Embed Size (px)
Citation preview

1 23
Folia GeobotanicaJournal of the Institute of Botany,Academy of Sciences of the CzechRepublic ISSN 1211-9520Volume 50Number 3 Folia Geobot (2015) 50:239-252DOI 10.1007/s12224-015-9218-8
An appraisal of eremophyte diversity andplant traits in a rocky desert of the Sahara
Lyès Bradai, M’hammed Bouallala,Noura Fatiha Bouziane, Sabah Zaoui,Souad Neffar & Haroun Chenchouni

An appraisal of eremophyte diversity and plant traitsin a rocky desert of the Sahara
Lyès Bradai & M’hammed Bouallala & Noura Fatiha Bouziane &
Sabah Zaoui & Souad Neffar & Haroun Chenchouni
Received: 4 July 2014 /Accepted: 5 July 2015 /Published online: 16 July 2015# Institute of Botany, Academy of Sciences of the Czech Republic 2015
Abstract This study aims to identify and describe com-munity characteristics and species traits of eremophyticvegetation in the Hammada of Debdeb (Illizi, AlgerianSahara). We recorded abundance, species richness, bio-diversity (Shannon index and evenness) and plant traits(life forms, dispersal types, morphological types, adap-tive strategies of Grime, Noy-Meir’s categories andphytogeographical types). The results revealed the pres-ence of 14 plant species in 11 families with the domi-nance of Asteraceae. The dominant plant taxa at sur-veyed sites of the Hammada desert were chamaephytes,perennials and anemochorous plants whereas the dom-inant phytogeographical origin of plants is the Saharo-Arabian type. According to Grime’s classification, mostof plant strategies were stress-tolerant, followed by thestress-tolerant ruderal type. In addition, arido-activespecies predominated over arido-passive plants. Theeremophytic vegetation of the Hammada is character-ized by low species density (Shannon index rangedbetween 1.12 and 2.84, evenness from 0.48 to 0.82).
The xerophytic species of the rocky desert under studyhave sparse and scattered spatial distribution with lowdensity. Plant traits of this eremophilous flora denote itsstrategies to adapt to adverse environmental conditionsof the Hammada desert including the hot hyper-aridclimate and the extremely poor rocky-substrate.
Keywords Drylands . Functional ecology . Life-historystrategies . Plant diversity . Rocky desert . Sahara .
Xerophytes
Introduction
Think of a desert, and the image that most likely comesto mind is of high sand dunes stretching endlessly in alldirections, in a landscape devoid of life, empty anddesolate. However, deserts are not like this everywhere.Many are mainly mountainous, and others are rocky.Much of the Sahara is a plateau; the surface is rocky,consisting of exposed horizontal bedrock, in placespartly covered by boulders or smaller rocks (Allaby2008; Goudie 2002).
Although deserts are barren, they are not devoid of life(Quinn 2009). There are many plants and animals thathave evolved to tolerate the harsh conditions of even thehottest, coldest, windiest, and driest deserts (Whitford2002). The Sahara Desert, which occupies 10 % of thearea of the African continent, is the largest hot desert onEarth (Goudie 2002; Laity 2009). The Algerian Sahara,which covers about 2 million km2 i.e. 85 % of thecountry’s surface, is well known for its extreme aridity.
Folia Geobot (2015) 50:239–252DOI 10.1007/s12224-015-9218-8
L. Bradai :M. Bouallala :N. F. Bouziane : S. ZaouiLaboratoire Bio-ressources sahariennes: préservation etvalorisation, Faculty of Natural and Life Sciences, Universityof Kasdi Merbah, 30000 Ouargla, Algeria
S. Neffar :H. Chenchouni (*)Department of Biological Sciences, University of Tebessa,12002 Tebessa, Algeriae-mail: [email protected]
M. BouallalaDepartment of Natural and Life Sciences, Faculty of Sciencesand Technology, University of Ahmed Draia,01000 Adrar, Algeria

Despite the vast extent of the Algerian Sahara, speciesrichness and endemism are low; approximately 1,200plant species are given by Ozenda (2004). Despite this,some adapted species survive with extraordinary formsof adaptations (Shmida et al. 1986). Within the Sahara,the vegetation cover is discontinuous, sparse and veryirregular. Plants use mainly places where water supply isa little less unfavourable than elsewhere (Ozenda 2004).
A variety of landscapes and geomorphological formsexist in the Sahara Desert; the most common includeErgs‘large wind-swept sand dunes’, Regs ‘extensive gravel-covered plains’, Dayas ‘shallow, seasonally inundatedbasins’, Wadis ‘ephemeral streams’, valleys, Hammadas‘rocky plateaus’ and Oases (Quézel 1965). Thisecoregion includes extensive rocky deserts ‘Hammadadeserts’, which are flat rocky plateaus with very monot-onous topography where the rock can outcrop into blocksor slabs (Goudie 2002), as in the cases of the Hammadaof Guir and the Hammada of Tindouf on the Algerian-Moroccan borders. These rocky hot deserts are the leastproductive ecosystems worldwide with the harshest ter-restrial ecological conditions for the survival of livingbeings. In an advanced stage of surface weathering, theterm Hammada desert refers to a landscape type wherethe rock surface may be covered with rock fragments, thefiner waste being transported by wind and water(Benhouhou et al. 2003; Bouallala 2013). Furthermore,the Hammada is a landscape type that does not allow alarge extension of vegetation even following relativelyheavy rainfall because most of the surface area is occu-pied by a solid rock (Adam 1962). Still, some species,such as Anabasis spp., Fagonia spp. or Fredolia spp., aredistributed across large areas, on which these speciesappear as the only permanent vegetation cover duringthe hot season (Huetz de Lemps 1970; Bouallala 2013).
In hot arid deserts, where only a relatively smallnumber of species survive, the classification of ecolog-ical communities and the understanding of involvedecological processes are easier to achieve than in areasof high diversity and ecological complexity (Cowlishawand Davies 1997). However, it has been demonstratedthat the composition of life forms in drylands reflects theresponse of vegetation to variation in certain environ-mental factors (El-Demerdash et al. 1995), such as lowrainfall (Noy-Meir 1973), especially for ephemeral spe-cies which are highly dependent on soil moisture andtexture (El-Demerdash et al. 1995), elevation (Burke2001), and topography (Vetaas 1993). The involvementof several factors in determining traits of arid vegetation
explains the difficulty to survey and classify desert plantcommunities, despite their low diversity and cover(Dasti and Agnew 1994). Nevertheless, methods havebeen developed to study desert vegetation as well asseveral numerical analyses to describe these desert plantcommunities (Olsvig-Whittaker et al. 1983; Shmidaet al. 1986; Maestre et al. 2012; Osman 2014).
It has even been proposed by Walter (1986) that thedescription of plant communities would be more effec-tive on the basis of geomorphological features ratherthan on the type of vegetation, especially for arido-passive (ephemeral) plants. Perennial species, by con-trast, form stable communities in different geomorpho-logical habitats encountered in the Sahara (Goldberg andTurner 1986), which provides the basis for a classifica-tion of plant communities into groups or life formsreflecting a combination of similar biological traits andsimilar responses to environmental factors (Grime 2006).
The reviewed literature reports several studies onSaharan plants (e.g. Quézel 1965; Quézel 1978;Ozenda 2004). However, very few studies investigatedthe functional characteristics of the flora living inHammada deserts, although most studies dealing withdesert vegetation attempt to describe plant diversity col-onizing studied habitats by at least assigning life forms tospecies. However, it is not always easy to achieve such asimple approach because adaptations of desertic speciesvary following the severity of local conditions.
It is within this context that the current study focuseson the characterization of vegetation of a particularSaharan biotope, which is the Hammada. Thus, it aimsto complement existing knowledge about the variabilityof flora distributed over various habitats and landscapesof the Algerian Sahara (Chehma et al. 2010). In fact, theSaharan vegetation is known to be organized followingdifferent types of habitats or landscapes (Benhouhouet al. 2003; Chehma et al. 2005). Natural habitats ofthe Algerian Sahara Desert are used for grazing, mainlyby camels farmed in extensive breeding mode, but withdifferent degrees of preference depending on plant rich-ness, vegetation cover and seasonal productivity of thehabitat (Chehma et al. 2005; Bouallala 2013).
Although several Hammadas exist in the Sahara andthe Arabian deserts, the vegetation of these vast areashas been very little investigated. The purpose of thisstudy is to perform a quantitative and qualitative assess-ment of the plant diversity in the Hammada of Debdeb(Illizi region, Algeria). In particular, our main aim is tocharacterize the plant community through plant traits,
240 L. Bradai et al.

which are a relevant tool for a better understanding ofthe relationship between vegetation and their environ-mental conditions, as well for assessing the state ofbiodiversity (McGill et al. 2006; Neffar et al. 2015).
Materials and methods
Study area
As part of the Algerian Sahara, the province ‘Wilaya’ ofIllizi extends over an area of 291,403 km2 with a perim-eter of 1,233 km. Its geographical coordinates lie betweenlatitudes 21°50′ N and 30°34′ N and longitudes 05°54′ Eand 12°00′ E. It is bordered to the east by Libya, to thewest by the province of Tamanrasset, to the north by theprovince of Ouargla and to the south by Niger (Fig. 1).
Based on climatic data spanning 22 years (1992–2013), provided by the weather station of Illizi (NOM2014), the annual average of temperatures is about 25.3± 8.0°C with a maximum in July of 34.9 ± 1.0°C and aminimum in January of 13.04 ± 1.8°C. Precipitation islow and irregular; the annual average is 30.92 ± 2.29mm. The wettest month is April with an average of 6.71± 3.23 mm, while the least rainy month is August with0.44 ± 1.2 mm. The relative humidity in the study area isoften less than 40%, with an annual average 28.9 ± 10.2%. Sandstorms with speed exceeding 100 km/h arecommon in March and April . Moreover, DeMartonne’s aridity index (De Martonne 1925) revealsa value of 0.89, indicating a hyper-arid climate wherethe drought lasts throughout the year based on theombrothermic diagram (Fig. 1).
Sampling and selection of study sites
The study was carried out in the municipality ofDebdeb, located 470 km north of the chief-town ofIllizi and near a border post to Libya. Because nopublished work is available on the description of plantspecies in this area and given the sparse physiognomy ofthe vegetation of the region (Ozenda 2004), we opted fora mixed sampling design in four different sites (Fig. 1).Site 1 (29°47′11.74″ N, 09°23′32.64″ E, elevation: 445m a.s.l.) is located west of Debdeb on the roadconnecting Debdeb to In-Amenas. Site 2 (29°39′31.80″ N, 09°23′57.62″ E, elevation: 480 m a.s.l.) islocated west of Debdeb on the road Debdeb–In-Amenas, at 20 km south of Site 1. Site 3 (30°00′
23.45″ N, 09°24′48.97″ E, elevation: 377 m a.s.l.) islocated 10 km north of the town of Debdeb. Site 4(30°04′22.73″ N, 09°25′34.43″ E, elevation: 358 ma.s.l.) is located 40 km north of the town of Debdeband at a distance of 30 km south of Site 3.
At the largest scale, these sites have similar geomor-phological and pedological characteristics, which con-sist of the same bedrock of the ‘dark palaeozoic lime-stone’ type, in which sandy soil accumulates in smallinterstices. However, at the landscape scale, differencesbetween sites are related to local geomorphologicalfeatures, grazing frequency and faunal diversity.
The sites were subjectively selected to represent thegeneral conditions of the Hammada desert. At each ofthe sites, vegetation was systematically sampled in tenquadrats of 36 m2 each (6 m × 6 m), chosen randomlywithin an area of 1 ha. In each quadrat, a relevé wasperformed during the month of March 2012. Samplingduring this season maximizes the number of speciesencountered because March coincides with theflowering phase of the majority of the eremophilousplants when rainfall is sufficient to trigger annual ger-mination and growth. In each quadrat, we recordedabundance by estimating the number ‘exact count’ ofindividuals present of each plant species (Gounot 1969).Species were identified according to Quézel and Santa(1962; 1963) and Ozenda (2004).
Data analyses
Abundance and diversity
To study ecological and functional characteristics ofplant species in each study site and the whole region,the following parameters and ecological indices wereestimated:
– Relative abundance (RA) expressed by the ratiobetween numbers of species belonging to the family/ the total number of inventoried species × 100;
– Spatial occurrence: frequency of occurrence (Occ)for a given species is equal to the ratio between thenumber of records (k) where the species was presentto the total (K) of ten surveys carried out (Faurieet al. 2003): Occ (%) = k/K × 100. Plant specieswere classified into five occurrence classes(Bensizerara et al. 2013): Class I: includes very rarespecies with occurrence frequency < 20%, Class II:the occurrence varies from 21 % to 40 % (rare
Eremophyte diversity and plant traits of a rocky desert “Hammada” 241

species), Class III: the frequency varies from 41 %to 60 % (frequent species), Class IV: the frequencyvaries from 61–80 % (abundant species), Class V:occurrence frequency > 80 % (species is very abun-dant and constant); and
– Species diversity: For each relevé and the wholestudy site, α diversity was estimated by three non-parametric indices (Magurran 2004), namely: (i)species richness (SR), estimated as the total numberof species present per sample, (ii) specific biodiver-sity was measured by Shannon's index (H) thatgives the ratio between the number of plant species
and their abundances: H ¼ �∑SR1 ðPi � log2Pi),
where Pi is the proportion of the ith species in agiven relevé or site, and (iii) Shannon’s evenness(E): E = H/Hmax was computed to estimate organi-zational distribution of plant population within eachrelevé and site community, where Hmax = log2SR.Evenness varies from 0, signifying the dominanceof one species, to 1, where all population densitiesare evenly distributed.
Plant characteristics
The identified taxa were characterized by plant attri-butes (morphological, physiological or phenologicalcharacteristics), most of them having an impact ongrowth, reproduction and survival of plants (Violleet al. 2007):
– Life forms: Plant were classified according tothe system established by Raunkiær (1934),who organized the vegetation according tothe position of survival organs (and thereforetheir meristems of growth) during theunfavourable periods. This allocation was per-formed using data from the flora of Algeria(Quézel and Santa 1962; 1963; Quézel 1965)and the database ‘eFlora’ of Tela Botanica(www.tela-botanica.org/eflore);
– Dispersal types: This classification is related tothe characteristics of diaspores and regenera-tion strategies (Médail et al. 1998). As thegeographical distribution of plants is closely
Fig. 1 Geographic locations of the study sites (solid squares) in the region of Debdeb (Illizi Province, Desert Sahara of Algeria), includingthe climatic diagram of Gaussen and Bagnouls for the period 1992–2013.
242 L. Bradai et al.

linked to their ability to spread over longdistances, van der Pijl (1982) identified sixcategories of diaspores according to internalor external, biotic or abiotic factors involvedin diaspore dispersion: anemochorous ,hydrochorous, zoochorous (endozoochorousand epizoochorous) , barochorous , andautochorous. Anemochorous, hydrochorousand zoochorous species are plants, which thedisperser factor of seeds is wind, water andanimals, respectively;
– Morphological types: According to different envi-ronmental adaptations and life cycle of plants, twomajor biological groups were identified: annual andperennial herbaceous, and woody perennials(Ghezlaoui et al. 2011);
– Types of Grime (C.S.R.): The classification ofGrime links biological traits to three broad levelsof stresses: environmental type, biotic disturbances,and inter- and intraspecific competitions. Plant spe-cies we identified were classified following Grime’scategories (Grime 2006). This classification in-cludes three primary strategies, namely strategy C(competitive), strategy S (stress-tolerant) and strat-egy R (ruderal); and secondary strategies, namelycompetitive ruderal (CR), competitive stress-tolerant (CS), and stress-tolerant ruderal (RS);
– Noy-Meir Types: The surveyed taxa were classifiedaccording to dryland vegetation model of Noy-Meir(1973) into two categories: ‘arido-passive’ are speciesthat do not display photosynthetic activity during thedry period, and ‘arido-active’ species (xeromorphicplants well-adapted to the Saharan climate), whichmaintain photosynthetic activity during the dry period,even with reduced rates. Plant species of the DebdebHammada were fitted to Noy-Meir’s types followingspecies list of Jauffret (2001); and
– Phytogeographical types: phytogeographical char-acteristics of the recorded species were determinedbased on the flora of Algeria and desert regions(Quézel and Santa 1962, 1963; Quézel 1965) andthe database ‘eFlora’ (www.tela-botanica.org/eflore). Thus, the inventoried flora was classifiedaccording to a phytogeographical system dividedinto six chorological types: (i) Mediterraneanplants, (ii) endemic species at Algerian or NorthAfrican scale, (iii) Holarctic plants (iv) Saharan‐Arabian plants, (v) plants of tropical origin, and(vi) cosmopolitan.
Statistical analyses
The Pearson's Chi-squared test (χ2) was applied to testthe null hypothesis of independence of categories of planttraits among the four study sites. Descriptive statistics,namely mean, standard deviation (SD), interquartilerange (IQR), and coefficient of variation (CV) of biodi-versity indices N, SR, H, Hmax, and E were given for thestudy sites based on data compiled in ten relevés per site.The homoscedasticity of datasets was verified usingBartlett's tests before examining differences in biodiver-sity indices between study sites. Non-parametric Kruskal-Wallis tests were performed to examine site differences invalues of N, SR, H, Hmax, and E.
The distribution of vegetation among the four studysites was analysed based on species densities using acorrespondence analysis (CA). The CA biplot was rep-resented on a symmetric plot using two factorial axesgiving the highest rate of inertia.
Statistical significance of all tests was determined at aprobability of 0.05. All statistical tests described abovewere performed with the Rcmdr package, a graphicaluser interface for R (Fox 2005).
Results
Flora composition and structure
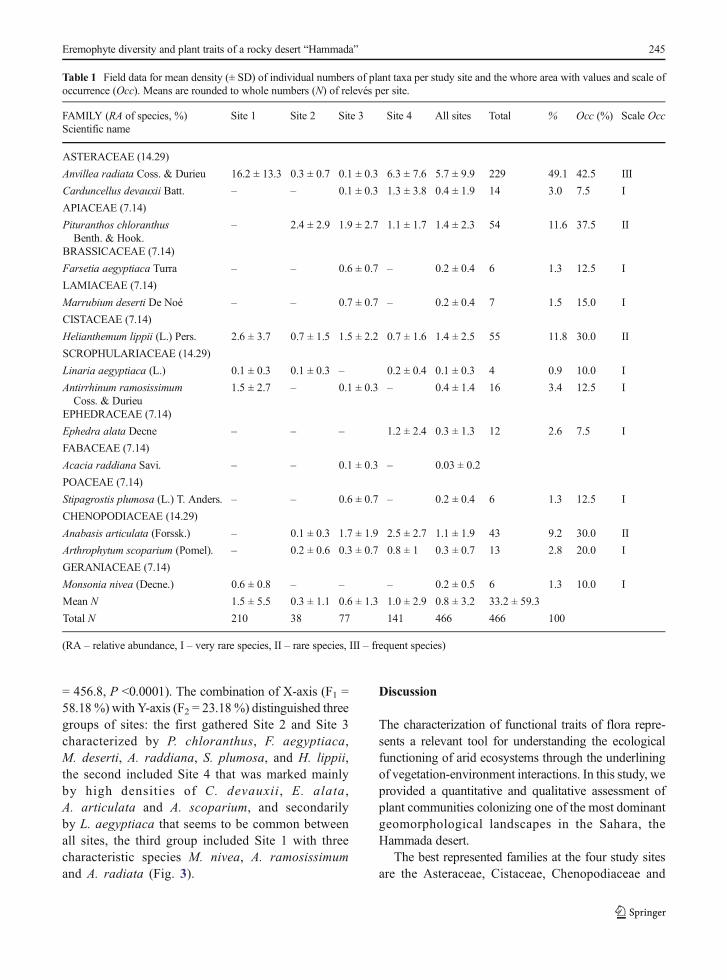
A total of 14 plant taxa were found on the Hammada ofDebdeb belonging to 14 genera and 11 families. Themajority of surveyed families included one species ex-cept the Asteraceae and Chenopodiaceae comprisingtwo species each, i.e. the equivalent of 14.29 % of thetotal census. On the 466 individuals sampled, theAsteraceae family predominates in the Hammada with52.1 % of individuals, followed by Apiaceae(Pituranthos chloranthus), Cistaceae (Helianthemumlippii) and Chenopodiaceae (Anabasis articulata andArthrophytum scoparium) with 11 % each. These fourfamilies comprise 87.6 % of the total plant density and42.9 % of the total species richness. The remainingfamilies reported rates lower than 4 % (Table 1). Thespecies Anvillea radiata is the most abundant with atotal of 229 individuals (49.1 %) and the most frequent(Occ = 42.5 %) in the study area. Its density was higherat Site 1 where it registered 16.2 ± 13.3 individuals perquadrat (36 m2). Besides this species, the occurrence ofPituranthos chloranthus, Anabasis articulata and
Eremophyte diversity and plant traits of a rocky desert “Hammada” 243

Helianthemum lippii in the various relevés was rare,with 37.5 %, 30.0 % and 30.0 %, respectively. The otherten species have very scarce presences. The Chi-squaredtest revealed a significant difference of percentagesof plant occurrence categories between study sites(χ2
6 = 18.2, P = 0.006).
Diversity parameters
Site 1 had the largest density of individuals with anaverage of 21.00 ± 16.27 individuals per relevé, follow-ed by Site 4 with 11.65 ± 11.42 individuals/relevé.According to data of the 10 phytoecological relevés,Site 3 totalled 11 plant species (average SR = 3.60 ±2.41 species/relevé), followed by Site 4 with a total of 8species (average SR = 3.00 ± 0.94 species/relevé), andSite 2 with 6 species (N = 3.80 ± 4.34, average SR = 1.20± 1.55 species/relevé). Following the biodiversity mea-sured in the ten relevés of each site, Site 3 had thehighest values with H = 1.49 ± 0.89 bits and E = 0.74± 0.39 per relevé, followed by Site 4 with H = 1.20 ±0.57 and E = 0.75 ± 0.21 per relevé. Furthermore, Site 2followed by Site 1 were less diversified with the lowestvalues of SR, H, Hmax, and E. In fact, values of CVof all parameters of diversity were higher at Site 2.In addition, outputs of Kruskal-Wallis test revealedthat values of N, SR, H, Hmax, and E varied signif-icantly (P < 0.05) between the four survey sites(Table 2).
Plant traits
Functional traits
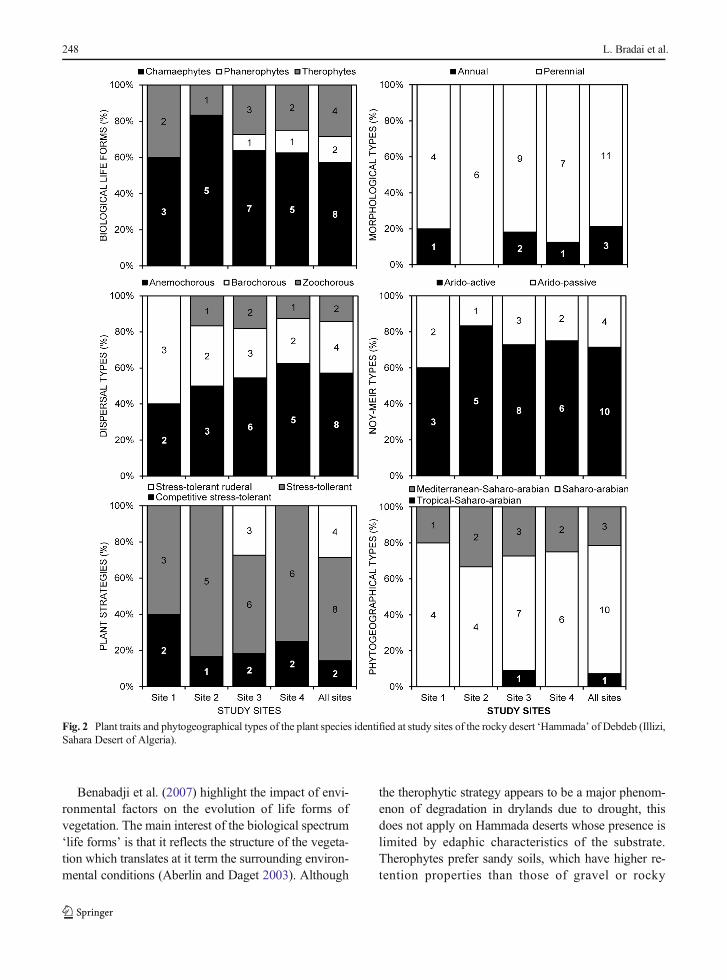
Chamaephytes were the predominant life form witheight species (i.e. 57.14 % of the flora), followed bytherophytes with four species (28.57 %) and two phan-erophytes (14.29 %; Table 3). Spatial occurrences of lifeforms revealed that chamaephytes dominated in the fourstudy sites with presences > 60 %. Therophytes weremainly found at Site 1 with 40 % of presences.The phanerophytes were present only at sites 3and 4 with 9 % and 13 %, respectively (Fig. 2).Chi-square test indicated that distribution of lifeforms on the study sites was statistically indepen-dent (χ2
6 = 8.35, P = 0.213). As for morphologicaltypes, perennials dominated in all sites with 71.4% against 28.6 % of annual plants (Fig. 2). The
distribution of morphological types over surveysites was statistically independent (χ2
3 = 4.76, P= 0.190), but the Chi-square test was significantfor the dispersal patterns of plants (χ2
6 = 15.71, P= 0.015). The results for this parameter showedthat anemochorous-type plants were dominant inall sampled sites, where their frequencies rangedbetween 40 % and 62.5 %. The frequencies ofbarochorous and zoochorus species were 28.5 %and 14.2 %, respectively (Fig. 2).
The results from primary plant strategies ofGrime showed that all species had the ‘stress-toler-ant’ type. A clear dominance of this type of strat-egy (S) was observed all over the study sites withrates exceeding 54 %. Secondary plant strategies ofGrime such as stress-tolerant ruderal species (RS)were moderately present (28.5 %), while competi-tive stress-tolerant plants (CS) were slightly repre-sented with only two species (14.3 %). The strategytypes CSR of Grime were statistically independentof the surveyed sites (χ2
6 = 6.64, P = 0.356).Finally, Noy-Meir classification of plants revealedthat ten species (71.4 %) were arido-actives andfour (28.6 %) were arido-passives. The arido-active plants dominated clearly in the four surveyedsites with percentages varying between 60 % and83.3 % (Fig. 2); however, the Chi-square test wasnot significant (χ2
3 = 4.76, P = 0.190).
Phytogeographical types
The flora of the surveyed Hammada had a typicalSaharo-Arabian phytogeographical affinity with 10taxa, i.e. 71.4 % of total plants having a Sharan-Arabian distribution (Table 3). Three species (21.4 %)had Mediterranean-Saharo-Arabian origin (M-SA). Thegroup of tropical-Saharo-Arabian was represented onlyby Acacia raddiana at Site 3 (7.1 %). No depen-dence was detected in the distribution of the phy-togeographical types of the plants across the studysites (χ2
6 = 2.09, P = 0.911).
Spatial distribution of plant species among sites
The two axes F1 × F2 of the correspondence analysis bi-plot totalled 81.36 % of cumulative inertia. The Chisquare test associated with CA revealed a high signifi-cant link between species densities and study sites (χ2
39
244 L. Bradai et al.
= 456.8, P <0.0001). The combination of X-axis (F1 =58.18 %) with Y-axis (F2 = 23.18 %) distinguished threegroups of sites: the first gathered Site 2 and Site 3characterized by P. chloranthus, F. aegyptiaca,M. deserti, A. raddiana, S. plumosa, and H. lippii,the second included Site 4 that was marked mainlyby high densities of C. devauxii, E. alata ,A. articulata and A. scoparium, and secondarilyby L. aegyptiaca that seems to be common betweenall sites, the third group included Site 1 with threecharacteristic species M. nivea, A. ramosissimumand A. radiata (Fig. 3).
Discussion
The characterization of functional traits of flora repre-sents a relevant tool for understanding the ecologicalfunctioning of arid ecosystems through the underliningof vegetation-environment interactions. In this study, weprovided a quantitative and qualitative assessment ofplant communities colonizing one of the most dominantgeomorphological landscapes in the Sahara, theHammada desert.
The best represented families at the four study sitesare the Asteraceae, Cistaceae, Chenopodiaceae and
Table 1 Field data for mean density (± SD) of individual numbers of plant taxa per study site and the whore area with values and scale ofoccurrence (Occ). Means are rounded to whole numbers (N) of relevés per site.
FAMILY (RA of species, %)Scientific name
Site 1 Site 2 Site 3 Site 4 All sites Total % Occ (%) Scale Occ
ASTERACEAE (14.29)
Anvillea radiata Coss. & Durieu 16.2 ± 13.3 0.3 ± 0.7 0.1 ± 0.3 6.3 ± 7.6 5.7 ± 9.9 229 49.1 42.5 III
Carduncellus devauxii Batt. – – 0.1 ± 0.3 1.3 ± 3.8 0.4 ± 1.9 14 3.0 7.5 I
APIACEAE (7.14)
Pituranthos chloranthusBenth. & Hook.
– 2.4 ± 2.9 1.9 ± 2.7 1.1 ± 1.7 1.4 ± 2.3 54 11.6 37.5 II
BRASSICACEAE (7.14)
Farsetia aegyptiaca Turra – – 0.6 ± 0.7 – 0.2 ± 0.4 6 1.3 12.5 I
LAMIACEAE (7.14)
Marrubium deserti De Noé – – 0.7 ± 0.7 – 0.2 ± 0.4 7 1.5 15.0 I
CISTACEAE (7.14)
Helianthemum lippii (L.) Pers. 2.6 ± 3.7 0.7 ± 1.5 1.5 ± 2.2 0.7 ± 1.6 1.4 ± 2.5 55 11.8 30.0 II
SCROPHULARIACEAE (14.29)
Linaria aegyptiaca (L.) 0.1 ± 0.3 0.1 ± 0.3 – 0.2 ± 0.4 0.1 ± 0.3 4 0.9 10.0 I
Antirrhinum ramosissimumCoss. & Durieu
1.5 ± 2.7 – 0.1 ± 0.3 – 0.4 ± 1.4 16 3.4 12.5 I
EPHEDRACEAE (7.14)
Ephedra alata Decne – – – 1.2 ± 2.4 0.3 ± 1.3 12 2.6 7.5 I
FABACEAE (7.14)
Acacia raddiana Savi. – – 0.1 ± 0.3 – 0.03 ± 0.2
POACEAE (7.14)
Stipagrostis plumosa (L.) T. Anders. – – 0.6 ± 0.7 – 0.2 ± 0.4 6 1.3 12.5 I
CHENOPODIACEAE (14.29)
Anabasis articulata (Forssk.) – 0.1 ± 0.3 1.7 ± 1.9 2.5 ± 2.7 1.1 ± 1.9 43 9.2 30.0 II
Arthrophytum scoparium (Pomel). – 0.2 ± 0.6 0.3 ± 0.7 0.8 ± 1 0.3 ± 0.7 13 2.8 20.0 I
GERANIACEAE (7.14)
Monsonia nivea (Decne.) 0.6 ± 0.8 – – – 0.2 ± 0.5 6 1.3 10.0 I
Mean N 1.5 ± 5.5 0.3 ± 1.1 0.6 ± 1.3 1.0 ± 2.9 0.8 ± 3.2 33.2 ± 59.3
Total N 210 38 77 141 466 466 100
(RA – relative abundance, I – very rare species, II – rare species, III – frequent species)
Eremophyte diversity and plant traits of a rocky desert “Hammada” 245

Apiaceae. These families are widespread in most conti-nental habitats of North Africa and across theMediterranean regions (Paradis and Piazza 2002). The
number of plant species found in the study Hammada,however, is very low, probably as a result of the severeenvironmental conditions prevailing in such ecosystems(Shmida et al. 1986). Indeed, this is a common charac-teristic of hot drylands in which the low biodiversity isassociated with the vastness of arid lands, low primaryproductivity and low plant population densities.
Our results highlighted significant differences in spe-cies diversity but close similarities in functional traitsacross sites. These differences may be attributed to somespatial peculiarities between sites such as: (i) the degreeof grazing; (ii) morphological characteristics of the pla-teau, that influence various edaphic factors includingquantities and depth of accumulated sand and hence soilmoisture; (iii) local differences in the effects of climateparameters, mainly rainfall and wind; and (iv) an unevendispersal of seeds that is related to all the previousfeatures. On the other hand, similarities in functionaltraits of plants denote that the eremophytes of Hammadaof Debdeb likely belong to the same plant communitythat is well adapted to rocky desert conditions.
The spatial differences are apparent in the CA plot(Fig. 3). However, because it is difficult to relate thedistinction of three groups of sites to particular geomor-phological (due to homogeneousness of the Hammada)or soil factors (which are not investigated in the currentstudy), we referred to plant traits of these groups todescribe the possible factors involved in thispartitioning. The dispersal types were taken as the maincriterion due to their dependency between sites (signif-icant Chi-square test). We speculate that grazing may berelated to x-axis because plants of Group 1 are mainlyzoochorous that have RS and CS strategies.Whereas they-axis may be related to a gradual abiotic stress as mostof plants of Group 2 were anemochorous with stress-tolerant strategy (S). The likely factor involved with thisaxis is soil moisture of the shallow sand restrained inrock interstices that is dependent of sand depth and windwhich accelerates evaporation and contributes in seeddispersion.
This study confirms the importance of chamaephytesand therophytes plant life forms in hot desert environ-ments (Osman et al. 2014). The abundance of chamae-phytes in the study area is probably due to their adapta-tions to aridity (Jauffret and Visser 2003; Chenchouni2012; Bouallala 2013). The absence of geophytes main-ly reflects the long period of drought in these drylands,but also the unfavourable edaphic conditions of theHammada (Gamoun et al. 2012) where generally
Table 2 Diversity indices of eremophytes recorded in theHammada of Debdeb in the Algerian Sahara Desert.
Parameters Site 1 Site 2 Site 3 Site 4 All sites
Number of individuals (N)
Mean 21.00 3.80 7.70 14.10 11.65
SD 16.27 4.34 6.11 7.37 11.42
IQR 28.25 7.25 7.00 10.75 14.25
CV 0.77 1.14 0.79 0.52 0.98
Total 210 38 77 141 466
Kruskal-Wallis (K2 = 11.68, d.f. = 3, P = 0.009)
Species richness (SR)
Mean 2.10 1.20 3.60 3.00 2.48
SD 1.29 1.55 2.41 0.94 1.83
IQR 1.00 2.50 3.75 2.00 3.00
CV 0.61 1.29 0.67 0.31 0.74
Total 5 6 11 8 14
Kruskal-Wallis (K2 = 8.65, d.f. = 3, P = 0.034)
Shannon index (H)
Mean 0.72 0.43 1.49 1.20 0.96
SD 0.50 0.73 0.89 0.57 0.78
IQR 0.34 0.69 1.06 0.91 1.61
CV 0.69 1.69 0.60 0.48 0.81
Total 1.12 1.66 2.84 2.41 2.56
Kruskal-Wallis (K2 = 10.41, d.f. = 3, P = 0.015)
Hmax
Mean 1.08 0.52 1.65 1.52 1.19
SD 0.66 0.84 1.03 0.47 0.87
IQR 0.58 1.19 1.44 1.00 2.00
CV 0.62 1.62 0.62 0.31 0.73
Total 2.32 2.58 3.46 3.00 3.81
Kruskal-Wallis (K2 = 9.33, d.f. = 3, P = 0.025)
Evenness (E)
Mean 0.55 0.25 0.74 0.75 0.57
SD 0.34 0.41 0.39 0.21 0.39
IQR 0.36 0.44 0.11 0.12 0.88
CV 0.62 1.66 0.53 0.28 0.69
Total 0.48 0.64 0.82 0.80 0.67
Kruskal-Wallis (K2 = 8.87, d.f. = 3, P = 0.031)
(SD – standard deviation, IQR – interquartile range, CV – coeffi-cient of variation, Total – value of the parameter in questioncomputed for all 10 replications in each site, K2 – statistical valueof Kruskal-Wallis test, d.f. – degrees of freedom, P – P-value).
246 L. Bradai et al.
shallow rocky substrates limit growth. Furthermore, aspersistent species, the chamaephytes are assumed to bebetter adapted to summer drought than are phanero-phytes because they exert an extreme stomatal control(Danin and Orshan 1990).
The growth of therophytes may be explained by thepresence of favourable conditions that trigger their ger-mination. When there is sufficient rain, these plantsparticularly grow in small interstices of rocks wheresand accumulates. In some locations, annual plants aremore abundant in depressions where sand accumulateswithin rock blocks or slabs (Gamoun et al. 2010). Thislife form is a characteristic of drylands (Daget 1980;Gamoun et al. 2012), where their density is controlledby variations of rainfall and temperature (El-Demerdashet al. 1995). The therophytic type appears to be anadaptive strategy to adverse conditions and a form ofresistance to arid climatic severity (Daget 1980). Neffaret al. (2013) show that therophytes have a limited eco-logical importance for rehabilitation and restorationprograms in arid rangelands due to their short lifecycle that lasts only few weeks under severeconditions. Daget (1980) demonstrated that the rate oftherophytes and its increase over time could be an
indicator of an increase in climatic aridity and/or habitatdegradation. Moreover, therophytes are more resistantto summer drought than hemicryptophytes andgeophytes because they spend long period as seedswhile others remain in the form of vegetativeorgans (Raunkiær 1934). In Saharan areas, thedecrease of hemicryptophyte numbers furthers thedevelopment of chamaephytes as the competitionfor soil moisture diminishes (Gamoun et al. 2010).Generally, the dominance of both chamaephytesand therophytes over other life forms seem to bea response to a hot dry climate, topographic vari-ations, human and animal interference (El-Demerdash et al. 1995; Neffar et al. 2013).
Regarding phanerophytes, their presence and theirnumbers in the arid environments is known to be low,except in the most favourable habitats which are thewadis and valleys. Their presence is reported all overarid rangelands except in Hammada deserts which arethe landscapes most exposed to wind and erosive factors(Shmida et al. 1986). Therefore, the presence of Acaciaraddiana in Hammada Debdeb seems to be accidentalconsidering the known distributional habitats of thespecies.
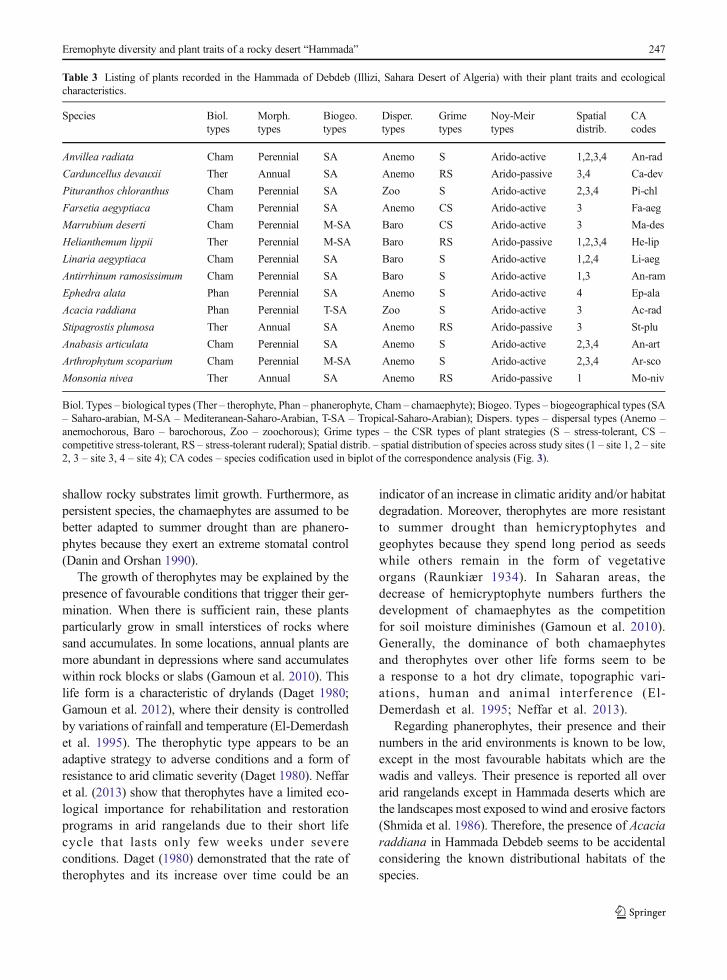
Table 3 Listing of plants recorded in the Hammada of Debdeb (Illizi, Sahara Desert of Algeria) with their plant traits and ecologicalcharacteristics.
Species Biol.types
Morph.types
Biogeo.types
Disper.types
Grimetypes
Noy-Meirtypes
Spatialdistrib.
CAcodes
Anvillea radiata Cham Perennial SA Anemo S Arido-active 1,2,3,4 An-rad
Carduncellus devauxii Ther Annual SA Anemo RS Arido-passive 3,4 Ca-dev
Pituranthos chloranthus Cham Perennial SA Zoo S Arido-active 2,3,4 Pi-chl
Farsetia aegyptiaca Cham Perennial SA Anemo CS Arido-active 3 Fa-aeg
Marrubium deserti Cham Perennial M-SA Baro CS Arido-active 3 Ma-des
Helianthemum lippii Ther Perennial M-SA Baro RS Arido-passive 1,2,3,4 He-lip
Linaria aegyptiaca Cham Perennial SA Baro S Arido-active 1,2,4 Li-aeg
Antirrhinum ramosissimum Cham Perennial SA Baro S Arido-active 1,3 An-ram
Ephedra alata Phan Perennial SA Anemo S Arido-active 4 Ep-ala
Acacia raddiana Phan Perennial T-SA Zoo S Arido-active 3 Ac-rad
Stipagrostis plumosa Ther Annual SA Anemo RS Arido-passive 3 St-plu
Anabasis articulata Cham Perennial SA Anemo S Arido-active 2,3,4 An-art
Arthrophytum scoparium Cham Perennial M-SA Anemo S Arido-active 2,3,4 Ar-sco
Monsonia nivea Ther Annual SA Anemo RS Arido-passive 1 Mo-niv
Biol. Types – biological types (Ther – therophyte, Phan – phanerophyte, Cham – chamaephyte); Biogeo. Types – biogeographical types (SA– Saharo-arabian, M-SA – Mediteranean-Saharo-Arabian, T-SA – Tropical-Saharo-Arabian); Dispers. types – dispersal types (Anemo –anemochorous, Baro – barochorous, Zoo – zoochorous); Grime types – the CSR types of plant strategies (S – stress-tolerant, CS –competitive stress-tolerant, RS – stress-tolerant ruderal); Spatial distrib. – spatial distribution of species across study sites (1 – site 1, 2 – site2, 3 – site 3, 4 – site 4); CA codes – species codification used in biplot of the correspondence analysis (Fig. 3).
Eremophyte diversity and plant traits of a rocky desert “Hammada” 247
Benabadji et al. (2007) highlight the impact of envi-ronmental factors on the evolution of life forms ofvegetation. The main interest of the biological spectrum‘life forms’ is that it reflects the structure of the vegeta-tion which translates at it term the surrounding environ-mental conditions (Aberlin and Daget 2003). Although
the therophytic strategy appears to be a major phenom-enon of degradation in drylands due to drought, thisdoes not apply on Hammada deserts whose presence islimited by edaphic characteristics of the substrate.Therophytes prefer sandy soils, which have higher re-tention properties than those of gravel or rocky
Fig. 2 Plant traits and phytogeographical types of the plant species identified at study sites of the rocky desert ‘Hammada’ of Debdeb (Illizi,Sahara Desert of Algeria).
248 L. Bradai et al.

substrates such as Hammadas (Olsvig-Whittaker et al.1983). It is noteworthy that the presence of annuals ismainly governed by moisture combined with adequatetemperatures.
The specific order of presence of phytogeographicaltypes in the Hammada is similar to the surveys carriedout by Bouallala (2013) in arid rangelands of WesternSahara Desert in Algeria. According to the classificationof Quézel (1978), the study area belongs to the Holarcticempire (Mesogean sub-empire), the Saharo-Arabian re-gion (Saharan sub-region) and the Northern Sahara do-main. In this large domain, several phytogeographicalelements of different origins exist and adapt to aridenvironmental conditions.
The dominance of Saharo-Arabian phytogeographictype in Hammada Debdeb corroborates results found inHammada deserts of South Morocco, where plant spe-cies with the Saharo-Arabian origin were dominant, andthe Mediterranean elements were present but with lowrate (Guinet and Sauvage 1954). Quézel (1995) attrib-uted such phytogeographical affinity of biodiversity tothe origins of theMediterranean flora that are influencedby climate changes, palaeogeographic variations andmajor tectonic movements.
In phytogeographical terms, it appears that the studyrocky desert is very little diversified. The low plantdiversity in such hostile environment is mainly due toclimate and substrate. Firstly, climate changes thatMediterranean Africa has undergone since theMiocene resulted in the migration of the tropical flora(Quézel 1960, 1983). Additionally, the current severeclimatic conditions are affecting both physiology anddistributional ranges of most xerophytes in hot desertsand are acting as a limiting factor for the establishmentof many other species. However, the current flora livingin the Sahara Desert is adapted to environmental condi-tions of aridity and drought, although it occupies aprominent place in vegetation cover, particularly inWadi beds and rocky/sandy soils (Bensaid et al. 1996).It is well known that the absence of strict-endemic taxaof the Hammada may be due to a lack of geographicbarriers of isolation (Bouallala 2013).
Wind-dispersed seeds were the most common dis-persal syndrome in the study flora (anemochorous, 57%). The dominance of this category is explained by thehigh frequency of winds (see climatic characterizationof study area). Indeed, the surface of Hammadas iscovered with rock fragments because the fine particleswere removed by wind and water. This obviously high-lights the important role of wind in the dispersion ofboth plants and particles of soil surface in Hammadadeserts. The low proportion of the zoochorous plant typeprobably returns to the low animal abundance in theseenvironments, which is generally represented by reptilesand certain insects because the latter are known to beimportant pollinators rather than good seed dispersers(Buchmann and Nabhan 2012). However, it is importantto emphasize that the diaspore may be dispersed by avariety of agents, making the classification of manyplants species into a single category on the basis of thedispersing agent almost impossible (van Rheede vanOudtshoorn and van Rooyen 1999).
Stable species coexistence depends upon reducedresource overlap, thus species develop adaptations todifferent forms of competition, abiotic stress or distur-bance (Grime 2006). From the point of view of Grime’sstrategies, the floristic composition of the studyHammada has a high rate of stress-tolerant taxa (68.8%). This is entirely logical given the climatic and soilconditions under which the plants are subjected. Indeed,Daget (1980) mentions that under such severe condi-tions of climate and soil, therophytic plants would liemuch better in stress-tolerant type strategies of Grime.
Fig. 3 Factorial bi-plot (F1 × F2) of the correspondence analysis(CA) applied to the distributions of plant species densities onsampled sites of rocky desert ‘Hammada’ of Debdeb in the Alge-rian Sahara Desert. (See Table 3 for species abbreviations). Plantspecies were abbreviated as the first two letters of the genusfollowed by the first three letters of the scientific name of thespecies.
Eremophyte diversity and plant traits of a rocky desert “Hammada” 249

Stress-tolerant ruderal species (RS) are 28.5 % of thetotal flora. The syndrome of stress-tolerant-ruderalityoccurs when disturbances persist, either in form of dras-tic climatic stresses or zoogenic-type disruptions mainlyrelated to the presence of livestock, which causes tram-pling of vegetation, uprooting of plants and/or pluckingof buds (Bonnet et al. 1999). Plant competitors (C) arepoorly represented (only two species) because they areadapted to high-productive environment, which is notthe case of rocky deserts. Furthermore, the occurrence ofRS plants may be a result of camel grazing inHammadas when herds transit between habitats. It isknown that annual plants use the RS strategy to avoidbeing grazed by a rapid phenological developmentwhereas ephemeral perennials apply this strategy toavoid grazing through morphological and/or biochemi-cal mechanisms of defence (Jauffret 2001).
The classification of Noy-Meir revealed the domi-nance of arido-active species (71.4 %) compared toarido-passives (28.6 %) in the rocky desert under study.The abundance of arido-active plants strongly decreaseswhen the environment is disturbed, as opposed to arido-passive plants that are abundant at all stages of degra-dation of a given habitat (Jauffret and Visser 2003).However, the study area including the Hammada land-scape is undisturbed and without any form of degrada-tion or human pressure. Once more, ephemeral charac-ters (arido-passive species) and persistent ones (arido-active species) represent thoroughly the two major strat-egies used by plants to withstand the dry season and aridconditions (Evenari et al. 1975), in particular throughspecific plant life forms like therophytes and chamae-phytes. Furthermore, the dominance of arido‐active spe-cies in the study area may be explained by the fact thatall arido‐passive plants are annuals, so their presencedepends on rainfall.
Conclusion
This study demonstrates the usefulness of plant traits inaiding the understanding of the organization of plantcommunities in extreme desert environments such asHammada deserts. The vegetation of the Hammadaunder study, characterized by the dominance of cham-aephytes followed by therophytes, is the expression ofthe arid conditions to which it is subject. The presence ofstress-tolerant and stress-tolerant ruderal species, espe-cially annuals, can be explained by current severe
climatic conditions rather than biotic factors, given theextreme aridity that plays a role of isolation barrier,preventing any plant immigration. Additionally, the func-tional attributes of ecological communities are actuallynot easy to investigate and understand under hyper-aridconditions, and in a more pronounced way in Hammadadeserts, given the rarity of the species on the one hand andseverity of climatic conditions on the other. This pointemphasizes the need to use well-adapted and customizedanalytical methods and sampling techniques with regardto the particularities and conditions of these habitats. Wehope this study may serve as a pioneer reference forfurther mid- and long-term investigations of the desertvegetation of North Africa.
Acknowledgements The authors express their sincere thanks toDr. Dylan W. Schwilk (Department of Biological Sciences, TexasTech University, USA) for having edited the English of ourmanuscript.
References
Aberlin JP, Daget P (2003) Establish and compare the life fromspectra of several plant groups. Rev Elev Med Vet Pays Trop56:57–61
Adam JG (1962) Itinéraires botaniques en Afrique occidentale.Flore et Végétation d'hiver de la Mauritanie Occidentale. LesPâturages: Inventaire des plantes signalées en Mauritanie. JAgr Trop Bot Appl 9:297–416
Allaby M (2008) Deserts (Ecosystem); Illustrations by RichardGarratt. Rev. ed. 337p
Benabadji N, Benmansour D, Bouazza M (2007) La flore de AinFezza dans l’ouest Algérien, Biodiversité et dynamique. SciTechnol 26:47–59
Benhouhou SS, Dargie TCD, Gilbert OL (2003) Vegetation asso-ciations in the Ougarta Mountains and dayas of the Guirhamada, Algerian Sahara. J Arid Environ 54:739–753
Bensaid S, Ait Mohand L, Echaib B (1996) Evolution spatio-temporelle des peuplements d'Acacia tortilis (Forsk.) Hayneraddiana (Savi) Brenan dans les monts Ougarta (Saharanord-occidental). Sécheresse 7:173–178
Bensizerara D, Chenchouni H, Bachir AS, Houhamdi M (2013)Ecological status interactions for assessing bird diversity inrelation to a heterogeneous landscape structure. Avian BiolRes 6:67–77
Bonnet V, Vidal E, Médail F, Tatoni T (1999) Analysediachronique des changements floristiques sur un archipelméditerranéen périurbain (Îles du Frioul, Marseille). RevEcol-Terre Vie 54:3–18
BouallalaM (2013) Etude floristique et nutritive spatio-temporelledes parcours camelins du Sahara Occidental algérien: Casdes régions de Béchar et Tindouf. PhD thesis, University ofOuargla, Algeria
250 L. Bradai et al.

Buchmann SL, Nabhan GP (2012) The forgotten pollinators.Island Press, Washington, DC
Burke A (2001) Classification and ordination of plant communi-ties of the Naukluft Mountains, Namibia. J Veg Sci 12:53–60
Chehma A, Djebar, MR, Hadjaiji F, Rouabeh L (2005) Étudefloristique spatio-temporelle des parcours sahariens du Sud-Est algérien. Sécheresse 16:275–285
Chehma A, Faye B, Bastienelli D (2010) Valeurs nutritionnellesdes plantes vivaces des parcours sahariens Algériens pourdromadaire. Fourrages 204:253–256
Chenchouni H (2012) Diversité floristique d’un lac du Bas-Saharaalgérien. Acta Bot Malacitana 37:33–44
Cowlishaw G, Davies JG (1997) Flora of the Pro-Namib DesertSwakop River catchment, Namibia: community classifica-tion and implications for desert vegetation sampling. J AridEnviron 36:271–290
Daget P (1980) Un élément actuel de la caractérisation du mondeméditerranéen: le climat.Naturalia MonspeliensaHS:101–126
Danin A, Orshan G (1990) The distribution of Raunkiaer lifeforms in Israel in relation to the environment. J Veg Sci 1:41–48
Dasti A, Agnew ADQ (1994) The vegetation of Cholistan andThai deserts, Pakistan. J Arid Environ 27:193–208
DeMartonne E (1925) Traité de Géographie Physique: 3 tomes. ParisEl-DemerdashMA, Hegazy AK, Zilay AM (1995) Vegetation-soil
relationships in Tihamah coastal plains of Jazan region, SaudiArabia. J Arid Environ 30:161–174
Evenari M, Schuze ED, Kappen L, Buschbom U, Lange OL(1975) Adaptive mechanisms in desert plants. In: VernbergF.J (Ed), Physiological Adaptation to the Environment.Cambridge University Press, pp 111–129
Faurie C, Ferra Ch, Medori P, Dévaux J, Hemptinne JL (2003)Ecologie: approche scientifique et pratique.Tec&Doc, Paris
Fox J (2005) The R Commander: A Basic Statistics GraphicalUser Interface to R. J Stat Softw 14:1–42
GamounM,BelgacemAO,Hanchi B, NeffatiM (2010)Variations dela diversité floristique en fonction du mode de gestion desparcours arides de la Tunisie méridionale. Ecol Medit 37:33–43
GamounM,OuledBelgacemA,Hanchi B, NeffatiM,Gillet F (2012)Effet du pâturage sur la diversité floristique des parcours aridesdu sud tunisien. Rev Ecol Terre Vie 67:271–282
Ghezlaoui B, Benabadji N, Benmansour D, Merzouk A (2011)Analyse des peuplements végétaux halophytes dans le ChottEl-Gharbi (Oranie-Algérie). Acta Bot Malacitana 36:113–124
Goldberg DE, Turner RM (1986) Vegetation change and plantdemography in permanent plots in the Sonoran Desert.Ecology 67:695–712
Goudie A (2002) Great warm deserts of the world: landscapesand evolution (Vol. 1). Oxford University Press
Gounot N (1969) Méthodes d’études quantitatives de lavégétation. Ed. Masson, Paris
Grime JP (2006) Plant strategies, vegetation processes, and eco-system properties. John Wiley & Sons, Sussex, UK
Guinet PH, Sauvage CH (1954) Les Hammadas sud marocaines.Botanique. Trav Inst Sci Chérifien 2:75–167
Huetz de Lemps A (1970) La végétation de la terre. Ed. Masson etCie, Paris
Jauffret S (2001) Validation et comparaison de divers indicateurs deschangements à long terme dans les écosystèmes méditerranéensarides: Application au suivi de la désertification dans le Sudtunisien. Doctoral Dissertation, Univ Aix-Marseille 3, France
Jauffret S, Visser M (2003) Assigning life-history traits to plantspecies to better qualify arid land degradation in PresaharianTunisia. J Arid Environ 55:1–28
Laity JJ (2009) Deserts and desert environments. John Wiley &Sons, Oxford
Maestre FT, Quero JL, Gotelli NJ, Escudero A, Ochoa V, Delgado-Baquerizo M, et al. (2012) Plant species richness and eco-system multifunctionality in global drylands. Science 335:214–218
Magurran AE (2004) Measuring biological diversity. Wiley-Blackwell, Malden, USA
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuildingcommunity ecology from functional traits. Trends Ecol Evol21:178–185
Médail F, Roche P, Tatoni T (1998) Functional groups inphytoecology: an application to the study of isolated plantcommunities inMediterranean France. Acta Oecol 19:263–274
Neffar S, Chenchouni H, Beddiar A, Redjel N (2013)Rehabilitation of degraded rangeland in drylands by Pricklypear (Opuntia ficus-indica L.) plantations: effect on soil andspontaneous vegetation. Ecol Balkan 5:63–83
Neffar S, Chenchouni H, Si Bachir A (2015) Floristic compositionand analysis of spontaneous vegetation of Sabkha Djendli inNorth-east Algeria. Plant Biosystems. doi:10.1080/11263504.2013.810181
NOM (2014) Rapport sur les données climatiques d’Illizi. Reportof the National Office of Meteorology ‘N.O.M.”, Illizi,Algeria
Noy-Meir I (1973) Desert ecosystems: environment and pro-ducers. Annu Rev Ecol S 4:25–51
Olsvig-Whittaker L, Shachak M, Yair A (1983) Vegetation pat-terns related to environmental factors in a Negev Desertwatershed. Vegetatio 54:153–165
Osman A, Al-Ghamdi F, Bawadekji A (2014) Floristic diversityand vegetation analysis of Wadi Arar: A typical desert Wadiof the Northern Border region of Saudi Arabia. Saudi J BiolSci 21:554–565
Ozenda P (2004) Flore et vegetation du Sahara. 3rd Edition. Ed.CNRS, Paris
Paradis G, Piazza C (2002) Contribution à l’étude de la flore et de lavégétation des îlots satellites de la Corse. 9e note: îlot deCapense (Centuri, Cap Corse). Le monde des plantes 477:1–6
Pijl, van der L (1982) Principles of dispersal in higher plants.Springer. Berlin, Heidelberg & New York.
Quézel P (1960) Flore et palynologie sahariennes. Quelques as-pec ts de leur signif icat ion biogéographique etpaléoclimatique. Bulletin IFAN 22:353–360
Quézel P (1965) La végétation du Sahara, du Tchad à laMauritanie. Ed. Masson, Paris
Quézel P (1978) Analysis of the flora of Mediterranean andSaharan Africa. Ann Missouri Bot Gard 65:479–534
Quézel P (1983) Flore et végétation de l'Afrique du nord, leursignification en fonction de l'origine, de l'évolution et desmigrations des flores et structures de végétation passées.Bothalia 14:411–416
Quézel P (1995) La flore du bassin méditerranéen: origine, mis enplace et endémisme. Ecol Medit 21:19–39
Quézel P, Santa S (1962) Nouvelle flore de l’Algérie et des régionsdésertiques méridionales. Vol 1. Ed. CNRS, Paris
Quézel P, Santa S (1963) Nouvelle flore de l’Algérie et des régionsdésertiques méridionales. Vol 2. Ed. CNRS, Paris
Eremophyte diversity and plant traits of a rocky desert “Hammada” 251

Quinn JA (2009) Desert Biomes. Greenwood Publishing,Portsmouth, NH, USA
Raunkiær C (1934) The life forms of plants and statistical plant.Geography Clarendon Press, Oxford
Rheede van Oudtshoorn, van K, Rooyen, van M (1999)Dispersalbiology of desert plants. Springer-Verlag, Heidelberg, Berlin
Shmida A, Evenari M, Noy-Meir I (1986) Hot desert ecosystems:an integrated view. In: Evenari M, Noy-Meir I, Goodall DW(eds). Ecosystems of the World, Vol. 12b. Hot Deserts andArid Shrublands. Elsevier, Amsterdam, pp 379–387
Vetaas OR (1993) Spatial and temporal vegetation changes along amoisture gradient in northern Sudan. Biotropica 25:164–175
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I,Garnier E (2007) Let the concept of trait be functional!.Oikos116:882–892
Walter H (1986) The Namib Desert. In: Evenari M, Noy-Meir I,Goodall DW (Eds), Ecosystems of the World: 12B Hot Desertsand Arid Shrublands. Elsevier, Amsterdam, pp 245–282
Whitford WG (2002) Ecology of desert system. Academic,London
252 L. Bradai et al.





























