovana. Gli episo-di primaverili, invece, coinvolgonodiluizioni ed incrementi di azotonitrico, e l’aumento in nitrati è
risultato correlato con la distribu-zione delle deposizioni di azoto. Laconcentrazione di SO4
2– è diminui-ta durante la maggior parte deglieventi.
Elevate lisciviazioni di NO3
in condizioni di bassa portata indi-cano che i suoli del bacino idrogra-fico nel Canton Ticino stanno rag-giungendo la saturazione di azoto.Questi dati sono tra i primi cheillustrano come l’azoto nitrico pos-sa portare a episodi di acidificazio-ne in Europa.
[RC]
stato valutato il potere tamponantedi 30 corsi d’acqua nel Canton Ti-cino (Svizzera), a bassa ed alta por-tata, lungo un gradiente sud/norddi deposizioni acide durante il2000.
In condizioni di bassa porta-ta, nessun corso d’acqua è risultatoacido, avendo valori di alcalinitàdi 29 eq. L-1 e pH 6,4. Tuttavia, aquesta portata l’azoto nitrico erapresente in concentrazioni tali dasuggerire la presenza di una fonteantropica; infatti i suoi valori diconcentrazione sono risultati cor-
ECOLOGIA DELLE ACQUE INTERNE > ANALISI BIBLIOGRAFICHE
A perspective on the key citations in freshwater benthic science, and the studies thatinfluenced themV.H. Resh, N.G. KobzinaJournal of the North American Benthological Society, 22: 341–351 (2003)V.H. Resh, Department of Environmental Science, Policy and Management, University of California, Berkeley, California 94720-3112 USA
Gli autori hanno ricercato nelScience Citation Index (SCI) gli ar-ticoli ed i libri che compaiono al-meno 12 volte nel Journal of theNorth American Benthological So-ciety (J-NABS) tra il 1995 ed il 2000,per individuare i lavori sulla scien-za del benthos maggiormente cita-
ti, sia in una rivista specializzata(J-NABS) sia nella letteratura scien-tifica generale (SCI). Entrambe lericerche indicano che il concettodel river continuum (Vannote et al.,1980. Canadian Journal of Fisheriesand Aquatic Sciences, 37: 130-137)rappresenta l’articolo più citato.
Testi di statistica generale, manua-li metodologici e di identificazionecostituiscono invece la maggior par-te dei libri più citati. Gli autori dei10 articoli più citati apparsi sul J-NABS commentano le idee che han-no maggiormente influenzato i pro-pri lavori. [PG]
ECOLOGIA DELLE ACQUE INTERNE > STATO ECOLOGICO DEI LAGHI > MACROINVERTEBRATI
The use of littoral mesohabitats and their macroinvertebrate assemblages in the ecologi-cal assessment of lakesJ. White, K. IrvineAquatic Conservation: Marine and Freshwater Ecosystems, 13: 331-351 (2003)J. White, Marine Institute, 80 Harcourt Street, Dublin 2, Ireland. E-mail: [email protected]
Storicamente, la valutazioneecologica dei laghi viene effettuatamediante analisi chimico-fisiche efitoplanctoniche delle acque pela-giche e lo stato trofico è determina-to da clorofilla a e fosforo totale,secondo il protocollo generaleOECD per la classificazione dei la-
ghi. Questa prassi andava/va di paripasso con l’opinione generale chela zona litorale dei laghi è troppoeterogenea da permettere un con-fronto significativo tra diversi la-ghi.
La recente Direttiva Europea2000/60/EC (Water Framework Di-
rective) richiede un approccio piùampio per la valutazione ecologicadei laghi, inclusa la necessità diincorporare un certo numero divariabili biologiche. Questo artico-lo descrive i risultati di un pro-gramma di monitoraggio finalizza-to alla determinazione di variazio-

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)64
ni –all’interno di uno stesso lago etra laghi diversi– delle comunità dimacroinvertebrati in mesohabitatdi 22 laghi, appartenenti a diversetipologie.
Campioni (in triplo) di ma-croinvertebrati prelevati da mesoha-bitat di 11 laghi, con substrati pre-valenti a pietre e ciottoli, indicanoche le comunità prelevate da unhabitat standard risultano confron-tabili e robuste. Campioni (in dop-pio) da mesohabitat diversi, a pie-tre, ciottoli e macrofite, prelevati da21 laghi risultano avere una mag-gior similarità all’interno di unostesso lago, piuttosto che tra laghi
diversi. I risultati dimostrano cheperfino in ambienti differenti, lastruttura di comunità dei macroin-vertebrati può essere un indicatorelago-specifico attendibile.
L’analisi multivariata di cam-pioni prelevati da 15 diversi me-sohabitat in uno stesso lago indicache la variazione all’interno di unlago può comunque venire identifi-cata da gruppi distinti di comunitàmacrobentoniche.
In conclusione, questo lavoroindica che le variazioni all’internodi un lago sono comprese nelle va-riazioni esistenti tra laghi apparte-nenti a diverse tipologie e che le
comunità di macroinvertebrati pos-sono essere un utile strumento nel-la valutazione di differenze ecolo-giche tra laghi. Il loro monitorag-gio, tuttavia, può essere disturbatoda un sostanziale “rumore di fon-do” dei dati ottenuti, riflettendo lacomplessità della struttura dellecomunità macrobentoniche dellezone litorali dei laghi. Si evidenziainoltre che l’inclusione dei macro-invertebrati nella valutazione eco-logica è più efficace quando vieneconfinata a definiti mesohabitat,piuttosto che ad un ampio interval-lo di mesohabitat all’interno di unsingolo lago. [RC]
ECOLOGIA DELLE ACQUE INTERNE > EUTROFIZZAZIONE
Man-made lakes in Mediterranean semi-arid climate: the strange case of Dr Deep Lakeand Mr Shallow LakeL. Naselli-FloresHydrobiologia, 506-509: 13-21 (2003)L. Naselli-Flores, Dipartimento di Scienze Botaniche, Università di Palermo, Via Archirafi 38, I-90123 Palermo. E-mail: [email protected]
La mancanza di ogni tipo diprotezione contro l’eutrofizzazionesta progressivamente comprometten-do la qualità delle acque delle ri-serve siciliane. Questi corpi idriciforniscono alla popolazione un ade-guato approvvigionamento di ac-qua potabile e soddisfano la mag-gior parte del fabbisogno irriguodell’agricoltura locale. Inoltre, rea-giscono al clima mediterraneo coneventi di esondazione e di seccasequenziali e prevedibili su basestagionale, la cui intensità varia in
modo vistoso secondo gli anni.Come conseguenza della sic-
cità estiva e della politica di soddi-sfare la domanda di acqua in tuttala stagione, le riserve subiscono gros-si decrementi di acqua, dando luo-go ad una riduzione del 90% rispet-to al volume di stoccaggio primave-rile. Quindi essi cominciano la sta-gione idrologica come corpi idriciabbastanza profondi da formareuna stratificazione stabile –anchese il termoclino generalmente spa-risce a metà della stagione estiva–
e, come conseguenza del vistoso ab-bassamento di livello, si trasforma-no in corpi idrici poco profondi.Questo duplice comportamento,dovuto ad un progressivo e cospi-cuo decremento della profondità,interferisce con le dinamiche deinutrienti e del fitoplancton, favo-rendo fenomeni di eutrofizzazionee contribuendo alla selezione dicianobatteri planctonici potenzial-mente pericolosi per la salute uma-na.
[ARM]
ECOLOGIA DELLE ACQUE MARINE > BIODIVERSITÀ - TASSONOMIA
Biodiversity, conservation, and the “Taxonomic impediment”A. GiangrandeAquatic Conservation: Marine and Freshwater Ecosystems, 13: 451-459 (2003)A. Giangrande, Dipartimento di Scienze e Tecnologie Biologiche e Ambientali, Università di Lecce, Laboratorio di Biologia Marina, 73100Lecce, Italy. E-mail: [email protected]
L’articolo mette in evidenzaquanto la tassonomia, come scien-
za autonoma, sia tenuta in scarsaconsiderazione; ne vengono inoltre
discusse le implicazioni per la bio-logia della conservazione

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 65
In tempi recenti si è verificatala tendenza a trascurare la tasso-nomia a livello di specie non solonelle ricerche di tipo ecologico, maanche nei processi di individuazio-ne e di motivazione della selezionedelle aree marine protette (MPA).Un criterio tradizionale per la scel-ta di una MPA è la conservazionedella biodiversità, ma la maggiorparte della MPA italiane sono statescelte senza studi iniziali dettaglia-ti sulla loro biodiversità, così chenon sono disponibili elenchi di spe-cie relative ai principali gruppi diinvertebrati.
L’identificazione degli orga-nismi a livello di specie all’internodelle comunità è uno dei più gros-si problemi in termini di tempo ecosti negli studi ecologici. Alcunistudi hanno suggerito che il lavo-ro a un livello tassonomico supe-riore alla specie non implica unaperdita di informazioni importan-te (sufficienza tassonomica). Essoporta comunque ad una inaccura-tezza della valutazione della bio-diversità, molto importante nellacomparazione di differenti aree, epuò condurre ad una esclusione apriori di alcune entità prima della
te la perdita di specie, un temaimportante non solo per gli ecologima anche per i politici. Sembra chemolte specie abbiano sovrapposi-zione di nicchie e, come si puòintuire, non sia essenziale la con-temporanea presenza di tutte le spe-cie. Al contrario, è stato afferrato ilruolo cruciale delle specie chiavenella biologia della conservazionecome uno strumento che aiuta amettere in evidenza le specie cherichiedono priorità per la loro pro-tezione.
L’attuale conoscenza dei si-stemi marini ha portato alla con-clusione che, prima di sviluppareteorie e disegni sperimentali, abbia-mo bisogno di una descrizione ap-propriata del sistema oggetto di stu-dio.
Una conoscenza di base dellabiodiversità generale in termini diricchezza in specie è ad esempioessenziale per proporre una MPA,insieme ad un esame dettagliatoche fornisca le liste tassonomichenecessarie per la caratterizzazionedei biotopi ed un set di dati diriferimento per comparazioni futu-re.
[ARM]
comprensione del loro ruolo eco-logico.
La tassonomia è stata sempreconsiderata una scienza marginaleanche durante il periodo descritti-vo di scoperta dell’ecologia e, tradi-zionalmente, ha ricevuto pochi sup-porti finanziari. Il risultato è statola produzione di molte identifica-zioni e segnalazioni sbagliate. Du-rante gli ultimi anni, l’approccioecologico sperimentale ha condot-to ad un miglioramento dei metodiscientifici, ma contemporaneamen-te ad una riduzione del numero ditassonomi esperti per molti gruppidi invertebrati. Lavori descrittivi,storicamente così comuni nell’areaMediterranea, sono ora consideratiobsoleti nonostante abbiano unvalore intrinseco.
Si è a lungo pensato che labiodiversità, e in particolare la ric-chezza in specie, abbia un’influen-za sulla variabilità temporale e sem-bra che gli sforzi per chiarire larelazione tra variabilità temporalee biodiversità, o per dimostrare lamancanza di tale relazione, potreb-bero continuare. Tale informazio-ne è essenziale al fine di mantene-re la funzione ecologica nonostan-
ECOLOGIA DELLE ACQUE MARINE > MACROBENTHOS
Hitch-hiking on floating marine debris: macrobenthic species in the Western Mediterra-nean SeaS. Aliani, A. MolcardHydrobiologia, 503: 59-67 (2003)S. Aliani, CNR, Istituto Scienze Marine - Sezione La Spezia, Forte Santa Teresa, 19036 Pozzuolo di Lerici (SP). E-mail:[email protected]
Si definiscono detriti marini imateriali solidi di origine umanascaricati direttamente in mare oche raggiungono il mare attraversoi corsi d’acqua. Gli effetti dei detritimarini sulla vita selvatica, sul turi-smo e sulla salute umana sono bendocumentati ed esiste una conside-revole letteratura scientifica a pro-
posito dei detriti plastici in mare esul fondo marino, che mette in ri-salto soprattutto il possibile impat-to sui mammiferi marini e sul turi-smo.
La dispersione di organismimarini e terrestri su oggetti galleg-gianti ha un interesse biogeografi-co ed ecologico. Per alcune specie
l’estensione dell’area di distribuzio-ne geografica è più probabilmentecollegata al trasporto di individuimaturi su zattere galleggianti piut-tosto che ad una dispersione attivao passiva di propaguli riproduttivi.La variabilità e la varietà dei mate-riali galleggianti è drammaticamen-te cresciuta negli anni recenti ed i

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)66
detriti marini sono stati ampiamen-te usati come una zattera da specieche si fanno trasportare.
L’articolo descrive i macroin-vertebrati bentonici che vivono suidetriti marini trasportati dal ventoe dalle correnti di superficie nelMar Mediterraneo occidentale. Leplastiche sono annoverate come
pesci vengono comunemente rinve-nuti in grosse buste di plastica. Vie-ne fornita una stima delle distanzeche possono essere coperte dallespecie che si fanno trasportare edel contributo del materiale galleg-giante alla dispersione teorica dellespecie.
[ARM]
costituenti della maggior parte de-gli oggetti di detrito a causa dellaloro bassa biodegradabilità; comun-que tra di essi si ritrovano anchevetro, lattine, reti da pesca e conte-nitori di polistirolo. Il macroben-thos che vive sul materiale galleg-giante comprende soprattutto mol-luschi, policheti e briozoi. Grandi
ECOLOGIA DELLE ACQUE MARINE > FAUNA ITTICA > SPECIE ESOTICHE
A hotspot of non-native marine fishes: evidence for the acquarium trade as an invasionpathwayB.X. Semmens, E.R. Bhule1, A.K. Salomon, C.V. Pattengill-SemmensMarine Ecology Progress Series, 266: 239-244, 2004B.X. Semmens, University of Washington, 24 Kincaid Hall, Department of Biology, Box 351800, Seattle, Washington 98195-1800, USA.
L’invasione delle specie esoti-che negli ecosistemi marini può es-sere ecologicamente dannosa ed eco-nomicamente costosa. Per incremen-tare al massimo l’efficacia dei pro-grammi tesi ad ostacolare l’invasio-ne delle specie non indigene è ne-cessario identificarne i “punti foca-li” e le fonti di introduzione.
Utilizzando una puntuale edampia banca dati dei pesci di mare,si è osservato che sulle scogliere
della Florida sudorientale (U.S.A.)è presente un sorprendente nume-ro di pesci esotici. Per spiegare ilfenomeno della presenza di pescimarini esotici in questa regione visono due ipotesi: l’introduzione at-traverso lo scambio dell’acqua dizavorra (acqua contenuta nei ser-batoi delle navi per aumentarne lastabilità) e le introduzioni dagli ac-quari. Per valutare il grado di coin-volgimento di questi due vettori
sono stati usati i dati relativi aimodelli di navigazione internazio-nale ed alle importazioni dei pescidi mare.
I risultati indicano che le in-troduzioni sono dovute al rilasciodagli acquari. Per prevenire ulte-riori rilasci e invasioni sono neces-sari l’educazione, l’estensione ed ilpotenziamento delle iniziative ri-volte agli acquariofili marini e al-l’industria degli acquari. [MA]
ECOTOSSICOLOGIA > MICROCOSMI ACQUATICI
Toxicity to freshwater organisms from oils and oil spill chemical treatments in laboratorymicrocosmsS. Bhattacharyya, P.L. Klerks, and J.A. NymanEnvironmental Pollution, 122: 205-215 (2003)S. Bhattacharyya, Department of Biology, University of Louisiana at Lafayette, PO Box 42451, Lafayette, LA 70504, USA
Gli autori hanno studiato latossicità ed i suoi cambiamenti tem-porali in microcosmi di acque cor-renti e stagni contenenti petroliogreggio della Luisiana del sud (SLC)o carburante diesel, e trattati conun depurante o un disperdente. Gliorganismi usati sono: Chironomustentans, Daphnia pulex e Oryzias la-
tipes.Per i saggi biologici è stato
usato un microcosmo acquatico (D.pulex e O. latipex) o fango da suolo(C. tentans) prelevati dopo 1, 7, 31 e186 giorni dal trattamento.
I risultati hanno mostrato che:SLC è meno tossico del diesel, gliadditivi chimici aumentano la tos-
sicità, il disperdente è più tossicodel depurante e la tossicità si ridu-ce fortemente al 186° giorno. Latossicità è risultata più elevata neisaggi con la specie bentonica ri-spetto a quelli con le altre due spe-cie che vivono nella colonna d’ac-qua. Un esperimento separato hamostrato che la sensibilità di C.

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 67
tentans è intermedia tra quella diTubifex tubifex e quella di Hyallelaazteca.
In conclusione, gli organismidi acqua corrente, soprattutto lespecie bentoniche, appaiono seria-
mente danneggiate dal petrolio nel-lo scenario del caso peggiore realiz-zato nei microcosmi. [ARM]
ECOTOSSICOLOGIA > DAPHNIA MAGNA
Is Cr(VI) toxicity to Daphnia magna modified by food availability or algal exudates? Thehypothesis of a specific chromium/algae/exudates interactionG. Gorbi, M.G. Corradi, M. Invidia, L. Rivara and M. BassiWater Research, 36: 1917-1926 (2002)G. Gorbi, Dipartimento di Scienze Ambientali, Università di Parma, Viale delle Scienze, I-43100 Parma. E-mail: [email protected]
Nell’articolo vengono esami-nati gli effetti a lungo termine sulciclo vitale di Daphnia magna Strausda parte del cromo esavalente inrelazione a differenti condizioni dinutrimento od alla presenza di es-sudati di Scenedesmus acutus; que-sti ultimi hanno mostrato di ridur-re la tossicità del cromo nei con-fronti dell’alga.
In presenza di abbondantidosi di cibo, una concentrazionepari a 14 g Cr(VI)/L ha avuto effettinegativi sulla sopravvivenza, sullacrescita e sulla fecondità.
Con basse dosi di cibo, gli
effetti sulla crescita non erano evi-denti, ma i dafnidi cambiavano lapropria strategia riproduttiva nor-malmente adottata in casi di caren-za di nutrimento e mostravano untasso giornaliero di produzione dineonati più elevato del controllo;comunque, i neonati erano piùpiccoli e meno resistenti alla caren-za di cibo.
L’effetto più rilevante del cro-mo è stato l’accorciamento dellavita media dei dafnidi, che è risul-tato indipendente dalle condizionidi nutrimento ed ha avuto luogoanche quando la crescita e il tasso
di produzione di neonati non era-no alterati.
Gli essudati algali sono risul-tati inefficaci nel ridurre la tossici-tà del cromo nei confronti dei daf-nidi, ma hanno causato una forteriduzione della fecondità. Ulterioriesperimenti condotti con S. acutushanno dimostrato che gli effetti didetossificazione osservati nell’algapotrebbero implicare una interazio-ne specifica cromo/alga/essudati, laquale ha luogo solamente quandole alghe sono preventivamente sog-gette ad uno stress a breve termineda cromo.
[PG]
ECOTOSSICOLOGIA > BATTERI BIOLUMINESCENTI
Evaluation of wastewater toxicity: comparative study between Microtox® and activatedsludge oxygen uptake inhibitionM. Gutiérrez, , J. Etxebarria and L. de las FuentesWater Research, 36: 919-924 (2002)M. Gutiérrez, GAIKER, Parque Tecnológico, Edif. 202, E-48170 Zamudio, Spain. E-mail: [email protected]
Il sistema Microtox® è unostrumento utilizzato di frequenteper lo screening tossicologico delleacque reflue che alimentano gliimpianti di trattamento. Allo statoattuale esiste una crescente contro-versia sull’impiego di questo siste-ma rispetto ad altri che utilizzano ifanghi attivi. Nell’articolo viene pre-sentato uno studio tossicologicocomparativo tra il Microtox® e leprove respirometriche. Le provesono state svolte con sette composti
organici e cinque composti inorga-nici tossici.
Il Microtox® ha mostrato unamaggiore sensibilità nei confrontidelle sostanze tossiche, ma è risul-tato, rispetto alle prove respirome-triche, meno rappresentativo circagli effetti sui fanghi attivi. Per esem-pio, i saggi condotti con LAS, untensioattivo di riferimento biodegra-dabile, hanno mostrato un effettotossico per il Microtox® ma, allostesso tempo, una buona biodegra-
dabilità e nessuna tossicità per laprova respirometrica. Ciò potrebbeessere dovuto dalla differente natu-ra del materiale biologico impiega-to, dal momento che il Microtox®
utilizza il microrganismo marinoVibrio fisheri, mentre le prove respi-rometriche utilizzano il consorziobatterico presente nei fanghi attivi.
In conclusione, per la valuta-zione della potenziale tossicità diun composto in un impianto ditrattamento delle acque reflue, il

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)68
materiale biologico preferenzial-mente impiegato dovrebbe essere il
fango attivo stesso. I risultati otte-nuti con qualunque altro materia-
le biologico sarebbero solo un’ap-prossimazione della realtà. [PG]
RINATURAZIONE E RIQUALIFICAZIONE AMBIENTALE > CORSI D’ACQUA
Assessing stream ecosystem rehabilitation: limitations of community structure dataS.S. Brooks, M.A. Palmer, B.J. Cardinale, C.M. Swan, S. RibblettRestoration Ecology, 10: 156-168 (2002)S.S. Brooks, Department of Biology, University of Maryland, College Park, MD 20742, U.S.A.
L’uso improprio del territorio,gli inquinanti, lo sfruttamento ed ilsovrappopolamento hanno banaliz-zato gli habitat fluviali e degradatola qualità delle acque in tutto ilmondo. Gli enti che si occupanodella gestione vengono sempre piùspesso incaricati di mitigare gli im-patti e ripristinare la “salute” deifiumi; tuttavia c’è una carenza dimetodi scientifici rigorosi e di unateoria sulla quale basare un validoprogetto di ripristino e monitorag-gio. Nonostante ciò, vengono rea-lizzati molti progetti di recupero ascala locale per stabilizzare l’ero-sione e migliorare l’eterogeneitàdegli habitat fluviali.
Molti tentativi di recupero sibasano sul paradigma secondo cuil’incremento della eterogeneità de-gli habitat conduce al recupero del-la biodiversità; tuttavia sono statirealizzati pochi studi che abbianoconsiderato la sola variazione diun parametro fisico indipendentedal mezzo al fine di isolare gli effet-ti dell’eterogeneità per sé.
Gli autori hanno condotto unesperimento in campo per simulare
il recupero dell’eterogeneità deglihabitat in un corso d’acqua pocoprofondo a fondo ciottoloso. Utiliz-zando un approccio sperimentalepiuttosto che una valutazione det-tagliata del lavoro di recupero, gliautori sono stati in grado di con-trollare le condizioni iniziali di ra-schi successivi, in modo che le ri-sposte degli organismi potessero es-sere attribuite senza ambiguità agliinterventi sull’eterogeneità.
Lo studio ha previsto la ma-nipolazione della variabilità dellagranulometria del substrato in al-veo e, conseguentemente, le carat-teristiche del flusso in corrispon-denza di interi raschi. Questi duefattori definiscono gradienti di va-riazione dell’eterogeneità degli ha-bitat a livelli tali da interessare lafauna di macroinvertebrati residen-te.
Ciononostante non è stato pos-sibile evidenziare differenze nellastruttura della comunità tra zonecon forti interventi sull’eterogenei-tà di habitat, rispetto a zone condeboli interventi.
L’analisi statistica indica che
le popolazioni di macroinvertebra-ti sono più sensibili alle specifichecondizioni di ogni singolo raschiopiuttosto che ai trattamenti sull’ete-rogeneità; ciò suggerisce che l’in-cremento dell’eterogeneità deglihabitat può essere una tecnica inef-ficace se lo scopo del ripristino èquello di promuovere la ricompar-sa dei macroinvertebrati in fiumidepauperati.
In presenza di una variabilitàmolto ampia tra i diversi raschi, iprogrammi di monitoraggio per pro-getti di recupero locali o per gliimpatti di sorgenti puntuali nonsono in grado di rilevare in modosoddisfacente i mutamenti gradua-li nella struttura della comunitàfino a quando le differenze tra ilsito di riferimento e quello trattatonon diventino molto grandi. Misu-re innovative di altri parametri qua-li le variabili di funzione dell’ecosi-stema (per esempio la produzione,la respirazione, la decomposizione)possono essere indicatori più ap-propriati di cambiamenti a scalalocale.
[ARM]
An assessment of a small urban stream restoration project in Northern CaliforniaA.H. Purcell, C. Friedrich, V.H. ReshRestoration Ecology, 10: 685-694 (2002)A.H. Purcell, Laurel Marcus & Associates, 3661 Grand Avenue, #204, Oakland, CA 94611, U.S.A.
I progetti di recupero fluvialesono diventati sempre più frequen-ti e risulterà impellente la necessitàdi valutazioni sistematiche post-pro-gettuali soprattutto per interventi
su piccola scala. Il presente studiodescrive come sia stato valutato ilrecupero di un tratto di 70 metri diun piccolo fiume urbano, il BaxterCreek (Pointset Park, El Cerrito,
California), usando valutazioni del-l’habitat, analisi biologiche e inda-gini sulle abitudini dei residenti.
Il recupero ha comportatol’apertura di un canale precedente-

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 69
mente interrato, l’impianto di vege-tazione riparia, la creazione di suc-cessioni di raschi e pozze e l’incre-mento della sinuosità. Campiona-menti replicati di macroinvertebra-ti bentonici in siti recuperati e inun sito non recuperato più a mon-te sono stati comparati usando variparametri, tra cui la ricchezza intaxa ed un indice biotico. Nella
sezione sottoposta all’intervento direcupero si è rilevato un migliora-mento sia della qualità biologicache di quella dell’habitat rispettoalla sezione non interessata dal pro-getto. Comunque, le condizioni del-l’habitat appena recuperato risul-tano di qualità più scadente se con-frontate con quelle di un corso d’ac-qua su cui l’intervento era stato
condotto dodici anni prima.Un’indagine tra gli abitanti
delle zone circostanti dimostra ilparticolare gradimento del risulta-to raggiunto nel sito sottoposto arecupero. L’approccio usato in que-sto progetto dimostrativo può esse-re applicato per altre valutazioni diinterventi di ripristino a piccola sca-la di fiumi urbani [ARM]
Ecological rehabilitation of the lowland basin of the river Rhine (NW Europe)P.H. Nienhuis, A.D. Buijse, R.S.E.W. Leuven, A.J.M. Smits, R.J.W. de Nooij, E.M. SamborskaHydrobiologia, 478: 53-72 (2002)P.H. Nienhuis, Department of Environmental Studies, Faculty of Science, University of Nijmegen, P.O. Box 9010, 6500 GL Nijmegen, TheNetherlands. Member of the Netherlands Centre for River Studies (NCR), P.O. Box 177, 2600 MH Delft, The Netherlands
Nel presente studio è stato ri-visitato lo stato del ripristino ecolo-gico del bacino del fiume Reno nel-la zona delle lowlands olandesi. Vie-ne presentata una digressione stori-ca riguardante soprattutto le misu-re di regolazione del fiume nel pas-sato.
Il corso inferiore del Reno ècaratterizzato da un bacino moltoantropizzato, da regimazioni forte-mente regolate, da acque e sedi-menti inquinati e da ecosistemi for-temente deteriorati.
Durante gli scorsi 25 anni laqualità delle acque e, di conseguen-za, la qualità dei sedimenti è consi-derevolmente migliorata portandoad un incremento della diversitàbiologica. Il recupero di ecosistemiabbandonati e disturbati è comin-ciato circa 15 anni fa. Le condizio-ni da cui non si può prescindereper attuare i progetti di recuperosono rappresentate dalla protezio-
ne contro le inondazioni e dal tra-sporto con le navi mercantili. Esi-stono teorie ecologiche olistiche chepromuovono la gestione della regi-mazione fluviale, ma i progetti direcupero sono legati soprattutto allepolitiche di pianificazione locali eregionali.
L’uso e l’applicazione di cri-teri ecologici giocano un ruolo sem-pre più importante negli schemi direcupero. Il ripristino ecologico èfocalizzato principalmente sullaconservazione della natura e sullestrategie di recupero, ad esempiosullo sfruttamento del potenzialeidrodinamico e morfodinamico deifiumi o sull’introduzione di un re-gime di pascolo semi-naturale at-traverso l’utilizzo di grandi erbivo-ri. In particolare, la creazione dinuovi alvei secondari contribuisceal recupero della diversità e dellaeterogeneità degli habitat ripari equindi della diversità specifica.
La valutazione di un certonumero di progetti di ripristino haportato alla conclusione che nonpuò essere fornito un giudizio ge-neralizzato riguardo al “successo”o al “fallimento” di specifiche mi-sure di recupero. Il bilancio gene-rale di 14 progetti valutati, su circa30 in fase di realizzazione, è positi-vo.
Un fiume altamente dinami-co come il Waal offre le miglioripossibilità per il recupero di spon-de sabbiose naturali e delle areegolenali. Sarà molto difficile ripri-stinare le foreste golenali a latifo-glie con le attuali condizioni digestione.
Il successivo recupero del bio-ta fluviale dipende da un decre-mento continuo dell’inquinamen-to, da un incremento della dinami-cità morfologica e dallo sviluppodegli habitat naturali originari.
[ARM]
Towards a decision support system for stream restoration in the Netherlands: an over-view of restoration projects and future needsP.F.M. Verdonschot, R.C. NijboerHydrobiologia, 478: 131-148 (2002)P.F.M. Verdonschot, Alterra, Department of Freshwater Ecology, P.O. Box 47, 6700 AA Wageningen
La riqualificazione fluviale èuna delle risposte al deterioramen-
to dei fiumi di pianura. Per operarescelte corrette nel ripristino fluvia-
le è necessario innanzitutto capirele complesse interazioni spaziali e

Rassegna scientifica Biol. Amb., 18 (n. 2, 2004)70
temporali tra le componenti fisi-che, chimiche e biologiche nell’eco-sistema fluviale. Molti concetti eco-logici sulla quadridimensionalità,la scala e la gerarchia nell’ecosiste-ma fluviale sono stati integrati nelmodello 5-S.
Questo modello fornisce il sup-porto teorico di un primo schemadi un sistema di supporto alle deci-sioni per il recupero fluviale. Lariqualificazione fluviale è in rapi-do sviluppo in Olanda. Nel 1991 sicontavano 70 progetti, nel 1993 era-
no 170 e questo numero è cresciutoa 206 nel 1998. Segnali positivinell’incremento del numero di pro-getti di riqualificazione sono l’au-mento della disponibilità finanzia-ria, degli studi di base, del migliora-mento dei processi di selezione deitratti di fiume e dell’ampiezza degliobiettivi e delle misure.
Segnali negativi sono, tra glialtri, che spesso le misure sono le-gate solo all’idrologia fluviale e allestrutture interne al fiume. Il baci-no idrografico non viene conside-
rato. Inoltre il collo di bottiglia èspesso rappresentato dall’economiae dagli accordi con la popolazionee/o con le organizzazioni. Final-mente è stato compiuto il primopasso verso un sistema di supportoalle decisioni per la riqualificazio-ne fluviale. Il sistema presentatofornisce solo informazioni basatesu quali decisioni dovrebbero esse-re adottate. “Dove e come” è neces-sario prendere queste decisioni ri-mane una sfida per il futuro.
[ARM]
RINATURAZIONE E RIQUALIFICAZIONE AMBIENTALE > SPECIE ESOTICHE
Exotic plant species as problems and solutions in ecological restoration: a synthesisC. D’Antonio, L.A. MeyersonRestoration Ecology, 10: 703-713 (2002)C. D’Antonio, Department of Integrative Biology, University of California, Berkeley, CA 94720-3140, U.S.A. Present address: Exotic andInvasive Weeds Research Unit, USDA-ARS, 920 Valley Road, Reno, Nevada 89512. E-mail: [email protected].
Le specie esotiche sono pro-gressivamente diventate un proble-ma gestionale rilevante in parchi eriserve e frequentemente creanocomplicazioni nei progetti di recu-pero. Allo stesso tempo, ci possonoessere circostanze nelle quali la lororimozione può avere conseguenzenegative impreviste, oppure nellequali è desiderabile il loro utilizzonella riqualificazione. Nell’artico-lo sono presentati i possibili effettiprodotti dalle specie esotiche, im-portanti durante le fasi di recupe-ro; viene inoltre proposto un filonedi ricerca che potrebbe incrementa-re la capacità di inviduare obiettivi
di gestione realistici. Poiché il con-tollo e l’utilizzo delle specie esoti-che possono essere oggetto di con-troversie, gli autori raccomandanodi considerare queste specie nel con-testo più ampio della struttura edella successione della comunità;inoltre, essi evidenziano gli ambitiin cui la ricerca ecologica potrebbeportare a sviscerare i problemi cheriguardano le specie esotiche ed ilripristino ambientale. Per esempio,per definire le priorità di rimozionee gli obiettivi realistici di gestione ènecessaria una comprensione dellapotenziale transitorietà delle specieesotiche in un sito e del ruolo che
particolari specie esotiche potreb-bero giocare nel modificare i pro-cessi che influenzano l’evoluzionedella successione ecologica. Allo stes-so modo, una maggiore compren-sione del ruolo ecologico delle spe-cie introdotte potrebbe aiutare aridurre le controversie circa il loroutilizzo nella riqualificazione. Nel-l’articolo, le generalizzazioni cheemergono dalla letteratura sull’eco-logia delle specie invasive sono mes-se in connessione con gli interessipratici del recupero, incluse le cir-costanze in cui risulta pratico usa-re specie esotiche nella riqualifica-zione ambientale. [ARM]
INTEGRITÀ ECOLOGICA > SPECIE INDICATRICI
Selecting indicator species to monitor ecological integrity: a reviewV. Carignan, M.-A. VillardEnvironmental Monitoring and Assessment, 78: 45-61 (2002)M.-A. Villard, Département de biologie, Université de Moncton, Moncton, New Brunswick, Canada. E-mail: [email protected]
Gli autori passano in rasse-gna le caratteristiche principali che
devono essere considerate al mo-mento della scelta di specie indica-
trici all’interno di un programmadi monitoraggio che abbia l’obietti-

Biol. Amb., 18 (n. 2, 2004) Rassegna scientifica 71
vo di mantenere o ripristinare l’in-tegrità ecologica. In primo luogosono esaminati i pro ed i contro deidifferenti approcci di gestione suiquali può essere impostato un pro-gramma di conservazione, giungen-do alla conclusione che una gestio-ne basata sugli ecosistemi sia lasoluzione più appropriata. Vengo-no quindi individuati i potenzialiindicatori di integrità ecologica a
diversi livelli dell’ecosistema, conparticolare enfasi sulle specie.
Gli autori concludono che,sebbene l’utilizzo di specie indica-trici resti un argomento controver-so, esso può mostrarsi utile qualora(1) nel programma di monitoraggiosiano incluse numerose specie rap-presentanti di vari taxa e differenticicli vitali , (2) la loro selezione siabasata su una solida base di dati
quantitativi della zona di interesse,e (3) si usi cautela nell’interpreta-zione dei trend di popolazione perdistinguere i reali segnali dalle va-riazioni che possono non esserecorrelate al deterioramento dell’in-tegrità ecologica. Infine, sono pre-sentati e discussi i differenti metodiche sono stati utilizzati per selezio-nare le specie indicatrici.
[PG]
TUTELA AMBIENTALE > ACQUE INTERNE
Community participation in conserving and managing inland watersW.D. WilliamsAquatic Conservation: Marine and Freshwater Ecosystems, 12: 315-326 (2002)W.D. Williams, Department of Environmental Biology, Adelaide University, South Australia 5005.
Molte organizzazioni interna-zionali e nazionali hanno enfatiz-zato la necessità della partecipazio-ne della comunità nella conserva-zione e nella gestione delle acqueinterne. La partecipazione dellacomunità è necessaria per tre ra-gioni principali: per attuare le mi-sure di gestione che sarebbero didifficile applicazione senza il sup-porto della comunità stessa; per pro-teggere le acque interne attraversoil supporto delle organizzazioni disalvaguardia ambientale; per mo-nitorare, recuperare e riqualificarei corpi idrici interni, attraversol’azione dei volontari.
È importante conservare e ge-stire le acque interne per il lorovalore ed i loro molteplici utilizzi.Per partecipare appieno alle misu-
re di conservazione e gestione, lacomunità ha bisogno di (a) ricono-scere l’importanza delle acque in-terne come parte del ciclo idrogeo-logico globale, (b) avere una certaconoscenza della natura e degli ef-fetti dei maggiori impatti antropicie (c) essere a conoscenza di alcunequestioni legali.
La comunità è eterogenea, masi possono riconoscere gruppi coninteressi simili. Essi variano da pic-coli gruppi di azione locali a orga-nizzazioni internazionali, passan-do attraverso gruppi nazionali. Essiprovvedono a diffondere e suppor-tare le azioni governative; altre or-ganizzazioni si oppongono e tenta-no di modificare le azioni del go-verno. Il coinvolgimento della co-munità può avvenire a vari livelli,
da una relativa passività ad un vi-goroso attivismo.
L’educazione ambientale del-la comunità dovrebbe cominciaresin dall’infanzia, proseguire nellescuole e nella altre istituzioni edu-cative e continuare per tutta la vita.L’informazione sulla conservazio-ne e gestione delle acque interne èdisponibile attraverso molte fontima una potente fonte moderna è ilWorld Wide Web.
Nell’articolo sono discussiquattro casi di studio, con partico-lare riguardo alla partecipazionedella comunità: Lake Washingtone Mono Lake negli Stati Uniti (ri-sultati positivi), il Mare di Aral inAsia centrale e il Lake Pedder inAustralia (risultati negativi).
[ARM]

Recensioni Biol. Amb., 18 (n. 2, 2004)72
Informazione&Documentazione
RECENSIONIRoberto Messori, Luciano Tosi
Gli insetti di Fly Line. Effi-mere, Tricotteri e Plecotteri
Edizioni Fly Line ecosistemi fluvia-li, Modena, 2003, pp. 416, euro 75,00(per i soci CISBA sconto del 25%)
Forse un biologo evoluzioni-sta parlerebbe di convergenza adat-tativa. Questo libro dimostra comel’interesse per gli insetti acquaticipossa accomunare due categorieapparentemente lontane tra loro, ibiologi ambientali ed i pescatoricon la mosca artificiale (Pam).
I Pam adottano uno stile par-ticolarmente elaborato di pesca, uti-lizzando come esche imitazioni diinsetti di cui i pesci si nutrono. Leimitazioni, fatte di piume, fili, pelied altri materiali assemblati sul-l’amo, vengono lanciate con oppor-tuna perizia dove si trova il pesce,il quale, se si farà ingannare inmaniera convincente, abboccherà.
In un saggio di quasi vent’an-ni fa dedicato alla sociobiologia(La sociobiologia, Editori Riuniti,1986), Vittorio Parisi faceva riferi-mento ai Pam per descrivere unesempio di trasmissione sociale,entro e tra le generazioni, per vianon genetica. L’apprendimento cul-turale di questa tecnica ha portato,nei secoli, alla creazione di svaria-te decine di mosche artificiali, ognu-na con un proprio nome e con unamorfologia ben precisa e codifica-ta. Una sorta di “evoluzione artifi-ciale”, con curiose analogie conquella naturale, ha fatto sì che leesche somigliassero sempre più ailoro modelli, fedelmente imitati nel-le loro caratteristiche morfologichee comportamentali.
Ecco, dunque, il significato diquesto libro per i Pam: conoscereda vicino gli insetti “veri”, i modellioriginali che rendono possibile larealizzazione del proprio desiderio,catturare pesci in quantità graziead una sfida basata non sulla forzafisica, bensì su un gioco di intuizio-ni. Per questo gli autori hanno ope-rato la scelta –felice– di un’impo-stazione corretta dal punto di vistascientifico, supportata dai maggioriesperti italiani dei tre ordini di in-setti: Fernanda Cianficconi (Uni-versità di Perugia) per i Tricotteri,Carlo Belfiore (Università di Napo-li Federico II) per gli Efemerotteri eRomolo Fochetti (Università dellaTuscia di Viterbo) per i Plecotteri.
Per quanto riguarda la strut-tura del volume –un’elegante pub-blicazione rilegata in carta patina-ta– dopo una breve introduzione,gli autori dedicano alcune paginealle generalità sugli Insetti e quindisi avventurano in tre ricchi capitoli,ciascuno dedicato ad un ordine.
Degli Efemerotteri vengonoriassunte le caratteristiche generalie presentate due chiavi di identifi-cazione delle forme immaginali alivello di famiglia, oltre alla siste-matica aggiornata (2003) delle spe-cie italiane. Ad ogni famiglia è de-dicato un paragrafo contenente ul-teriori chiavi di determinazione deigeneri (forme immaginali), per cia-scuno dei quali sono descritte lespecie presenti in Italia e le loroprincipali caratteristiche (morfolo-gia, distribuzione geografica, ecolo-gia). L’ultima parte del capitolo èdedicata alle ninfe acquatiche perle quali si riporta una chiave didiscriminazione delle famiglie eduna breve descrizione delle stesse.
Il capitolo dedicato ai Tricot-
teri, dei tre l’ordine più rilevanteper numero di specie (circa 410censite in Italia), è suddiviso inuna parte generale –che affronta letematiche di morfologia (una parteè dedicata alle ali), l’etologia e lasistematica (sono riportate le chia-vi per la determinazione delle fami-glie e dei generi delle forme imma-ginali)– e in una parte speciale, incui vengono descritte in manieradettagliata le 17 famiglie, i relativigeneri e le specie più comuni chevivono nel nostro Paese.
Infine, una settantina di pagi-ne è dedicata ai Plecotteri, con notesulla morfologia di larve e adulti,sull’etologia, i suggerimenti per laraccolta, nonché le chiavi di deter-minazione di famiglie e generi.Come per gli altri ordini, per cia-scuna famiglia sono descritti i ge-neri e le principali specie presentiin Italia.
Nel complesso, il volume ri-porta informazioni aggiornate cheriguardano oltre 320 specie dellafauna italiana. Un ulteriore puntodi forza è rappresentato dalle nu-
Per usufruire dello sconto previsto per i soci CISBA su alcuni volumi, chiedere alla segreteria ([email protected]) l’attestato da allegare all’ordine.

Biol. Amb., 18 (n. 2, 2004) Recensioni 73
Informazione&Documentazione
merosissime (480) fotografie inedi-te a colori ad alta definizione, siadelle forme adulte che di quellelarvali. Il materiale iconografico ècompletato da 250 disegni.
I biologi ambientali che, permotivi legati al biomonitoraggio,hanno familiarità con i macroin-vertebrati del benthos, potranno tro-vare in questo libro un’utile inte-grazione delle guide di identifica-zione tradizionalmente utilizzate,ma soprattutto l’occasione di allar-gare le proprie conoscenze entomo-logiche prendendo confidenza conle loro forme alate.
Sfogliando il libro viene dariflettere –per certi versi con preoc-cupazione– sul fatto che una pub-blicazione di buon livello come
questa provenga da un ambito nonpropriamente legato al mondo del-la ricerca scientifica. Senza dubbioil libro cerca di colmare un vuotodi conoscenza, degli invertebratiacquatici e delle relative forme ala-te, che si è creato in questi ultimianni anche a causa della mancan-za di un progetto organico che mo-bilitasse risorse a favore della ricer-ca di base ed applicata su questiorganismi. Oggi i tempi sarebberomaturi per l’avvio della revisione eper la pubblicazione aggiornatadella collana di guide della faunaacquatica del CNR, comparse a par-tire dalla fine degli anni ’70. L’ap-plicazione della direttiva europeasulle acque, che richiede una valu-tazione delle biocenosi acquatiche
in quanto elementi di qualità deicorpi idrici, non può infatti pre-scindere dalla disponibilità di unquadro di conoscenze valide ed at-tuali sulla nostra fauna.
Per concludere con una notadi ottimismo, è confortante pensareche la spinta che ha originato laconvergenza adattativa tra biologiambientali e Pam, di cui si parlavain apertura, non è tanto l’oggettodel libro fin qui discusso, quanto,in ultima analisi, un ambizioso so-gno comune: il recupero e la tuteladei nostri fiumi e, più in generale,di tutti i corpi idrici.
Per ordinazioni: Fly Line, c.p.30 - 41041 Casinalbo (MO), tel./fax059 573663 [email protected], www.flylinemagazine.com.
Pietro Genoni
Sergio Zerunian
Condannati all’estinzione?Biodiversità. Biologia, minac-ce e strategie di conservazionedei pesci d’acqua dolce indi-geni in Italia
Edagricole, Bologna, 2002 (220 pp.,E 49,50)
I Pesci d’acqua dolce costitui-scono un gruppo faunistico conmolte specie endemiche in Italia. Illibro documenta in modo puntualelo stato dell’ittiofauna indigena del-le acque dolci italiane e denuncia,con forza, ma anche con rigorescientifico, le cause che hanno pro-dotto la critica situazione attuale.
La situazione dei vertebratiitaliani desta non poche preoccu-pazioni, per il consistente numerodi specie minacciate di scomparsa.Applicando infatti i criteri dell’Unio-ne Mondiale per la Conservazione,su 494 specie di vertebrati presentinel nostro territorio, 338 (circa il68%) sono inserite in un’ipotesi di
Lista Rossa; di queste, 199 (circa il40%) sono considerate minacciatee classificate nelle categorie “in pe-ricolo critico”, “in pericolo” e “vul-nerabile”. Tra i diversi gruppi, quellodei pesci d’acqua dolce (Ciclostomie Osteitti) è nella condizione piùcritica: su 48 specie presenti, 41(circa 85%) sono inserite nella Li-sta Rossa e le specie minacciatesono il 64,6%.
Non è però un lavoro di soladenuncia: il libro contiene un’ac-curata descrizione della morfolo-gia, della biologia, dell’ecologia del-le specie e del loro habitat. I testisono corredati da tavole a coloridelle specie di eccellente qualità efedeltà curate da Titti de Ruosi.
Un intero capitolo, dedicatoalle strategie di conservazione, in-dica in modo preciso le strade dapercorrere per evitare perdite irre-versibili come le estinzioni. Un uti-lissimo strumento, quindi, non soloper i ricercatori, ma anche per leamministrazioni locali e gli Enticompetenti in materia di pesca e diripopolamento, che devono attuarele migliori forme di gestione del
patrimonio naturale costituito daipesci d’acqua dolce.
Condannati all’estinzione? ponequindi solide basi scientifiche perla gestione dell’ittiofauna d’acquadolce italiana ed è un fondamenta-le punto di riferimento per suggeri-re una corretta impostazione di con-creti programmi finalizzati alla con-servazione delle specie e delle co-munità ittiche indigene.
Paolo Turin

Recensioni Biol. Amb., 18 (n. 2, 2004)74
Informazione&Documentazione
Sergio Zerunian
Piano d’azione generale perla conservazione dei Pescid’acqua dolce italianiQuaderno di Conservazione dellaNatura n° 17, Ministero dell’Am-biente e della Tutela del Territorioed Istituto Nazionale per la FaunaSelvatica “A. Ghigi”, 2003 (123 pp.)
Questo lavoro riprende alcu-
ne parti del volume Condannati al-l’estinzione? Biodiversità, biologia,minacce e strategie di conservazionedei Pesci d’acqua dolce indigeni inItalia, già pubblicato dal medesimoautore nel 2002, e le sviluppa nelladirezione dei principi della conser-vazione della natura.
È il primo lavoro di sintesiche ha come oggetto la conserva-zione dell’ittiofauna d’acqua dolceitaliana e, in particolare, dei suoi
preziosi endemismi; costituisce per-ciò anche un riferimento bibliogra-fico fondamentale per questo tema.
Oltre alla trattazione di tuttele principali minacce antropicheper i pesci d’acqua dolce, sono in-dividuati 8 taxa considerati di mag-giore d’interesse conservazionistico(Lampreda padana, Storione cobi-ce, Trota macrostigma, Carpione delFibreno, Trota marmorata, Carpio-ne del Garda, Panzarolo, Ghiozzodi ruscello). Per ciascuno di essi èfornito un quadro aggiornato dellabiologia, dell’ecologia e degli inter-venti di conservazione fin qui even-tualmente realizzati. Sono infineindividuate azioni capaci di inver-tire la tendenza in atto e scongiura-re il pericolo di estinzione per lespecie maggiormente a rischio.
Il Quaderno può essere richie-sto, da parte di Enti pubblici, al-l’Istituto Nazionale per la FaunaSelvatica (Via Cà Fornacetta 9,
mia, habitat, biologia, areale italia-no, rapporti con l’uomo e conser-vazione. Di particolare pregio è laraffigurazione a colori delle specie,eseguita da Titti De Ruosi.
È riportato il quadro aggior-nato della sistematica dell’ittiofau-na italiana; particolarmente utilile chiavi dicotomiche che facilita-no la determinazione delle specie.Completa il testo un CD-rom (conil testo in italiano e in inglese) dipratica consultazione, utile ancheper applicazioni didattiche.
Il volume può essere richie-sto, da parte di Enti pubblici, al-l’Istituto Nazionale per la FaunaSelvatica (Via Cà Fornacetta 9,40064 Ozzano dell’Emilia – BO); abreve sarà anche possibile scaricar-lo in formato pdf dal sito http://w w w . m i n a m b i e n t e . i t / S i t o /settori_azione/scn/pubblicazioni/qcn.asp.
Può anche essere acquistato
in riedizione integrale (E 28,00) da:Grandi e Grandi Editori, Strada Pro-vinciale 14, 230 – Savignano sulPanaro (MO); fax 059.796202,[email protected].
Paolo Turin
Sergio Zerunian
Pesci delle acque interned’Italia
Quaderno di Conservazione dellaNatura n. 20, Ministero dell’Am-biente e della Tutela del Territorioed Istituto Nazionale per la FaunaSelvatica “A. Ghigi”, 2004 (257 pp.)
Il volume è la versione “qua-derno” dell’Iconografia dei Pescidelle acque interne d’Italia, di S.Zerunian e T. De Ruosi (2002, Mi-nistero dell’Ambiente e della Tute-la del Territorio ed Istituto Nazio-nale per la Fauna Selvatica “A.Ghigi”).
Tutte le 63 specie indigenenelle acque interne italiane (4 Ci-clostomi e 59 Pesci Ossei) sono trat-tate ordinatamente con una suddi-visione in paragrafi che facilita lalettura del testo: sistematica, geone-
40064 Ozzano dell’Emilia – BO),oppure potrà essere fra breve scari-cato, da tutti gli utenti, in formatopdf dal sito: http://www.minam-biente.it/Sito/settori_azione/scn/pubblicazioni/qcn.asp
Paolo Turin

Biol. Amb., 18 (n. 2, 2004) Recensioni 75
Informazione&Documentazione
Vezio Puccini, Elvira Tarsitano
Parassitologia urbanaCittà, animali e salute pubblica
Edagricole, Bologna (XIV+344 pag.,69 illustrazioni. E 44,50)
Nelle città attuali, la tenden-za è quella di costruire ambienti divita che tengano conto solo delleesigenze dell’uomo, senza prenderecoscienza che, accanto alla popola-zione umana, si trovano una vastapopolazione animale portata dal-l’uomo stesso (animali domestici)ed una che invade nicchie da luicreate artificialmente (animali si-nantropici: insetti, uccelli, topi, ecc.).La situazione di sovraffollamentoconduce tutte queste popolazioniad adattarsi, mutandone il compor-tamento e introducendo nelle cittàparassiti patogeni. Si registra così
Maurizio Borin
FitodepurazioneSoluzioni per il trattamentodei reflui con le pianteEdagricole, Bologna (VIII+198 pag.,85 illustrazioni. E 24,00)
La fitodepurazione può esse-re considerata a pieno titolo unavera e propria “eco-tecnologia”d’avanguardia per il trattamentodei reflui, con basso fabbisogno tec-nologico ed energetico.
Già conosciuta ed applicataall’estero, è ancora poco diffusa inItalia dove manca soprattutto unabuona conoscenza delle prestazio-ni, tale da supportare efficacemen-te la corretta progettazione e gestio-ne degli impianti.
Il volume presenta le applica-zioni più significative della fitode-purazione e fornisce un contributoa risolvere tale problema.
Il manuale raccoglie e orga-nizza i risultati di una ricerca mul-tidisciplinare, gran parte dei qualioriginali e ottenuti da una vastaattività condotta presso l’Universi-tà di Padova, fa il punto sui diversiaspetti della materia finora acqui-siti ed espone con taglio praticodiverse soluzioni idonee per am-bienti differenti, con un occhio diriguardo ai problemi legati alla scel-ta e alla gestione delle specie vege-tali, un aspetto veramente crucialeper un buon funzionamento del-l’impianto.
La facilità di consultazione ela snellezza della trattazione ren-dono il testo uno strumento essen-
ziale per tecnici, progettisti, deciso-ri pubblici e funzionari con compe-tenze ambientali.
un aumento delle patologie trasmes-se dagli animali di città o che sipossono contrarre durante i pic-nic, il campeggio e nelle gite ‘fuori-porta’. Questo utile manuale mettea fuoco le problematiche sanitariespecifiche e propone soluzioni permonitorarle e controllarle.
Il volume presenta il lavorodi specialisti di numerose discipli-ne che hanno individuato strategieprecise per il controllo sanitariodelle zoonosi parassitarie attraver-so piani di programmazione sani-taria e urbanistica, ma anche attra-verso un controllo ‘partecipato’ del-la comunità. Sono trattati l’inqui-namento, la sovrapposizione conmalattie degli animali e dell’uomo,la gestione di aree verdi o degrada-te, la gestione faunistica. Il volumeconsente di impostare interventi acarattere preventivo e di controllo erappresenta un punto di riferimen-
to per coloro che operano nel cam-po della salute pubblica e delladisinfestazione, siano essi sanitari,amministratori pubblici, tecnici oresponsabili di aziende.

Recensioni Biol. Amb., 18 (n. 2, 2004)76
Informazione&Documentazione
Francesco Petretti
Gestione della fauna
Il management delle popola-zioni animali negli ambientinaturali, agricoli e urbanizzatiEdagricole, Bologna (370 pag., 62illustrazioni. E 38,50)
L’Italia vanta ben 57.344 di-verse specie d’interesse faunistico,un patrimonio da salvaguardareche, come tutte le risorse, va ammi-nistrato saggiamente. Ma gestirequesto fondamentale patrimonio ècosa da veri professionisti ed esper-ti, un vero e proprio “mestiere” dovel’improvvisazione è bandita. Il ma-nuale, curato da uno dei maggioriesperti del settore, è un’opera com-pleta che, associando agli aspettididattici quelli più pratici e di mag-gior interesse, fornisce un validoaiuto ai professionisti della gestio-
ne faunistica. Tutti gli argomentiaffrontati nel testo si riconduconoad un profilo professionale emer-gente e per il quale vi è un’interes-sante domanda: il tecnico della fau-na. Ad esso sono dedicate numero-se parti del volume e un capitoloche spiegano come avvicinarsi allaprofessione e come rapportarsi conil mondo del lavoro, avendo comemissione la gestione della fauna inuna prospettiva attiva di conserva-zione e gestione dell’ambiente na-turale. Il volume, infatti, nato dallelezioni tenute all’Università di Ca-merino per la Prima Scuola di spe-cializzazione in gestione dell’am-biente naturale e delle aree protet-te, avvicina concretamente i tecnicidella fauna alle problematiche rea-li. L’ampiezza, la profondità e lachiarezza espositiva lo rendono in-teressante anche per un pubblicopiù vasto, sensibile alle tematichedel rapporto uomo-natura.
L’autore, biologo, collabora
Marco Paci
Ecologia forestaleElementi di conoscenza dei si-stemi forestali
Edagricole, Bologna (310 pag., 161illustrazioni. E 39,50)
Le foreste non sono un pozzodi San Patrizio da cui estrarre risor-se a tempo indeterminato, bensì eco-sistemi, il cui eccessivo sfruttamentoha creato squilibri su scala planeta-ria. L’ecologia forestale nasce pro-prio dalla necessità di capirne ilfunzionamento, per attuarne unagestione ecologicamente sostenibile.
Questa seconda edizione, com-pletamente aggiornata, fornisce leconoscenze su struttura e funzio-namento dei sistemi forestali e aiu-ta a porre le fondamenta per un
lo ecologico svolto dall’uomo nellevarie epoche, apre questo lavoroscientifico che insegna a difenderea denti stretti la biodiversità, fonda-mentale per la collettività umana.
corretto approccio gestionale. Inquesto senso, la conoscenza deimeccanismi che consentono all’al-bero di approvvigionarsi di acquadal suolo o di fissare il carbonioatmosferico, rappresenta solo laparte di un percorso gerarchico chedall’individuo porta alla comunitàvivente, all’ecosistema e al paesag-gio. Insomma, un insieme di ecosi-stemi interagenti fra loro: foreste,campi coltivati, incolti, pascoli, fiu-mi, laghi e centri abitati.
Il libro tratta con particolareattenzione la componente arboreadegli ecosistemi forestali e rappre-senta una base conoscitiva impre-scindibile per chi, mediante la sel-vicoltura, dovrà intervenire su strut-tura e dinamismo dei boschi. Unapiacevole premessa storica sui rap-porti uomo-bosco, in cui si affron-tano le problematiche legate al ruo-
con le trasmissioni televisive Quarke Geo & Geo in qualità di esperto.Docente all’Università di Cameri-no, autore di numerosi documenta-ri e di libri di argomento naturalisti-co, è stato direttore dei programmiper la biodiversità del WWF Italia.

Biol. Amb., 18 (n. 2, 2004) Recensioni 77
Informazione&Documentazione
Henri Tachet, Philippe Ri-choux, Michel Bournaud, Phi-lippe Usseglio-Polatera
Invertébrés d’eau douce. Sy-stematique, biologie, écologie
CNRS Editions (www.cnrseditions.fr), Paris, 2003 (2000), 587 pag., E.51,30
Negli ultimi decenni del seco-lo scorso si è potuto osservare, siain Europa che in Italia, il progres-sivo e profondo mutamento a cui èandato incontro l’approccio allostudio e alla conservazione degliambienti fluviali. Anche il legisla-tore, per proprio conto, ha iniziatoa comprendere l’importanza di ve-rificare lo stato di qualità delle ac-que superficiali sulla base delle ri-sposte fornite dai loro primi fruito-ri: le comunità acquatiche.
La diffusione di nuovi metodidi indagine basati sullo studio de-gli indicatori, prevalentemente rap-presentati dai macroinvertebratibentonici per i corsi d’acqua cor-rente, ha richiesto l’adozione diguide pratiche che consentisserouna introduzione all’argomento,immediata e disponibile anche peri meno esperti. In tale scenario il“Tachet”, come veniva chiamato ilfamoso compendio dalla moltitudi-ne di biologi ambientali dedita allostudio dei corsi d’acqua, ha senzadubbio rivestito un ruolo di prima-ria importanza per la semplicità diutilizzo con la quale è stato conce-pito.
A distanza di venti anni dallasua prima edizione, il “Tachet” èstato rinnovato, ampliato e rivistoin tutte le sue parti. Il manualepratico ed immediato che per anniha costituito uno strumento indi-spensabile alla conoscenza dellecomunità di macroinvertebrati del-le acque dolci è stato, in effetti,completamente riscritto, fino ad as-
sumere le vesti di un vero e propriotrattato a carattere scientifico e di-vulgativo.
L’edizione 2003, in lingua ori-ginale, comprende un corpo cen-trale costituito da diciannove capi-toli dedicati alla classificazione dialtrettanti gruppi animali, un’am-pia introduzione (oltre 40 pagine)ed un capitolo di chiusura dedica-to all’approccio ecologico (circa 30pagine).
Oltre alla parte in cui si af-fronta la classificazione degli orga-nismi, ciascun capitolo è articolatoin vari paragrafi che collocano dalpunto di vista sistematico ciascungruppo considerato, ne descrivonola morfologia e l’anatomia, gliaspetti legati alla biologia e al-l’ecologia, riportano infine le tecni-che di identificazione, nonché labibliografia di riferimento. Per faci-litare la classificazione, ciascun ta-xon viene descritto nei suoi trattiessenziali e illustrato, con disegnidal tratto semplice ed essenziale,sin nei particolari anatomici dimaggior interesse per la determina-zione sistematica, com’è nello stiledegli autori. Le figure d’insieme chene scaturiscono, precedono e intro-ducono la classica chiave dicoto-mica che consente di giungere, disolito, fino a livello di famiglia e digenere, in alcuni casi fino a quellodi specie.
La chiave dicotomica, contra-riamente a quanto avveniva nellavecchia edizione, è stata estesa atutti i gruppi trattati, ed è correda-ta, nei vari percorsi, da una serie didisegni schematici di grande detta-glio che guidano agilmente il letto-re alla determinazione sistematicadei taxa studiati.
Nella convinzione che l’iden-tificazione di un organismo, sebbe-ne sia un preliminare fondamenta-le, perde di interesse se non con-temporaneamente affiancata dalsignificato ecologico che questi as-
sume, gli autori hanno pensato dielaborare una tabella a caratterinumerici, riprodotta per ogni grup-po sistematico considerato e ripor-tata a fine diagnosi, che consentedi delineare i tratti essenziali, rela-tivamente a biologia, fisiologia edecologia, di ogni taxon studiato.Partendo dalle Spugne per giunge-re agli Insetti è stato individuatoun numero massimo di 22 trattiper ciascun organismo considera-to, comprendenti, in totale, 119 dif-ferenti modalità di comportamen-to.
È nella parte introduttiva deltesto che viene illustrato, sin neiminimi particolari, il metodo de-scrittivo ideato dagli autori, da cuiscaturisce la tabella dei tratti com-portamentali di ogni singolo taxon.Ed è ancora nella parte introdutti-va che il lettore può trovare altrenotizie utili sugli aspetti legati al-l’ambito spaziale dei corsi d’acqua,dalla regione al microhabitat, e aquello più ecologico dei macroin-vertebrati. Anche in questa partedel volume non si lesinano grafici,schemi e disegni per meglio illu-strarne i contenuti.
E ancora, nel capitolo dedica-to all’approccio ecologico, l’ultimo,

Recensioni Biol. Amb., 18 (n. 2, 2004)78
Informazione&Documentazione
sono illustrate, seppure in manierasintetica, le possibilità offerte dal-l’uso della tabella dei tratti com-portamentali di ciascun gruppo, conuna serie di esempi applicativi nel-l’indagine sperimentale.
L’opera del Tachet si collocaad un livello di interesse che coin-volge sia lo specialista, che puòtrarre da essa informazioni sul va-
sto panorama dei macroinvertebra-ti, che lo studioso o l’idrobiologonon specializzati, che si avvicina-no allo studio dei corsi d’acqua,trovando in esso un valido stru-mento di lavoro. Ma come si leggeper mano di Albert Louis Roux,nella prefazione, uno dei maggioriobbiettivi dell’opera è quello di giun-gere ad un livello tassonomico che
riveli le informazioni ecologiche trale più pertinenti per comprendere ivari aspetti dell’ambiente in cui vi-vono i macroinvertebrati, con par-ticolare riguardo agli aspetti dellaqualità delle acque. (Per acquistarloin Italia: Libreria della natura, C.soMagenta, 48 – 20123 Milano, tel 0248003159; ai soci CISBA sconto 8%).
Gilberto N. Baldaccini
Corrado Battisti
Frammentazione ambien-tale, connettività, reti eco-logiche. Un contributo teori-co e metodologico con partico-lare riferimento alla fauna sel-vatica.
Provincia di Roma, Assessorato allepolitiche agricole, ambientali e pro-tezione civile, 2004, 248 pag.
La frammentazione degli am-bienti naturali, ovvero il processodi origine antropica attraverso ilquale un ambiente naturale vienesuddiviso in frammenti più o menoisolati, costituisce una delle più gra-vi minacce alla diversità biologica.
La risposta a tale problemaviene cercata in genere nella crea-zione di “reti ecologiche”, che favo-riscano la connessione tra diversiframmenti di ambienti naturali. Apartire dalle Direttive 1979/409/CE(Uccelli) e 1992/43/CE (Habitat), lanecessità di costituire reti ecologi-che è stata recepita anche in diver-si atti normativi e di pianificazio-ne, a livello europeo, nazionale eregionale.
Purtroppo però, il concetto direte ecologica ha assunto negli annisignificati assai diversi tra loro. Unprimo significato è quello di siste-ma interconnesso di unità ecosistemi-che, che contribuisca in modo si-
gnificativo a conservare la biodi-versità. Una seconda lettura del con-cetto, molto in uso nelle ammini-strazioni pubbliche, vede la rete eco-logica come un insieme integrato diaree protette, inserite in un sistemacoordinato di infrastrutture e servi-zi. Un terzo significato infine, uti-lizzato da chi segue un approcciourbanistico-territoriale, è quello disistema integrato di aree su cui realiz-zare azioni di conservazione e di valo-rizzazione delle risorse naturali e cul-turali, per la promozione dello svi-luppo socio economico locale.
I problemi causati da questadiversità di interpretazioni sonoaggravati dal fatto che, come riferi-sce Battisti, l’enfasi assegnata alsettore della pianificazione delle retiecologiche non corrisponde ad unaanaloga conoscenza dei problemidi conservazione da parte di chiredige i piani, anche a causa di uncoinvolgimento spesso marginaledelle professionalità naturalistiche,le sole capaci di interpretare edanalizzare i fattori e i processi eco-logici.
L’Assessorato alle politicheagricole, ambientali e protezionecivile della Provincia di Roma haquindi ritenuto opportuno pubbli-care un volume che affronta il temadella frammentazione ambientalee della connettività. Lo scopo del-l’iniziativa, come dice l’assessoreFiliberto Zaratti nella prefazioneal libro, è di “fornire strumenti co-
noscitivi e metodologici per avviareun percorso, condiviso con diverseprofessionalità, che investa la com-plessità dei sistemi territoriali, eco-logici, urbanistici e socio-economi-ci”.
Il testo è composto da tre par-ti. Nella prima viene descritto ilprocesso di frammentazione am-bientale e ne sono illustrate le con-seguenze a livello di popolazioni,comunità ed ecosistemi. Nella se-conda parte viene discusso il con-tributo che le aree protette possonodare alla conservazione della bio-diversità, nei paesaggi frammenta-ti. Sono inoltre illustrate le oppor-tunità offerte e le limitazioni deidiversi approcci utilizzati per ripri-stinare la connettività nelle aree

Biol. Amb., 18 (n. 2, 2004) Recensioni 79
Informazione&Documentazione
frammentate, considerando anchela necessità di adottare approccispecie-specifici. Nella terza parte,infine, viene illustrato un approc-cio alla individuazione di reti eco-logiche specie o gruppo specifiche,che può essere utilizzato da enti dipanificazione territoriale, come leprovince.
Molto ricco di esempi e di casidi studio tratti da una vasta lettera-tura, il volume pubblicato dallaProvincia di Roma offre numerositemi di riflessione per chi è impe-gnato, con diverse competenze di-sciplinari, nella pianificazione ter-ritoriale. È quindi auspicabile unasua ampia diffusione non solo tra i
biologi e i naturalisti, ma anche tragli architetti, gli urbanisti e gli in-gegneri che si occupano di territo-rio.
È infatti necessario che vengarivolta una specifica attenzione allaconservazione della connettivitàambientale, non solo nell’ambitodella pianificazione delle aree pro-tette. La vera partita, infatti, si gio-ca nell’ambito della valutazione diimpatto ambientale e, ancor più,nella valutazione ambientale stra-tegica istituita dalla direttiva Diret-tiva 2001/42/CE.
Infatti le decisioni più impor-tanti per evitare che proceda laframmentazione degli ambienti na-
turali sono prese proprio nell’am-bito dei piani e programmi riguar-danti i settori agricolo, forestale,della pesca, energetico, industriale,dei trasporti, della gestione dei ri-fiuti e delle acque, delle telecomu-nicazioni, turistico, della pianifica-zione territoriale e della destinazio-ne dei suoli, che costituiscono ilcampo di applicazione della Diret-tiva.
Il libro può essere richiesto a:Provincia di Roma, DipartimentoV, Servizio 1°, Via Tiburtina 691 -00159 Roma (Tel. 06 67663301, Fax06 43562126; [email protected]
Massimo Leone
RECENSIONI BREVII volumi di seguito segnalati possono essere ordinati per posta ordinaria a NHBS Mailorder Bookstore, 2-3 Wills Road, Totnes,Devon TQ9 5XN, United Kingdom, oppure all’indirizzo di posta elettronica nhbs@nhbs. co.uk. Il prezzo è espresso in sterline(GBP); prima dell’ordine, controllare le condizioni più aggiornate e ulteriori informazioni sui singoli volumi sul sito http://www. nhbs.com.
Heavily modified water bodies. Iden-tification and Designation IncludingCase Studies in EuropeEd. by E Kampa and W HansenSpringer, 2004, 320 pag. GBP 69, hard-cover.
Handbook of ecological indicatorsfor assessment of ecosystem healthEd. by SE Jorgensen, R Costanza and FXu.Comprehensive account of ecologicalindicators for evaluating the health ofa wide variety of ecosystems. It presentsa conceptual framework for selecting,evaluating and validating ecologicalindicators of ecosystem health and ap-plies this framework in a series of chap-ters on major ecosystem types, includ-ing coastal areas, forests, wetlands, fish-eries and agricultural land.Lewis, USA, 2004, 500 pages. GBP 85,hardcover.
Towards a theory of the functioningof aquatic ecosystemsA.F. Alimov
Backhuys, Netherlands, 2003, 130pages, 54 Figs, 23 tabs. GBP 45,softcover.
The functional assessment of wet-land ecosystemsEd. by E Maltby, U Digdy and C Baker.Woodhead, 2004, 520 pages. GBP238.53, CD.
Bioassessment of freshwater ecosys-tems. Using the Reference ConditionApproachRobert C Bailey, Richard H Norris andTrefor B Reynoldson.Aquatic ecosystem assessment is a rap-idly developing field, and one of thenewer approaches to assessing the con-dition of rivers and lakes is the Refer-ence Condition Approach. This is a sig-nificant advancement in biomonitoringbecause it solves the problem of tryingto locate nearby control or referencesites when studying an ecosystem thatmay be degraded. This book describesthe basic methods involved in select-ing and sampling appropriate refer-
ence sites, comparing test sites to ap-propriate reference sites using predic-tive modeling, and determiningwhether or not test sites are in the ref-erence condition. This provides a rapidassessment method that can deal witheverything from large-scale, nationalissues to local-scale problems with thesame approach, and often parts of thesame database.Kluwer, Netherlands, 2003, 184 pages.GBP 58.50, hardback.
Lake and reservoir managementEd. by SE Jorgensen, H Loffler, W Rastand M Straskraba.Elsevier, Netherlands, Developments inWater Science 54. 2004, 350 pp. GBP100, hardcover.
A practical guide to the managementof saline lagoonsEd. by N Symes and P Robertson.This is a practical handbook that isbased on the management experiencesat ten man made lagoon sites in theeast and south of England. The book

Recensioni brevi Biol. Amb., 18 (n. 2, 2004)80
Informazione&Documentazione
ing communication between research-ers, biologists, environmental engineersand others managing or studyingriverine environments. Summarisesstream classification, stream rehabili-tation and methods for determininginstream flow requirements. Includesthe most recent rapid developments inthe area of stream rehabilitation andthe setting of environmental flows. Anew Windows based set of revised pro-grammes of the computer packageAQUAPAK will be available.Wiley, 2004, 443 pages, figs, tabs. GBP32.50, softcover.
Assembly rules and restoration ecol-ogy. Bridging the gap between theoryand practiceEd. by Vicky M Temperton, Richard JHobbs, Tim Nuttle and Stefan Halle.Understanding how ecosystems are as-sembled - how the species that makeup a particular biological communityarrive in an area, survive, and interactwith other species - is key to success-fully restoring degraded ecosystems.Yet little attention has been paid to theidea of assembly rules in ecologicalrestoration, in both the scientific lit-erature and in on-the-ground restora-tion efforts. This book addresses thatshortcoming, offering an introduction,overview, and synthesis of the poten-tial role of assembly rules theory inrestoration ecology.Shearwater, USA, Science and Practiceof Ecological Restoration, 2004, 528pages, photos, figs, tabs. GBP 31.50,softcover.
Nature’s operating instructions. Thetrue biotechnologiesEd. by Kenny Ausubel and JP Harpi-gnies.Visionary leaders in fields such asbiomimicry (mimicking nature in orderto restore nature and serve human endsharmlessly), “living machines” thatbreak down toxics biologically, and therestoration of natural capital, the con-tributors’ guiding principles includediversity, kinship, symbiosis, reciproc-ity, and community. Sector by sector -from energy and agriculture to trans-portation, industrial production, andland management - the biotechnologiesdescribed here are examples of hownature has already created intelligent
design that we can emulate and adapt.California UP, USA, 2004, 288 pages.GBP 10.95, softcover.
Botanical latin. History, Grammar,Syntax, Terminology & VocabularyWT Stearn.First published in 1966, this book hasbecome accepted worldwide as thestandard work of reference on this im-portant subject. With an illustratedguide to descriptive terminology inboth English and Latin, plus an exten-sive vocabulary of terms taken fromcurrent botanical usage, this is the mostaccurate, up-to-date and comprehen-sive book on the subject ever produced,‘This remarkable work has evokedworldwide esteem and affection. Ency-clopedic and erudite, extraordinarilyinteresting and packed with detail.’The Journal of the Royal HorticulturalSociety.David & Charles, 2004, 546 pages, b/willustrations. GBP 19.99, paperback.
Towards a thermodynamic theory forecological systemsEd. by Sven E Jorgensen and YuriSvirezhev.Presents a consistent and complete eco-system theory based on thermodynamicconcepts. The introduction of the con-cept of exergy makes it possible to givea comprehensive explanation of ecosys-tem reactions and growth patterns. Thetrophic chain, global energy and radia-tion balance and pattern and the reac-tions of ecological networks are all ex-plained by the use of exergy.Pergamon Elsevier, 2004, 450 pp. GBP56.50, softcover.
Monitoring stream and watershedrestorationEd. by P Roni.Provides a comprehensive, practicalresource for developing monitoringand evaluation programmes for fresh-water habitat restoration at variousscales. Its unique emphasis is on howto monitor effects at habitat unit (site),reach and watershed scales. Numerousexamples are given, particularly of res-toration activities for coolwater biotaand salmonid fishes. However, theprinciples discussed are relevant world-wide.CABI, 2004, 300 pp. GBP 40, softcover.
provides guidance in the techniquesof both restoration management andmaintenance management of salinelagoons. It emphasises the need foradequate survey, monitoring and man-agement planning and describes suit-able techniques. The book is aimed ateveryone involved in saline lagoonmanagement, including site managers,reserve wardens, rangers, contractorsas well as those advising in its manage-ment. Useful techniques are illustratedusing case studies.RSPB, 2003, 90 pages. GBP 24, soft-cover.
Insect diversity conservationMichael J Samways.A contemporary global synthesis of therapidly developing and important fieldof insect conservation biology. Insectsplay important roles in terrestrial eco-logical processes and in maintainingthe world as we know it. They presentparticular conservation challenges, es-pecially as a quarter face extinctionwithin the next few decades. This text-book addresses the ethical foundationof insect conservation, and asks why weshould concern ourselves with it. Afteraddressing threats, from invasive al-ien plants to climate change, the bookexplores ways insects and their habi-tats are prioritised, mapped, monitoredand conserved. Landscape and speciesapproaches are considered.Cambridge UP, 2004, 250 pages, 120illus, 100 figs. GBP 30, softcover.
Managing priority habitats for inver-tebratesProvides detailed information on thehabitat requirements of 18 inverte-brate groups. The material is also ar-ranged into 32 UK priority habitats.2004, GBP 54.05 (CD).
Geomorphology and river manage-ment. Application of the river stylesframeworkG Brierley and K Fryirs.Blackwell, February 2005, 368 pp. GBP39.95, softcover.
Stream hydrology. An introduction forecologistsND Gordon, TA McMahon, BL Finlay-son, CJ Gippel and RJ Nathan.Provides a common language for improv-

BIOLBIOLBIOLBIOLBIOLOGIAOGIAOGIAOGIAOGIAAMBIENTAMBIENTAMBIENTAMBIENTAMBIENTALEALEALEALEALE ISTRUZIONI PER GLI AUTORIISTRUZIONI PER GLI AUTORIISTRUZIONI PER GLI AUTORIISTRUZIONI PER GLI AUTORIISTRUZIONI PER GLI AUTORI
Manoscritti. I lavori proposti per la pubblicazione, compatibilmentecon il loro contenuto, devono essere suddivisi in: introduzione, mate-riali e metodi, risultati, discussione, eventuali ringraziamenti,bibliografia, tabelle, figure. Qualora il lavoro sia già stato pubblicatoo sottoposto all’attenzione di altri editori, la circostanza deve esserechiaramente segnalata: in tal caso il lavoro potrà essere preso inconsiderazione solo per la recensione nella sezione Informazione &Documentazione.
Titolo e Autori. Il titolo deve essere informativo e, se possibile, conci-so; deve essere indicato anche un titolo breve (massimo cinquantacaratteri) da utilizzare come intestazione delle pagine successive allaprima. Il titolo deve essere seguito dal nome (per esteso) e dal cogno-me di tutti gli autori. Di ogni autore (contrassegnato da un richiamonumerico) deve essere riportato l’indirizzo postale completo dell’isti-tuto nel quale è stato svolto lo studio. Il nome dell’autore referenteper la corrispondenza con la redazione e con i lettori deve esserecontrassegnato anche da un asterisco; il suo indirizzo di posta ordina-ria deve essere seguito anche dal numero di telefono, di fax e dall’in-dirizzo di posta elettronica; soltanto tramite quest’ultimo verrannoinviate le bozze per la correzione.
Riassunto, abstract e parole chiave. Il riassunto (lunghezza massi-ma 250 parole) deve sintetizzare lo scopo dello studio, descrivere gliesperimenti, i principali risultati e le conclusioni; deve essere seguitodalle parole chiave, separate da una barra obliqua. Devono esserealtresì riportati in lingua inglese il titolo e un abstract (massimo 250parole), seguiti dalle key words separate da una barra obliqua.
Figure e tabelle. Le figure, con la loro didascalia al piede e numeratecon numeri arabi, non devono essere inserite nel testo, ma in fogliseparati alla fine del testo. È gradita l’indicazione, nel testo, dellaposizione preferita per l’inserzione di ciascuna figura. Anche le tabel-le devono essere riportate in fogli separati, alla fine del dattiloscritto;devono essere complete di titolo e numerate con numeri romani.Occorre curare titoli, legende e didascalie in modo da rendere letabelle e le figure autosufficienti, comprensibili cioè anche senzaconsultare il testo. Per le figure (grafici, disegni o fotografie di buonaqualità), si raccomanda agli autori di verificare con opportune riduzio-ni l’aspetto finale e la leggibilità delle scritte, tenendo conto chesaranno stampate riducendone la base a 80 mm (una colonna) o 170mm (due colonne). Non inviare fotografie o grafici a colori senzaessersi accertati che la loro stampa in bianco e nero assicuri comunquel’agevole riconoscibilità delle diverse sfumature o retinature. Nellascelta degli accorgimenti grafici privilegiare sempre la facilità e imme-diatezza di lettura agli effetti estetici.
Bibliografia. Al termine del testo deve essere riportata la bibliografiain ordine alfabetico. Ad ogni voce riportata nella bibliografia devenecessariamente corrispondere il riferimento nel testo e viceversa.Per il formato tipografico e la punteggiatura, attenersi strettamente aiseguenti esempi:DUTTON I.M., SAENGER P., PERRY T., LUKER G., WORBOYS G.L., 1994. An
integrated approach to management of coastal aquatic resources. Acase study from Jervis Bay, Australia. Aquatic Conservation: marineand freshwater ecosystems, 4: 57-73.
HELLAWELL J.M., 1986. Biological indicators of freshwater pollution andenvironmental management. Elsevier Applied Science Publishers,London and New York, 546 pp.
PULLIAM H.R., 1996. Sources and sinks: empirical evidence and popu-lation consequences. In: Rhodes O.E., Chesser R.K., Smith M.H.
(eds.), Population dynamics in ecological space and time. The Uni-versity of Chicago Press, Chicago: 45-69.
CORBETTA F., PIRONE G., (1986-1987) 1988. I fiumi d’Abruzzo: aspettidella vegetazione. In: Atti Conv. Scient. “I corsi d’acqua minoridell’Italia appenninica. Aspetti ecologici e gestionali”, Aulla (MS),22-24 giugno 1987. Boll. Mus. St. Nat. Lunigiana 6-7: 95-98.
Proposte di pubblicazione. Il manoscritto su supporto cartaceo deveessere inviate a:Redazione di Biologia Ambientale,c/o Giuseppe Sansoni, V.le XX Settembre 148 - 54033 Carrara (MS)Il manoscritto deve essere accompagnato da una copia su supportomagnetico; in alternativa, quest’ultima può essere inviata all’indiriz-zo di posta elettronica [email protected] manoscritti saranno sottoposti alla lettura di revisori scientifici; entrodue mesi l’autore indicato come referente per la corrispondenza verràinformato delle decisioni della redazione. Per evitare ritardi nellapubblicazione e ripetute revisioni del testo, si raccomanda vivamenteagli autori di prestare la massima cura anche alla forma espositiva chedeve essere concisa, chiara, scorrevole e in buon italiano, evitandoneologismi superflui. Tutte le abbreviazioni e gli acronimi devonoessere definiti per esteso alla loro prima occorrenza nel testo. I nomiscientifici delle specie devono essere sottolineati (saranno convertiti incorsivo prima della stampa). I dattiloscritti, compreso il materialeillustrativo, non saranno restituiti, salvo esplicita richiesta dell’autoreall’atto dell’invio del materiale. La redazione si riserva il diritto diapportare ritocchi linguistici e grafici e di respingere i manoscritti chenon rispettano i requisiti delle presenti norme per gli autori. Leopinioni espresse dagli autori negli articoli firmati non rispecchianonecessariamente le posizioni del C.I.S.B.A.
Bozze ed estratti. Le bozze di stampa saranno inviate all’autoreindicato come referente per la corrispondenza, che deve impegnarsiad una correzione molto accurata e al nuovo invio alla redazione entro5 giorni; trascorso tale periodo, il lavoro può essere pubblicato con lesole correzioni dell’editore. All’autore referente per la corrisponden-za sarà inviato il numero della rivista e, tramite posta elettronica, ilfile dell’estratto in formato PDF, utilizzabile per riprodurre il numerodesiderato di estratti.
Formato dei file. Oltre al manoscritto vanno inviati, su supportomagnetico, i relativi file. Per assicurare la compatibilità con i program-mi di videoscrittura e di impaginazione, il file contenente il testo vainviato in triplice versione: formato solo testo (*.TXT), rich text format(*.RTF) e WinWord (*.DOC, preferibilmente salvato nel formato dellasua penultima versione commerciale). I grafici devono essere in bian-co e nero ed essere sempre accompagnati dalla tabella dei dati diorigine; per quelli realizzati con fogli elettronici inviare il file conte-nente sia i grafici che i dati di origine (preferibilmente salvato nellapenultima versione commerciale di Excel) al fine di consentirne ilridimensionamento o eventuali modifiche al formato, volte a miglio-rarne la leggibilità. I file delle figure al tratto vanno inviati preferibil-mente in formato *.TIF; quelli delle fotografie preferibilmente informato *.JPG. Per formati di file diversi da quelli sopra indicati,precisare il software utilizzato. Importante: i grafici e le illustrazioniinseriti in un file DOC come utile corredo alla lettura del manoscritto,non sono sufficienti per la realizzazione tipografica (comportano unaperdita di nitidezza e difficoltà in fase di impaginazione); è perciònecessario inviare sempre anche i grafici e le figure come file indi-pendenti. Per ogni chiarimento tecnico contattare Giuseppe Sansoni(tel./fax 0585 841592, e-mail [email protected] ).

BIOLBIOLBIOLBIOLBIOLOGIAOGIAOGIAOGIAOGIAAMBIENTAMBIENTAMBIENTAMBIENTAMBIENTALEALEALEALEALE
Spediz. in abbon. post, art. 2, comma 20/c, L. 662/96, filiale Reggio EmiliaTassa pagata - Taxe perçue
Volume 18Numero 2Dicembre 2004
Info
rmaz
ione
&do
cum
enta
zion
e
SOMMARIO
RECENSIONI
Lavo
ri or
igin
ali
RASSEGNASCIENTIFICA
3 FERRARINI A., ROSSI P., MIGLIAZZI M., ROSSI O.,GIAVELLI G. - L’analisi da remoto del paesaggiomediante finestre mobili: il caso di studio della ValBaganza (Parma)
11 SFRISO A., FACCA C., CEOLDO S., GHETTI P.F. -Variazioni delle concentrazioni di nutrienti e deiflussi di sedimentazione nella parte centrale dellalaguna di Venezia
19 FACCA C., SFRISO A., GHETTI P.F. - Abbondanzae diversità del fitoplancton e delle diatomeebentoniche in laguna di Venezia
25 ZERUNIAN S. - Proposta di un Indice dello StatoEcologico delle Comunità Ittiche viventi nelle ac-que interne italiane
31 GAITER S., BODON M., ROCCA D. - Applicazionedella metodica di indagine biologica, basata sullafauna bentonica e sulla componente organica delsedimento, in acque di sorgente ed emergenzeassimilabili
48 PICCININI A. - Il Siluro tra mito e realtà
53 Rassegna scientifica (a cura di P. Genoni)
72 Recensioni





























