Embed Size (px)
Citation preview

doi 101098rsif20110154 1665-1672 first published online 13 May 20118 2011 J R Soc Interface
Steve Alpern Robbert Fokkink Marco Timmer and Jeacuterocircme Casas predator search for preyAmbush frequency should increase over time during optimal
Referenceshttprsifroyalsocietypublishingorgcontent8641665fullhtmlref-list-1
This article cites 32 articles 7 of which can be accessed free
Email alerting service hereright-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top
httprsifroyalsocietypublishingorgsubscriptions go to J R Soc InterfaceTo subscribe to
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
J R Soc Interface (2011) 8 1665ndash1672
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Author for c
doi101098rsif20110154Published online 13 May 2011
Received 19 MAccepted 8 A
Ambush frequency should increaseover time during optimal predator
search for preySteve Alpern1 Robbert Fokkink2 Marco Timmer2
and Jerome Casas3
1Department of Mathematics London School of Economics London WC2A 2AE UK2Faculty of Electrical Engineering Mathematics and Information Technology Delft
University PO Box 5031 2600 GA Delft The Netherlands3Institut de Recherches sur la Biologie de lrsquoInsecte University of Tours UMR CNRS 6035
37200 Tours France
We advance and apply the mathematical theory of search games to model the problem facedby a predator searching for prey Two search modes are available ambush and cruisingsearch Some species can adopt either mode with their choice at a given time traditionallyexplained in terms of varying habitat and physiological conditions We present an additionalexplanation of the observed predator alternation between these search modes which is basedon the dynamical nature of the search game they are playing the possibility of ambushdecreases the propensity of the prey to frequently change locations and thereby renders itmore susceptible to the systematic cruising search portion of the strategy This heuristic expla-nation is supported by showing that in a new idealized search game where the predator isallowed to ambush or search at any time and the prey can change locations at intermittenttimes optimal predator play requires an alternation (or mixture) over time of ambush andcruise search Thus our game is an extension of the well-studied lsquoPrincess and Monsterrsquosearch game Search games are zero sum games where the pay-off is the capture time andneither the Searcher nor the Hider knows the location of the other We are able to determinethe optimal mixture of the search modes when the predator uses a mixture which is constantover time and also to determine how the mode mixture changes over time when dynamicstrategies are allowed (the ambush probability increases over time) In particular we estab-lish the lsquosquare root law of search predationrsquo the optimal proportion of active search equalsthe square root of the fraction of the region that has not yet been explored
Keywords predatorndashprey search game ambush foraging behaviour
1 INTRODUCTION
When searching for a potentially mobile prey a preda-tor can adopt a behaviour lying between two extrememodes (i) active or cruising search seeking to maxi-mize the area searched in a given time period or (ii)sit-and-wait or ambush search remaining still in sucha way so as to maximize the probability of detectingany motion of the prey The earlier literature was syn-thesized and abstracted in the study of Schoener [1]which analyses the predator behaviour of these twotypes Since then considerable literature describesspecies that alternate between the two search modesin various mixtures The traditional explanation ofthe alternation of strategies is as a response to chang-ing external (eg weather) or internal (eg hunger)parameters observable by the predator The thesis ofthis article is that there is an alternative explana-tion of the observed alternating behaviour that holds
orrespondence (salpernlseacuk)
arch 2011pril 2011 1665
even in a constant internal and external environmentthe possibility of an ambush decreases the propensity ofthe prey to frequently change locations thereby makingit more susceptible to detection through systematicsearch (cruising)
We come to this and other more precise conclusionsthrough the analysis of a new Ambush Search Gamethat extends the well-known lsquoPrincess and Monsterrsquosearch game [2] by giving the Monster (the predator)an additional strategy at any time namely the possi-bility of adopting the ambush mode When in theambush mode he detects and captures the Princess(prey) if she moves but simply wastes time (coveringno new ground in search) if she stays still For thisreason the Princess will be stationary except for inter-mittent lsquodashesrsquo to new random locations With thismodel we are able to determine as a function of certainexogenous parameters the ideal (minimax search time)stationary mixture of cruising versus ambushing for thepredator In a fully dynamic version of the game we can
This journal is q 2011 The Royal Society
1666 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
further determine the ideal mixture (cruising prob-ability) for the predator to adopt when the prey hasbeen excluded by search from a fraction of the searchregion Our answer is what we call the Square RootLaw of Predation Adopt the cruising mode with prob-ability equal to the square root of the unexploredfraction of the search region This implies in particularthat when beginning the search the cruising modeshould predominate but when the prey location hasbeen narrowed to a small area the ambush modeshould predominate This discourages the prey frommoving to escape back into the full region
A related attempt to model the ambushndashcruisesearch mixture has been recently presented in Zoroaet al [3] in a more geometrical context On the preyside the question of when to move to a new location(though with more information than we allow) hasbeen studied by Broom amp Ruxton [4]
2 SEARCH GAMES WITH MOBILE HIDER
The game-theoretic background to the problem weconsider lies in the search game with mobile hider morecolourfully named by Isaacs [2] as the Princess andMonster Game This is a zero sum game between asingle predator (the Monster) and a single mobile prey(the Princess) with the capture time T as the pay-off(obviously the Monster is the minimizer) The twoplayers move about in a known search region Q untilthe first time T that the Princess comes within the Mon-sterrsquos detection radius If the search region is a networkthe game ends when the two players meet (detectionradius is zero) The Monster has a maximum speedtaken as 1 which is also the rate at which he exploresnew territory The mathematical theory of Princess andMonster Games was begun in Zeliken [5] and Alpern[6] where the search region was considered a circle andextended [7] to games on networks or regions of n-dimen-sional space Alternative approaches to the latterproblem were given by Lalley amp Robbins [8] and Garnaev[9] Further work on related search games was carried outby Kikuta amp Ruckle [10] Pavlovic [11] and Alpern et al[12] (see also the monographs [1314])
As far as we are aware the only biological appli-cation of Princess and Monster Games is [1516] inthe context of a contest between a wasp (Monster) ofthe genus Sympiesis (Hymenoptera Eulophidae) anda leafminer larvae (Princess) of Phyllonorycter blancar-della (Lepidoptera Gracillariidae) The search region Xis a roughly planar area between the two surfaces of anapple leaf where the feeding activity of the larvae pro-duces a so-called windows randomly placed translucentand whitish small spots in which only the upper cuti-cula remains while the plant tissue underneath iseaten up The wasp is attracted in flight by the visualappearance of the mines lands on the leaf and thusstarts the game Leaf vibrations cause the larvae tocease any feeding and become lsquoalertrsquo The wasp vio-lently inserts its ovipositor nearly exclusively in thefeeding windows as the rapid escape reactions ofthe larva command blazing fast attacks unfeasible ifthe wasp has to pierce thick tissues A successful
J R Soc Interface (2011)
search ends with the larva being parasitized and thewasp laying an egg on it Thus this is a discrete locationapproximation of the Princess and Monster Game
The search game that we introduce in this article addsan extra strategy for the Monster (predator) namely anambush mode that he may adopt at any time whichresults in immediate capture if the Princess is movingat that time The presence of this strategy keeps thePrincess still most of the time except for intermittentshort (actually instantaneous) dashes to new randomlocations On the other hand if the Princess is stationaryduring a time interval when the Monster is ambushingthe Monster effectively does no searching and accom-plishes nothing in this time interval The possibility ofambush behaviour in the Princess and Monster Gamewas first considered by Alpern amp Asic [1718] in thecontext of lsquobottlenecksrsquo in the search region where theMonster might do well to wait in ambush such as atthe central node of the lsquofigure eightrsquo network Ourinterpretation here is not that the Monster will wait ata particular point of the search region but rather thathe will hover or perch somewhere above it where he cansurvey the whole region (or a part of itmdashsee sect46)
We point out that there is a related literature onwhat are called ambush games where the mobile hider(prey) has to move between two known locations (ega known migration pattern) and the Searcher (ambush-ing predator) has to choose locations at which toambush (intercept) the mobile prey This problem ismore geometrical than ours in that the Searcher hasto pick locations rather than as in our model speedor cruisendashambush alternation The literature for suchambush games [19ndash22] may well have applicationsto the relationship between predator location andmigratory patterns of prey For example it is suggestedby Walter [23] that colonies of Eleonorarsquos falcon areplaced to intercept migrations
There is another literature on pursuit-evasion gameswithin the field of differential games (see Isaacs [2])which has applications to the next stage of thepredatorndashprey interaction what happens between thedetection of prey and capture of prey This is an impor-tant problem but not one that we address here Weassume that the predator wishes to minimize the timeto detect (find) the prey keeping our analysis firmlywithin the field of search games
3 RELATIONSHIP WITH THEBEHAVIOURAL AND ECOLOGICALLITERATURE
Nearly all of the many works done on the searching andescaping modes of predator and prey have been done onevolutionary and ecological timescales (see Helfman [24]and Cooper [25] for reviews and Michel amp Adams [26] forone of the latest studies)This ranges from studies focusingon the matching between the searching modes and thegrouprsquos phylogeny to studies highlighting the switch ofsearching modes according to internal state or externalconditions such as temperature obstacles and habitatcomplexity to prey densities In this context speed doesnot vary within a single interaction between a predator
Ambush frequency should increase over time S Alpern et al 1667
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
and a prey By contrast exceedingly few studies focus onthe ethological time scale at which search occurs andduring which motivational or environmental factors donot change much For the same reasons most studiesdo not address the problem of information acquisition bythe opponents during the search game A monographabout the ethology of predation [27] already reported alack of quantitative ethological studies on the geometricaland speed components of predatorndashprey interactions andlamented that many results were based on anecdotes Thisremains basically true as of today
A quantitative assessment of the predator speedduring the interaction has been however carried out ina few cases often related to pursuitndashevasion gamesIn the first case [2829] the bimodal attacking speed dis-tribution observed in wolf spiders is interpreted as anoptimization strategy trading speed on the one handand production of unwanted information used by preyfor escape on the other hand spiders in ambush orwalking slowly towards prey do not produce a largerecognizable aerodynamical signature so they mayachieve capture before revealing their presence to theprey spiders attacking at full speed do produce a lotof such information for the prey but their speed maysuffice to overtake even an escaping prey spiders run-ning at an intermediate speed produce sufficientinformation to be spotted and are not quick enoughto get their prey so these hunting speeds are thereforeavoided In that example speed is nearly constantduring a single pursuit The second case is a searchingstrategy the saltatory search This strategy consists ofmaking small hops or short dashes followed by shortpauses and is thus a mixture of speeds displayedduring a single interaction [3031] It is common infishes and birds predators look for prey when theypause and move bit-by-bit producing a kind ofpseudo-systematic pattern of search Pausing enablesthem also to attend to other tasks such as lookingout for top predators [30] Saltatory search in fisheshas been explained using hydrodynamical forcesacting against cruise search [32] Other functionalexplanations for this behaviour have been given relatedto information acquisition by the predator rather thanby the prey as in the invertebrate example Either theradius of perception is nearly similar to the length ofthe move in the case of fishes [32] or speed itself is ahandicap to sensory acquisition such as for stabilizingvisual images of potential prey in birds and lizards[33] These hypotheses are backed up by recent workshowing that speed decreases acuity [34ndash36] In sum-mary saltatory search lies in the middle of acontinuum between ambush and cruise search The pro-portion of either behaviour remains however constantduring the saltatory search in contrast to our modelVarying speed during an interaction when preybecome cornered or when prey are difficult to subduehas been observed [27]
4 THE AMBUSH SEARCH GAME
This section describes and analyses our modification ofthe Princess and Monster Game which we call the
J R Soc Interface (2011)
Ambush Search Game The Hider (modelling theprey or the Princess in the version of Isaacs [2] andconsidered female) is stationary at a random locationof a search region X of measure (or area) 1 except forintermittent times of her choosing when she can relo-cate (in a quick dash which we take to beinstantaneous) to a new random location The Searcher(modelling the predator or the Monster in Isaacs [2]and considered male) can explore X at a maximumspeed (or discovery rate) of 1 This means that if theSearcher never ambushes he can search the wholeregion in unit time In this case if the Hider does notmove she will be found in an expected time of 12 Inany time interval the Searcher may ambush (inwhich case the Searcher will certainly detect amoving Hider but not do any exploring) or he mayadopt active cruising search (in which case he willexplore a maximum amount of new territory but willnot catch a moving Hider) We assume that when theHider successfully moves (to a new random location)this event is detected by an active Searcher (perhapsthrough the vibration of a leaf in the leafminer case)but the new location of the Hider cannot be ascertainedThus the information available to the Searcher is thesame as at the start of the game when he arrived atthe patch namely that the Hider is uniformly distribu-ted over the search space X Hence we say that in thisevent the original game is repeated Note that when wesay that the Hiderrsquos movement is detected by theSearcher we do not mean that the Hider is capturedonly that the Searcher knows that the Hider is nolonger contained in the smaller region in which he wasknown to be located prior to the move Of course ifthe move occurs during a period of ambush the gameends
A pure search strategy for the Searcher isfully described by a function s(t) with s(0) frac14 0 and0 s0(t) 1 where s(t) is the fraction of X that hehas searched up to time t assuming the Hider has notmoved In so-called saltatory or alternating searchperiods of ambush are characterized by derivativess0(t) frac14 0 and periods of cruising by s0(t) frac14 1 We alsoallow intermediate speeds between 0 and 1 for whichwe have two interpretations In the probabilisticinterpretation we assume that the Searcher maydivide an interval containing time t into many tinytime intervals on each of which he cruises with prob-ability s0(t) and ambushes with probability 1 2 s0(t)So if the Hider moves the Searcher will be in anambush interval with probability 1 2 s0(t) in whichcase she will be caught The other interpretationwhich we shall adopt henceforth is that s0(t) is simplythe speed at which the Searcher is moving throughthe search region (and also his rate of exploring newterritory) In this interpretation we assume that hisability to detect prey decreases linearly with hisspeed so that when moving at speed s0(t) he detectsa prey movement with probability 1 2 s0(t) Theassumption that visual acuity (or other measures oflsquoaccuracyrsquo) is decreasing in predator speed is wellfounded in neuroethological terms in diverse situationssuch as pollination or crypsis [36]
01 02 03 04 05 06 07 08 09 100
10
20
a
m
Figure 1 Optimal Hider moving time methaTHORN (Online version incolour)
05 06 07 08 09 10
08
09
10
a
T
Figure 2 Worst case capture time TethaTHORN (Online version incolour)
1668 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To evaluate the efficiency of a particular search strat-egy s(t) we suppose that the Hider plans to move to anew random location at some time t frac14 m 0 assumingshe is not found by the Searcher before then What isthe expected capture timeT frac14TethsmTHORN in this situationThis timeT satisfies the following equation
T frac14TethsmTHORN
frac14ethm
0t s0ethtTHORN dt thorn eth1 sethmTHORNTHORNethm thorn s0ethmTHORN TTHORN eth41THORN
The first term corresponds to the case where theHider is caught by a moving Searcher before time mbecause the probability of her being found in a smalltime interval [tt thorn Dt] is approximately s0(t) Dt Thesecond term corresponds to the case where the Hiderhas not been found by time m which occurs with prob-ability 1 2 s(m) In this case the capture time is atleast m for sure Furthermore the Hider is successfulin her move at time m with probability s0(m) inwhich case the search game is played again so she getsan additional expected time of T
Our aim is to find the (minimax) search functionsethtTHORN [ C 2 which gives the least expected capture timeT for the worst (highest) case of the Hiderrsquos choice ofm that is
maxm
TethsmTHORN frac14 mins[S
maxm
TethsmTHORN
We expect (and show later) that against the optimalsearch function s the expected capture time will be thesame for all Hider moving times m We first analyse thecase where the Searcher is restricted to a constantsearch speed a (that is s(t) is of the form at) and findthe optimal speed a in this case However the expectedcapture time with this restriction always depends on mso there is no minimax strategy of constant speedThen we analyse quadratic search functions and findan optimal solution
There is an additional technical question (which thereader can disregard for the moment) as to what hap-pens if the Hider is always relocating which weinterpret as moving at time m frac14 0 In this limitingcase the Hiderrsquos expected lifetime T is 0 unless theSearcher is putting all his effort into active search sothat s0(0) frac14 1 and there is no probability of ambushIn this limiting case we have
Teths 0THORN frac14 limm0
TethsmTHORN frac14 11 s00eth0THORN
if s0eth0THORN frac14 1eth42THORN
Since the second derivative arises in this context werequire that our search strategies s belong to the space Sof C2 functions functions with a continuous secondderivative which we now take as the predator purestrategy space
41 Restricted optimization for constant-speedpredators
Suppose we restrict the Searcher to searching with a con-stant speed a 1 so that at time t he has searched a
J R Soc Interface (2011)
fraction of the search space X given by s(t) frac14 at Asusual this assumes that t m that the Hider has notyet moved Substituting at for s(t) in equation (41) wehave an expected capture timeT frac14TethamTHORN satisfying
T frac14 etham 1THORNethmetha 1THORN aethm thornTTHORNTHORN thorn 12
am2
or T frac14 2m am2
2a thorn 2a2 m thorn 2
eth43THORN
For fixed speed a the Hider (prey) maximizesthe capture time T by moving after timemethaTHORN frac14 etha thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffia2 thorn 1p
1THORN=a2 as drawn in figure 1Note that against a full-speed predator the preyshould always be moving so as to remain at a randomlocationmdashthere is no downside to moving causedby ambush Thus the worst case capture timeTetha methaTHORNTHORN when the Searcher moves at speed a isTethaTHORN frac14 eth
ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
thorn a2 1THORN=etha3ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
THORN with mini-mum at a frac14
ffiffiffi3p
=2 ≃ 087 of TethaTHORN frac14 4ffiffiffi3p
=9 ≃ 077 asdrawn in figure 2
Summarizing our results from this section we havethe following
Proposition 41 If the Searcher (predator) isrestricted to moving at a constant speed a (interpretedas a constant mixture over time of ambush and cruisesearch) his best speed is a frac14
ffiffiffi3p
=2 ≃ 087 which guar-antees an expected capture time not exceeding4ffiffiffi3p
=9 ≃ 077 against any moving time m of the Hider
The interpretation of this result is that a predatorwho cannot keep track of how long it has been searchingshould employ active search about 87 per cent of thetime and ambush about 13 per cent of the time Ofcourse this result assumes that an ambushing Searcheralways detects a moving prey regardless of the preyrsquoslocation A more realistic version of this result without
02 04 06 08 10 12 14 16 18 200
02
04
06
08
10
t
s
Figure 3 Optimal fraction sethtTHORN of space searched by time t (Online version in colour)
Ambush frequency should increase over time S Alpern et al 1669
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
certain detection of moving prey by an ambushingpredator is given in sect46
42 An indifference-inducing search strategy s
In this section we determine a particular search strategysethtTHORN for which the Searcher is indifferent to the time mwhen the Hider moves This is a common heuristic for sol-ving zero sum games but is not a sufficient condition foroptimality We show in sect43 that this strategy is indeedthe pure minimax strategy It turns out that the Searchercan reduce the worst case expected capture time from the07698 he could ensure using constant speed down to23 frac14 06666 if he uses a quadratic function ratherthan a linear one Since s(0) frac14 0 (at time 0 he hassearched none of the space) there is no constant termso we assume that s(t) has the form
sethtTHORN frac14 ct2 thorn bt eth44THORN
for unknown constants c and b Substituting this quadra-tic form of s into the expected capture time formula (41)gives
T frac14 16
m2eth3bthorn 4cmTHORN ethethbthorn 2cmTHORNethm thornTTHORN
methbthorn 2cm 1THORNTHORNethcm2 thorn bm 1THORN
or 0 frac14 23
c ceth2cT thorn 1THORN
m3
thorn 12
b beth2cT thorn 1THORN cbT
m2
thorn eth2cT Tb2 thorn 1THORNm thorn ethbT TTHORN
Solving for b c and T so that the coefficients ofpowers of m are all zero (independence of m) gives theoptimal search strategy sethtTHORN frac14 eth1=4THORNt2 thorn t as shownin figure 3
The method of deriving the formula for sethtTHORN givenhere avoids the need for differential equations and inany case the required constancy TethsethtTHORNmTHORN frac14 2=3 canbe verified directly from equation (41) Our model pre-dicts a slowing down of predator speed
43 Proof that s is optimal
We now show that the pure search strategysethtTHORN frac14 t2=4thorn t is the optimal (minimax) search func-tion Since TethsmTHORN does not depend on m it is enough
J R Soc Interface (2011)
to show that for any other search function s frac14 s(t)there is some m such that TethsmTHORN TethsmTHORN To thisend we need the following simple result We adoptthe notation t frac14 ts to denote the time taken for a strat-egy s(t) to explore all of the region X (eg ts frac14 2)
Proposition 42 For any search strategy s [ Ss = seither
1 s0(0) frac14 1 and s00(0) 12 or2 sethtTHORN sethtTHORN for all t in some interval (01) In this
case either (i) ts 2 or (ii) there is a least timet1 0 with setht1THORN setht1THORN and s0etht1THORN frac14 s0etht1THORN
Proof Since s0(0) 1 for all s [ S if condition 1fails then the first sentence of condition 2 must holdIf part 2(i) fails so that ts 2 then by the Intermedi-ate Value Theorem there is a time t2 such thatsetht2THORN frac14 setht2THORN Then since seth0THORN frac14 seth0THORN frac14 0 it followsfrom the Mean Value Theorem that there is a timet1 0 such that s0etht1THORN frac14 s0etht1THORN Since the set of sucht1 is compact there is a least such time B
Figure 4 illustrates five curves from top to bottom ta type 1 curve t 2 t28 the minimax curve s the type2(ii) curve t23 with 2t13 frac14 1 2 t12 or t1 frac14 67 frac14086 the type 2(i) curve t25 with ts 2
Proposition 43 The strategy sethtTHORN frac14 t t2=4 is theminimax pure Searcher strategy That is
mins[S
maxm
TethsmTHORN frac14 maxm
TethsmTHORN frac14 23
Proof We show that for each type of strategy s inproposition 42 we can determine a Hider move time msuch that TethsmTHORN 2=3 If s is of type 1 of proposition42 then Teths 0THORN frac14 1 eth1 s00eth0THORNTHORN 2=3 If s is of type2(i) of proposition 42 Teths 2THORN Teths 2THORN frac14 2=3 If sis type 2(ii) take m frac14 t1 where t1 is the time given byproposition 42 part 2(ii) For this value of m definefs(T ) to be the linear function given by the right sideof equation (41) We claim that the line fs(T ) has ahigher slope and is higher at T frac14 0 than fsT so itintersects the diagonal at a higher point Since T frac14T(sm) is the solution of T frac14 fs (T ) and T frac14 2=3 is thesolution of fsethTTHORN frac14 T we have TethsmTHORN T frac14 2=3
0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)

J R Soc Interface (2011) 8 1665ndash1672
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Author for c
doi101098rsif20110154Published online 13 May 2011
Received 19 MAccepted 8 A
Ambush frequency should increaseover time during optimal predator
search for preySteve Alpern1 Robbert Fokkink2 Marco Timmer2
and Jerome Casas3
1Department of Mathematics London School of Economics London WC2A 2AE UK2Faculty of Electrical Engineering Mathematics and Information Technology Delft
University PO Box 5031 2600 GA Delft The Netherlands3Institut de Recherches sur la Biologie de lrsquoInsecte University of Tours UMR CNRS 6035
37200 Tours France
We advance and apply the mathematical theory of search games to model the problem facedby a predator searching for prey Two search modes are available ambush and cruisingsearch Some species can adopt either mode with their choice at a given time traditionallyexplained in terms of varying habitat and physiological conditions We present an additionalexplanation of the observed predator alternation between these search modes which is basedon the dynamical nature of the search game they are playing the possibility of ambushdecreases the propensity of the prey to frequently change locations and thereby renders itmore susceptible to the systematic cruising search portion of the strategy This heuristic expla-nation is supported by showing that in a new idealized search game where the predator isallowed to ambush or search at any time and the prey can change locations at intermittenttimes optimal predator play requires an alternation (or mixture) over time of ambush andcruise search Thus our game is an extension of the well-studied lsquoPrincess and Monsterrsquosearch game Search games are zero sum games where the pay-off is the capture time andneither the Searcher nor the Hider knows the location of the other We are able to determinethe optimal mixture of the search modes when the predator uses a mixture which is constantover time and also to determine how the mode mixture changes over time when dynamicstrategies are allowed (the ambush probability increases over time) In particular we estab-lish the lsquosquare root law of search predationrsquo the optimal proportion of active search equalsthe square root of the fraction of the region that has not yet been explored
Keywords predatorndashprey search game ambush foraging behaviour
1 INTRODUCTION
When searching for a potentially mobile prey a preda-tor can adopt a behaviour lying between two extrememodes (i) active or cruising search seeking to maxi-mize the area searched in a given time period or (ii)sit-and-wait or ambush search remaining still in sucha way so as to maximize the probability of detectingany motion of the prey The earlier literature was syn-thesized and abstracted in the study of Schoener [1]which analyses the predator behaviour of these twotypes Since then considerable literature describesspecies that alternate between the two search modesin various mixtures The traditional explanation ofthe alternation of strategies is as a response to chang-ing external (eg weather) or internal (eg hunger)parameters observable by the predator The thesis ofthis article is that there is an alternative explana-tion of the observed alternating behaviour that holds
orrespondence (salpernlseacuk)
arch 2011pril 2011 1665
even in a constant internal and external environmentthe possibility of an ambush decreases the propensity ofthe prey to frequently change locations thereby makingit more susceptible to detection through systematicsearch (cruising)
We come to this and other more precise conclusionsthrough the analysis of a new Ambush Search Gamethat extends the well-known lsquoPrincess and Monsterrsquosearch game [2] by giving the Monster (the predator)an additional strategy at any time namely the possi-bility of adopting the ambush mode When in theambush mode he detects and captures the Princess(prey) if she moves but simply wastes time (coveringno new ground in search) if she stays still For thisreason the Princess will be stationary except for inter-mittent lsquodashesrsquo to new random locations With thismodel we are able to determine as a function of certainexogenous parameters the ideal (minimax search time)stationary mixture of cruising versus ambushing for thepredator In a fully dynamic version of the game we can
This journal is q 2011 The Royal Society
1666 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
further determine the ideal mixture (cruising prob-ability) for the predator to adopt when the prey hasbeen excluded by search from a fraction of the searchregion Our answer is what we call the Square RootLaw of Predation Adopt the cruising mode with prob-ability equal to the square root of the unexploredfraction of the search region This implies in particularthat when beginning the search the cruising modeshould predominate but when the prey location hasbeen narrowed to a small area the ambush modeshould predominate This discourages the prey frommoving to escape back into the full region
A related attempt to model the ambushndashcruisesearch mixture has been recently presented in Zoroaet al [3] in a more geometrical context On the preyside the question of when to move to a new location(though with more information than we allow) hasbeen studied by Broom amp Ruxton [4]
2 SEARCH GAMES WITH MOBILE HIDER
The game-theoretic background to the problem weconsider lies in the search game with mobile hider morecolourfully named by Isaacs [2] as the Princess andMonster Game This is a zero sum game between asingle predator (the Monster) and a single mobile prey(the Princess) with the capture time T as the pay-off(obviously the Monster is the minimizer) The twoplayers move about in a known search region Q untilthe first time T that the Princess comes within the Mon-sterrsquos detection radius If the search region is a networkthe game ends when the two players meet (detectionradius is zero) The Monster has a maximum speedtaken as 1 which is also the rate at which he exploresnew territory The mathematical theory of Princess andMonster Games was begun in Zeliken [5] and Alpern[6] where the search region was considered a circle andextended [7] to games on networks or regions of n-dimen-sional space Alternative approaches to the latterproblem were given by Lalley amp Robbins [8] and Garnaev[9] Further work on related search games was carried outby Kikuta amp Ruckle [10] Pavlovic [11] and Alpern et al[12] (see also the monographs [1314])
As far as we are aware the only biological appli-cation of Princess and Monster Games is [1516] inthe context of a contest between a wasp (Monster) ofthe genus Sympiesis (Hymenoptera Eulophidae) anda leafminer larvae (Princess) of Phyllonorycter blancar-della (Lepidoptera Gracillariidae) The search region Xis a roughly planar area between the two surfaces of anapple leaf where the feeding activity of the larvae pro-duces a so-called windows randomly placed translucentand whitish small spots in which only the upper cuti-cula remains while the plant tissue underneath iseaten up The wasp is attracted in flight by the visualappearance of the mines lands on the leaf and thusstarts the game Leaf vibrations cause the larvae tocease any feeding and become lsquoalertrsquo The wasp vio-lently inserts its ovipositor nearly exclusively in thefeeding windows as the rapid escape reactions ofthe larva command blazing fast attacks unfeasible ifthe wasp has to pierce thick tissues A successful
J R Soc Interface (2011)
search ends with the larva being parasitized and thewasp laying an egg on it Thus this is a discrete locationapproximation of the Princess and Monster Game
The search game that we introduce in this article addsan extra strategy for the Monster (predator) namely anambush mode that he may adopt at any time whichresults in immediate capture if the Princess is movingat that time The presence of this strategy keeps thePrincess still most of the time except for intermittentshort (actually instantaneous) dashes to new randomlocations On the other hand if the Princess is stationaryduring a time interval when the Monster is ambushingthe Monster effectively does no searching and accom-plishes nothing in this time interval The possibility ofambush behaviour in the Princess and Monster Gamewas first considered by Alpern amp Asic [1718] in thecontext of lsquobottlenecksrsquo in the search region where theMonster might do well to wait in ambush such as atthe central node of the lsquofigure eightrsquo network Ourinterpretation here is not that the Monster will wait ata particular point of the search region but rather thathe will hover or perch somewhere above it where he cansurvey the whole region (or a part of itmdashsee sect46)
We point out that there is a related literature onwhat are called ambush games where the mobile hider(prey) has to move between two known locations (ega known migration pattern) and the Searcher (ambush-ing predator) has to choose locations at which toambush (intercept) the mobile prey This problem ismore geometrical than ours in that the Searcher hasto pick locations rather than as in our model speedor cruisendashambush alternation The literature for suchambush games [19ndash22] may well have applicationsto the relationship between predator location andmigratory patterns of prey For example it is suggestedby Walter [23] that colonies of Eleonorarsquos falcon areplaced to intercept migrations
There is another literature on pursuit-evasion gameswithin the field of differential games (see Isaacs [2])which has applications to the next stage of thepredatorndashprey interaction what happens between thedetection of prey and capture of prey This is an impor-tant problem but not one that we address here Weassume that the predator wishes to minimize the timeto detect (find) the prey keeping our analysis firmlywithin the field of search games
3 RELATIONSHIP WITH THEBEHAVIOURAL AND ECOLOGICALLITERATURE
Nearly all of the many works done on the searching andescaping modes of predator and prey have been done onevolutionary and ecological timescales (see Helfman [24]and Cooper [25] for reviews and Michel amp Adams [26] forone of the latest studies)This ranges from studies focusingon the matching between the searching modes and thegrouprsquos phylogeny to studies highlighting the switch ofsearching modes according to internal state or externalconditions such as temperature obstacles and habitatcomplexity to prey densities In this context speed doesnot vary within a single interaction between a predator
Ambush frequency should increase over time S Alpern et al 1667
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
and a prey By contrast exceedingly few studies focus onthe ethological time scale at which search occurs andduring which motivational or environmental factors donot change much For the same reasons most studiesdo not address the problem of information acquisition bythe opponents during the search game A monographabout the ethology of predation [27] already reported alack of quantitative ethological studies on the geometricaland speed components of predatorndashprey interactions andlamented that many results were based on anecdotes Thisremains basically true as of today
A quantitative assessment of the predator speedduring the interaction has been however carried out ina few cases often related to pursuitndashevasion gamesIn the first case [2829] the bimodal attacking speed dis-tribution observed in wolf spiders is interpreted as anoptimization strategy trading speed on the one handand production of unwanted information used by preyfor escape on the other hand spiders in ambush orwalking slowly towards prey do not produce a largerecognizable aerodynamical signature so they mayachieve capture before revealing their presence to theprey spiders attacking at full speed do produce a lotof such information for the prey but their speed maysuffice to overtake even an escaping prey spiders run-ning at an intermediate speed produce sufficientinformation to be spotted and are not quick enoughto get their prey so these hunting speeds are thereforeavoided In that example speed is nearly constantduring a single pursuit The second case is a searchingstrategy the saltatory search This strategy consists ofmaking small hops or short dashes followed by shortpauses and is thus a mixture of speeds displayedduring a single interaction [3031] It is common infishes and birds predators look for prey when theypause and move bit-by-bit producing a kind ofpseudo-systematic pattern of search Pausing enablesthem also to attend to other tasks such as lookingout for top predators [30] Saltatory search in fisheshas been explained using hydrodynamical forcesacting against cruise search [32] Other functionalexplanations for this behaviour have been given relatedto information acquisition by the predator rather thanby the prey as in the invertebrate example Either theradius of perception is nearly similar to the length ofthe move in the case of fishes [32] or speed itself is ahandicap to sensory acquisition such as for stabilizingvisual images of potential prey in birds and lizards[33] These hypotheses are backed up by recent workshowing that speed decreases acuity [34ndash36] In sum-mary saltatory search lies in the middle of acontinuum between ambush and cruise search The pro-portion of either behaviour remains however constantduring the saltatory search in contrast to our modelVarying speed during an interaction when preybecome cornered or when prey are difficult to subduehas been observed [27]
4 THE AMBUSH SEARCH GAME
This section describes and analyses our modification ofthe Princess and Monster Game which we call the
J R Soc Interface (2011)
Ambush Search Game The Hider (modelling theprey or the Princess in the version of Isaacs [2] andconsidered female) is stationary at a random locationof a search region X of measure (or area) 1 except forintermittent times of her choosing when she can relo-cate (in a quick dash which we take to beinstantaneous) to a new random location The Searcher(modelling the predator or the Monster in Isaacs [2]and considered male) can explore X at a maximumspeed (or discovery rate) of 1 This means that if theSearcher never ambushes he can search the wholeregion in unit time In this case if the Hider does notmove she will be found in an expected time of 12 Inany time interval the Searcher may ambush (inwhich case the Searcher will certainly detect amoving Hider but not do any exploring) or he mayadopt active cruising search (in which case he willexplore a maximum amount of new territory but willnot catch a moving Hider) We assume that when theHider successfully moves (to a new random location)this event is detected by an active Searcher (perhapsthrough the vibration of a leaf in the leafminer case)but the new location of the Hider cannot be ascertainedThus the information available to the Searcher is thesame as at the start of the game when he arrived atthe patch namely that the Hider is uniformly distribu-ted over the search space X Hence we say that in thisevent the original game is repeated Note that when wesay that the Hiderrsquos movement is detected by theSearcher we do not mean that the Hider is capturedonly that the Searcher knows that the Hider is nolonger contained in the smaller region in which he wasknown to be located prior to the move Of course ifthe move occurs during a period of ambush the gameends
A pure search strategy for the Searcher isfully described by a function s(t) with s(0) frac14 0 and0 s0(t) 1 where s(t) is the fraction of X that hehas searched up to time t assuming the Hider has notmoved In so-called saltatory or alternating searchperiods of ambush are characterized by derivativess0(t) frac14 0 and periods of cruising by s0(t) frac14 1 We alsoallow intermediate speeds between 0 and 1 for whichwe have two interpretations In the probabilisticinterpretation we assume that the Searcher maydivide an interval containing time t into many tinytime intervals on each of which he cruises with prob-ability s0(t) and ambushes with probability 1 2 s0(t)So if the Hider moves the Searcher will be in anambush interval with probability 1 2 s0(t) in whichcase she will be caught The other interpretationwhich we shall adopt henceforth is that s0(t) is simplythe speed at which the Searcher is moving throughthe search region (and also his rate of exploring newterritory) In this interpretation we assume that hisability to detect prey decreases linearly with hisspeed so that when moving at speed s0(t) he detectsa prey movement with probability 1 2 s0(t) Theassumption that visual acuity (or other measures oflsquoaccuracyrsquo) is decreasing in predator speed is wellfounded in neuroethological terms in diverse situationssuch as pollination or crypsis [36]
01 02 03 04 05 06 07 08 09 100
10
20
a
m
Figure 1 Optimal Hider moving time methaTHORN (Online version incolour)
05 06 07 08 09 10
08
09
10
a
T
Figure 2 Worst case capture time TethaTHORN (Online version incolour)
1668 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To evaluate the efficiency of a particular search strat-egy s(t) we suppose that the Hider plans to move to anew random location at some time t frac14 m 0 assumingshe is not found by the Searcher before then What isthe expected capture timeT frac14TethsmTHORN in this situationThis timeT satisfies the following equation
T frac14TethsmTHORN
frac14ethm
0t s0ethtTHORN dt thorn eth1 sethmTHORNTHORNethm thorn s0ethmTHORN TTHORN eth41THORN
The first term corresponds to the case where theHider is caught by a moving Searcher before time mbecause the probability of her being found in a smalltime interval [tt thorn Dt] is approximately s0(t) Dt Thesecond term corresponds to the case where the Hiderhas not been found by time m which occurs with prob-ability 1 2 s(m) In this case the capture time is atleast m for sure Furthermore the Hider is successfulin her move at time m with probability s0(m) inwhich case the search game is played again so she getsan additional expected time of T
Our aim is to find the (minimax) search functionsethtTHORN [ C 2 which gives the least expected capture timeT for the worst (highest) case of the Hiderrsquos choice ofm that is
maxm
TethsmTHORN frac14 mins[S
maxm
TethsmTHORN
We expect (and show later) that against the optimalsearch function s the expected capture time will be thesame for all Hider moving times m We first analyse thecase where the Searcher is restricted to a constantsearch speed a (that is s(t) is of the form at) and findthe optimal speed a in this case However the expectedcapture time with this restriction always depends on mso there is no minimax strategy of constant speedThen we analyse quadratic search functions and findan optimal solution
There is an additional technical question (which thereader can disregard for the moment) as to what hap-pens if the Hider is always relocating which weinterpret as moving at time m frac14 0 In this limitingcase the Hiderrsquos expected lifetime T is 0 unless theSearcher is putting all his effort into active search sothat s0(0) frac14 1 and there is no probability of ambushIn this limiting case we have
Teths 0THORN frac14 limm0
TethsmTHORN frac14 11 s00eth0THORN
if s0eth0THORN frac14 1eth42THORN
Since the second derivative arises in this context werequire that our search strategies s belong to the space Sof C2 functions functions with a continuous secondderivative which we now take as the predator purestrategy space
41 Restricted optimization for constant-speedpredators
Suppose we restrict the Searcher to searching with a con-stant speed a 1 so that at time t he has searched a
J R Soc Interface (2011)
fraction of the search space X given by s(t) frac14 at Asusual this assumes that t m that the Hider has notyet moved Substituting at for s(t) in equation (41) wehave an expected capture timeT frac14TethamTHORN satisfying
T frac14 etham 1THORNethmetha 1THORN aethm thornTTHORNTHORN thorn 12
am2
or T frac14 2m am2
2a thorn 2a2 m thorn 2
eth43THORN
For fixed speed a the Hider (prey) maximizesthe capture time T by moving after timemethaTHORN frac14 etha thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffia2 thorn 1p
1THORN=a2 as drawn in figure 1Note that against a full-speed predator the preyshould always be moving so as to remain at a randomlocationmdashthere is no downside to moving causedby ambush Thus the worst case capture timeTetha methaTHORNTHORN when the Searcher moves at speed a isTethaTHORN frac14 eth
ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
thorn a2 1THORN=etha3ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
THORN with mini-mum at a frac14
ffiffiffi3p
=2 ≃ 087 of TethaTHORN frac14 4ffiffiffi3p
=9 ≃ 077 asdrawn in figure 2
Summarizing our results from this section we havethe following
Proposition 41 If the Searcher (predator) isrestricted to moving at a constant speed a (interpretedas a constant mixture over time of ambush and cruisesearch) his best speed is a frac14
ffiffiffi3p
=2 ≃ 087 which guar-antees an expected capture time not exceeding4ffiffiffi3p
=9 ≃ 077 against any moving time m of the Hider
The interpretation of this result is that a predatorwho cannot keep track of how long it has been searchingshould employ active search about 87 per cent of thetime and ambush about 13 per cent of the time Ofcourse this result assumes that an ambushing Searcheralways detects a moving prey regardless of the preyrsquoslocation A more realistic version of this result without
02 04 06 08 10 12 14 16 18 200
02
04
06
08
10
t
s
Figure 3 Optimal fraction sethtTHORN of space searched by time t (Online version in colour)
Ambush frequency should increase over time S Alpern et al 1669
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
certain detection of moving prey by an ambushingpredator is given in sect46
42 An indifference-inducing search strategy s
In this section we determine a particular search strategysethtTHORN for which the Searcher is indifferent to the time mwhen the Hider moves This is a common heuristic for sol-ving zero sum games but is not a sufficient condition foroptimality We show in sect43 that this strategy is indeedthe pure minimax strategy It turns out that the Searchercan reduce the worst case expected capture time from the07698 he could ensure using constant speed down to23 frac14 06666 if he uses a quadratic function ratherthan a linear one Since s(0) frac14 0 (at time 0 he hassearched none of the space) there is no constant termso we assume that s(t) has the form
sethtTHORN frac14 ct2 thorn bt eth44THORN
for unknown constants c and b Substituting this quadra-tic form of s into the expected capture time formula (41)gives
T frac14 16
m2eth3bthorn 4cmTHORN ethethbthorn 2cmTHORNethm thornTTHORN
methbthorn 2cm 1THORNTHORNethcm2 thorn bm 1THORN
or 0 frac14 23
c ceth2cT thorn 1THORN
m3
thorn 12
b beth2cT thorn 1THORN cbT
m2
thorn eth2cT Tb2 thorn 1THORNm thorn ethbT TTHORN
Solving for b c and T so that the coefficients ofpowers of m are all zero (independence of m) gives theoptimal search strategy sethtTHORN frac14 eth1=4THORNt2 thorn t as shownin figure 3
The method of deriving the formula for sethtTHORN givenhere avoids the need for differential equations and inany case the required constancy TethsethtTHORNmTHORN frac14 2=3 canbe verified directly from equation (41) Our model pre-dicts a slowing down of predator speed
43 Proof that s is optimal
We now show that the pure search strategysethtTHORN frac14 t2=4thorn t is the optimal (minimax) search func-tion Since TethsmTHORN does not depend on m it is enough
J R Soc Interface (2011)
to show that for any other search function s frac14 s(t)there is some m such that TethsmTHORN TethsmTHORN To thisend we need the following simple result We adoptthe notation t frac14 ts to denote the time taken for a strat-egy s(t) to explore all of the region X (eg ts frac14 2)
Proposition 42 For any search strategy s [ Ss = seither
1 s0(0) frac14 1 and s00(0) 12 or2 sethtTHORN sethtTHORN for all t in some interval (01) In this
case either (i) ts 2 or (ii) there is a least timet1 0 with setht1THORN setht1THORN and s0etht1THORN frac14 s0etht1THORN
Proof Since s0(0) 1 for all s [ S if condition 1fails then the first sentence of condition 2 must holdIf part 2(i) fails so that ts 2 then by the Intermedi-ate Value Theorem there is a time t2 such thatsetht2THORN frac14 setht2THORN Then since seth0THORN frac14 seth0THORN frac14 0 it followsfrom the Mean Value Theorem that there is a timet1 0 such that s0etht1THORN frac14 s0etht1THORN Since the set of sucht1 is compact there is a least such time B
Figure 4 illustrates five curves from top to bottom ta type 1 curve t 2 t28 the minimax curve s the type2(ii) curve t23 with 2t13 frac14 1 2 t12 or t1 frac14 67 frac14086 the type 2(i) curve t25 with ts 2
Proposition 43 The strategy sethtTHORN frac14 t t2=4 is theminimax pure Searcher strategy That is
mins[S
maxm
TethsmTHORN frac14 maxm
TethsmTHORN frac14 23
Proof We show that for each type of strategy s inproposition 42 we can determine a Hider move time msuch that TethsmTHORN 2=3 If s is of type 1 of proposition42 then Teths 0THORN frac14 1 eth1 s00eth0THORNTHORN 2=3 If s is of type2(i) of proposition 42 Teths 2THORN Teths 2THORN frac14 2=3 If sis type 2(ii) take m frac14 t1 where t1 is the time given byproposition 42 part 2(ii) For this value of m definefs(T ) to be the linear function given by the right sideof equation (41) We claim that the line fs(T ) has ahigher slope and is higher at T frac14 0 than fsT so itintersects the diagonal at a higher point Since T frac14T(sm) is the solution of T frac14 fs (T ) and T frac14 2=3 is thesolution of fsethTTHORN frac14 T we have TethsmTHORN T frac14 2=3
0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)

1666 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
further determine the ideal mixture (cruising prob-ability) for the predator to adopt when the prey hasbeen excluded by search from a fraction of the searchregion Our answer is what we call the Square RootLaw of Predation Adopt the cruising mode with prob-ability equal to the square root of the unexploredfraction of the search region This implies in particularthat when beginning the search the cruising modeshould predominate but when the prey location hasbeen narrowed to a small area the ambush modeshould predominate This discourages the prey frommoving to escape back into the full region
A related attempt to model the ambushndashcruisesearch mixture has been recently presented in Zoroaet al [3] in a more geometrical context On the preyside the question of when to move to a new location(though with more information than we allow) hasbeen studied by Broom amp Ruxton [4]
2 SEARCH GAMES WITH MOBILE HIDER
The game-theoretic background to the problem weconsider lies in the search game with mobile hider morecolourfully named by Isaacs [2] as the Princess andMonster Game This is a zero sum game between asingle predator (the Monster) and a single mobile prey(the Princess) with the capture time T as the pay-off(obviously the Monster is the minimizer) The twoplayers move about in a known search region Q untilthe first time T that the Princess comes within the Mon-sterrsquos detection radius If the search region is a networkthe game ends when the two players meet (detectionradius is zero) The Monster has a maximum speedtaken as 1 which is also the rate at which he exploresnew territory The mathematical theory of Princess andMonster Games was begun in Zeliken [5] and Alpern[6] where the search region was considered a circle andextended [7] to games on networks or regions of n-dimen-sional space Alternative approaches to the latterproblem were given by Lalley amp Robbins [8] and Garnaev[9] Further work on related search games was carried outby Kikuta amp Ruckle [10] Pavlovic [11] and Alpern et al[12] (see also the monographs [1314])
As far as we are aware the only biological appli-cation of Princess and Monster Games is [1516] inthe context of a contest between a wasp (Monster) ofthe genus Sympiesis (Hymenoptera Eulophidae) anda leafminer larvae (Princess) of Phyllonorycter blancar-della (Lepidoptera Gracillariidae) The search region Xis a roughly planar area between the two surfaces of anapple leaf where the feeding activity of the larvae pro-duces a so-called windows randomly placed translucentand whitish small spots in which only the upper cuti-cula remains while the plant tissue underneath iseaten up The wasp is attracted in flight by the visualappearance of the mines lands on the leaf and thusstarts the game Leaf vibrations cause the larvae tocease any feeding and become lsquoalertrsquo The wasp vio-lently inserts its ovipositor nearly exclusively in thefeeding windows as the rapid escape reactions ofthe larva command blazing fast attacks unfeasible ifthe wasp has to pierce thick tissues A successful
J R Soc Interface (2011)
search ends with the larva being parasitized and thewasp laying an egg on it Thus this is a discrete locationapproximation of the Princess and Monster Game
The search game that we introduce in this article addsan extra strategy for the Monster (predator) namely anambush mode that he may adopt at any time whichresults in immediate capture if the Princess is movingat that time The presence of this strategy keeps thePrincess still most of the time except for intermittentshort (actually instantaneous) dashes to new randomlocations On the other hand if the Princess is stationaryduring a time interval when the Monster is ambushingthe Monster effectively does no searching and accom-plishes nothing in this time interval The possibility ofambush behaviour in the Princess and Monster Gamewas first considered by Alpern amp Asic [1718] in thecontext of lsquobottlenecksrsquo in the search region where theMonster might do well to wait in ambush such as atthe central node of the lsquofigure eightrsquo network Ourinterpretation here is not that the Monster will wait ata particular point of the search region but rather thathe will hover or perch somewhere above it where he cansurvey the whole region (or a part of itmdashsee sect46)
We point out that there is a related literature onwhat are called ambush games where the mobile hider(prey) has to move between two known locations (ega known migration pattern) and the Searcher (ambush-ing predator) has to choose locations at which toambush (intercept) the mobile prey This problem ismore geometrical than ours in that the Searcher hasto pick locations rather than as in our model speedor cruisendashambush alternation The literature for suchambush games [19ndash22] may well have applicationsto the relationship between predator location andmigratory patterns of prey For example it is suggestedby Walter [23] that colonies of Eleonorarsquos falcon areplaced to intercept migrations
There is another literature on pursuit-evasion gameswithin the field of differential games (see Isaacs [2])which has applications to the next stage of thepredatorndashprey interaction what happens between thedetection of prey and capture of prey This is an impor-tant problem but not one that we address here Weassume that the predator wishes to minimize the timeto detect (find) the prey keeping our analysis firmlywithin the field of search games
3 RELATIONSHIP WITH THEBEHAVIOURAL AND ECOLOGICALLITERATURE
Nearly all of the many works done on the searching andescaping modes of predator and prey have been done onevolutionary and ecological timescales (see Helfman [24]and Cooper [25] for reviews and Michel amp Adams [26] forone of the latest studies)This ranges from studies focusingon the matching between the searching modes and thegrouprsquos phylogeny to studies highlighting the switch ofsearching modes according to internal state or externalconditions such as temperature obstacles and habitatcomplexity to prey densities In this context speed doesnot vary within a single interaction between a predator
Ambush frequency should increase over time S Alpern et al 1667
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
and a prey By contrast exceedingly few studies focus onthe ethological time scale at which search occurs andduring which motivational or environmental factors donot change much For the same reasons most studiesdo not address the problem of information acquisition bythe opponents during the search game A monographabout the ethology of predation [27] already reported alack of quantitative ethological studies on the geometricaland speed components of predatorndashprey interactions andlamented that many results were based on anecdotes Thisremains basically true as of today
A quantitative assessment of the predator speedduring the interaction has been however carried out ina few cases often related to pursuitndashevasion gamesIn the first case [2829] the bimodal attacking speed dis-tribution observed in wolf spiders is interpreted as anoptimization strategy trading speed on the one handand production of unwanted information used by preyfor escape on the other hand spiders in ambush orwalking slowly towards prey do not produce a largerecognizable aerodynamical signature so they mayachieve capture before revealing their presence to theprey spiders attacking at full speed do produce a lotof such information for the prey but their speed maysuffice to overtake even an escaping prey spiders run-ning at an intermediate speed produce sufficientinformation to be spotted and are not quick enoughto get their prey so these hunting speeds are thereforeavoided In that example speed is nearly constantduring a single pursuit The second case is a searchingstrategy the saltatory search This strategy consists ofmaking small hops or short dashes followed by shortpauses and is thus a mixture of speeds displayedduring a single interaction [3031] It is common infishes and birds predators look for prey when theypause and move bit-by-bit producing a kind ofpseudo-systematic pattern of search Pausing enablesthem also to attend to other tasks such as lookingout for top predators [30] Saltatory search in fisheshas been explained using hydrodynamical forcesacting against cruise search [32] Other functionalexplanations for this behaviour have been given relatedto information acquisition by the predator rather thanby the prey as in the invertebrate example Either theradius of perception is nearly similar to the length ofthe move in the case of fishes [32] or speed itself is ahandicap to sensory acquisition such as for stabilizingvisual images of potential prey in birds and lizards[33] These hypotheses are backed up by recent workshowing that speed decreases acuity [34ndash36] In sum-mary saltatory search lies in the middle of acontinuum between ambush and cruise search The pro-portion of either behaviour remains however constantduring the saltatory search in contrast to our modelVarying speed during an interaction when preybecome cornered or when prey are difficult to subduehas been observed [27]
4 THE AMBUSH SEARCH GAME
This section describes and analyses our modification ofthe Princess and Monster Game which we call the
J R Soc Interface (2011)
Ambush Search Game The Hider (modelling theprey or the Princess in the version of Isaacs [2] andconsidered female) is stationary at a random locationof a search region X of measure (or area) 1 except forintermittent times of her choosing when she can relo-cate (in a quick dash which we take to beinstantaneous) to a new random location The Searcher(modelling the predator or the Monster in Isaacs [2]and considered male) can explore X at a maximumspeed (or discovery rate) of 1 This means that if theSearcher never ambushes he can search the wholeregion in unit time In this case if the Hider does notmove she will be found in an expected time of 12 Inany time interval the Searcher may ambush (inwhich case the Searcher will certainly detect amoving Hider but not do any exploring) or he mayadopt active cruising search (in which case he willexplore a maximum amount of new territory but willnot catch a moving Hider) We assume that when theHider successfully moves (to a new random location)this event is detected by an active Searcher (perhapsthrough the vibration of a leaf in the leafminer case)but the new location of the Hider cannot be ascertainedThus the information available to the Searcher is thesame as at the start of the game when he arrived atthe patch namely that the Hider is uniformly distribu-ted over the search space X Hence we say that in thisevent the original game is repeated Note that when wesay that the Hiderrsquos movement is detected by theSearcher we do not mean that the Hider is capturedonly that the Searcher knows that the Hider is nolonger contained in the smaller region in which he wasknown to be located prior to the move Of course ifthe move occurs during a period of ambush the gameends
A pure search strategy for the Searcher isfully described by a function s(t) with s(0) frac14 0 and0 s0(t) 1 where s(t) is the fraction of X that hehas searched up to time t assuming the Hider has notmoved In so-called saltatory or alternating searchperiods of ambush are characterized by derivativess0(t) frac14 0 and periods of cruising by s0(t) frac14 1 We alsoallow intermediate speeds between 0 and 1 for whichwe have two interpretations In the probabilisticinterpretation we assume that the Searcher maydivide an interval containing time t into many tinytime intervals on each of which he cruises with prob-ability s0(t) and ambushes with probability 1 2 s0(t)So if the Hider moves the Searcher will be in anambush interval with probability 1 2 s0(t) in whichcase she will be caught The other interpretationwhich we shall adopt henceforth is that s0(t) is simplythe speed at which the Searcher is moving throughthe search region (and also his rate of exploring newterritory) In this interpretation we assume that hisability to detect prey decreases linearly with hisspeed so that when moving at speed s0(t) he detectsa prey movement with probability 1 2 s0(t) Theassumption that visual acuity (or other measures oflsquoaccuracyrsquo) is decreasing in predator speed is wellfounded in neuroethological terms in diverse situationssuch as pollination or crypsis [36]
01 02 03 04 05 06 07 08 09 100
10
20
a
m
Figure 1 Optimal Hider moving time methaTHORN (Online version incolour)
05 06 07 08 09 10
08
09
10
a
T
Figure 2 Worst case capture time TethaTHORN (Online version incolour)
1668 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To evaluate the efficiency of a particular search strat-egy s(t) we suppose that the Hider plans to move to anew random location at some time t frac14 m 0 assumingshe is not found by the Searcher before then What isthe expected capture timeT frac14TethsmTHORN in this situationThis timeT satisfies the following equation
T frac14TethsmTHORN
frac14ethm
0t s0ethtTHORN dt thorn eth1 sethmTHORNTHORNethm thorn s0ethmTHORN TTHORN eth41THORN
The first term corresponds to the case where theHider is caught by a moving Searcher before time mbecause the probability of her being found in a smalltime interval [tt thorn Dt] is approximately s0(t) Dt Thesecond term corresponds to the case where the Hiderhas not been found by time m which occurs with prob-ability 1 2 s(m) In this case the capture time is atleast m for sure Furthermore the Hider is successfulin her move at time m with probability s0(m) inwhich case the search game is played again so she getsan additional expected time of T
Our aim is to find the (minimax) search functionsethtTHORN [ C 2 which gives the least expected capture timeT for the worst (highest) case of the Hiderrsquos choice ofm that is
maxm
TethsmTHORN frac14 mins[S
maxm
TethsmTHORN
We expect (and show later) that against the optimalsearch function s the expected capture time will be thesame for all Hider moving times m We first analyse thecase where the Searcher is restricted to a constantsearch speed a (that is s(t) is of the form at) and findthe optimal speed a in this case However the expectedcapture time with this restriction always depends on mso there is no minimax strategy of constant speedThen we analyse quadratic search functions and findan optimal solution
There is an additional technical question (which thereader can disregard for the moment) as to what hap-pens if the Hider is always relocating which weinterpret as moving at time m frac14 0 In this limitingcase the Hiderrsquos expected lifetime T is 0 unless theSearcher is putting all his effort into active search sothat s0(0) frac14 1 and there is no probability of ambushIn this limiting case we have
Teths 0THORN frac14 limm0
TethsmTHORN frac14 11 s00eth0THORN
if s0eth0THORN frac14 1eth42THORN
Since the second derivative arises in this context werequire that our search strategies s belong to the space Sof C2 functions functions with a continuous secondderivative which we now take as the predator purestrategy space
41 Restricted optimization for constant-speedpredators
Suppose we restrict the Searcher to searching with a con-stant speed a 1 so that at time t he has searched a
J R Soc Interface (2011)
fraction of the search space X given by s(t) frac14 at Asusual this assumes that t m that the Hider has notyet moved Substituting at for s(t) in equation (41) wehave an expected capture timeT frac14TethamTHORN satisfying
T frac14 etham 1THORNethmetha 1THORN aethm thornTTHORNTHORN thorn 12
am2
or T frac14 2m am2
2a thorn 2a2 m thorn 2
eth43THORN
For fixed speed a the Hider (prey) maximizesthe capture time T by moving after timemethaTHORN frac14 etha thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffia2 thorn 1p
1THORN=a2 as drawn in figure 1Note that against a full-speed predator the preyshould always be moving so as to remain at a randomlocationmdashthere is no downside to moving causedby ambush Thus the worst case capture timeTetha methaTHORNTHORN when the Searcher moves at speed a isTethaTHORN frac14 eth
ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
thorn a2 1THORN=etha3ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
THORN with mini-mum at a frac14
ffiffiffi3p
=2 ≃ 087 of TethaTHORN frac14 4ffiffiffi3p
=9 ≃ 077 asdrawn in figure 2
Summarizing our results from this section we havethe following
Proposition 41 If the Searcher (predator) isrestricted to moving at a constant speed a (interpretedas a constant mixture over time of ambush and cruisesearch) his best speed is a frac14
ffiffiffi3p
=2 ≃ 087 which guar-antees an expected capture time not exceeding4ffiffiffi3p
=9 ≃ 077 against any moving time m of the Hider
The interpretation of this result is that a predatorwho cannot keep track of how long it has been searchingshould employ active search about 87 per cent of thetime and ambush about 13 per cent of the time Ofcourse this result assumes that an ambushing Searcheralways detects a moving prey regardless of the preyrsquoslocation A more realistic version of this result without
02 04 06 08 10 12 14 16 18 200
02
04
06
08
10
t
s
Figure 3 Optimal fraction sethtTHORN of space searched by time t (Online version in colour)
Ambush frequency should increase over time S Alpern et al 1669
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
certain detection of moving prey by an ambushingpredator is given in sect46
42 An indifference-inducing search strategy s
In this section we determine a particular search strategysethtTHORN for which the Searcher is indifferent to the time mwhen the Hider moves This is a common heuristic for sol-ving zero sum games but is not a sufficient condition foroptimality We show in sect43 that this strategy is indeedthe pure minimax strategy It turns out that the Searchercan reduce the worst case expected capture time from the07698 he could ensure using constant speed down to23 frac14 06666 if he uses a quadratic function ratherthan a linear one Since s(0) frac14 0 (at time 0 he hassearched none of the space) there is no constant termso we assume that s(t) has the form
sethtTHORN frac14 ct2 thorn bt eth44THORN
for unknown constants c and b Substituting this quadra-tic form of s into the expected capture time formula (41)gives
T frac14 16
m2eth3bthorn 4cmTHORN ethethbthorn 2cmTHORNethm thornTTHORN
methbthorn 2cm 1THORNTHORNethcm2 thorn bm 1THORN
or 0 frac14 23
c ceth2cT thorn 1THORN
m3
thorn 12
b beth2cT thorn 1THORN cbT
m2
thorn eth2cT Tb2 thorn 1THORNm thorn ethbT TTHORN
Solving for b c and T so that the coefficients ofpowers of m are all zero (independence of m) gives theoptimal search strategy sethtTHORN frac14 eth1=4THORNt2 thorn t as shownin figure 3
The method of deriving the formula for sethtTHORN givenhere avoids the need for differential equations and inany case the required constancy TethsethtTHORNmTHORN frac14 2=3 canbe verified directly from equation (41) Our model pre-dicts a slowing down of predator speed
43 Proof that s is optimal
We now show that the pure search strategysethtTHORN frac14 t2=4thorn t is the optimal (minimax) search func-tion Since TethsmTHORN does not depend on m it is enough
J R Soc Interface (2011)
to show that for any other search function s frac14 s(t)there is some m such that TethsmTHORN TethsmTHORN To thisend we need the following simple result We adoptthe notation t frac14 ts to denote the time taken for a strat-egy s(t) to explore all of the region X (eg ts frac14 2)
Proposition 42 For any search strategy s [ Ss = seither
1 s0(0) frac14 1 and s00(0) 12 or2 sethtTHORN sethtTHORN for all t in some interval (01) In this
case either (i) ts 2 or (ii) there is a least timet1 0 with setht1THORN setht1THORN and s0etht1THORN frac14 s0etht1THORN
Proof Since s0(0) 1 for all s [ S if condition 1fails then the first sentence of condition 2 must holdIf part 2(i) fails so that ts 2 then by the Intermedi-ate Value Theorem there is a time t2 such thatsetht2THORN frac14 setht2THORN Then since seth0THORN frac14 seth0THORN frac14 0 it followsfrom the Mean Value Theorem that there is a timet1 0 such that s0etht1THORN frac14 s0etht1THORN Since the set of sucht1 is compact there is a least such time B
Figure 4 illustrates five curves from top to bottom ta type 1 curve t 2 t28 the minimax curve s the type2(ii) curve t23 with 2t13 frac14 1 2 t12 or t1 frac14 67 frac14086 the type 2(i) curve t25 with ts 2
Proposition 43 The strategy sethtTHORN frac14 t t2=4 is theminimax pure Searcher strategy That is
mins[S
maxm
TethsmTHORN frac14 maxm
TethsmTHORN frac14 23
Proof We show that for each type of strategy s inproposition 42 we can determine a Hider move time msuch that TethsmTHORN 2=3 If s is of type 1 of proposition42 then Teths 0THORN frac14 1 eth1 s00eth0THORNTHORN 2=3 If s is of type2(i) of proposition 42 Teths 2THORN Teths 2THORN frac14 2=3 If sis type 2(ii) take m frac14 t1 where t1 is the time given byproposition 42 part 2(ii) For this value of m definefs(T ) to be the linear function given by the right sideof equation (41) We claim that the line fs(T ) has ahigher slope and is higher at T frac14 0 than fsT so itintersects the diagonal at a higher point Since T frac14T(sm) is the solution of T frac14 fs (T ) and T frac14 2=3 is thesolution of fsethTTHORN frac14 T we have TethsmTHORN T frac14 2=3
0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)

Ambush frequency should increase over time S Alpern et al 1667
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
and a prey By contrast exceedingly few studies focus onthe ethological time scale at which search occurs andduring which motivational or environmental factors donot change much For the same reasons most studiesdo not address the problem of information acquisition bythe opponents during the search game A monographabout the ethology of predation [27] already reported alack of quantitative ethological studies on the geometricaland speed components of predatorndashprey interactions andlamented that many results were based on anecdotes Thisremains basically true as of today
A quantitative assessment of the predator speedduring the interaction has been however carried out ina few cases often related to pursuitndashevasion gamesIn the first case [2829] the bimodal attacking speed dis-tribution observed in wolf spiders is interpreted as anoptimization strategy trading speed on the one handand production of unwanted information used by preyfor escape on the other hand spiders in ambush orwalking slowly towards prey do not produce a largerecognizable aerodynamical signature so they mayachieve capture before revealing their presence to theprey spiders attacking at full speed do produce a lotof such information for the prey but their speed maysuffice to overtake even an escaping prey spiders run-ning at an intermediate speed produce sufficientinformation to be spotted and are not quick enoughto get their prey so these hunting speeds are thereforeavoided In that example speed is nearly constantduring a single pursuit The second case is a searchingstrategy the saltatory search This strategy consists ofmaking small hops or short dashes followed by shortpauses and is thus a mixture of speeds displayedduring a single interaction [3031] It is common infishes and birds predators look for prey when theypause and move bit-by-bit producing a kind ofpseudo-systematic pattern of search Pausing enablesthem also to attend to other tasks such as lookingout for top predators [30] Saltatory search in fisheshas been explained using hydrodynamical forcesacting against cruise search [32] Other functionalexplanations for this behaviour have been given relatedto information acquisition by the predator rather thanby the prey as in the invertebrate example Either theradius of perception is nearly similar to the length ofthe move in the case of fishes [32] or speed itself is ahandicap to sensory acquisition such as for stabilizingvisual images of potential prey in birds and lizards[33] These hypotheses are backed up by recent workshowing that speed decreases acuity [34ndash36] In sum-mary saltatory search lies in the middle of acontinuum between ambush and cruise search The pro-portion of either behaviour remains however constantduring the saltatory search in contrast to our modelVarying speed during an interaction when preybecome cornered or when prey are difficult to subduehas been observed [27]
4 THE AMBUSH SEARCH GAME
This section describes and analyses our modification ofthe Princess and Monster Game which we call the
J R Soc Interface (2011)
Ambush Search Game The Hider (modelling theprey or the Princess in the version of Isaacs [2] andconsidered female) is stationary at a random locationof a search region X of measure (or area) 1 except forintermittent times of her choosing when she can relo-cate (in a quick dash which we take to beinstantaneous) to a new random location The Searcher(modelling the predator or the Monster in Isaacs [2]and considered male) can explore X at a maximumspeed (or discovery rate) of 1 This means that if theSearcher never ambushes he can search the wholeregion in unit time In this case if the Hider does notmove she will be found in an expected time of 12 Inany time interval the Searcher may ambush (inwhich case the Searcher will certainly detect amoving Hider but not do any exploring) or he mayadopt active cruising search (in which case he willexplore a maximum amount of new territory but willnot catch a moving Hider) We assume that when theHider successfully moves (to a new random location)this event is detected by an active Searcher (perhapsthrough the vibration of a leaf in the leafminer case)but the new location of the Hider cannot be ascertainedThus the information available to the Searcher is thesame as at the start of the game when he arrived atthe patch namely that the Hider is uniformly distribu-ted over the search space X Hence we say that in thisevent the original game is repeated Note that when wesay that the Hiderrsquos movement is detected by theSearcher we do not mean that the Hider is capturedonly that the Searcher knows that the Hider is nolonger contained in the smaller region in which he wasknown to be located prior to the move Of course ifthe move occurs during a period of ambush the gameends
A pure search strategy for the Searcher isfully described by a function s(t) with s(0) frac14 0 and0 s0(t) 1 where s(t) is the fraction of X that hehas searched up to time t assuming the Hider has notmoved In so-called saltatory or alternating searchperiods of ambush are characterized by derivativess0(t) frac14 0 and periods of cruising by s0(t) frac14 1 We alsoallow intermediate speeds between 0 and 1 for whichwe have two interpretations In the probabilisticinterpretation we assume that the Searcher maydivide an interval containing time t into many tinytime intervals on each of which he cruises with prob-ability s0(t) and ambushes with probability 1 2 s0(t)So if the Hider moves the Searcher will be in anambush interval with probability 1 2 s0(t) in whichcase she will be caught The other interpretationwhich we shall adopt henceforth is that s0(t) is simplythe speed at which the Searcher is moving throughthe search region (and also his rate of exploring newterritory) In this interpretation we assume that hisability to detect prey decreases linearly with hisspeed so that when moving at speed s0(t) he detectsa prey movement with probability 1 2 s0(t) Theassumption that visual acuity (or other measures oflsquoaccuracyrsquo) is decreasing in predator speed is wellfounded in neuroethological terms in diverse situationssuch as pollination or crypsis [36]
01 02 03 04 05 06 07 08 09 100
10
20
a
m
Figure 1 Optimal Hider moving time methaTHORN (Online version incolour)
05 06 07 08 09 10
08
09
10
a
T
Figure 2 Worst case capture time TethaTHORN (Online version incolour)
1668 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To evaluate the efficiency of a particular search strat-egy s(t) we suppose that the Hider plans to move to anew random location at some time t frac14 m 0 assumingshe is not found by the Searcher before then What isthe expected capture timeT frac14TethsmTHORN in this situationThis timeT satisfies the following equation
T frac14TethsmTHORN
frac14ethm
0t s0ethtTHORN dt thorn eth1 sethmTHORNTHORNethm thorn s0ethmTHORN TTHORN eth41THORN
The first term corresponds to the case where theHider is caught by a moving Searcher before time mbecause the probability of her being found in a smalltime interval [tt thorn Dt] is approximately s0(t) Dt Thesecond term corresponds to the case where the Hiderhas not been found by time m which occurs with prob-ability 1 2 s(m) In this case the capture time is atleast m for sure Furthermore the Hider is successfulin her move at time m with probability s0(m) inwhich case the search game is played again so she getsan additional expected time of T
Our aim is to find the (minimax) search functionsethtTHORN [ C 2 which gives the least expected capture timeT for the worst (highest) case of the Hiderrsquos choice ofm that is
maxm
TethsmTHORN frac14 mins[S
maxm
TethsmTHORN
We expect (and show later) that against the optimalsearch function s the expected capture time will be thesame for all Hider moving times m We first analyse thecase where the Searcher is restricted to a constantsearch speed a (that is s(t) is of the form at) and findthe optimal speed a in this case However the expectedcapture time with this restriction always depends on mso there is no minimax strategy of constant speedThen we analyse quadratic search functions and findan optimal solution
There is an additional technical question (which thereader can disregard for the moment) as to what hap-pens if the Hider is always relocating which weinterpret as moving at time m frac14 0 In this limitingcase the Hiderrsquos expected lifetime T is 0 unless theSearcher is putting all his effort into active search sothat s0(0) frac14 1 and there is no probability of ambushIn this limiting case we have
Teths 0THORN frac14 limm0
TethsmTHORN frac14 11 s00eth0THORN
if s0eth0THORN frac14 1eth42THORN
Since the second derivative arises in this context werequire that our search strategies s belong to the space Sof C2 functions functions with a continuous secondderivative which we now take as the predator purestrategy space
41 Restricted optimization for constant-speedpredators
Suppose we restrict the Searcher to searching with a con-stant speed a 1 so that at time t he has searched a
J R Soc Interface (2011)
fraction of the search space X given by s(t) frac14 at Asusual this assumes that t m that the Hider has notyet moved Substituting at for s(t) in equation (41) wehave an expected capture timeT frac14TethamTHORN satisfying
T frac14 etham 1THORNethmetha 1THORN aethm thornTTHORNTHORN thorn 12
am2
or T frac14 2m am2
2a thorn 2a2 m thorn 2
eth43THORN
For fixed speed a the Hider (prey) maximizesthe capture time T by moving after timemethaTHORN frac14 etha thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffia2 thorn 1p
1THORN=a2 as drawn in figure 1Note that against a full-speed predator the preyshould always be moving so as to remain at a randomlocationmdashthere is no downside to moving causedby ambush Thus the worst case capture timeTetha methaTHORNTHORN when the Searcher moves at speed a isTethaTHORN frac14 eth
ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
thorn a2 1THORN=etha3ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
THORN with mini-mum at a frac14
ffiffiffi3p
=2 ≃ 087 of TethaTHORN frac14 4ffiffiffi3p
=9 ≃ 077 asdrawn in figure 2
Summarizing our results from this section we havethe following
Proposition 41 If the Searcher (predator) isrestricted to moving at a constant speed a (interpretedas a constant mixture over time of ambush and cruisesearch) his best speed is a frac14
ffiffiffi3p
=2 ≃ 087 which guar-antees an expected capture time not exceeding4ffiffiffi3p
=9 ≃ 077 against any moving time m of the Hider
The interpretation of this result is that a predatorwho cannot keep track of how long it has been searchingshould employ active search about 87 per cent of thetime and ambush about 13 per cent of the time Ofcourse this result assumes that an ambushing Searcheralways detects a moving prey regardless of the preyrsquoslocation A more realistic version of this result without
02 04 06 08 10 12 14 16 18 200
02
04
06
08
10
t
s
Figure 3 Optimal fraction sethtTHORN of space searched by time t (Online version in colour)
Ambush frequency should increase over time S Alpern et al 1669
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
certain detection of moving prey by an ambushingpredator is given in sect46
42 An indifference-inducing search strategy s
In this section we determine a particular search strategysethtTHORN for which the Searcher is indifferent to the time mwhen the Hider moves This is a common heuristic for sol-ving zero sum games but is not a sufficient condition foroptimality We show in sect43 that this strategy is indeedthe pure minimax strategy It turns out that the Searchercan reduce the worst case expected capture time from the07698 he could ensure using constant speed down to23 frac14 06666 if he uses a quadratic function ratherthan a linear one Since s(0) frac14 0 (at time 0 he hassearched none of the space) there is no constant termso we assume that s(t) has the form
sethtTHORN frac14 ct2 thorn bt eth44THORN
for unknown constants c and b Substituting this quadra-tic form of s into the expected capture time formula (41)gives
T frac14 16
m2eth3bthorn 4cmTHORN ethethbthorn 2cmTHORNethm thornTTHORN
methbthorn 2cm 1THORNTHORNethcm2 thorn bm 1THORN
or 0 frac14 23
c ceth2cT thorn 1THORN
m3
thorn 12
b beth2cT thorn 1THORN cbT
m2
thorn eth2cT Tb2 thorn 1THORNm thorn ethbT TTHORN
Solving for b c and T so that the coefficients ofpowers of m are all zero (independence of m) gives theoptimal search strategy sethtTHORN frac14 eth1=4THORNt2 thorn t as shownin figure 3
The method of deriving the formula for sethtTHORN givenhere avoids the need for differential equations and inany case the required constancy TethsethtTHORNmTHORN frac14 2=3 canbe verified directly from equation (41) Our model pre-dicts a slowing down of predator speed
43 Proof that s is optimal
We now show that the pure search strategysethtTHORN frac14 t2=4thorn t is the optimal (minimax) search func-tion Since TethsmTHORN does not depend on m it is enough
J R Soc Interface (2011)
to show that for any other search function s frac14 s(t)there is some m such that TethsmTHORN TethsmTHORN To thisend we need the following simple result We adoptthe notation t frac14 ts to denote the time taken for a strat-egy s(t) to explore all of the region X (eg ts frac14 2)
Proposition 42 For any search strategy s [ Ss = seither
1 s0(0) frac14 1 and s00(0) 12 or2 sethtTHORN sethtTHORN for all t in some interval (01) In this
case either (i) ts 2 or (ii) there is a least timet1 0 with setht1THORN setht1THORN and s0etht1THORN frac14 s0etht1THORN
Proof Since s0(0) 1 for all s [ S if condition 1fails then the first sentence of condition 2 must holdIf part 2(i) fails so that ts 2 then by the Intermedi-ate Value Theorem there is a time t2 such thatsetht2THORN frac14 setht2THORN Then since seth0THORN frac14 seth0THORN frac14 0 it followsfrom the Mean Value Theorem that there is a timet1 0 such that s0etht1THORN frac14 s0etht1THORN Since the set of sucht1 is compact there is a least such time B
Figure 4 illustrates five curves from top to bottom ta type 1 curve t 2 t28 the minimax curve s the type2(ii) curve t23 with 2t13 frac14 1 2 t12 or t1 frac14 67 frac14086 the type 2(i) curve t25 with ts 2
Proposition 43 The strategy sethtTHORN frac14 t t2=4 is theminimax pure Searcher strategy That is
mins[S
maxm
TethsmTHORN frac14 maxm
TethsmTHORN frac14 23
Proof We show that for each type of strategy s inproposition 42 we can determine a Hider move time msuch that TethsmTHORN 2=3 If s is of type 1 of proposition42 then Teths 0THORN frac14 1 eth1 s00eth0THORNTHORN 2=3 If s is of type2(i) of proposition 42 Teths 2THORN Teths 2THORN frac14 2=3 If sis type 2(ii) take m frac14 t1 where t1 is the time given byproposition 42 part 2(ii) For this value of m definefs(T ) to be the linear function given by the right sideof equation (41) We claim that the line fs(T ) has ahigher slope and is higher at T frac14 0 than fsT so itintersects the diagonal at a higher point Since T frac14T(sm) is the solution of T frac14 fs (T ) and T frac14 2=3 is thesolution of fsethTTHORN frac14 T we have TethsmTHORN T frac14 2=3
0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)

01 02 03 04 05 06 07 08 09 100
10
20
a
m
Figure 1 Optimal Hider moving time methaTHORN (Online version incolour)
05 06 07 08 09 10
08
09
10
a
T
Figure 2 Worst case capture time TethaTHORN (Online version incolour)
1668 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To evaluate the efficiency of a particular search strat-egy s(t) we suppose that the Hider plans to move to anew random location at some time t frac14 m 0 assumingshe is not found by the Searcher before then What isthe expected capture timeT frac14TethsmTHORN in this situationThis timeT satisfies the following equation
T frac14TethsmTHORN
frac14ethm
0t s0ethtTHORN dt thorn eth1 sethmTHORNTHORNethm thorn s0ethmTHORN TTHORN eth41THORN
The first term corresponds to the case where theHider is caught by a moving Searcher before time mbecause the probability of her being found in a smalltime interval [tt thorn Dt] is approximately s0(t) Dt Thesecond term corresponds to the case where the Hiderhas not been found by time m which occurs with prob-ability 1 2 s(m) In this case the capture time is atleast m for sure Furthermore the Hider is successfulin her move at time m with probability s0(m) inwhich case the search game is played again so she getsan additional expected time of T
Our aim is to find the (minimax) search functionsethtTHORN [ C 2 which gives the least expected capture timeT for the worst (highest) case of the Hiderrsquos choice ofm that is
maxm
TethsmTHORN frac14 mins[S
maxm
TethsmTHORN
We expect (and show later) that against the optimalsearch function s the expected capture time will be thesame for all Hider moving times m We first analyse thecase where the Searcher is restricted to a constantsearch speed a (that is s(t) is of the form at) and findthe optimal speed a in this case However the expectedcapture time with this restriction always depends on mso there is no minimax strategy of constant speedThen we analyse quadratic search functions and findan optimal solution
There is an additional technical question (which thereader can disregard for the moment) as to what hap-pens if the Hider is always relocating which weinterpret as moving at time m frac14 0 In this limitingcase the Hiderrsquos expected lifetime T is 0 unless theSearcher is putting all his effort into active search sothat s0(0) frac14 1 and there is no probability of ambushIn this limiting case we have
Teths 0THORN frac14 limm0
TethsmTHORN frac14 11 s00eth0THORN
if s0eth0THORN frac14 1eth42THORN
Since the second derivative arises in this context werequire that our search strategies s belong to the space Sof C2 functions functions with a continuous secondderivative which we now take as the predator purestrategy space
41 Restricted optimization for constant-speedpredators
Suppose we restrict the Searcher to searching with a con-stant speed a 1 so that at time t he has searched a
J R Soc Interface (2011)
fraction of the search space X given by s(t) frac14 at Asusual this assumes that t m that the Hider has notyet moved Substituting at for s(t) in equation (41) wehave an expected capture timeT frac14TethamTHORN satisfying
T frac14 etham 1THORNethmetha 1THORN aethm thornTTHORNTHORN thorn 12
am2
or T frac14 2m am2
2a thorn 2a2 m thorn 2
eth43THORN
For fixed speed a the Hider (prey) maximizesthe capture time T by moving after timemethaTHORN frac14 etha thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffia2 thorn 1p
1THORN=a2 as drawn in figure 1Note that against a full-speed predator the preyshould always be moving so as to remain at a randomlocationmdashthere is no downside to moving causedby ambush Thus the worst case capture timeTetha methaTHORNTHORN when the Searcher moves at speed a isTethaTHORN frac14 eth
ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
thorn a2 1THORN=etha3ffiffiffiffiffiffiffiffiffiffiffiffiffi1 a2p
THORN with mini-mum at a frac14
ffiffiffi3p
=2 ≃ 087 of TethaTHORN frac14 4ffiffiffi3p
=9 ≃ 077 asdrawn in figure 2
Summarizing our results from this section we havethe following
Proposition 41 If the Searcher (predator) isrestricted to moving at a constant speed a (interpretedas a constant mixture over time of ambush and cruisesearch) his best speed is a frac14
ffiffiffi3p
=2 ≃ 087 which guar-antees an expected capture time not exceeding4ffiffiffi3p
=9 ≃ 077 against any moving time m of the Hider
The interpretation of this result is that a predatorwho cannot keep track of how long it has been searchingshould employ active search about 87 per cent of thetime and ambush about 13 per cent of the time Ofcourse this result assumes that an ambushing Searcheralways detects a moving prey regardless of the preyrsquoslocation A more realistic version of this result without
02 04 06 08 10 12 14 16 18 200
02
04
06
08
10
t
s
Figure 3 Optimal fraction sethtTHORN of space searched by time t (Online version in colour)
Ambush frequency should increase over time S Alpern et al 1669
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
certain detection of moving prey by an ambushingpredator is given in sect46
42 An indifference-inducing search strategy s
In this section we determine a particular search strategysethtTHORN for which the Searcher is indifferent to the time mwhen the Hider moves This is a common heuristic for sol-ving zero sum games but is not a sufficient condition foroptimality We show in sect43 that this strategy is indeedthe pure minimax strategy It turns out that the Searchercan reduce the worst case expected capture time from the07698 he could ensure using constant speed down to23 frac14 06666 if he uses a quadratic function ratherthan a linear one Since s(0) frac14 0 (at time 0 he hassearched none of the space) there is no constant termso we assume that s(t) has the form
sethtTHORN frac14 ct2 thorn bt eth44THORN
for unknown constants c and b Substituting this quadra-tic form of s into the expected capture time formula (41)gives
T frac14 16
m2eth3bthorn 4cmTHORN ethethbthorn 2cmTHORNethm thornTTHORN
methbthorn 2cm 1THORNTHORNethcm2 thorn bm 1THORN
or 0 frac14 23
c ceth2cT thorn 1THORN
m3
thorn 12
b beth2cT thorn 1THORN cbT
m2
thorn eth2cT Tb2 thorn 1THORNm thorn ethbT TTHORN
Solving for b c and T so that the coefficients ofpowers of m are all zero (independence of m) gives theoptimal search strategy sethtTHORN frac14 eth1=4THORNt2 thorn t as shownin figure 3
The method of deriving the formula for sethtTHORN givenhere avoids the need for differential equations and inany case the required constancy TethsethtTHORNmTHORN frac14 2=3 canbe verified directly from equation (41) Our model pre-dicts a slowing down of predator speed
43 Proof that s is optimal
We now show that the pure search strategysethtTHORN frac14 t2=4thorn t is the optimal (minimax) search func-tion Since TethsmTHORN does not depend on m it is enough
J R Soc Interface (2011)
to show that for any other search function s frac14 s(t)there is some m such that TethsmTHORN TethsmTHORN To thisend we need the following simple result We adoptthe notation t frac14 ts to denote the time taken for a strat-egy s(t) to explore all of the region X (eg ts frac14 2)
Proposition 42 For any search strategy s [ Ss = seither
1 s0(0) frac14 1 and s00(0) 12 or2 sethtTHORN sethtTHORN for all t in some interval (01) In this
case either (i) ts 2 or (ii) there is a least timet1 0 with setht1THORN setht1THORN and s0etht1THORN frac14 s0etht1THORN
Proof Since s0(0) 1 for all s [ S if condition 1fails then the first sentence of condition 2 must holdIf part 2(i) fails so that ts 2 then by the Intermedi-ate Value Theorem there is a time t2 such thatsetht2THORN frac14 setht2THORN Then since seth0THORN frac14 seth0THORN frac14 0 it followsfrom the Mean Value Theorem that there is a timet1 0 such that s0etht1THORN frac14 s0etht1THORN Since the set of sucht1 is compact there is a least such time B
Figure 4 illustrates five curves from top to bottom ta type 1 curve t 2 t28 the minimax curve s the type2(ii) curve t23 with 2t13 frac14 1 2 t12 or t1 frac14 67 frac14086 the type 2(i) curve t25 with ts 2
Proposition 43 The strategy sethtTHORN frac14 t t2=4 is theminimax pure Searcher strategy That is
mins[S
maxm
TethsmTHORN frac14 maxm
TethsmTHORN frac14 23
Proof We show that for each type of strategy s inproposition 42 we can determine a Hider move time msuch that TethsmTHORN 2=3 If s is of type 1 of proposition42 then Teths 0THORN frac14 1 eth1 s00eth0THORNTHORN 2=3 If s is of type2(i) of proposition 42 Teths 2THORN Teths 2THORN frac14 2=3 If sis type 2(ii) take m frac14 t1 where t1 is the time given byproposition 42 part 2(ii) For this value of m definefs(T ) to be the linear function given by the right sideof equation (41) We claim that the line fs(T ) has ahigher slope and is higher at T frac14 0 than fsT so itintersects the diagonal at a higher point Since T frac14T(sm) is the solution of T frac14 fs (T ) and T frac14 2=3 is thesolution of fsethTTHORN frac14 T we have TethsmTHORN T frac14 2=3
0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)
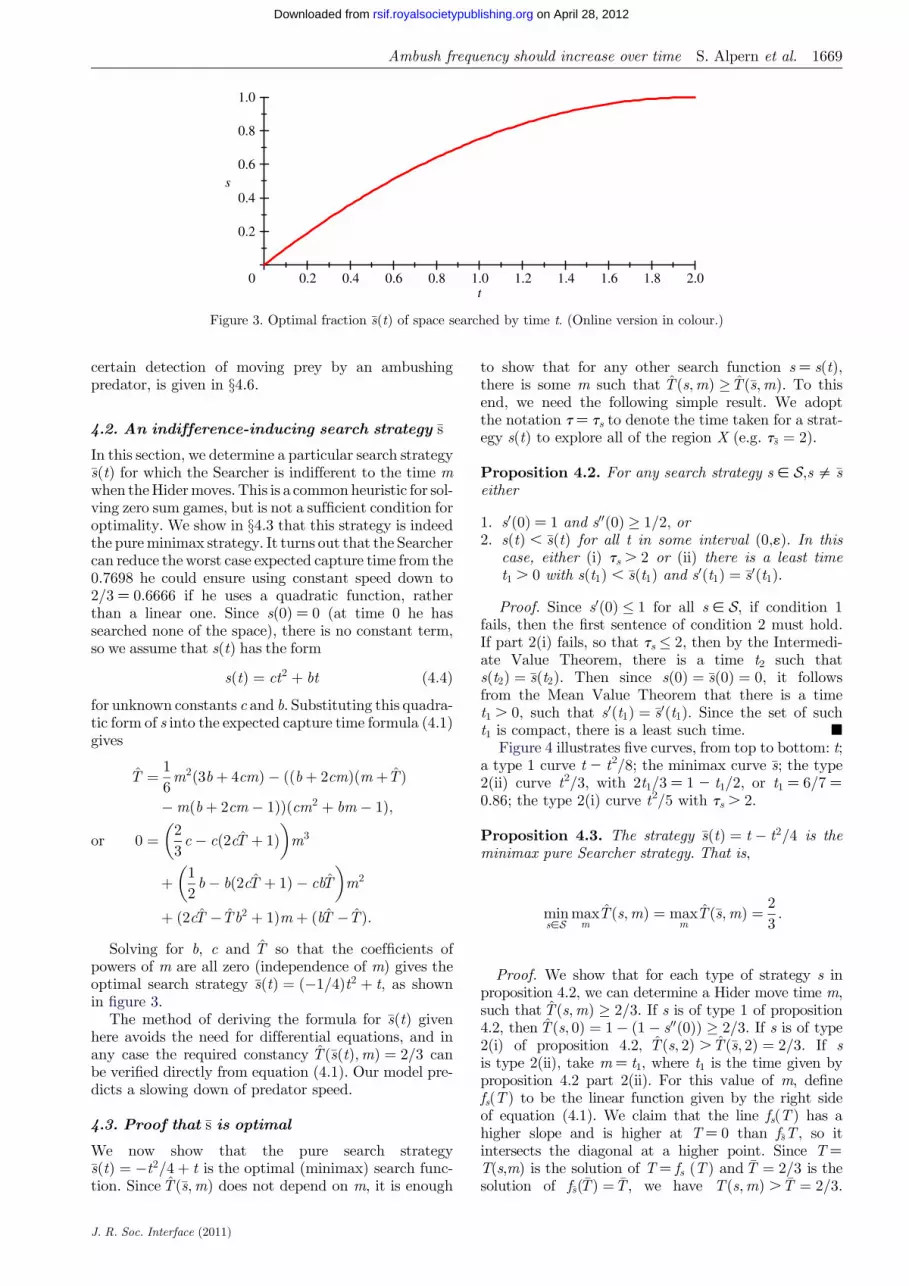
02 04 06 08 10 12 14 16 18 200
02
04
06
08
10
t
s
Figure 3 Optimal fraction sethtTHORN of space searched by time t (Online version in colour)
Ambush frequency should increase over time S Alpern et al 1669
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
certain detection of moving prey by an ambushingpredator is given in sect46
42 An indifference-inducing search strategy s
In this section we determine a particular search strategysethtTHORN for which the Searcher is indifferent to the time mwhen the Hider moves This is a common heuristic for sol-ving zero sum games but is not a sufficient condition foroptimality We show in sect43 that this strategy is indeedthe pure minimax strategy It turns out that the Searchercan reduce the worst case expected capture time from the07698 he could ensure using constant speed down to23 frac14 06666 if he uses a quadratic function ratherthan a linear one Since s(0) frac14 0 (at time 0 he hassearched none of the space) there is no constant termso we assume that s(t) has the form
sethtTHORN frac14 ct2 thorn bt eth44THORN
for unknown constants c and b Substituting this quadra-tic form of s into the expected capture time formula (41)gives
T frac14 16
m2eth3bthorn 4cmTHORN ethethbthorn 2cmTHORNethm thornTTHORN
methbthorn 2cm 1THORNTHORNethcm2 thorn bm 1THORN
or 0 frac14 23
c ceth2cT thorn 1THORN
m3
thorn 12
b beth2cT thorn 1THORN cbT
m2
thorn eth2cT Tb2 thorn 1THORNm thorn ethbT TTHORN
Solving for b c and T so that the coefficients ofpowers of m are all zero (independence of m) gives theoptimal search strategy sethtTHORN frac14 eth1=4THORNt2 thorn t as shownin figure 3
The method of deriving the formula for sethtTHORN givenhere avoids the need for differential equations and inany case the required constancy TethsethtTHORNmTHORN frac14 2=3 canbe verified directly from equation (41) Our model pre-dicts a slowing down of predator speed
43 Proof that s is optimal
We now show that the pure search strategysethtTHORN frac14 t2=4thorn t is the optimal (minimax) search func-tion Since TethsmTHORN does not depend on m it is enough
J R Soc Interface (2011)
to show that for any other search function s frac14 s(t)there is some m such that TethsmTHORN TethsmTHORN To thisend we need the following simple result We adoptthe notation t frac14 ts to denote the time taken for a strat-egy s(t) to explore all of the region X (eg ts frac14 2)
Proposition 42 For any search strategy s [ Ss = seither
1 s0(0) frac14 1 and s00(0) 12 or2 sethtTHORN sethtTHORN for all t in some interval (01) In this
case either (i) ts 2 or (ii) there is a least timet1 0 with setht1THORN setht1THORN and s0etht1THORN frac14 s0etht1THORN
Proof Since s0(0) 1 for all s [ S if condition 1fails then the first sentence of condition 2 must holdIf part 2(i) fails so that ts 2 then by the Intermedi-ate Value Theorem there is a time t2 such thatsetht2THORN frac14 setht2THORN Then since seth0THORN frac14 seth0THORN frac14 0 it followsfrom the Mean Value Theorem that there is a timet1 0 such that s0etht1THORN frac14 s0etht1THORN Since the set of sucht1 is compact there is a least such time B
Figure 4 illustrates five curves from top to bottom ta type 1 curve t 2 t28 the minimax curve s the type2(ii) curve t23 with 2t13 frac14 1 2 t12 or t1 frac14 67 frac14086 the type 2(i) curve t25 with ts 2
Proposition 43 The strategy sethtTHORN frac14 t t2=4 is theminimax pure Searcher strategy That is
mins[S
maxm
TethsmTHORN frac14 maxm
TethsmTHORN frac14 23
Proof We show that for each type of strategy s inproposition 42 we can determine a Hider move time msuch that TethsmTHORN 2=3 If s is of type 1 of proposition42 then Teths 0THORN frac14 1 eth1 s00eth0THORNTHORN 2=3 If s is of type2(i) of proposition 42 Teths 2THORN Teths 2THORN frac14 2=3 If sis type 2(ii) take m frac14 t1 where t1 is the time given byproposition 42 part 2(ii) For this value of m definefs(T ) to be the linear function given by the right sideof equation (41) We claim that the line fs(T ) has ahigher slope and is higher at T frac14 0 than fsT so itintersects the diagonal at a higher point Since T frac14T(sm) is the solution of T frac14 fs (T ) and T frac14 2=3 is thesolution of fsethTTHORN frac14 T we have TethsmTHORN T frac14 2=3
0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)

0 02 04 06 08 10 12 14 16 18 20 22 24
02
04
06
08
10
t
s
Figure 4 Curves s frac14 t type 1 sethtTHORN type 2(ii) type 2(i) from top (Online version in colour)
1670 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
To demonstrate our first claim observe that
f 0sethTTHORN f 0sethTTHORN frac14 ethsethmTHORN sethmTHORNTHORNs0ethmTHORN 0
For the second claim observe that fs(0) is the expectedtime tofind a stationaryHider in the truncated formwherethe capture time is taken to be m if it is not found beforethen Since s finds such a Hider by any given time with ahigher probability than does s it follows that this expectedtruncated capture time is higher for s than for s B
44 The square root rule
If the Searcher is adopting the optimal (minimax cap-ture time) pure strategy sethtTHORN frac14 etht t2=4THORN then thetime at which a fraction u frac14 1 sethtTHORN of the space isunexplored is given by
u frac14 1 t t2
4
or t frac14 2 2
ffiffiffiup
So the optimal search speed (or cruising probability)is given by
dsdtfrac14 1 t
2frac14 1 2 2
ffiffiffiup
2frac14
ffiffiffiup
Proposition 44 When the prey location is restrictedto an unsearched fraction u of the search region theminimax strategy for the predator is to adopt a speedor cruising probability of the square root of u
45 Mixed strategies and noisy predators
So far we have taken the perspective of the predator(Searcher) and found the pure strategy s which mini-mizes the worst case capture time If we take a gametheoretic viewpoint which includes the perspective ofthe prey (Hider) there are two cases which we calllsquosilent predatorrsquo and lsquonoisy predatorrsquo In the silent pred-ator case the Hider only knows that time since she lastmoved as she cannot hear the speed of the Searcher andhence does not know how much territory has beensearched In the noisy predator case she can hear thespeed of the Searcher and can conclude the amount ofterritory s that has been searchedmdashand can base herstrategy on this parameter The noisy game is one oflsquoperfect informationrsquo in that each player knows the por-tions of the otherrsquos strategy that has already beenplayed Such games typically (in finite versions) have
J R Soc Interface (2011)
pure strategy solutions and indeed we find such asolution for the noisy version of the ambush game
We have shown in proposition 43 that the Searcher canensure an expected capture time not exceeding 23 byusing the pure strategy s and that this is the best possibleestimate A question that naturally arises is whether or nothe can do better (lower estimate on expected search time)byemployingmixed strategiesTheanswer to this questiondepends on the information available to the Hiderwhether the game is lsquonoisyrsquo or lsquosilentrsquo The terms noisy orsilent predator refer to the information available to theprey as to the speed of the predator and hence as to theamount of territory the predator has already searched Inthe noisy version if the prey knows that the unsearchedterritory is very small then the predator has an addedincentive to move to avoid being found very soon
First we consider the lsquosilent predatorrsquo model wherethe Hider knows only the time t since she last movedFor this version consider the two pure Searcher strat-egies s1(t) frac14 t 2 (18)t2 and s2(t) frac14 t 2 (38)t2 oneither side of the minimax strategy s frac14 t eth1=4THORNt2The mixed strategy consisting of s1 with probability513 and s2 with probability 813 has a maximumexpected search time (against any moving time m) of6091 which is less than the pure strategy minimaxtime of 23 So in the silent predator model theSearcher can improve on the expected search time of23 by adopting mixed strategies The optimal mixtureof pure strategies is not known
Next we consider the lsquonoisy predatorrsquo model whereat every time t0 the Hider knows the prior Searcherpath s(t) t t0 In the noisy predator game a Hiderstrategy is a function H S R where the movingtime is m frac14 H(s) such that for any sa sb [ S we have
m frac14 H ethsaTHORN frac14 H ethsbTHORN if saethtTHORN frac14 sbethtTHORNfor t m
eth45THORN
which simply means that the Hider cannot predict theSearcherrsquos future motion
Proposition 45 In the noisy predator game wherethe Hider knows the Searcherrsquos motion up to the currenttime the value is 23 and the pure Searcher strategy s isthe optimal mixed strategy The Hider also has an opti-mal pure strategy H
Proof The Hider moves at time m frac14 HethsTHORN with H asfollows If s0(0) frac14 1 and s00(0) 14 then HethsTHORN frac14 0Otherwise let H ethsTHORN be the least time t that
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)
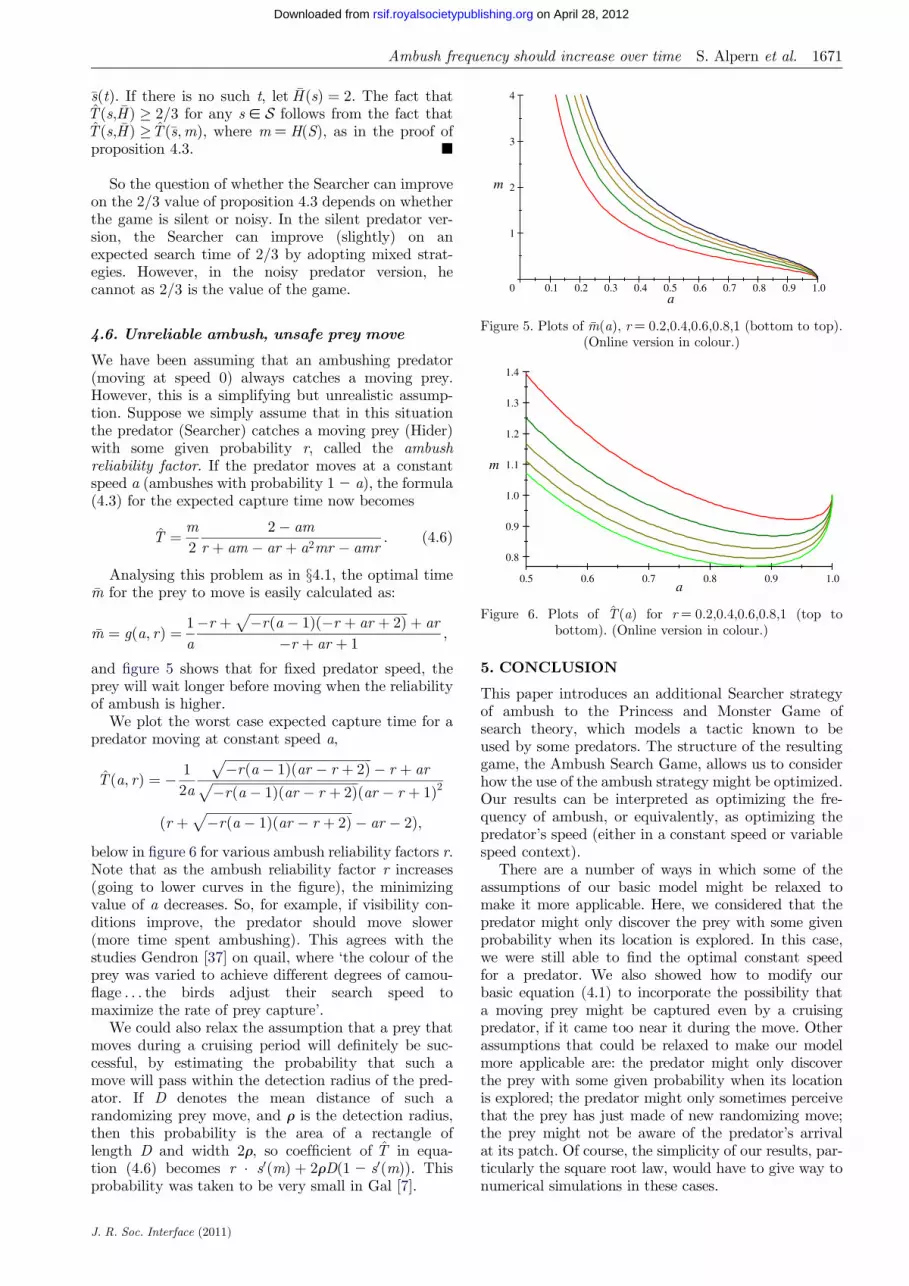
01 02 03 04 05 06 07 08 09 100
1
2
3
4
m
Ambush frequency should increase over time S Alpern et al 1671
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
sethtTHORN If there is no such t let H ethsTHORN frac14 2 The fact thatTeths H THORN 2=3 for any s [ S follows from the fact thatTeths H THORN TethsmTHORN where m frac14 H(S) as in the proof ofproposition 43 B
So the question of whether the Searcher can improveon the 23 value of proposition 43 depends on whetherthe game is silent or noisy In the silent predator ver-sion the Searcher can improve (slightly) on anexpected search time of 23 by adopting mixed strat-egies However in the noisy predator version hecannot as 23 is the value of the game
a
Figure 5 Plots of methaTHORN r frac14 020406081 (bottom to top)(Online version in colour)
05 06 07 08 09 10
08
09
10
11
12
13
14
a
m
Figure 6 Plots of TethaTHORN for r frac14 020406081 (top tobottom) (Online version in colour)
46 Unreliable ambush unsafe prey move
We have been assuming that an ambushing predator(moving at speed 0) always catches a moving preyHowever this is a simplifying but unrealistic assump-tion Suppose we simply assume that in this situationthe predator (Searcher) catches a moving prey (Hider)with some given probability r called the ambushreliability factor If the predator moves at a constantspeed a (ambushes with probability 1 2 a) the formula(43) for the expected capture time now becomes
T frac14 m2
2 amr thorn am ar thorn a2mr amr
eth46THORN
Analysing this problem as in sect41 the optimal timem for the prey to move is easily calculated as
m frac14 getha rTHORN frac14 1ar thorn
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethr thorn ar thorn 2THORN
pthorn ar
r thorn ar thorn 1
and figure 5 shows that for fixed predator speed theprey will wait longer before moving when the reliabilityof ambush is higher
We plot the worst case expected capture time for apredator moving at constant speed a
Tetha rTHORN frac14 12a
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p r thorn arffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
retha 1THORNethar r thorn 2THORNp
ethar r thorn 1THORN2
ethr thornffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiretha 1THORNethar r thorn 2THORN
p ar 2THORN
below in figure 6 for various ambush reliability factors rNote that as the ambush reliability factor r increases(going to lower curves in the figure) the minimizingvalue of a decreases So for example if visibility con-ditions improve the predator should move slower(more time spent ambushing) This agrees with thestudies Gendron [37] on quail where lsquothe colour of theprey was varied to achieve different degrees of camou-flage the birds adjust their search speed tomaximize the rate of prey capturersquo
We could also relax the assumption that a prey thatmoves during a cruising period will definitely be suc-cessful by estimating the probability that such amove will pass within the detection radius of the pred-ator If D denotes the mean distance of such arandomizing prey move and r is the detection radiusthen this probability is the area of a rectangle oflength D and width 2r so coefficient of T in equa-tion (46) becomes r s0(m) thorn 2rD(1 2 s0(m)) Thisprobability was taken to be very small in Gal [7]
J R Soc Interface (2011)
5 CONCLUSION
This paper introduces an additional Searcher strategyof ambush to the Princess and Monster Game ofsearch theory which models a tactic known to beused by some predators The structure of the resultinggame the Ambush Search Game allows us to considerhow the use of the ambush strategy might be optimizedOur results can be interpreted as optimizing the fre-quency of ambush or equivalently as optimizing thepredatorrsquos speed (either in a constant speed or variablespeed context)
There are a number of ways in which some of theassumptions of our basic model might be relaxed tomake it more applicable Here we considered that thepredator might only discover the prey with some givenprobability when its location is explored In this casewe were still able to find the optimal constant speedfor a predator We also showed how to modify ourbasic equation (41) to incorporate the possibility thata moving prey might be captured even by a cruisingpredator if it came too near it during the move Otherassumptions that could be relaxed to make our modelmore applicable are the predator might only discoverthe prey with some given probability when its locationis explored the predator might only sometimes perceivethat the prey has just made of new randomizing movethe prey might not be aware of the predatorrsquos arrivalat its patch Of course the simplicity of our results par-ticularly the square root law would have to give way tonumerical simulations in these cases
1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)

1672 Ambush frequency should increase over time S Alpern et al
on April 28 2012rsifroyalsocietypublishingorgDownloaded from
Mixtures of cruising and ambushing as we find hereare known to exist in other domains of search theoryand practice for example when a convict escapes fromprison the police search the known hide-outs (activesearch) but also put up roadblocks to prevent the escapeefrom switching between them (ambush) In conclusionour model raises new functional questions regarding thedynamics of the search phase of many predatorndashpreyinteractions and adds a new dimension to the generaltheory of search games with mobile hiders
SA and RF were supported by NATO CollaborativeResearch Grant CLG 983583
REFERENCES
1 Schoener T W 1971 Theory of feeding strategies AnnuRev Ecol Syst 2 369ndash404 (doi101146annureves02110171002101)
2 Isaacs R 1965 Differential games New York NY Wiley3 Zoroa N Fernandez-Saez M J amp Zoroa P 2011 A foraging
problem sit-and-wait versus active predation Eur JOperatRes 208 131ndash141 (doi101016jejor201008001)
4 Broom M amp Ruxton G 2005 You can runmdashor you canhide optimal strategies for cryptic prey against pursuitpredators Behav Ecol 16 534ndash540 (doi101093behecoari024)
5 Zeliken M 1972 On a differential game with incompleteinformation Soviet Math Doklady 13 228ndash231
6 Alpern S 1974 The search game with mobile hideron the circle In Differential games and control theory(eds E O Roxin P T Liu amp R L Sternberg) pp181ndash200 New York NY Marcel Dekker
7 Gal S 1979 Search games with mobile and immobilehider SIAM J Control Optim 17 99ndash122 (doi1011370317009)
8 Lalley S P amp Robbins H E 1987 Asymptotically minimaxstochastic search strategies in the plane ProcNatl Acad SciUSA 84 2111ndash2112 (doi101073pnas8482111)
9 Garnaev A Y 1992 A remark on the Princess and Mon-ster search game Int J Game Theory 20 269ndash276(doi101007BF01253781)
10 Kikuta K amp Ruckle W 1994 Initial point search onweighted trees Naval Res Log 41 821ndash831 (doi1010021520-6750(199410)416821AID-NAV322041061030CO2-Y)
11 Pavlovic L 1995 A search game on the union of graphswith immobile hider Naval Res Log 42 1177ndash1199(doi1010021520-6750(199512)4281177AID-NAV322042080530CO2-R)
12 Alpern S Fokkink R Lindelauf R amp Olsder G 2008 ThelsquoPrincess and Monsterrsquo game on an interval SIAM J ControlOptim 47 1178ndash1190 (doi101137060672054)
13 Garnaev A Y 2000 Search games and other applications ofgame theory Lecture Notes in Economics and MathematicalSystems vol 485 p 145 Berlin Germany Springer
14 Alpern S amp Gal S 2003 The theory of search games andrendezvous Kluwer International Series in OperationsResearch and Management Sciences p 319 BostonGermany Kluwer
15 Meyhofer R Casas J amp Dorn S 1997 Vibration-mediatedinteractions in a hostndashparasitoid system Proc R Soc LondB 264 261ndash266 (doi101098rspb19970037)
16 Djemai I Meyhofer R amp Casas J 2000 Geometricalgames in a hostndashparasitoid system Am Nat 156257ndash265 (doi101086303388)
J R Soc Interface (2011)
17 Alpern S amp Asic M 1985 The search value of a networkNetworks 15 229ndash238 (doi101002net3230150208)
18 Alpern S amp Asic M 1986 Ambush strategies in searchgames on graphs SIAM J Control Optim 24 66ndash75(doi1011370324002)
19 Ruckle W H 1981 Ambushing random walks II continu-ous models Oper Res 29 18ndash120
20 Woodward I D 2003 Discretization of the continuousambush game Naval Res Log 50 515ndash529 (doi101002nav10066)
21 Baston V J amp Kikuta K 2004 An ambush game with anunknown number of intruders Oper Res 52 597ndash605(doi101287opre10400122)
22 Baston V J amp Kikuta K 2009 Technical notemdashan ambush game with a fat infiltrator Oper Res 57514ndash519 (doi101287opre10800569)
23 Walter H 1979 Eleonorarsquos falcon adaptations to prey andhabitat in a social raptor Chicago IL University ofChicago Press
24 Helfman G S 1990 Mode selection and mode switching inforaging animals Adv Study Behav 19 249ndash298 (doi101016S0065-3454(08)60205-3)
25 Cooper W E 2005 The foraging mode controversyboth continuous variation and clustering of foragingmovements occur J Zool 267 179ndash190 (doi101017S0952836905007375)
26 Michel M J amp Adams M M 2009 Differential effects ofstructural complexity on predator foraging behaviorBehav Ecol 20 313ndash317 (doi101093behecoarp005)
27 Curio E 1976 The ethology of predation BerlinGermany Springer
28 Dangles O Ory N Steinmann T Christides J P ampCasas J 2006 Spiderrsquos attack versus cricketrsquos escapevelocity modes determine success Anim Behav 72603ndash610 (doi101016janbehav200511018)
29 Casas J Steinmann T amp Dangles O 2008 The aerody-namic signature of running spiders PLoS ONE 3 e2116(doi101371journalpone0002116)
30 OrsquoBrien W J Evans B I amp Browman H I 1989 Flex-ible search tactics and efficient foraging in saltatorysearching animals Oecologia 80 100ndash110 (doi101007BF00789938)
31 Kramer D L amp McLaughlin R L 2001 The behavioralecology of intermittent locomotion Am Zool 41137ndash153 (doi1016680003-1569(2001)041[0137TBEOIL]20CO2)
32 Dowling N A Hall S J amp Mitchell J G 2000 Foragingkinematics of barramundi during early stages of develop-ment J Fish Biol 57 337ndash353 (doi101111j1095-86492000tb02176x)
33 Andersen J P Stephens D W amp Dunbar S R 1997Saltatory search a theoretical analysis Behav Ecol 8307ndash317 (doi101093beheco83307)
34 Gendron R P amp Staddon J E R 1983 Searching forcryptic prey the effect of search rate Am Nat 121172ndash186 (doi101086284049)
35 Gendron R P amp Staddon J E R 1984 A laboratorysimulation of foraging behavior the effect of search rateon the probability of detecting prey Am Nat 124407ndash415 (doi101086284281)
36 Chittka L Skorupski P amp Raine N E 2009 Speedndashaccuracy tradeoffs in animal decision making TrendsEcol Evol 24 400ndash407 (doi101016jtree200902010)
37 Gendron R 1986 Searching for cryptic prey evidence foroptimal search rates and the formation of search images inquail Anim Behav 34 898ndash912 (doi101016S0003-3472(86)80076-8)





























