Embed Size (px)
Citation preview


AN INTRODUCTION
N E U R O L O G Y
C. JUDSON HERRICK
PROFESSOR OF NE U ROLOGY IN THE UNIVE RSITY O F CHICAGO
ILLUSTRATED
PHILADELPHIA AND LONDON
W. B. SAUNDERS COMPANY1916

Copyright , 1 9 1 5 , by W. B . Saunders Company
l ows$
W. B . SAUNDERS COMPANY

PREFACE
THERE are two groups of functions perform ed by the nervoussystem wh ich are of general interest ; these are, first , the physiological adjustm ent of the body as a who le to its env ironm entand the correlation of the activities of its organs am ong themselves
, and , in the second place, the so- called h igher functionsof the cerebral cortex related to the consc ious life. The secondof these groups of functions cannot be stud ied apart from the
first,for the enti re conscious experience depends for its materials
upon the content of sense, that is , upon the sensory data receivedby the lower brain centers and transm itted through them to the
cerebral cortex . Since the organ ization of these lower centers isextrem ely com plex , and since even the sim plest nervous processes involve the interaction and cooperat ion of several of thesem echanism s
,it fol lows that an understanding of the workings
of any part of the nervous system requires the m astery of a largeam ount of rather intricate anatom ical detail .Fortunately
,the know ledge of the precautions wh ich m ust be
observed in order to m aintain the nervous system in healthyworking order is not d ifficu lt of acqu isition (though surprisinglyfew people seem to have gained it) , just as any one can learn tooperate an autom ob i le, even though quite ignorant of the engineering problem s involved in its design and construction . In
formation regard ing these matters of practical hygiene is readi lyavai lab le,‘and it is not the primary purpose of th is book to sup
1 GUL ICK , LUTHE R H . 1907 . The E fficient Life,New York .
GU L ICK, LUTHE R H . 1908 . Mind and Work,N ew York .
J EWETT FRANCISGU LICK , 1898 . Control of Body and Mind,N ew York ,
Ginn Co Adapte d for use in the gra ded schools .LUGA RO ,E . 1909 . Modern Problem s in Psychiatry
,Manchester
Univers ity Press . A book wri t ten espec ially for physicians,but full of
stim ulating ideas for every educated reader .
S'
rrL E s, P . G . 19 14 . The N ervous System and Its Conservation,
Philadelphia, W . B . Saunders Company .

12 PREFACE
ply it . But to understand the actual inner operation of thenervous m echanism s is a much m ore difficult m atter
, and th isknow ledge cannot be acquired without arduous and sustainedstudy of the pecul iar form relations of the nervous organs andtheir complex interconnections ; and inform ation of th is sort isindispensable for a grasp of the principles of nervous organization
,and especial ly for an intelligent treatm ent of nervous d is
eafim.
The study of neurology is , therefore, intrins ically d ifficultif one is to advance beyond its m ost superfic ial phases ; the m oreso if the student is not wel l grounded in general b iology and at
least the elem ents of the general anatom ical structure of the vertebrate body . To these inherent d ifficulties there is added a
purely artificial obstac le in the form of a cum bersom e and con
fused term inology wh ich has grown up during several centuries ofanatom ical study of the brain,
in the early stages of wh ich littleor no com prehension of the functional significance of the partsd iscovered was possible, and fanc iful or bizarre nam es were givenwi thout reference to the mutual relationsh ip of parts .The prob lem s wh ich at present ch iefly occupy the attention of
neurologists are of two sorts—first, to discover the regionallocal ization w ith in the nervous system of the nerve- cel ls andfibers wh ich serve particular types of function or
,briefly
, arch itecture, and second , to d iscover the chem ical or other changeswh ich take place during the process of nervous function,
that is ,the metabol ism of the nervous tissues . The first of these problem s is at present further advanced than the second ; the largerpart of th is work is , therefore, devoted to a description of arch itectural relations . Without a knowledge of these relations ,m oreover , the prob lem s of metabo lism are, in large measure,
m eaningless .I t is im possible to understand clearly the form of the brain,
and especial ly the relations of its internal structures , from verbaldescriptions m erely . Pictorial i l lustrations and the various brainm odels wh ich are on the market are of great assistance ; but actual laboratory experience in d issecting the brain and
,if possi
ble, the study of m icroscopic preparations of selected parts ofit are ind ispensable for a thorough mastery of the subject .The brains of the sheep , d og, and cat are easily obtained , and

PREFACE 13
are so sim i lar to the human brain in all respects,save the
smal ler relative size of the cerebral cortex , that they can readi lybe used for such stud ies . Before dissection the brain should becarefully rem oved from the skul l and hardened by imm ersion fora few days in a solution Of formal in (to be obtained at anydrugstore and d i luted w ith water in the proportion of one partformalin to nine parts water) . Several neuro logical laboratory
guides have been published , and one Of these should be followedin the d issection .
1
This work is designed as an introduction in a l iteral sense .
Several very excel lent manuals and atlases Of neurology are
avai lable, and to these the reader is referred for the i l lustrationsand m ore detai led descriptions necessary to com plete the ratherschem atic outline here presented . The larger medical textbooks Of anatom y and physiology are, however, often very d i fficult for the beginner , ch iefly on account Of the lack Of correlationOf the structures described and their functions . Th is littlebook has been prepared in the hope that it w i l l help the studentto learn to organize h is know ledge in definite functional patterns earlier in h is work than is often the case, and to appreciatethe significance Of the nervous system as a working m echanismfrom the beginning of his study .
The structure and functions Of the nervous system are Of
interest to students in several d ifferent field s—med ic ine, psy
chology,sociology , education
, general zoology , com parativeanatom y
,and physiology , am ong others . The view- points and
special requirem ents of these various groups are,of course
,
different ; nevertheless the fundam ental principles of nervousstructure and function are the sam e
,no m atter in what
field the princ iples are applied,and the aim here has been ' to
l BU RRHO LDE R , J . F . 1 9 12 . The Anatomy of the Brain , Second Edition,Chicago , G . P . E ngelhard CO . (Dissec t ion of the brain of the sheep . )FISKE , E . W . 19 13 . An E lem entary Study of the Brain Based on theDissection of the Brain of the Sheep , N ew York , The Macm illan Com pany .
HARDE STY, I . 1902 . N eurologicalTechnique, The Uni versity of ChicagoPress . (Dissection of the human brain by m eans O f transverse grosssection)s, m ethods O f m icroscop i c preparation, and l is ts of neurological
term sHE RRICK , C . J UDSO N and CROSBY , E L IZAB ETH . 19 15 . A Laboratory
Outline i n N eurology , privately printed by the authors at the University O fChicago . (Dissec t ion of the d ogfish , sheep , and human brains , and d ircetions for study of prepared m i croscop i c sections Of the human brain . )

14 PRE FACE
present these principles rather than any detai led applicationOf them . In the selection Of subject matter and m ode Of
treatm ent the author has been fortunate in hav ing the adviceOf m any experienced teachers in several Of these fields
,who
have read the manuscript Of th is work or Of selected chaptersand whose suggestions have contributed greatly to its value .E spec ial acknowlegem ent Of generous assistance Of th is sortshould be made to Doctors G . W . Bartlem ez , R . R . Bensley ,Harvey A . Carr
,C . M . Ch i ld
,G . E . Coghi l l , Mabel R . Fernald
,
Joseph W . Hayes,Mary Stevens Hayes
,F . L . Landacre
,John
T . McManis, and R . E . Sheldon .
The materials presented in th is book are arranged in three
groups : ( 1 ) Chapters I to VI I d iscuss the m ore general neurological topics ; (2) Chapters VII I toXVI I I com prise a brief account Ofthe form of the nervous system and the functional significance Ofits ch ief subd ivisions in general , fol lowed by a rev iew O f the arch itectural relations Of the m ore im portant functional system s ; (3)Chapters XIX to XXI are devoted to the cerebral cortex and itsfunctions . Readers whose ch ief interest lies in the general neurological questions m ay om it m uch of the detai l com prisedw ith in the second group Of chapters or use these for referenceonly . TO fac i l itate ready reference the general index has beenprepared with espec ial care
,and w ith it is com b ined a brief
glossary Of som e m ore comm only used technical term s . In the
text some of the m ore special topics , which may be om itted ifa briefer presentation is desired , are printed in sm al ler type.
C . J U DSON HE RRICK .
CHICAGO , ILL .,
October, 1 9 15 .

CO NTENTS
CHAPTER IBIOLOGICAL INTRODUCTION
CHAPTER IITHE N E RVOUS FUNCTIO NS
CHAPTER III'
THE N EU RON
CHAPTE R IVTHE R EFLEX C IRCUITS
CHAPTER v
THE R ECE PTORS AND E FFECTO RS
CHAPTE R VITHE GENE RA L PHYSIO LOGY O F THE N E RVOUS SYSTEM
CHAPTE R VIITHE G EN E RA L ANATOMY AND SU BDIVISIO N O F THE N E RVOUS SYSTEM 106
CHAPTE R VIIITHE SPINA L CORD A ND ITS N E RVE S
CHAPTER IXTHE M EDULLA OB LO NGATA A ND CE RE BE LL U M 143
CHAPTER XTHE CE REBRUM 160

16 CONTE NTS
CHAPTER XITHE GEN E RA L SOMATIC SYSTE MS OF CO NDUCTION PATHS
CHAPTER XIITHE VESTIB ULAR APPA RATUS A ND CE RE BE LLUM .
CHAPTER XIIITHE AUDITO RY APPA RATUS
CHAPTER XIVTHE VISUA L APPA RATUS .
CHAPTER XVTHE OLFACTORY APPA RATUS .
CHAPTER XVITHE SYMPATHETIC N E RVOUS SYSTEM
CHAPTER XVIITHE VISCE RAL AND GUSTATO RY A PPA RATUS 234
CHAPTER XVIIIPAIN AND PLE ASURE . 249
CHAPTE R XIXTHE STRUCTURE OF THE CE RE B RAL CO RTEx
CHAPTE R XX
THE FUNCTIONS O F THE CE RE BRAL CO RTEx .
CHAPTER XXITHE E VO LUTIO N AND SIGN IFICANCE O F THE CERE BRAL CoRTEx .
INDEX AND G LOSSARY

INTRODUCTION TO NEURO LOGY
CHAPTER I
B IO LO G ICAL INTR ODUCTION
THE l iving body is a little world set in the m idst Of a largerworld . I t leads in no sense an independent l ife, but its continued welfare is conditioned upon a nicely balanced adjust
ment between its own inner act ivities and those of surroundingnature, som e of wh ich are beneficial and som e harm ful . The
great prob lem Of neurology is the determ ination of the exactpart wh ich the nervous system plays in this adjustm ent .This prob lem is by no m eans sim ple. The search for its
solution w i l l lead us, in the first place,back to an exam ination
Of som e of the fundamental properties of the ' sim plest livingsubstance, of protoplasm itself ; and in the last analysis it w i llinvo lve a consideration of the highest m ental capacities Of thehuman race and Of the physiological apparatus through wh ichthese capacities com e to expression . We shall first take up thenature of th is adjustm ent on the lower biological levels .All Of the infinitely d iverse form s Of l iving things have cer
tain points in comm on,so that one rarely has any doubt
whether a given Object is alive or dead . N evertheless , theprec ise definition of l ife itself proves very difficult . HerbertSpencer
,in h is “Principles of B iology, after many pages
Of close argum ent and rather form idable verbal gymnastics ,arrived at th is formula : Life is “
the definite com bination Of
heterogeneous changes , both simultaneous and successive, incorrespondence w ith external coexistences and sequences” ;or
, more briefly,
“The continuous adjustm ent Of internal re
lations to external relations .” A som ewhat sim i lar idea was
2 17

18 INTRODUCTION TO NEUROLOGY
subsequently more sim ply expressed by the late C . L . Herrickin the proposition.“Life is the correlation of physical forcesfor the conservation Of the ind ividual” ; and this, in turn,
may
be cast in the m ore general form ,Life is a system Of forces
maintained by a continuous interchange of energy between thesystem and its environm ent, these forces being so correlatedas to conserve the identity Of the system as an individual andto propagate it . A certain m easure Of m od ifiability in the character of the system ,
w ithout loss Of its individual ity,is not ex
cluded .
NO one Of these definitions , or any other which has been sug
gested , is ful ly satisfactory ; but biologists generally agree thatthe comm on characteristics of living beings can best be ex
pressed in the present state of our know ledge in term s Of theiractions
,their behavior . The properties comm only ascribed to
any Object are in last analysis nam es for its behavior,and the
so- called vital properties are very special form s of energy transform ation.
N ow ,the ch ief difference between a corpse and a living body
consists in the fact that the forces of surrounding nature tendto the d isintegration Of the dead body, wh ile in the l iving bodythese forces are uti lized for its upbui lding. If
,then
,the vital
process is essential ly a Special type Of mutual interaction between the bodi ly mechanism and the forces of the surroundingworld
,Of the correspondence between the organism and the
environment,to use the Spencerian phrase, it follows that the
living body cannot be studied by itself alone. Quite the con
trary, the analysis Of the environm ental forces upon wh ich thelife of the body depends and of the parts Of the body itselfin thei r relations to these external forces is the very kernel ofthe problem Of l ife .
The m easure Of the fulness Of life in any organism is twofold . In the first place
,the life is m easured by the am ount Of
energy wh ich the organism can assim i late from surroundingnature and incorporate into its own organization . This entersthe body ch iefly in the form Of chem ical potential energy in foodeaten, air breathed , and so on ,
and can be quantitatively d eterm ined and stated in the form of standard units Of energy,such as calories or foot-pounds of work . This measures the

BIOLOGICAL INTRODUCTION 19
working capacity Of the mach ine,but gives little insight into
the real value Of the work done. In the second place,l ife may
be m easured in term s Of the extensity or number and d iversityOf environmental relations . ~ Th is takes account Of the rangeor working d istance of the organi zation and
,in general , of the
efficiency Of the work done. For evidently the organism wh ichhas few and sim ple relations w ith the environm ent
,so that it
can adjust itself to only a sm al l range of external conditions,
is less efficient than one wh ich has m any diverse relationsh ipsand an extensive series Of possible adjustm ents
,even though the
actual am ount of energy expended may be vastly greater in theform er than in the latter case. The first Of these standards is atolerably satisfactory m easure Of the vegetative functions ofthe body
,som etim es less happily term ed the “organic functions .
”
We have no word in comm on use which covers precisely thegroup Of activities embraced by our second standard Of m easurem ent
,though the term s “animal functions
,
” “somatic orexteroceptive activities” are som etimes used in about th issense.
Let us now endeavor to illustrate the last topic a little m oreconcretely . We are standing on a h i lltop overlooking a
meadow , through wh ich‘ runs a m ountain brook
,and beyond
the valley is another range Of rugged h i lls . In the fence- cornernear us is a patch of daisies and clover w ith a honey-bee buzz ingfrom flower to flower . A plowboy is crossing the field
,and at
our elbow an artistic friend is busy w ith sketch ing pad and
brushes . Here are four th ings wh ich have this at least incomm on
,that they are al ive—d aisy, bee, plowboy , artist .
There can be no doubt about their vitality,but how differently
they respond to the sunshine,the rain
,and the other forces of
nature.
The daisy expands in the vivifying l ight Of the summer sun,
the energy Of whose actinic rays is used to bui ld up living protoplasm and vegetable fiber from the inert substances of air andsoi l . Its vitality
, m easured in term s of energy transforma
tion,is great ; yet how lim ited its range Of life, how helpless -in
the face Of the storm s Of adversity wh ich are sure to buffet it .R ooted to its station
,it can only assim i late what food is brought
to it and it cannot flee from scorch ing w ind or b lighting frost .

20 INTRODUCTION To N EUROLOGY
The honey-bee leads a m ore free and varied l ife. Instead ofpassively and blind ly waiting for such bane or b lessing as fatemay bring, she hurries forth , strong Of wing and with sensesalert , to gather the dai ly m easure Of honey and pollen . The
senses of touch , sight , and smel l open realm s Of nature foreverc losed to the plant, and enable her to seek food in new fieldswhen the local supply is exhausted , as wel l as to avoid enem iesand m isfortunes . With the approach Of the storm ,
she fl ies toshelter in a hom e which she and her sisters have prepared withconsumm ate ski ll . Yet in th is provision for the future in h iveand wel l- stocked honeycomb there is little evidence Of intelli
gent foresight or rational understanding of the purposes forwh ich they work . Though so m uch m ore hi gh ly organized thanthe plant
,the honey-bee is to a very large extent bl indly fol low
ing out the inborn im pulses of her hereditary organization and
she has no clear understanding Of what she does, much less whyshe does it . There is som e evidence Of intelligent adaptation inher behavior
,but the part played by this factor in her li fe as a
who le is probably very sm al l com pared w ith the blind inbo rnim pulses wh ich dom inate m ost of her activities . Like the plant
,
the bee’s reactions are determ ined ch iefly by the past evolutionary h istory Of the spec ies , wh ich has shaped the innateorganization of the body and fixed its typical m odes Of response to stimulation. But the bee l ives m uch m ore in the
present than does the plant ; that is, she can vary her behaviormuch m ore widely in response to the needs Of the m om ent . As
for the future she knows naught Of it .The farmer s boy wh istles as he goes about h is work . He
,
too, has a certain innate endowm ent , including the wholerange of h is vegetative functions , together with an instinctivelove Of sport and m any other inborn aptitudes . Th is is hisinh eritance from the past . By these instincts and appetites heis , as Dewey says
,pushed from beh ind” through the per
form ance Of m any b lindly im pulsive acts . He is a creature Of
the present, too, his whole nature overflowing with the j oy Of
living. But he also looks into the future and hastens throughthe daily tasks that he may Obtain the coveted hour of sunsetto fish in the brook . He fli cks Off the heads Of the daisies withh is wh ip- stock and remarks in passing,
“This meadow is

BIOLO GICAL INTRODUCTION 21
choking up with wh ite-weed . The boss wi ll have to plow it upnext year and replant it .” The extraord inary natural beautyOf the place is, however, unnoticed am id the round of dai lywork and sim ple pleasure.
The art ist looks out upon the same scene,but through what
different eyes $ The mass of wh ite daisies and the rocky knol lbeyond ruddy w ith sheep sorrel suggest to him no waste Of
valuable pasture land,but a harm ony of color and grace Of form
upon wh ich he feasts h is soul . The esthetic delights of theforest
, the sky, the brook , and the overhanging crag beyond arefor him unm ixed w ith any uti litarian motive.
Each Of these four organism s occupies , in one sense, thesam e environm ent ; but it is evident that the factors of thisenvi ronment w ith wh ich each com es into active Vi tal relationsare immeasurab ly different . They correspond with or are at
tuned to quite different energy com plexes, though the cor
respondence or interaction is very real in each case. Th is hasbeen stated very sim ply by Dr. Jennings when he says thatevery spec ies Of organism has its characteristic “action system ,
”
i . e., a habitual m ode of reaction to its environm ent which is
determ ined whol ly or in part by its inherited organization .
E very animal and every plant has,accord ingly,
‘
a definiteseries Of characteristic m ovem ents wh ich it can m ake in re
sponse to external stim ulation . This is all that Jennings m eansby the “
action system . We hum ans are no exception to th isrule Of life. We m ove along in a m ore or less stereotyped way,through m ore or less fam i liar grooves, in our dai ly work .
Much Of this work is routine, done about as m echanically as
the flower unfolds its petals to the m orning sun or the honeybee gathers in her store of honey . Th is is our action system .
Of course, we have m uch else to do besides th is routine, and ouractual value to the community is in largemeasure determ ined byour ab i lity to vary th is routine in adaptation to new situationsas they arise. Even the daisy has a l ittle Of th is capac ity forindependently variable action ; the insect has m ore ; and m an
’spreem inence in the world is d ue primari ly to h is larger powersOf adapting h is reactions not only to the needs of the moment ,but to probab le future contingencies , i . e.
, Of varying h isinborn action system by intel ligently d irected choices .

2 INTRODUCTION To NEUROLOGY
Th is d istinction between the b lind working Of a stereotypedaction system whose character is determ ined by the inheritedbodi ly structure
,on the one hand , and ind ividual ly acquired
variab le adaptive actions (wh ich may or m ay not be intelli
gently performed) , On the other hand , is very fundamental, and
we shal l have occasion to return to it . Most animal activitiescontain both Of these factors
,and it is Often very difficult
to analyze a given exam ple of behavior into its elem ents,but the d istinction is nevertheless im portant . Plant l ife ischaracterized by the dom inance of invariable types Of reactionwh ich are determ ined by innate structure ; these in their m ostelem entary form s give us, in fact , the so- cal led vegetative functions . These sam e functions predom inate in the lowest anim alsalso ; but in the h igher animals
,as we shal l see
,there are two
rather distinct l ines of evo lutionary advance . In one l ine theinnate stereotyped functions are very h igh ly specialized , leadingup to a com plex instinctive m ode Of l ife ; in the other line thesefunctions are subordinated to a h igher developm ent Of the ind ividual ly acquired variable functions
,leading up to the intelli
gence and doci l ity of the h igher mammals,including the human
race.
The d istinction between plants and anim als is very difficultto draw and
,in fact
,there are numerous groups Of organism s
wh ich at the present time occupy an am biguous position, suchas the slim e m olds . The botanists claim them and cal l themMyxomycetes ; the zoOlogists also describe them under the nam e
Mycetozoa ; sti l l other naturalists frankly give up the problemand assign them to an interm ediate kingdom ,
neither vegetablenor animal
,wh ich they call the Protista. As chi ldren we prob
ab ly considered the ch ief d istinction between plants and ani
mals to be the abi lity Of the latter to m ove freely about ; but oneOf the first lessons in our elem entary biology was the correctionOf th is notion by the study Of sedentary animals and m oti leplants . N evertheless
,I fancy that in the broad view the ch i ld
ish idea has the root of the matter in it . The plants and sedentary animals m ay have their vegetative functions of internaladjustm ent never so h igh ly special ized and yet remain relatively low in the bio logical scale because thei r relations w iththe envi ronm ent are necessari ly lim ited to the smal l ci rcle w ithin

BIOLOGICAL INTRODUCTION 23
wh ich they first take root,whereas the power of locom otion
carries w ith it , at least potential ly, the abi lity to choose betweenmany m ore envi ronm ental factors . I t is only the free-m ovinganimals that have anyth ing to gain by looking ahead in the
world,and here only do we find well- developed distance recep
tors, i . e.,sense organs adapted to respond to im pressions from
Objects rem ote from the body . And the d istance receptors , aswe shall see, have dom inated the evolution of the nervous system in vertebrates and determ ined the l ines it should fol low .
The net result of th is d iscuss ion can be briefly stated . Thed ifferences between various kinds Of organism s are, in the m ain
,
incidental to the extent and character Of their relations with theforces Of surrounding nature. A species wh ich can adjust itselfto few elements Of its environm ent we cal l low ; one that canadapt itself to a w ide range Of envi ronm ental cond itions in a
great variety of ways we cal l higher. The suprem acy Of thehum an race is directly d ue to our capacity for d iversified l iving.
If man finds h im self in an unfavorab le cl im ate,he may either
m ove to a more congenial local ity or adapt h is m ode Of life byartificial aids
,such as cloth ing, houses, and fire. And in these
adaptations he is not l im ited to a narrow range of inheritedinstincts , l ike the h ive Of bees , but h is greater powers of Observation and reflection enable him to discover the general uniform ities Of natural process (he cal ls these laws of nature) andthus to forecast future events
'
and prepare h im self for them in
telligently . In other words , to return to our original point Ofview
,our advantage in the struggle for existence l ies in our
ab i lity to correlate our bod ily activities w ith a wide range Of
natural forces so as to make use of these forces for our good ratherthan our hurt . (Of course, it should be borne in m ind that th isform ula makes no pretense Of being an exhaustive account Ofhuman faculty ; but only that, in so far as b iological evolutionaryfactors have Operated in the human realm ,
they act in accordance with th is principle.) The apparatus by which these external adjustm ents are effected and by wh ich the inner parts of thebody are kept in working order is the nervous system .

CHAPTER I I
THE NERVOUS FUN CTIONS
THE body is com posed of organs and tissues , the organs beingparts w ith particular functions to perform and the tissues beingthe cel lular fabric Of wh ich the organs are com posed . Thetissues (wh ich m ust be stud ied m icroscopically) are class ified
,
som etim es in accordance with the general functions wh ich theyserve, such as the nervous and m uscular tissues
,and som etim es
w ith reference to the form s and arrangements of their com ponent cel ls . An i llustration Of the latter m ethod Of treatm ent isfurnished by the epithel ial tissues
,wh ich are th in sheets Of
cel ls,som etim es arranged in one layer (sim ple epithelia) , som e
tim es in several layers (stratified epithelia) . E pithelial tissuesmay perform the m ost d iverse functions .All l iving substance (protoplasm ) possesses in some m easure
the d istinctive nervous functions Of sensitivity and conductivity,that is
,it responds in a characteristic fash ion to certain exter
nal forces (stim uli) , and when thus stimulated at one point them ovem ent or other response may be effected by som e rem otepart . This last feature im plies that som e form Of energy isconducted from the site of the stim ulus to the part m oved .
O rdinary protoplasm also possesses the power Of correlation,
that is,of com b ining a num ber of individual reactions to stimu
lation in diverse special adjustm ents .The one- celled animals and all plants lack the nervous sys
tem ent irely ; nevertheless they are ab le to m ake h igh ly com plexadjustm ents . The leaves
,roots
, and stem s Of the h igher plantshave individual functions wh ich are
,however, bound together
or integrated into a very perfect unity. In animals, as con
trasted with plants , we see a further differentiation Of parts ofthe body for special functions , and at the same tim e a m ore perfeet correlation of part w ith part and integration Of the wholefor rapid and diversified reactions of the entire body . The
24

26 INTRODUCTION TO N EUROLOGY
pulse outward from the center to (5 ) the effector apparatus,
consisting Of the organs of response (m uscles, glands) and the
term inals Of the efferent nerves upon them .
N O part Of the nervous system has any significance apart fromthe peripheral receptor and effector apparatus w ith which it isfunctionally related . Th is is true not only Of the nervous m echanism Of all physiological functions, but even O f the centers concerned w ith the h ighest manifestations of thought and feeling of
wh ich we are capable, for the m ost abstract m ental processesuse as their necessary instruments the data Of sensory experiencedirectly or indirectly
,and in m any , if not all
,cases are inti
mately bound up w ith som e form Of peripheral expression .
The neurologist’s problem is to d isentangle the inconceivab ly
com plex interrelations Of the nerve-fibers which serve all the
manifo ld functions Of adjustm ent Of internal and external relations ; to trace each functional system of fibers from its appro
priate receptive apparatus (sense organ) to the centers Of correlation ; to analyze the innum erable nervous pathways by whichthese centers are connected w ith each other (correlation tracts) ;and
,final ly
,to trace the courses taken by all outgoing im pulses
from these correlation centers to the peripheral organs Of response (m usc les
, glands, etc ., or
,collectively
,the effectors) .
Th is is no sim ple task . I f it were possib le to find an educatedman who knew noth ing Of electricity and had never heard of a
telegraph or telephone, and if th is m an were assigned the dutyOf m aking an investigation Of the telegraph and telephone system s O f a great c ity w ithout any outside assistance whatever
,
and Of preparing a report upon all the physical equipm ent w ithdetailed m aps Of all stations and c ircuits and with an explanation Of the m ethod of operation of every part
,h is task would be
sim ple com pared wi th the prob lem of the neurologists . Thehuman cerebral cortex alone contains som e 9280 m i ll ion nervecells
,m ost of wh ich are provided w ith long nerve—fibers wh ich
stretch away for great d istances and branch in different d irections , thus connecting each cel l w ith m any different nervecenters . The total num ber O f possible nervous pathways is
,
therefore,inconceivably great .
Fortunately for the neurologists , these interconnecting ner
vous pathways do not run at random ; but, just as the wires

THE NERVOUS FUNCTIONS 27
entering a telephone exchange are gathered together in greatcables and distributed to the switchboards in accordance wi th acarefully elaborated system ,
SO in the body nerve-fibers of likefunction tend to run together in separate nerves or w ith in thebrain in separate bundles cal led tracts . N otwithstand ing thecom plexi ty of organization of the nervous organs, the larger andm ore im portant functional system s Of nervous pathways havebeen successfully analyzed , and the courses of nervous dischargefrom the various receptors to the appropriate centers Of adjustm ent
,and from these (after manifold correlations with other sys
tem s) to the organs of response, are fairly wel l known . The
acquisition of th is knowledge has required several centuries Ofpainstaking anatom ical and physiological study, and muchremains yet to be done.
The external form s Of the brain and other parts of the nervoussystem are dependent m ainly upon the arrangements Of the
nerve- cel ls of which they are com posed (for the characteristics Ofthese cel ls see Chapter I I I ) , and these arrangem ents
,in turn,
are correlated w ith the functions to be performed . The functional connections Of the nerve- cells can be investigated best bythe m icroscopical study of the tissues com b ined w ith physiological experim entation . From th is it fol lows that the study Of the
gross anatom y, the m icroscopical anatom y (histology) , and thephysiology Of the nervous system should go hand in hand so faras th is is practicable.
A study of the com parative anatomy Of the nervous systemshows that its form is always correlated w ith the behav ior of theanim al possessing it . The sim plest form Of nervous system con
sists Of a diffuse network Of nerve- cells and connecting fibersdistributed am ong the other tissues Of the body . Such a ner
vous system is found in some jelly-fishes and in parts Of thesym pathetic nervous system of h igher animals . Animals whichpossess this d iffuse type of nervous system can perform onlyvery simple acts
,ch iefly total m ovements of the whole body
or general movem ents Of large parts Of it , with relatively smal lcapacity for refined activities requiring the cooperation ofmany d ifferent organs . But even the lowest animals wh ichpossess nerves Show a tendency for the nervous net to be con
d ensed in som e regions for the general control of the activities
28 INTRODUCTION TO NEUROLOGY
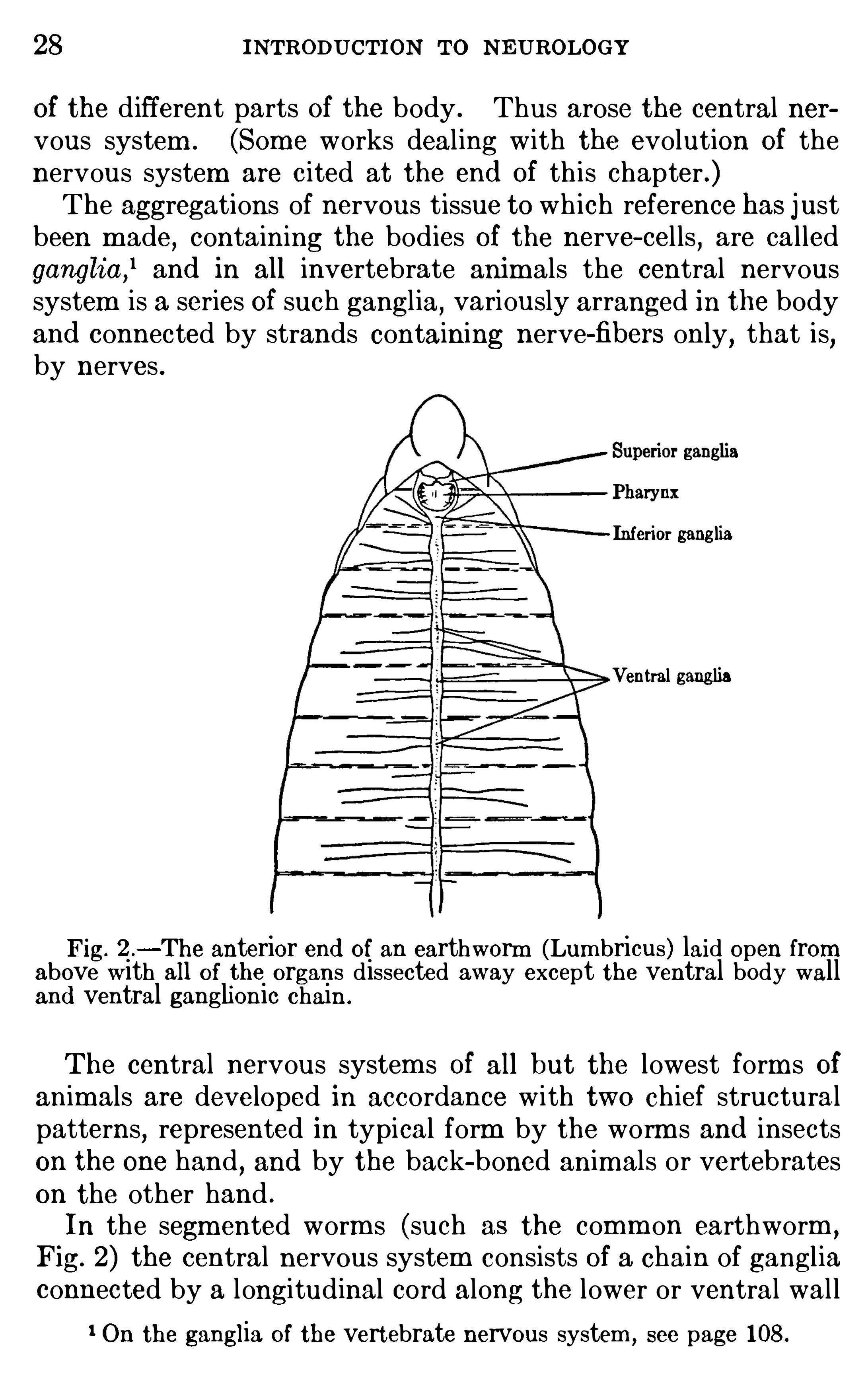
Of the d ifferent parts Of the body . Thus arose the central nervous system . (Some works deal ing with the evolution Of thenervous system are c ited at the end Of th is chapter .)The aggregations of nervous tissue to wh ich reference has just
been m ade, containing the bodies Of the nerve- cel ls,are called
ganglia,land in all invertebrate anim als the central nervous
system is a series Of such gangl ia,variously arranged in the body
and connected by strands containing nerve-fibers only,that is
,
by nerves .
Inferior ganglia
Ventral ganglia
Fig. 2 .—The anterior end of an earthworm (Lumbricus) laid open from
above with all of the organs dissected away except the ventral body walland ventral ganglionic chain .
The central nervous system s Of all but the lowest form s Ofanimals are developed in accordance with two chief structuralpatterns
,represented in typical form by the worm s and insects
on the one hand , and by the back-boned animals or vertebrateson the other hand .
In the segm ented worm s (such as the comm on earthworm ,
Fig. 2) the central nervous system consists of a chain of gangl iaconnected by a longitudinal cord along the lower or ventral wal l
1 On the ganglia of the vertebrate nervous system , see page 108.

THE NE RVOUS FUNCTIONS 29
of the body. Each of these ganglia is connected by m eans Ofperipheral nerves w ith the Skin and muscles of its own segm ent,and each j oint Of the body with its contained ganglion (ventral
ganglion) has a certain m easure of physio logical independence SOthat it can act as a unit . Th is is a typical segm ented nervoussystem . At the head end Of the body the ventral ganglionicchain d ivi des around the pharynx and m outh
,and there are
enlarged ganglia above and below the pharynx . The superi or
ganglia (supra-esophageal ganglia) are sometim es cal led thebrain
,and th is organ dom inates the local activi ties of the several
segments,enab ling the anim al to react as a whole to external
influences .The nervous system s of crustaceans (crabs and their all ies) ,
spiders,and insects have been derived from the type just
described . In these animals the segm ents Of the body are m oreor less united in three groups, constituting respectively the head,thorax
,and abdom en,
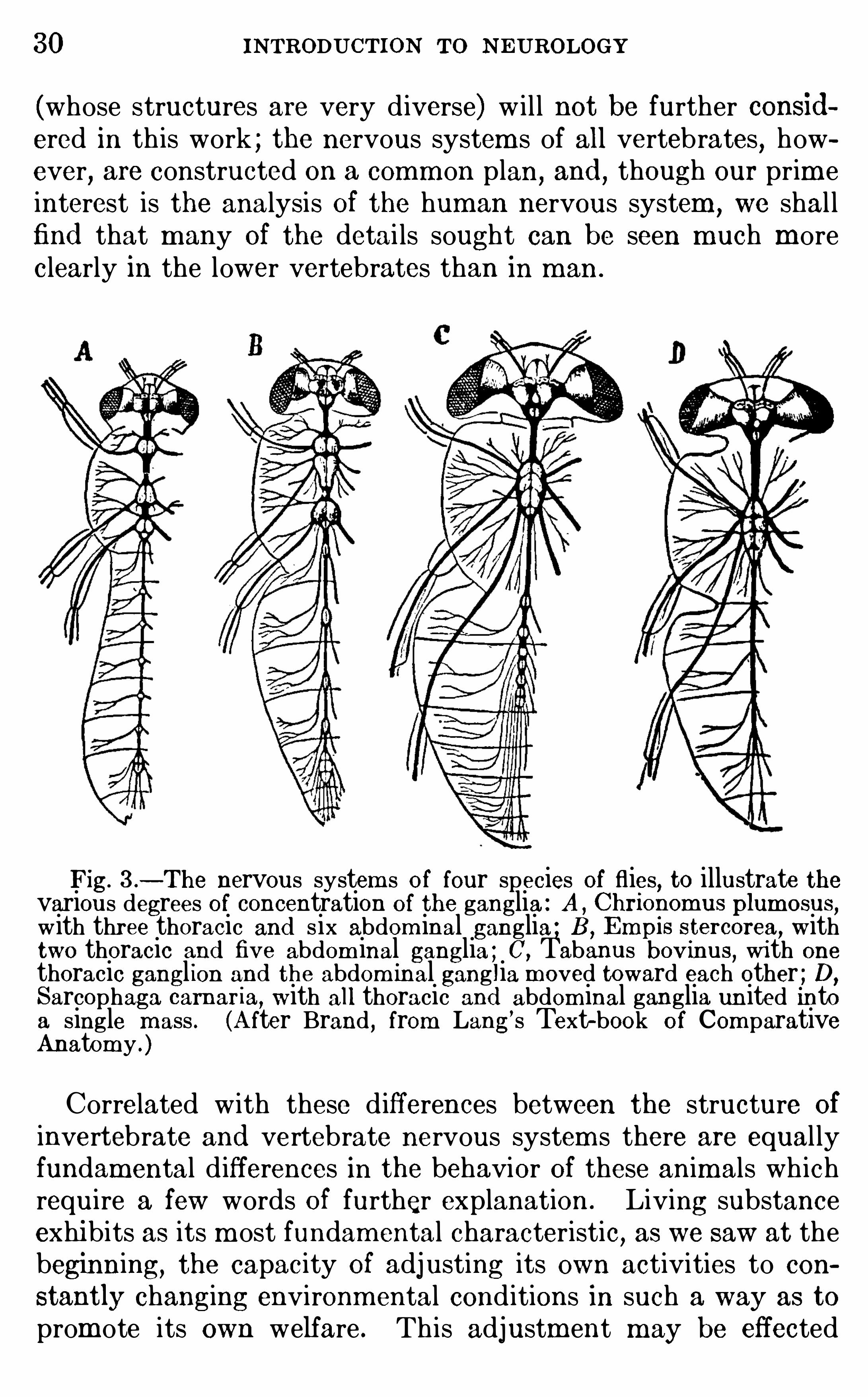
and the ganglia Of the central nervoussystem are m odified in a characteristic way in each Of theseregions . Figure 3 i l lustrates the nervous system s Of four SpeciesOf fl ies
,show ing different degrees of concentration Of the gangl ia.
In all cases the head part (brain) is greatly en larged , and isarranged , as in worm s, in gangl ia above and below the m outhand esophagus . The other ganglia are d iversely arranged, fromthe sim ple condition (A ) where there are three thoracic gangl ia,one for each pair Of legs, and six abdom inal gangl ia, through intermed iate stages (B and C) , to the highest form (D) , where allof the gangl ia of both thorax and abdom en are united in a singlethoracic mass .The type of nervous system just described is found throughout
the highest groups Of invertebrate animals,as in insects and
Spiders,and is constructed on a total ly different plan from that
Of all of the vertebrate or back- boned animals . In this lattergroup we have, instead of a segm ented chain of ventrally placedsol id ganglia, a hol low tube of nervous tissue wh ich extendsalong the back or dorsal wal l of the body and constitutes thespinal cord and brain . The cavi ty or lumen Of th is tube extendsthroughout the entire length of the central nervous system
,
form ing the ventricles Of the brain and the central canal Of thespinal cord . The detai ls of the invertebrate nervous system s
30 INTRODUCTION TO N EUROLOGY
(whose structures are very d iverse) w i l l not be further considered in th is work ; the nervous system s of all vertebrates
,how
ever, are constructed on a comm on plan,and
,though our prim e
interest is the analysis Of the human nervous system,we shal l
find that many Of the detai ls sought can be seen much moreclearly in the lower vertebrates than in m an .
Fig. 3 .—The nervous system s of four species of flies, to illustrate the
various degrees of concentration of the ganglia : A , Chrionomus plum osus,
with three thoracic and six abdom inal ganglia ; B ,Empis stercorea, with
two thoracic and five abdom inal ganglia ; C,Tabanus bovinus, with one
thoracic gangl ion and the abdom inal ganglia m oved toward each other ; D,
Sarcophaga cam aria, with all thoracic and abdom inal ganglia un ited intoa single m ass . (A fter Brand , from Lang’s Text-book of Com parativeAnatomy .)
Correlated with these d ifferences between the structure Of
invertebrate and vertebrate nervous system s there are equal lyfundam ental d ifferences in the behavior of these anim als wh ichrequire a few words of further explanation . Living substanceexhibits as its m ost fundam ental characteristic, as we saw at thebeginning, the capacity Of adjusting its own activities to con
stantly changing environmental cond itions in such a way as to
prom ote its own welfare . Th is adjustm ent m ay be effected

THE NERVOUS FUN CTIONS 31
in two ways,both of wh ich are universal ly present and which
throughout the remainder Of th is work we shal l cal l the invariableor innate behavior and the vari able or individual ly m odifiablebehavior .Every animal reaction,
then,contains these two factors, the
invariab le and the variable or individual ly m odifiab le. The
first factor is a function of the relatively stab le organization Of
the particular living substance invo lved . The pattern of th isorganization is inherited , and these characteristics Of the be
havior are, therefore, comm on,except for relatively sl ight devia
tions,to all m em bers Of the race or species ; they are rigidly
determ ined by innate bodi ly organization so arranged as tofaci litate the appropriate reactions
,in an invariab le m echanical
fash ion, to every kind Of stimulation to wh ich the organismis capab le of responding at all. In the strictly vegetative functions
,in all true reflexes (as these are defined on page and in
purely instinctive activities in general th is factor of behavior isdom inant .But in addition to this invariable innate behavior
,all organ
ism s have som e power to m odify their characteristic action system s in adaptation to changed environm ental relations . Th isind ividual m od ifiability is known as b iological regulation , a process which has Of late been very careful ly stud ied . We cannothere enter into the problem s connected with form regulation,
that is,the power Of an organism to restore its norm al form after
muti lation or other injury . On regulation in behavior referencesh ould be made to the works of Jennings and Ch i ld . In lowerorganism s Jennings recognizes three factors in the regulation Of
behavior : First,the occurrence of definite internal processes ;
these form part of the invariable hered itary action system re
ferred to above. Second,interference w ith these processes
causes a change Of behavior and varied m ovem ents,subjecting
the organ ism to m any d ifferent cond itions . Th ird,one Of these
conditions m ay rel ieve the interference with the internal processes , SO that the changes in behavior cease and the rel ievingcondition is thus retained . Lack Of oxygen,
for instance, wouldinterfere w ith an anim al ’s internal processes ; th is leads it to m oveabout ; if final ly it enters a region plentifully supplied wi th oxygen, the internal processes return to normal
,the m ovement

32 INTRODUCTION TO N EUROLOGY
ceases, and the animal again settles down to rest . If th is regulatory process is oft repeated another factor enters
,viz .
,the
faci litation of a given adjustment by repetition . Thus arisephysiological hab its or acquired automatism s .The m ore h ighly com plex form s Of ind ividual m od ifiability
are term ed associative m em ory and intelligence, and the latterOf these is by definition consciously performed . Whether consciousness is present in the sim pler form s Of “associativem em ory”
as these are dem onstrated by students Of animal behav ior inlower anim als cannot be positively determ ined . In the behaviorof lower animals there are no criteria which enab le us to tel lwhether a given act is consciously performed or not, and , therefore
,the lower lim its Of intelligence in the animal kingdom are
prob lematical . In other words,the manifestations Of variab le
behavi or form a graded series from the sim ple regulatory phenom ena Of unicel lular organism s , as i llustrated above
,to the
highest human intel ligence, so far as these express them selvesobjectively.
In mankind,where intel ligent behavior is dom inant, the
stereotyping of the adjustments by repetition (true habit forma
tion) may also take place, and in th is case the acquired au
tomatism s are sometim es said to arise by “lapsed intelligence,”
that is,an act wh ich has been consciously learned may ulti
mately come to be perf ormed m echanical ly and nearly or quiteunconsciously. Much of the process of elem entary educationis concerned w ith the estab lishm ent Of such habitual reactions tofrequently recurring Situations . How far
“lapsed intelligence”
is represented in the SO- called instincts Of other anim als is sti l la debated question (see p .
Am ong the invertebrate anim als, the insects and their allies
possess a bodi ly organization wh ich favors the performance ofrelatively few m ovem ents in a very perfect fash ion, that is , theaction system is sim ple but h igh ly perfected w ith in its own range.
Their reflexes and instincts are very perfectly performed,but
the num ber Of such reactions which the anim al can make is rathersharply l im ited and fixed by the inherited bodi ly structure.
Their behavior is dom inated by the invariable and innate factors and they cannot
‘
read ily adapt them selves to unusual conditions . The vertebrates l ikewise have many elem ents of their
34 INTRODUCTION TO NEUROLOGY
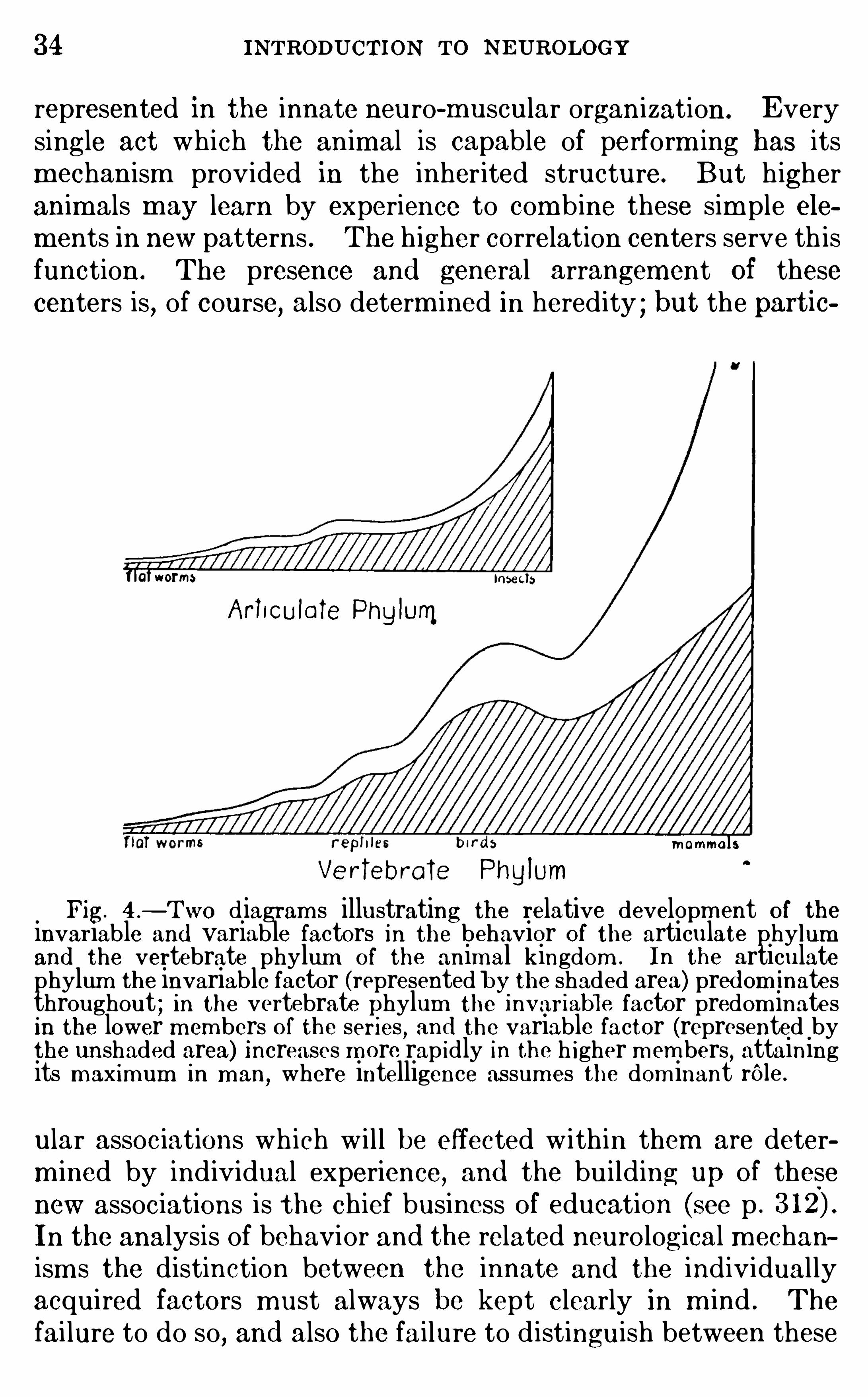
represented in the innate neuro-m uscular organization . Everysingle act which the animal is capable of perform ing has itsm echanism provided in the inherited structure. But h igheranim als m ay learn by experience to com b ine these sim ple elements in new patterns . The h igher correlation centers serve th isfunction . The presence and general arrangement Of thesecenters is , of course, also determ ined in heredity ; but the partic
wormsArluculole Phy urn
mammalsVe rlebrOTe Phglum
Fig. 4 .
—Two diagram s illustrating the relative developm ent of theinvariable and variable factors in the behavior Of the articulate phylumand the vertebrate phylum O f the animal kingdom . In the articulatephylum the invariable factor (represented by the shaded area) pred om inatesthroughout ; in the vertebrate phylum the 1nvariable factor predom inatesin the lower m embers of the seri es
,and the variable factor (represented by
the unshaded area) Increases m ore rap idly In the h igher m em bers,attaining
its m ax imum in m an,where intelligence assum es the dom inant rOle .
ular assoc iations wh ich w i l l be effected w ithin them are determ ined by ind ividual experience, and the bui lding up of thesenew associations is the chief business O f education (see p .
In the analysis of behavior and the related neurological m echanism s the distinction between the innate and the individual lyacquired factors must always be kept clearly in m ind . The
fai lure to do so, and also the failure to d istinguish between these

THE N ERVOUS FUNCTIONS 35
two factors and the acquired autom atism s (p . is responsiblefor m uch confusion in the current discussions of instinct .
In the nom enclature Of the correlation centers there is considerablediversity of usage. In describing the adjustm ents m ade by these centersneurologists frequently use the words coordination ,
correlation,and associ
ation in about the sam e sense ; but the adjustm ents made in those centerswhich lie closer to the receptors or sense organs are physiologically O f different type from those m ade in the centers related m ore closely to theeffector apparatus . In recognition Of this fact the following usage has beensuggested to m e by Dr . F . L . Landacre and will be ad opted in thi s work :The term correlation is applied to those com binations Of the afferent
im pulses within the sensory centers which provide for the integration of
these impulses into appropriate or adaptive responses ; in other words, the
correlation centers determ ine what the reaction to a given com bination of
stim uli will be. Nervous im pulses from different receptors act upon the
correlation centers, and the reaction which follows will be the resultant ofthe interaction of all of the afferent im pulses (and physiological traces orvestiges of previous sim ilar responses) involved in the process . When thisresultant nervous di scharge passes over into the m otor centers and pathways
, the final comm on paths (see p . 62) innervated will lead to a responsewhose character Is determ ined by the organization Of the particular m otorcenters and paths actuated .
To the term coord ination we shall give a restricted significance, applyingit only to those processes em ploying anatom ically fixed arrangements of themotor apparatus which provide for the co-working Of ‘particular groups Ofmuscles (or other effectors) for the performance of defini te adaptively usefulresponses . Every reaction—even the sim plest reflex—involves the com
b ined action Of several different muscles , and these m uscles are so innervated as to facilitate their concerted action in this particular m ovem ent .These are called synergic m uscles . Coordination involves those adjustm ents which are m ade on the effector side of the reflex are (p . Thi sis the sense in which the term is applied by Sherrington in the followingpassage (Integrative Action of the N ervous System ,
p .
“R eflex coOrd ination m akes separate muscles whose contractions actharmoniously
,e. g.
, on a lever, contract together, although at separateplaces , so that they assist toward the sam e end . In other words , it excitessynergic muscles . But it in m any cases does more than that. Where twomuscles would antagonize each other’ s action the reflex are, instead Of
activating m erely one Of the two , causes when it activates the one depressionof the activity (tonic or rhythm ic contraction) of the other. The latter isan inhibitory effect .”The m otor paths and centers in general are m ore Sim ply organized than
are the sensory paths and centers . The nervous discharges through thesemotor system s are very direct and rapid . Com p lex nervous reactionsrequire m ore tim e than sim ple reflexes, and this delay or central pause ischiefly In the correlation centers rather than in the efferent coord inationm echanism s (see pp . 98,The word associ ation m ay be reserved for those higher correlations
where plasticity and m od ifiabili ty are the dom i nant features of the responseand whose centers are separated from the peripheral sensory apparatus bythe lower correlation centers which are devoted to the stereotyped Invar iable reflex responses . Correlation m ay be m echanically determ ined by

36 INTRODUCTION TO NEUROLOGY
innate structure, or there m ay be som e sm all m easure of individual mod ifiability
,but when the m od ifiabi li ty com es to be the dom inant characteristic,
so that the result of the stim ulus cannot be readily pred icted with m echanical precis ion ,
the process m ay be called association. The intelligent typesof reaction and all higher rational processes belong here, and the cerebralcortex is the chief apparatus em ployed .
The boundaries between the three types of centers just d istingu ishedare not always sharply drawn ,
especially In their sim pler form s , though Ingeneral they are easily distinguished . The m echanisms Of Coordinationare neurologically sim p ler than those of correlation and association
,and In
general they are developed in the m ore ventral parts Of the brain andspinal cord , that 18 , below the lim iting sulcus of the em bryoni c brain (p .
The correlation and association centers are developed in the m ore dorsalparts of the brain and cord
, and the greater part of the thalam us and cerebral hem ispheres is com posed of tissues Of this type. Nevertheless
, the d i s
tinctions here drawn are fundam entally physiological rather than anatomical, and coordination centers m ay be developed in the dorsal parts of thebrain,
as in the case of the cerebellum and probably also the corpus striatumof m amm als (though not the striatum of lower vertebrates) .
Summary.
—The functions wh ich characterize the nervoussystem have been derived from those Of ordinary protoplasmby further development Of three Of the fundamental protoplasm ic propert ies—viz .
,sensitivity
,conductivi ty
,and correlation .
The m ost prim itive form Of nervous system known is diffuse andlocal in its action,
but in all the m ore h igh ly developed form s thech ief nervous organs tend to be centralized for ease of generalcorrelation and control . Most of the types of nervous system sfound in the animal kingdom are represented in two distinct anddivergent lines Of evolution, one adapted especial ly wel l for thereflex and instinctive m ode of life and found in the worm s, insects , and their allies , and the other found in the vertebrates andculm inating in the hum an brain w ith its remarkable capacityfor individual ly acquired and conscious functions .
LITERATUREBAR KE R, L . F . 1901 . The N ervous System and Its Constituent N eu
rones, N ew York .CHILD ,C . M . 19 1 1 . The R egulatory Processes in Organism s , Journal of
Morphology , vol . xxii , pp . 17 1—222 .
EDINGE R,L. 1908 . TheRelations of Com parative Anatom y to Com par
ative Psychology, Jour. Com p . Neur. , vol . xviii , pp . 437—457 .
HERRICK, C . JUDSO N . 19 10 . The E volution of Intelligence and ItsOrgans
, Science, N . S.,vol . xxxi , p . 7—18 .
19 10 . The Re lations of the entral and Peripheral Nervous System sin Phylogeny
,Anat. R ecord
,vol . iv, pp . 59—6 9 .

THE NERVOUS FUNCTIONS 37
JE NNIN GS, H . S. 1905 . The Method Of R egulation in Behavior and inOther Fields, Jour . Exp. ZoOl . , vol . ii, pp . 473—494 .
1 906 . Behavior of the Lower Organi sm s, N ew York .
JLEWANDOWSKY, M . 1907 . Die Funktionen d es zentralen N ervensystem s
,
ena.
LO E B , J . 1900 . Com parative Physiology of the Brain and Com parativePsychology, N ew York .
PARKE R,G . H . 1909 . The Origin of the N ervous System and Its Ap
propriation of E ffectors, Pop. Sci . Monthly,vol. lxxv, pp . 56—64, 137—146 ,
253—263 , 338- 345 .
19 14 . The Origin and E volution of the N ervous System ,Pop. Sci .
Monthly,vol . lxxxiv
,pp . 1 18—127 .
PA RME L E E,M . 19 13 . The Science Of Human Behavior, N ew York .
SHE RRIN GTON , C . S. 1906 . The Integrative A ction of the N ervous System , New York .
VE RWORN ,M . 1899 . General Physiology , London .
WASHBURN,MA RGA RET F . 1908 . The An imal Mind
,N ew York .
WATSON ,J . B . 19 14 . Behavior
, An Introduction to Com parative Psychology, N ew York .
YE RKE S, R . M . 1905 . Concerning the Genetic R elations of Types ofAction
,Jour. Com p . N eur. , vol. xv, pp . 132—137 .

CHAPTER I I I
THE NEURON
As we have seen in the last chapter,the functions Of irrita
bility,conduction
,and correlation are the most d istinctive fea
tures of the nervous system . Like the rest of the body,the
nervous tissues are com posed Of cel ls,the irritabi l ity of whose
protoplasm is Of diverse sorts in adaptation to d ifferent functional requirements . Each sense organ,
for instance,is irri
table’
to its own adequate stimulus only (see pp . 25,
The
functions of correlation and integration Of bodi ly actions cannotbe carried on by the nerve- cel ls as individuals , but they are
effected by various types of connections between the differentcel ls in the nerve- centers . The character Of any particular correlation ,
in other words,is a function Of the pattern in accord
ance w ith which the nerve-cel ls concerned are connected w itheach other and w ith the end - organs Of the reflexarcs invo lved .
The conducting function of nerve- cel ls is,perhaps
,their m ost
striking peculiarity , and their very special form s are d ue largelyto the fact that thei r business is to connect remote parts Of thebody so that these parts can coOperate in com plicated m ovements .
Not all of the cells which com pose the central nervous system are nervecells . The brain and spinal cord are surrounded by three connective-tissuem em branes (dura m ater, arachnoid
,and pia m ater, in the aggregate
term ed m eninges ) whose functions are chiefly protective and nutritive ;from the inner m em brane, the pia m ater
,blood-vessels
, and strands ofconnective tissue extend into the true nervous substance. In addition tothese non-nervous elem ents which grow into the central nervous systemfrom without
,the substance of the brain and sp inal cord contains a sup
porting fram ework composed of ependym 1 and neuroglia or glia cells whichdevelop from the pr im itive em bryonic nervous system (the neural tube, seepp . 1 06 . but are not known to perform nervous functions , thoughnutritive and other functions have been ascribed to them (see p .
The true nerve- cells are called neurons . There has been a
long controversy regarding the way in which the neurons of the38

THE N EURON 39
adult body are developed from the cel ls of the em bryonic nervoussystem ; but it is now generally accepted that each neuron isdeveloped from a single em bryonic cel l (known as a neurob last) ,and that in the adult body each neuron has a certain m easure Ofanatom ical and physiological d istinctness from all Of the others .The very young nerve- cel l (neuroblast) is oval in form and is
com posed Of a nucleus and its surrounding protoplasm (cytoplasm ) but in further developm ent it rapid ly elongates by theoutgrowth of one or m ore fibrous processes from the cel l body,SO that the m ature neuron may be regarded as a protoplasm icfiber with a thickening som ewhere in its course wh ich is the cel lbody Of the original neuroblast and contains the cel l nucleusand a part only of its cytoplasm (th is part being cal led theperikaryon) , the rem ainder Of the cytoplasm com posing the
fibrous processes , that is , the nerve-fibers. The cel l body of
the mature neuron is som etim es loosely term ed the nerve- cell,
though the latter term should strictly include the entire neuron .
The im portance Of the conducting function is reflected In the elon
gated form s Of the neurons and In the peculiar protoplasm ic strueture Of the nerve-fibers . The function Of the cel l body Is ch ieflynutritive ; the entire neuron dies if the cel l body is destroyed .
Each neuron may be regarded as essential ly an elongated conductor
,and these uni ts are arranged in chains in such a way that
a nervous im pulse is passed from one to another in series . Sincethe arrangem ent is such that the nervous im pulse usuallypasses through the series in only one direction (see the typicalreflex are
,Fig. 1
,p . each neuron has a receptive function
at one end and di scharges its im pulse at the other end . Th is iswhat is m eant by the polari ty of the neuron (see pp . 52 and
The sim pler form s O f neurons are bipolar,w ith one or m ore
processes known as d end ri tes conducting nervous im pulses towardthe cel l body
,and (usually) only one process , the axon or neurite,
conducting away from the cell body . The dendri tes are usual lyshort, and in this case their structure is sim i lar to that of the cel lbody. But where the dendrites are long, as in the neurons Of thespinal and cranial gangl ia (Figs . 1 , they m ay have thesam e structure as the axon . The axons are the axis- cyl indersOf the longer nerve-fibers and are structural ly very different fromthe protoplasm Of the cel l body
,being com posed ch iefly of
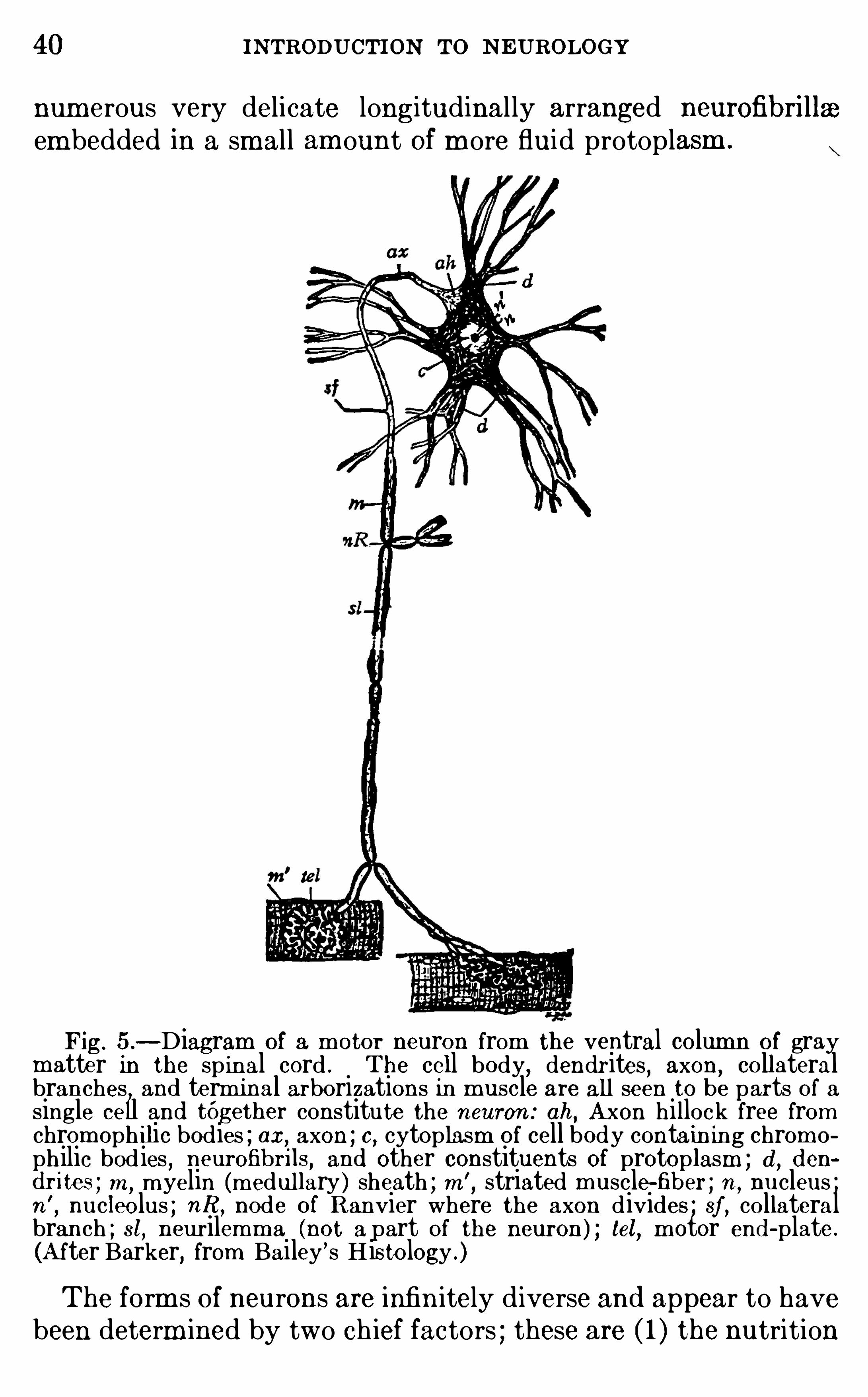
40 INTRODUCTION TO N EUROLOGY
num erous very delicate longitudinal ly arranged neurofibri llaa
em bedded in a small am ount Of m ore fluid protoplasm .
Fig. 5 .—Diagram of a m otor neuron from the ventral column of gray
matter in the sp inal cord . The cell body,dendrites , axon
, collateralbranches and term inal arborizations in m uscle are all seen to be parts of asingle cell and tOgether constitute the neuron : ah, Axon hillock free fromchrom oph ilic bodies ; ax, axon ; c, cytoplasm of cell body contain ing chrom o
philic bodies,neurofibrils, and other constituents of protoplasm ; d , d en
d r ites ; m ,m yelin (m edullary ) sheath ; m ’
, striated m uscle-fiber ; n ,nucleus ;
n’
,nucleolus ; nR , node of Ranvier where the axon divides ; sf , collateral
branch ; st, neur ilemm a (not a part of the neuron) ; tel, m otor end -plate.
(A fter Barker, from Bailey’s His tology .)
The form s of neurons are infin itely diverse and appear to havebeen determ ined by two ch ief factors ; these are ( 1 ) the nutrition

42 INTRODUCTION TO N EUROLOGY
Neurons can function only when connected together in chains,
so that the nervous im pulse can be passed from one to the other .In any such chain the neuron first to be excited is called theneuron of the first order
,and the succeeding mem bers of the
series neurons Of the second , th ird , fourth order, and so forth .
A ll reflexes require an afferent neuron whi ch conducts the ner
vous im pulse from the receptor to the center , one or m ore efferent neurons conducting from the center to the organ of response,
Fig. 7 .—The body Of a pyram idal neuron from cerebral cortex
,
stained by N issl ’s m ethod,illustrating the arrangem ent O f the chrom ophilic
substance and the form of the nucleus : a,Axon ; b, chrom ophilic bod ies
surrounding the nucleus ; 0 , a m ass O f chrom e hilic substance in the angleform ed by the branching of a dendrite ; d , nuc eus of a neuroglia cell (not apart of the neuron) . (Af ter RamOn y Cajal . )and usually one or m ore neurons intercalated between thesewi thin the center itself (see pp . 25
,56
,Figure 1 , p . 25
,
i llustrates the sim plest possible connection Of neurons in a reflexarc Of the spinal cord
,involving only two elements . The
afferent neuron sends its dendrite to the skin and its axon intothe spinal cord , where the nervous im pul se is taken up by thedendrites Of the efferent neuron,
wh ich in turn transm its it to amuscle. Figures 5 to 9 i l lustrate the form s of other neurons.

THE NEURON 43
The different dendri tes Of a neuron may be physiologicallyall alike
,or they m ay spread out in different directions to receive
nervous im pulses of d iverse sorts from d ifferent sources . Sim ilarly the axon may di scharge its nervous im pulse into a singlenerve center or peripheral end -organ,
or it may branch , thusconnecting wi th and stimulating to activity two or m ore diversefunctional mechani sm s . In other words
,a given neuron m ay be
a link in a chain of some sim ple nervous c ircuit (Fig. or itmay be adapted to col lect nervous im pulses from d ifferentsources and discharge them into a single final common path , orin the th ird place it may receive nervous im pulses of one or m orefunctional sorts and then discharge its own nervous energy intoseveral rem ote parts Of the nervous system . Th is , in brief, is them echani sm Of correlation, and i l lustrations Of these differenttypes Of connection wi l l be found in the follow ing chapters . If
animal reactions were sim ple responses SO arranged that a givenstimulus could produce only one kind Of m ovem ent
,the only
nervous m echanism required would be a single neuron transm itting the excitation from the point of stim ulation to the organ of
response,as a cal l bel l m ay be rung by pul ling a bel l cord . But
the actual reactions are always m ore com plex than th is , so thatseveral neurons must be connected in series w ith various d ivergent pathways Of nervous d ischarge wh ich reach d ifferentcorrelation centers
,all Of wh ich must coOperate in the final
response . I llustrations Of som e of these com plicated reflexmechanism s w i ll be found in Chapter IV .
Neurons with short dendri tes and a single long axon are them ost comm on form and were term ed Type I by Golgi (Fig.
In som e cases (Fig. 9 ) the axon also is very Short,breaking up
in the imm ed iate neighborhood of the cell body ; these are the
Type I I neurons Of Golgi and appear to be adapted for the d iffusion and summation Of stim ul i with in a nerve center. The
neurons of the spinal and cranial gangl ia form a third type.
In embryonic development they begin as bipolar cells wi th a
dendritic process at one end and an axonal process at the
Opposite end of the cel l body ; but in the course Of further developm ent (Fig. 10) the two processes approach each other andfinal ly unite for a short distance into a single stem ,
which thenseparates into an axon and a h igh ly special form of dendrite
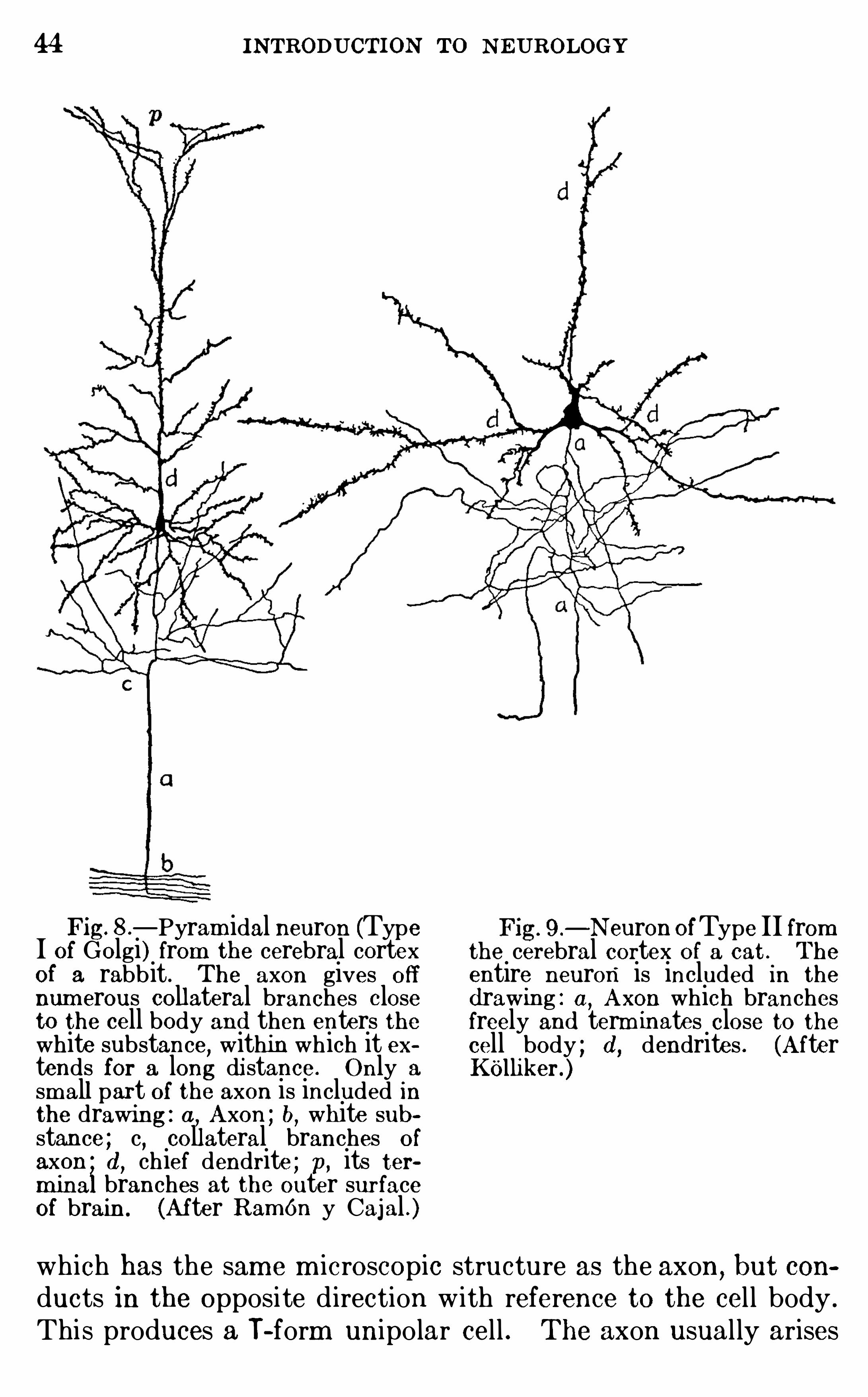
44 INTRODUCTION TO NEUROLOGY
Fig—Pyram idal neuron (Type
I of Golgi) from the cerebral cortexof a rabbit . The axon gives off
num erous collateral branches closeto the cell body and then enters thewhite substance
,within which it ex
tends for a long di stance. Only a
sm all part of the axon is included inthe drawing : a, Axon ; b, white sub
stance ; c, collateral branches of
axon d, chief dendrite ; p, its ter
m inal branches at the outer surfaceof brain. (After RamOn y Cajal . )
9 .—N euron OfType II from
the cerebral cortex of a cat . The
entire neuron is included in the
d rawing : a,A xon which branches
freely and term inates close to thecell body ; d , dendrites . (A fterKOlliker.)
which has the sam e m icroscopic structure as the axon,but con
ducts in the opposite d irection w ith reference to the cel l body .
Thi s produces a T- form unipo lar cell . The axon usual ly arises
THE NE URON 45
from the cel l body ; it may arise from the base Of one Of the d end rites or, rarely, from the apex of the chi ef dendrite (Fig.
N eurons d iffer in internal structure,as well as in form
,from
the other cells of the body . The m ost im portant Of these pecu
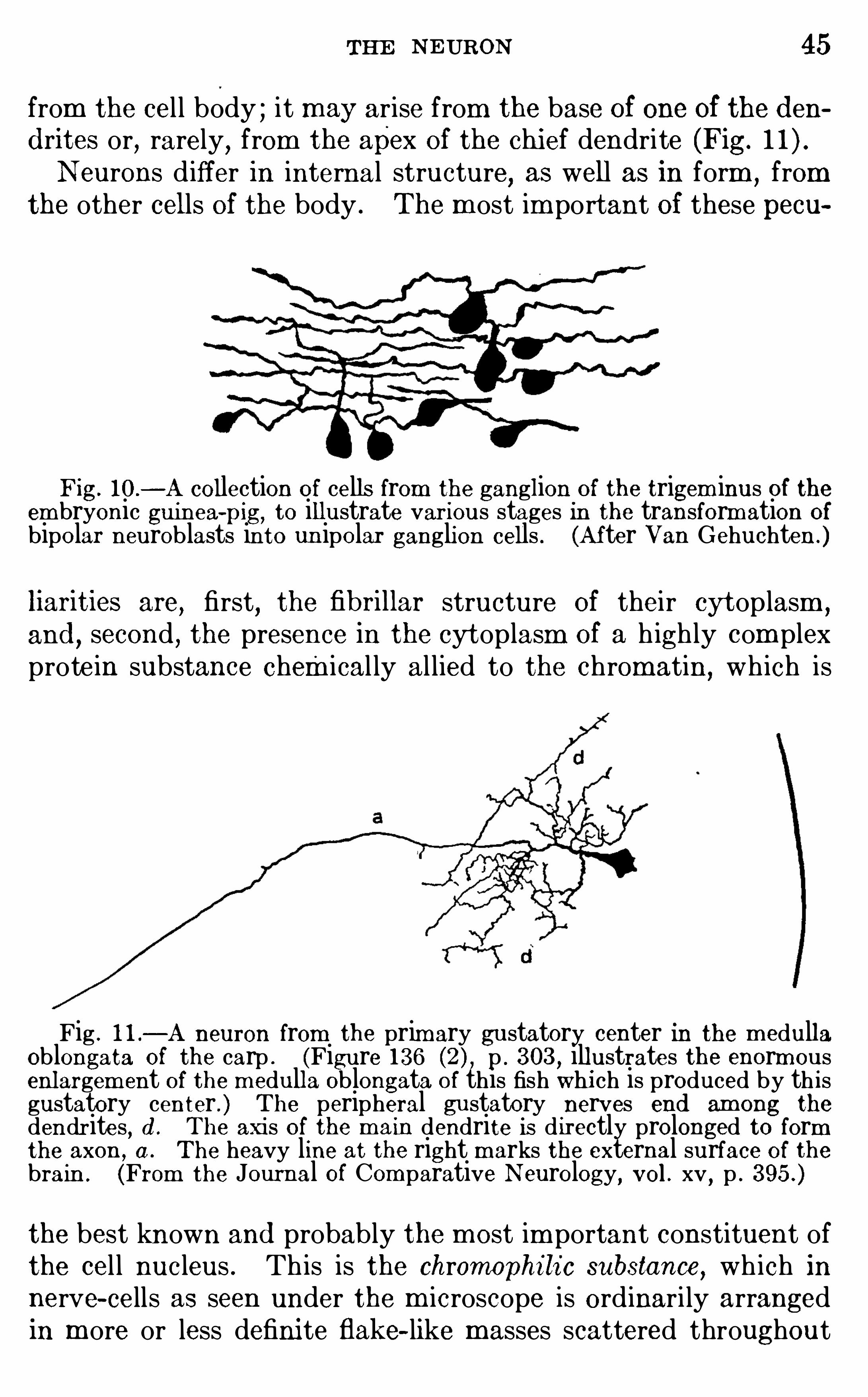
Fig. 10 .
—A collection of cells from the ganglion of the trigem inus of theem bryonic guinea-
pig, to illustrate various stages in the transformation ofbipolar neuroblasts into un ipolar ganglion cells . (After Van Gehuchten .)
liarities are,
first,the fibrillar structure Of their cyt oplasm ,
and,second , the presence in the cytoplasm Of a h igh ly com plex
prote in substance chemical ly al lied to the chromatin,wh ich is
Fig . 1 1 .—A neuron from the prim ary gustatory center in the m edulla
Oblongata of the carp . (Figure 136 p . 303 , illustrates the enorm ousenlargem ent of the m edulla Oblongata of this fish whi ch is produced by th isgustatory center . ) The peripheral gustatory nerves end am ong thedendrites, d . The axis O f the m ain dendrite is d irectly prolonged to formthe axon , a . The heavy line at the right m arks the external surface of thebrain . (From the Journal Of Com parative N eurology, vol . xv , p .
the best known and probably the m ost im portant constituent ofthe cel l nucleus . Th is is the chromophi li c substance
,wh ich in
nerve- cells as seen under the m icroscope is ordinari ly arrangedin m ore or less defini te flake- like m asses scattered throughout

46 INTRODUCTION TO N EUROLOGY
the cytoplasm of the cel l and extend ing out into the larger d end rites (see Figs . 6 , These m asses were first careful ly investigated by N issl , who devised a special staining m ethod for thatpurpose ; they are, accordingly , Often called the N issl bodies
,and
som etim es tigroid bod ies . They never occur in the axon nor
in a spec ial conical protuberance Of the cel l body (the axonh i l lock) from wh ich the axon arises (see Fig. 5
,ah
,and Fig. 6
,c) .
The neurOfibrils are very del icate strands Of denser protoplasmfound in all parts Of the neuron except the nucleus . They are bym any regarded as the specific conducting elements Of the neuron
,
though the evidence for th is is not conclusive. They ram ifythroughout the cytoplasm (Fig. passing through the cel lbody from one process to another .The longer nerve-fibers are usually enveloped by a th ick wh ite
gl istening sheath Of m yel in,a fat- like substance secreted by the
nerve-fibers them selves . Th is myelin sheath,or m edullary
sheath , is a part Of the neuron w ith wh ich it is related and thefibers wh ich possess it are called m yelinated or m edul lated fibers ;these fibers com pose the wh ite m atter Of the brain and a largepart Of the peripheral nerves (see Fig. There may be, in ad
d ition,in the case Of the peripheral nerves an outer sheath , the
neuri lemma (prim itive sheath or sheath of Schwann) . This is ath inner nucleated mem brane, not a part of the neuron to wh ichit is attached , but form ed from surrounding cells .The function Of the myelin sheath has O ften been regarded as
sim ply that Of an insulating substance to prevent the overflowand loss Of the nervous im pulse conducted by the axon, butthere is som e evidence that this sheath plays an im portant partin the chem ical processes involved in the act of nervous conduetion . The neurilemma is likewise Often spoken Of as a protectingmembrane. Whether it has any other function in the normall ife Of the nerve-fiber is unknown ; but if a nerve-fiber is by accident severed from its cel l body, it is known that the nuclei ofthe neuri lemma play a very im portant part in the degenerationand regeneration Of the severed fiber and the restoration of itsnorm al function .
As has been suggested , nerve-fibers cut Off from thei r cel lbodies imm ediately d ie and degenerate . But in the case Of
peripheral nerves the neuri lemma nuclei do not d ie ; and , appa

THE NEURON 47
rently w ith the ai d Of these nuclei , a new nerve-fiber may underfavorable condi tions grow out from the central stum p of the Cutnerve, and final ly the entire nerve may regenerate . In the cen
Fig . 12 .
—Cell from the ventral gray colum n of the human spinal cord ,Illustrat ing the arrangem ent of the neurofibrils : ax, Axon ; lu,
interfibrillar spaces occupied by chromophilic substance ; n ,
nucleus ; x, neurofibrilspassmg from one dendrite to another ; y, sim ilar neurofibrils pas sing throughthe body of the cell . (A fter Bethe, from Heidenhain
’ s Plasm a und Zelle) .
tral nervous system,where the neuri lemma is absent or greatly
reduced, the regeneration Of such injured nerves takes place
wi th great difficulty , if at all.
48 INTRODUCTION TO N EUROLOGY
I t is possible by a special m ethod Of staining devi sed byMarch ito d ifferentiate myelinated fibers wh ich are in process Of degeneration from the norm al fibers with wh ich they may be m ingled .
Th is m ethod has Often perm itted a m uch m ore precise determ ination of the exact course Of the fibers Of a given peripheral
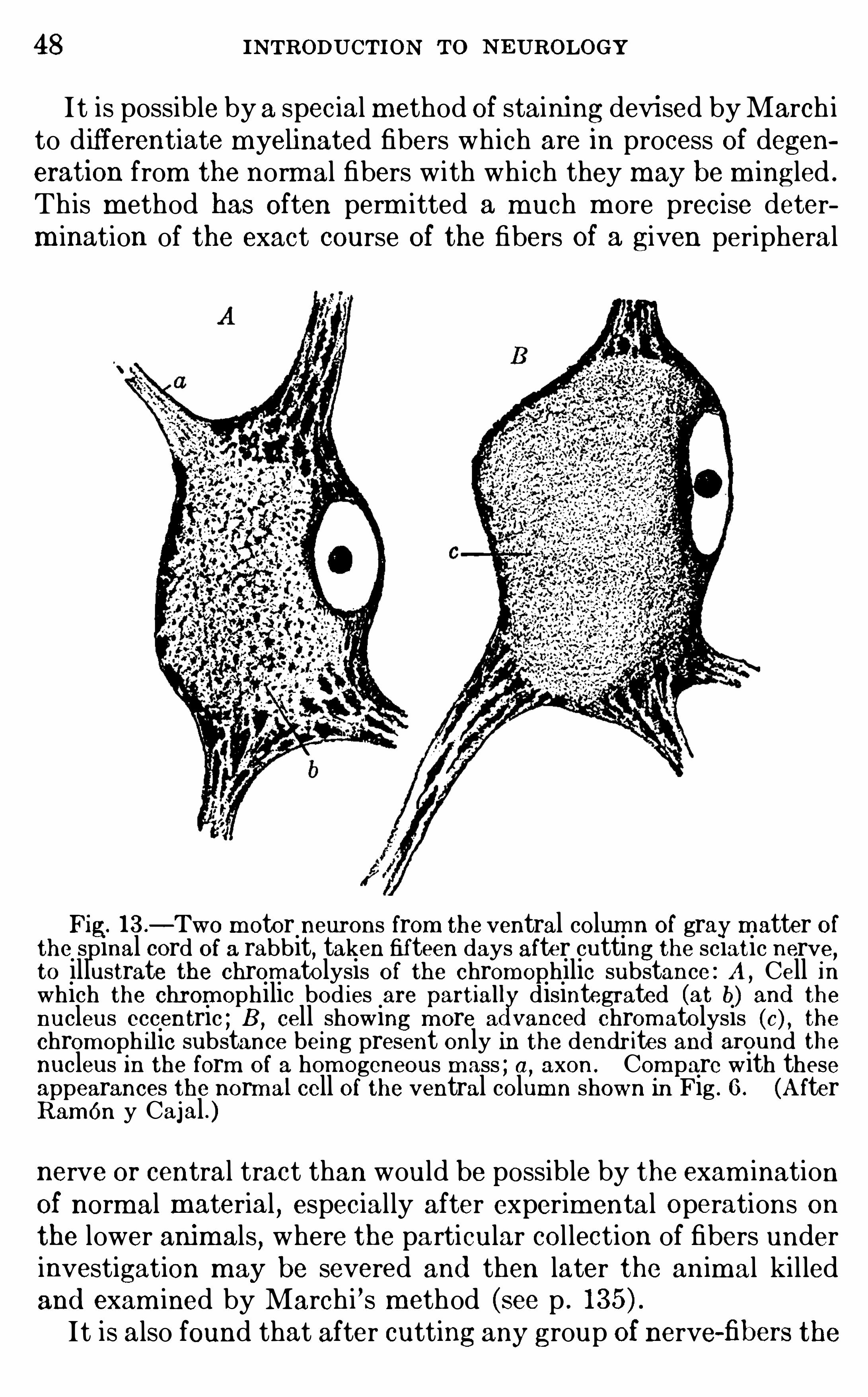
Fig. l 3 .—Two m otor neurons from the ventral colum n of gray matter of
the spinal cord Of a rabbit, taken fifteen days after cutting the sciatic nerve,to illustrate the chrom atolysis O f the chromophilic substance : A , Cell inwhich the chrom ophilic bod ies are partiall disintegrated (at b) and thenucleus eccentric ; B , cell showing m ore a vanced chrom atolysis (c) , thechrom oph ilic substance being present only in the dendr ites and around thenucleus in the form of a hom ogeneous m ass ; a,
axon . Com pare with theseappearances the norm al cell Of the ventral column shown in Fig . 6 . (A fterRamon y Cajal . )nerve or central tract than would be possible by the exam inationOf normal material
,especially after experim ental Operations on
the lower animals , where the particular col lection of fibers underinvestigation m ay be severed and then later the animal kil ledand exam ined by March i ’s method (see p .
It is also found that after cutting any group Of nerve-fibers the

50 INTRODUCTION TO NEUROLOGY
Of any part Of the neuron affects the welfare Of the whole, andthe destruction Of the nucleus and cel l body destroys the entireneuron
,but such injuries d o not directly affect adjacent
neurons .6 . The neuron is a functional unit or
,better
,the functional
unit Of the nervous system .
Fig. 14 .—N eurons from the trapezoid body of the m edulla Oblongata Of a
cat, illustrating d ifferent form s Of synapse : a, Delicate pericellular netaround the cell body of a neuron which is not shown ; b, coarser endings ; c,still coarser net ; d , calyx—like envelope . In b
,c,and d , at the left of the
figure, the globular cell body of the neuron Of the second order is shadedwith lighter stipple than the term inals of the axon of the neuron of the firstorder. (A fter Veratti, from E dinger’s Vorlesungen . ) (It should be notedthat in this account we d o not follow Veratti ’s interpretation of thesestructures , but that Of Held , Ramon y Cajal, and the majority of otherneurologists .)
These six propositions are accepted in their entirety by manyneurologists ; but it should be clearly understood that all Of
them are controverted by others . The fourth proposition,in
particular, has been the subject Of violent attack (see the d iscussion of the synapse below) . The neuron
,m oreover
,is a
functional unit (proposition 6 ) in only a rather lim ited sense(see p . W ithout further d iscussion of the merits Of these
THE NEURON 51
controversial questions,it m ay be regarded as general ly accepted
that all of the preceding propositions have som e measure of
factual basis,though different neurologists would give various
interpretations and m odifications of some Of them .
The place where the axon Of one neuron com es into physiological relation with another neuron is known as the synapse.
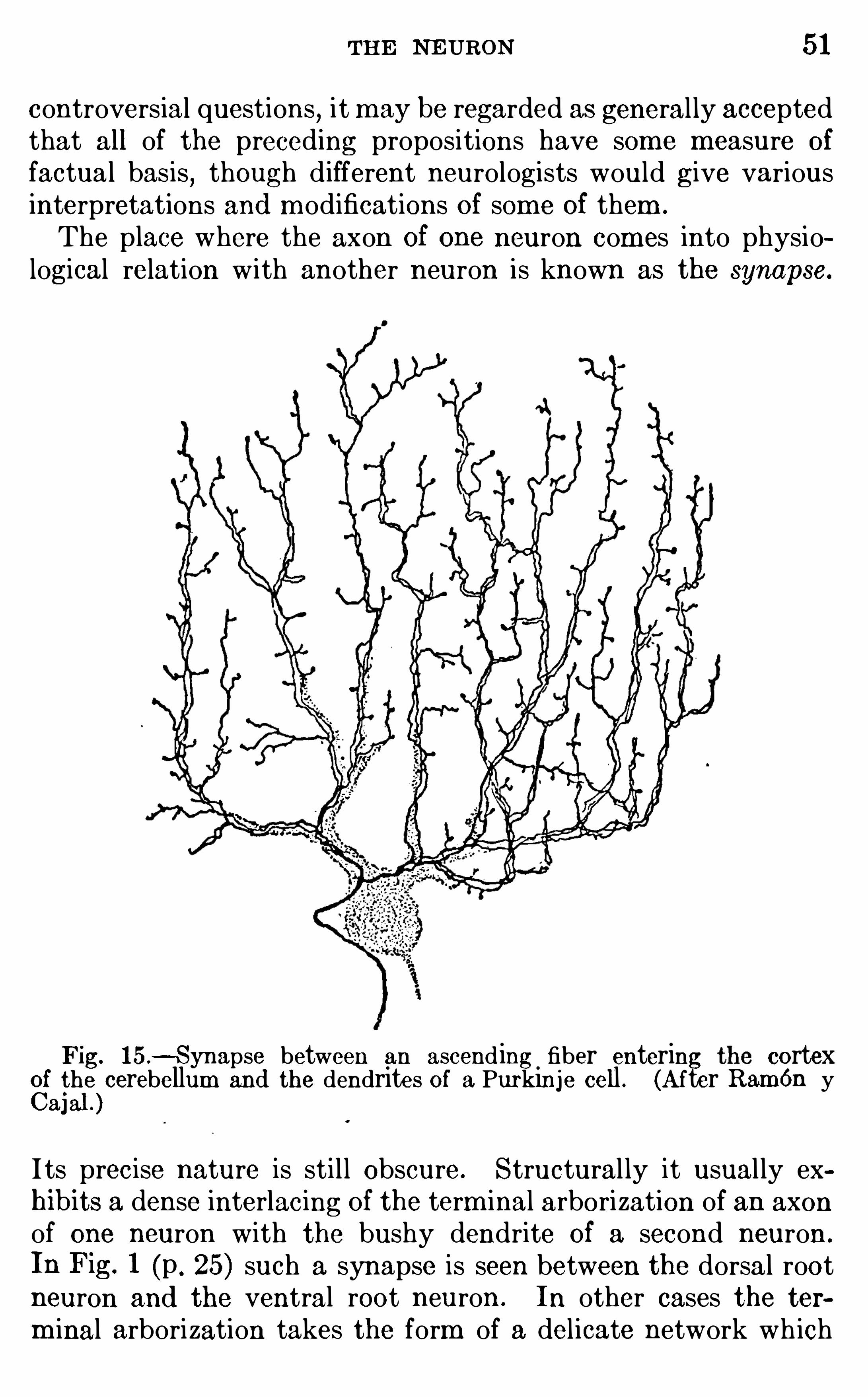
Fig. 15 .—Synapse between an ascending fiber entering cortex
Cf i
gle cerebellum and the dendrites of a Purkinje cell . (After Ram én y
a]
I ts precise nature is sti l l Obscure. Structural ly it usual ly exhibits a dense interlacing of the term inal arborization of an axonof one neuron with the bushy dendrite Of a second neuron .
In Fig. 1 (p . 25) such a synapse is seen between the dorsal rootneuron and the ventral root neuron . In other cases the term inal arborization takes the form of a delicate network wh ich

52 INTRODUCTION TO NE UROLOGY
twines around the cel l body of the second neuron or of a calyxlike expansion or coarse-meshed reticulum closely envelopingthe cel l body (Fig. Another form of synapse is Seen in Fig.
15 from the cortex of the cerebel lum . The body and largerdendrites Of a single cortical neuron Of the type known as
Purkinje cells (see p . 19 1 ) are shown in gray, and the term inalbranches Of an afferent neuron are seen twin ing about the d end ritic branches Of the Purkinj e cel l
,thus form ing a very int im ate
uni on. Sim i lar synapses are found in the cerebral cortex (p .
Figure 16 i llustrates a type of synapse also found in the
Fig. 16 .-A basket cell from the cerebellar cortex Of a rat, illustratin
the discharge of a single neuron,B , by synaptic connection with the cc
bodies of several Purkinje neurons, A ,by bas ket-hke endings of the axon :
A , cells of Purkin je ; a , the basket-like synapse on the body of a Furkin e
cell ; B , the basket cell ; b, term inus Of the axon ; c, axon Of basket cc(A fter Ramon y Cajal ; cf . Fig. 89
,p .
cerebellaf cortex . A single basket cel l , B,has a short axon
whose branches form synapses around the bodies Of a largenumber of Purkinje cells
,thus diffusing and greatly strength
ening the nervous d ischarge (see p . 192 and Fig. 89 , b) . Forstill other types Of synapse see Figs . 61 , 89 , 98, 104 , 109 , 126 .
The synapse has been a crucial point in recent d iscussionsregard ing the general physiology Of the nervous system ,
m anyneurologists believing that it is the most im portant part of thereflex circui ts (see, for instance, on the theory of sleep , p .
The doctrine of the polarization of the neuron (p . 39 ) im plies
THE NEURON 53
that at the synapse there must be a reversal Of the po larity w ithreference to the cel l body as the nervous im pulse passes overfrom an axon to a dendrite.
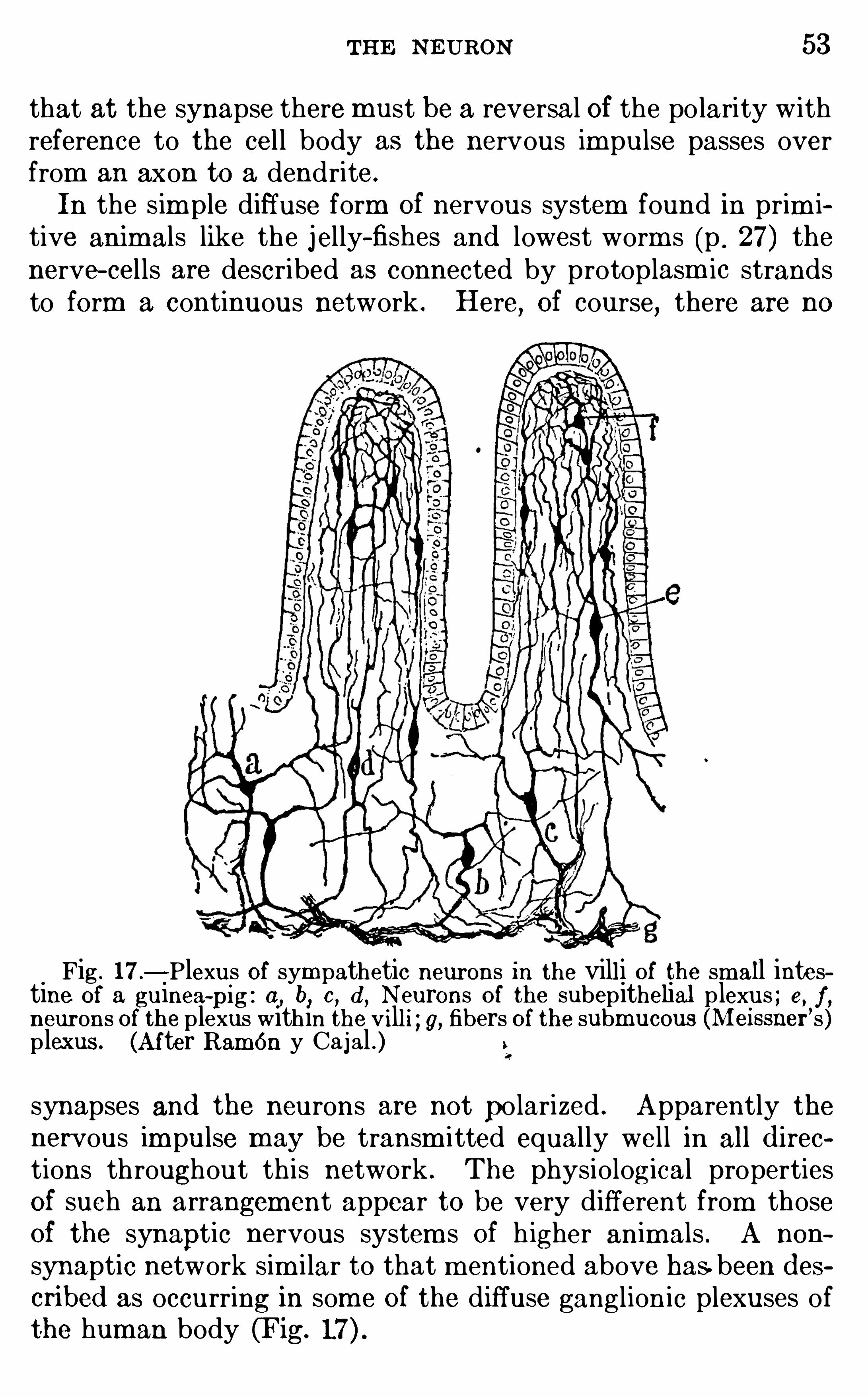
In the sim ple diffuse form Of nervous system found in prim itive anim als like the jelly-fishes and lowest worm s (p . 27) thenerve- cel ls are described as connected by protoplasm ic strandsto form a continuous network . Here
, Of course, there are no
Fig . 17—Plexus Of sym pathetic neurons in the vill i Of the small intestine of a guinea-
pig : a, b, c, d , Neurons of the subepithelial p lexus ; e, f ,neurons of the plexus within the vill i ; g, fibers of the submucous (Meissii er’s )p lexus . (Af ter Ramon y Cajal . )synapses and the neurons are not po larized . Apparently thenervous im pulse may be transm itted equally well in all d i rec
tions throughout this network . The physiological propertiesOf such an arrangement appear to be very different from thoseOf the synaptic nervous system s Of h igher animals . A non
synaptic network sim i lar to that m entioned above has.been d escribed as occurring in som e Of the diffuse ganglionic plexuses ofthe human body (Fig.

54 INTRODUCTION TO N EUROLO GY
In the synaptic system s,as found in all high ly differentiated
nervous centers, the m aj ority of neurologists teach that at thesynapse the two neurons invo lved are sim ply in contact and thatthe nervous im pulse passes from one to the other across a veryshort gap in the conducting substance. Others believe thatthey have dem onstrated very delicate protoplasm ic threadswh ich bridge this gap, thus estab lish ing continui ty Of the con
ducting substance across the synapse . Good h istological preparations Show
,however
,in som e
'
Of the m ost intimate synapsesknown where the axon ends directly on the cel l body of the second neuron that there is a distinct cellular mem brane aroundthe term inals Of the fibers Of the first order and a second cellu
lar m em brane enveloping the body of the neuron of the secondorder, so that continui ty Of the ordinary protoplasm of theneurons here seem s to be quite im possible
,so far as our present
techni c is adequate to decide the question .
l
The following im port ant points regarding the synapse seemto be estab lished :1 . Unim peded protoplasm ic continuity across the synapse has
not been c learly estab lished,and in som e cases there is clearly
a m embranous barrier interposed between the two neurons .But the exact nature Of this barrier is unknown and it by no
m eans fol lows that the synaptic m em brane is an inert substance.
It m ay be com posed of living substance Of a different naturefrom that of the other protoplasm Of the neurons .2 . The transm ission Of the nervous im pulse across the synapse
invo lves a delay greater than that found in the nerve-fiber orthe cel l body . Th is suggests that there is som e sort of an Ob
struction here wh ich does not occur elsewhere in the reflex arc(see p .
3 . The synapse is m ore susceptible to certain toxic substances,
such as nicotin,than is any other part Of the reflex are.
4 . Though a nerve-fiber seem s to be capab le Of transm ittingan im pulse in either d irection
,the nervous im pulse can pass the
synapse only in one d irection,viz .
,the d irection of normal dis
charge from the axon of one neuron to the dendrite Of another .
1 For an illustration of such a synapse see BA RTL EME z, G . W .,
’Mauthner sCell and the Nucleus Motorius Tegm enti , Jour . Comp . Neur ., vol. xxv,
19 15 , Figs . 1 1 , 12, and 13 , pp . 1 26—1 28 .

THE NE URON 55
The synapse, therefore, acts as a sort of valve, to use a crudeanalogy, and appears to be one of the factors (not necessarilythe only one, see p . 97) in establish ing the polarity Of the neuron .
5 . Observations upon injured neurons show that the degenerations caused by the severance of their fibrous processes (whetherthese be m anifested as degeneration of the fibers or as chroma
tolysis, see p . 49 ) or by the destruction Of the cel l bodies fromwh ich the fibers arise cannot croSs the barriers interposed by thesynapses .Summary.
—In this chapter the form and internal structure Ofneurons have been briefly reviewed and the present status of theneuron doctrine is summ arized on p . 49 . The synapse is theplace where the nervous im pulse is transm itted from one neuronto another, and it is regarded as of the utm ost physio logicalimportance, its m ost im portant features being presented brieflyon p . 54 . The doctrine of the polarization of the neuron teachesthat nervous im pulses are received by the dendritic processesand transm itted outward from the cel l body through the axon.
LITE RATUREAPATHY , S. 1898 . Ueber N curofibrillen, Proc . Internat : ZoologicalCongress , Cam bridge, pp . 125—14 1 .
BA RKE R,L . F . 190 1 . TheN ervous System and Its Constituent Neurones,
N ew York.
BETHE,A . 1904 . Der h eutige Stand d er Neurontheorie, Deutsch .
m ed . Woch ., No . 33 .
GO LGI, C . 1 882—1885 . Sulla fina anatom ia degli organi centrali d elsistem a nervoso, R iv . Sperim . di Freniatria, vols . viii , ix, and xi .
1907 . La dottrina d el neurone, Teoria e fatti,Arch . Fisiol . , vol. iv,
pp . 187—2 15 .
HE IDE N HA IN,M . 19 1 1 . Plasm a und Zelle, 2 Lieferung (in Bard eleben ’
s
Handbuch d er Anatom ie d es Menschen,Bd . Jena .
HIS,W . 1889 . Die Neuroblasten un d deren Entstehung im em bry
oualen Mark , Leipzig .
ME YE R, ADO LF . 1898 . Critical R eview of the Data and General Meth
od s and Deductions of Modern N eurology,Jour . Com p . N eur., vol . viii ,
pp . 1 13—148 and 249—3 13 .
N ISSL,F . 1903 . Die Neuronenlehre und ihre Anh anger, Jena.
R AMON Y CA J AL, S. 1 909 . Histologie d e System e N erveux
,Paris .
WA LDE YE R,W . 189 1 . Ueber eini ge neuere Forschungen im Gebiete
d er Anatom i c d es Centralnervensystem s,Deutsch . m ed . Woch .
,Bd . 17 .

CHAPTER IV
THE REFLEX CIRCUITS
THE cellular uni t Of the nervous system,as we have seen
,is
the neuron. Neurons,however
,never function independently
,
but only when j oined together in chains whose connections arecorrelated with the functions wh ich they serve . Accordingly,the m ost im portant unit Of the nervous system
,from the phys
iological standpoint, is not the neuron,but the reflex ci rcuit, a
chain Of neurons consisting of a receptor or sensory organ,a cer
relating center or adjustor, and an effector or organ Of response,together w ith afferent and efferent nerve-fibers which serve as
conductors between the center and the receptor and effectorrespectively (see p . In a reflex c ircuit the parts must beso connected that upon stimulation of the receptive end -organ a
useful or adaptive response follows,such
,for instance
,as the
imm ed iate j erking away Of the hand upon accidentally touch inga hot stove .A reflex act, as this term is usually defined by the physio logists,
is an invariab le m echanically determ ined adaptive response tothe stimulation Of a sense organ,
invo lving the use Of a center Ofcorrelation and the conductors necessary to connect th is centerw ith the appropriate receptor and effector apparatus . The act
is not Voluntari ly perform ed,though one may becom e aware Of
the reaction during or after its perform ance.
The term “reflex” is Often popularly very loosely applied , butas generally used by physiologists it invo lves the rather com plexnervous function above described . If an electric shock is applied directly to a muscle or to the m otor nerve wh ich innervatesthat muscle
,the muscle w i ll contract, but th is di rect contraction
is not a reflex act . Many acquired m ovements have becom e sohabitual as to be perform ed quite automatically, such as theplay of the fingers Of an expert pianist or typist ; but these
56
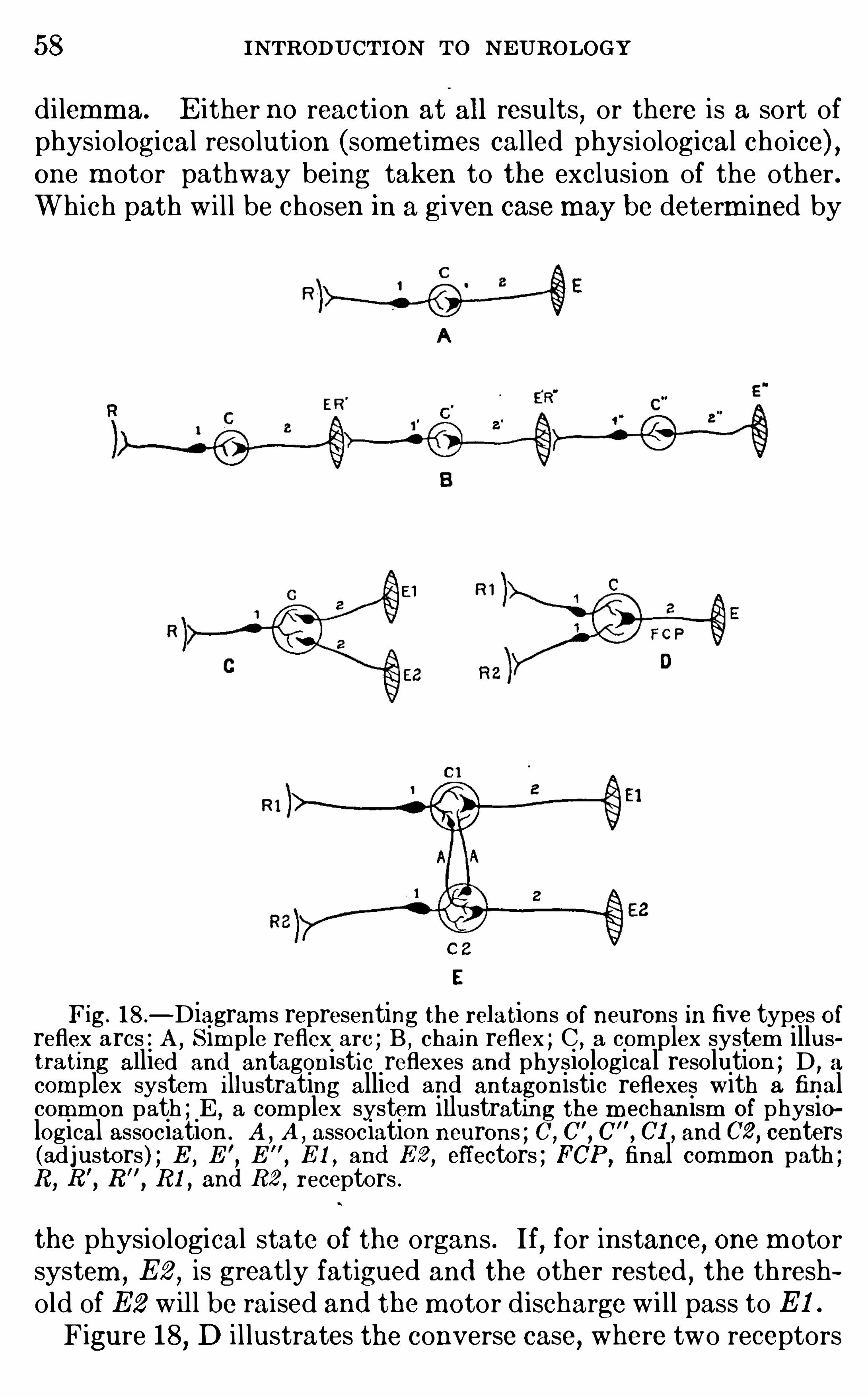
58 INTRODUCTION TO N EUROLOGY
di lemma. E ither no reaction at all results,or there is a sort of
physio logical reso lution (sometimes cal led physiological choice) ,one m otor pathway being taken to the exclusion of the other .Which path w i l l be chosen in a given case may be determ ined by
Fig. 18 .—Diagram s representing the relations of neurons in five types of
reflex arcs : A,Sim ple reflex are ; B , chain reflex ; C , a com plex system illus
trating allied and antagonistic reflexes and physiological resolution ; D ,a
com plex system illustrating allied and antagonistic reflexes with a finalcomm on path ; E , a com plex system illustrating the m echan ism of physiological association . A , A , association neurons ; C, C
'
, C$
, CI , and C2 , centers(adjustors) ; E , E
’
,E
”
,E 1 , and E 2 , effectors ; FCP ,
final common path ;R , R
'
, R”, RI , and R2 , receptors .
the physiological state of the organs . If, for instance, one m otorsystem
,E 2
,is greatly fatigued and the other rested , the thresh
old of E2 w il l be raised and the m otor d ischarge w i ll pass to E 1 .
Figure 18, D i l lustrates the converse case, where two receptors

THE RE FLEX CIRCUITS
discharge into a single center, which , in turn,
by m eans of a finalcomm on path (FCP ) excites a single effector (B ) . If the two receptors upon stim ulation normally call forth the same response
,
they w i l l reinforce each other if simultaneously stimulated,the response wi l l be strengthened, and we have another type Ofal lied reflex . But there are cases in wh ich the stimulation ofRI and R2 (Fig. 18
,D) would natural ly call forth antagonistic
reflexes . Here,if they are Sim ultaneously stimulated, a phys
iological di lemma w i l l again arise wh ich can be resolved only byone or the other afferent system getting control of the final common path .
Figure 18, E i llustrates sti l l another form of com bination ofreflexes . Here there are connecting tracts (A ,
A ) between thetwo centers so arranged that stimulation of either of the tworeceptors (RI and R2 ) m ay cal l forth a response in either one oftwo effectors (E 1 and E 2) . These responses m ay be al lied orantagonistic , and much m ore com plicated reflexes are here posSible than in any of the preced ing cases .A few i l lustrations of the practical Operation of these typesof reflex circuits w i ll be given here and many other cases are
cited throughout the following discussions . A case‘
of a sim plereflex has already been mentioned in the sudden twitch of thehand in response to a painful stim ulation of the Skin . The
sim plest possib le m echanism Of th is reaction involving only twoneuronsis shown in Fig. 1 (p . In actual practice, however,the arrangement figured is one elem ent only of a m ore com plexreaction (see p . Figure 19 i llustrates a m ore usual formof this type Of reaction
,where a series of three or m ore neurons
is involved and at least two cerebral centers . An auditory impulse com ing to the brain from the ear through the VII I cranialnerve term inates in a primary acoustic center in the superiorO l ive (a deep nucleus of the m edulla Ob longata, see p .
where it is taken up by an intercalary neuron of the second orderand transm itted to the nucleus Of the VI nerve. The result is acontraction of the external rectus muscle of the eyeball , turningthe eye toward the side from wh ich the aud itory stim ulus wasreceived . So far as th is reaction alone is concerned
,it is a sim ple
reflex , but in practice the external rectus m uscle of one eye isnever contracted apart from the other five muscles of that eye
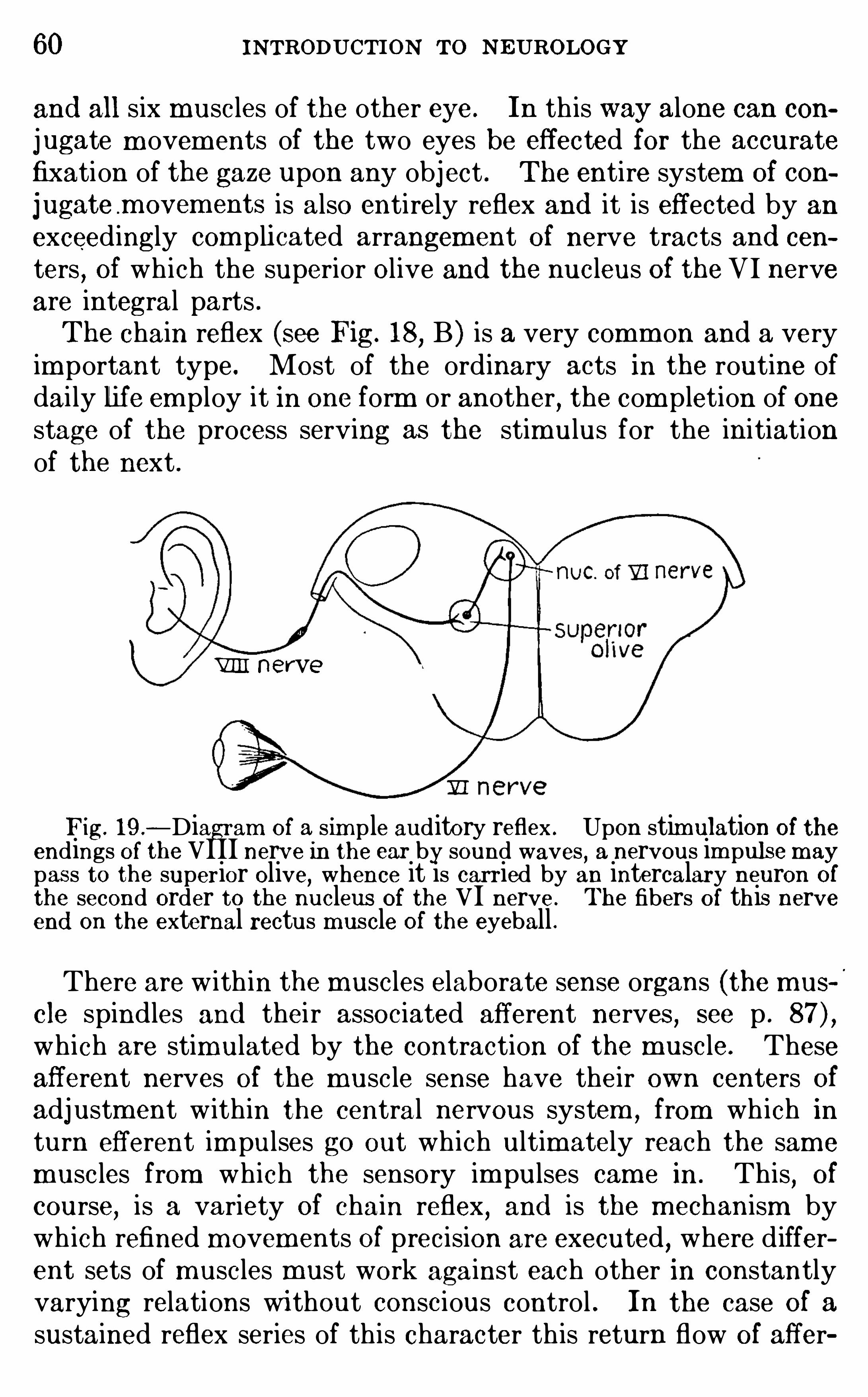
60 INTRODUCTION TO N EUROLOGY
and all six muscles of the other eye. In th is way alone can con
jugate m ovem ents of the two eyes be effected for the accuratefixation of the gaze upon any object . The entire system of con
jugatem ovem ents is also entirely reflex and it is effected by anexceedingly com pli cated arrangement of nerve tracts and centers
, of wh ich the superior O live and the nucleus of the VI nerveare integral parts .The chain reflex (see Fig. 18
,B ) is a very comm on and a very
im portant type . Most of the ordinary acts in the routine ofdai ly life em ploy it in one form or another, the com pletion of onestage of the process serving as the stimulus for the initiationof the next .
VIII nerve
Fig. 19 .
—Diagram of a sim p le auditory reflex . Upon stim ulation of theendings Of the VIII nerve in the ear by sound waves , a nervous im pulse maypass to the superior olive, whence it is carried by an intercalary neuron of
the second order to the nucleus of the VI nerve . The fibers of this nerveend on the external rectus muscle of the eyeball .
There are w ith in the muscles elaborate sense organs (the m us
cle spindles and thei r associated afferent nerves,see p .
which are stim ulated by the contraction of the muscle. Theseafferent nerves of the m uscle sense have their own centers ofadjustment w ithin the central nervous system ,
from wh ich inturn efferent im pulses go out wh ich ultimately reach the sam e
musc les from which the sensory im pulses cam e in . This,of
course,is a variety of chain reflex
,and is the m echanism by
wh ich refined m ovem ents Of precision are executed , where d ifferent sets of m uscles must work against each other in constantlyvarying relations wi thout conscious control . In the case of a
sustained reflex series of this character th is return flow of affer

THE RE FLEX CIRCUITS 6 1
ent im pulses of the m uscle sense,tendon sense, etc .
,exerts a
constant influence upon the center wh ich receives the ini tialstimulus
,so that th is center is constantly under the comb ined
influence of the external stimulus which sets the reflex in m otionand the internal stim uli arising from the m uscles them selves(propri oceptors, see p . 86) wh ich control its course. In
‘
th iscase there is a true physio logical circuit rather than an are orsegm ent of a circuit
,as is comm only im plied in the expression
“reflex are.
” Thi s case is typical of the com plex reflexes of thebody in general , and for this and other considerations we followthe usage of Dewey (1893) and term the m echanism of a com
plete reflex a“reflex circuit” rather than an arc (see C . J .
Herrick,19 13
,and p .
I t has been suggested by Loeb also that m any instincts aresim ply com plex chain reflexes . Even in animals whose behav ior
\ is so com plex asabird s
,a careful analysis of the cycle Of nest
bui lding and rearing of young reveals m any c lear illustrat ions ofthis principle (see the works of F . H. Herri ck
,cited at the end of
th is chapter) . Each step in the cycle is a necessary antecedentto the next , and if the series is interrupted it is often necessaryfor the b irds to go back to the beginning of the cycle. Theycannot make an intelligent adjustm ent m idway of the series .The com plex circuit i llustrated by Fig. 18
,C presents two
possible types of reaction, either all ied or antagonistic reflexes .The form er case is illustrated again by the sudden m ovem ent ofthe hand in response to a painful stim ulation of the skin. Th isis brought about, as we saw in considering the sim ple reflex , by acontraction of the arm m uscles . But the m usc les which m ovethe elbow- joint are not, when the arm is at rest, entirely flaccid .
Both flexors and extensors are always contracted to a certaindegree , one balanced against the other . N ew at the sam e tim e
that the sensory stimulus from R (see Fig. 18, C ) causes the contraction of the flexor m uscle
,E 1
,it also causes the relaxation of
the antagonistic extensor, E 2 the two efferent im pulses cooperating to effect the avoiding reaction as rapid ly as possib le. In
the antagonistic reflexes Of our third type the physiological resolution invo lved in the selection of one or the other possiblereaction always involves a delay in the response unti l one m otorpathway dom inates the system to the exclusion of the other .
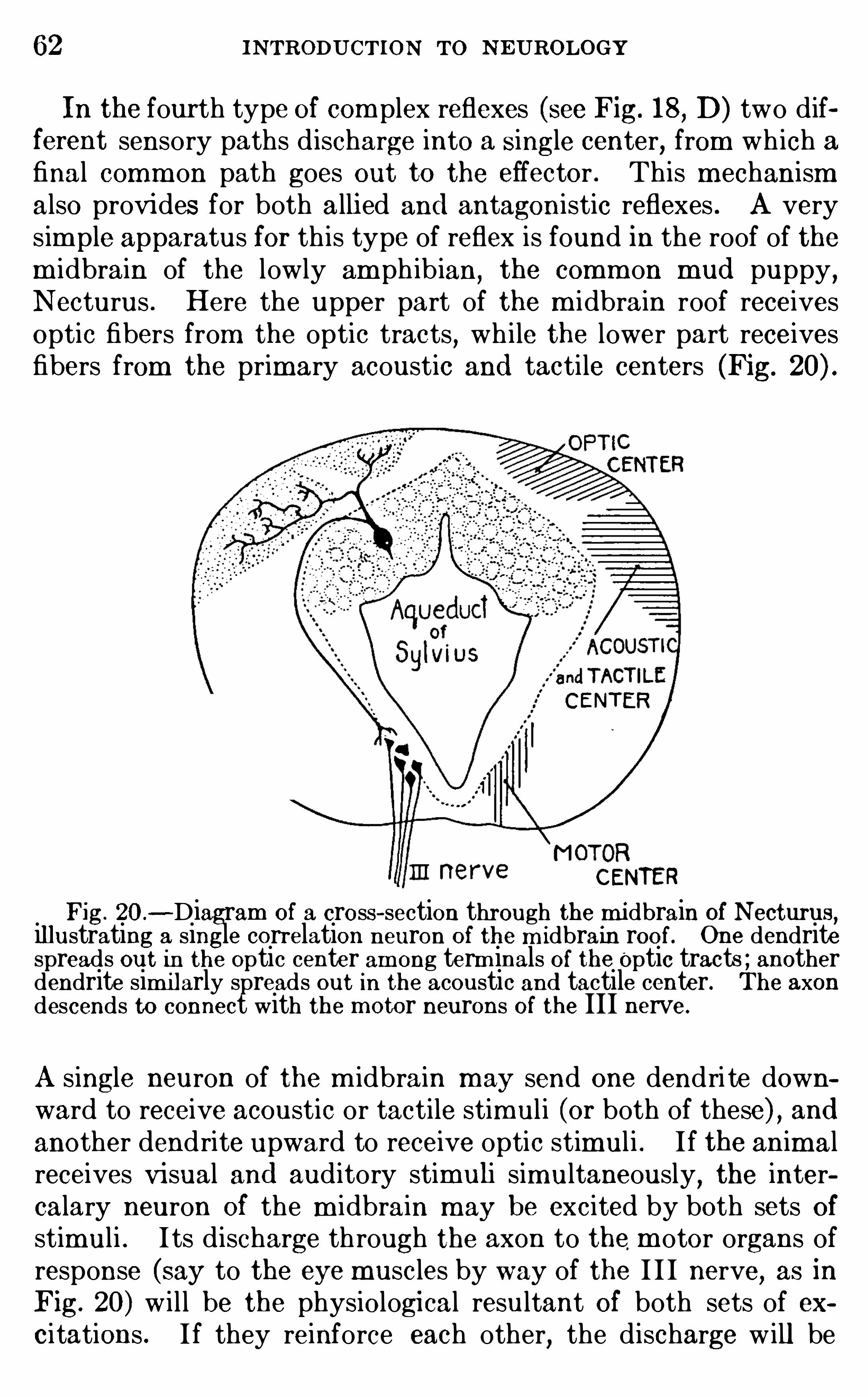
62 INTRODUCTION To N EUROLOGY
In the fourth type of com plex reflexes (see Fig. 18,D) two dif
ferent sensory paths d ischarge into a Single center, from wh ich afinal comm on path goes out to the effector . This m echanismalso provi des for both alli ed and antagonistic reflexes . A verysim ple apparatus for th is type of reflex is found in the roof of them idbrain of the lowly am ph ib ian
,the comm on mud puppy
,
Necturus . Here the upper part of the m idbrain roof receivesoptic fibers from the optic tracts , wh i le the lower part receivesfibers from the primary acoustic and tacti le centers (Ifig.
CENTER
Fig. 20 .—Diagram of a cross—section through the m idbrain of Necturus,
illustrating a single correlation neuron of the m idbrain roof . One dendritespreads out in the optic center am ong term inals of the Optic tracts ; anotherdendrite sim i larly spreads out in the acoustic and tactile center. The axondescends to connect with the m otor neurons of the III nerve.
A single neuron of the m idbrain may send one dendri te downward to receive acoustic or tactile stim ul i (or both of these) , andanother dendrite upward to receive optic stim ul i . I f the anim alreceives vi sual and auditory stimuli simultaneously
,the inter
calary neuron of the m idbrain m ay be excited by both sets ofstimuli . I ts discharge through the axon to them otor organs ofresponse (say to the eye muscles by way of the I I I nerve, as inFig. 20) w i l l be the physiologi cal resultant Of both sets of excitations . I f they reinforce each other
,the discharge w ill be
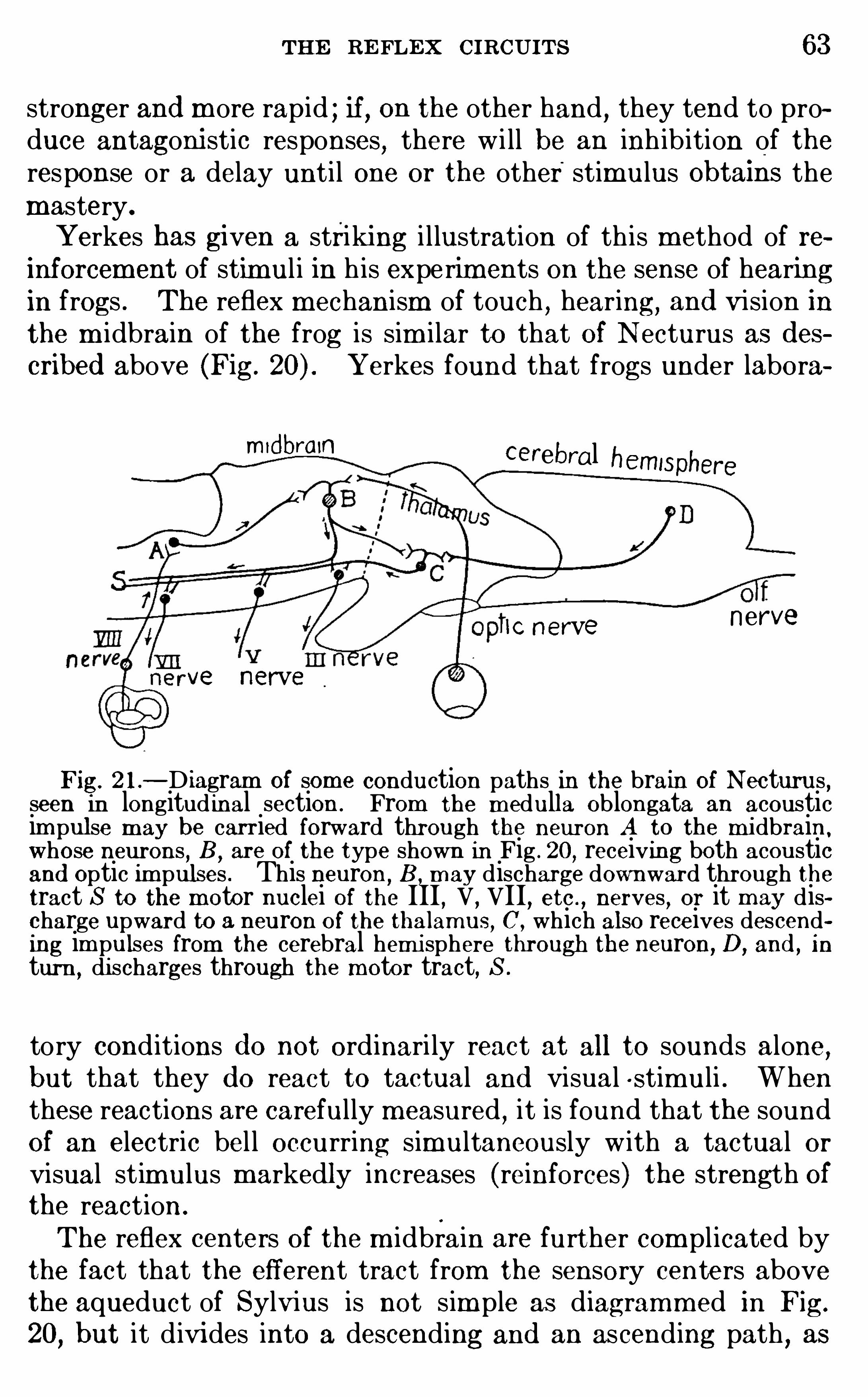
THE RE FLEX CIRCUITS 63
stronger and m ore rapid ; if, on the other hand , they tend to produce antagoni stic responses, there w i ll be an inh ib ition of theresponse or a delay unti l one or the other stimulus ObtaIns themastery.
Yerkes has given a striking i l lustration of th is m ethod of reinforcement of stimul i in h is expe rim ents on the sense of hearingin frogs . The reflex m echanism of touch , hearing, and vision inthe m idbrain of the frog is sim i lar to that of N ecturus as d escribed above (Fig. Yerkes found that frogs under labora
mid brain cerebral hemisphere
lC nerve nerve
ve nerve
Fig. 2 1—D iagram of som e conduction paths In the brain of Necturus ,seen in longitud inal section . From the m edulla Oblongata an acousticim pulse may be carried forward through the neuron A to the m idbrainwhose neurons
,B
,are of the type shown in Fig . 20 , receiving both acoustic
and optic im pulses . This neuron,B , m ay discharge downward through the
tract S to the m otor nuclei of the III,V
,VII
, etc . , nerves , or it m ay discharge upward to a neuron of the thalam iis
, C , which also receives descending im pulses from the cerebral hem i sphere tlirough the neuron, D, and , i n
turn , di scharges through the motor tract, S.
tory cond itions d o not ordinari ly react at all to sounds alone,
but that they d o react to tactual and visual s timuli . Whenthese reactions are carefully m easured
,it is found that the sound
of an electric bel l occurring simultaneously w ith a tactual orvisual st im ulus markedly increases (reinforces) the strength ofthe reaction .
The reflex centers of the m idbrain are further com plicated bythe fact that the efferent tract from the sensory centers abovethe aqueduct of Sylvius is not sim ple as diagramm ed in Fig.
20,but it d ivides into a descending and an ascending path , as

64 INTRODUCTION TO N EUROLOGY
shown by the neuron B of Fig. 21 . The descending pathconnects directly w ith m otor centers, including the oculom otor,bulbar
,and spinal m otor nuclei (Fig. 21
, S) , wh ile the ascendingpath enters the thalamus
,where associations of a sti l l higher
order are effected through the thalam ic neuron, C. Here again isintroduced a physiological choice or d ilemma ; the response is nota sim ple m echanical resultant of the interacting stimuli , but itscharacter may be influenced by variable physiological states .The invariable type of action is replaced by a relatively variableor lab i le type (see p . In the thalam us the nervous im pulseis again subj ected to m odification under the influence of a sti l l
greater variety of afferent im pulses,for these centers receive all
sensory types found in them idbrain,and
,in addition
,im portant
descending tracts from the cerebral hem ispheres (in lower vertebrates the latter are ch iefly O lfactory) .The m ore com plicated associations are effected by arrange
m ents of correlation tracts and centers i l lustrated in the sim plestpossib le form by Fig. 18
,E . The m ode of operation of such
a system may be i l lustrated by an exam ple : A col lie d ogwh ich Ionce owned acquired the hab it of round ing up my neighbor
’sSheep at very unseasonable tim es . The sight Of the flock in thepasture (stimulus RI , Fig. 18, E ) led to the pleasurab le reaction(E 1 ) Of chasing the sheep up to the barnyard . I t becam e necessary to break up the hab it at once or lose a valuable d og at the
hands of an angry farm er w ith a shotgun . Accord ingly, Iwalked out to the pasture with the d og. She at once brought inthe sheep of her own accord and then ran up to m e w ith everyexpression of canine pride and self—satisfaction, whereupon Iimm ediately gave her a severe wh ipping (stim ulus R2) . Th iscalled forth the reaction (E2 ) of running home and h iding in herkennel . The next d ay (the d og and I having m eanwh i le withmutual forgiveness again arrived at friendly relations) we took awalk in a d ifferent d irection ,
in the course of wh ich we unex
pected ly met another flock of sheep . At sight of these the d ogimm ediately, w ith no word from me
, put her tai l between herlegs , ran hom e as fast as possible, and h id in her kennel . Herethe stimulus RI led not to its own accustomed response
,E 1
,
but to E2 , evidently under the influence of vestigeal traces of theprevious day ’s experience, wherein the activities of C1 and C2

66 INTRODUCTION To NEUROLOGY
reflex path ; but the stimulus is rarely sim ple and the nervousd ischarge irrad iates m ore or less w idely, so that the activity isby no means l im ited to the part which gives the act its reflexpattern . Moreover, neither the stimulus com plex nor the character of the irradiation wi ll be repeated exactly in any h igheranimal , so that theprecise nature of the response cannot in anycase be infallibly predicted except under experim ental conditions(and not always then) .Our picture of the reflex act in a h igher animal wi ll
,then
,
include a view of the whole nervous system in a state of neuraltension. The stimulus d isturbs the equi librium at a defin itepo int (the receptor) , and the wave of nervous discharge thus setup irradiates through the com plex lines determ ined by the neuralconnections of the receptor. If the stimulus is weak and the
reflex path is sim ple and wel l insulated, a sim ple response m ay
follow immediately. Under other conditions the nervous discharge may be inhib ited before it reaches any effector, or it may
irrad iate widely,producing a very com plex reflex pattern. In
the former case the neural equi librium w i ll be only local lydisturbed ; in the latter case alm ost the who le nervous systemmay part ic ipate in the reaction,
a part focal and sharply definedand the rest marginal , diffuse, and exercising m ore or less ofinh ibitory or reinforcing control on the final reaction.
The studies of Herrick and Cogh i l l have shown that in thedevelopm ent of the nervous system Of Am ph ibia the first reflexcircuits to come to maturi ty are made up of rather com plexchains of neurons so arranged as to perm it of only one type ofresponse
, vi z .,a total reaction (the swimm ing m ovem ent) , from
all possible form s of stim ulation,and that in successive later
stages th is general ized type is gradual ly replaced by a series ofSpecial reflexes involving m ore d iversified m ovem ents . Parallelw ith th is process the h igher correlation centers are developedfor the integration of the several special reflexes into com plexaction system s . The sim ple reflex are
,as i l lustrated in Fig.
1 (p . wh ich is adapted for the execution of a singlem ovem entin response to a part icular stimulus
,is the final stage in th is
developm ental process, whose initial stages are much m orecom plex and diffuse arrangements of neurons adapted for totalreactions of a m ore general sort.

THE RE FLEX CIRCUITS 67
We have just described the m echanism s of certain reflexes .The question at once arises
,In what sense do we know the
mechanism of a nervous reaction? Certainly not in the sensethat we understand all of the factors involved in nervous conduetion and correlation . But we do have a practical know ledge Of
the comb inations of neurons necessary to effect certain definiteresults
,much as the practical electrician may be ab le to wind a
dynam o or bui ld a telephone,even though h is know ledge of the
theory of electricity be very smal l .Summary.
—The reflex arcs or reflex circuits rather thanthe neurons of wh ich these circuits are com posed are, fromthe physiological standpo int , the m ost im portant units of thenervous system . Reflex acts are to be
'
d istinguished , on the
one hand, from the Sim pler non-nervous reactions known as
tropism s and taxes , and, on the other hand
,from voluntary
acts and acquired automatism s . Many instincts are chainreflexes of very com plex sorts
,the com pletion of one reaction
serving as the stimulus for the next, and so on in series. Thesim plest true reflex requires a receptor
,a center or adjustor
,
an effector,and the afferent and efferent conductors wh ich put
the center into physio logical relation w ith the receptor and theeffector respectively . Five types Of reflex circuits were d istin
guished (see Fig. 18) and i llustrations of them given . All of thereflex centers are interconnected by system s of fibers
,either in
the form of definite tracts or else by m ore d iffuse connectionsin the neuropil . Localization of cerebral function is , therefore,only approximate
,w ith the possib i lity of all sorts of intercon
nection of d ifferent reflex system s as occasion m ay require .This is the neuro logical basis of the greater plasticity ofbehavior of higher vertebratesas contrasted w ith invertebratesand lower vertebrates .
LITE RATUREDEWE Y
,J . 1893 . The Reflex Arc Concept in Psychology , Psycho] .
R eview, vol . iii , p . 357 .
HE RRICK, C . JUDSON . 19 13 . Som e R eflections on the Origin and Significance of the Cerebral Cortex , Jour . of An imal Behavior, vol . ii i , pp . 222
HERRICK , C . JUDSON and CO GHILL , G . E . 19 15 . The Developm ent ofReflexMechanism s in Am blystoma
,Jour . Com p . Neur .
,vol. xxv, pp . 65—85 .

68 INTRODUCTION TO NEUROLOGY
HE RRICK,F . H . 1905 . The Hom e Life of Wild B irds . Revised edition,
New Yor
1907 . Analysis of the Cycli cal Instincts of Birds, Science, N . S., vol.
xxv,pp . 725 , 726 ; and Jour. Com p . N eur .
, vol . xvii , pp . 194, 195 .
190 The B lending and Overlap of Instincts, Science, N . S., vol .xxv
,pp . 781 , 782 ; and Jour . Com p . N eur .
,vol. xvii , pp . 195—197 .
The Relation of Instinct to Intelligence in B irds, Science,N . S. ,
.
vol . xxvi i , pp . 847—850 .
HOUGH , TH. 19 15 . The Classification of Nervous Reactions, Science,N . S ,
vol . xli , pp . 407—4 18 .
JE N N IN GS, H . S. 1905 . The Bas is for Taxis and Certain Other Term s Inthe Behavior of Infusoria, Jour . Com p . N eur .
,vol . xv
,pp . 138—143 .
1906 . The Behavior of Lower Organism s, N ew York .
LO E B,J . 1900 . Com parative Physiology of the Brain and Com parative
Psychology, N ew York .
19 12 . The Mechanistic Conception of Life, Chi cago.
SHE RRIN GTON ,C . S. 1906 . The Integrative A ction of
agthe Nervous Sys
tem , N ew York .
YE RKE S, R . M . 1905 . TheSense of Hearing in Frogs, Jour. Com p . Neur .,
vol . xv, pp . 279—304 .

CHAPTER V
THE RECEPTORS AND EFFE CTORS
IN the further study of the nervous system as the apparatusof adjustm ent between the activities of the body and those Of
environing nature, our first task is the analysis of the receptors
(that is, the sense organs) ; for these are the only places throughwh ich the forces of the world outside can reach the nervous system in order to excite its activity .
Theworld is so full of a num ber of thingsI’
m surewe should all be as happy as kings .
But in order to attain th is fortunate result it is necessary that weshould be able to discrim inate the essential from the unim portantelem ents of th is environing com plex
,and to adjust our own be
havior in relation thereto .
Protoplasm in its sim plest form is sensitive to some sorts ofm echanical and chem ical stim ulation . In fact
,as we have seen
,
all of the so- called nervous functions are im plicit in und ifferen
tiated protoplasm . But the bodi es of all but a few of the lowestorganism s are protected by som e sort Of a shel l or cuticle fromexcessive stim ulation from the outside
,and individual parts of
the surface are then differentiated in such a way as to be sensitive to only one group of excitations wh ile remaining insensitiveto all other form s . Thus arose the sense organs , each of whichconsists essential ly of special ized protoplasm wh ich is h igh lysensitive to som e part icular form of energy manifestation, butrelatively insensitive to other form s Of stimulation . Each senseorgan possesses, in addition
,certain accessory parts, adapted to
concentrate the stimul i upon the essential sensitive protoplasm ,
to intens ify the force of the stimulus, or to so transform the69

70 INTRODUCTION To NEUROLO GY
energy of the stimulus as to enab le it to act m ore efficiently uponthe essential end - organ.
Sherrington states the distinctive characteristic of the senseorgans in th is form ,
“The main function of the receptor is
,there
fore,to lmIIer the threshold of exci tabi li ty of the are for one kind
of stimulus and to heighten i tfor all others .
”The selective func
tion of the receptors is i l lustrated by a consideration of thed ifferent form s of vibratory energy wh ich pervade the envi ronment in which we live .There are
,first
,rhythm ically repeated mechanical im pacts
perceived through the sense of touch . Th is series of tacti lesensations extends from a single isolated contact at one extreme
to rhythm ically repeated contacts touching the skin as fre
quently as 1552 vibrations per second .
A second series of v ibratory phenomena is presented by them echani cal vibrations of the surroundingm edium perceived sub
jectively as sound . Out of the entire series of such vibrations ofall possib le frequencies the human ear is sensitive to a series ofapproxim ately ten octaves from '
about 30 (in some cases 12) toabout (in som e cases vibrations per second (wavelengths from 1228 cm . or 40 ft . to cm . or .5 inch in length ) .To all other vibrations it is insensitive. W ith in this range theaverage human ear can d iscrim inate som e different pitchqual ities (Titchener) .Subjectively, the series of tone sensations is broken up into
a number of octaves, and it is found that a given tone of themusical scale is excited by vibrations of exactly twice the frequeney wh ich excites the correspond ing tone of the next loweroctave . By analogywith th is arrangem ent all series of physicalv ibrations are sometim es spoken of as d ivi sible into octaves,the octave being defined as those vibration frequencies wh ichlie between a given rate and twice that rate or half thatrate .
A th ird type of vibratory phenom ena is presented by themuchm ore rapid series of so- called ethereal vibrations
,or waves in
imm aterial m edia. The lower m embers of thi s series are theHertz ian electri c waves ; the h igher members are the tic- rays .Between these extrem es lie waves perceived as radiant heat
,the
light waves, and the ultra-Violet rays Of the spectrum . Th is

THE RE CE PTORS AND E FFE CTORS 7 1
series of ethereal vibrations m ay extend farther indefini tely bothdownward and upward
,but of its ultimate lim its we have no
know ledge.
There is no human sense organ whi ch can respond directly tothe electric waves
,the ultra-violet rays, and the x- rays .
These have, accordingly, remained whol ly unknown to us unti lrevealed indirectly by the researches of the physical laboratories.Som e ten octaves of th is series are contained in the so lar spectrum
,from an infra-red wave length of about .1 mm . to an ultra
violet wave length of .00035 mm . The light from m etallic arcsand from incandescent gases has, however, been found to containwave lengths as short as .00006 mm . The hum an eye is sensitive to som ething over one octave of th is series (waves from.0008 to .0004 mm . in length , whose rates lie betweenand b i llions of vibrations per second) , with six octavesin the infra- red and three in the ultra-violet . The lower m em
bers of this'
series of Vibrations of the solar spectrum ,and to a
less extent the h igher also , are capab le of stim ulating the tem
perature organs of the skin .
Thus it appears that of the com plete series of ethereal vibrations, we can sense directly only about one octave by the eye anda num ber of others through the sense organs for tem perature inthe skin, whi le to the lowest and highest m em bers of the seriesour sense organs are entirely insensitive . The sensitivi ty of theskin to these vibrations is lim ited subjectively to a smal l rangeof tem perature sensations
,whi le the retinal excitations give us
subjectively an extens ive series of sensations of color and brightness . The human eye can di scri m inate from 150 to 230 purespectral tints
,bes ides various degrees of intensity and purity of
tone, making a total of between and possib lediscrim inations by the visual organs (von KIi es) . Som e of thepreceding data are summarized in the tab le1 on page 73 .
.
1
In the preparation of th is table I have been assisted by Professor R . A .
MIJhkan, of the University of Chi cago ,
whose kindness I gratefully ac
knowledge. The figures given are based upon the formula
and the velocity of transm ission is taken as 3 x 101° cm . per second . Theactual veloc ity of li ght waves as worked out experimentally by Michelson18 kilom eters per second .
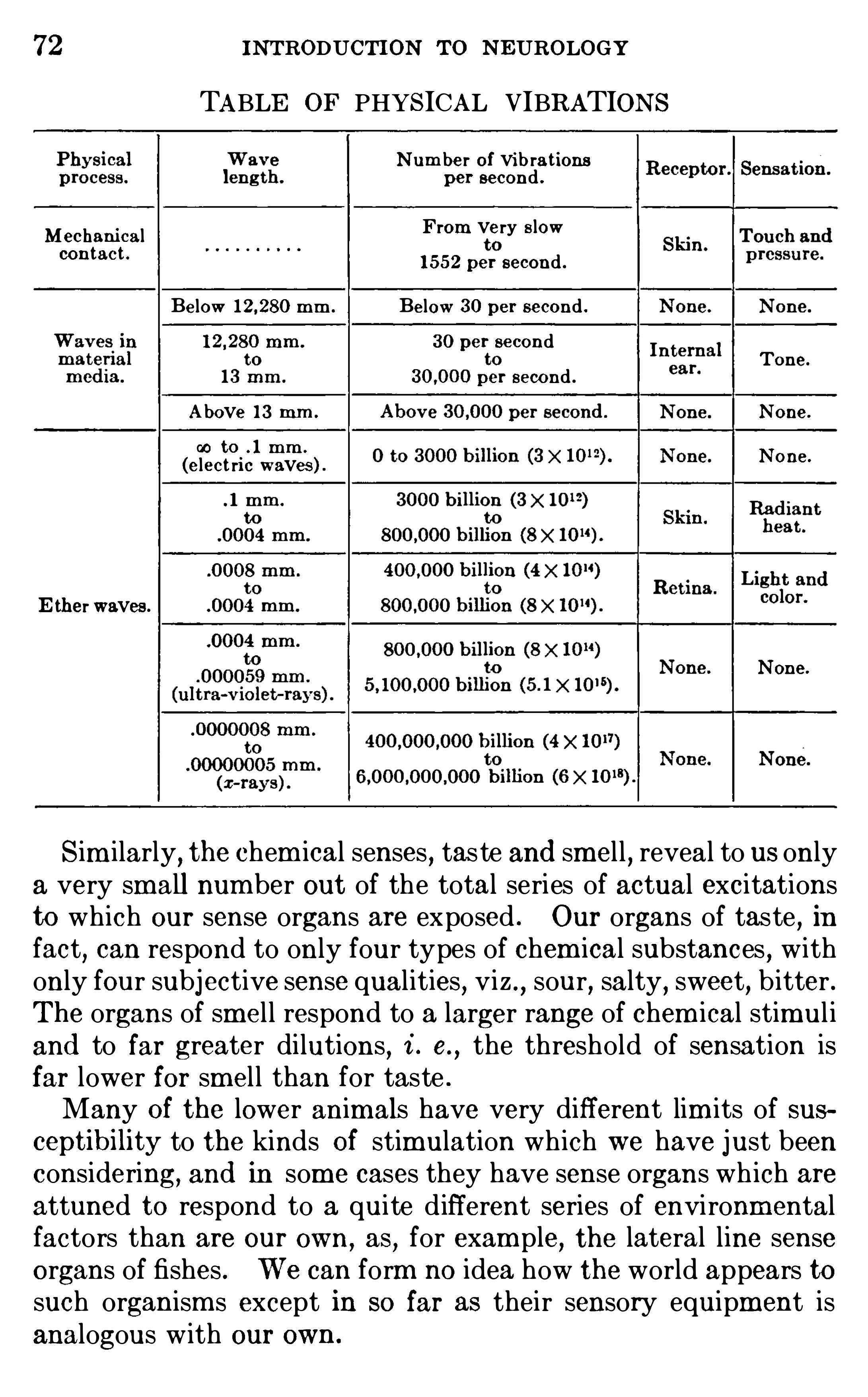
72 INTRODUCTION TO NEUROLOGY
TABLE OF PHYSICAL VIBRATIONSN um ber of vib rations
per second .From very s lowMechani cal
contac t .1 552 per second .
Below 1 2 280 m m . Below 30 per second .30 per second
Tone.
per second .A bove 13 mm . None.
00 to . 1 mm .
(elect ric waves ) . None.
E ther waves .
None.
Sim ilarly, the chem ical senses , taste and smel l , reveal to us onlya very small number out of the total series of actual excitationsto which our sense organs are exposed . Our organs of taste , infact
,can respond to only four types of chem ical substances , with
only four subjective sense qualities, viz ., sour, salty , sweet, b itter .The organs of smel l respond to a larger range of chem ical stimul iand to far greater di lutions , i . e.
, the threshold of sensation isfar lower for sm el l than for taste .
Many Of the lower animals have very different lim its of susceptibility to the kinds Of stimulation wh ich we have just beenconsidering, and in some cases they have sense organs wh ich areattuned to respond to a quite d ifferent series of envi ronmentalfactors than are our own,
as,for exam ple
,the lateral line sense
organs of fishes . We can form no idea how the world appears tosuch organism s except in so far as their sensory equipment isanalogous w ith our own.

4 INTRODUCTION To N EUROLOGY
w indows so as to look out d irectly into new fields ; but we haveincreased the range of vision through the old w indows, m uch as atelescope brings rem ote Objects near and as a periscope enab lesthe observer to see around a corner . To the developm ent ofthe cerebral cortex we owe the acquisition of these new powerswh ich have opened to us the realm s of electric v ibrations
,
ultra- violet rays , and many other natural phenomena to wh ichour unaided sense organs are quite insensitive.
Chi ldren in the kindergarten are taught that there are five
senses . In real ity, there are m ore than twenty different senses .Som e Of the sense organs are stim ulated by external objects andhence are term ed exteroceptors ; others are stim ulated by internalexcitations of the v isceral organs and are term ed interoceptors.
Stil l further classifications have been suggested , to wh ich reference w i ll be made short ly .
,
Here we must first consider thecriteria in accordance with wh ich the various senses are d istinguished .
The analysis and c lassification of the senses is by no m eansso simple a task as one m ight at first suppose. It is true thatordinari ly we do not confuse a thing
‘
Seen w ith a sound heard ;but, on the other hand , we do constantly confuse savors w ithodors , and it often requires refined physiological experimentationto determ ine whether the organ of taste or the organ Of sm el l isthe source of the sensory excitation in question. Most of thecomm on
“flavors” of food are, in real ity , Odors and are perceivedby the organ Of sm el l only . A bad cold wh ich closes the posterior nasal passages makes all food taste alike” for th is reason .
In real ity,as we have already seen
,there are only four tastes
recognized by the physiologists , viz . , sweet, sour, salty, and
b itter .Confusion has arisen in the attem pts to analyze these two
senses from the fact that different physiologists have used different definitions Of a sense.
”One author
,who defines these
senses in term s of the physical agents wh ich excite them,says
that taste is stimulated by liquids and sm ell by vapors,and that
,
accordingly, aquatic animals,whose nostri ls are fi lled with water
,
have by definition no sense of sm ell . Other authors separatethese senses accord ing to the organ stimulated , the excitation ofthe nose being smell
,that of the taste-buds being taste, regard

THE RE CE PTORS AND E FFE CTORS 75
less of the nature of the exciting substance or of the subjectivequal ity of the sensation .
There are,in reality
,four different factors which must be
taken into account in defining a“sense. ( 1 ) Doubtless w ith us
human fo lk the m ost im portant criterion is direct introspectiveexperience
,the psychologi cal cri teri on. Ordinari ly th is is ad e
quate,but, as we have just seen,
there are som e cases whereit alone cannot be depended upon to distinguish between twosenses . (2) The adequate stimul i of the various senses exh ibitcharacteri stic physical or chem ical differences, the physi cal
cri teri on. Th is factor,too
,must be careful ly investigated or we
may be led astray . (3) The data of anatomy and experimentalphysio logy m ay differentiate structurally the receptive organsand conduction paths Of the several types Of sensation,
the ana
tom i cal cri terion . (4) Final ly, the type of response varies in a
characteristic way for the different senses, the physiologi cal
cri teri on.
The fourth criterion has been applied to solve the problem ofthe reason for the developm ent of two very different types ofsense organs and cerebral connections for the senses of sm el l andtaste
,both of wh ich are chem ical senses w ith sim i lar subjective
qual ities . It has been pointed out by Sherrington that taste isan interoceptive sense, cal ling forth vi sceral responses w ith in thebody, wh ile sm el l is
,in part at least
,an exteroceptive sense, being
excited by Objects at a distance from the body and calling forthm ovements of locom otion carrying the whole body toward or
away from the source of the odorous emanations . Thus theform of the response is here the d istinctive factor, and incidentalto this feature the organs of sm el l are sensitive to far smallerquantities of the stimulating substance than are the taste-buds .Parker and Stabler have shown that the human organ of sm el lis sensitive to alcohol at a d i lution tim es greater than thatnecessary to stimulate the organs of taste (see p .
I t is im possible in the present state of our knowledge to frame
adequate definitions of all the senses in term s Of any one of
these four criteria alone , although it is a reasonab le hope thatth is m ay at some future tim e be attained . Even when all of
these criteria are taken into account , it is by no means easy todeterm ine how many separate senses the normal human being

76 INTRODUCTION To NEUROLOGY
possesses . Not only is there a considerable number of senseorgans not represented at all in our trad itional list of five senses
,
but several of these five are com plex . Thus,the internal ear
includes two quite d istinct organs—the coch lea, wh ich serves asa receptor for sounds
,and the labyrinth
,whose sem icircular
canals serve as the ch ief sense organs concerned in the regulation of bod i ly position and the maintenance of equi librium
,func
tions wh ich are quite d istinct from hearing. The skin,too
,
serves not only as the ch ief organ of touch , but also the add itional functions of response to warm , cold , and painful im pressions, besides som e other m ore obscure sensory activities
,such
as tickle.
An acceptable classification of the sense organs or receptorsof the body must take account of their anatom ical relations
,of
the nature of the physical or chem ical forces wh ich serve as theadequate stimul i
,of the subjective qualities wh ich we experience
upon their excitation,and of the character of the physiological
responses wh ich comm only fol low their stimulation. The lastpoint has been too much neglected .
In fact,the m ost fundam ental d ivision of the nervous sys
tem wh ich we have,cutting down through the entire bodi ly
organization ,is based upon this physiological criterion. From
th is standpoint we divide the nervous organs into two greatgroups : ( 1 ) a somatic group pertaining to the body in generaland its relations w ith the outer environment
,and (2) a vi sceral ,
splanchnic , or interoceptive group . The latter group com prisesthe nerves and nerve-centers concerned ch iefly w ith digestion,
respiration,circulation
,excretion,
and reproduction . These are
intimately related w ith the sym pathetic nervous system and
those parts of the central nervous system directly connectedtherew ith
,though the m ore high ly special ized members of th is
group are independent of the sym pathetic system . The somatic
group com prises the greater part of the brain and spinal cord andthe cranial and spinal nerves, or, briefly, the cerebro- spinal nervous system as distinguished from the sym pathetic system (seep . Th is is the m echanism by wh ich the body is able to adjust its own activities d irectly in relation to those of the outsideworld—to procure food
,avoid enem ies, and engage in the
pursuit of happiness .

THE RE CE PTORS AND E FFE CTORS 77
The organs belonging to each of these two groups do much oftheir work independently of the other group, i . e.
,visceral stimul i
cal l forth visceral responses and external or somatic stimulical l forth somatic responses . Nevertheless, the two groups oforgans are by no m eans entirely independent
,for external excita
tions m ay produce strong visceral reactions, and conversely .
Thus,the sight of luscious fruit (exteroceptive stimulus) natu
ral ly cal ls forth m ovements of the body (somatic responses) to goto the desired Obj ect and seize it . But if one is hungry, them outh may water in anticipation, a purely v isceral response.
On the other hand , the strictly visceral (interoceptive) sensationof hunger is apt to set in m otion the exteroceptive reactionsnecessary to find a dinner .Sherrington, whose analysis w ith som e m odifications is here
adopted,recogni zes three types of sense organs or receptors :
( 1 ) the interoceptors, or visceral receptive organs, wh ich respondonly to stimulation arising w ith in the body, ch iefly in connectionw ith the processes Of nutrition,
excretion,etc . ; (2) the extero
ceptors, or somatic sense organs, wh ich respond to stimulationarising from objects outside the body ; (3) the propri oceptors, asystem of sense organs found in the muscles, tendons, j oints,etc .
, to regulate the m ovem ents cal led forth by the stimulationof the exteroceptors . Th is th ird group is real ly subsidiaryto the somatic group , or exteroceptors, and w i l l be consideredm ore in detai l below .
The proprioceptive sense organs are deeply embedded in thetissues and are typically excited by those activities of the bodyitself wh ich arise in response to external stimulation. The
proprioceptors then excite to reaction the‘
sam e organs of response as the exteroceptors and regulate thei r action by reinforcement or by com pensation or by the maintenance of musculartone . All reactions concerned with m otor coordination, withmaintenance Of posture or attitude of the body, and withequi librium involve the proprioceptive system .
The im portant point to bear in m ind here is that stimulationof the vi sceral sense Organs typically cal ls forth visceral responses ,i . e.
, adjustm ents wholly w ith in the body, wh i le stimulation ofthe somatic (exteroceptive) sense organs typically calls forthsomatic responses, i . e.
, a readjustment of the body as a whole

78 INTR ODUCTION TO N EUROLOGY
w ith reference to its environm ent . This is a very fundam entald istinction . These two functions are quite diverse and the
organization of these two parts Of the nervous system shows corresponding structural d ifferences .The internal adjustm ents of the v isceral system s are effected
by a nicely balanced m echanism of local and general reflexes soarranged that m ost of their work is done quite m echanical ly andunconsciously . The taking of food and its prelim inary masticartion are generally voluntary acts whose various processes areormay be
—control led at wi ll . But once the food has passed intothe esophagus, the further work of swal lowing, digestion, and
assim i lation is no longer under d irect control . The presence ofa m orsel of food in the upper part of the esophagus excites themuscular m ovem ents necessary for the com pletion Of the act ofswallow ing, which no act of w i l l can prevent or m odify . In fact
,
any attem pt at conscious interference or regulation is apt toresult in an incoordination of the m ovem ents involved
,and
sputtering or gagging may result .The mechanism s involved in these processes are inborn and
require no practice for their perfect perform ance. They are
innate, invariable, and essentially sim i lar in all m embers of arace or species . They are, m oreover , nicely adapted to them ode of life characteristic of the species . In a carnivorous anim al the whole physiological mach inery of nutrition is differentfrom that of a herbivorous animal . These physiological andstructural peculiarities by wh ich each species of animal isadapted to its m ode of life have been brought about by naturalselection and other evolutionary factors . This is not absolutelytrue of all visceral actions ; some are acquired and m odifiable.
But as a general ru le th is is their type .
Some of the somatic actions are likewi se innate and relativelyfixed in character. Thi s is true of most of the proprioceptivereactions and of many of the exteroceptive as wel l . Fish can
swim as soon as they are hatched ch icks just out of the shell havean instinctive tendency to peek at all smal l objects on theground .
But in m ost of these cases (of wh ich innum erable instancesm ight be cited) som e practice is necessary before perfect responses are attained ; and a vei'y large proportion Of the exteroceptive acts are not innate , but acquired by long and often ardu

THE RE CE PTORS AND E FFE CTORS 79
ous experi ence . In higher vertebrates, as a rule,all but the
sim plest and m ost elem entary exteroceptive activi ties are ind ividual ly acqui red , variab le, non- hereditary
,plastic behavior
types . The elem ents of whi ch these acts are made up are,of
course, necessari ly present in the inherited reflex pattern ; butthe pattern according to which these elem ents are com b ined isnot wholly predete rm ined in the hereditary organization of thespecies (pp . 3 1
,
With these principles in m ind,let us now undertake an anal
ysis of the human receptors and of the nervous end -organs related to their effectors
,or organs of response. The fol low ing list
is by no means com plete and is in som e parts m erely provisional .
I. SOMATIC RECEPTORS
These are concerned wi th the adjustm ent of the body to external orenvironm ental relations .
A . THE EXTE ROCE PTIVE G ROUPThe sense organs of thi s group are stimulated by objects outside the
body and typically call forth reactions of the whole body, such as locom o
tion,or of its parts
,so as to change the relation of the body to i ts environ
m ent. Th is group includes a system of general cutaneous sense organs ,som e organs of deep sensibility
, and som e of the higher sense organs . The
cutaneous exteroceptors com prise a very com p lex system whose analysishas proved very d ifli cul t. The conclusions presented in the paragraphswhich follow are based chiefly upon the observations of von Frey , HenryHead , and Trotter and Davies . The correlation of the data of physiologicalexperim ent with the anatom ical structure of the cutaneous end -organs isstill som ewhat problematical and the assignm ent of end—organs here to thevar ious cutaneous senses should be regarded as provi sional rather than as
demonstrated .
1 . O rgans of Touch and Pressure.—These fall into two groups , those
for deep sensibility (pressure) and those for cutaneous sensibili ty (touch ) .The d ee pressure sense is served by nerve—endings throughout the tissues
of the bodyand is preserved intact after the loss of all cutaneous nerves .Most of the functions of the deep sensory nerves belong to the proprioceptive and interoceptive series (see below ) , but som e exteroceptive f unctionsare here present also . The latter are probably related chiefly to the
Pacin ian corpuscles and sim ilar encapsulated end -organs . The Paciniancorpuscle has a central nerve—fiber enclosed in a.firm lamellated connectivetissue sheath (Fig . By these end -organs relatively coarse pressurem ay be discrim inated and locali zed (exteroceptive function ) , and m ovementsof muscles and joints can be recogni zed (proprioceptive function) . The
sensory fibers concerned with the deep pressurexsense are distributed throughthe m uscular branches of the spinal nerves in company with the m otorfibers . The po int stimulated can be localized wi th a fair degreeof accuracy,
80 INTRODUCTION To N EUROLOGY
but there is no discrim ination of two compass points app lied sim ultaneouslyto the overlying skin. The two points will appear as one stim ulus
, evenwhen widely separated .
The cutaneous organs of tacti le sensi bi li ty are of several kinds,whose
precise functions are still obscure. There are two principal groups of these,those arranged in the hair bulbs at the roots of the hairs and those on the
hairless parts, such as the lips, the alm s of the hands, and the soles of thefeet. The latter are m ore highly ifferentiated endings and are organs ofthe m ost refined active touch .
Most of the surface of the body is m ore or less hairy,though many of
these hairs may be sofine as to escape observation . The hairs are the most
Fig. 22 .—Pacinian corpuscles from the peritoneum of a cat . (Af ter Sala,
from Bohm -Davidoff-Huber’s Histology .)
important sources of excitation of the first group of cutaneous sense organs,and the sens itiveness of the hair- clad parts is greatly reduced after the hairis shaved . The threshold of excitation to touch of the skin about the baseof a hair is from three to twelve times higher than that of a sim ilar excitation applied to the hair itself. The innervation of the hair bulbs is verycom p lex and varies greatly for d ifferent animals and for the d ifferent kindsof hairs on the sam e body
,so that no general description is possible.
Miss Vincent has shown that the large vibri ssae of the rat receive theirnerve-supply from two sources . A large nerve bundle pierces the deep

82 INTRODUCTION To N EUROLOGY
Under the hairless parts of the skin there are special tactile bodies, suchas Meissner ’s corpuscles . These are generally found in the deep layer ofthe skin (derm is ) and in the underlying tissues, either as free skein—liketerminal arborizations of cutaneous nerves or as sim ilar more elaborateendings enclosed in connective- tissue capsules . Figures 24 and 25 illustratethe m ost highly d ifferentiated form of these endings
, the Meissner corpuscles . Merkel ’s corpuscles (Fig. 26 ) are probably simpler organs of thi ssystem .
Stratum lucidumSt ratum
corium
Blood ~vesselsand nerves
Fig . 24.
—Section through the hum an skin ,illustrating the four layers
of the ep iderm is and the papillae of the derm is or corium . A corpuscle of
Meissner Is seen within one of the dermal papillae . (From Cunningham ’sAnatom y .)
All form s of cutaneous sensibility (touch , tem perature, and pain) whenstudied phys iologically are found to be localized in small areas or sensoryspots
, each of whi ch has a specific sensibility to one only of the cutaneoussensory qualities . The intervening parts of the skin are insensitive . An
imm ense am ount of physiological and clinical observation has been devotedto the analysis of cutaneous sensibility , including the experim ental di vi sion
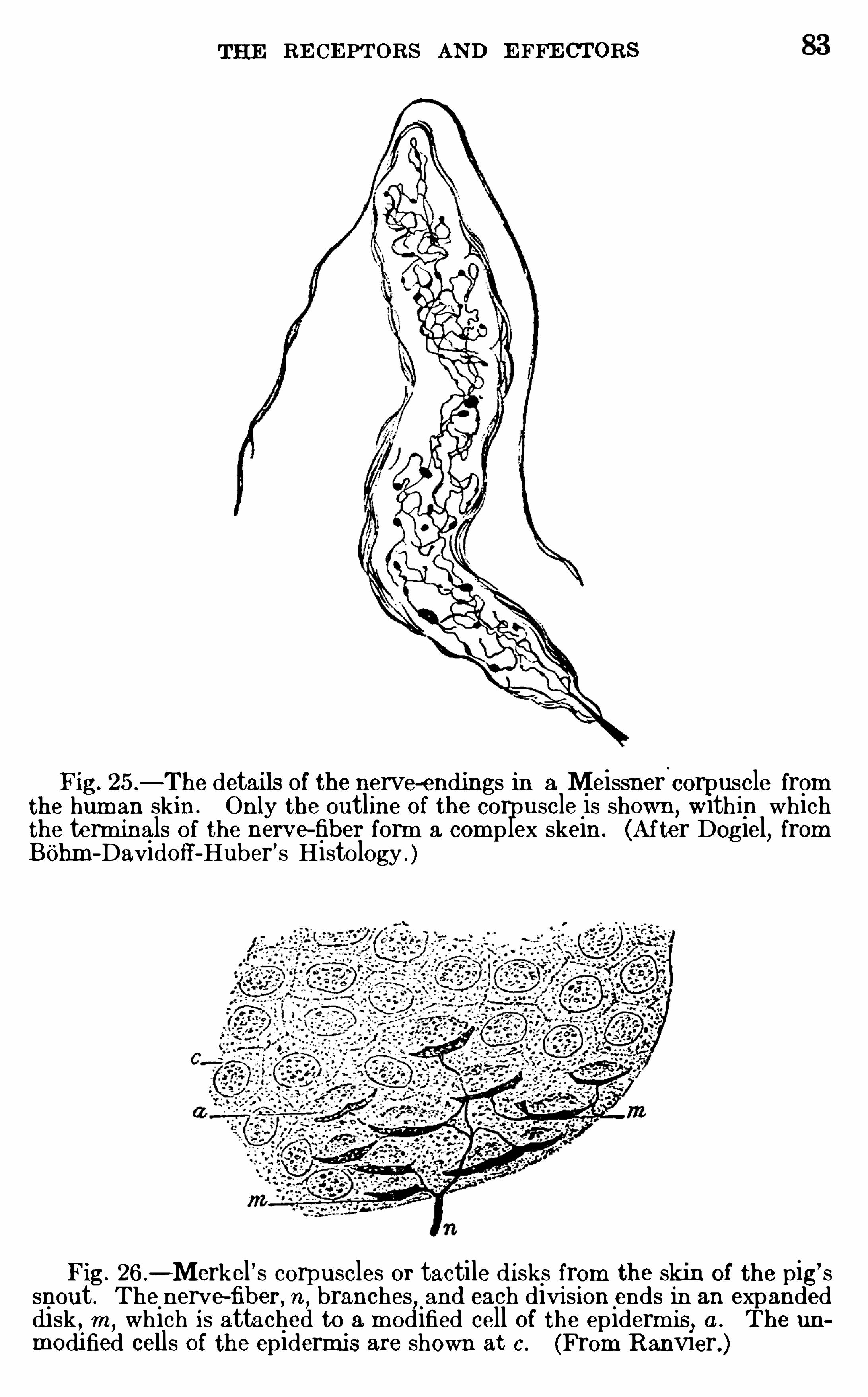
RE CE PTORS AND E FFE CTORS
Fig. 25 .—The details of the nerve-endings in a Meissner corpuscle from
the hum an skin. Only the outline of the corpuscle is shown, w ithin whichthe term inals of the nerve fiber form a com p lex skein . (After Dogiel, fromBOhm -Davidoff-Huber ’s Histology . )
Fig. 26 .-Merkel ’s corpuscles or tactile di sks from the skin of the pig
’ssnout . The nerve—fiber, n, branches, and each divi sion ends in an expandedd isk , m , which is attached to a m odified cell of the epiderm is , a . The numodified cells O i the epidermi s are shown at c . (From R anvier.)

84 INTRODUCTION TO N EUROLOGY
of cutaneous nerves in their own bodies by Head , Trotter, and Davies forthe purpose of studying m ore critically the d istribution of the varioussensory functions in and around the anesthetic areas produced by theinjuries and the phenom ena accom panying the restoration of these functionsduring the regeneration of the nerves . But general agreem ent has not yetbeen reached on all questions .Head and hi s colleagues are of the Op inion that all form s of cutaneous
sensibility (touch , tem perature, and pain) are grouped in two series eachserved by different nerve-fibers and end -organs ; these he term s “protopathic and
“ep icritic” sensibility . Protopathi c sensi bi li ty is subjectively
general di ffuse sensibility of a prim itive form . Its sense organs are arrangedin definite spots, and yet these sensations have no clear local reference or
Sign ; that is, the spot stim ulated cannot be accurately localized . There areseparate spots for touch , heat, cold, and pain ; these spots being generallygrouped near the hair bulbs . In fact, the hairs are the m ost im portanttactile organs of this system and the other sense qualities belonging here areintim ately associated with the roots of the hairs. Epicri tic sensi bi lity is
Fig. 27.—End -bulb of Krause from the conjunctiva of man .
.
Thenerve-end mg form s a globular skein within a deli cate connectrve-t1ssue
capsule. (A fter Dogiel . )a more refined sort of discrim ination, and is regarded as a later evolutionarytype . It includes light touch
,on the hairless parts of the body particularly,
an d the discrim ination of the interm ediate degrees of tem perature. Cutarneous localization and the discrim ination of the distance between two pointssim ultaneously stim ulated (the “
com pas s test ” ) are functions of this system ; but pain sensibility is not included , this being wholly protopathic .
Trotter and Davies repeated som e of Head ’ s experim ents and , whileconfirm ing m ost of his observations , they were led to som ewhat differentconclusions . They d o not regard the protopathic and epicritic series asserved by distinct system s of nerves , but as different physiological phases ofthe sam e system s of nerve-fibers and end -organs .2 . End -organs for SensibiIity to Col d .
3 . End -organs for Sens ib ility to Heat.—Physiological experim entshows that warm th and cold are sensed by different parts of the skin (thewarm Spots and the cold spots respectively) , and Head is of the Opinion thateach of these types of sensibility may be present in an epicritic and a protopathic form . What end -organs are involved here is by no m eans certain .
The m argin of the corneawas found by von Frey to be sensitive to pain and

THE RE CE PTORS AND E FFE CTORS 85
cold only . The free nerve-endings found here he assum es to be pain receptors and the end -bulbs of Krause (Fig . 27) to be cold receptors . y an
analogous argum ent he assum es that the “genital corpuscles Of Dogiel andsom e sim ilar endings widely distributed in the skin are warm th receptors .By som e other physiologists these types of corpuscles are regarded as belonging to the tactile system . Stimulation of the somatic nerves Of deep sensibili ty causes no tem perature sensations . (For tem perature sensations in theviscera see p .
4 . End -organs for Pain —Som e physiolo
gists believe that there are separate nerveendings for pain ; others regard pain as a
quality whi ch m ay be present in any sense,and not as itself a true sensation (pp . 249
The free nerve-endings am ong the cellsof the epiderm is are regarded by von Freyas the pain receptors
,because these endings
alone are present in som e parts of the bodywhere susceptibility to pain is the only sensequality present, such as the dentin and pulpof the teeth (Fig. the cornea
,and the
tym panic m em brane of the ear (J . G .Wilson ) .Sim i lar endings are found throughout the
epiderm is (Fig. 29 ) and in many deep struetures . The nerves of deep sensibility of the
som atic sensory type m ay also carry painfulimpressions . (For visceral pain see pp . 243,
A ccording to Head, cutaneous pain iswholly of protopathic type, and in case of in
jury to the peripheral nerves it disappearsand reappears in regeneration simultaneouslywi th the protopathic type of tactile and temperature sensation . Thi s cutaneous pain isnot accurately localizable unless epicritic cu
taneous sensibility is also present.5 . End -organs of G eneral Chem i cal Sen
sibili ty.—In m an this type of sensibility is
found only on m oist ep ithelial surfaces, suchas the m outh cavity ; but in fishes it m ay bepresent over the entire surface of the body .
The sense organ is probably the free nerveterm inals am ong the cells of the epithelium ,
never special sense organs li ke taste-buds,for these when present in the skin belongto a quite different system . Coghill has
Fig. 28 .
—Longitudinalsection Of a tooth of afish ,G obius, showing nerve term inals : d , Dentin ; n
,
nerve-fibers entering the
cavity of the dentin and
ending free . (A fter R etz ius
,from Barker ’s N er
vous System . )
recently shown that the supposed sensitivity of the amphibian skin to acidsis really d ue to a destructive action of the reagents upon the epithelium , and
the entire question of d iffuse chem ical sensibility requires further study .
6 . O rgans of Hearing—The stimulus is m aterial vibrations whosefrequency ranges from 30 to per second (see p . The receptor isthe spiral organ (organ of Corti) in the cochlea of the ear (see p . and
perhaps also the sensory spots in the saccule and utricle . There are two
form s of auditory sensations : ( 1 ) noise, stimulated by sound concussions orIrregular m ixtures of aerial vibrations ; (2) tone, stimulated by sound wavesor perIod ic aerial vibrations .
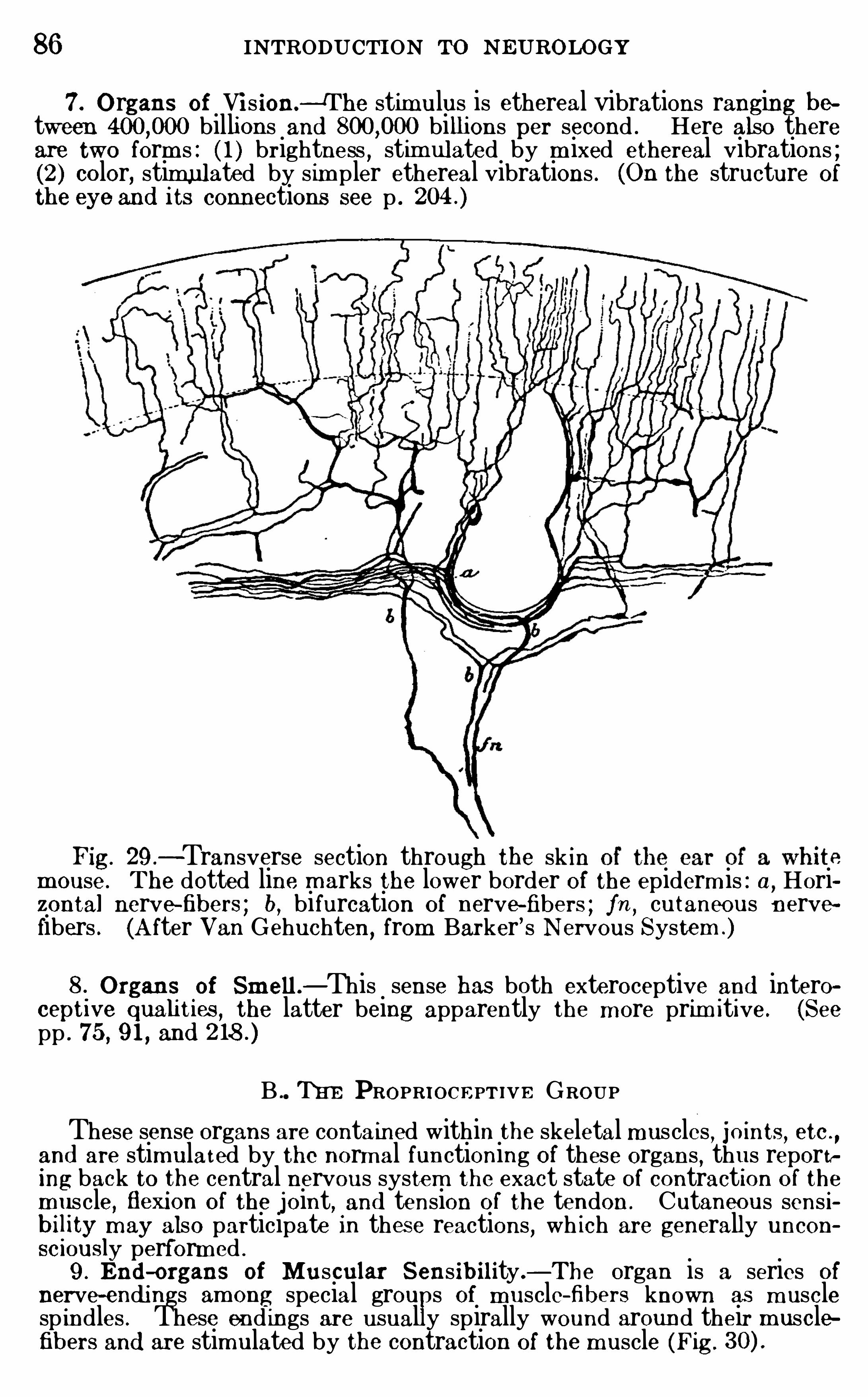
86 INTRODUCTION TO NEUROLO GY
7 . O rgans of Vi s ion—The stimulus is ethereal vibrations ranging between billions and billi ons per second . Here also thereare two form s : ( 1 ) brightness , stim ulated by m ixed ethereal vibrations ;(2 ) color, stimulated by sim pler ethereal vibrations . (On the structure Of
the eye and i ts connections see p .
Fig. 29 .—Transverse section through the sk in
m ouse . The dotted line m arks the lower border Of the epiderm is : a, Horizontal nerve-fibers ; b, bifurcation of nerve-fibers ; fn ,
cutaneous nervefibers . (A fter Van Gehuchten, from Barker
’ s Nervous System .)
8.O rgans of Sm ell .—This sense has both exteroceptive and intero
ceptive quahties , the latter being apparently the m ore prim itive. (Seepp . 75 , 9 1 , and
B TE E PROPRIOCE PTIVE GROUPThese sense organs are contained within the skeletal muscles
, Jmnts , etc .
,
and are stimulated by the norm al functioning of these organs , thus reporting back to the central nervous system the exact state Of contraction of the
m uscle, flexion of the joint, and tension of the tendon . Cutaneous sensibility m ay also participate in these reactions , wh ich are generally uncon
sc iously perform ed .
9 . End -organs of Muscular Sensib ility .—The organ is a series Of
nerve-endi ngs am ong special grou s of muscle-fibers known as m usclesp indles . These end ings are usually spirally wound around their musclefibers and are stim ulated by the contraction of the m uscle (Fig .
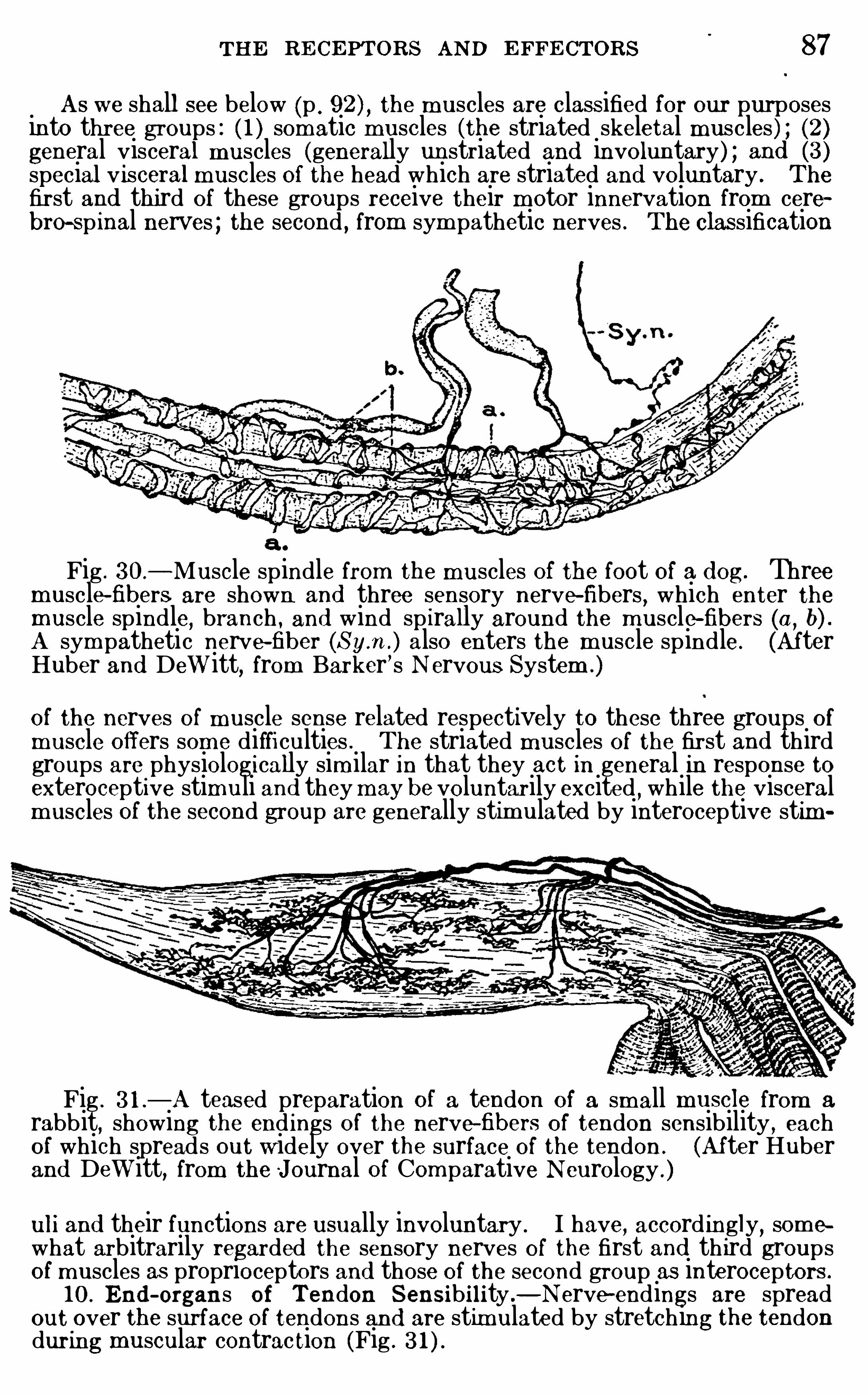
THE RE CE PTORS AND E FFE CTORS 87
As we shall see below (p . the m uscles are classified for our purposesinto three groups : ( 1 ) somatic muscles (the striated skeletal muscles ) ; (2)general visceral muscles (generally unstriated and involuntary) ; and (3 )special visceral muscles Of the head which are striated and voluntary . The
first and third Of these groups receive their m otor innervation from cerebro-sp inal nerves ; the second , from sym pathetic nerves . The classification
Fig . 30 .
—Muscle spindle from the muscles Of the foot of a d og. Threemuscle-fibers are shown and three sensory nerve- fibers
,which enter the
muscle spindle,branch , and wind sp irally around the m uscle-fibers (a ,
b) .A sym pathetic nerve-fiber also enters the muscle sp indle. (AfterHuber and Dewitt
,from Barker ’s N ervous System .)
of the nerves of m uscle sense related respectively to these three groups ofmuscle offers som e difficulties . The striated muscles of the first and thirdgroups are physiologically sim ilar in that they act in general in response toexteroceptive stimuli and theym ay be voluntar ily excited , while the visceralm uscles of the second group are generally stim ulated by interoceptive stim
Fig . 3 1 .
—A teased preparation O f a tendon of a sm all muscle from a
rabbit, showing the end ings of the nerve-fibers O f tendon sensibility , eachof which spreads out widely over the surface of the tendon . (Af ter Huberand Dewitt, from the J ournal Of Com parative N eurology . )
uli and their functions are usually involuntary . I have, accord ingly, som e
what arbitrarily regarded the sensory nerves of the first and th ird groupsof muscles as proprioceptors and those Of the second group as interoceptors .
1 0 . End - organs of Tendon Sensibi lity .—N erve—endings are spread
out over the surface of tendons an d are stimulated by stretching the tendonduring muscular contrac tion (Fig .
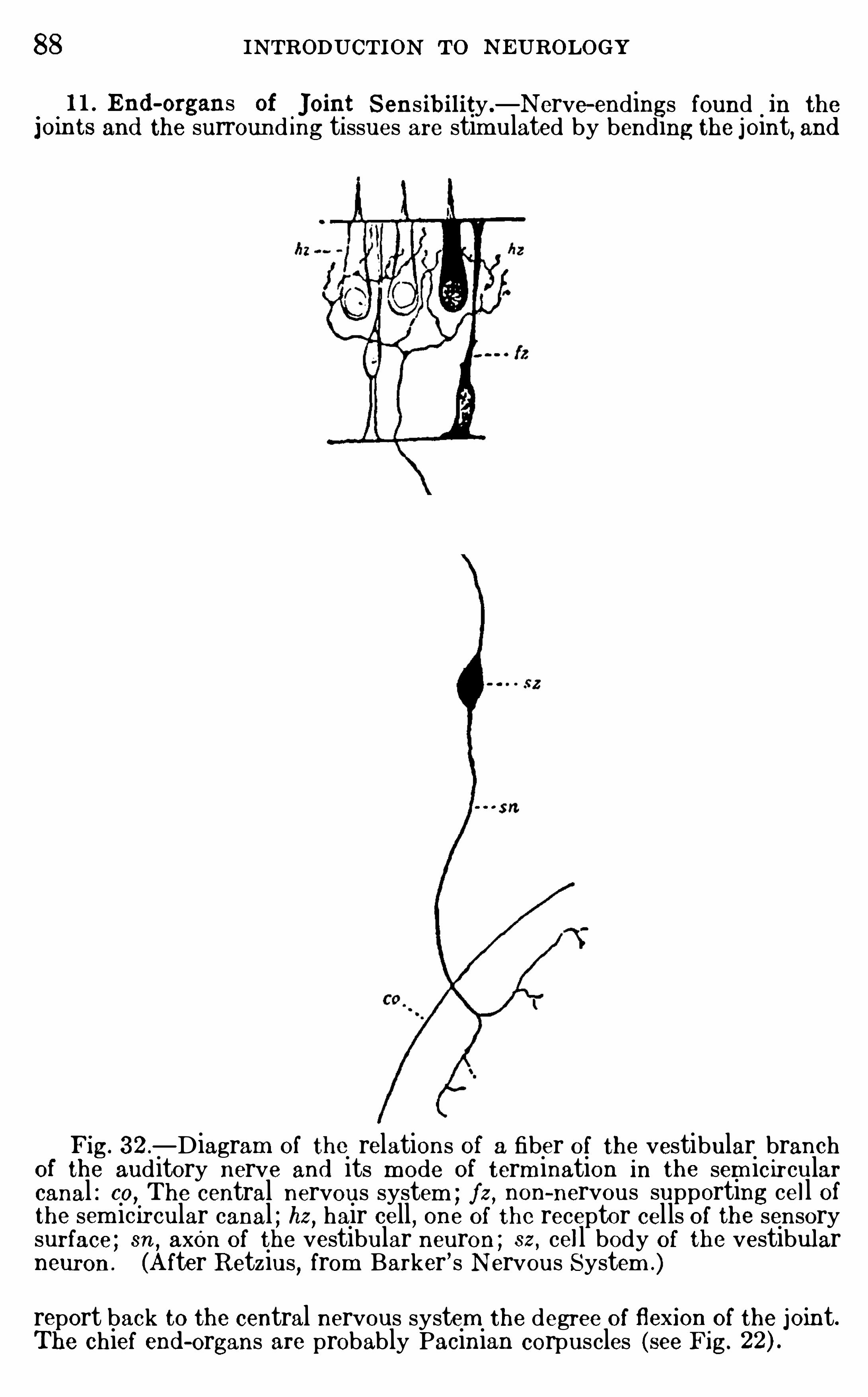
88 INTRODUCTION TO N EUROLOGY
0
1 1 . End -organs of.Joint Sens ibi lity.—Nerve—endings found in the
j om ts and the surround ing ti ssues are stim ulated by bending the joint, and
Fig. 32 .—D iagram Of relations of a fiber Of the vestibular branch
of the auditory nerve and its m ode of term ination in the sem icircularcanal : co, The central nervous system ; fz , non-nervous supporting cell ofthe sem icircular canal ; hz, hair cell , one Of the receptor cells of the sensorysurface ; sn , axon of the vestibular neuron ; sz , cell body of the vestibularneuron . (A fter R etzius, from Barker
’s N ervous System .)
report back to the central nervous system the degree of flexionof the joint.The chief end -organs are probably Pacinian corpuscles (see Fig.
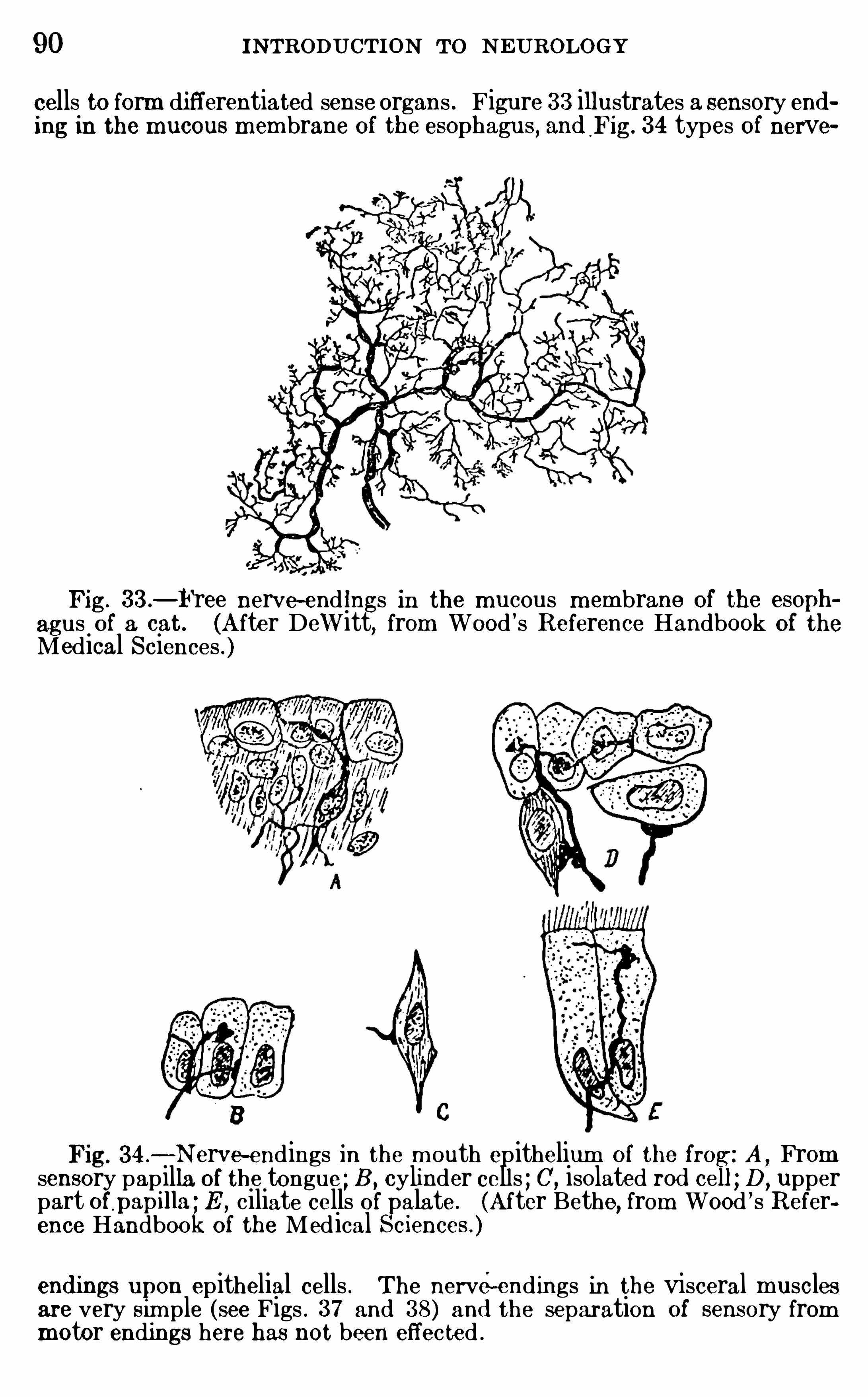
90 INTRODUCTION TO N EUROLOGY
cells to form di fferentiated sense organs . Figure 33 illustrates a sensory ending in the mucous mem brane Of the eSOphagus, and Fig. 34 types of nerve
Fig. 33.—Free nerve-end ings in the mucous m embrane of the esoph
agus of a cat. (After Dewitt, from Wood ’s Reference Handbook Of theMedical Sciences . )
34 .—Nerve-endings in the mouth epithelium of the fro A
,From
sensory papilla of the tongue ; B , cylinder cells ; C’, isolated rod cei D, upper
part Of,papilla
’ E ,ciliate cells of alate. (After Bethe, from Wood ’s R efer
ence Handbookof the Medical eiences . )
end ings upon epithelial cells . The nerve-endings in the visceral musclesare very Sim ple (see Figs . 37 and 38) and the separation of sensory frommotor endings here has not been effected .

THE RE CE PTORS AND E FFE CTORS 91
B . SPE CIA L VISCE RA L GROUP .22 . O rgans of Taste.
—These are excited by chem ical stimulation oftaste-buds on the tongue and pharynx by sweet
,sour
,salty
,or bitter sub
stances . In m an this is a strictly interoceptive sense ; but in som e fishestaste—buds are scattered over the outer body surface in addition to them outhcavity
,and thus m ay serve as exteroceptors also . The organ is a flask
shaped collection of specialized epithelial cells of two sorts, supporting andspecific sensory elem ents (Fig. There is a double innervation
,partly
by perigemm al fibers whose end ings surround the bud , and partly by intragemm al fibers which penetrate the bud and arborize in intimate relationwith the specific sensory cell s .23 . O rgans of Sm ell .—These are excited by chem ical stimulation of the
specific Olfactory mucous membrane of the nose. The number of substances
Fig. 35 .
—Taste—bud from the side wall of a circum vallate papilla of thetongue : ( 1 , Taste-pore ; b, nerve-fibers , som e Of which enter the taste—bud(intragemm al fibers ) , while others end freely in the surrounding epithelium(perigemm al fibers) . (After Merkel—Henle. )
wh ich m ay act as stim uli is greater than in the case Of taste-buds , the number Of subjective qualities is also greater, and the d iscrim ination thresholdis m uch lower (see pp . 75 and The peripheral organ of sm ell is aspecific sensory epithelium within the nose, whose sensory cells give risedirectly to the fibers Of the olfactory nerve
,this being the only peripheral
nerve Of the human body whose fibers arise from superficially placed cellbodies (Fig .
That the Olfactory system was originally an interoceptive sense seem sclear ; but in all vertebrates living at the present tim e the visceral responsesto sm ell are less im portant than the somatic reactions . The sense of sm ell
is the leading exteroceptor in m ost lower vertebrates, and this function hasbeen secondarily derived from the prim ary visceral function . We have seenabove that the sense of taste in som e fishes has secondarily acquired exteroceptive functions ; and in the case Of sm ell th is secondary change has beencarried stil l further until the exteroceptive function has com e to dom inate

92 INTRODUCTION TO NEUROLOGY
Fig. 36 .—Cells from the O lfactory mucous m em brane : A from the frog,
B and C from man . The supporting cells are non-nervous . The O lfactoryhairs Of the olfactory cells project out
'
into the m ucus O f the nose, and are
probably the specific receptors . The central process at the base of eacholfactor cell is prolonged into a fiber of the o lfactory nerve (not shownin the gure) , which extends inward to the brain (cf. Fig. 104 , p .
(After Schulte and Brunn .)
the prim itive interoceptive, though the latter has by no means been en
tirely obliterated .
III. SOMATIC EFFE CTORS
24 . End -organs on Striated Skeletal Muscles .—This m otor end
plate is a com lex term inal arborization Of the m otor nerve-fiber, associ
ated with an e evated granular m as s Of protoplasm and a collection O f
nuclei Of the muscle—fiber (see Fig. 5 , tel, p .
The somatic m uscles whose innervation is here under consideration are
derived em bryologically from the som ites, or prim ary m esodermal segm entsof the em bryo
,while the visceral m uscles have a different origin. They are
under the d irect control of the will and are concerned ch iefly with locom otion or other m ovem ents wh ich change the relations O f the body to itsenvironm ent. They are typically stim ulated to action through the ex
teroceptive sense organs . They m ake up the bulk of the musculature of
the trunk and lim bs and are represented in the head only in the externalm uscles of the eyeball and a part of the m uscles of the tongue.
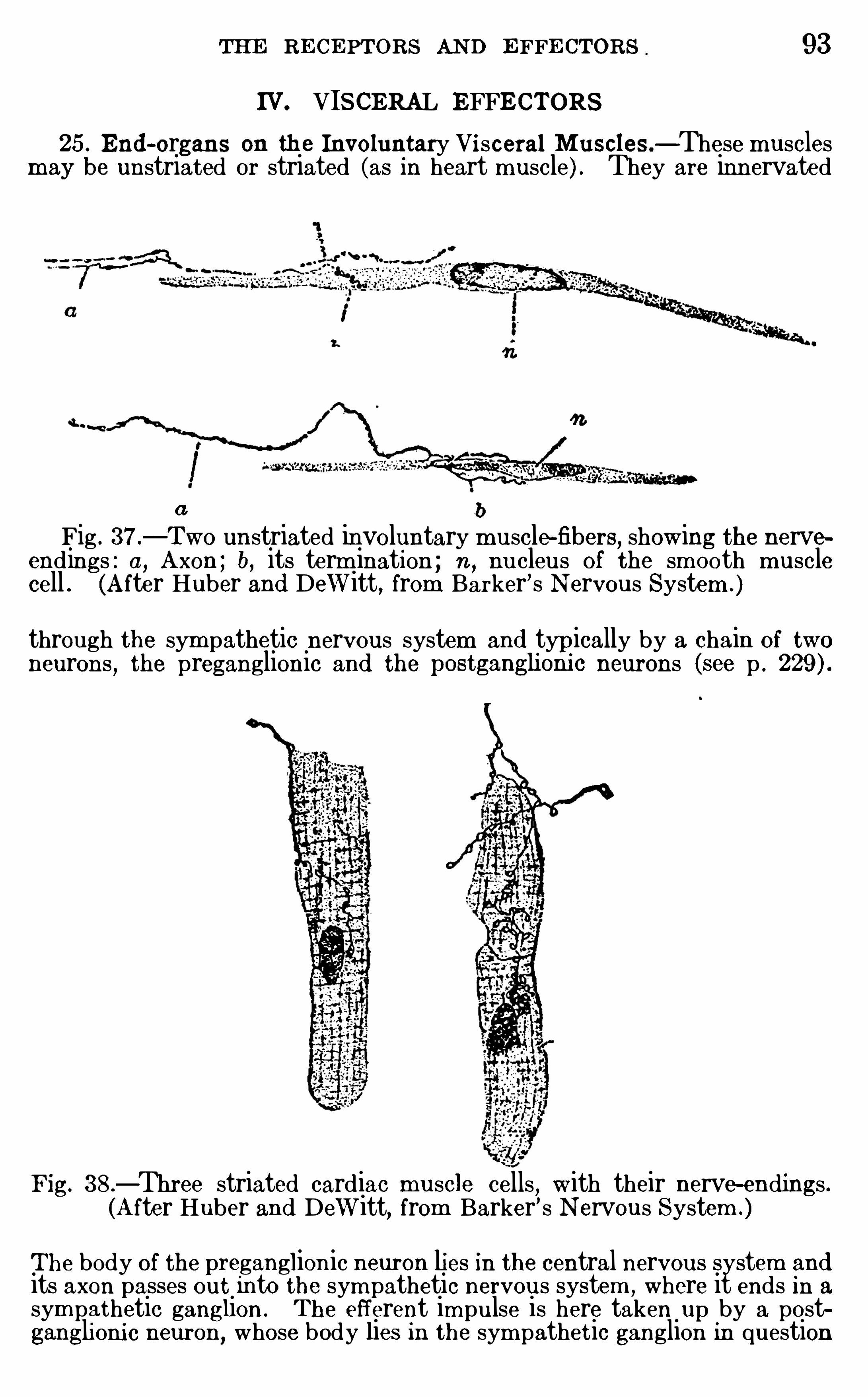
THE RE CE PTORS AND E FFE CTORS . 93
IV VISCERAL EFFECTORS25 . End -organs on th e Involuntary Vi sceral Muscles—These muscles
may be unstriated or striated (as in heart muscle) . They are innervated
a
Fig . 37 .—Two unstriated involuntary m uscle-fibers, showing the nerve
endings : a, Axon ; b, its term ination ; n ,nucleus of the smooth muscle
cell . (A fter Huber and Dewitt, from Barker’s Nervous System .)
through the sym pathetic nervous system and typically by a chain of twoneurons , the preganglionic and the postganghom c neurons (see p .
Fig. 38.—Three striated cardiac muscle cells
, with their nerve-endings .(After Huber and Dewitt, from Barker’ s N ervous System .)
The body of the preganglionic neuron lies in the central nervous system andits axon passes out into the sym pathetic nervous system , where it ends in a
sym pathetic ganglion . The efferent im pulse is here taken up by a postganglioni c neuron, whose body lies in the sym pathetic ganglion in question

94 INTRODUCTION To N EUROLOGY
and whose axon passes onward through a sym pathetic nerve to end In the
appropriate effector . The nerve-endings of this system are sim ple or
branched free term inals ehd ing on the surface of the muscle—fiber (Fig.
in the case of heart muscle the fibers usually have expanded tips (Fig.
26 . End -organs on G land s .—The innervation of these organs is in m ostrespects sim ilar to that of the involuntary m uscles last described . A finep lexus of unm yelinated fibers Of sym pathetic origin envelops the sm allerglands and pervades the larger ones ; these are beli eved 1n som e cases to bethe excite-glandular fibers .27 . Special Vi sceral Motor End -organs .
—The nerves of these m uscleshave no connection with the sym pathetic nervous system . These effectorsare striated m uscles which may act under the direct control of the w ill .In their evolutionary origin they are derived from the m uscles of the gillsOf the lower vertebrates , and they are developed em bryologically from theventral unsegm ented m esoderm and not from the prim itive m esoder malsegm ents which give rise to the som atic m uscles . They are found only 1n
the head and neck and their nerve-endings are Sim ilar to those of the
striated muscles of the som atic series .
Summary.
-We have seen that the ch ief function of the senseorgans is to lower the threshold of excitabi lity Of the body indefinite places to particular kinds Of stimulation, and thus toeffect an analysis Of the forces of nature so far as these concernthe welfare of the body . The nature of th is analysis of the environing energy com plex was i l lustrated by a review Of the waysin wh ich the body may respond to different kinds of vibrations .The senses
,as th is word is comm only used
,were d istinguished
by four cri teria,term ed briefly the psychological , physical , ana
tom ical,and physiological . Then followed a physiological c lassi
fication Of the receptors and effectors of the human body .
LITERATUREBARKE R
,L . F . 190 1 . TheN ervous System an d Its ConstituentNeurones ,
New York .CO GHIL L , G . E . 19 14 . Correlated Anatom ical and Physiological Studiesof the Growth of the N ervous System of Am phibia. I . The Afferent System Of theTrunk ofAm blystom a
,Jour . Com p . N eur .
,vol . xxiv, pp . 16 1—233 .
VON FRE Y , M . 1897 . Untersuchungen fi ber d ie Sinnesfunctionen d er
m enschlichen Haut, Abhangl . kgl . Sachs . G esellseh .,Bd . 40 (Math -Phys .Classe, Bd .
HE AD, H., R IVERS, W . H . R .
,and SHERREN , J . 1 905 . The Afferent
N ervous System from a N ew A spect, Brain ,vol . xxviii , pp . 99—1 15 .
HE RRICK , C . JUDSON . 1903 . On the Morphologi cal and PhysiologicalClassification Of the Cutaneous Sense Organs of Fishes, Am er . N aturalist,
vol . xxxv ii , pp . 3 13 3 18 .
1908 . On the Phylogenet ic Differentiation O f the Organs of Sm elland Taste, Jour . Com p . N eur .
, vol. xv iii, pp . 157- 166 .
—. 19 14 . End -organs, N ervous, Wood ’s R eference Handbook of the
Med 1cal ScIences, 3d ed .,vol. iv, pp . 20—27 , New York.

THE RE CE PTORS AND E FFE CTORS 95
HERTz, A . F . 19 1 1 . The Sensibility Of the A lim entary Canal, London .
HUBE R , G . C . 1900 . Observations on Sensory N erve—fibers in VisceralNerves and on their Modes of Term inating
,Jour . Com p . N eur .
, vol . x,pp . 134- 15 1 .
HUB E R,G . C .
,and DEWIT
’
I‘
,LYD IA
,M . A . 1897 . A Contribution on
the Motor N erve—endings in the Muscle-spindles , Jour . Com p . N eur .,vol.
vii,pp . 169—230 .
1900 . A Contribution on the Nerve Terminations in Neuro—tendinous End -organs , Jour . Com p . N eur .
,vol . x
,pp . 159—208 .
PA RKE R, G . H . 19 12 . The R elation of Sm ell,Taste
, and the Comm onChem ical Sense in Vertebrates, Jour . A cad . Nat . Sci .,Phila.,
2 Ser., vol.
xv,pp . 22 1—234 .
PA RKE R,G . H.
, and STAB LE R , E LE AN OR M . 19 13 . On Certain Distinctions Between Taste and Sm ell , Am er . Jour. Physiol . , vol . xxxi i, pp . 230
240.
R IVE RS, W . H . R . and HEAD ,H . 1908 A Hum an E xperim ent In N erve
Division,Brain
,vol . xxxi
, p. 323 .
SHE LDON , R . E . 1909 . The Reactions of the Dogfish to Chem ical Stimuli , Jour . Comp . N eur .
,vol . xix
,pp . 273—3 1 1 .
SHERRIN GTON , C . S. 1906 . The Integrative Action of the N ervous System , N ew York .
TROTTE R, W.,and DAVIE S
,H . M . 1909 . E xperim ental Studies in the
Innervation of the Skin,Jour. Of Physiol .
,vol . xxxvii i, pp . 134- 246 .
VINCE NT, STE LLA B . 19 13 The Tactile Hair of the White Rat, Jour .Com p . N eur. , vol. xxiii, pp 1
19 13a. The Functionof the Vibrissze in the Behavior of the WhiteR at, Behavior Monographs , vol . i , No . 5 , pp . 7- 8 1 .
WATSO N , J . B . 19 15 . Behavior,An Introduction to Com parative Psy
chology, Chapters XI—XIV, N ew York .
WILSO N, J . G . 19 1 1 . The N erves and N erve—endings 1n the Mem brana
Tympani of Man,Am er. Jour. Anat.
, vol. xi , pp . 10 1 1 12 .

CHAPTER VI
THE GENERAL PHYSIO LO GY OF THE NERVOUS
SYSTEM
THE functions of the body are generally effected by chem icalchanges with in its protoplasm . These chem ical changes in theaggregate we term
“metabol ism and they general ly invo lve a
rather slow interchange of the chem ical substances of food andwaste materials between the cytoplasm and the lym ph wh ichsurrounds the cel ls and between the cytoplasm and the protoplasm of the nucleus (karyoplasm ) . The rate of m etabo lism isdependent upon m any factors, one Of wh ich is the tim e requiredfor the passage Of soluble substances through the cell m em braneand through the nuclear membrane which separates the cytoplasm from the karyoplasm .
In the nerve- cells both Of these sorts of chem ical interchangeare faci litated by the form and internal structure of the cel l .As we have already seen (p . the w idely branch ing dendritespresent a large surface for the absorption Of food materials fromthe surrounding lym ph and the elim ination Of waste. The
specific nervous functions involve the consum ption of l iving substance
,both in the cel l body and in the nerve—fibers . Th is is
in part an oxidation process, and th is phase of the activity can
be rough ly m easured by the am ount of carbon dioxid elim inated .
Unti l very recently it was not possible to secure any evidence ofC0 2 production in nerve-fibers ; in view of this and of the furtherfact that nerve-fibers seem to be less susceptible to fatigue thannerve- cells and synapses
,m any physiologists assum ed that
nervous conduction is not a chem ical process,but perhaps som e
sort Of m olecular vibration . The conduction of a nervous impulse through a living nerve-fiber is accom panied by an electricchange, the so- cal led negative variation,
wh ich by som e physiologists has been identified with the nervous im pulse itself .Th is and other com plicated theories of nervous transm ission
96

98 INTRODUCTION To N EUROLOGY
nerves it may be as rapid as 125 m eters per second . This rateof conduction of the nervous im pulse in peripheral nerves varies
greatly w ith different animals,w ith d ifferent nerves in the same
anim al,and in the same nerve under different physiological con
d itions .
The reacl ion time required for the perform ance O f various reflex actscan be very accurately m easured
, and it is found that the time Of even thesim plest reflex is considerably greater than is required for the transm issionO f the nervous irnpu lse through the conductors involved . The average rateof conduction m human nerves is probably about 120 m eters per second , andthe sim p lest reaction tim es which have beenm easured In psychologi cal laboratories vary between and second (from to for reactionsto touch , and from to for reactions to sound ) . The total time
required for transm ission of the nervous im pulse through the nerve-fibersinvolved in these reactions need not exceed second , whence it appearsthat the greater part of the reaction tim e is otherwise consum ed . A part Ofthis excess tim e is required to overcom e the inertia of the end -organs(receptor and effector) , and the remainder is used in the central nervoussystem . Th is “cen tral pause is characteristic O f all reflexes and ,
in fact,has a profound sign ificance in connection with the evolu t ion of the higherassociational functions of the brain . The introduction O f further com plexity'in the reaction
,of whatever sort, usually lengthens the tim e of the central
pause,though long traini ng in making a d iscr im inative reaction m ay re duce
th is pause alm ost to the tim e O f a sim ple reaction .
Many attempts have been m ade to determ ine the central tim e of reac
tions of differen t degrees of com plexity by substracting from the total time
in each case the probable tim e, requ ired for the peripheral processes and bysubtracting the total tim e required for the sim pler reactions from the totaltim e taken in m ore com plex discrim inative reactions . But further analysis(particularly m ore critical in trospection) has shown that in these hum an
reactions the problem is too com plex to be resolved by th is m ethod (seeLadd and Woodworth , 19 1 1 , p .
The sim pler reflexes O f lower vertebrates can be stud ied physiologically,and these give data which are m uch m ore readily analyzed than the m orecom plex hum an reactions . In the case O f the sim plest reflex obtainable inthe spinal cord of the frog
, the central pause was estim ated by Wundt to beonly second , i . c.
, all of the tim e required for the reaction except thisin terval was used in the peripheral appara tus . But in a crossed reflex
,
where the reac t ion occurs on the oppos i te. side of the body from the stim ulus
,the increased com plexi ty of the central process consum ed second
addi t ional .Miss Buchanan w ith m ore accura te.m e thods of study
,finds in the
frog that the central tim e varies be tween .014 and .02 1 second . She alsom easured the addi tional latent time required for a crossed reflex
,and found
i t to be of the sam e order o f m agnitude as the latent tim e O f the sim plereflex (instead of half as m uch as in Wund t
’
s experim ents ) , that is , thecrossed reflex required about twice the latent tim e in the spinal cord as theuncrossed reflex . It is assum ed that this central pause in the uncrossedreflex 1s consum ed ch iefly in the synapses between the peripheral sensoryand the peripheral m otor neurons , and that only one such synapse is 1nvolved in each sim ple reflex connect ion (a two-neuron circuit, see Fig. 1 ,

THE GE N E RAL PHYSIOLOGY OF THE N E RVOUS SYSTE M 99
p . but in the crossed reflex two such synapses are involved (a threeneuron circuit such as the pathway from d .r.2 to v.r .1
' through correlationneuron 1 in Fig . 6 1
,p . and the introduction of the second synapse
doubles the t im e . It is , therefore, assum ed that it requires in the frogbetween .01 and .02 second for the nervous im pulse to pas s the synapsebetween two neurons in a reflex circuit.
Turn ing now to the activities of the nerve- cel l body,it wi ll
be recalled (p . 45) that here the chrom oph i lic substance is generally scattered throughout the cytoplasm in the form of the“N issl bodies .” This substance is very sim ilar to that of the
chromatin of the nucleus,from wh ich it is said to be derived
during the developm ent and functional activity Of the neuron .
During the resting state Of the cell it and other reserve m aterialsaccumulate in the cytoplasm ; and now
,when the cell is stimu
lated to activity,the energy thus stored up may be liberated
alm ost instantly because the chem ical substances necessary forthe reaction are w idely diffused throughout the entire mass ofthe cytoplasm .
The function of neurons, as com pared w ith that Of m ost othercells of the body , may,
therefore,be described as of the explo
sive type . A word of explanation w il l render the analogy c lear.In ordinary com bustion ,
oxygen is supplied to the surface Of the
burning material,say a b laz ing log, and the chem ical process
of burning goes on only as fast as the superfic ial parts can beoxid ized and rem oved . But explosive substances are chem ically so constituted that as soon as com bustion begins oxygen isl iberated in the interior Of the material and the process of oxidation takes place almost instantaneously throughout the
entire m ass . Sim i larly in the nerve- cell,the processes Of m etab
olism are not dependent upon the slow interchange Of substancesthrough the nuc lear m em brane between the cytoplasm and the
nuclear plasm ; but the chrom oph i lic substance d istributedthrough the cytoplasm perm its Of m uch m ore rapid responses .The organization Of the protoplasm of the nerve—cell is suchthat a very sm al l st im ulus m ay l iberate a large am ount of energyw ith explosive suddenness . The energy thus l iberated doesnot all leave the cell
,but part of it is directed into the axon,
wh ich is thereby excited to conduct a nervous impulse to theappropriate end -organ or to the next synapse, and thence to asecond neuron .
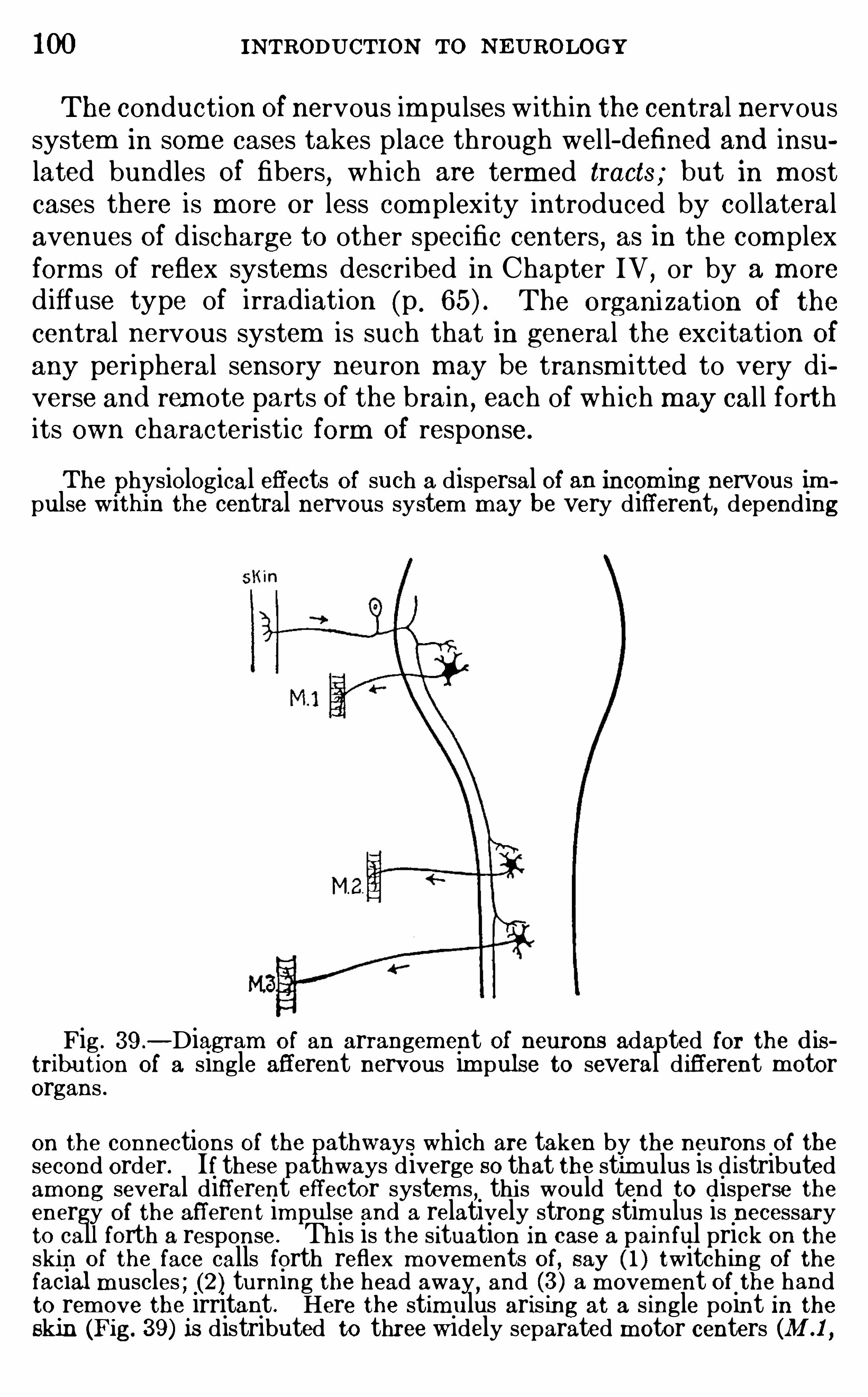
100 INTRODUCTION To NEUROLOGY
The conduction Of nervous im pulses w ith in the central nervoussystem in som e cases takes place through wel l- d efined and insulated bundles Of fibers, wh ich are term ed tracts ; but in m ostcases there is m ore or less com plexity introduced by collateralavenues of d ischarge to other specific centers
,as in the com plex
form s of reflex system s described in Chapter IV,or by a m ore
d iffuse type of irradiation (p . The organization Of the
central nervous system is such that in general the excitation Of
any peripheral sensory neuron m ay be transm itted to very d iverse and rem ote parts Of the brain
, each of wh ich may cal l forthits own characteristic form of response.
The physiological effects of such a dispersal Of an incom ing nervous impulse w 1th1n the central nervous system may be very different, depending
Fig . 39 .—Diagram of an arrangem ent of neurons adapted for the d is
tribution of a single afferent nervous im pulse to several different m otororgans .
on the connections of the pathways which are taken by the neurons Of thesecond order. If these pathways diverge so that the stimulus is d istributedamong several different effector system s , thi s would tend to disperse theenergy Of the afferent impulse and a relatively strong stimulus is necessaryto call forth a response . This is the situation in case a painful prick on theskin of the face calls forth reflex m ovem ents of, say ( 1 ) twitching of thefacial m uscles ; (2 ) turning the head away, and (3 ) a m ovem ent of the handto rem ove the irritant . Here the stim ulus aris ing at a single point in theskin (Fig. 39 ) is distributed to three widely separated m otor centers (M .1
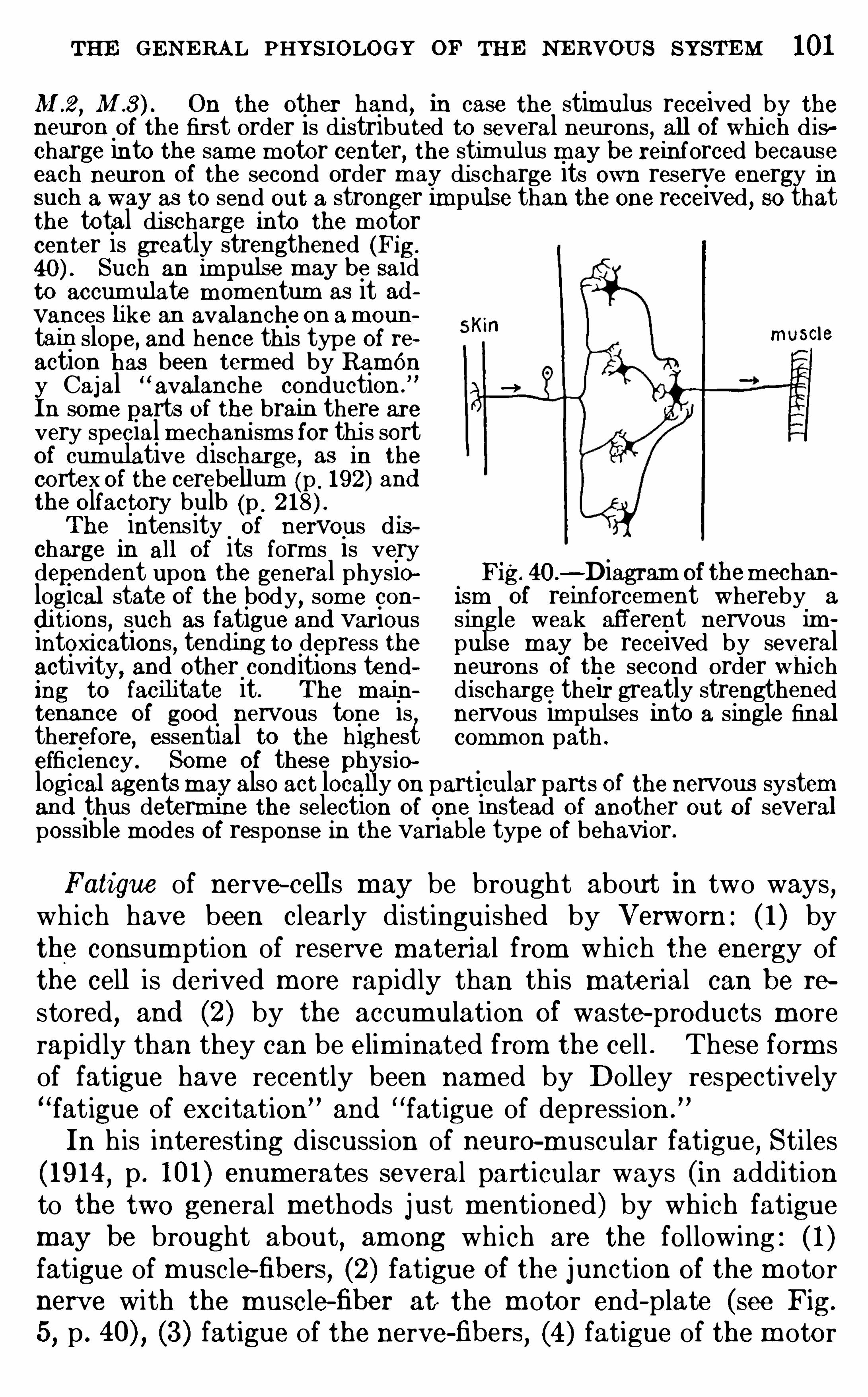
THE GEN E RA L PHYSIOLOGY OF THE NERVOUS SYSTEM 101
M .2 , M On the other hand, in case the stim ulus received by theneuron of the first order is distributed to several neurons , all of whi ch d ischarge into the sam e m otor center
, the stimulus m ay be reinforced becauseeach neuron of the second order may di scharge its own reserve energy insuch a way as to send out a stronger impulse than the one received , so thatthe total discharge into the m otorcenter is greatly strengthened (Fig.
Such an im p may be saidto accum ulate m om entum as it advances like an avalanche on am oun
tain slope, and hence this type of reaction has been term ed by Ramony Cajal “
avalanche conduction .
”
In som e parts of the brain there arevery special m echani sm s for thi s sortof cumulative discharge, as in the
cortex Of the cerebellum (p . 192) andthe olfactory bulb (p .
The intensity of nervous d ischarge in all Of its form s is very
muscle
dependent upon the general physiological state of the body, som e con
d i tions, such as fatigue and var iousintoxi cations , tending to depress theactivity
,and other conditions tend
ing to facilitate it. The m ain
tenance of good nervous tone is,
therefore, essential to the highest
Fig. 40.—Diagram of them echan
ism of reinforcem ent whereby a
single weak afferent nervous impulse may be received by severalneurons O f the second order whi chdi scharge their greatly strengthenednervous im pulses into a single finalcomm on path .
efficiency . Som e of these ph sio
logi cal agents may also act loca y on particular parts of the nervous systemand thus determ ine the selection of one instead of another out of severalpossible m odes of response in the variable type of behavior .
Fatigue of nerve- cell s m ay be brought about in two ways,
wh ich have been clearly distinguished by Verworn : ( 1 ) bythe consum ption of reserve materi al from which the energy ofthe cell is derived m ore rapidly than th is material can be re
stored , and (2) by the accumulation of waste-products m orerapid ly than they can be elim inated from the cell . These form sof fatigue have recently been nam ed by Dolley respectively“fatigue of exc itation
”and
“fatigue of depress ion .
”
In h is interesting d iscussion of neurO-muscular fatigue, Sti les( 19 14, p . 101 ) enum erates several part icular ways (in addi tionto the two general methods just mentioned) by wh ich fatiguemay be brought about
, am ong wh ich are the following : ( 1 )fatigue of muscle-fibers
, (2) fatigue Of the junction of the m otornerve with the m uscle-fiber at the m otor end - plate (see Fig.
5, p . (3) fatigue of the nerve-fibers , (4 ) fatigue of the m otor

102 INTRODUCTION To N EUROLOGY
nerve- cells, (5) fatigue of the synapses between the nerve- cel ls,
(6 ) fatigue of the sense organs and afferent apparatus, (7) fatigueof the centers of voluntary control . The first
,second
,fourth ,
and fifth types comm only play a part in ordinary fatigue, theth ird is insignificant , and the sixth and seventh may be present .The synapses and the m otor end - plates are probably especiallysusceptible to fatigue of depression by toxic substances, and them uscle-fibers and nerve- cell bodies to fatigue of excitation byconsum ption of thei r m aterial .A resting neuron when exc ited to activity at first increases in
size by reason of the stim ulus given to general m etabol ic activity . The first signs of fatigue result from the exhaustion of theoxygen supply of the cells ; then fol lows the consum ption of the
reserve food materials,ch iefly those represented in the chrom o
ph ilic substance,with consequent shrinkage of the N issl bodies .
In extrem e fatigue the ultim ate dissolution and death Of the cel lm ay be hastened by the accum ulation of toxic products of cel lmetabolism .
It appears to be well established by numerous experim ental studies thatat the beg inning Of functional activity both the nucleus and the cytoplasmof the resting neuron are enlarged
,and that with the onset of fatigue
there is a shr inkage,especially of the nucleus, with vacuolation Of the
cytoplasm and solution of the N issl bod ies d ue to the consum ption of the
chrom ophilic substance during activity . The neurofibrils are also said tobe m odified during functional activity . A fter excessive activity they becom e m ore slender and apparently increase in num ber
,while during rest and
after hibernation of those anim als wh ich have this habit the neurofibrilsbecom e thicker and less num erous .Cells whose chrom oph ilic substance has been consum ed by activefunction m ay after rest return to the norm al form ; but if the excitation becarried beyond the stage of norm al fatigue, recovery Of the neuron is impossible and it gradually disintegrates, resulting in the permanent enfeeblem en t of the nervous system .
The Observations O f Dolley have suggested to him that the volum e of thenucleus bears a constant relation to the volum e of the cytoplasm in allresting nerve-cells of the sam e type . In varying functional states of excitation an d depression this m ass relation is disturbed in accordance with theform ula : A c t ivity finally results in a disturbance of the norm al nucleuscy toplasm ic relation in favor of the cytoplasm ( fat igue of excitation) , whiledepression resulting from accum ulated toxins finally results in a disturbanceof this relation in favor of the nucleus . In short
,the depression of the
neuron by any form of in toxicat ion or otherwise gives the converse p ictureof struc tural changes from that presented by fatigue of excitation .
Most of the physio logical work wh ich has been done upon fatigue has beendirected toward the iso la t ion of special toxic substances such as in Dollcy ’
s
schem e would prod uce “ fatigue of depression . It has been shown that

THE GE NE RAL PHYSIOLOGY OF THE N E RvOUS SYSTE M 103
prolonged m uscular exertion produces toxin s (carbon dioxid , lactic acid , andothers) which are dissolved in the blood and exert a profound depressinginfluence upon all of the tissues Of the body . If the blood of a fatiguedanim al be in jected into or transfused with a perfectly fresh an imal of thesam e species
,the latter imm ediately m anifests all the signs of fatigue .
It is often taught that a change O f work is physiologically equivalent tocomplete rest. It is true that
,so long as one is well within the lim its of
extrem e fatigue,a change of work wil l prolong efficiency far beyond that
which would be possible in continuous activity of a single nervous or mus
cular mechanism . N evertheless experim ent shows that m ental efficiencyis greatly im paired In extrem e m uscular fatigue, and ,
conversely,muscular
power i s greatly weakened after long sustained m ental work . Glandularsecretions are also apparently often reduced 1n extrem e fatigue, thus , forinstance
,reducing the efficiency of the digestive organs . These effects are
doubtless d ue to the accum ulation O f toxic products in the blood,producing
a true “fatigue of depression ” throughout the entire body .
It has been suggested that the local feelings of m uscular fatigue are d ueto excitations of the organs of the muscular sense in the m uscle sp indles (p .
but the evidence for this does not seem very convincing.
The experim ents of Dolley suggest to h im ,further
, that the m ore highlydifferentiated nerve- centers are m ore susceptible to the structural al terations Of fatigue than are those of the lower reflex system s . It i s a wellknown fact that sustained m ental work produces the subjective evidencesof fatigue m ore prom ptly than does m uscular work
, and that during severemental train ing one is m ore apt to go “stale ” than during physical training .
Th is principle has been widely recognized in the provision O f short working hours and frequent hol idays for pup ils and teachers in our schools ; itshould be still fur thcr extended
,especially In comm ercial and professional
life. Its neglect Is in large m easure responsible for the prevalence of neu
rasthenia and other form s of nervous breakdown .
The early fatigue of the higher voluntaIy centers is particularly evidentin young ch ildren,
where continuous sustained attention is im possible exceptfor veg;short periods . By training, these periods can be greatly length
ened e nervous m echan ism involved here probably being the acqu isiti on of a wider range of associations related with the subject which occupiesthe focus of attention
,so that indi vidual neurons or system s of neurons
which participate In the functional com p lex m ay be temporarily rested whileother related system s are brought into max imum activ i ty
,without thereby
interrupting the continuous progress of the train of thought.
The neurological basis of sleep is at present whol ly unknown,
though thephysiologi cal phenom ena seem to be In many respectsanalogous w ith those of fatigue . Of the various theories wh ichhave been suggested , the two wh ich have exc ited greatest interestare : ( 1 ) the belief that some solub le toxin is produced duringwaking hours wh ich induces sleep by a process sim i lar to that ofthe
“fatigue of depression ,
”and (2 ) the doctrine Of the retraction
of the neuron,which teaches that during sleep (and accord ing
to som e authors in less measure during fatigue also) the dendritesof the neurons retract toward their cel l bodies and away from

104 INTRODUCTION To NEUROLOGY
contact wi th the axons of other neurons with wh ich they are insynaptic union
,thus increasing the resistance to nerve cond uc
tion at the synapse.
Many physio logical experiments show that,though the pred is
position to sleep m ay be brought about by the accumulation of
toxins in the blood or by,
other general causes, the actual fal l ingasleep is accom panied by a fall in blood-pressure
,wh ich may be
the essential factor in sleep . Fatigue of the vasom otor centerhas been suggested as the real physiological cause of sleep . No
adequate proof of any of these theories has been brought forward .
The numerous theories regarding the neurological processestaking place in the cerebral cortex during the progress Of suchmental functions as attention
,association of ideas , etc .
,
are l ikew ise as yet entirely unproved . It has been suggestedthat during cerebral function the resistance of some pathwaysmay be d im ini shed by the am eboid outgrowth of the dendri tesso as to effect m ore intimate synaptic union w ith the physiologically related neurons, wh i le the resistance of other paths may
be increased by the retraction of dendrites from their synapses .O thers believe that the neuroglia m ay participate in the processby thrusting out ameboid processes between the nervous term inals in the synapses and thus increasing the resistance .Lugaro has suggested a different interpretation
,in accordance
w ith wh ich during sleep there is a general ly diffused extensionof all nervous processes , thus providing for the uniform diffusionof incom ing stim uli , wh i le in the state of attention all of theseprocesses retract save those wh ich are directed in some definitedirection,
thus narrowing the stream of nervous discharge so as tointensify it and direct it into the appropriate centers . There isno di rect evidence for any of these theories , and the scientifically correct attitude toward them is frank ly to adm it that at
present we do not know what physiological processes are involved in any Of these functions .Summary—The form s assum ed by neurons are shaped in
part by thei r nutritive requi rem ents and in part by their functional connections . The m etabol ism of nervous protoplasm
, as
m easured by its C0 2 output, is found to be as active in nervefibers as in the cell bodies . In a nerve-fiber the m etabo lic

CHAPTER VI I
THE G ENERAL ANATOMY AND SUBD IVISION OF THE
NER VO US SYSTEM
ON m erely topographic grounds the nervous organs are dividedinto the central nervous system ,
or axial nervous system,com pris
ing the brain and spinal cord,and the peripheral nervous system ,
includ ing the cranial and spinal nerves , their gangl ia and peripheral end—organs , and the sym pathetic nervous system . The
nerves are sim ply conductors , putting the end - organs into physr
iological connection with thei r respective centers . The generalform of the hum an central nervous system and its connectionsw ith the peripheral nerves are seen in Fig. 4 1 . The nervesconnected with the spinal cord are the spinal nerves
,those con
nected w ith the brain are the cranial or cerebral nerves,and
both of these system s of nerves together are cal led the cerebrospinal nerves
,in contrast w ith the sym pathetic nerves
,wh ich
latter m ay or m ay not be connected w ith the central nervoussystem (see p .
The central nervous system is the.great organ of correlation
and integration of bodily processes . I ts prim itive form in vertebrates is a sim ple tube
,and th is is the form shown in an early
human embryo (see Fig. 46,p . The original tubular form
is but li ttle m od ified in the trunk region of all vertebrates , wherethe spinal cord (m edulla Spinalis) is form ed by a tolerably uniform th ickening Of the lateral walls of the tube (see Figs . 41 ,But in the head region the brain (encephalon) is form ed by thevery unequal th ickening of d ifferent parts of the wal ls of the tubeand by various fold ings brought about thereby . The generalarrangem ent of the human central nervous system at successivestages of developm ent is seen in Figs . 47—5 1 .
The external form of the brain has been shaped by the spacerequirements of the nerve- cells and fibers wh ich make up itssubstance . A group of nerve- cells wh ich perform s a Singlefunction is often spoken of as the “
center” of that function ; but
- I CER VICAL NE RVE
MIDDLE OER VICAL SYMPA THETI0GANGLION
INFERIOR CER VICAL SYMPA
fNGLIAm p 003 »
I
l GANGL ION
LUMBAR NER VB
3AORAL NE R VE
FIL UM TERMINALE
Fig. 41 .—The human central nervous system from the ventral side
,
illustrating also its connections w ith the cerebro—spinal nerves and wi ththe sym pathe t ic nervous system ,
the lat ter drawn in black . (A fter A llenThom pson and Rauber
,from Morris ’ Anatom y . )

108 INTRODUCTION TO NEUROLOGY
it should be borne in m ind that thi s does not im ply that thi sfunction resides exclusively in that place . These functions areall m ore or less com plex and the “center is usually the regionwhere various nervous im pulses are received and redistributed ;it is
,therefore
,roughly analogous w ith the sw itchboard of an
electric plant .The nerve-fibers wh ich conduct nervous im pulses toward a
given center are called afierent, and those wh ich conduct awayfrom the center are called efferent w ith reference to that center.Most of the peripheral nerves are m ixed, in the sense that theycarry both afferent and efferent fibers w ith reference to thecentral nervous system . The efferent fibers may excite m ovem ent in musc les (m otor fibers) or secretion in glands (excito
glandular fibers) ; other efferent fibers which check the actionof the organ to which they are d istributed are cal led inh ibitoryfibers . The afferent fibers of the peripheral nerves are oftencal led sensory fibers
,though it must be borne in m ind that their
excitation is not always fo l lowed by sensations or other consciousprocesses .The vertebrate nervous system when exam ined in the fresh
cond ition is found to be made up of wh ite matter (substantiaalba) and gray matter (substantia grisea) , the wh ite mattercontaining ch iefly nerve-fibers with myelin sheaths (see p . 46)and the gray matter nerve- cel l bodies and unmyelinated fibers.The centers are
,therefore
, generally gray in color and the intervening parts of the central nervous system are whi te.
A group of nerve-cells constituting a center as above described is O ftencalled a “
nucleus ,” a term which has nothing to d o with the nuclei of theind ividual cells (see p . 39 ) Of which the center is com posed . Som e criticalwriters use the word “nidulus” (originally suggested by C . L . Herrick) or“nidus” (S itzka) for such a center, thus avoiding the ambiguity in the use
of the wor nuc leus . The term “ganglion” is also som etim es used for nuclei
or centers within the brain (ganglion habenulaa, ganglion inte ed unculare,but this usage is objec tionable, for the use of the wor ganglion in
vertebrate neurology should be restricted to collections of neurons outsidethe central nervous system ,
such as the ganglia Of the cranial and spinalnerves and the sym pathetic ganglia.
A nucleus from which nerve~fibers ar ise for conduction to som e rem otepart of the nervous system is called the nucleus of origin of these fibers ;conversely, a nucleus into which nervous im pulses are discharged by fibersarising elsewhere is the term inal nucleus Of those fibers . Any correlationcenter is, therefore, a term inal nucleus for its afferent fibers and a nucleusof origin for its efferent fibers .

1 10 INTRODUCTION To NEUROLOGY
brain are indicated ; and Fig. 44 shows an enlarged side v iewof the brain and the sensory roots of the cranial nerves .
n d J F
Fig. 42 .—Dissection of the brain and cranial nerves of the d ogfish ,
Scyllium catulus . The right eye has been removed . The cut surfacesof the cart ilaginous skull and spinal column are dotted . cl .I—cl .5 , Branch ial (gill) clefts ; ep.
, epiphysis ; exl .rect. , external rectus m uscle of the
eyeball ; gl .ph ., gIOSSOpharyngeal nerve ; hor.can . , horizontal sem icircular
canal ; hy.mnd .Vl I, hyomandibular branch of the facial nerve ; i nf .obl . ,inferior Oblique m uscle ; i nt.rect.
,internal rectus m uscle ; lat.vag. ,
lateralline branch of the vagus nerve ; mmLV, m andibular branch of the trigem inalnerve ; mx.V, maxil lary branch of trigem inus ; olf .cps .
,olfactory capsule ;
alf .s .,olfactory sac ; oph .V.VII, superficial ophthalm ic branches of thetrigem inal and facial nerves ; p at/t . , trochlear nerve (patheticus ) ; pt.V1 1 ,
palatine branch of facial nerve ; s .abt. , superior obhque m uscle ; sp.co .,
sp inal cord ; spi n ,spiracle ; s .rect.
, superior rectus muscle ; vag.,vagus nerve ;
vest,vestibule . (After Marshall and Hurst, from Parker and Haswell ’s
Zoology.)
In fishes there is a system of smal l sensory canals w idely d istributed under the skin . These contain sense organs som ewhatSim ilar to those in the sem icircular canals of the internal ear, and

ANATOMY AND SUBDIVISION OF NERVOUS SYSTEM 1 1 1
their functions are probab ly interm ed iate between those of theorgans of touch in the Skin and those of the internal ear, respond
r. ophthal . superfic. V
r. ophthal . superfic. VII
11. te rminal isOlfacto ry bulb r. ophthal. profundusV
Opti c nerve (II. II)
Gang. gemculi VII
Gang. later . VIIr. prespirac. VII
Spiracler . hyomand ib. VII
r. lateralis Xr branchialis Xr. intestinalis X
Fig . 43 .
—Diagram of brain and sensory nerves of the. sm ooth d ogfish
,Mustelus canis
,from above . Natural size . The R oman num erals
refer to the cranial nerves . The olfactory part of the brain is dotte d , theVisual centers are Shaded with oblique. cross-hatching
,the acoustico—lateral
centers with hori zontal lines , the v isceral sensory area wi th vertical lines,and the general cutaneous area is lef t unshaded . O n the right side thelateral line nerves are drawn in black
,the o ther nerves are unshaded .

1 12 INTRODUCTION To NEUROLOGY
ing to water vibrations of Slow frequency and probab ly assistingin the orientation of the body in space. These are the lateralline canals . They are innervated by Special roots of the VI I andX pairs of cranial nerves (the lateralis roots of these nerves) ,wh ich are drawn in b lack in Figs . 43 and 44 . The other nervesare lightly shaded or wh ite. The lateral l ine organs and theirnerves are entirely absent in h igher vertebrates (see p .
The lateral line nerves and the acoustic nerve (VI I I pair) infishes term inate in a comm on center w ithin the brain (the acousti co—lateral area) , wh ich is shaded with horizontal cross-hatching in Figs . 43 and 44 . The nerves of general cutaneous sensibility also term inate in a particular region wh ich is unshaded
r . byomand ibularis VIISpiracler. palatinus VII
Ganglion geniculi VII
Fig . 44 .—The sam e brain as Fig. 43 seen from the side and slightly enlarged .
and marked “general cutaneous area. The visceral nerves
from the gi lls , stomach , etc .,all enter a single
“Visceral area,
”
wh ich is shaded w ith vertical l ines . The eye is also connectedwi th a spec ial region in the m idbrain,
the“optic lobe,
” which isShaded with Obl ique cross- hatch ing ; and the nose is connectedw ith a part of the forebrain wh ich is stippled .
We may, therefore, recognize in th is fish a nose brain an
eye brain, an“ear brain,
”a“Visceral brain,
”and a skin
brain,
each of these peripheral organs having enlarged primaryterm inal nuclei wh ich make up definite parts of the brain sub
stance. Remembering that the prim itive brain was a sim pletubular structure
,we observe that each one of the ch ief sense
organs and each group of sim i lar sense organs sends sensory

1 14 INTRODUCTION To NEUROLOGY
If in the hum an brain we dissect away the cerebral cortex andthe cerebel lar cortex and the wh ite matter imm ediately con
Nucleus lentiformis Capsula interns (pars lenticulo—thalamica)Nucleus caudatus
atumlaterals
olfactorius Corpus pmcalcCor. geniculatum medialeCol liculus superiorColhculus inferior
Tuber cinereum Nervas tmchleans.
Corpus mamillare Brachiurn oomunctwumN . oculomotoriusBasis pedunculiPons
Nervus trigeminus (portio major )Nervus trigeminus (portio minor)
I
Corpus ponw-bulbareOliva$
Fasciculus circumolivaris pyramidis $ F35010111113 Spinocel‘ebellafl s
spinalis
Fig. 45 .
—Left lateral aspect of a human brain from which the cerebralhem isphere (with the exception of the corpus striatum ,the olfactory bulb
and tract, and a small portion of the cortex adjacent to the latter) and thecerebellum (excepting its nucleus dentatus ) have been removed . The
brain stem (segm ental apparatus , palaeéncephalon) includes everythinghere shown with the exception of the strip of cortex above the tractusolfactorius and the nucleus dentatus . Wi th in its substance, however, arecertain cortical dependencies (absent in the lowest vertebrates ) , whichhave been developed to facilitate comm unication between the brain stemand the cerebral cortex . The chi ef of thes e are found in the thalamus ,bas is pe dunculi, and pons . Com pare this figure w ith the side view of theintact brain
,Fig. 54 (Modified from Cunningham ’s Anatomy .)
nected therewith we have the form shown in Fig. 45 . Th is isthe human brain stem .

ANATOMY AND SUBDIVISION O F NERVOUS SYSTEM 1 15
The cerebellum appears in the evolutionary h isto ry of thevertebrate brain much earlier than the cerebral cortex ; itsfunctions are wholly reflex and unconscious (see pp . 158, 186)and are concerned ch iefly with m otor coord ination,
equi libration,
and,in general , the orientation of the body and its m em bers
in space. Its activities are of the invariable, innate, structural lypredeterm ined type (see pp . 22
,31
, The cerebral cortex,on the other hand
,is the organ of the h ighest and m ost plastic
correlations,wh ich are in large m easure ind ividually acquired .
It attains its m axim um size in the human brain .
In recognition of the late phylogenetic origin of the cerebralcortex E dinger has cal led the entire brain stem and cerebel lumthe old brain (palaeencephalon) , and the cerebral cortex and
parts of the brain developed in relation therewi th the new brain(neencephalon) .The term inology of the brain is in great confusion . Most of
the m ore obv ious parts were nam ed before their functions wereknown,
the same part often receivingmany di fferent nam es,and
som etimes the sam e nam e being applied to very d iflerent parts .To rem edy th is Situation the German Anatom ical Society in1895 publ ished an official li st of anatom ical term s wh ich is knownas the Basle N om ina Anatom ica (comm only abbreviated as
B . N . Each of these term s has a clearly defined significanceand they are now very widely used , though many anatom istscontinue to use some older and unoffic ial nam es . The B . N . A .
term s or their English equivalents are used in this work, save ina few cases which are Specifically m entioned . The term inologyof the brain is based upon the em bryological researches of Professor His, and can best be outl ined by reviewing the form of thehum an brain at a few selected stages of developm ent .
The B . N . A . term inology was developed with exclusive reference to thehuman body . The nam es ofm any parts of the bodies of other animals thanm an and of m icroscop ic structures in general are not included . The nam esof thi s li st are all used and defined In W . Krause ’ s Handbuch d er Anatom icd es Menschen
,Leipzig
,1 905 , and in m ost of the recent Am erican and
E ngli sh text-books O f anatom y . At the end ofKrause ’
3 book 18 a Very complete li st of synonym s , including m ost of the anatom ical term s in use and
their B . N . A . equivalents .Following the exam ple of m any other recent anatom ists , we shall In this
work replace the B . N . A . term ‘‘anterior” (on the front or belly side) by the
word “ventral,
”and the B . N . A . term posterior” (on the back side) by

1 16 INTRODUCTION To NEUROLOGYthe word “dorsal . The head end of the body wi ll be referred to as the“anterior or
“cephalic” end ; the other end of the body as the “posterior”or
“caudal” end . The term s upper” or
“higher and“lower will refer
to the relations in the erect hum an body . In the nom enclature of the
m edulla Oblongata (see p . 122 ) and of the thalamus (p . 167 ) our usagedeparts Slightly from that of the B . N . A . R egarding the nami ng of fibertracts see page 128.
Figure 46 i llustrates the form of the brain in a very earlyhuman embryo . Its tubular form is very evident
, and in the
fclcncephalonstriatum
Me nnupha/an
Masencep‘u /on
Rhanb’encepba/on
flbombm cep/wmflexure
Fig. 46 .—An enlarged m odel of the brain of a human em bryo mm .
long (about two weeks old ) . The outer surface is shown at the left, andon the right the inner surface after division of the m odel in the m edianlane . The Anterior neuropore m arks a point where the neural tube
Is st ill 0 en to the surface of the body . The Pall ium is the region fromwhich the cerebral cortex will develop . The Opti c recess m arks theportion of the lateral wall of the Di encepha lon from whi ch the hollowOptic vesicle has evaginated . (Af ter His, from Prentiss
’
Em bryology .)
brain the diameter of the tube is but little greater than thatof the spinal cord . The walls are th in and the cavity wide .
In a slightly older em bryo the form is Shown in Fig. 47, and
Fig. 48 i llustrates diagramm atically the m edian section of anem bryo of about the sam e age as that shown in Fig. 47
, uponwhich the regions as defined by the B . N . A . are indicated . The

Tek ncephalon.
(End -brai n)
Rhinenccphalom' lerm inalir strwtum
(Ollad ory~brai n)
Fig. 49 .
—Vertical m edian section of a m odel of the brain of a hum an
em bryo mm . long : 1 , Optic recess, m arking the attachm ent of theoptic vesicle ; 2 , ridge form ed by the optic chiasma ; 3 , optic ch iasm a ;
4, infundibular recess . The lim iting sulcus is visible in the m odel , thoughnot nam ed
,running upward from the Optic recess between the thalamus and
the hypothalamus . (Af ter His, from Sobotta’
s Atlas of Anatom y .)
Thalamusithalamus (Corpus pineale)Metathalamns
orpora geniculata)
Corpora quad rlgeminaCorpus striatum .
Ped uncnlus cerebri
Rh inencephalonPars optics hypothalam i FOB“ rhombmd ea
Ch lasma opticum '
l
'
Hypophysi s Med ul la OblongataPars mammaris hypothalam i
Pons [Varoll]
Fig. 50 .—A vertical median section of a m odel of the brain of a hum an fetusin the third month . (After His , from Spalteholz
’s Atlas .)
1 18
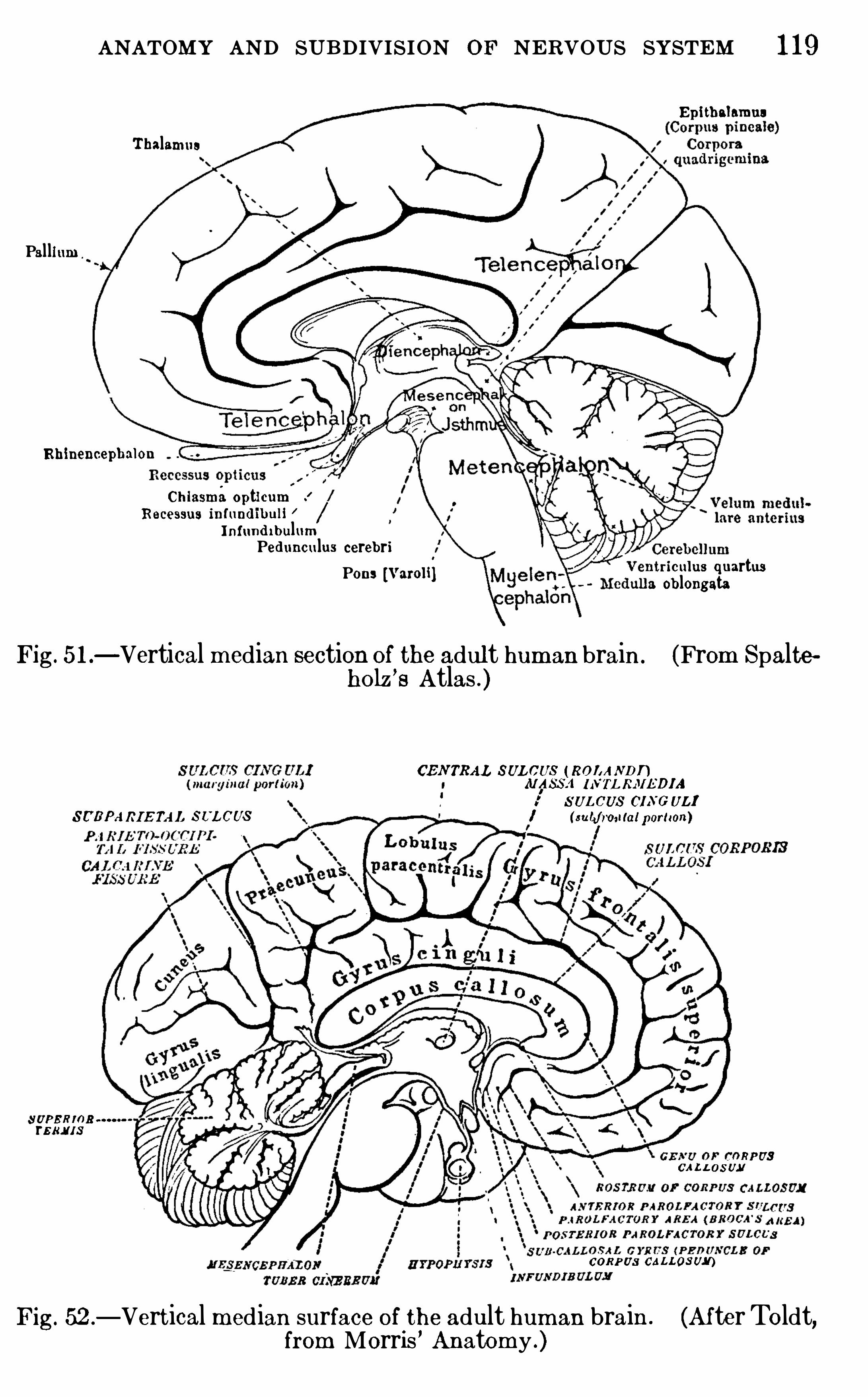
ANATOMY AND SUBDIVISION OF NERVOUS SYSTEM 1 19
Epi tbalamus
Thalamus
Pal lium
Rhinenoepbalon
Ch iasmaOpti cumRecessus infund ibull
Infund i bulumPed unculus cerebri
Pons [Varoli ]riculus quartus
a Oblongata
Fig. 5 1 .—Vertical median section of the adult human brain . (From Spalte
holz’
s Atlas .)
CENTRAL SUL CUS ( 120 /A ND”MASS/1 IIVTL RI‘IEDIA
,o'
SUL CUS CIA'
G UL$
SUBPARIE TA L SL‘
L CUS I ( sub/mu{atportion)
SUPERmsrsm as
k rvCAL L OSUM
CORP US CALLosvy )
Fig. 52 .—Vertical m edian surface of the adult hum an brain . (AfterToldt,
from Morris ’ Anatomy .)

120 INTRODUCTION To NEUROLOGY
The brain as a who le is the encephalon, and its chief d ivisions
are indicated by prefixes having a topograph ic Significance ap
pli ed to this word . In Fig. 48 the ventral part of the neural tubeis Shaded to indicate the region in wh ich the m otor centers of theadult brain are found . The unshaded part of the figure ind icates the region devoted to the primary sensory centers and the
Opti c chiasmaOlfactory bulb
Left corpus mami llareOlfactory tract
Substantia perforataposteriorPed unculus cereb
Optic tract
Trochlear nervenerve
geal nerveHypoglossal n
Medulla OblongataMedul la spinalis (cut) Hyposloseal nerve
Fig. 53 .- Ventral View of the adult human brain . Compare Fig. 41 .
(From Cunningham ’s Anatomy . )
correlation centers related to them . The sensory and m otorregions are separated in early em bryo logic stages by a longitud inal lim iting sulcus (the sulcus l im itans) . Com parison withthe figures of later stages which follow Shows that the suprasegm ental structures are developed wholly from the sensoryregion . Figures 49 and 50 i llustrate later stages of develop

122 INTRODUCTION To NEUROLOGY
The isthmus is a sharp constriction which separates the braininto two m aj or d ivisions
,the rhom bencephalon behind and the
'
cerebrum in front . In the B . N . A .
’
tab le the isthmus is regardedas a transverse segm ent or ring ; it m ight better be regardedSim ply as a plane of separation between the rhom bencephalonand cerebrum . In the tab le the m edulla Oblongata is regardedas synonym ous w ith m yelencephalon,
that is,the region between
the pons and the spinal cord . The o lder usage, wh ich is sti l lw idely current, regards the m edul la Ob longata as includingeverything between the isthmus and the Spinal cord except thecerebe llum dorsal ly and the fibers and nuclei of the pons and m idd le peduncle of the cerebellum ventrally . Th is is the old or segmental part of the rhombeneephalon,
and the cerebellum and
pons fibers related to it are added to this prim itive m edullaOblongata. The older usage is preferab le to the B . N . A . divisionand w i ll be adopted here
,for the medulla Oblongata as here
defined is a structural and functional unit, whose form is notm odified in those animals wh ich alm ost total ly lack the cerebellum and its m iddle peduncle. Them idbrain (mesencephalon)is the least m od ified part of the neural tube in the adult brain .
The betweenbrain (diencephalon) has three principal d ivisions :( 1 ) be low is the hypothalam us ; (2) above is the epithalamus ;(3) between these is the thalam us wh ich includes the thalam usand m etathalam us of the tab le (see p . The hypothalam usand epithalam us are high ly developed in the lowest vertebratesand are related to the olfactory apparatus ; in these brains thethalam us proper is very smal l
,th is part increasing in size in the
h igher anim als parallel w ith the evolution of the cerebral cortex .
The thalam us proper is real ly a sort of vestibule to the cerebral cortex ; all nervous im pulses wh ich reach the cortex , exceptthose from the olfactory organs, enter it through the thalam us .The endbrain (telencephalon) includes the cerebral hem ispheresand a very sm all part of the prim itive unmodified neural tubeto wh ich the hem ispheres are attached
,th is being the pars
Optica hypothalam i of the tab le or, better, the telencephalonmedium .
I f now we com pare this subd ivi sion of the human brain w ithour rough functional analysis of the fish brain (p . we noticethat the “
ear brain”
(acoustico- lateral area) , skin brain” or

ANATOMY AND SUBDIVISION OF NERVOU S SYSTEM 123
face brain (general cutaneous area) , and visceral brain (visceral area) are all contained in the rhombeneephalon, whose segmental or stem port ion is made up of these centers and thecorrespond ing m otor centers . The same relations hold in the
human brain,and in both cases the cerebel lum (and in m an the
pons in the narrower sense in wh ich I use that term ) is added asa suprasegm ental part . In both cases the “
eye brain” includes
the retina of the eye, the optic nerve, and a part of the roof ofthe m idbrain . In the fish a very small part of
,
the thalamus
(not indicated on Figs . 43 and 44 ) also receives fibers from the
optic nerve . In man this optic part of the thalam us is greatlyenlarged , form ing so large a part of that structure in fact thatthe thalam us as a whole is often cal led the optic thalamus . I tShould be rem em bered
,however
,that even in man the optic
centers com prise only a part of the thalamus . The“nose brain”
of the fish com prises m ost of the cerebral hem ispheres (all exceptthe small “somatic area” of Fig. and all of the epithalamusand hypothalam us . In man these parts remain essential ly unchanged , but the
“somatic area” of the hem isphere has greatlyenlarged to form the large corpus striatum and the enorm ouscerebral cortex
,the latter form ing the suprasegmental apparatus
of the telencephalon,and greatly m od ifyi ng the form relations
of all adjacent parts .The details of the developm ent of the brain lie outside the
scope of this work,as also do the anthropological questions grow
ing out of the statistical study of brain weights1 and measurements . These and many other topics of fundamental im portance are presented in a very interesting way in Donaldson
’sbook on The Growth of the Brain .
Summary.
—In all vertebrates the central nervous system isfundam ental ly a hollow dorsal tube in wh ich the primary seg
mentation is subord inated to the development of im portantlongitudinal correlation tracts and centers . Th is tube is en
larged at the front end to form the brain . The vertebrate brainmay be d ivided on physiological grounds into great d ivi sions,
1 The weight of the brain is exceedingly variable, even in a homogeneouspopulation . The average weight of the norm al ad ult E uropean male brain15 comm only stated to be 1360 gram s (48 and that of the female 1250
gram s (44

124 INTRODUCTION To NEUROLOGY
first the brain stem,or primary segmental apparatus ; and second
the cerebellum and cerebral cortex,or suprasegm ental apparatus .
The brain stem and cerebel lum are devoted ch iefly to reflex andinstinctive activities and constitute the “old brain of Ed inger .The cerebral cortex is devoted to the h igher associations andindividually acquired activities and is called the “
new brainby E dinger . N o nervous im pulses can enter the cortex w ithoutfirst passing through the reflex centers of the brain stem .
In fishes the form of the brain is Shaped alm ost whol ly by thedevelopm ent of the reflex centers
,and here these m echanism s
can best be studied,each of the m ore obvious parts of the brain
being dom inated by a single system of sensori-m otor reflex cir
cuits . The sam e pattern is preserved in the human brain,but
much distorted by the add ition of the centers of higher correlation .
The term inology of the brain now in m ost comm on use isbased on its em bryological developm ent, wh ich is briefly re
v iewed .
LITERATU REBARKER
,L . F . 1907 . Anatom ical Term inology , Philadelph ia.DONA LDSON ,H . H . 1899 . The Growth of the Brain, a Study of the N er
vous System in R elation to E ducation, New York .
E DIN GE R, L . 1 908 . The Relations of Com parative Anatom y to Comparative Psychology , Jour . Comp . N eur .
,vol . xviii , pp . 437—457 .
HE RRICK,C . JUDSO N . 19 10 . The Morphology of the Forebrain in
Am phibia and Reptilia, Jour . Com p . N eur . , vol . xx , pp . 4 13—547 .
HIS, W . 1895 . Die anatom ische N om enclatur : N om ina Anatom ica,Archiv f . Anat . und Physiol . , Anat . Abt .
,Supplem ent-Band .
h.
J OHNSTON , J . B . 1906 . The Nervous System of Vertebrates, Philad elp Ia.
1909 . The Central Nervous System of Vertebrates, E rgebnisse undFortschritte d er Zoologie, Bd . 2 Heft 2
,pp . 1—170 .
1909 . The Morphology Of the Forebrain Vesicle in Vertebrates , J our.Com p . Neur ., vol . xix, pp . 457—539 ; also im portant papers on the sam e sub
j cet in later volum es of The Journal of Com parative N eurology .
KE IB E L,F . , and MALL
,F . P . 19 12 . Manual of Hum an Em bryology,
Philadelphia,vol . ii
,pp . 1—156 .
KRAU SE,W . 1905 . Handbuch d er An atom ic d es Menschen
,m it einem
Synonym enregister, auf Grundlage d er neuen Baseler anatom ischen N om en
clatur , Leipzig .
R ETZIUS,G . 1896 . Das Menschenhirn
,2 vols . , Stockholm .
SHER RIN GTO N ,C . S. 1906 . The Integrative Action of the Nervous Sys
tem, New York .
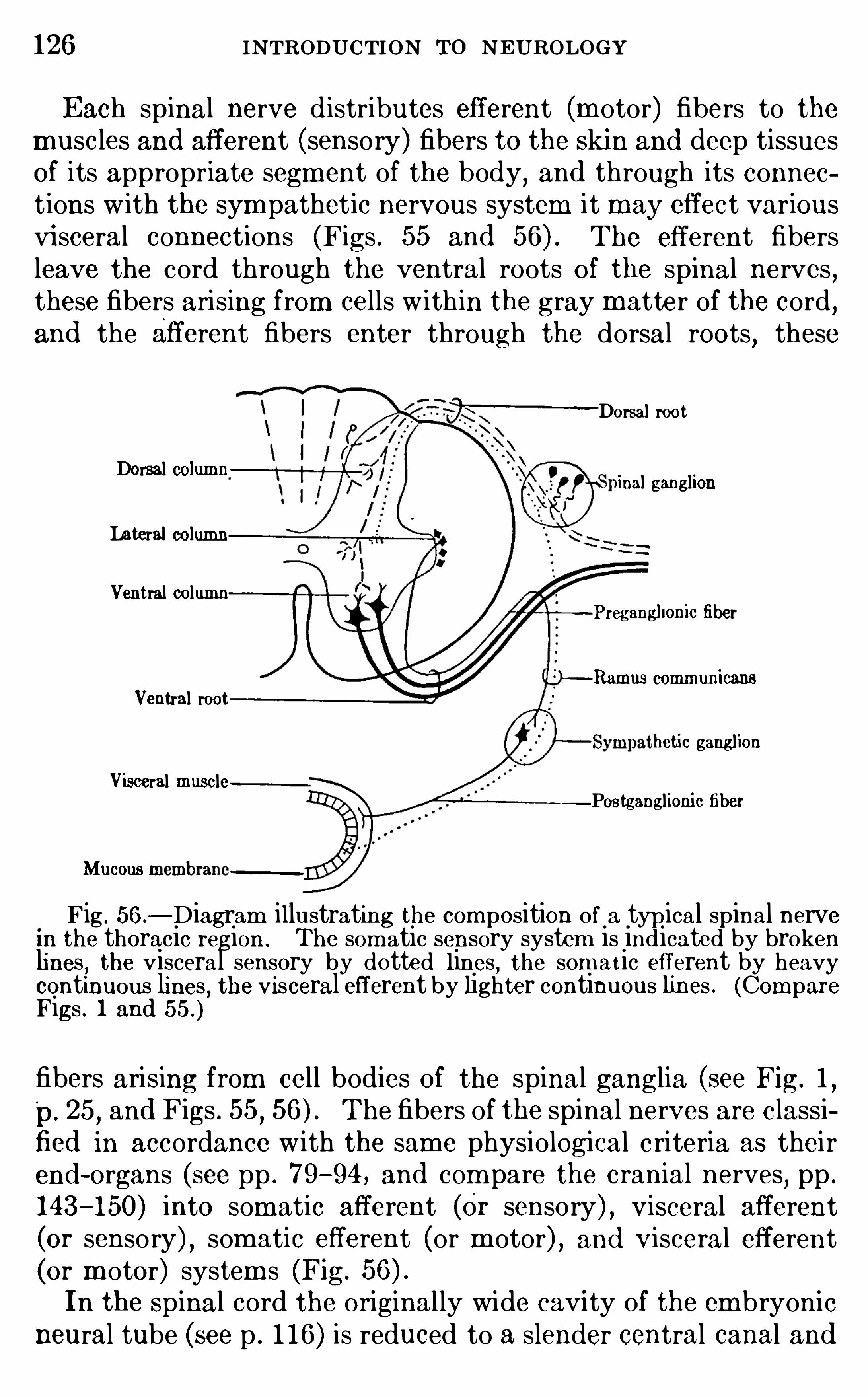
126 INTRODUCTION TO NEUROLOGY
Each spinal nerve d istributes efferent (m otor) fibers to themuscles and afferent (sensory) fibers to the Skin and deep tissuesof its appropriate segm ent of the body
,and through its connec
t ions w ith the sym pathet ic nervous system it may effect variousvi sceral connections (Figs . 55 and The efferent fibersleave the cord through the ventral roots of the spinal nerves ,these fibers arising from cells w ith in the gray matter of the cord
,
and the afferent fibers enter through the dorsal roots, these
Dorsal root1Dorsal co umn
inal ganglionLateral columnVentral column
Pregangllonic fiber
Ramus comm unicans
Ventral roothetic ganglion
Visceral muscle—Postganglionic fiber
Mucous
Fig . 56 .—Diagram illustrating the com position of a typ ical spinal nerve
in the thoracic region . The som atic sensory system is indicated by brokenlines , the visceral sensory by dotted lines , the som at ic efferent by heavycontinuous lines , the v isceral efferent by lighter continuous lines . (Com pareFigs . 1 and
fibers arising from cell bodies of the spinal ganglia (see Fig. 1,
p. 25,and Figs . 55 , The fibers of the spinal nerves are classi
fied in accordance w ith the same physiological criteria as thei rend - organs (see pp . 79—94 , and com pare the cranial nerves
,pp .
143—150) into somatic afferent (Or sensory) , visceral afferent(or sensory) , som atic efferent (or m otor) , and Visceral efferent(or m otor) system s (Fig.
In the Spinal cord the original ly wide cavi ty of the embryonicneural tube (see p . 1 16) is reduced to a slender central canal and

THE SPINAL CORD AND ITS NE RVE S 127
the walls of the tube are th ickened . The nerve- cells retain theirprim ary position bordering the central canal
,thus form ing a
mass of central gray matter wh ich is rough ly H- shaped in crosssection . Th is gray m atter on each side is accum ulated in theform of two massive longitudinal ridges , a dorsal colum n
(columna d orsali s,or posterior horn) , whose neurons receive
term inals of the sensory fibers of the dorsal roots,and a ventral
colum n (columna ventrali s, or anterior horn) whose neurons giverise to the fibers of the ventral roots .The wh ite m atter of the Spinal cord is superficial to the gray
and is made up of sensory and m otor root fibers of spinal nerves ,ascend ing and descending correlation fibers putting d ifferentparts of the cord into functional connection
,and longer ascend
ing and descend ing tracts by wh ich the spinal nerve- centersare connected w ith the h igher association centers of the brain .
In general , the shorter fibers lie near to the central gray and thelon7er tracts m ore superficially .
The wh ite m atter wh ich borders the gray in the spinal cordis m ore or less m ingled w ith nerve- cells and fine unm yelinatedendings, and thus shows under low powers of the m icroscope a
reticulated appearance. Th is is the reticular formation (processus reticularis) of the cord (see pp . 65
,158
,and Fig.
Imm ed iately surrounding the reticular formation and part lyem bedded with in it are m yelinated fibers belonging to neuronsintercalated between the sensory and the motor roots, wh ichrun for relatively short distances in an ascending or descendingd irection for the purpose of putting all levels of the cord intofunctional connection in the performance of the m ore com plexspinal reflexes . These fibers form the deepest layer of thewhite matter and are term ed the fasci culi propri i (dorsal is,lateralis
,and ventralis
,see Fig. These fascicles are also
called ground bundles and fundam ental columns .In the narrow space between the ventral fissure and the cen
tral canal (see Fig. 58) there is a bund le of nerve-fibers wh ichcross from one side of the spinal cord to the other . This is theventral comm issure. A sim i lar but smal ler dorsal comm issurecrosses imm ediately above the central canal .
There is considerable confusion in the term inology in use in the furtheranalysis of the sp inal white m atter, and the usage which follows d i ffers

128 INTRODUCTION To NEUROLOGYin som e respects from m ost of the classical descriptions , no two of whichagree am ong them selves . We shall lim it the appli cati on of the termfuniculus to the three m ajor divisions of the whi tematter of each half of thespinal cord , viz . , the dorsal funi culus bounded by the dorsal fissure and thedorsal root, the lateral funiculus lying between the dorsal and ventralroots , and the ventral funiculus between the ventral root and the ventralfissure (Fig.
E ach funiculus m ay be divided in a purely topograph i c sense Into
fasci culi or collections of nerve fibers which occupy the sam e general reg ionin the cross- section of the cord , such as the fasc i culus grac i li s of G 0 11 and thefasciculus cuneatus of Burdach (which together m ake up the greater partof the funi culus dorsali s , see Figs . 57 and and the superficral ventrolateral fasciculus of Gowers (including am ong other tracts the spine?tectal tract and the ventral spino- cerebellar tract ofFig. These fas c i culiare usually m ixed bundles containing tracts of di verse functional types .
Dorsal rootDorsal funiculusDorsal columnLateral funiculusLateral columnVentral columnVentral funiculus
Ventral rootFig . 57 .
—Diagram of a cross- section through one-half of the spinalcord to Illustrate the arrangem ent of the funiculi of white m atter and thecolumns of gray m atter.
The true physiological units of the Spinal white m atter are the tracts, i . e.,
collections of nerve-fibers of sim ilar functional type and connections .These tracts by som e neurologists are term ed fasciculi ; and , like the othertracts of the central nervous system , they are, in general , nam ed in accordance with the term inal relations of their fibers
,the nam e of the location of
their cells of origin preceding that of their place of discharge in a hyphenatedcom ound word . Thus
, the tractus cortico- spinalis arises from cells of thecere ral cortex (p . and term inates in the spinal cord , and the tractusspino-cerebellaris arises in the spinal cord and term inates in the cerebellum(p . But
,as already stated , there is no uniform ity in the nom enclature
of these tracts and no two authorities agree exactly in the term inologyadopted . Moreover, few Of the tracts have clearly defined anatom ical lim its ,in most cases the fibers of d ifferent system s being m ore or less mingled .
The appearance of a cross- section through the spinal cord inthe lower cerv ical (neck) region, after staining so as to reveal thearrangem ent of both the nerve- cel ls and the nerve-fibers, is seen

130 INTRODUCTION To NEUROLOGY
Tr. vestibule-spin.
Rad ix ventral isFig. 59 .
—Diagram Of a cross- section through the human spinal cord atthe level of the fifth cervical nerve, to illustrate arrangem ent O f the fibertracts in the white m atter and Of the nerve-cells in the gray m atter of theventral column . On the right side the area O ccupied by the dorsal graycolumn (posterior horn) is stippled ; on the left side som e Of the groups Ofcells of the ventral gray column (anterior horn) are indicated . In the whitem atter the outlines Of som e of the m ore im portant tracts are schematicallyindicated , ascend ing fibers on the right side and descending fibers on
.
the
left. The sam e area Of white m atter is in som e cases shaded on both Sidesof the figure. This indicates that ascend ing and descend ing fibers aremingled in these regions . A list of the tracts here illustrated follows . The
nam es here employed in som e cas es differ from those of the O fficial German
Anatom ical Society list (see p . the B . N . A . term s here being italicized .
ASCENDING TRACTSFasciculus graci lis (colum n of Goll) and fasci cu lus cuneatus (colum n Of
Burdach . ) These are m ixed bundles which in the aggregate m ake up the
greater part of the dorsal funiculus (O ld term ,posterior columns ) . They are
m ade up chiefly of the ascend ing branches of dorsal root fibers (see Fig .
those in the gracilis from the sacral,lum bar, and lower thoracic nerves (S,
L , T5 and those in the cuneatus from the Upper thoracic and cervicalnerves (TI—4, C) , as indicated in the figure . These fas ciculi term inateres
gectively in the nuclei of the fas ciculus gracilis (clava) and cuneatus
(tu erculum cuneatum ) at the lower end of the m edulla Oblongata (cf . Fig .and conduct chiefly im pulses of the proprioceptive reflexes and those
concerned with sensations of posture , spatial discrim ination,and the co
ordination O f m ovem ents of precision (see pp . 137 ,Fasc iculus dorso—lateralis (tract of Lissauer, Lissauer’ s zone) , made up
chiefly Of unm yel inated fibers from the dorsal roots , together with myelinated correlation fibers of the fasciculus proprius sys tem .
Tractus spino- cerebellaris dorsal is (fas ciculus cerebello-spi nali s, directcerebellar tract, Flechs ig
’
s tract) . These fibers arise from the neurons Of thenucleus d orsali s (Clarke ’s colum n O f gray m atter between the dorsal andventral gray columns in the thoracic region , also called St illing ’s nucleus )of the sam e side and en ter the cerebellum by way of its inferior peduncle(corp us restiforme) .Tractus spino-cerebellaris ventralis (part O f Gowers ’ tract, or thefascicu
lus antero—lalerali s sufierficia lis of the B . N . A These fibers also arise fromthe nucleus dorsalis of the sam e side (A . N . Bruce) in the lower levels Of the

THE SPINAL CORD AND ITS N E RVES 131
spinal cord and enter the cerebellum by way of its superior peduncle (brachium conj unctivum ) .The spinal lem niscus . Under th is nam e are included several tracts to
the mi dbrain and thalam us . These fibers arise from neurons O f the dorsalgray column,
cross in the ventral comm i ssure, and ascend in the lateral andventral funiculi Of the Opposite side, partly superficially m ingled with thoseof the ventral sp ino-cerebellar tract and partly deeper in the fasciculusproprius . This system of fibers includes a tractus spino—tectalis to the roo f(tectum ) of the m idbrain and a tractus sp ino-thalam icus to the ventral andlateral nuclei of the thalamus . The deeper fibers Of the latter tract arearranged in two groups, the tractus spino-thalam icus lateralis for sensoryim pulses Of tem perature and pai n
,and the tractus spino-thalam icus ven
tralis for sensory im pulses Of touch and pressure (see p . 138,Tractus spino-O livaris, fibers arising from the entire length of the sp inal
cord and term inating in the inferior O live (Goldstein ) .
DESCE NDIN G TRACTSTractus cortico—spinalis (fasci cu lus cerebro-sp
'inalis , pyrami dal tract) .This system Of fibers conducts voluntary m otor im pulses from the precentralgyrus of the cerebral cortex to the motor centers of the sp inal cord . It divides at the upper end of the spinal cord into two tracts , the larger divisionimm edi ately crossing through the decussation Of the pyram ids to the Opposite side Of the spinal cord , where it becom es the tractus cortico-spinalislaterali s (fasciculus cerebro—spinalis lateralis , lateral or crossed pyram idaltract) . A sm aller number Of these fibers pass downward into the spinalcord from the m edulla Oblongata without decussation to form the tractuscortico-spinalis ventrali s (fasci culus cerebro-spinalz
’
s anterior , d irect pyram idal tract, colum n Of Turck) . These fibers cross in the ventral comm issure a few at a tim e throughout the upper levels of the cord , and finally term inate in relation with the m otor neurons of the Opposite side. Both partsof the pyram idal tract, therefore, decussate before their fibers term inate .
Tractus rubro—spinalis (tract of Monakow ) , from the nucleus ruber Of themidbrain to the spinal cord , for thalam ic and cerebellar reflexes .
Tractus O livo-sp inalis (Helw ig’
s bundle, tractus triangularis) , fibers d escending from the inferior O li ve of the m ed ulla Oblongata to the lower cervical or upper thoracic segm ents of the s inal cord .
Tractus tecto—sp inali s (predorsal bun le, tract Of Lowenthal) , from the
roof (tectum ) of the m idbrain to the spinal cord , chiefly for Optic reflexes .Tractus vestibulo- spinalis , from the prim ary centers of the ves tibular
nerve in the m edulla Oblongata to the spinal cord , for equilibratory reflexes.The two tracts last m entioned
,together with several others , com pose the
fasc iculus m arginalis ventralis .
THE FASCICULUS PROPRIU SThe fasciculus proprius system Of fibers (also called ground bundles , bas is
bundles , and fundam ental bundles ) com prises chi efly short ascend ing anddescending fibers arising from neurons of the Spinal gray m atter, for intrinsicinal reflexes . In general , these fibers border the gray pattern ,
but in theoreal fun iculus som e are aggregated in the tractus septo—m arginalis and thefasciculus interfascicularis (comm a tract, tract of Schul t ze) , these two tractsalso containing descending branches of the dorsal root fibers . Some fibersOf the fasciculus proprius ventralis lie ad jacent to the ventral fissure and areterm ed the fasciculus sulco-m arginali s , these fibers form ing the directcontinuation into the cord Of the fas ciculus longitudinalis medial is (posteriorlongitud inal bundle) Of the brain (see pp . 185 ,

132 INTRODUCTION To NEUROLOGY
The sensory nerves which enter the Spinal cord com e eitherfrom the deep tissues or from the Skin
,and both Of these types Of
nerves carry fibers of very diverse functional sorts belonging tothe som atic sensory group , in addition to visceral fibers wh ichwi l l not be considered here. I t wi ll be recalled (see pp . 77
,79 )
that the general somatic sensory group includes : (1 ) proprioceptive system s
,concerned wi th m otor coordination and the orien
tation of the body and its m em bers in space (muscle sense, tend on sense, and (2 ) exteroceptive system s, concerned w iththe relations of the body to its envi ronm ent (touch , tem perature,and pain sensibi lity) . The first of these system s is servedchiefly by the deep nerves , and the second chiefly by the cutaneous nerves
,though th is is not rigidly true. In particular it
should be noted that,even though the Skin be com pletely anes
thetic , the nerves of deep sensibi lity can sti ll respond not onlyto their proprioceptive functions, but also to the ordinary clinicaltests for the exteroceptive qualities Of touch , tem perature, andpain
,though w ith a higher threshold than in the case Of the
cutaneous end - organs of these senses .Henry Head and h is colleagues have also separated the cuta
neous fibers into a protopath ic group (including cutaneous pain,
a diffuse non- localizable tacti le sensibi lity,and the d iscrim ina
tion of extrem e degrees Of tem perature) and an epicritic group( light touch , cutaneous locali zation
,d iscrim ination of inter
m ediate degrees Of tem perature and som e others) ; but there isd ifference of opinion as to whether these groups represent twodistinct sets of nerve-fibers or d ifferent stages in regenerationor d ifferent types of end - organs Of the sam e fibers (see p .
U pon entering the Spinal cord all of these functional typesof fibers effect two sorts Of connections : ( 1 ) for intrinsic spinalreflexes, and (2) for the transm ission of thei r im pulses upward tothe h igher centers of the brain. We Shal l first take up the in
trinsic connections .The sim plest Of these intrinsic connections is the direct m otor
reflex i llustrated by Fig. 1 (p . but there are many m orecom plex form s Of the connection between the dorsal and ventral roots , som e of wh ich are indicated in Figs . 60 and 6 1 . In
general , there is at least one neuron of the gray matter of thespinal cord interpolated between the dorsal and the ventral root
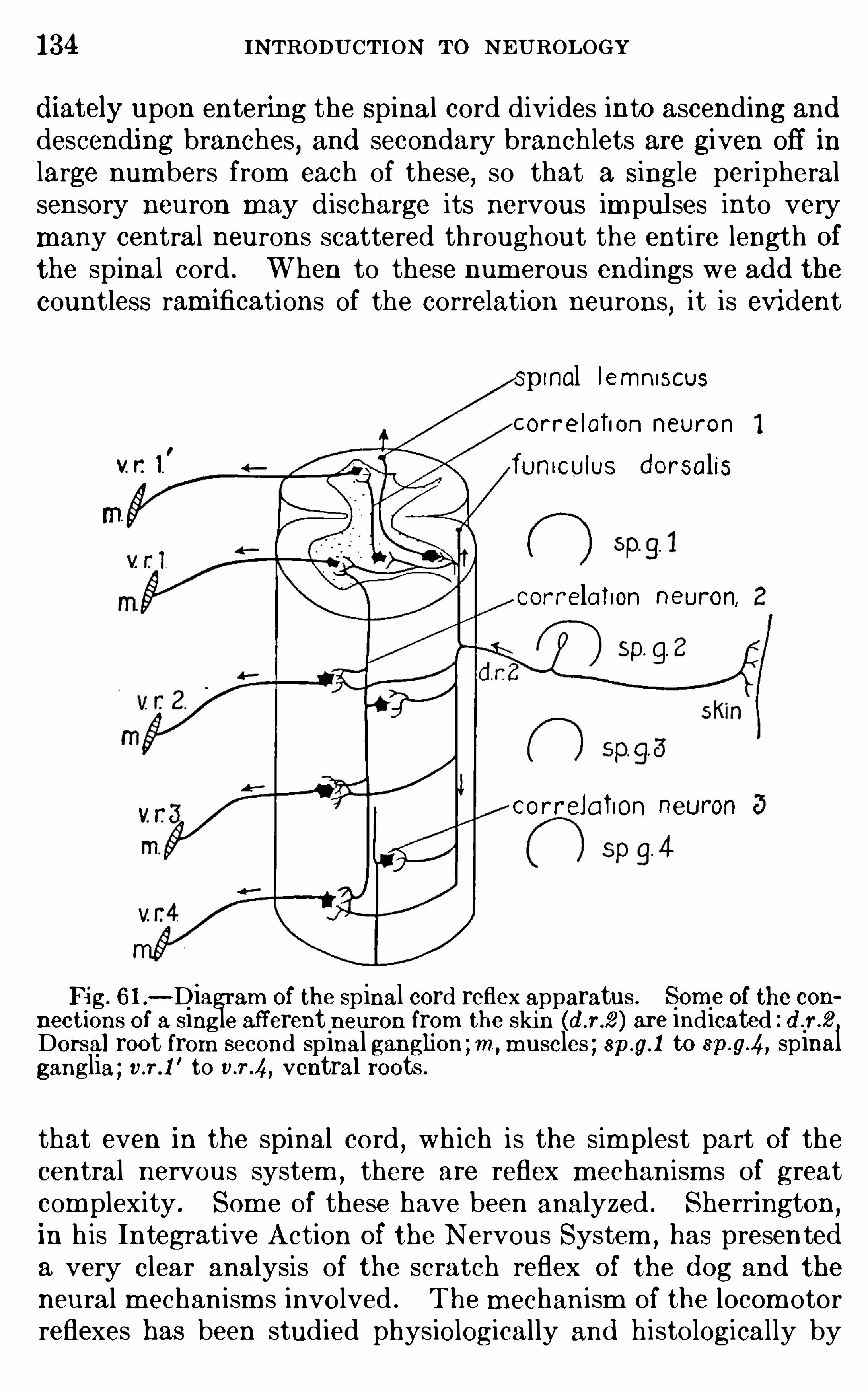
134 INTRODUCTION TO NEUROLOGY
d iately upon entering the Spinal cord divides into ascending anddescendi ng branches, and secondary branch lets are given off in
large num bers from each of these,so that a Single peripheral
sensory neuron may d ischarge its nervous im pul ses into verymany central neurons scattered throughout the entire length Ofthe spinal cord . When to these num erous endings we ad d thecountless ram ifications of the correlation neurons
,it is evident
p inal Iemnnscus
orre lahon neuron
funICU lUS d orsalis
Correlation neuron,
correlaluon neuron
Fig. 6 1 .—Diagram of the Spinal cord reflex apparatus . Som e of the con
nections of a single afferent neuron from the skin are indicated : d .r .2 ,Dorsal root from second sp inal ganglion ; m , m uscles ; 8p.g.1 to sp.g.4, spinalganglia ; v.r.1
'to v.r .4, ventral roots .
that even in the spinal cord,wh ich is the sim plest part of the
central nervous system ,there are reflex mechanism s of great
com plexity . Som e of these have been analyzed . Sherrington,
in h is Integrative Action of the N ervous System ,has presented
a very clear analysis of the scratch reflex Of the d og and the
neural mechanism s involved . The m echanism Of the locom otorreflexes has been studied physiologically and histologically by

THE SPINAL CORD AND ITS NERVE S 135
Steiner, Philippson,Polimanti
,Herrick and Cogh i l l, and very
many others .Our m ost precise knowledge of the arrangement of the afferent
and efferent myelinated fibers in the Spinal roots has been gainedby the application of March i ’s m ethod (p . 48) to the study ofdegenerations fol low ing acc idental .and experim ental injuries .Nerve-fibers wh ich have been cut Off from thei r cel ls Of origindegenerate w ith in about two weeks after the injury . It is ,therefore
,possible by the m icrOSCOpic study Of a d ivided nerve
wi th March i ’s m ethod (wh ich stains only the degenerating myelinated fibers) to determ ine on which side Of the injury are thecells of origin from which these fibers arise.
Figure 62 i llustrates the effects of section Of the spinal rootsm ade at four different places . In the first case section of the
m ixed trunk peripheral ly of the union Of the dorsal and ventralroots is followed by degeneration Of all Of the m yelinated fibersOf the nerve—trunk ,
showing that the cell bodies of all Of thesefibers lie centrally Of the injury . In the second case, section of
the ventral root c lose to the spinal cord is followed by degeneration of all the fibers Of thi s root without d isturbance of thoseof the dorsal root
,Show ing that the ventral root fibers arise as
axons of cells with in the spinal cord . In the thi rd case sectionOf the dorsal root fibers peripherally Of the ganglion and beforethei r union w ith those of the ventral root results in the degeneration of all Of the fibers of the m ixed nerve which arise in thespinal ganglion (sensory fibers) , without loss of any m otor fibersfrom the ventral root . In the fourth case section of the dorsalroot on the central Side of the ganglion is followed by degeneration of all m yelinated fibers Of the central stum p Of th is root
,but
not Of the peripheral part of the root or the spinal gangl ion.
This shows that the cells of origin Of these fibers lie in the spinalganglion and not
, l ike those of the ventral root, wi th in the spinalcord . The peripheral processes of these ganglion cells
,there
fore, are dendrites , and the centrally di rected processes wh ichcom pose the dorsal roots are axons (cf. Fig. 1
,p . 25
,and Fig.
56,p .
Another useful m ethod for the solution Of problem s Of th ischaracter is the study Of the fine structure of the cell bodies ofthe neurons after such experim ental lesions as those just d es
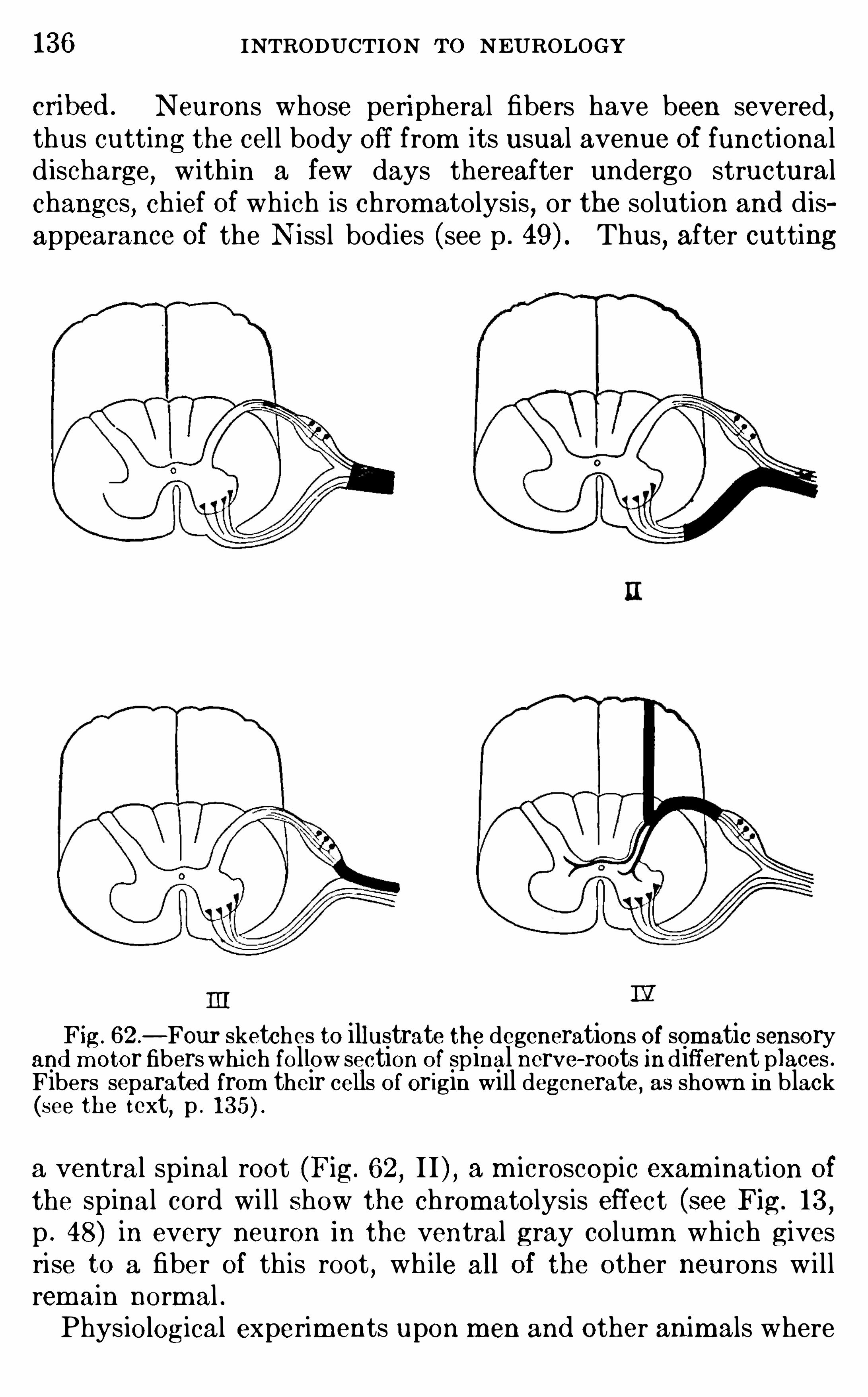
136 INTRODUCTION TO NEUROLOGY
cri bed . Neurons whose peripheral fibers have been severed ,thus cutting the cel l body off from its usual avenue Of functionald ischarge, with in a few days thereafter undergo structuralchanges, ch ief Of wh ich is chromatolysis
,or the solution and dis
appearance Of the N issl bodies (see p . Thus, after cutting
Fig . 62 .—Four sketches to illustrate the degenerations of somatic sensory
and m otor fibers whi ch follow section of sp inal nerve- roots in different places .Fibers separated from their cells Of origin will degenerate, as shown in black(see the text, p .
a ventral spinal root (Fig. 62,I I ) , a m icroscopic exam ination Of
the spinal cord wi l l Show the chromatolysis effect (see Fig. 13,
p . 48) in every neuron in the ventral gray column wh ich givesrise to a fiber Of this root , wh i le all Of the other neurons w i l lrem ain norm al .Physiological experim ents upon m en and other anim als where

138 INTRODUCTION To NEUROLOGY
only out Of a larger num ber of paths by wh ich afferent spinalim pulses may be d ischarged into it (see p .
The remaining proprioceptive fibers of the spinal roots ared irected upward in the dorsal funiculus
, Of wh ich they form the
larger part . At the point where the spinal cord passes overinto the m edulla Ob longata they term inate, and after a synapsehere the neurons Of the second order carry the im pulse across tothe Opposite side Of the brain and upward toward the thalam usin a tract known as the medial lemniscus or fillet (Fig.
After another synapse here,a final neuron may carry the nervous
im pulse forward to the cerebral cortex . Th is m edial lemniscussystem is largely concerned w ith unconscious m otor adjustm entsinvolving the muscles of the trunk and lim bs . Disturbance Of
its functions produces motor incoordination (ataxia) , but notnecessarily any great loss Of exteroceptive sensations . SO far as
its functions com e into consciousness, they are recognized as sensations of position
,spatial localization, and m otor contro l .
I I . E xteroceptive System s—The central course Of the extero
ceptive fibers Of the spinal nerves is quite different from thatjust described . Almost immed iately after entering the spinalcord these fibers term inate among the neurons of the dorsal
gray column . Af ter a synapse here the fibers of the second ordercross to the opposite side of the spinal cord , and here turn and
ascend in theWh ite matter of the lateral and ventral funicul i ,where they form thespinal lemniscus, or tractus spino- thalam icus .Some fibers Of the spinal lem niscus ascend throughout the entirelength of the Spinal cord
,m edul la Ob longata, and m idbrain, to
end in the thalamus . In the upper part of their course thesefibers accom pany those of the m edial lemniscus already d escribed .
Co llateral connections are effected between the ascend ingfibers of the spinal lem niscus and the various m otor nuclei Of thebrain for d ifferent cranial reflexes
,such as turning the eyes in
response to a cutaneous stimulation on the hand . But thei rfinal term inus is in the thalamus
,and after a synapse here the
nervous im pulse may be carried forward to the cerebral cortexby neurons of the th ird order . The spinal lem niscus system isthe chief ascend ing pathway for nervous im pulses giving rise toconsciousness Of touch , tem perature, and pain from the trunk
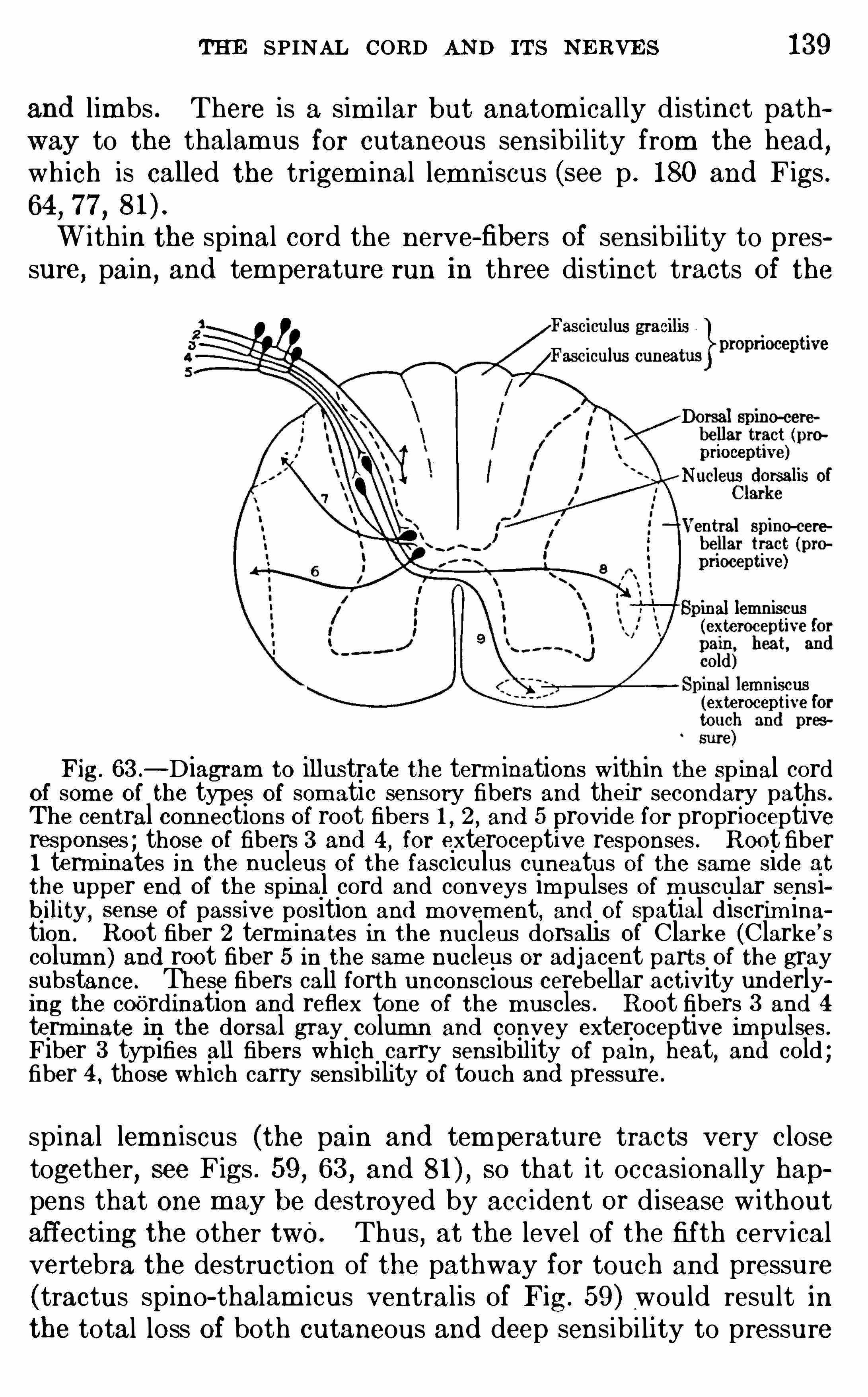
THE SPINAL CORD AND ITS NERVE S 139
and limbs . There is a sim i lar but anatom ical ly d istinct pathway to the thalamus for cutaneous sensibi lity from the head ,wh ich is called the trigem inal lemni scus (see p . 180 and Figs .64
,77,Within the spinal cord the nerve-fibers Of sensib ility to pres
sure, pain,and tem perature run in three distinct tracts Of the
Fig. 63 .—Diagram to illustrate the term inations within the Sp inal cord
of som e Of the types of som atic sensory fibers and their secondary paths .The central connections Of root fibers 1 , 2 , and 5 provide for proprioceptiveresponses ; those Of fibers 3 and 4 , for exteroceptive responses . R oot fiber1 terminates in the nucleus O f the fasciculus cuneatus of the sam e side at
the upper end Of the spinal cord and conveys im pulses Of m uscular sensibility,
sense of passive position and m ovem ent, and of Spatial discrim ina
tion . Root fiber 2 terminates in the nucleus dorsalis of Clarke (Clarke’ scolumn) and root fiber 5 in the sam e nucleus or adjacent parts Of the graysubstance. These fibers call forth unconscious cerebellar act ivity underlying the coordination and reflex tone of the m uscles . R oot fibers 3 and 4
term inate in the dorsal gray column and convey exteroceptive impulses .Fiber 3 typ ifies all fibers which carry sensibility Of pain
,heat, and cold ;
fiber 4 , those which carry sensibility of touch and pressure .
spinal lemniscus (the pain and tem perature tracts very c losetogether, see Figs . 59 , 63 , and so that it occasionally happens that one m ay be destroyed by accident Or d isease w ithoutaffecting the other two. Thus
,at the level of the fif th cerv ical
vertebra the destruction of the pathway for touch and pressure(tractus spino- thalam icus ventralis of Fig. 59 ) would result inthe total loss Of both cutaneous and deep sensibi li ty to pressure

140 INTRODUCTION To NEUROLOGY
over the whole Of the opposite Side of the body below the levelof the injury
,but there would be no d isturbance of either tem
perature or pain sensibi lity . Sim i larly, by an injury of the tractus spino- thalamicus laterali s, pain or tem perature sensibi litym ight be lost w ith no disturbance of pressure sense . (For thedescription of a case Of th is sort see p .
Such com binations Of symptom s as just described couldnot occur from any form Of injury to the peripheral nerves, forin these nerves the various kinds Of fibers are all m ingled in thelarger trunks , so that one functional com ponent cannot be ihjured w ithout involvem ent Of the others also . And at the firstdivision of these trunks into deep and superficial branches eachbranch also carries all or nearly all of the functional system s(see pp . 79—84
,
The return pathway for m otor nervous im pulses from the
cerebral cortex is the cortico—spinal tract or pyram idal tract(Fig. whose fibers descend without interruption from the
precentral gyrus of the cerebral cortex (see p . 283) to the spinalcord
,where they form the lateral and ventral cortico-Spinal
tracts (Fig. The various reflex centers of the brain stemalso send m otor fibers downward into the cord for the excitationOf m ovem ents of the trunk and limbs . The tecto—spinal tract(Fig. 59 ) is such a path
,leading from the Optic and acoustic
centers Of the m idbrain,as is also the vestibulo- spinal tract,
leading from the vestibular nuclei Of the m edul la Ob longata(p . 176, Fig. 83 , neuronSummary.
—The spinal nerves are segm ental ly arranged and
are nam ed after the vertebrae adjacent to wh ich they em ergefrom the spinal canal of the vertebral column . Each nervearises by a series of dorsal rootlets afferent in function and a
series Of ventral rootlets efferent in function . Most of the graymatter of the spinal cord is massed in two longitudinal colum nson each Side
,for somatic sensory and som atic m otor functions
respectively . These are separated by an intermediate regioncontaining the vi sceral sensory and m otor centers and vari ouscorrelation neurons . The wh ite matter Of the cord is superficialto the gray and contains myelinated fibers for various kinds Ofcorrelation
,besides root—fibers of the spinal nerves . The white
matter is divided topograph ically into funiculi and fascicul i and

142 INTRODUCTION To NEUROLOGY
according to Henry Head,protopath ic and epicritic functional
system s Of fibers . AS soon as the peripheral nerve-fibers haveentered into the spinal cord they are segregated into proprioceptive and exteroceptive groups, and each of these again intoparticular functional tracts . There are connections for localSpinal reflexes, reflexes Of the brain stem and cerebel lum
,and
for the cerebral cortex . The Spino- cerebel lar tracts and thedorsal funiculi are proprioceptive in function, and the spinallemni scus carries Spino- thalam ic tracts Of the system s Of touch ,temperature, and pain sensibi lity for the cerebral cortex .
LITERATU REBA RKE R , L . F . 1901 . The Nervous System and Its Constituent Neu
rones, N ew York .
BROUWE R , B . 19 15 . Die biologische Bedeutung d er Derm atom erie.
Beitrag zur Kenntnis d er Segm entalanatom ie und d er Sensibili tatsleitungim Ruckenm ark und In d er Medulla Oblongata, Folia Neuro-b i ologi ca Bd .
9 , pp . 225—336 .
BRUCE , A . 190 1 . A Topographic Atlas Of the Spinal Cord , London.
BRUCE, A . N . 19 10 . TheTract of Gowers , Quart . Journ . Exp. Physiol .,
vol . iii , pp . 39 1—407 .
HE AD,H.
,R IVE RS, W . H . R .
, and SHE RRE N,J . 1905 . The Afferent
Nervous System from a New A spect, Brain ,vol . xxviii , pp . 99—1 15 .
HE AD,H.
, and THOMPSON ,T. 1906 . The Grouping of the Afferent
Im pulses Within the Spinal Cord,Brain,
vol . xx ix , p . 537 .
HE RRICK, C . JUDSO N , and COGH ILL , G . E . 19 15 . The Developm ent ofR eflex Mechanism s in Am blystom a
,Jour . Com p . N eur . vol . xxv
,pp . 65—85 .
PHILIPPSON,M . 1905 . L
’
autonom ie et la centrali sation dans le system enerveux d es animaux
,Paris .
POL IMAN'I‘I, O . 19 1 1 . Contributi alla fisiologia d el sistema nervoso cen
trale e d el m ovim ento d ei pesci , ZOO] . J ahrb .
,Abt . f . ZOO] . u . Physiol ., Bd .
30, pp . 473- 7 16 .
R IVE RS, W . H . R ., and HE AD , H . 1 908. A Hum an Experim ent in NerveD ivision
,Brain
,vol . xxxi , p . 323 .
SHE RRIN GTON ,C . S. 1906 . The Integrative Action of the Nervous Sys
tem N ew York .
STE IN E R,J . 1885 . Die Functionen d es Centralnervensystem s und ihre
Phylogenese . I . Abte ilung . Untersuchungen fi ber d ie Physiologic d esFroschh irns, Braunschweig .
1888 . Idem . II . Abteilung,Die Fische.
1900 . Idem . IV . Abteilung, R eptilien-Ruckenm arksreflexe, Verm ischtes .
1886 . Ueber d as Centralnervensystem d er grunen E idechse nebstweiteren Untersuchungen fi ber d as d es Haifisches, Sitzb . k . Akad . Wiss .,Berlin
,p . 541 .

CHAPTER IX
THE MEDULLA OBLONG ATA AND CEREBELLUM
THE brain contains a series of pri mary sensory and m otorcenters related to the cranial nerves (see p . the correlationm echanism whi ch serves these sensori-m otor centers
,and an
extensive system of conduction pathways between the brain and
spinal cord and between the various correlation centers of thebrain itself .The brain is divided into two principal parts by a constriction
in front Of the cerebellum and pons , the isthm us (see p .
Above this level lies the cerebrum and below it the rhom bencephalon,
com prising the m edul la Ob longata or bulb and thecerebel lum . The m edulla Ob longata contains the prim arycenters concerned with m ost of the Sim pler cerebral reflexes ,especially those of the visceral , general cutaneous , auditory, andproprioceptive system s (see pp . 1 12 and The cerebel lumis a suprasegmental apparatus developed phylogenetical ly and
em bryologically out of the m ore prim itive bulbar nuc lei of thevestibular nerve, i . e.
, out Of the acoustico—lateral area of fishes
(Figs . 43 and 44,pp . 1 1 1
,1 12
,and Fig.
The O lfactory nerve (I pair) , the so- cal led Optic nerve ( I Ipair) , and the auditory nerve (VI I I pair) are special sensorynerves , whose central connections w i l l be described m ore indetai l below . The rem aining nine pairs Of cranial nerves Of thehuman body m ay be briefly summ arized as fol lows :
The oculom otor nerve (III pair) , trochlear nerve (IV pair) , and abducens(VI pair) contain the somati c m otor fibers and fibers of m uscle sense relatedto the six muscles which m ove the eyeball . The III pair also contain s v i sceral motor fibers for the ciliary ganglion ,
from which are innervatedthe muscles of the ciliary process and m s with in the eyeball, i . e .
, them usclesof accomm odation . The trigem inal nerve (V pair) supplies general sensibility to the skin and deep tissues O f the face and the m o tor innervation of the
m uscles of mastication . The facial nerve (VII pair) innervates the tas tebuds of the anterior twO-thirds of the tongue (Spec ial visceral sensory fibers) ,the sublingual and submaxillary salivary glands (general visceral efferent
143

144 INTR ODUCTION TO NEUROLOGYfibers) , and the muscles related with the hyoid bone and the superficialfacial m uscles or muscles Of facial expression
,these two groups O f muscles
belonging to the series of special visceral muscles (p . The glossepharyngeal nerve (IX pair ) supplies fibers to the taste- buds on the posteriorthird of the tongue (special visceral sensory ) , also general sensibility to thisregion,
m otor fibers for the stylopharyngeus muscle (special visceral m otor) ,and excite-
glandular fibers for the parotid salivary gland (general visceralefferent) . It also cooperates with the vagus nerve in innervating the skinabout the external auditory canal (by the auricular branch Of the vagus) .The vagus nerve (X pair ) is very com plex . In addition to the general somatic sensory fibers of the auricular branch
,whi ch have just been m en
tioned,it contains ge‘neral visceral sensory fibers from the pharynx
,lungs
,
stomach,and other abdom inal viscera, and visceral efferent fibers of several
sorts to the pharynx,esophagus , stomach
,intestines , lungs, heart, and
arteries . The peripheral and central courses Of m ost Of these functionalsystem s have been accurately determ ined
,but are far too com plex for sum
m ary here. The accessory nerve (XI pair ) contains two parts : ( 1 ) thebulbar part
,which should be regarded as nothing other than detached
filam ents of thevagus, for all of these fibers peripherally join vagus branches ,(2 ) the spinal part, which arises by num erous rootlets from the upper levelsof the spinal cord and participates in the innervation of two of the m usclesof the shoulder (the trapezius and sternocleidom astoid m uscles) . The human hypoglossus nerve (XII pair) is am odified derivative Of the first sp inalnerve of lower vertebrates . It has lost its sensory fibers and innervatesa special part of the tongue musculature.
All of the nerves of the preced ing l ist except the I , I I , I I I , andIV pairs connect w ith the m edulla Oblongata. In the d ogfish
we have seen that th is region of the brain presents Special em inences wh ich form respectively the term inal nuclei of theacoustic (and lateral line) , cutaneous
,and visceral ( including
gustatory) sensory system s (see p . 1 12 and Figs . 42 Theprimary m otor centers lie ventral ly of these sensory areas .The cranial nerves are usual ly described in our text-books
as if they were segm ental units l ike the spinal nerves (see p .
Th is was,in fact, the prim itive condition ; but in all
vertebrate animals this segmental pattern has been greatlym odified in such a way as to faci litate the d ischarge into thebrain Of all sensory fibers of like physiological type into a singlecenter . These physiological system s are, accordingly, the m ostuseful units of structure in the cranial nerves . E ach cranialnerve may contain several Of these functional system s , and no
two pairs Of cranial nerves have the sam e com position . The
com ponents of the cranial nerves , l ike those Of the spinal nerves(p . are nam ed in accordance with the same physiologicalcriteria as their end -organs (see pp . 79
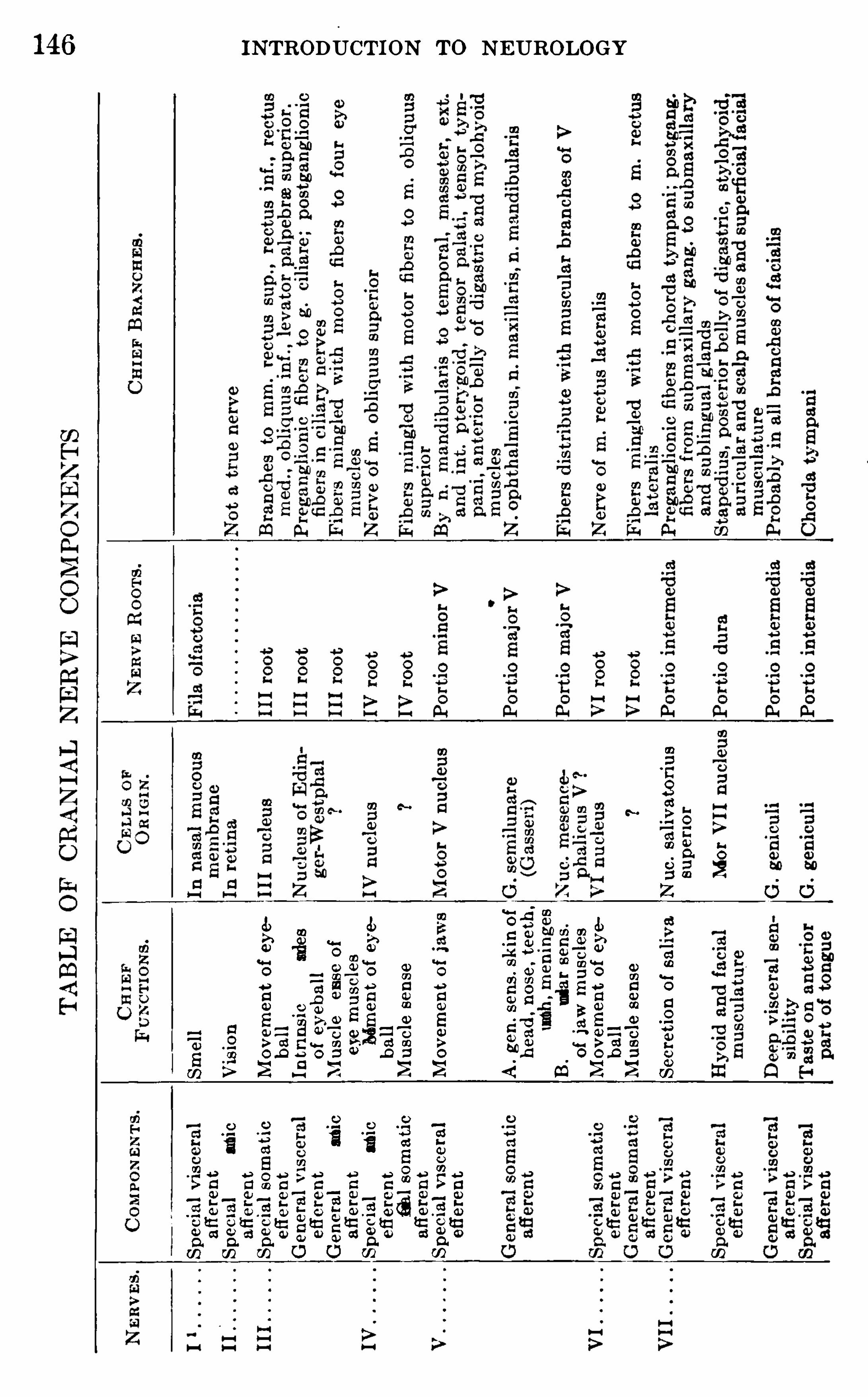
INTRODUCTION TO NEUROLOGY146
.
O.
O
80
0
2
m=
t
8a>=em
69
Z
N.
O
O
QO
EO
>0
2
emaem
zen
po
ke
M
o
a
no
n-9
52
9
3
«o
356
zapo
ho
mo
81
59
8o
u
nce-g
nu
zen
ro
am
o
a
ao
Eo
>0
2
n
ew
;Sm
n
ap-o
we
13
8m
;
36
o
Bogo
ta
o
fi
aw
12
250
gen
o
me
£
38
8
36
o
Sona
ta
2
588g
en
o
me
m
ggen
o
me
om
u
wSOm
36
o
e
man
a
te
3588
1:
0
o
Sona
ta
a
re
36
o
em
aom
fizoo
m
omawm
=
S
the
we
Ba
.“
«0
.
mcow
516
25“.
m
mOw
aE
£25
5
Ho
me»
6
q.
fiao
n
O
o
Exm.
mcom.
dew
é.

THE ME DULLA OBLONGATA AND CEREBE LLUM
00
000
80
8800
0
00
000
0000
00
0
0
00
00
80
000
0
000
0
000000
0
0
0
m00m0
00
00
00
820
800
00
0
80:w00w0ua~
00
00
000
9
0
0
800
0
2
0
000
200
§E
00
0
00
0
8
0
00
00000
00
80
m
00
00
00
50
0w0>
0
8000
0
8000
0
000
00
00
0
8000
0
00
0
on.
0
80
00
800
0-0
00
000
000
0w0>
0
00
00
05
000
00
000
0
0
0
000
0
80
.
80>
0
00
m0
80
m
0
00
00
80
0
bflfiou
m
08000
0
000
0
007
00
.
0
00
3
000000000
£0
000
80
:
000
m
00
w0
50~
000
0
00.
000m
0
800
00
00
00
0
0
8000
>
0000
000000
£0
5000
m
80
m
60>
00
0
0—00
100.
0
0
800
.
mg
0
0
80
m
000
000
0000
00
0
0000
0
8000
000
0
00
00
00
500
000
8000
000
00000
.
0
00w0
b00
m
Nu
0
00
w0>u0000
§00
0
0
80
m
000
0
00
0
00
00
00
00
.
0
800
300000
0
”
0000
00
0
00
000
0
0
800
830
0
800000000
m
00
0—00
00
0
0>.
0
Z
0000
0000»
0
0>.
0
Z
000
_0000
Q
0
80
Q
0
00.
0
000
000»
0
000
0
000
000»
:N
H
0
00
8
m0~
Hum0
00m
Sn
0
0
5
N
500
00
m
um0
3000
N
bow00
m
N0
00
0
2
N0
00
0
2
0
2.
0
00
m
NH
.
0
00
m
0
000
NH
.
m00
m
0
00.
um
2
0
000
NH
0
00
0
2
0
100
20
00
0
:
00
m
0
10
0
50
0000,
500
m
30
0
50000
000
580000
000
0
000
0
000
000
0
0
0
000
3000
00
0
8
00
0
8
00
0
00
5
0
00000
0
0
00
0—0000
0
.
m
00000000
.0
0
00
0
8000
0
1
0
um
00
0
0
0
80
m
0
00
um
0.
0
000
0
000000
0
0
0
50
0—0
80
800
000
0.
0
0
0
00
80.
00
00
0
0
9
00
80000
000
£00
000
£0
5000
800
0000.
0.0
0
00
500
000
00
;
00
00
00
00
0
0
00
00
0
8000
0
2
w
0
000
?
000
0
00.0
0
0
000—0
000
00
00
0
800
E.
S00
D
0
00
00
000
0
0.
0
000
0
000000
0
0
0
00000
0
0
00
00
0
3
800000
0
0.
0
0
00
00
0
00
00
00
0
0
8
0
00000
000
0
00
Mm
800
000
0
0.
0
L0000
0.
0
000
.
00
0
00
5000
0
0
00
89
02
0
000
380.
0
0
Z
N0
0
.
0
0
8_00..0
Q
0
00
0000
.
0
0
000
580.
0
0
2
0000
0
50
800
.
0
000
0
0
0,
0000
0
0
00
000
20
0
800
0000
0
00
0
0
800
00
000
5
0
0
000
0»
00
000
0,
0
0
000
0
00—
0
00
0w
.
00
0000
000
0
0»
00
025
00
>0
00
000
0
00
800
0
000
?00
0
25
m
>0
0
>~
0
00
Qw
a
sss
s
00
80
0
30
800
00
000
5

148 INTRODU CTION TO N EUROLOGY
4 . Special somati c efi'
erent nerves, supplying two groups of h igh ly spec ialized somat ic muscles, namely, the externa l eye m uscles and a part of thetongue muscles . They ar1se from a ventro-med ia l seri es of motor nuclei andare represented i n the III, IV ,
VI, and XII pa irs .
5 . General visceral afferent nerves , innervating visceral m ucous surfacesw i thout hi gh ly d ifferent iated sense organs . They d istribu te through thesympathetic nervous system and are represented in the VII, IX, and X pairsand perhaps 1n some others .6 . Specia l v iscera l afi'
erent nerves , for the innervation of spec ialized senseorgans serv ing the senses of taste and sme l l . The gustatory fibers arerepresented in the VII, IX,
and X pa irs . The o l factory nerve ( I pa ir ) ispr
oibably a more h igh ly different iated member of th is group (see pp. 9 1
an
7 . Genera l v isceral efferent nerves , for unstriped invo luntary visceralmuscles
,heart mus cle
, glands , etc .
,d istribut ing through the sym pathetic
nervous system . These fibers (preganglionic fibers of Langely, p . 229 ) arepresent in the III
,VII IX
,X , and XI pa1rs .
8 . Special visceral efiereh t nerves supplying hi ghly speciali zed striatedmuscles of a d ifierent origin (both embryo logi ca lly and phylogenetica lly)from the striated trunk muscles . These m uscles are connected w i th theviscera l or facia l skeleton of the head and are derived from the mus clesof fishes . These nerves in the ad ul t body resemb le those of t e som ati cmotor system ,
save that they arise from a d ifferent series of motor nuclei inthe brain (the ventro—latera l motor co lumn) . They have no connectionwi ththe sym patheti c nervous system and are represented in the V, VII , IX, X,
and X1 pairs .In the preced ing Tab le of Nerve Components (pages 146 , 147) the sev
era l cranial nerves are ana lyzed and compared w ith a typi cal spinal nerve.
The var ious functi onal system s of the head tend to be concentrated in one or a few c ran i a l nerves for ease of central correlation
,and even in case a given system i s represented in several
nerves, the fibers of th i s system m ay converge within the brainto connect with a compact center . Thi s i s wel l i l lustrated by thegustatory and acoustico—lateral system s of the crani al nerves ofthe fish , Meni d ia
,as shown in Fig. 65 . Here the gustatory sys
tem ( indi cated by c ross- hatching) i s present in the VII, IX, and
X c rani al nerves,and al l of these fibers, together with other
vi sceral fibers , converge within the brain to enter thevi sceral sensory area in the vagal lobe (lob.X Sim i lar ly
,the lateral l ine
components of the V I I and X nerves and the VI I I (printed insol id black) converge to enter the acoustico—lateral area in thetubercu lum acusticum The general cutaneous fibersenter by the V and X nerves
,and al l of these fibers enter the
spinal V tract
Ino
the paragraphs wh ich fo llow the chief centra l connect ions (termina lnuclei of the sensory systems and nuclei of origin of the motor systems , see
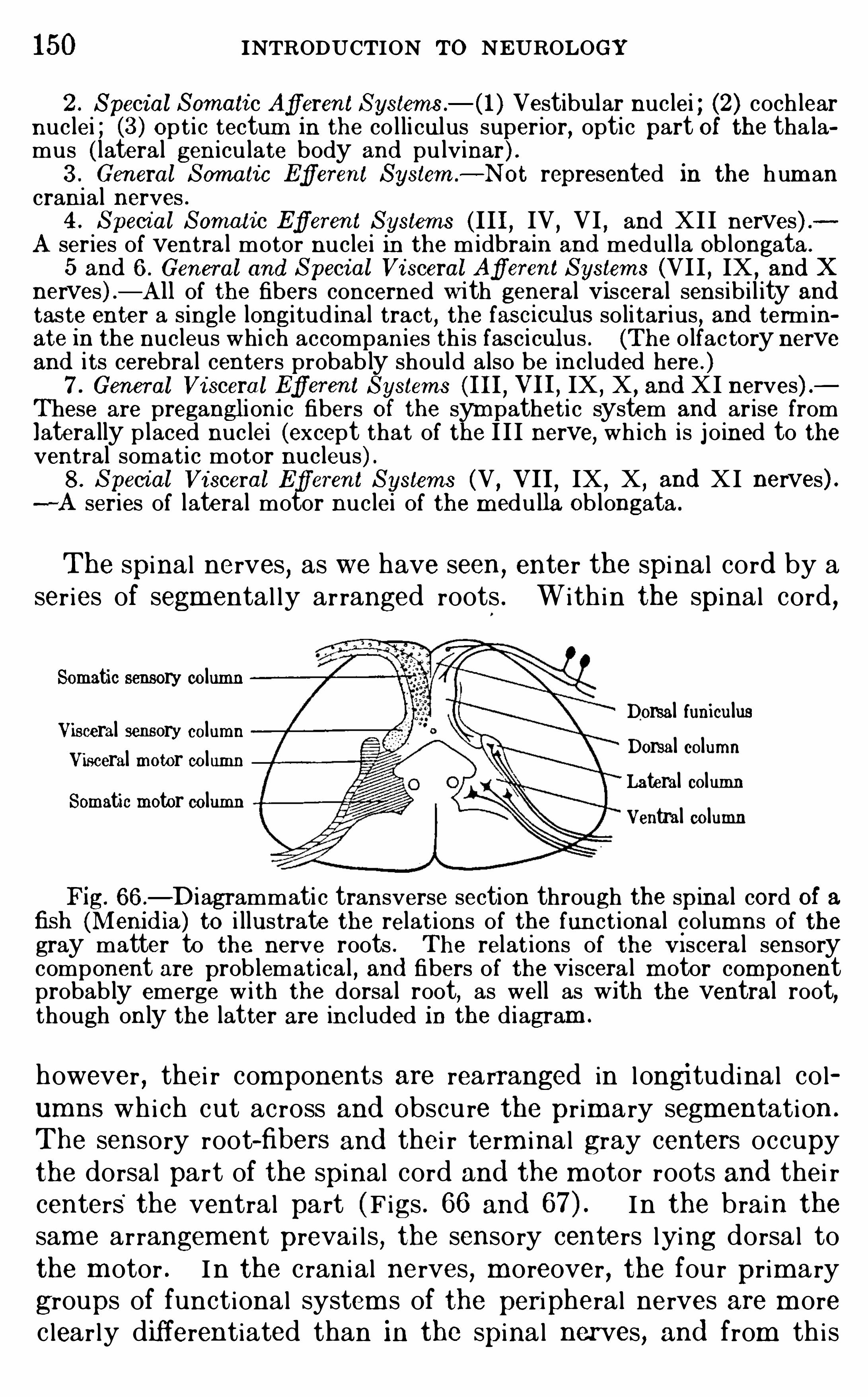
150 INTRODUCTION TO NEUROLOGY2 . Special Somati c AfferentSystems . Vestibular nuclei ; (2) coch lear
nuclei ; (3) Op t i c tectum in the co l li culus superior, optic part Of the tha lamus ( lateral gen iculate body and pulvinar ) .3 . General Somatic E’
flerent System—Not represented in the hum ancranial nerves .4 . Special Somati c Efierent Systems (III, IV , VI , and XII nerves) .
A series of ventra l motor nuclei in the midbrain and medul la Ob longata .
5 and 6 . General and Special Vi sceral AflerentSystem s (VI I , IX ,and X
nerves) —A ll of the fibers concerned w ith general Visceral sens ib i l ity andtaste enter a s ingle longitud inal tract , the fasciculus so li tarius, and term inate in the nucleus whi ch accompanies th is fasciculus . (The o lfac tory nerveand i ts cerebra l centers probab ly should also be included here . )7 . General Vi sceral Efi
'
erent Systems (III, VII, IX , X , and XI nerves) .These are pregangl ion i c fibers Of the sym patheti c system and arise fromlatera lly placed nuclei (except that of the I I I nerve, wh i ch is j o ined to theventral somat i c motor nucleus) .8 . Speci al Vi sceral Efierent Systems (V ,
VII,IX , X , and XI nerves) .—A series of lateral motor nuclei of the m edulla Ob longata .
The spinal nerves, as we have seen,enter the spi nal cord by a
ser ies of segmental ly arranged roots . Within the spinal cord ,
Somatic sensory columnDorsal funiculus
Visceral sensory columnDorsal column
Visceral motor columnLateral column
Somatic motor columnVentral column
Fig. 66 .—D iagrammati c transverse section through the spinal cord of a
fish (Meni d ia) to i llustrate the relat ions of the funct ional co lumns of thegray matter to the nerve roots . The relat ions of the V i scera l sensorycomponent are prob lemat ica l , and fibers of the V iscera l m otor componentprobab ly emerge w i th the dorsa l root, as well as Wi th the ventral root,though on ly the latter are included in the d iagram .
however , thei r components are rearranged in longitudinal columns whi ch cut across and Obscu re the prim ary segmentation .
The sensory root-fibers and thei r term inal gray centers occupythe dorsal part of the spinal cord and the m otor roots and thei rcenters the ventral part (Figs . 66 and In the brain thesame arrangem ent prevai l s , the sensory centers lying dorsal tothe m otor . I n the c rani al nerves , m oreover , the fou r prim ary
groups of functional system s of the peripheral nerves are m orec learly differentiated than in the spinal nerves, and from th i s

THE MEDULLA OBLONGATA AN D CERE BE LLUM 15 1
i t fo l lows that thei r prim ary centers are correspondingly h ighlydeveloped and di stinct . The m edu l la Oblongata , in fact, i sd ivided into fou r longi tudinal columns related respectively tothe great prim ary groups of functi onal system s . In fishes,where the am ount of correlat ion ti ssue i s less than in man
,these
four primary columns appear as wel l- d efined ri dges in the wal lof the fourth ventri c le.
An enlarged view Of the medu l l a Oblongata of the stu rgeon,
which i s very sim i lar to that of the d ogfish , i s seen in Fig. 68,
which also i l lustrates the arrangem ents Of the prim ary sensoryand m otor centers in cross- section at several different levels .
Somatic sensory columnVisceral sensory columnVisceral motor columnSomatic motor column
Fig. 67 .—Diagramm at ic transverse section through the hum an spinal
cord . Compare F igs . 56 to 59 and note the relatively greater s ize of thedorsa l gray col umns and dorsa l funi culi in m an than in the fish (Fig .
Th is is correlated with the greater importance in m an Of the ascend ingconnections between the cord and the brain (see p.
F igu re 69 shows a c ross- section through the m edu l la Oblongatain the regi on Of the vagus nerve in another fish
,the sea- robin .
In al l of these cases the fou r pr inc ipal functional system s (seepp. 76 and 79—94) are arranged in l ongi tudinal colum ns fromthe dorsal to the ventral su rface in the order : som ati c sensory ,visceral sensory
,vi sceral m otor
,and som ati c m otor centers , as
indi cated diagramm ati cal ly on the left side of Fig. 69 . The
arrangement of the per ipheral nerve-fibers of these system s i sindi cated on the right side. F igu re 70 illustrates a c ross- sectionthrough the corresponding regi on of the m edu l la Oblongata in an
ear ly hum an embryo,where the same general arrangement of
the sensori-m otor centers i s evident .

152 INTRODUCTION TO NEUROLOGY
N .
Lobus liners lateralis
Fasc. long. med
Lobus visceralis
Fig. 68 .—The medul la Ob longata and cerebellum of the lake sturgeon
(Acipenser rubicund us) to show the longitud ina l co lumns wh i ch have beend ifferent iated in correlation w ith the periphera l functional systems . Compare Figs . 43 and 44 and note that the “
L obus lincaz latera lis ” and Tu
berculum acusticum O f th is figure together correspond to the acousticolateral area ” of the d ogfish . A is a dorsal v iew w ith the m embranous roofOf the fourth ventricle removed to show the longitud ina l co lumns w i th inthe ventricle . B
,C , and D are sketches of cross- sec tions at the levels
ind icated in wh ich the four funct ional columns are d iagrammat i cal ly shaded ,the somat ic motor by wh i te c ircles
,the visceral motor by wh i te rectangles,
the v iscera l sensory by ob l ique cross-hatching,and the somatic sensory by
t i ca l cross-hatch ing . The Roman numera ls refer to the crania l nerves .
(From Johnston ’s N ervous System of Vertebrates . )

154 INTRODUCTION TO NE UROLOGY
general vi sceral m otor nuc lei by sm al l dots, the spec ia l vi sceralm otor nuc lei by large dots , the visceral sensory nuc lei by double
Nuc. IIImed .
Nue. IV
Velum med ul Nuc. moseno.Vlarc superius
Nuo. mot. V
Nuc. sensor. VNuc. VI
Nuc. mot VII
Fig. 7 1 .—Dorsal view of the human midbrain and m edulla Ob longata
after the removal of the cerebel lum and the roof of the fourth ventricle,w i th the pos i tions of the cran ial nerve nuclei projected upon the surface .
The motor nuc lei are ind icated on the left s ide and the sensory nuclei on theright . The somati c motor nuc lei are ind i cated by circles , the general v iscera l eflerent nuclei by small dots
,and the special v isceral efferent nuclei by
large dots . The general somat ic sensory area is ind icated by horizonta l l ines ,the v isceral sensory area by doub le cross-hatch ing
, and the specia l somati csensory area by open stipple . (Compare Figs . 77 , 86 , and
n .1 V, N ervus troch learis ; nuc .com .Caj al, the commissura l nucleus ofR am én y Caj al ; nuc .III E -W.
, the sma l l - celled v iscera l motor nucleus of theI I I nerve, or nuc leus of E d inger-W’
estphal ; nc I1 lat.,lateral nucleus of
I I I nerve ; nuc . III m eal,med ia l nuc leus of III nerve ; nnc . IV,
nucleus of IVnerve ; nuc .mesenc.V,
mesencephal i c nucleus of V nerve ; nuc .mot.V, motornucleus of V nerve ; nuc .mot.VIl , ch ief motor nucleus of VII nerve ; nucs al.inf nucleus sa l ivatorius infer ior ; nucleus sal ivator ius superior ;nuc .sensor.V,
ch ief sensory nucleus of V nerve ; nuc .VJ , nucleus of VI nerve ;nuc . XII nucleus of XII nerve ; n .V,
nervus trigeminus .
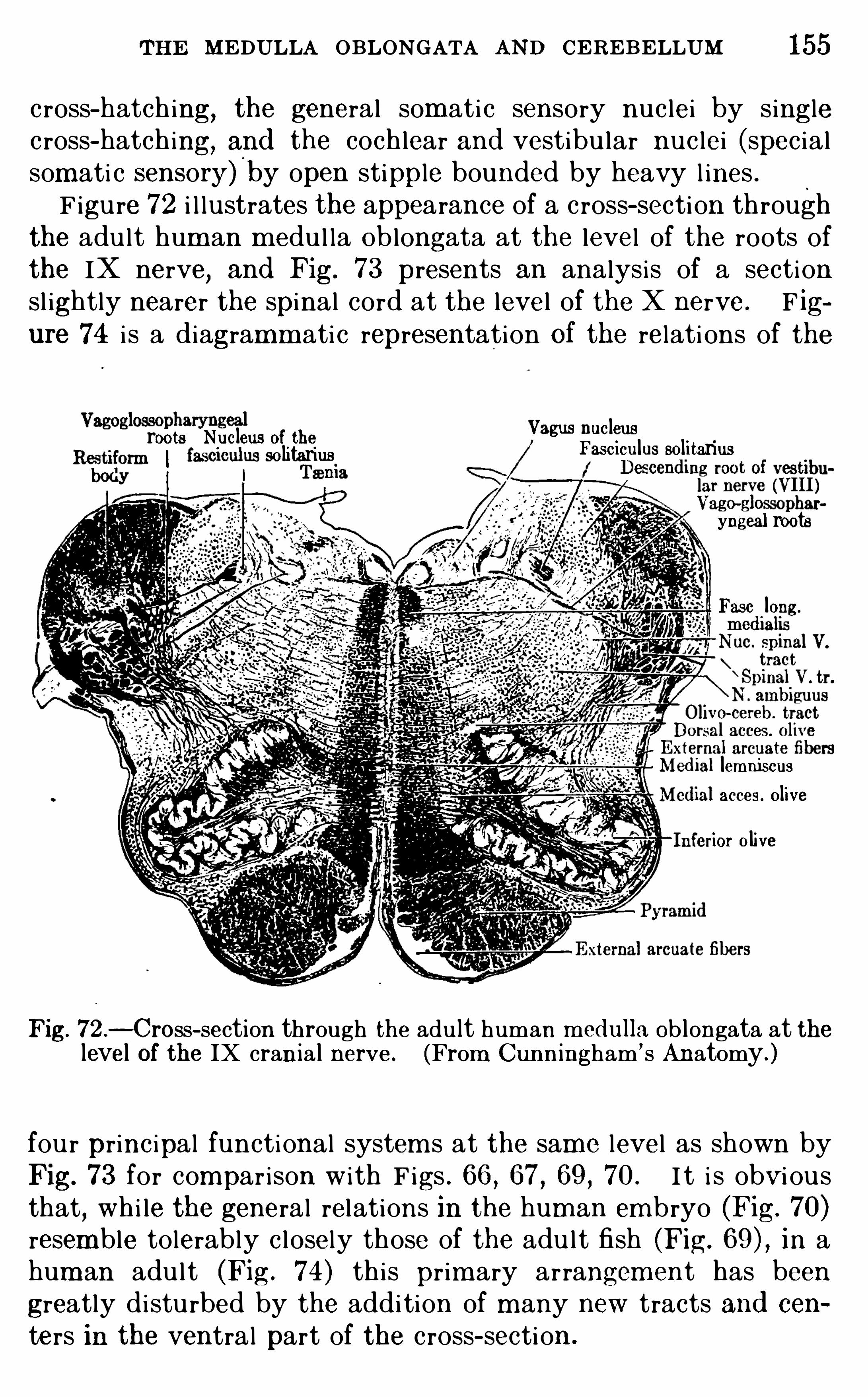
THE MEDULLA OBLONGATA AND CEREBELLUM 155
c ross- hatch ing, the general som ati c sensory nuc lei by singlec ross-hatching, and the cochlear and vestibu lar nuc lei (spec ia lsom ati c sensory)
'
by open stipple bounded by heavy l ines .F igure 72 i l lustrates the appearance of a c ross- section through
the adu l t hum an m edu l la Oblongata at the level of the roots ofthe IX nerve
,and Fig. 73 presents an analysis of a section
sl ightly nearer the spinal cord at the level of the X nerve . F igure 74 i s a diagramm ati c representation Of the relati ons of the
Vagus nucleusroots Nucleus of theFasc iculus sol i tarius
soli tarius
yngeal roots
Fasc long.
lemniscusacces. oliveior olive
Fig. 72 .—Cross- section through the adult human m ed ulla Ob longata at thelevel of the IX crania l nerve . (From Cunningham
’ s Anatomy. )
four princ ipal functional system s at the same level as shown byFig. 73 for compar ison with F igs . 66 , 67 , 69 , 70 . I t i s obvi ousthat
,whi le the general relations in the hum an em bryo (Fig. 70)
resemble tolerably c l osely those of the adu l t fish (Fig. in ahum an adu l t (Fig. 74) th is pr im ary arrangem ent has been
greatly di sturbed by the addi tion of m any new tracts and cen
ters in the ventral part of the cross- section .
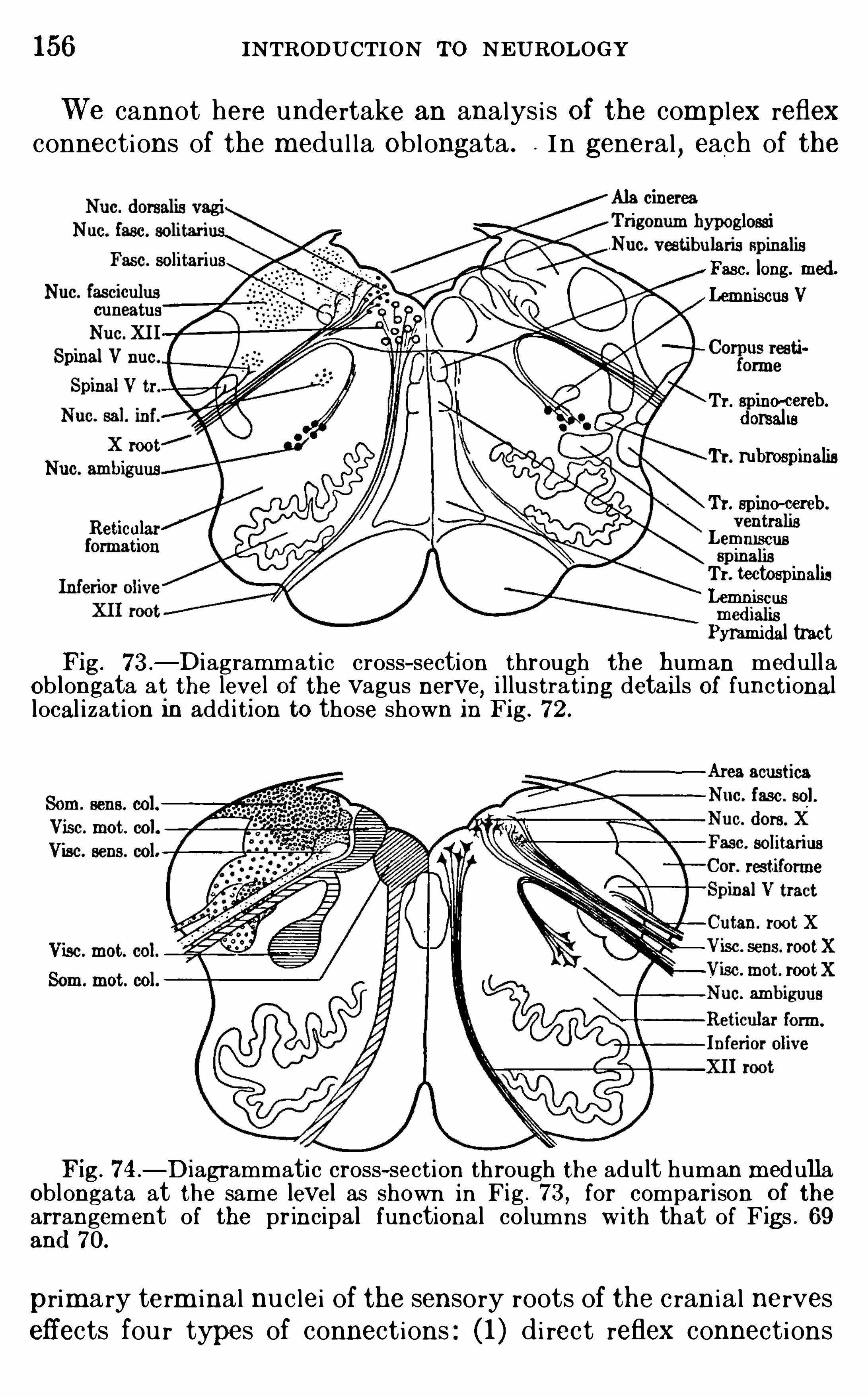
156 INTRODUCTION To NEUROLOGY
We cannot here undertake an analysi s Of the complex reflexconnections of the medu l la Oblongata . I n general , each of the
Fasc. long. med .
Lemniscus V
Tr. spino-cereb.
d orsal isTr. rubrospinalisTr. spine-cereb.
Lemmscus
Tr. tectospinalisLemniscus
Pyrami d al tractFig. 73 .
—Diagrammat ic cross- section through the human medul laOb longata at the level Of the vagus nerve, i llustrating detai ls Of functionallocal izat ion In add i t i on to those shown in Fig. 72 .
Fig. 74 .
—D iagrammatic cross-section through the adult human m edullaOblongata at the same level as shown in Fig . 73 , for comparison
.
of thearrangement of the principal functiona l co lumns w i th that of F igs . 69and 70.
primary term inal nuc lei of the sensory roots of the c ranial nerveseffects fou r types of connections : ( 1 ) di rect reflex connections

158 INTRODUCTION To NEUROLOGY
the centers of correlati on . As has al ready been indi cated, all ofthese centers are interconnected by correlati on neu rons sim i larto those of the spinal cord (F igs . 60, These neu rons arel oosely arranged in the spaces between the sensory and m otorgroups of nuc lei , thi s ti ssue bei ng termed the reti cular formation
(thi s regi on i s also called the tegm entum,see pp. 65, 127 and
Figs . 69 , But the chief centers of correlat ion of the brainstem are found in spec i al ly enlarged nuc lei of the m i dbrain andthalam us
,som e of whi ch are m entioned in the next chapter .
I n its m ore ventral parts the m edu l la Oblongata contains anumber of large correlat ion centers and important conducti on
pathways between rem ote parts of the brain . Of the form er,the
largest are the infer i or ol ives (F igs . 72 , 73, deeply bu riedm asses of gray m atter arranged in the form of a hol low shel l ofcomplex shape on each side of the m edian plane. The O l ivesreceive fibers from the thalam us and spinal cord and dischargeinto the cerebel lum (olive- cerebel lar fibers of Fig. Theirfunctions are unknown .
The cerebellum has al ready been referred to as a great suprafsegm ental mechani sm of unconsc i ous m otor coordination . I ti s connected with the under lying brain stem by three pai rs ofstalks or pedunc les , two of whi ch j oin the m edu l la Oblongata andone the m idbrain . The infer i or pedunc le (restiform body)connects with the dorsal m argin of the m edu l la Oblongata andcarr ies fibers into the cerebel lum from the spinal cord and oh
longata. The m i ddle pedunc le (brachium ponti s) connectsw i th the pons and m ost of i ts fibers convey impu l ses from the
nuc lei of the pons to the cerebellum . The superi or pedunc le(brach ium conj unctivum ) connects with the cerebral pedunc lein the floor of the m i dbrain and contains ch iefly fibers whichdescend from the cerebel lum ,
c ross the m i dplane under theaqueduct of Sylvius, and term inate i n or near the red nuc leus(Fig. 75
,nuc leus ruber) . The internal structu re and connec
ti ons of the cerebel lum wi l l be further considered on page 186 .
Summary.
—The rhom bencephalon inc ludes the m edu l laOblongata and cerebel lum ,
that i s,al l parts of the brain bel ow
the i sthm us . All of the cran ia l nerves except the fi rst fou r pai rsconnect with the medu l la Oblongata . An analysi s of the functional components of the cranial nerves shows that they can

0
THE MEDUL LA OBLONGATA AND CEREBELLUM 159
best be understood by consider ing each functional system offibers as a uni t and studying the connections Of each componentseparately . These connecti ons are summ ari zed in a table on pp.
146,147 . The m edu l la Oblongata of lower vertebrates and of
the hum an em bryo i s seen to be composed chiefly of the prim arycenters related to these functi onal components of the per ipheralnerves
,arranged in l ongi tudinal colum ns in the order from dorsal
to ventral surfaces on each si de of som ati c sensory,vi sceral
sensory,visceral m otor
,som ati c m otor centers . The sam e
arrangement appears in the adu lt human Oblongata , thoughsom ewhat di storted by the presence of large m asses of correlati on ti ssue and Of large conducti on tracts whi ch are not presentin the l ower vertebrates . The sensory centers of the Oblongataare connected local ly with the adj acent m otor centers and alsoby longer tracts with the spinal cord, cerebel lum ,
and thalam us .The latter fibers constitute the bu lbar lemni scus, of whi ch severalfunctional components can be distingu i shed, them ost importantbeing the trigem inal lemni scus for general cutaneous sensibi l i tyand the lateral or acousti c lemni scus for auditory sensibi l ity.
The cerebel lum i s a propr ioceptive center developed out of thevestibu lar area of the m edulla Oblongata .
LITERATU REThe detai ls of the structure and fun ct ions of the parts mentioned in thi s
and the fo l lowing chap ters w il l be found fully presented in the standard textbooks of hum an anatomy and phys iology and m the med i ca l text-books ofneuro logy, and al l of thi s li terature up to the year 1899 is summ ar i zed In
Barker ’s Nervous System and Its Const ituent N eurones . See also W .
von Bechterew,Die Funktionen d er N ervencentra Jena, 1 908 to 19 1 1 ,
3 vo ls . For d iscuss ions Of comparative neuro logy and the evolut ion Of the
nervous system ,reference may be made to art icles in the neuro logi ca l
j ourna ls,especia lly the J ournal of Comparative Neurology; see a lso the
Bib liograph ies on pp. 36 , 124 , 193 , and 223 , and the fo llow ing works :HE RRICK , C . JUDSON . 1899 . The Cranial and First Spina l N erves of
Menid ia : A Contribut ion Upon the Nerve Components of the Bony Fishes,Jour . Comp . N euro logy, vo l . ix. p, p . 1 53—455 .
19 13 . Brain Anatomy,Wood ’
5 Reference Handbook of the Med ica lSciences
,3d cd .
,vo l . i i
,pp . 274—342 .
19 14 . Cranial N erves,ib id .
,vo l . i ii
,pp . 32 1 - 339
toii N STON , J . B . 1906 . The N ervous System of Vertebrates , Phi lad el
p a1909 . The Centra l N ervous System O f Vertebrates , Spengel
’sE rgebnisse und Fortschr i tte d er Zoologie, Bd . 2 Heft 2
,Jena .
E DIN GER,L . 1 908 . Vor lesungen tiber d en Bau d er nervesen Zentral
organe, 7th Auflage, Band 2 , Verg lei chende Anatom ic d es Geh irns, Leipz ig.
19 1 1 . Idem , 8th Auflage, Band 1 .

CHAPTER X
THE CEREBRUM
THE cerebrum inc ludes al l of the brain ly ing in front of thei sthm us
,that i s
,the m idbrain (m esencephalon) , betweenbrain
(diencephalon) , and cerebral hem i spheres (telencephalon) , thetwo last compr i sing the forebrain (prosencephalon) . I t con
tains the pr im ary sensory centers of the olfactory nerves ( I
pai r) , the sensory correlation centers Of sm el l and sight, the
primary motor and sensory centers of the ocu l om otor and trochlear nerves (II I and I V pai rs) for m ovem ents of the eyes, and al lof the most important higher correlation centers of the brain .
These higher correlation centers m ake up by far the larger partof i ts substance in the hum an brain,
though in fishes the converserelati on prevai l s , with the prim ary sensor i-m otor centers and thesimpler correlati on m echani sm s m ak ing up the larger part (seeF igs . 43 , 44 , pp. 1 1 1
,
The mesencephalon (m i dbrain) i s that part of the brain i nwhi ch the early embryon i c neu ral tube (F igs . 46—5 1 , pp. 1 16
1 19 ) has been least m odified in the adu l t . The ventral partof the m i dbrain,
i . e., the part lying ventral ly of the ventr i c le,
whi ch i s here term ed the aqueduct of Sylvius , i s cal led the cerebral pedunc le ; the dorsal part i s the corpora quadrigem ina , theupper pai r of these fou r em inences being the super i or col l icu l i ,and the l ower pai r the inferior col l i cu l i (see Fig. 7 1 , p.
The corpora quadrigem ina contain important correlationcenters, the superior col l i cu lus ch iefly vi sual (p. 209 ) and theinferi or col l i cu lus ch iefly audi tory (p. The cerebralpedunc le, as the nam e impl ies , contains the great ascending anddescending fiber tracts between the forebrain above and the
m edu l la Oblongata, cerebel lum , and spinal cord bel ow . The
arrangement of som e of these tracts can be seen i n Fig. 75 . The
cerebral pedunc le also contains the nuc lei of o rigin for the m otorfibers of the I I I and I V pai rs of cranial nerves and several m asses
160

162 INTRODUCTION To NEUROLOGY
whol ly to various types Of correlati on. I t has three m ain divisions
,the thalam us , the epi thalam us , and the hypothalam us, of
whi ch the two last are dom inated by the O l factory apparatus
(see p.
The epi thalamus consi sts of the mem branous chor ioid plexuswhi ch form s the roof of the th i rd ventri c le (Fig. 7 the pinealbody or epiphysis (Fig. the habenu la (m arked trigonum
Genu of corpus cal losumCorpus cal losum (cut)
eepti pel lucid iSeptum pel luci d umCaud ate nucleusFornixForamen interventriculareAnterior commissureAnt. tubercle of thalamusMassa intermed ia
Non-ventricular part 0t a amus
ulvinar
incal bod yBrachium conjunctivum
LingulaMed ul la Oblongata
Fig. 76 .—A d issection Of the brain from above to expose the tha lamus andcorpus striatum . (From Cunn ingham ’s Anatomy) .
habenu laeon Fig. and the stria m edu l lari s , a fiber tract whi chconnects the olfactory centers of the cerebral hem i spheres withthe habenu la (Figs . 78, The habenu la i s a center for the correlation of olfactory sensory impu l ses with the various som ati csensory centers of the dorsal part of the thalam us . The pinealbody of som e l ower vertebrates i s a sense organ ,
apparentlyvisual in function and known as the parietal eye (p. in

THE CE RE BRUM 163
man its pr im ary sensory function i s lost and i t is said to produce an important internal secretion whose physi ologi cal valuei s sti l l Obscure.
The hypothalamus inc ludes the tuber c inereum and mamm il
lary bodies (see F igs . 53, 78, and these structures beingO l factory centers, and the hypophysi s or pi tu i tary body (whi chhas been rem oved from the spec im en shown in Fig. 53
,i ts point
of attachment being the infundibu lum ) . The hypophysi s i s a
glandu lar organ whi ch produces an internal secret ion of greatimportance i n m aintaining the proper balance of the m etabol i cactivities of the body. The hypothalam us i s an important center for the correlation of olfactory impu lses wi th vari ous vi sceralfunctions , inc luding probably the sense of taste.
The thalamus i s in the hum an brain chiefly a sort of vestibu lethrough whi ch the system s of som ati c sensory nervous impulsesreach the cerebral cortex. There are, however , two parts of thethalam us whi ch shou l d be c lear ly distingu i shed . The ventral
part contains ch iefly m otor coordination centers . It i s feeblydevel oped in the hum an brain, where i t i s term ed the subthalamus (not to be confused , as is often done, with the hypothalamus
,see F igs . 78, 79 , and The dorsal part of the
’
thalam us,
in i ts tu rn, contains two di stinct types Of sensory correlationcenters : (1 ) pr im i tive sensory reflex centers , chiefly in the m edial
group of thalam i c nuc lei ; (2 ) the m ore lateral nuc lei whi ch formthe cort ical vestibu le to whi ch reference was m ade above.
These lateral nuc lei are sometimes cal led the new thalam us(neothalam us) in distinction from al l of the other thalam i cnuc lei whi ch form the old thalam us (palaaothalamus) .The centers whi ch compri se the new thalam us m ake up by far
the larger part of the thalamus in the human brain and inc ludethe fol lowing nuc lei : the lateral
,ventral , and posteri or nuc lei
(for general cutaneous and deep sensibi l i ty) receiving the
spinal , trigem inal , and m edial lemni sc i ; the lateral gen i cu latebody and pu lvinar (vi sual sensib i li ty) receiving the opti c tracts ;the medial geni cu late body (audi tory sensibi l i ty) receiving thelateral or acousti c lemn i scus . The lateral and medial gen ienlate bodies compr i se the m etathalam us Of the B . N . A . (see p.
121 and Fig. 50, p. whi ch in th is work are desc ribed as
part of the thalam us .
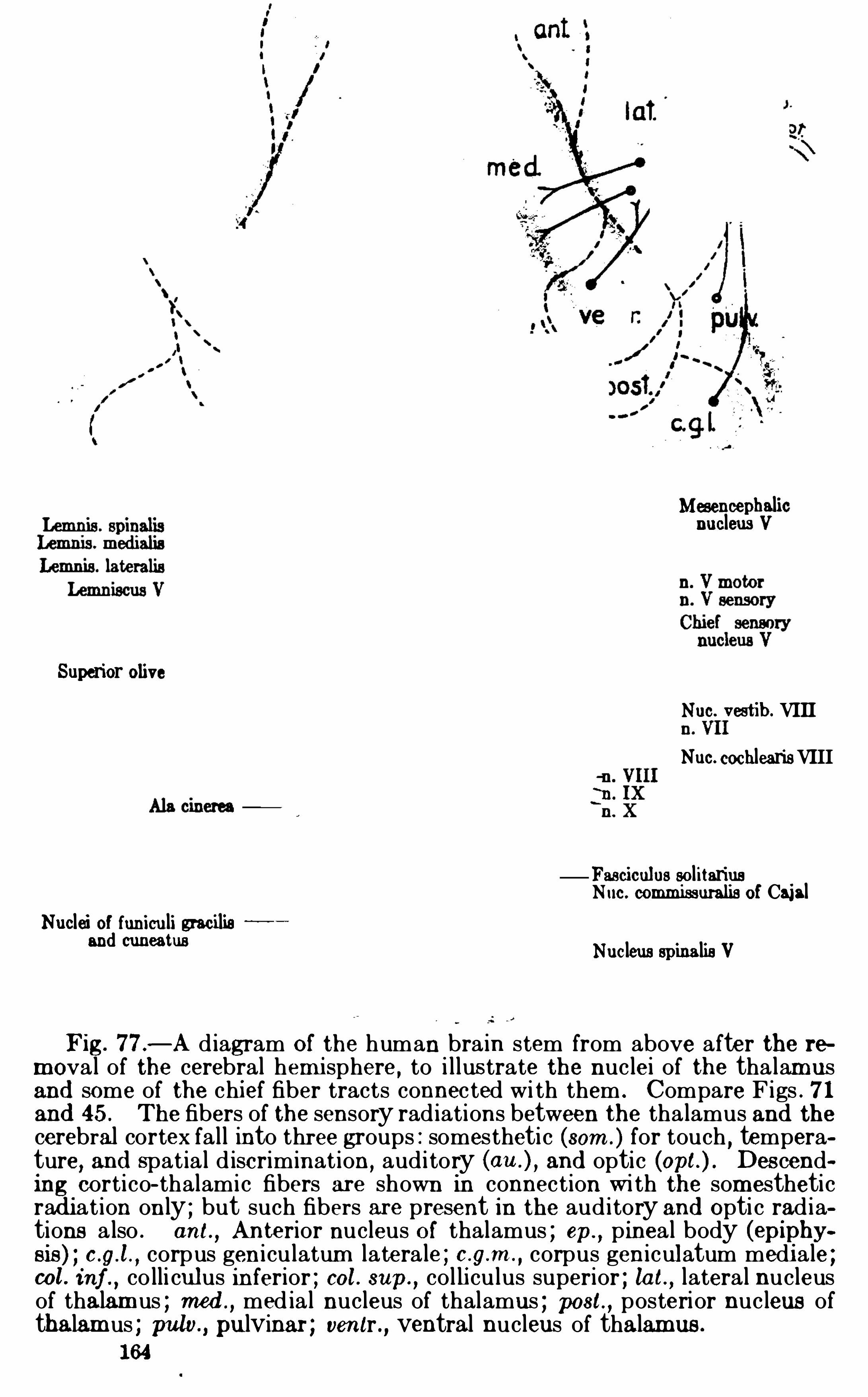
Mesencephalicnucleus V
Superior oliveNuc. vestib. VIIINuc. cochlearisVIII
Fasciculus sol itariusNuc. commissuralis of Cajal
Nuclei of funiculi w oi lieand cuneatus
Nucleus spinalis V
Fig. 77 .—A d iagram Of the human bra in stem from above after the t e
mova l of the cerebral hemi sphere , to i l lus trate the nuclei of the tha lam usand some of the chief fiber tracts connec ted wi th them . Compare Figs . 7 1and 45 . The fibers of the sensory rad iations between the tha lamus and thecerebral cortex fal l into three groups : somesthetic (som . ) for touch , temperature
,and spat ia l d iscriminat ion, aud i tory and optic Descend
ing corti co- thalamic fibers are shown in connection wi th the somestheticrad iat ion on ly ; but such fibers are present in the aud itory and Opti c rad iations a lso . ant.
, Anterior nucleus O f thalamus ; ep.
,pineal body (epiphy
sis ) ; corpus geni culatum latera le ; corpus geniculatum medi ale ;col . inf .
,co lli culus inferior ; col . sup.
,co l li culus superior ; lat. , lateral nucleus
of thalam us ; med ,med ia l nucleus of thalamus ; post , posterior nucleus of
thalam us ; puta , pulvinar ; ventr ., ventral nucleus of thalamus .
1 64
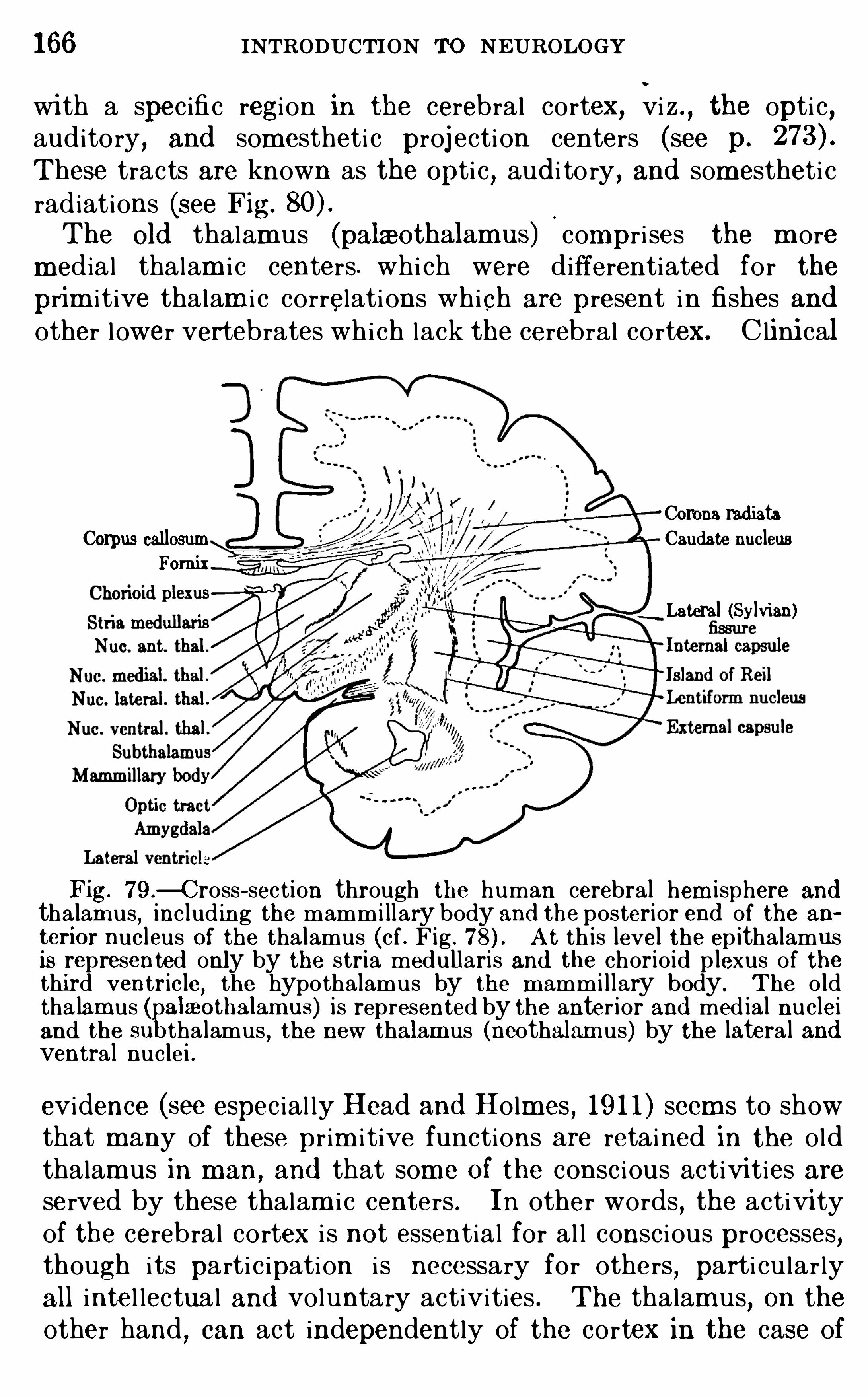
166 INTRODUCTION TO NEUROLOGY
wi th a spec ifi c regi on in the cerebral cortex, vi z . , the opti c ,auditory
,and somestheti c proj ecti on centers (see p.
These tracts are known as the Opti c , aud i to ry, and som estheti cradiations (see Fig.
0
The Old thalam us (palmothalamus) compri ses the m ore
medial thalam i c centers. whi ch were differentiated for the
prim i t ive thalam i c correlati ons which are present in fishes andother lower vertebrates whi ch lack the cerebral cortex. C lini cal
Corona rad iataCorpus callosum Caud ate nucleus
Fom ix
Lateral (Sylvian)
Fig. 79 .—C ross—section through the human cerebral hem isphere and
thalam us,includ ing the mammillary body and the posterior end of the an
terior nucleus Of the thalamus (cf . Fig . A t th is level the epithalamusis represented only by the stria medullaris and the chorio id plexus of thethi rd ventricle
, the hypothalamus by the m ammillary body. The o ldtha lamus ( alaaothalam us) is represented by the anter ior and med ial nucleiand the su tha lamus
, the new thalamus (neothalam us) by the latera l andventral nuclei .
evidence (see espec ial ly Head and Holm es,19 1 1 ) seem s to show
that m any of these pr im i t ive functions are retained in the oldthalam us in man,
and that som e Of the consc i ous activi ties areserved by these thalam i c centers . In other words
,the activi ty
of the cerebral cortex i s not essential for al l consc i ous processes,though i ts parti c ipation i s necessary for others
, parti cu lar lyall intel lectual and voluntary activities . The thalamus
,on the
other hand,can act independently of the cortex in the case of

THE CE RE BRUM 167
painfu l sensibi l i ty and the enti re series of pleasurable and painfu l qual i ties ; for the thalam i c centers when isolated from thei rcorti cal connections are found to be concerned m ain ly withaffective exper ience, and destructive lesions whi ch involve thecortex alone do not disturb the painfu l and affective quali t ies ofsensation (see p.
The relat ions of the thalam i c nuc lei and of som e Of the tractsconnected with them are shown as seen from above in Fig. 77
and in a section paral le l with the m edian plane in Fig. 78 .
THE DIENCEPHALO NI . Epithalamus .
1 . Chorio id plexus Of the thi rd ventricle.
2 . P inea l body (epiphys is ) .3 . Habenula (receives the stria medullaris from the o lfactory centers
and sends fibers to the cerebral peduncle) .I I . Thalam us .
1 .Dorsa l part.( 1 ) Med ial group of nuclei .
(a) Med ia l nucleus (receives fibers from the O lfactory areaand neothalamus and from the trigeminal lemni scussends fibers to the O lfactory area
,corpus striatum ,
subtha lamus,and probab ly cerebral cortex) .
(b) Anterior (or dorsal ) nucleus (receives fibers from the
mammillary body and sends fibers to the corpus striatum ) .
(2) Lateral group Of nuclei (neothalamus) .(a) Lateral , ventral , and posterior nuclei (receive the
m ed ial , spinal , and trigemina l lemnisci ; conn ect withparietal and fronta l cortex by ascendingand descending somestheti c project ion fibers ) .
(b) Pulvinar and lateral geni culate body (receive Opti ctracts ; connect wi th occip ital cortex by ascend ingand descend ing Op ti c projec ti on fibers) .
(c) Med ia l geni culate body (receives the latera l or acoust i clemn iscus ; connects w ith temporal cortex by as cending and descend ing aud i tory projec t ion fibers) .
[The two geni culate bod ies = m etathalamus , B . N . A .]2 . Ventral part , or subtha lamus (a m otor coordination center re
ceiving fibers from the dorsal part O f the thalamus , from the
corpus striatum and from the pyramida l tract ; sends fibers tothe pedunculus cerebri ; compr ises the body of Luys , Forel $8field H2, and some adjacent gray matter ; is cont inuous behi ndwi th the substant ia nigra of the cerebra l peduncle) .
III . Hypothalamus .1 . Tuber cinereum (o lfacto-v isceral correlat ion center) .2 . Mammil lary body (receives fibers from the olfactory centers ; sends
fibers to the cerebra l peduncle and nucleus anterior thalam i ) .3 . Hypophys is .

168 INTRODUCTION To NEUROLOGY
Some of these centers are seen in cross- section in Fig. 79 . The
preceding analysi s of the diencephalon, which differs in some
respects from that of the B . N . A . (p. i s summ arized in theaccompanying table (p. whi ch inc ludes also a few of them ore important fiber tracts connected with each nuc leus .In front of the thalamus lie the corpus striatum and olfactorycenters (see Fig. 45
, p. and above these l ast two i s spreadthe great expanse of the cerebral cortex or pal l ium . The corpus
stri atum consi sts of m asses of gray m atter separated by sheetsof white m atter
,an arrangement whi ch gi ves a striated appear
ance in section .
In studying the comparative anatomy of the cerebral hem ispheres wefind the corpus striatum wel l devel oped in som e l owervertebrates whi ch lack the cerebral cortex, and very highly d eveloped in others , l ike repti les and bi rds, where the cortex i s
present , though very sm al l . In these an im al s the corpus striatum appears to be a reflex center of great importance and ofhigher order than the thalam us ; and the d fferentiation of thisapparatus seem s to have been a necessary precursor of the elaboration of the cerebral cortex as we find i t i n the m amm al s .The functions of the m amm al ian corpus striatum are very
Obscu re. I t i s connected by both ascending and descendingfibers with various nuc lei of the thalam us and cerebral pedunc le,and also wi th the cerebral cortex. Ramon y Caj al i s of theopini on that the m amm al ian str iatum functi ons chiefly to re
inforce the descending m otor impu lses which leave the cerebralcortex, these system s of fibers gi ving Off col lateral branches asthey traverse i t, and the stri atum i tself sending important d escending tracts into the thalamus and cerebral pedunc le.
The white m atter Of the corpus str iatum consists part ly of thefibers already m entioned as passing between i t and the thalam usand cortex, but chiefly of fibers passing between the cortex and
deeper parts of the brain stem ,having no functi onal connection
wi th the striatum i tself . These are called projection fibers .They are part ly ascending and descending fibers passing betweenthe thalam us and the cortex (the opti c , audito ry, and som estheticprojection system s, or radiat ions , whi ch have al ready beenmentioned , p. and partly descending m otor projectionfibers of the cort i co- spinal or pyram i dal tract (p. 140 and Fig.

170 INTRODUCTION To NEUROLOGYGyms frontalis inf.
Cyrus cinguliCorpus callosumExternal capsuleLateral ventricleCaud ate nucleus
Anterior limb ofinternal capsuleColumn of fornixLenti form nucleus
Posterior limb ofinternal capsule
PulvinarHabenula
Lateral ventricl
Caud ate nucleusParieto-occip. fimure
Cuneus
Fig. 80.—Longitud inal section through the hum an cerebral hem isphere
pass ing throu the internal capsule, some of the fiber systems of wh i ch arenumbered as sted below :
1 . Fronta l thalamic tracts between the med ial nucleus Of the thalamusand the fronta l lobe Of the cerebral cortex .
2 . Frontal pontile tract between the frontal lobe of the cortex and thepons .
3 . Cortico-oculomotor tract from the motor cortex to the nucleus of theoculomotor nerve .
4 . Cort i co—bulbar trac ts from the m otor cortex to them otor nuclei of them edul la Ob longata .

THE CERE BRUM 171
the sensory projection system s of sight, hearing, and generalsensib i l ity into the cerebral cortex. The subdivisi on of thediencephalon i s summ ari zed in the table on p. 167 . The cor
pus str iatum in l ower vertebrates i s an important reflex center ;in man i ts functi ons seem to be subsidiary to those of the cerebral cortex for the m ost part . I t consi sts of two chief m asses of
gray m atter, the caudate and lentiform nuc lei,with sheets of
whi te m atter between and within these m asses . The chief system s of fibers of the white m atter are accum u lated in the internalcapsu le which l ies between the lentiform nuc leus lateral ly and thecaudate nuc leus and thalam us m edial ly . Through the internalcapsu le run the proj ection fibers whi ch connect the cerebral cortex wi th the l ower parts of the brain stem ,
inc luding the sensoryradiations from the thalam us and the descending system s to the
pons and brain stem and the great pyram i dal tract, whi ch is thevoluntary m otor path from the cortex to the spinal cord .
L ITERATU REHEAD
,H.
, and HOLMES, G . 19 1 1 . Sensory Disturbances from CerebralLes ions, Brain ,
vo l . m d v,pp . 109 -254 .
HE RRICK,C . JUDSON . 1 9 13 . A rti cle Brain Anatomy, in Wood
’s Reference Handbook of the Medi ca l Sciences , 3d ed .
, vo l . ii , pp . 274—342 .
J OHNSTON ,J . B . 1906 . The Nervous System Of Vertebrates , Philad el
v. MONAKOW, C . 1895 . E xperimentelle und pathologische-anatom ischeUntersuchungen t
'
i ber d ie Haubenregion ,d en Sehh i
'
i gel und d i e Regio subthalam ica
,Arch . f . Psychiat.
, B d . 27 .
SACHS, E . 1909 . On the Structure and Functiona l Relat ions of the Op ti cTha lam us , Brain, vo l . xxxi i , pp . 95- 186 .
5 . Cort i co-rubri c tract from the motor cortex to the nucleus ruber .6 to 10. Pyram idal trac t (tractus cort ico- sp inalis ) from them otor cortex
to the spinal cord , w ith the fo llowing parts6 . To the cervi cal sp inal cord for the m uscles of the shoulder.7 . To the cervica l cord for the m uscles Of the arm .
8 . To the cervica l cord for the muscles of the hand .
9 . TO the l umbar cord for the muscles of the leg.
10 . To the lumbar cord for the muscles of the foot .1 1 . Somesthet ic rad iat ions from the latera l and ventra l nucl ei of the
tha lamus to the cerebral cortex .
1 2 . O ccipito-temporal pont ile tract to the pons , and temporo—thalaim c
trac t to‘ the tha lamus .13 . Aud itory rad iat ion from the m ed ial geni culate bod y to the superior
temporal gyrus .
14 . Opt i c rad iation from the pulvinar and lateral geni culate body to thecuneus in the occipital lobe of the cortex .

CHAPTER XI
THE GENERAL SOMATIC SYSTEMS OF CONDUCTION
PATHS
IN thi s and the fol lowing chapters we shal l review the conduction pathways fol lowed by som e of the ch ief sensori-m otorsystem s and add some fu rther detai l s to the general desc riptional ready given,
beginning with the m ore general i zed som ati csensory functi ons .C l inical neurologi sts have long been i n the habi t of grouping
together the different form s of deep and cutaneous sensib i l i tyunder the term “
general sensib i l i ty .
”The m ore refined re
searches of recent students (espec i al ly Sherr ington, Head ,Trotter and Davies, B rouwer , see the bibl iographies on pp.
94 and 142) have given us a m uch m ore prec i se analysi sof these system s
,as al ready explained . The peripheral
nerves of deep sens ibi l i ty (exc lusive of those devoted tostr ictly visceral functions) are anatom i cal ly di stinct fromthose of cutaneous sensibi l i ty. Physi ologi cal ly, the nerves ofdeep sensib i l i ty are devoted chiefly to propr i oceptive functions(m usc le sensib i l i ty , j o int sensib i l i ty, and the nerves ofcutaneous sensibi l i ty ch iefly to exteroceptive functions (touch ,temperature, and pain) ; but th i s holds only approxim ately, fornerves of deep sensibi l i ty may also serve the exteroceptive functions of pressure and painfu l response to overstim u lation,
though wi th a higher stim u lus threshold than in the sk in,and the
cutaneous nerves also partic ipate to some extent in the propri oceptive functions of spatial orientation of the body and i ts mem
bers (see pp. 77 if . and
Exteroceptive System s .
—The nerves serving the functionsof touch , pressure, temperatu re, and pain of the body andl imbs , whether derived from the skin or the deep tissues, immediate ly after thei r entrance into the spinal cord term inate i n
172
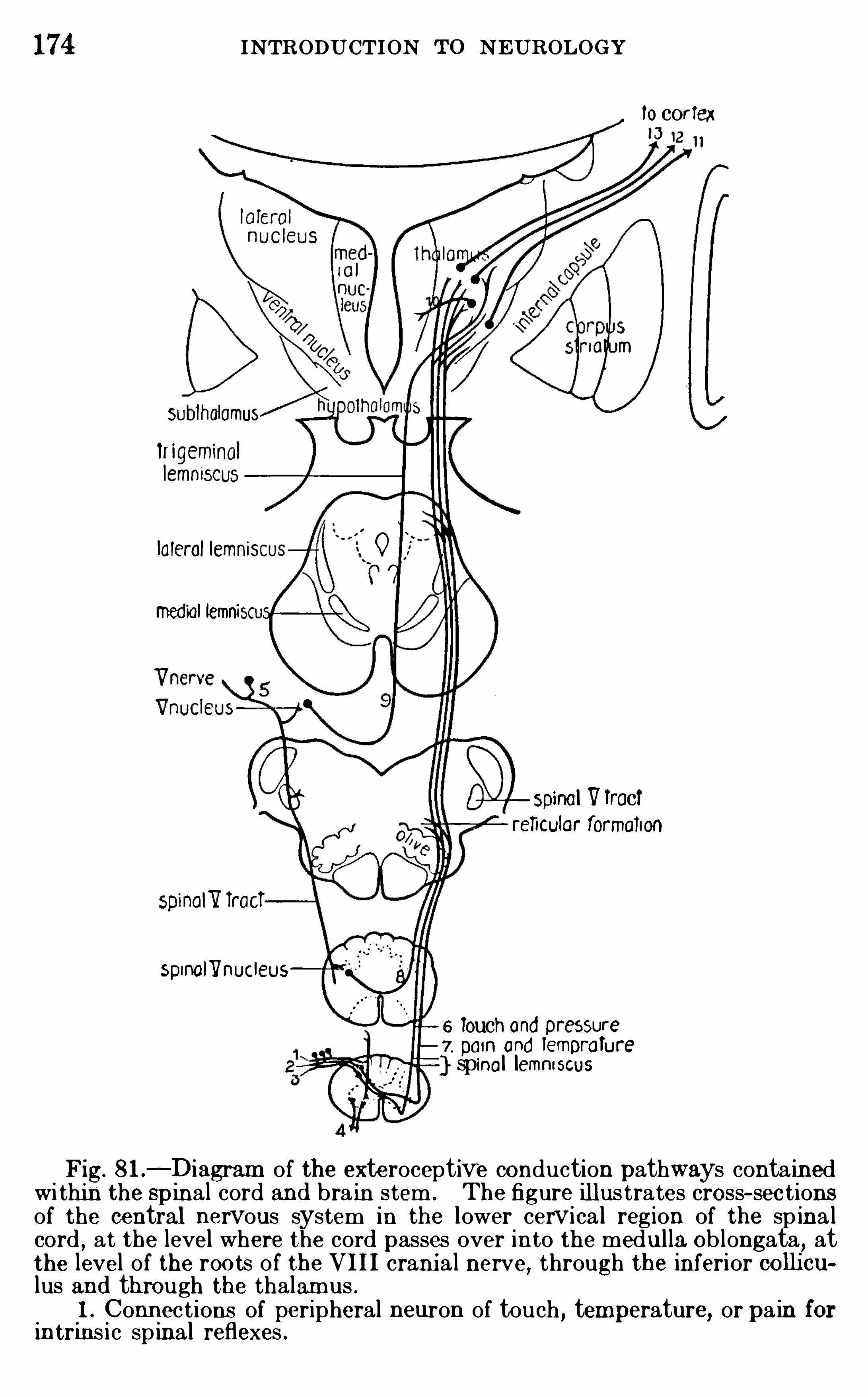
174 INTRODUCTION NEUROLOGY
locortex
Sublholamus
lalerol Iemnis
med ial lemni
Spinal VTractreficolar formation
spinalllnucleus
6 Touch and Dresserepom ond lemorafurespinal lemniscus
Fig. 81 .—Diagram of the exteroceptive conduction pathways conta ined
wi thin the spinal cord and brain stem . The figure ill us trates cross- sec tionsof the central nervous system in the lower cervica l region of the sp ina lcord , at the level where the cord passes over into the medul la Ob longata, atthe level of the roo ts of the VIII crani a l nerve
,through the inferior collicu
lus and through the thalamus .1 . Connect ions of peripheral neuron of touch , temperature, or pa in for
intrins i c spinal reflexes .
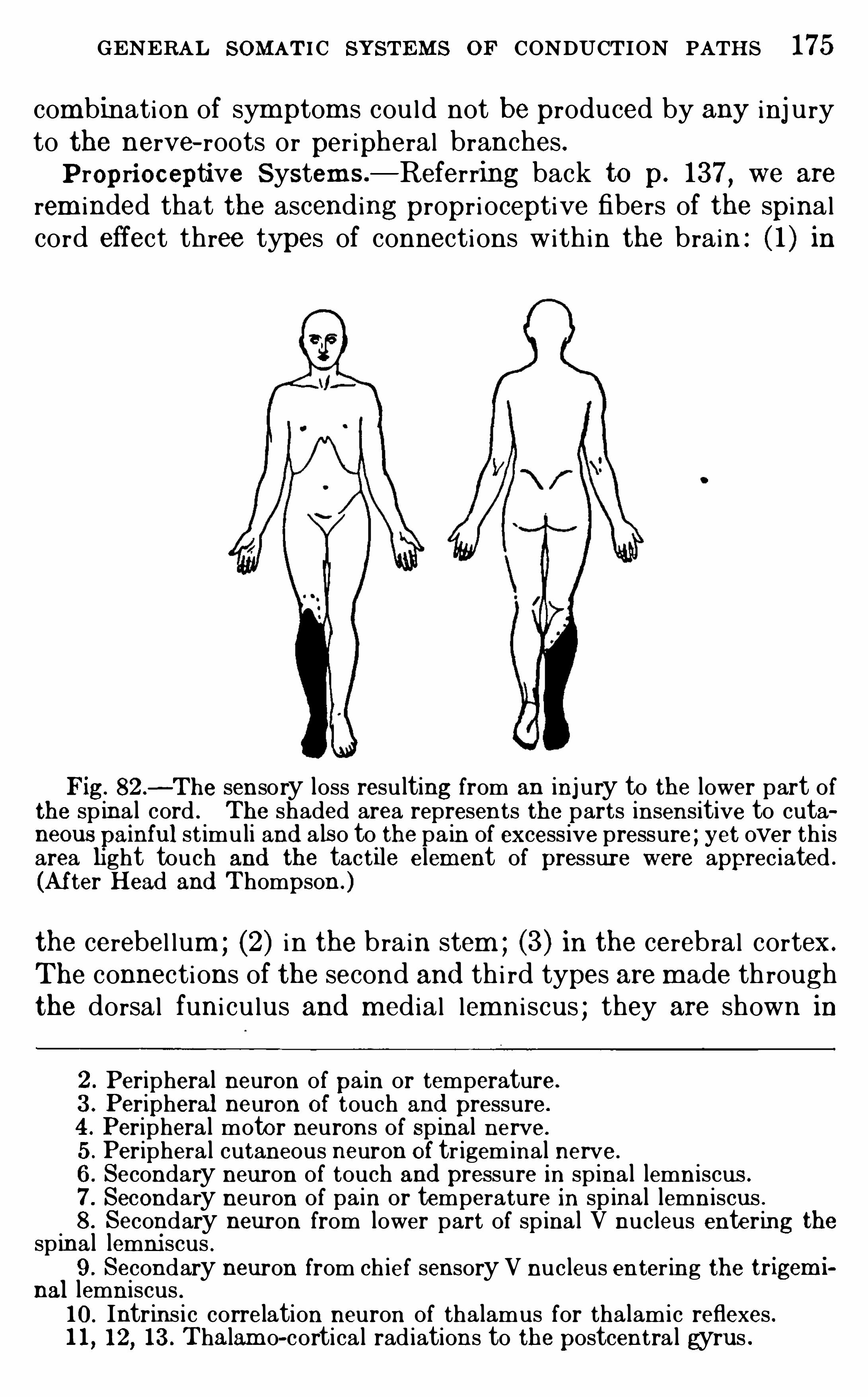
GENERAL SOMATIC SYSTEMS or CONDUCTION PATHS 175
combination of sym ptom s cou l d not be produced by any inj uryto the nerve- roots or per ipheral branches .P ropri oceptive System s .
—Referring back to p. 137,we are
rem inded that the ascending propr i oceptive fibers of the spinalcord effect three types of connections with in the brain : ( 1 ) in
Fig. 82 .—The sensory loss resulting from an injury to the lower part of
the spina l cord . The shaded area represents the parts insensitive to cutaneous ainful st imuli and a lso to the pa in of excess ive pressure ; yet over th isarea light touch and the tactile elem ent of pressure were appreciated .
(After Head and Thompson .)
the cerebel lum ; (2) i n the brain stem ; (3) in the cerebral cortex.
The connections of the second and th i rd types are m ade throughthe dorsal funi cu lus and m edial lemni scus ; they are shown in
2 . Peripheral neuron of pa in or temperature.
3 . Peripheral neuron Of touch and pressure.
4 . Periphera l motor neurons of Spinal nerve.
5 . Periphera l cutaneous neuron Of tr igeminal nerve.
6 . Secondary neuron of touch and pressure in spina l lemniscus .
7 . Secondary neuron of pain or temperature in spinal lemn iscus .8 . Secondary neuron from lower part of spina l V nucleus entering the
spinal lemni scus .9 . Secondary neuron from ch ief sensory V nucleus enter ing the trigem i
nal lemniscus .10 . Intrins i c correlation neuron of thalamus for thalamic reflexes .l l , 12, 13 . Thalam o-cortica l rad iations to the postcentra l gyrus .
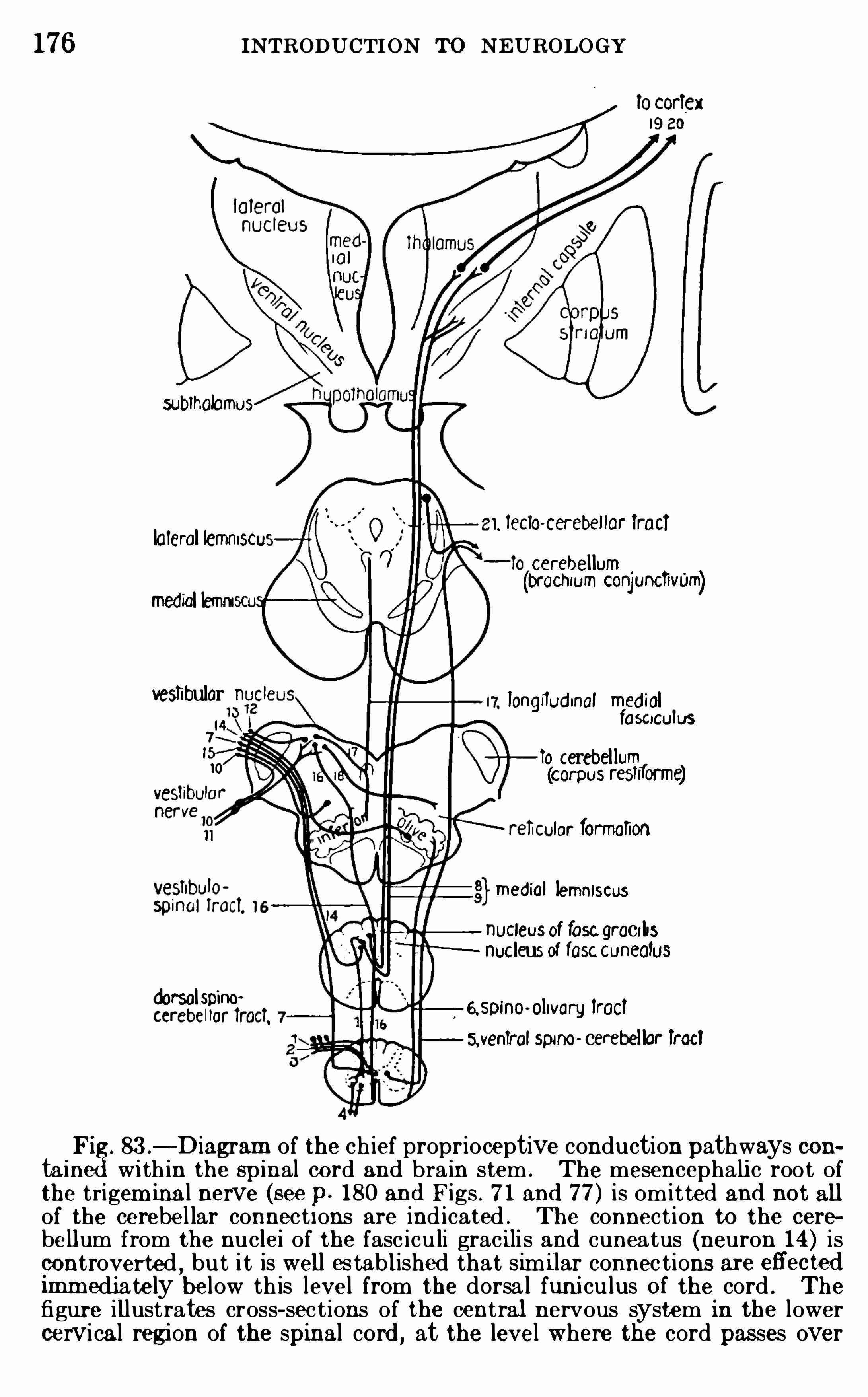
176 INTRODUCTION To NEUROLOGY
subiholomus
1, ieclo-cerebellor iracilateral lemmSCUo cerebel lum(bl
'
OCl'
llUm conjuncfivum)med ial lemui
vestibular longflud inal med ialfaSCiCuluS
To cerebel lum(corpus restiforme)
lar formation
med ial lemniscusucleusof fasc graci lisnucleusof iascCuneoius
d orsalSDiflO'
espino-olivary lroclaveniral spino cerebel lar iroci
Pi 83 .—D iagram of the ch ief proprioceptive conduction pathways con
ta in wi th in the spinal co rd and bra in stem . The mesencephali c root ofthe trigem inal nerve (see p. 180 and Figs . 7 1 and 77 ) is omitted and not allof the cerebe l lar connect ions are ind icated . The connect ion to the cerebe llum from the nuclei of the fasc icu li grac i li s and cuneatus (neuron 14 ) iscontroverted
,bu t i t is well es tablished that sim i lar connec tions are efl'
ectedimm ediate ly below th is level from the dorsa l funi culus Of the cord . Thefigure illustrates cross- sec t ions of the central nervous system in the lowercervical region of the spinal cord , at the level where the cord pas ses over

178 INTRODUCTION To NEUROLOGY
with the propri oceptive group of reactions . These may be
unconsc i ous reflexes of m otor coordination and the m aintenanceof equil ibr ium ,
or they m ay com e into consc i ousness as sensations of posi tion and orientati on of the body and i ts parts and ofspatial di scr im i nation . Pu rely exteroceptive stim u l i , whethertransm i tted by the deep nerves or by the cutaneous nerves , m ay
be carr ied for a few segm ents in the dorsal funi cu l i (Fig. 81 ,
neuron but they are soon fi ltered off into the gray m atter of
the dorsal column,and after a synapse here they are sorted into
functional ly di stinct tracts on the opposi te side of the cord .
The tacti le elem ents of the m ixed peripheral root fibers enteringthe dorsal funi cu lus are drawn off later than are the elements fortherm al and painfu l sensib i l i ty ; and some of the componentscomm only reckoned with cutaneous exteroceptive sensibi l i tyremain in the dorsal funi cu lus for i ts enti re length . These are
chiefly two-point di sc r im inati on,and disc rim ination Of si ze,
shape, form ,and texture of su rfaces . These all involve a com
pari son and discr im ination i n consc i ousness of spatial factorsand are
,therefore, bound up with those fibers whi ch serve the
propr ioceptive reflexes , wh i ch are unconsc i ous spatial adj ustments .Som e pecu l iar com binations of sym ptom s ari se from the factthat
,whereas the ascending proprioceptive impu l ses (so far as
these are consc i ously perceived ) pass up in the dorsal fun i cu lusof the same side for the enti re length of the cord , the impu l sesOf the exteroceptive impu lses , within a few segments of thei r
po int of entrance into the cord , are transferred to the opposi te
s ide to ascend in the spinal lemn i scus tracts . F rom th i s i t follows that a local i zed central inj ury involving the dorsal graycolumn and dorsal funi cu lus of one side on ly wi l l cut off al lascending propri oceptive impu l ses whi ch pass through the dorsal funiculus from l ower level s on the sam e side of the body as thelesion
,and at the sam e tim e wi l l abol i sh both propr i oceptive and
exteroceptive functions in a c i rcum scr ibed region of the same
side of the body whose exteroceptive neurons Of the fi rst orderd ischarge into the inj ured part of the dorsal gray column .
F igure 84 i l lustrates the loss Of sensibi l i ty to painfu l stim u l iresult ing from a tum or in the cervi cal regi on of the spinal cord .
Tacti le, temperature, and deep sens ibi l i ty were also profoundly
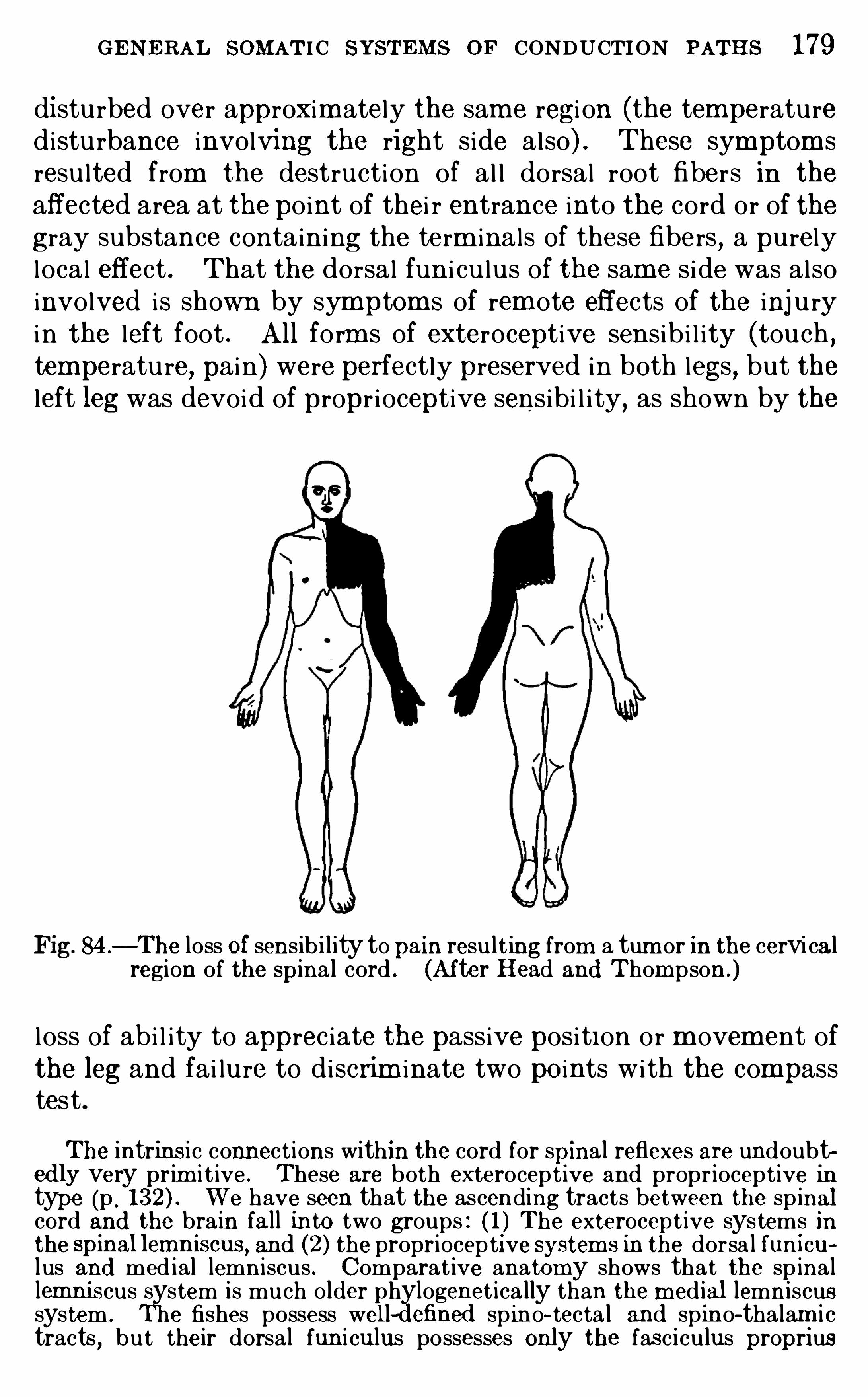
GENERAL SOMATIC SYSTEMS or CONDUCTION PATHS 179
disturbed over approxim ately the sam e regi on (the temperaturedistu rbance involving the right side also) . These symptom sresu lted from the destruction of al l dorsal root fibers in the
affected area at the point of thei r entrance into the cord or of the
gray substance containing the term inals of these fibers , a purelylocal effect . That the dorsal fun i cu lus of the sam e side was alsoinvolved i s shown by symptom s of rem ote effects of the inj u ryin the left foot . All fo rm s of exteroceptive sensib i l i ty (touch ,temperatu re, pain) were perfectly preserved in both legs, but theleft leg was devoid of propr i oceptive sensibi l i ty, as shown by the
84 .—The loss of sens ib i l ity to pain result ing from a tum or in the cervi calregion of the spina l cord . (After Head and Thom pson.)
l oss of abi l i ty to apprec iate the pass ive positi on or m ovement ofthe leg and fai lure to discrim i nate two po ints with the compasstest .
The intrins i c connections within the cord for spina l reflexes are undoubted ly very prim i t ive . These are both exterocep t ive and propr ioceptive intype (p . We have seen that the ascending tracts between the spinalcord and the brain fall into two groups : ( 1) The exterocept ive systems inthe spinal lemniscus, and (2 ) the propriocep ti ve system s in the dorsa l funi culus and med ial lemniscus . Comparat ive anatomy shows that the spinallemniscus system is much o lder phylogenet ica lly than the m ed ial lemniscussystem . The fishes possess well—d efined spino- tecta l and spino—thalami ctracts , but their dorsal funi culus possesses only the fasciculus propri us

180 INTRODUCTION To N EU ROLOGY
fibers (cf . Figs . 66 , 67 , pp . 150, 15 1 ) and they lack the m ed ial lemni scusal together . The spino- cerebel lar tracts, on the other hand , are very anc ientand are present from the lowest to the hi ghest vertebrates .
These considerations suggest that the first fibers to pass from the spinalcord to the h i gher centers of the brain , and presumab ly the first sensoryim pulses from the spina l nerves to be conscious ly perceived , were those oftouch and temperature transmitted through the spina l lemniscus . (Pa inis probab ly also very prim i t ive as a conscious experience, bu t it is doubtfulwhether i t is represented in the sp ina l lemniscus of lower form s ; see p.
The proprioceptive im pulses in lower vertebrates are coord inated qui te unconscious ly in the bra in stem and cerebellum , and it is only in the higherform s that th is system of nervous im pulses reaches the tha lam us ( throughthe medi al lemn iscus ) and cerebra l cortex for conscious contro l . C lin icalevidence shows that the med ia l lemniscus connect ions in m an are concernedw i th the conscious adj ustmen ts of the pos i tions and orientat ion in space ofthe body and i ts members and wi th spat ial d iscrim inations of various sorts ,rather than wi th the senses of touch and pressure as externa lly projected .
The innervat ion of the organs of m uscular sensib i l ity and tendon sensibili ty in the head is not as ful ly known as in the case of those of the trunkand l imbs , as above described . Sherrington and Tozer have recentl
shown that such organs are present in the muscles whi ch move the eyeb
and that their nerves accompany the m otor fibers of the III, IV , and VIcrania l nerves ; °but of the centra l connections of thes e sensory nerve-fibersof the eye muscles noth ing is known . It is suggested by the researches ofJohnston
,W il lems , and many others that the j aw m uscles , whi ch receive
their motor innervation from the motor V nucleus (nucleus m asticatorius ) ,receive their senso ry innervation from the m esencepha li c nucleus of the Vnerve, whose posi t ion a long the latera l border of the aqueduct of Sylviusis seen in Figs . 7 1 , 75 , and 77 . But recent stud ies of E dgeworth haveshown that these muscles a lso receive sensory fibers from the semilunaror Gasserian gangli on of theV nerve, and the quest ion requi res further investigation . Poss ib ly the sensory fibers from the Gasserian ganglion to the
m uscular branches of the V nerve conduct impress ions of deep sens ib ili tyof pressure and pain of the exterocep tive type, wh i le those from the mesencephal i c V nucleus innervate the muscle sp ind les for true proprioceptivesensib i li ty.
The fibers of the ch ief sensory root of the V nerve in part end in the ch iefsensory V nucleus near the level of thei r entrance into themedulla Ob longata(Figs . 7 1 , 77) and in part pass downward through the who le length of them ed ul la Ob longata and upper leve ls of the Spina l cord as the spina l V trac t(Figs . 64, 7 1 , 72 , I t is suggested by clini cal and comparat ive evidencethat the spinal V tract and its nucleus are connected wi th a phl geneticallyo ld type of reaction to touch
,temperature, and pain, probab ly chiefly reflex,
whi le the chi ef nucleus is concerned w i th the more recently acquired d iscrim inations of these systems w i th m ore d i rec t corti cal connect ions . Thefibers of the trigeminal lemnis cus (p . 157 ) fo l low two separate tracts aris ingfrom thos e two parts of the sensory V nucleus , only the upper one of whi chis shown in Fig . 77 , though both are shown in Fig . 81 (neurons 8 and
Motor Path s—Throughout the length of the spi nal cord andbrain stem the ascending fibers Of both exteroceptive and pro
prioceptive sensibi l i ty gi ve off collateral branches into the reticu

182 INTRODUCTION To N EUROLOGY
In very young amph ibian embryos Coghill l finds a st i ll Sim pler cond itionwh ich is probab ly also more prim itive . In the spinal cords of these larvaethe ind ividual neurons of themotor tegm entum give rise both to fibers Of thelongi tud inal conduction tract of m otor coord ination (fasciculus propriusventra l is ) and to peripheral fibers of the ventral roots , the latter aris ing asco llatera ls of the longitud inal axons . In o lder larvae separate neurons havebeen differentiated for these two functions of per ipheral conduction andlongitud inal conduct ion . The steps in the embryo logic development andprobab le evolu tion of the more complex centers of adjustment have beenbriefly reviewed by Herrick and Cogh ill (see p.
Summary—The Old c l ini cal concept general sensibi l i tyhas recently been analyzed into a number of components, them ost fundam ental divi si on being the distincti on between a
group of exteroceptive and a group of propr i oceptive system s .The exteroceptive system s are transm i tted from the spinalcord to the brain through a complex tract, the spi nal lem ni scus
,
w i thin whi ch there are separate pathways for the three qual itiesof sensation,
touch,temperature, and pain . These sensati on
qual ities com e into consc i ousness with a dist inct peripheral orexternal reference. The propri ocept ive system s (m usc le senseand al l ied types) are transm i tted to the brain through the dorsalfuni cu lus of the sam e side of the cord
,the m edial lemni scus of
the opposite side, the thalam us , and the som estheti c radiati onsto the cerebral cortex ; and also through the spino- cerebel lartracts to the cerebel lar cortex. Most of these reactions ofspatial adj ustment do not com e into consc i ousness at al l
,but
some appear subj ectively as sensations of postu re, bodi ly m ovement, and spatial d i scr im ination . The cerebel lum i s the greatc learing house for these and al l other afferent system s whi chare concerned in the propri oceptive functions, so fa r as these areunconsc i ously perform ed .
1 COGHILL , G . E . 19 13 . The Prim ary Ventral Roots and Somati c MotorColumn of Am blystoma , Jour . Comp . N eur .
,vol . xxi i i , pp. 12 1—144 .
19 14 . Correlated Anatomica l and Phys io logica l Stud ies of theG ro
iv
fit
li
gg
ghe Nervous System of Amphib ia, Jour . Comp . Neur.
,vo l . xxiv,
pp.

CHAPTER X I I
THE VESTIBULAR APPARATUS AND CEREBELLUM
THE general som ati c sensory system s considered in the lastchapter inc lude som e of the m ost pr im i t ive reflex mechani sm s .These fal l into two groups—the exteroceptive system s and thepropr i oceptive system s (pp. 77—89 )—and each Of these groupscompri ses , in addition to its prim i t ive general i zed m embers
,
certain so- cal led organs of spec ial or h igher sense . The Spec ia lexteroceptive sense organs are the organ of hearing (p. 195)and the organ of vi sion (p. The spec ia l propri oceptivesense organs are the sem i c i rcu lar canals of the internal ear ; andthose wi l l next be desc ribed
,together with thei r central m echan
i sm s in the medu l la Oblongata and cerebel lum .
The Vestibular Apparatus—The internal ear contains twoqu i te di stinct groups of sense organs , the organ of hearing in thecochlea and the vestibu lar organs (utri c le, saccu le, and sem icircular canal s) , both of whi ch are suppl ied by theVII I c rani al nerve,whi ch accordingly has two parts, the cochlear and the vestibular nerves . The sem i c i rcu lar canal s are the m ost highly spe
cialized end—organs of the propr ioceptive ser ies and are con
cerned chiefly wi th the m aintenance of bodi ly equ i l ib rium . The
general structu re of the internal ear i s described on p. 195 ; herewe need merely m ention that the three sem i c i rcu lar canals
(ductus sem i c i rcu lares) of each ear lie approxim ately at rightangles to each other , as shown diagrammati cal ly in Fig. 85 , and
each canal i s d i lated at one end to form the ampu l la , wi th inwhi ch i s a patch of sensory epi thel ium from whi ch hai rs proj ectinto the contained flu i d (see F igs . 32 and A m ovem ent ofthe head in any di recti on wi l l cause a flow of the flu i d in one orm ore of these canals in each ear
,whi ch in tu rn wi l l exc i te a
nervous impu l se in the hai r- cel l s of the corresponding ampu l lae.
These nervous impu l ses wi l l be transm i tted to the vestibu larcenters of the brain
,where theywi l l be so analyzed as to cal l forth

184 INTR ODUCTION To N EU ROLOGY
the appropr iate reaction to them ovement whi ch has exc i ted thepart i cu lar sem i c i rcu lar canal s i nvolved .
The fibers of the vestibu lar nerve enter themedu l l a oblongataimmediately beh ind the pens and term inate i n a vestibular nu
Fig. 85 .—Diagram of the posi tion of the sem i circular canals in the head ,
as seen from beh ind . On each s ide it w ill be seen that the three canals liein planes at right angles to one another . The externa l or horizonta l canals(E ) of the two s ides lie in the same plane. The anterior cana l of one s ide(A ) lies in a plane parallel to that of the posterior cana l (P ) of the otherside. (After Ewald .)
cleus which form s an em i nence on the floor of the fou rth ventri c lein th i s regi on (F igs . 71 , Thi s nuc leus has fou r subdivi si ons
,
as fol lows :
N ucleus nerv i vestibuli m ed ia l is (of Schwalbe, a lso cal led nucleus dorsalis and princ ipa l nuc leus ) .
N ucleus nervi ves t ibul i latera l is (of Dei ters) .N ucleus nervi vest ibul i super ior (of Bechterew) .Nucleus nervi vestibuli spinal is .
The arrangement of these nuc lei and of some of thei r secondary connections i s shown in Fig. 86 . Som e of these connectionsare made wi th the m otor nuc lei and reti cu lar form ation of the

186 INTRODUCTION To NEUROLOGY
lus by which fibe rs descend to the spinal cord (ch ieflyfor tu rning m ovements of the head by the neck m usc les) andascend to the m i dbrain . The last-m enti oned fibers connectch iefly wi th the nuc lei of the m otor
.
nerves for the eye m usc les
(I II , I V , and V I pai rs of c ranial nerves) , thus providing forthe conj ugate m ovem ents of the eyes whi ch accompany headm ovem ents (in thi s way , for instance, enabl ing one to keep the
gaze fixed upon a stationary object whi le the head i s m oving,cf. p.
I t wi l l be noticed that there i s no important pathway fromthe vestibu lar nuc leus to the thalam us and cerebral co rtex, fo rthe equ i l ibratory reactions exc i ted from the sem i c i rcu lar canalsare norm al ly unconsc i ously perfo rm ed . Thi s i s i n m arkedcontrast with the connecti ons Of the
'
cochlear nerve, for the auditory reactions are Often consc i ously di rected (p. Therei s,however , an important connection w i th the cerebel lum
,
partly di rectly by root fibers of the vestibu lar nerve and part lyby secondary fibers from the supe ri or and lateral vestibu larnuc lei (Fig. The cerebel lum i s , accordingly , an importantcenter of adj ustment for the propr ioceptive reflexes, and to thi sou r attenti on w i l l next be di rected .
Th e Cerebel lum .—Thi s important organ i s an over lord whi ch
dominates the proprioceptive functi ons of the body in som e
what the sam e way that the cerebral cortex di rects and controlsthe exteroceptive reactions . Both of these organs are secondarily added to the m ore prim i t ive segm ental structu res of thebrain stem
,that is
,they are suprasegm ental (p.
The correlation centers of the brain stem,and parti cu larly
those of the cerebral cortex,analyze the afferent impu l ses enter
ing the brain and determ ine what parti cu lar reactions are ap
propriate in each si tuation. Af ter the character of the m ovem ent has been determ ined in th is way
, the proprioceptive system s cooperate in i ts execution ,
and the cerebel lum i s the cen
tral coo rdination stati on for the propri oceptive reactions .N one Of i ts activi ties com e into consc i ousness .The cerebel lum , therefore, i s intim ately connected with al l
sensory cente rs whi ch are concerned in the adj ustment of thebody in space and m oto r contro l in general . The m aintenanceof bodi ly equ i l ibrium i s the m ost important of these functions,

THE VE STIBULAB APPARATU S AND CERE BE LLUM 187
and the sem i c i rcu lar canals of the internal ear (pp. 89 , 196) arethe receptive organs, whi ch are of chief importance in these reacti ons . Comparative and em bryologi cal studies Show that thecerebel lum was devel oped as a di rect outgrowth from the pri
m ary centers for the sem i c i rcu lar canals in them edu l la oblongata
(the acoustico—lateral area of fishes,Fig. and even in the
hum an body root fibers from the vestibu lar branch of the VI I Icran ia l nerve enter the cerebe l lum di rectly . Neurons of thesecond order also enter the cerebe l lum from the vestibu larnuc leus
,as wel l as from the spinal cord and from pract ical ly a l l
of the som ati c sensory centers of the brain ; there i s al so a veryimportant path from the cerebral cortex.
The hum an cerebel lum consists of a m edian l obe, the worm
(verm i s) , and two larger cerebel lar hem i spheres . The verm i sreceives fibers chiefly from the som ati c sensory centers of thebrain stem and spinal cord , and i t alone i s well devel oped inl ower vertebrates (from fishes to bi rds, see Fig. The cerebel lar hem i spheres vary in si ze in different m amm als in proportion to the si ze of the cerebral cortex, being, therefore, m uchlarger in man than in any other anim al . Thei r appearance fromthe ventral s ide i s seen in Fig. 53 . The cerebel lum i s attachedto the brain stem by three stalks or pedunc les on each side, thesuperior pedunc le (brachium conj un ctivum ) , the m i dd le pe
d uncle (brach ium ponti s) , and the i nfer i or pedunc le (corpusrestiform e) .F igure 87 i l lustrates diagramm ati cal ly the ch ief pathways
whi ch enter the cerebel lum , and Fig. 88 those by which nervousimpu l ses leave i t . We cannot here describe these connectionsin detail
,but can ment ion a few only of thei r general features .
The cerebe l lum ,as al ready stated
,receives afferent impu l ses
from al l of the impo rtant som ati c sensory centers and also fromthe cerebral cortex. The afferent fibers from the brain stementer by the superi or and inferi or pedunc les . The pons i s anem inence under the upper part of the m edu l la oblongata (Fig.
53) which contains gray centers (the ponti le nuc lei ) . F ibers
pass into the ponti le nuc lei from the assoc iation centers of thecerebral cortex by way of the cort i co-ponti le tracts , and from the
m oto r areas of the cerebra l cortex by way of col lateral branchesfrom the cort i co—spinal tract as i t passes through the pens .
i.
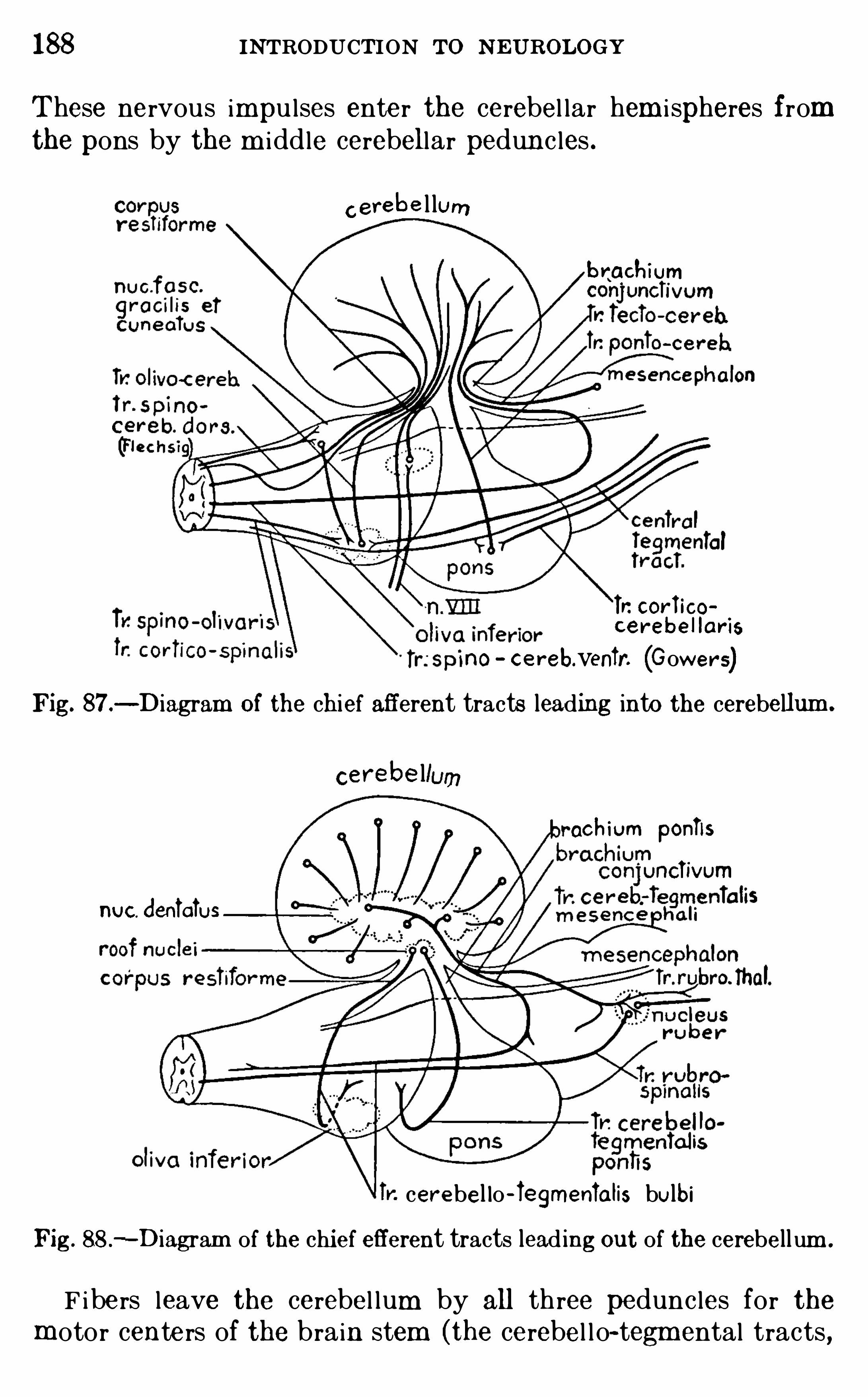
188 INTRODUCTION To N EUROLOGY
These nervous impu lses ente r the cerebel lar hem i spheres fromthe pons by the m i ddle cerebellar pedunc les .
c erebe l lum
brachium
Tr. cor i icocerebe lIar is
no ce reb.venir. (Gowers)Fig. 87.
-Diagram of the chief afferent tracts leading into the cerebellum .
cerebe llum
ochiom ponlls
ncTivum
nuc d enialus
cerebel lo -Tegmenlalis bulbiFig. 88 .
—Diagram of the chief efferent tracts lead ing out of the cerebell um .
F ibers leave the cerebel lum by all three pedunc les for themotor centers of the brain stem (the cerebel l o- tegm ental tracts ,
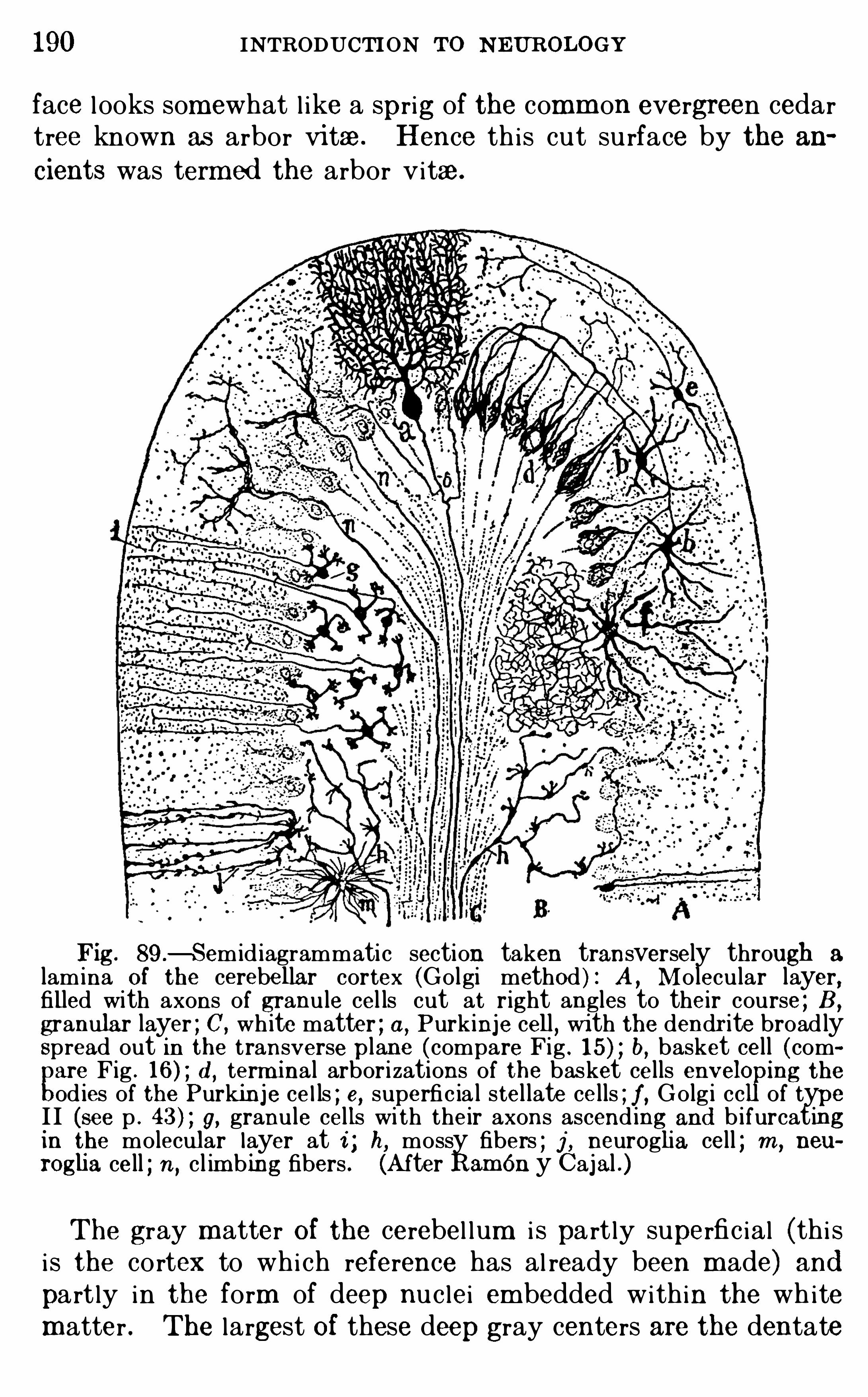
190 INTRODUCTION To N EUROLOGY
face l ooks somewhat l ike a sprig of the comm on evergreen cedartree known as arbor vitae. Hence th is cut su rface by the an
cients was termed the arbor vitae.
Fig . 89 .—Semid iagrammatic section taken transversely through a
lamina Of the cerebellar cortex (Go lgi method ) : A , Mo lecular layer,filled with axons of granule cel ls cut at right angles to their course ; B ,
granular layer ; C, wh ite matter ; a,Purkinje cell, with the dendrite broad ly
Spread out in the transverse plane (compare Fig . b,basket cell (com
pare Fig . d
, term inal arborizations of the basket cells envelo ing the
od ies of the Purkin je ce l ls ; e, superfic ia l stel late cells ; f , Go lg i cc of typeI I (see p . g, granule cells wi th their axons ascend ing and b ifurcatingin the mo lecular layer at i ; h, mossy fibers ; j , neuroglia cell ; m , neu
roglia cell ; n , cl im bing fibers . (A fter RamOn y Cajal .)
The gray matter Of the cerebel lum i s part ly superfic ial (thi si s the cortex to whi ch reference has al ready been m ade) andpart ly in the form of deep nuc lei em bedded within the wh itematter . The largest Of these deep gray centers are the dentate

THE VESTIBULAR APPARATUS AND CERE BE LLUM 19 1
nuc lei with in the cerebel lar hem i spheres . Within the verm i sare other sm aller centers , called the roof nuc lei , because they lieimmediately above the fou rth ventri c le (nuc lei embol iform i s,globosus, and fastigi i , see Fig. Som e of the afferent fiberswhi ch enter the cerebe l lum end i n these nuc lei
,but m ost of them
end in the cortex. The efferent fibers, on the other hand, ari se
from the deep nuc lei,espec ia l ly the dentate nuc lei (Fig.
The cerebe l lar cortex has three distinct layers . External tothe central whi te m atter (Fig. 89
,C) i s a wide layer composed of
very m inute granu le cel l s (Fig. 89,B ) densely crowded together ,
with scanty cytoplasm ,short
,c law- like dendrites
,and slender
unm yel inated axons whi ch ascend to the supe rfic ia l m olecu larlayer (Fig. 89
,A ) , where they bifurcate (thei r branches runn ing
lengthwise of the folium ) and end am ong the dendri tes of thePurkinj e cel l s
,to be desc ri bed imm ediately . The m i ddle layer
of the cortex i s composed of a single row of Purki nje cel l s (Fig.
89 , a) ; these have large gl obose bo d i es w i th m ass ive bushydendri tes di rected outward and s lender axons di rected inward .
These axons are mye l inated and constitute the chief efferent
pathway from the cortex ; they do not , however , leave the cerebel lum
,but end in the deep gray nuc lei (chiefly the dentate
nuc lei ) , from whi ch other neurons carry the impu lses out of thecerebel lum . The dendrites of the Purk inje cel l s are w idely ex
pand ed transversely to the length of the fo l ium ,but are very
narrow in the opposi te di rection ; thus each cel l comes intocontact wi th the largest possible num ber of axons of the
granu le cel l s whi ch run lengthwise of the folium . The outerm ost Or m olecu lar layer contains the dendrites of the Purk injecells
,term ini of the axons of the granu le cel l s and of other
fibers, and a sm all num ber of neu rons with short axons , am ong
whi ch are the basket cel l s i l lustrated in F igs . 16 and 89 , b.
Afferent fibers term inate in the cerebe l lar cortex in two ways .They may pass di rectly out to the m olecu lar layer as ascendingor c l im bing fibers
,where they end in very intim ate relati on with
the dendri tes of the Purk inje cel l s (F igs . 15 and 89 , n) , or theymay end as m oss fibers (Fig. 89
,h) am ong the cel l s of the gran
ule layer . Here the granu les take up the nervous impu l ses anddel iver them to the dendr i tes of the Purkinje cel ls . RamOn
y Caj al i s of the opini on that the m oss fibers are the term inals

192 INTR ODUCTION To N EU ROLOGY
of the afferent fibers of the i nferi or cerebel lar pedunc le, and thatthe ascending fibers are the term inals of the fibers from the
m i ddle pedunc le (brachium ponti s) .Since each fiber from the i nfer ior pedunc le branches exten
sively and reaches m any granu le cel l s in wi dely separated partsof the cerebel lum ,
and since the axon of each granu le cel l-reachesthe dendri tes of a very large number of Pu rk inje cel l s, a singleincom ing nervous impu l se may exc i te a very large num ber ofPu rkinje cel ls , and thus i ts physio logi cal effect may be greatlyenhanced . The same resu l t i s also secured by the action of thebasket cel l s (Fig. 89 , b) and other form s of neu rons with shortaxons with in the cortex (Fig. 89
,e, f ) , each of wh ich may
discharge powerfu l impulses di rectly upon several Pu rk injecells . The axons of the Purkinje cell s them selves also give Off
collateral fibers into the granu lar layer , whose neu rons d i scharge back into the Purkinje cel l s again . In al l of these .
ways provi sion i s m ade for the diffusion,summ ation
,and re
inforcem ent of stim u l i du ring the process of thei r transm i ss ionthrough the cerebel lar cortex, and also for prol ongation of m otorreactions whi ch wou ld otherwise soon subside
,and for the m ain
tenance of m uscular tone.
Thi s type of reacti on has been term ed avalanche conducti on
(see p. and i ts mechan i sm here i s sim i lar to that found inthe Olfacto ry bu lb (p. but m uch m ore complex. I t i s probable that the rec iprocal relat ion between the cerebel lum and thecerebral cortex i s Of a sim i lar sort, all cort ical activi ties exc i tingalso the cerebel lum and drawing therefrom additi onal nervousenergy as needed to m aintain the tone of the reacting m echanism ; and voluntary m ovements exc i ted by the corti co- spinalor pyram idal tract from the cerebral cortex (see p. 283 ) areunder espec ial ly di rect propri oceptive contro l from thi s source.
The relationships of the centers of the brain stem ,the cerebral
cortex, and the cerebel lum m ay be i l lustrated som ewhat c rudelyby the analogy of the three chief departments of the nationa l
governm ent. The reflex centers of the brain stem correspondto the legi s lative branch of governm ent
,determ in ing in advance
by virt ue of their i nnate structure what actions m ay appropriately be perform ed in each particu lar type of frequently recu rring situation. The cerebral cortex i s a sort of gl orified j udic i al

194 INTRODUCTION To N E UROLOGY
FE RRIER , D .
, and TURNER ,W . A . 1895 . A Record of Experiments Ill us
trative of the Sym ptomatology and Degenerat ions Fo llowing Lesions of theCerebellum , Phil . Trans . Roy. Soc. London for 1894, vo l . c lxxxv B , pp.
755—76 1 .
GEHUCHTE N , A . VAN . 1 904 . Le corps restiforme et les connexionsbulbO-cerebelleuses, Le Névraxe, vo l . vi .
1905 . Les ped oncules cerebelleuses superieurs, Le N évraxe, vo l . vu .
GO LDSTE IN ,K . 19 10 . Ueber d ie aufste igende Degeneration und Quer
schni ttsunterbrechung d es R ii ckenmarks (Tractus spino-cerebellaris posterior, Tractus spino-o livaris , Tractus spino-tha lam i cus) , Neurol. Centra lb latt
, No . 17 .
HE RRICK , C . JUDSON . 19 14 . The Cerebel lum of N ecturus and OtherU rodele Amphib ia
,J our . Comp . N eur .
,vo l . xxiv
,pp . 1—29 .
HE RRICK, C . L . 189 1 . Illustrations of the A rch i tecton i c of the Cerebellum , Jour . Comp . Neur . , vo l . 1, pp . 5—14 .
LEWANDOWSKY , M . 1907 . Die Funk tionen d es zentra len N ervensys
tems , Jena .
LU CIAN I , L . 1893 . Das Kleinh irn,Leipz i g .
RUSSE LL,J . S. RIE SEN . 1895 . E xperim enta l Researches into the Func
tions of the Cerebellum , Ph il . Trans . Roy. Soc . L ondon,vo l . c lxxxv B ,
pp. 819—86 1 .
SCHAPER , A . 1894 . Die morpho logische und h isto logische Entwi ckelungd es Kle inhi rns d er Teleost ier
, Morph . J ahrb .
, Bd . xxi .SHE RR IN GTO N ,
C . S. 1909 . On P lastic Tonus an d Propr iocep tive Reflexes , Quar t . Jour . E xp . Physio l .
,vo l . ii
,p . 109 .
WILSO N , J . GO RDON , and PIKE ,F . H. 19 12 . The E ffects of Stimulat ion
an d E xt irpat ion O f the Labyr inth of the Ear, and Their Relation to the
Motor System , Phi l . Trans . Roy. Soc . London,vo l . cc ii i B
, pp. 127—160 .

CHAPTER XII I
THE AUDITORY APPARATUS
THE hum an organ of hearing consists of the external ear,
bounded with in by the drum m embrane (tym pani c m embrane,m em brana tym pan i ) ; the m i dd le car
,a cavi ty fi l led with ai r
which comm uni cates with the pharynx through the auditory
Tympanic cavi ty, with chain of ossic lesSem icircular d uct
Utric leDuctus end olymphaticus
Ductus coch
Auricula
Aud itory tubeMembrana tympani
Recessus epitympanicusExternal acoustic meatus
Fig. 90.—Diagramm atic view of the parts of the hum an
Cunningham’s Anatomy.)
or Eustachean tube and contains the audi tory ossi c les ; and theinternal ear, a complex bony cham ber , the bony labyrinth ,within whi ch is them em branous labyr inth containing the specificreceptors or sensory su rfaces of the internal ear (Fig. The
1 95

196 INTRODUCTION To N EUROLOGY
tympani c membrane receives the ai r waves whi ch form the
physical stim ul i of sound (pp. 70 and These vibrations arethen transm i tted (and at the sam e
' time intens ified) by the au
d i tory ossi c les of the m i ddle ear to the l iqu i d within the bonylabyr inth .
Themembranous labyr inth i s of approxim ately the same shapeas the bony labyrinth , but sm al ler , so that there i s a space between the membranous labyrinth and the enc l osing bony wal l .Thi s space i s fi l led with l iqu i d
,the peri lymph , and the mem
branous labyrinth i s also fi l led with l iqu i d,the endolym ph . I n
Fig. 90 the per i lym phati c space i s pr inted in black and the endolymphati c space in white. The parts of the membranous labyrinth are shown diagramm ati cal ly in Fig. 9 1 .
Recessus utriculi
‘Ampul la of superior semicircular d uct
Ampul la of lateral d uctDuctus reuniens Crus commune
Ductus end olymphaticus Ductus utriculosaccularisAmpull
Saccus end o
Fig. 9 1 .—Diagrammatic representation of the parts of the membranous
labyrinth . (From Cunn ingham’s Anatomy. )
Them embranous labyrinth is a c l osed sac whi ch has four chiefparts : ( 1 ) the utri c le ( recessus utri cu l i ) , wi th a patch of sensoryepi thelium , the m acu la utri cu l i ; (2) the three sem i c i rcu lar canals(ductus sem ic i rcu lares ) , each of whi ch comm uni cates at bothends w i th the utri c le and has at one end a di lation (ampu l la)containing a patch of sensory epi thelium , the c ri sta ; (3) thesaccu le (saccu lus) connected by a narrow ductus utriculosaccularis with the utri c le and contain ing a patch Of sensory epithe
l ium , the m acu la sacculi ; (4) the ductus cochleari s, whi ch com
muni cates by a narrow ductus reun iens with the saccu le and i sSpi ra lly wound to fit the bony cochlea
,whi ch i s shaped like a snai l
shell . The ductus cochlearis (old name,scala media) i s trian

198 INTRODUCTION TO N EUROLOGY
sensory epitheli um , part of whose cells are supporting elements of d iversesorts and part (the hair cells) are specific receptors . The term ini of thecoch lear nerve arbor ize around the bod ies of the hair cel ls in the same waythat fibers of the vestibular nerve are related to the hai r cel ls of the cristzn
of the semicircular cana ls (Fig. 32, p. The m embrana tectoria isa del i cate gelat inous mass resting upon the sp ira l organ and in timatelyconnected w ith the hairs of the hair cel ls . Its shape has been very carefully stud ied by Hardesty.
Many detai ls of the structure of the spiral organ, or organ of Corti , andthe whole quest ion of the mode of its funct ioning are st i ll controverted .
Our present kn owledge of the h isto logica l organ izat ion of the bas ilar m em
brane shows that it is structural ly in capab le of serving the funct ion of toneana lysis in the way postulated by Helm holtz ’s theory. Based upon im
portant add itions to our know ledge of the minute structure of the organ
ep. s.sp.
Fig . 94 .
—Section through the apica l turn of the coch lea of the pig atabou t ful l term , show ing outer aud i tory hairs embedded in the membranatectoria : E pithe l ium of spira l sulcus ; inner hai r cel ls ;inner pillar ; m .bas .
, basement membrane ; m . isel . , membrana tectoria ; lab.
vest. , lab ium ves t ibulare ; n .each ., coch lear nerve ; outer ha ir -cel l ; s .sp.
,
sulcus spiralis . (A fter C . W . Prentiss . )
of Corti and cl in ica l observations upon d iseased cond i t ions, several d ifferenttheor ies of the mechan ism of tone analys is have recently been expressed .
Among the more important of these researches are those of ‘Shambaugh .
Th i s author has demonstrated that the ha irs of the ha ir cel ls do not terminate freely in the endolym ph , as commonly figured
,but that they are
firm ly attached to the under surface of the tector ial membrane . Th ismembrane has a semigelat inous texture and is capab le of taking up sympathet i ca l ly the v ibrations of the endo lym ph wi th in wh i ch it floats .The developm ent of the tectorial membrane has recentlybeen restud ied by
Prent iss and Hardesty . I t first appears as a thin cut i cular plate developedover the free ends of the co lumnar cel ls wh i ch form the inner or axia l par t ofthe ep i thel i um of the basement membrane. In the adu lt ear i t reta ins itsattachment to the lab ium vest ibulare along the axial border of the duc tuscoch leari s, but becomes free from the cel ls wh ich form the lin ing of the

THE AUDITORY APPARATUS 199
sulcus sp ira lis (Fig . Prentiss claim s that it is in part formed from theembryoni c cell s wh i ch develop into the spira l organ, and that its connectionw i th the spira l organ is reta ined in the adult (Fig. butHardesty ( 1 9 15 )says that the cel ls of the embryoni c spiral organ contribute li tt le or noth ingto the format ion of the tector ia l membrane and that thi s membrane is freefrom the spira l organ in the adul t . Prentiss describes the membrane as
grow ing in th i ckness by the secret ion of a cuticulum formed between theends of the epi thel ial cells
,thus giv ing to the mature membrane a cham
bered or honey- comb structure. Hardesty,however
,regards i t as produced
by fibri ls growing out from the free ends of the epitheli al cel ls lying between the embryoni c spira l organ and the axis of the cochlea, these fibrilsbeing embedded in a gelatinous matrix .
Shambaugh concludes that the tectorial membrane takes the part of aphys ical resonator by respond ing in i ts var ious parts to tones of d ifferentpitch , depend ing on the si ze of the membrane, tones of hi gher
pitch being
taken up by the hair cells located near the beginn ing of the basa co il , thoseof lower pi tch by the cel ls near the apex of the coch lea, where the tectorialmembrane attains its maxim um size. The stimu lation of the ha ir cells iseffected only through the med ium of their projecting hairs, these beingexcited by vibrations of the tectorial membrane to wh ich they are attached .
In fishes the organs of the interna l ear are intimately associated w i th anextens ive ser ies of subcutaneous canals conta ining num erous sense organsand w i th naked cu taneous sense organs of the same type, the entire complexforming the system of latera l line sense organs (see p . 1 10 and Fig.
The nerves whi ch in fishes supply the latera l l ine sense organs ( lateralisroots of the VII and X cran ia l nerves) and the organs of the internalear (VIII nerve) are intimately associated and terminate together in the
acoustico- lateral area of the m ed ulla ob longata (Figs . 43 and 44 , pp. 1 1 1 ,and a l l of these end -organs have the same type of structure as those
of the hum an interna l ear (Fig . 32, p .
The interna l ears of fishes are essent ia lly s imilar to those of m an savethat they lack the coch lea and the organ of Corti . They posses s a smallsense organ in the saccule, the lagena, supplied by a Special branch of theVII I nerve (Fig . 95 , RL ) , from wh ich the coch lea of h igher vertebrates hasbeen developed . The researches of Parker have shown that fishes hear
,
though there is no ev idence that they’ possess the power of tone analys is ,and the sense organs of the saccule are the essentia l receptors for soundwaves . The sense organs of the lateral line system are said by Parker to besens itive to water vibrations of s lower frequency than the sound waves towh ich the ear responds , wh ile Hofer is of the opinion that these organs are
st imulated only by stream ing movements of the water in wh i ch the animalsl ive. Probab ly the lateral l ine organs a lso part icipate in the equi libratoryreactions of the fish .
Though our knowledge of the functions of the various parts of the acoustico—latera l system of fishes is st i l l very im perfect
,it is ev ident that a ll of
these organs are both structura lly and phys io logica l ly of common type, andi t is probab le that they have had a common evo lut ionary origin from a moregeneralized form of cutaneous tact i le organ . Th is is the exp lanation of theintimate associat ion in the human ear of sense organs of so d iverse functionsas the coch lea for hear ing and the semicircular cana ls for equil ibrat ion
, the
form er being an exterocep tor whose react ions may be v iv id ly consc ious , andthe latter being a proprioceptor whose react ions are a lm ost entire ly unconsciously performed . For further cons ideration of the semicircular canalsand their centra l connections see p. 183 .

200 INTRODU CTION TO NEUROLOGY
In the human body the cochlear and vestibu lar nerves arevery intim ately assoc iated , but the embryologi cal studies of
Fig. 95 .—Diagram of the acoustico- lateral system of nerves w ith their
peripheral end -organs, as seen from the right Si de, in a fish , the commonSilver- s ides, Menid ia (X The relations here figured were reconstructed from serial sect ions by project ion upon the sagittal p lane. Forthe relat ions between the acoustico—lateral nerves and the other systemsof nerves in th is fish
,see the more deta iled chart from wh i ch thi s was drawn
off,in the J ournal of Comparative N euro logy, vo l . ix, 1899 , plate 1 5 ; cf .
also Fig . 65,p . 149
,of th is book . The dotted outline represents the posi
t ion of the bra in ,the lateral l ine canals are shad ed w i th cross-hatch ing, the
internal ear is stippled , and the nerves are drawn in b lack . The organsof the lateral line system are drawn as b lack d isks when naked on the
surface of the sk in , and as b lack c ircles when lying in the canal s . NAA ,
Anterior nasal aperture ; NA P , posterior nasal aperture ; N 0 L ,o lfactory
nerve ; N OPT,Optic nerve ; RAA , nerve of superior ampulla ; RAE , nerve
of latera l ampul la ; RAP , nerve of inferior ampulla ; R BUC,ramus bucca
lis of facia l nerve ; RL , nerve of the lagena (rud imentary spira l organ) ;R LAT, ramus lateral is of the vagus ; R 08 ,
ramus o hthalm icus superficialis of the facia l nerve ; R MAN EX,
ramus mand i ularis externus ofthe facial nerve ; R SAC, nerve of the sacculus ; RU , nerve of the utri culus ;T
, acoustico—lateral area . (After Herrick , from Wood ’s Reference Handbook of the Med i ca l Sciences .)
Streeter and others have m ade i t pl ain that these two nerves arereal ly m ore distinct than was formerly supposed . The periph

202 INTRODU CTION To N EUROLOGY
sensory spots in the saccu le, dem onstrates the relatively late
phylogeneti c origin of the cochlear system from the vest ibu lar ,and has suggested to som e physio logi sts that even in man thesetwo system s are not whol ly d istinct , and that the sense organs inthe saccu le may also function as a sound receptor . I t i s c lear ,however
,that tone analysi s i s effected only in the cochlea .
The central connections of the cochlear and vestibu lar nervesare fundamental ly different . The vestibu lar nerve term inatesin reflex centers of the medu l la oblongata and cerebel lum (p.
185 ) wi th no important cortical connecti ons , whi le the coch learnerve has , i n addition to important reflex connections in the oblongata and m i dbrain,
the m uch stronger ascending pathway Ofthe lateral lemni scus di rectly to the m edial gen i cu late body ofthe thalamus, and thence to the temporal lobe of the cerebralcortex (see p. 157 and F igs . 75 , 77 , 80, Som e of the fibersof the lateral lemni scus are interrupted in the infer i or co l l icu lus,whi ch is an im portant auditory reflex center .Summary.
—The hum an ear has three parts : (1 ) the externalear
,for rece iving sound waves from the ai r ; (2) the m i ddle ear,
for intensifying the vibrations and transm i tting them to (3) theinternal car
,whi ch i s filled with l iqu id and contains sense organs
of uncertain functi on in the utri c le and saccu le,sense organs for
equ i librat ion in the sem ic i rcu lar canals,and the Spi ral organ
(organ of Corti ) i n the cochlea for tone analysi s . The spi ralOrgan i s a compli cated epi thel ial structure resting on a basem entm embrane and consi sting of supporting cel l s of diverse k inds,the hai r cel l s (which are the spec ific receptors and receive theendings of thefibers of the cochlear nerve) , and the tectoria l m em
brane. Sham baugh i s of the opin i on that the tectorial m em branei s capable of responding in i ts various parts to different vibrationfrequenc ies, and that the hai r cel l s are stim u lated through thei rhai rs whi ch are attached to the tectorial m em brane.
I n fishes the organ of hearing i s m uch simpler than in man,the
sem ic i rcu lar canals are, however , sim i lar , and there i s , in addition, an elaborate system of lateral l ine sense organs whose functions seem to be intermediate between the tacti le and audi toryorgans . It i s probable that these three system s of sense organswere der ived phylogenetical ly from some m oregeneral i zed formof cutaneous tacti le organ. This accounts for the intim ate as

THE AUDITORY APPARATU S 203
soc i ation in the hum an ear of organs of so diverse functions as thesem i c i rcu lar canals and the cochlea .
The central connecti ons of the vestibu lar and cochlear nervesare very different , the form er effecting chiefly reflex connectionsfor equ i l ibration in the m edu l la ob longata and cerebel lum ,
and
the latter both reflex connections in the brain stem and cort icalconnections through the lateral lem ni scus
,m edial gen i cu late
body Of the thalam us and auditory radiations, for consc i oussensations of hearing.
IJ TE RATU RE
EWALD, J . R . 1892 . Phys io logische Untersuchungen uber das Endorgan d es Nervus octavus
,W iesbaden
,J . F . Bergmann .
HARDESTY,I . 1908 . On the N ature of the Tector ia l Membrane and Its
Probab le ROle in the Anatomy of Hear ing,Am er . Jour . Anat.
,vo l . vi ii .
19 15 . On the Proport ions,Development , and A ttachm ent of the
Tectoria l Membrane, Amer . Jour . An at. , vo l . xv i ii .VON HE LMHO LTZ , H. L . T. 1896 . Die Lehre von d en Tonempfind ungen,
Ausgabe 5 , Braunschweig .
HO FE R,B . 1908 . Stud ien uber d ie Hauts innesorgane d er Fische,
Berichte kgl . Bayerischen Bio logischen Versuchsstation in Miinchen,Bd . 1 ,
p. 1 15 .
KA PPE RS, C . U . ARIE NS. Kurze Skizze d er Phylogenetischen Entw icklung d er Oktavus und Lateralisbahnen mit Beri i cksichtigung d er neuestenE rgebnisse
,Zentralbl . f . Phys io l ., B d . 23 , 1909 .
PARKE R,G . H. 1903 Hearing and A l l ied Senses 1n Fishes , U . S. Fish
Commiss ion Bulletin for 1 902 , Wash ington .
1903 . TheSense ofHearing In Fishes , Am er . N atura li st, vo l . xxxvn .
1905 . The Func tion of the L ateral L ine O rgans in Fishes , Bul . of theBureau of Fisheries for Wash ington .
PRE NTISS,C . W . 19 13 . On the Development of the Membrana Tectoria
w i thIéteference to I ts Structure and A ttachm ents , Amer . Jour . Anat ., vo l .
xha o . 4 .
RETZIUS, G . 1884 . Das G ehororgan d er W irbelt iere , Stockholm .
SCHON EMANN , A . 1904 . Die Topograph ie d es mensch li chen G ehoror
ganes , VVu sbad en .
SHAMBAU GH , G . E . 1 907 . A Restudy of the M inute Anatomy of Struetures in the Cochl ea w ith Conclus ions Bear ing on the So lu t ion of theProb lem of Tone Perception
,Am . Jour . Anat . , vo l . vn .
1908 . The Membrana Tectoria and the Theory of Tone Perception ,
Arch . O to logy, vo l . xxxv i i .
19 10 . Das Verhaltn is zw ischen d er Membrana Tectoria und d emCort ischen O rgan , Zelts . f . Ohrenheilk .
,Bd . 62 .
Bd
.—6
. 19 12 . U eber d en Bau und d ie Funkt ion d er Crista Am pullaris , Ib id .,
5 .
STRE ETE R,G . L . 1907 . On the Development of the Membranous Laby
rinth and the A coust ic and Facia l N erves in the Human Embryo , Am er .Jour . Anat . , vol . v i .WATSON , J . B . 1 9 14 . Behav ior , An Introduct ion to Comparative Psy
chology, New York , Chapter XII .
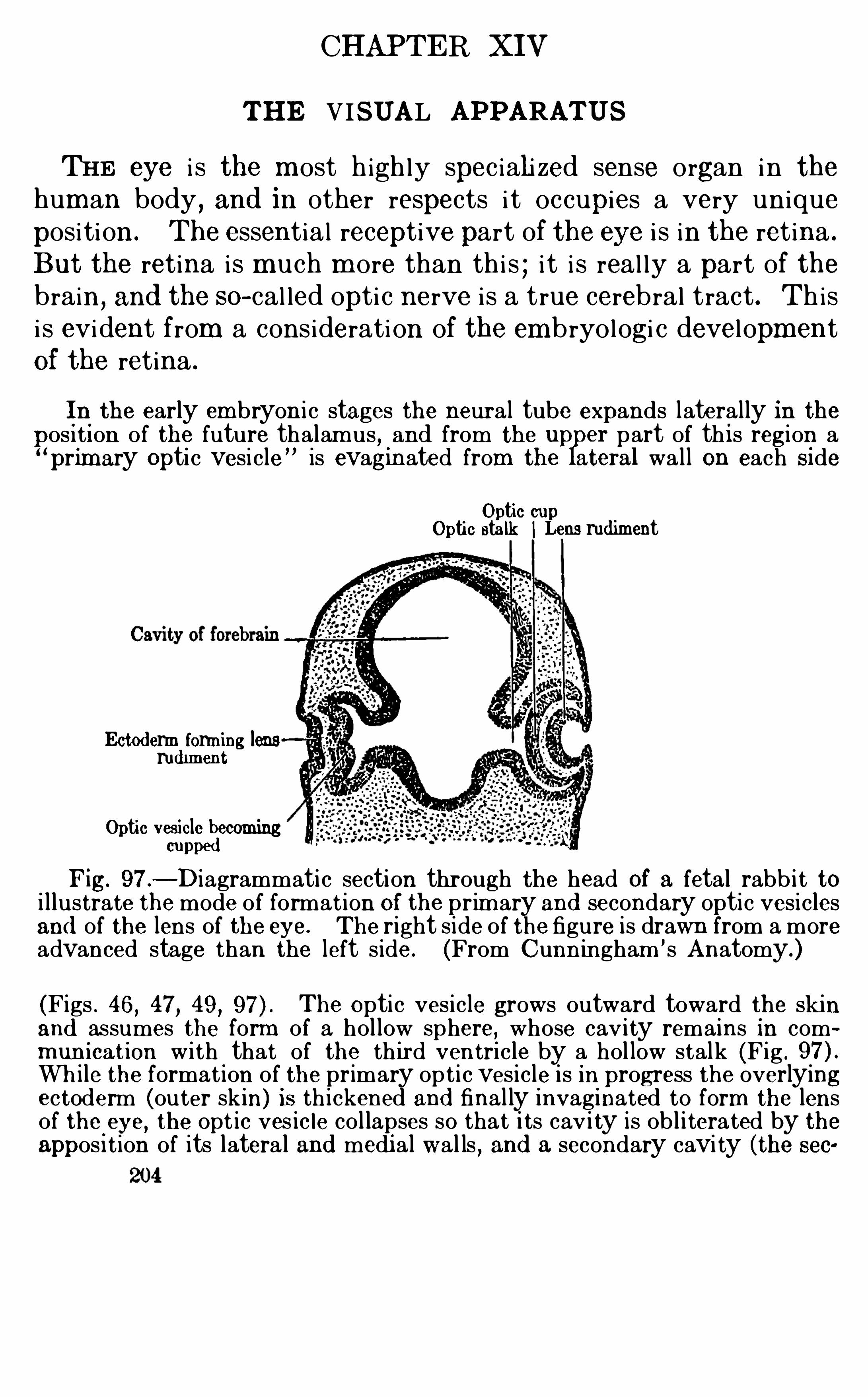
CHAPTER XIV
THE VISUAL APPARATUS
THE eye i s the m ost h ighly spec iali zed sense organ in thehuman body , and in other respects i t occupies a very unique
positi on . The essential receptive part of the eye i s in the retina .
But the retina i s m uch m ore than thi s ; i t i s real ly a part of thebrain
,and the so- cal led opti c nerve i s a true cerebral tract . Th is
i s evident from a considerati on of the embryologi c devel opmentOf the retina .
In the early embryon i c stages the neural tube expands lateral ly in thepositi on of the future thalam us, and from the up
lper part of th i s region a
primary Optic vesi cle” is evaginated from the ateral wa l l on each Si de
Optic cupOptic stalk ILens rud iment
Ectod erm form ing lensrud iment
Optic vesicle becomingcupped
Fig. 97 .—Diagrammatic section through the head of a fetal rabb i t to
i l lustrate the mode of format ion of the prim ary and secondary optic vesi clesand of the lens of the eye. The right side of the figure is drawn from a moreadvanced stage than the left s ide. (From Cunningham ’s Anatomy.)
(Figs . 46 , 47, 49 , The Optic ves icle grows outward toward the sk inand assumes the form of a hol low sphere, whose cav ity remains in commun ication w ith that of the th ird ventricle by a hol low stalk (Fig .
Wh i le the format ion of the primary op ti c ves i cle is in progress the overlyingectoderm (outer skin) is th i ckened and final ly invaginated to form the lensof the eye, the opt ic ves ic le co l lapses so that i ts cav i ty is ob l i terated by theappos i tion of its latera l and medi al wal ls , and a secondary cavi ty (the sec
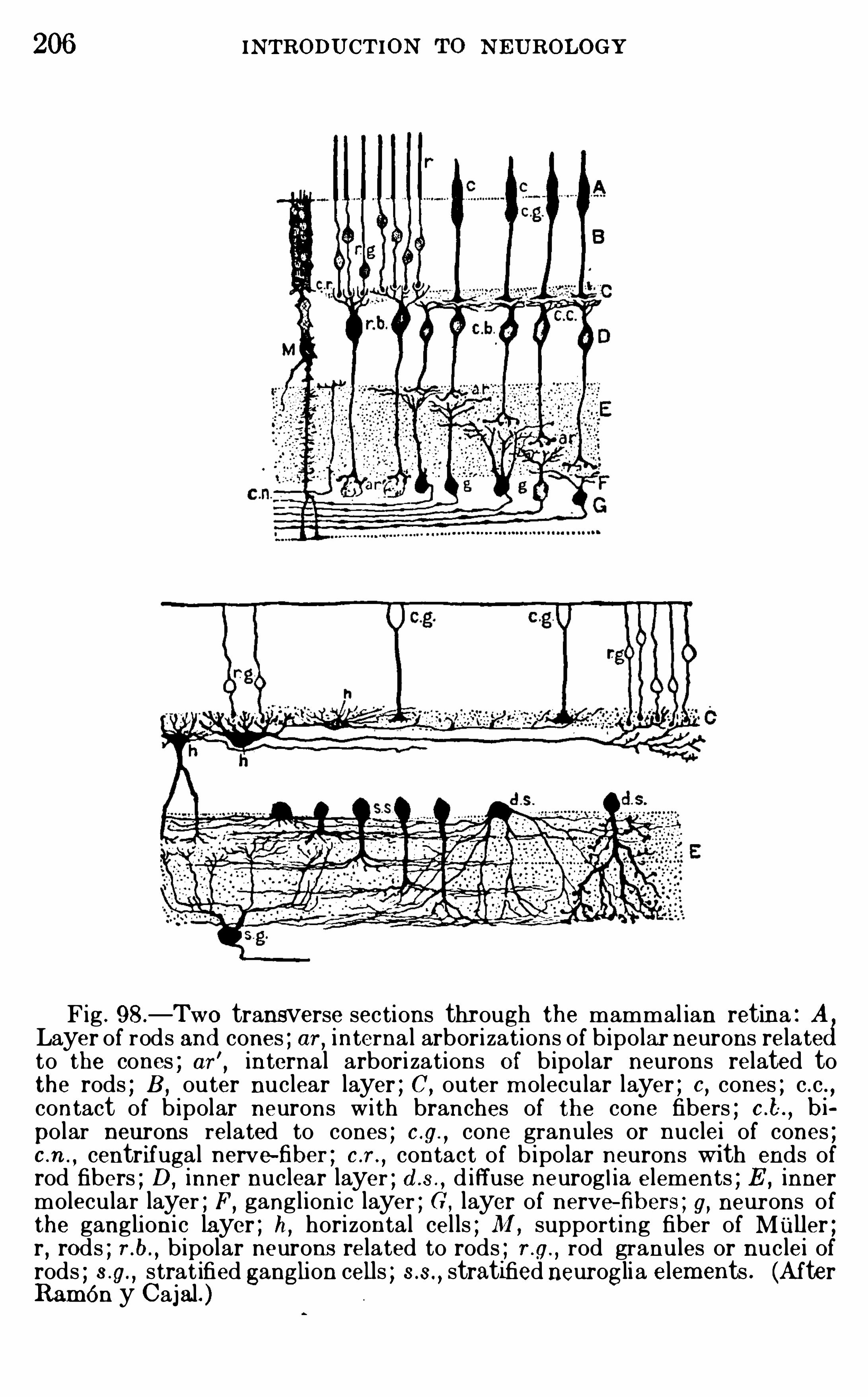
206 INTRODU CTION N EU ROLOGY
Fig . 98.—Two transverse sections through the mammal ian retina : ALayer of rods and cones ; or , interna l arborizations of b ipo lar neurons related
to the cones ; ar', internal arbori zations O f b ipo lar neurons related tothe rods ; B ,
outer nuclear layer ; C,outer m olecular layer ; 0 , cones ; c .c . ,
contact of b ipolar neurons w i th branches of the cone fibers ; c .t bi
po lar neurons related to cones ; c .g.
,cone granules or nuclei of cones ;
c .n . , centrifugal nerve-fiber ; c .r .,contact of b ipo lar neurons with ends of
rod fibers ; D,inner nuclear layer ; d .a.
,d iffuse neurog l ia elements ; E ,
innermolecular layer ; F,
ganglioni c layer ; G , layer of nerve-fibers ; g, neurons ofthe gangli oni c layer ; h, horizontal cel ls ; M ,
support ing fiber of Muller ;r , rods ; r .b.
, b ipolar neurons related to rods ; r .g.,rod granules or nuclei of
rods ; s .g., strat ified ganglion cells ; stratified neuroglia elements . (After
Ram én y Cajal. )
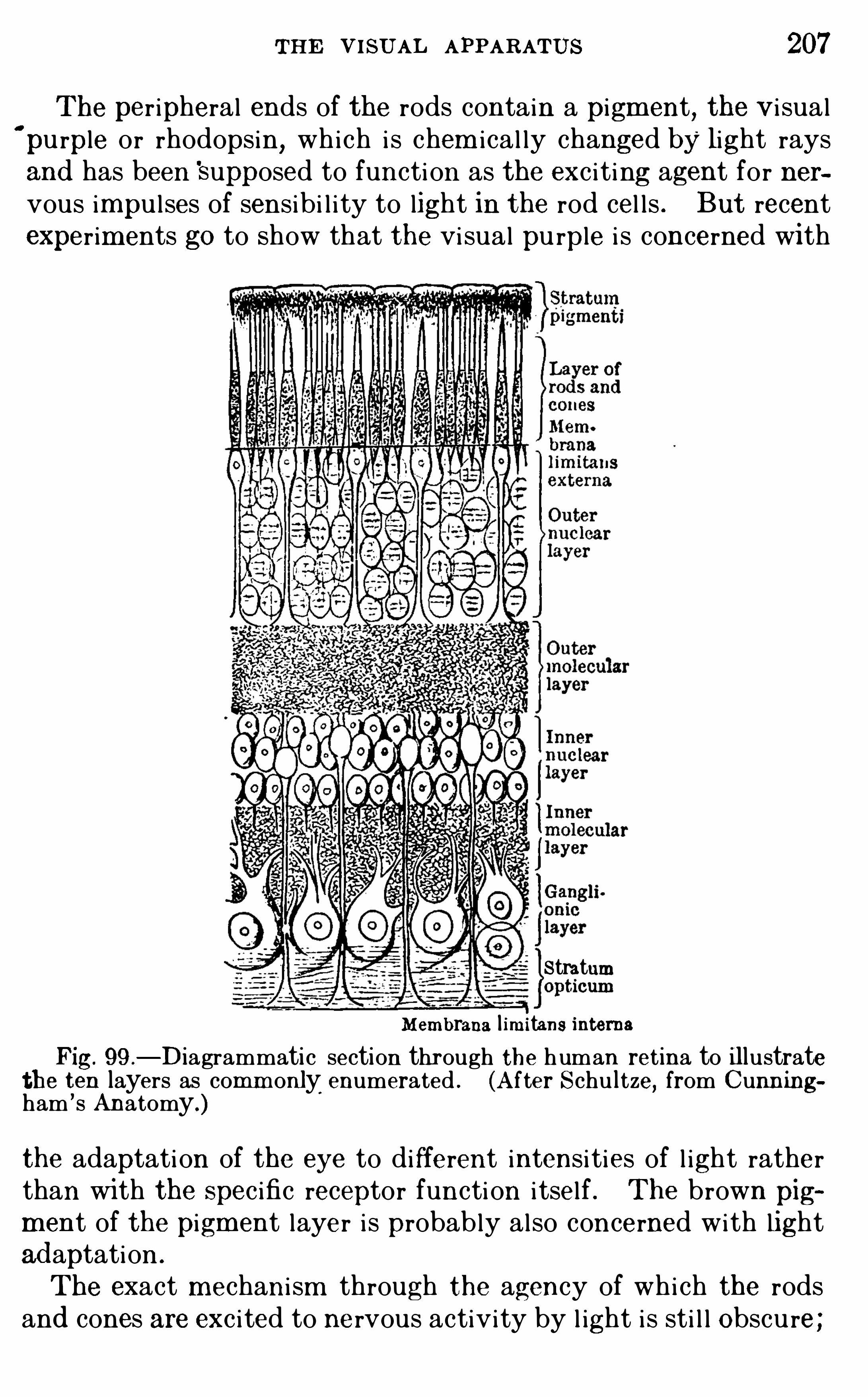
THE VISU AL APPARATUS 207
The per ipheral ends of the rods contain a pigm ent, the V i sual
‘
pu rple or rhodopsin, which i s chem ical ly changed by light raysand has been Supposed to functi on as the exc i t i ng agent for nervous impu l ses of sensibi l i ty to light in the rod cel l s . But recentexperim ents go to Show that the vi sual pu rple i s concerned with
StratumMembrana l im itans interns
Fig. 99 .
—Diagrammatic section through the human retina to il lustratethe ten layers as comm only
,
enumerated . (A fter Schultze, from Cunn ingham ’s Anatomy.)
the adaptati on of the eye to different intensi ties of l ight ratherthan wi th the Spec ific receptor functi on i tsel f . The brown pigm ent of the pigment layer i s probably also concerned with lightad aptation .
The exact mechani sm through the agency of whi ch the rodsand cones are exc i ted to nervous activity by l ight i s sti l l Obscu re ;
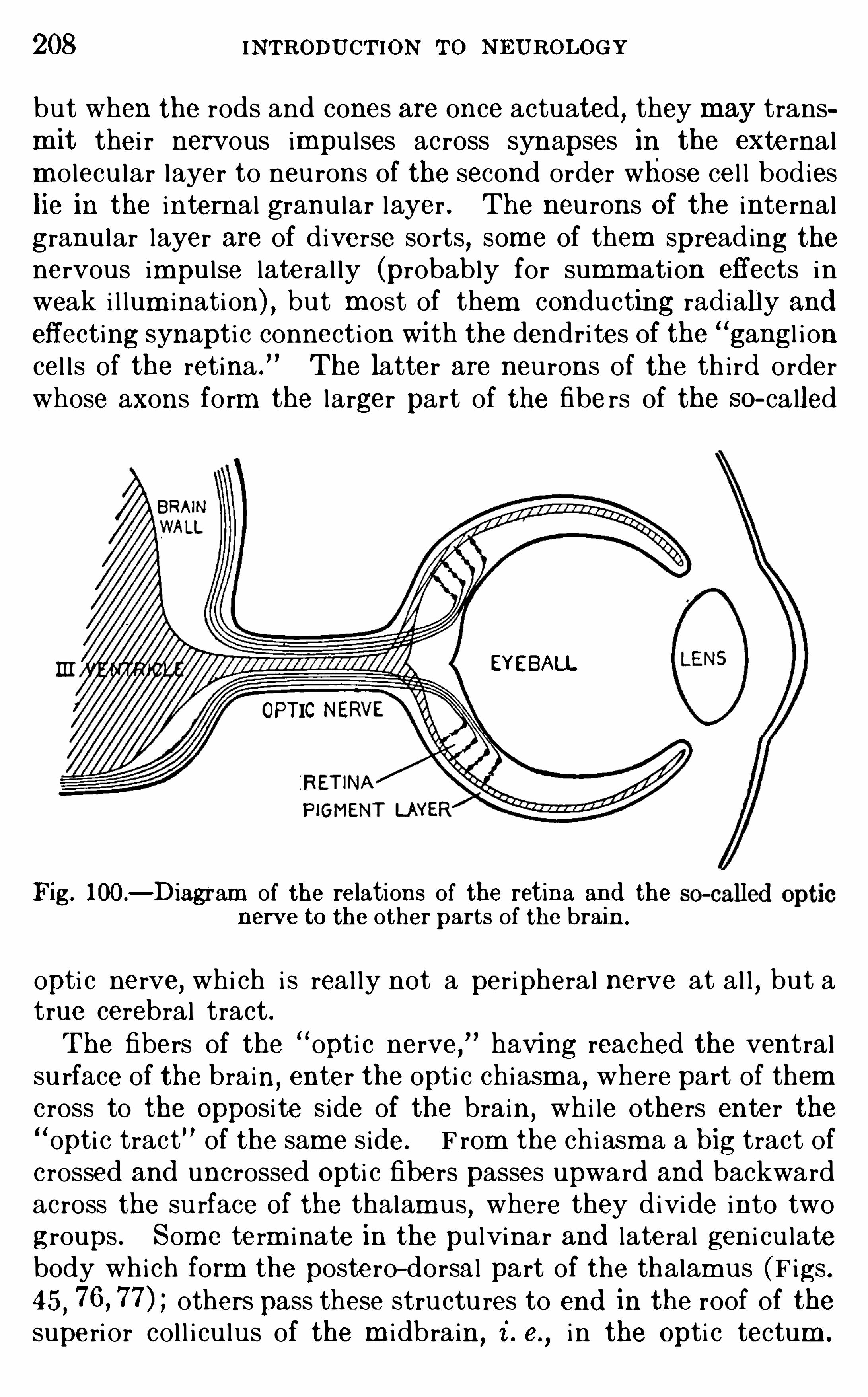
208 INTRODU CTION To NEUROLOGY
but when the rods and cones are once actuated , they may transm it thei r nervous impu l ses ac ross synapses in the externalm olecu lar layer to neu rons of the second order whose cel l bodieslie in the internal granu lar layer . The neu rons Of the internal
granu lar layer are of diverse sorts, som e of them spreading thenervous impu l se lateral ly (probably for summ ation effects inweak i l lum inati on) , but m ost of them conducting radially andeffecting synapti c connection wi th the dendri tes of the
“gangl i on
cel l s of the retina .
”The latter are neurons of the th i rd order
whose axons form the larger part of the fibe rs of the SO- called
EYEBALL
OPTIC NE
Fig. 100—Diagram of the relations of the retina and thenerve to the other parts of the brain .
opti c nerve, whi ch i s real ly not a peripheral nerve at al l , but atrue cerebral tract .The fibers of the “opti c nerve, having reached the ventral
su rface of the brain, enter the opti c chiasma , where part of themc ross to the Opposi te Side of the brain
,whi le others enter the
“opti c tract
” of the sam e s ide. F rom the chi asm a a big tract ofc rossed and uncrossed Opti c fibers passes upward and backwardacross the su rface of the thalam us
,where they divide into two
groups . Some te rm inate in the pu lvinar and lateral geni cu latebody which form the poste ro- dorsa l part of the thalam us (Figs .45
, others pass these structu res to end in the roof of thesuperi or col l i cu lus of the m i dbrain,
i . e.,i n the Opti c tectum .

210 INTRODU CTION TO NEUROLOGY
Gen i cula
Fig. 101 .

THE VISU AL APPARATUS 2 1 1
l obe of the fish brain (F igs . 43 , Here vi sual impressionsc are brought into physiologi cal relations with those of the tactualand auditory system s received by the lemn i sc i . The chief efferent pathway from th is center i s by way of the under lying cerebral pedunc le (Fig. Here reflex connections are effecteddi rectly with the nuc lei of the I I I and I V cran ia l nerves for theeye m usc les , and through the fasc i cu lus longi tudinali s mediali swith the centers for al l other cranial and Spinal m usc les . Th isfasc i cu lus i s a strong bundle composed of both descending andascending fibers whose functi on i s the general coordinati on ofreflex m otor responses, and in parti cu lar those of the conj ugatem ovem ents of the two eyes (see p.
The accomm odation of the eye for distance i s effected bychanges in the cu rvature of the lens, and the adaptation fordifferences in i l lum ination i s effected in part by changes in thediameter of the pupi l (th is i s in addi tion to the changes in theretinal pigm ent referred to on p. 207 and to changes in the rodsand cones and other neurons of the retina which may be exc i tedby the centr ifugal fibers from the brain to the retina referred toon p. The nerves contro l l ing the m ovem ents of the lensand the pupi l lary reactions bel ong to the vi sceral m otor system .
They leave the central nervous system in part through the oculom otor nerve and in part (for di lati on of the i r i s) from the l owercervi cal region of the spinal cord . The latter fibers pass by wayof roots of spinal nerves into the superior cervi cal sympatheti cgangl i on (p. 234 and Fig. 41
, p. 107) and then forward to theeyebal l . We cannot here enter into further detai l s of the mechanism of accomm odation or of the d iopteric apparatus and theaccessory parts of the eye ; see the larger text- books of anatom yand physiology .
The thalam i c connections of the opti c tracts in the l owestvertebrates are very insignificant, col laterals of these fibers
Fig. 101 .—A d iagram of the visual tract, i l lustrating the s ignificance of
the partial decussat ion of nerve—fibers in the op ti c ch iasma so as to ensurethe representation in the cerebral cortex of nervous im pulses excited by ohjects on the opposite half of the body only. III O cul omotor nerve ; L ,
med ial lemni scus ; M , mammillary bod ies ; RN , red nucleus (nucleus ruber) ;SN , _b lack substance (substantia ni gra) TG , opt ic tract to corpora quadr igem ina (cf. Fig. (From Starr ’s Nervous Diseases .)
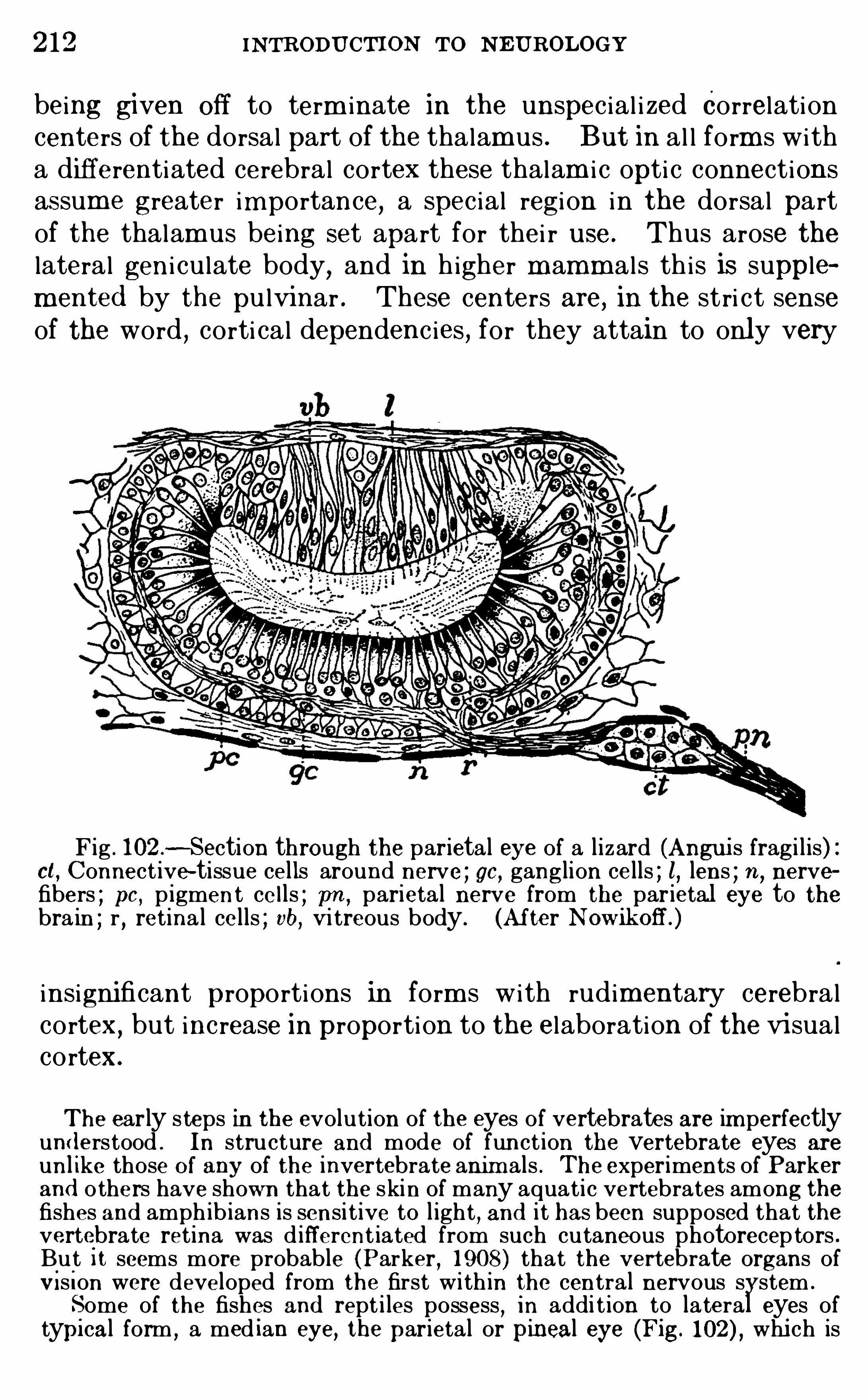
2 12 INTRODU CTION To N E UROLOGY
being given off to term inate in the unspec ial i zed Correlationcenters of the dorsal part of the thalam us . But in al l form s witha differentiated cerebral cortex these thalam i c opti c connectionsassum e greater importance, a spec ial regi on i n the dorsal partof the thalam us being set apart for thei r use . Thus arose thelateral geni cu late body, and in h igher m amm als th is is supplem ented by the pu lvinar . These centers are, in the stri ct senseof the word , corti cal dependenc ies , for they attain to only very
vb I
Fig . 102—Section through the parietal eye of a l izard (Angui s fragi l is) :ct, Connect ive-tissue cells around nerve ; gc, ganglion cells ; l, lens ; 71 , nervefibers ; pc , pigment ce l ls ; pn ,
parietal nerve from the parietal eye to thebra in ; r , retina l cel ls ; vb, Vi treous body. (Af ter
insignificant proportions in form s with rudimentary cerebralcortex, but increase in proportion to the elaboration of the vi sualcortex.
The early steps in the evolution of the eyes of vertebrates are im perfectlyund erstood . In structure and mode of funct ion the vertebrate eyes areun l ike those O f any of the invertebrate anim als . The experiments of Parkerand others have shown that the skin of many aquati c vertebrates among thefishes and amph ib ians is sensi t ive to l ight
, and it has been supposed that thevertebrate retina was d ifferent iated from such cutaneous photorecep tors .Bu t 1t seems more probable (Parker , 1908) that the vertebrate organs ofV i s ion were deve loped from the first wi th in the central nervous s stem .
Some of the fishes and rept i les possess , in addi t ion to lateral,
eyes oftypi cal form , a med ian eye, the parietal or pineal eye (Fig. whi ch is

214 INTRODU CTION To N EUROLOGY
N U E L. J . P . 1904 . La Vis ion, Bibl iotheque Internationa le d e PsychologieExpérim enta le Normal et Patho logique, Paris .PARKE R , G . H. 1908 . The O rigin of the Lateral Eyes of Vertebrates ,
Am er . Nat .,vo l . xli i , pp. 601—609 .
1909 . The Integum entary N erves of Fishes as Photoreceptors andTheir Significance for the Origin of the Vertebrate Eyes, Am er. Jour . ofPhys io l . , vo l . xxv,
pp . 77—80.
RAMON Y CAJAL, S. 1894 . Die Retina d er Wirbeltiere, W iesbaden .
SCHAF E R,E . A . Text-book of Phys io logy
,vol . i i
,pp . 752—76 1 , 1026
1 148 .
VINCENT,S. B . 19 12 . The Mamm al ian Eye, Jour . An im a l Behavior,
vo l . i i,pp . 249—255 .
WATSON , J . B . 19 14 . Behav ior, an Introduction to Comparative Psychology, New York. Chapter XI .

CHAPTER XV
THE O LFACTORY APPARATUS
THE olfactory part of the brain as a whole i s som etimes cal ledthe rh inencephalon . In fishes (p. 1 12 and F igs . 43 , 44) alm ost thewhole of the cerebral hem i sphere i s devoted to thi s function,
and
as we pass up the scale of an imal l ife m ore and m ore non—O lfactory centers are added to the hem i sphere in the corpus str iatum and cerebral cortex, unti l in m an thenon-O lfactory part ofthe hem i sphere overshadows the rh inencephalon . The complexform of the hum an cerebral hem i sphere cannot be adequatelyunderstood apart from a knowledge of thi s evolut ionary h i story,whi ch has been studied with great care by comparative neurolo
gi sts. Them etam orphosi s of the vertebrate cerebral hem i spherefrom a simple olfacto ry reflex apparatus in the lower fishes to the
great organ of the higher m ental processes upon whi ch al l hum ancu lture depends i s a very dram ati c hi story, i nto which , unfortunately, we cannot here enter .Sm el l i s evidently the dom inant sense in m any of the l ower
vertebrates . That thi s i s the case in the d ogfish i s shown by theenorm ous devel opm ent of the O l factory centers of the brain, towhi ch reference has j ust been m ade . And in m ost of the labo ratory m amm als
,such as the rat and the d og, the sense of sm el l
sti l l plays a very m uch m ore important part in the behaviorcomplex than in m an and other primates, whose o lfactory organsare in a reduced condition .
The nervus term inalis is a Slender ganglionated nerve found associatedwi th the o lfactory nerve in most classes of vertebrates from fishes to m an .
Its fibers,wh i ch are unmyel inated , reach the m ucous membrane of the
nose, though the precise method of their ending is unknown . They passinward in company w i th those of the o lfactory nerve as far as the o lfactorybulb . Here they separate from the o lfactory fibers and enter the cerebra lhemisphere between the attachment of the olfactory bulb and the lam inaterm inalis (Fig. 43 , p . W i th in the bra in they have been fo llowedbackward through the entire length of the O lfactory area and hypothalamus ,but their cerebra l connect ions have never been accurately determ ined .
The function of th is nerve is likewise who l ly unknown .

216 INTRODU CTION To NEUROLOGY
The O lfactory cerebral centers fal l into two groups : ( 1 ) thereflex centers of the brain stem and (2) the olfactory cerebralcortex. The arrangements of the olfactory reflex centers andthei r connecting tracts are essentia l ly sim i lar in plan in al l vertebrate brains (except in som e aquati c m amm als, l ike the dol
ph in, which lack olfactory organs altogether) . The olfactorycerebral cortex
,on the other hand , i s very diversely devel oped
in different groups of vertebrates . There i s no true cerebralcortex in fishes ; in amphibians (parti cu larly in the frog) theolfactory cerebral cortex begins to emerge from the general
Fig.103 . Dissec t ion of the right O lfactory bulb and nerve on the
lateral wa ll of the nasa l cav i ty. (From Wood’ s ReferenceHandbook of the
Medi cal Sciences )
o lfactory reflex centers ; in repti les there i s a wel l- fo rm ed olfac
tory cortex of simple hi stologi c pattern and the beginnings of thenon—O lfactory cortex ; in bi rds the O l facto ry apparatus i s reducedand the non—olfactory cortex i s som ewhat m ore extensive thanin repti les ; in m amm als both the olfactory cerebral cortex and
the non-olfactory cortex attain thei r m aximum dim ensions, theformer in the l owest m em bers of thi s group and the latter in thehighest .The cerebral cortex as a whole i s som etim es called the
pallium . That portion of the pal l ium whi ch i s related with theo lfacto ry apparatus was d ifferentiated earl ier in vertebrate evo

2 18 INTRODU CTION TO N E U ROLOGY
termed a glomeru lus, whi ch also receives one or m ore dendri tesfrom the Olfactory neurons of the second order , or m i tral cel l s .The gl omeru lus
,therefore
,contains the fi rst synapse i n the ol
factory pathway . The axons of the m i t ral cel l s form the ol
factory tract and di scharge into the olfactory area, or secondaryolfactory nuc leus
,at the base of the olfactory bu lb . These axons
give off col latera l branches whi ch discharge am ong very sm al lneurons of the olfactory bu lb , the granu le cel l s, whose chief
processes are di rected peripheralward , to end am ong dendr itesof the m itra l cel ls .A ttenti on has al ready been cal led (pp. 75 and 9 1 ) to the factthat
,though sm ell and taste are both chem i cally exc i ted Senses
,
the olfactory organs can be exc i ted by m uch m ore di lute soluti ons of the st im u lating substances than can the gustatory or
gans . The l oweri ng of the threshold for olfactory stim u l i hasbeen effected by several m eans
,am ong whi ch we m ay mention
the fol lowing : Whereas in the taste- buds there i s a synapsebetween the spec ific receptor cel l s and the per ipheral nerve-fiber
(Fig. 35 , p. there i s no such synapse in the olfactory organ,
the peripheral receptor ce l l gi ving ri se di rectly to the O l factorynerve-fiber (Fig. In the second place, the per ipheral
gustatory nerve-fiber di scharges central ly into several neu ronsof the prim ary gustatory center in the m edu l la oblongata ; butm any peripheral olfacto ry fibers enter a single gl om eru lus
,where
they are engaged by dendrites from only one or two m i tral cel l s,thus provid ing for the summ ation of stim u l i in each m i tral cel l .Again, the col lateral di scharge from the olfactory tract into thegranu le cel ls (whi ch are very num erous) carries the di schargefrom the m i tral cel ls back again into these cel l s and thus reinforces thei r di scharge (see pp. 101 , By these and otherdevi ces exceedingly feeble peripheral stim u l i m ay gi ve ri se tovery strong exc i tati ons in the olfactory centers .The fibers of the olfactory tract reach the olfactory area , or
secondary center , by three paths whi ch Spread out from the
base of the olfactory bu lb and are known as the m edial,inter
m ediate , and lateral olfactory striae (these are shown but notnamed on Fig. 53
, p. The O l factory area has var ious subdivi sions (Fig. the m ost important of whi ch are : ( 1 ) thelateral olfactory nuc leus (or gyrus) whi ch receives the lateral

THE OLFACTORY APPARATUS 19
olfacto ry stria and extends backward di rectly into the tip of thetemporal lobe of the cerebral cortex (uncus) , where the ventrolateral ends of the h ippocampus and the hippocampal gyrus cometogether ; (2) the m edial olfacto ry nuc leus
,inc luding the sub
callosa l gyrus (Fig. 52, p. 1 19 ) and septum ,
whi ch receive them edial olfactory stria ; (3) the inte rm ediate olfactory nuc leus
,
whi ch occupies the anterior perforated substance (F igs . 53 , 105 )and receives the interm ediate olfactory str ia . These nuc lei areal l important reflex centers, where o lfactory stim u l i are combined
Olfactory bulbMed ial ol factory gyrus (stria)
O lfactory tractLateral olfactory gyms
L imen insular
Anterior perforatedUncus (hippocampal Bu cc
gyrus)Hippocampal gyrus
Fig. 105 .—Brain of a hum an fetus at the beginning of the fifth m onth
cm . long) , il lustrat ing the olfacto ry centers vis ib le on the ventralsurface. (After Retz ius , from Morris ’ An atomy. )
w i th other sensory impressions , each nuc leus having i ts own particular reflex pattern . The interm ediate nuc leus (also cal ledtubercu lum olfactorium and by E dinger lobus parolfactor ius) i sbetter devel oped in m any other m amm als than in m an
,and i s
probably espec ially concerned w i th the feeding reflexes of thesnout or m uzz le, inc luding sm el l , touch , taste, and m uscu larsensibi l i ty
,a physiologi cal complex which Edi nger has cal led
collectively the “oral sense .
”Thi s complex of m uzz le reflexes
has probably played a very important rele in the earl ier stagesof the evolutionary history of the correlation centers of the
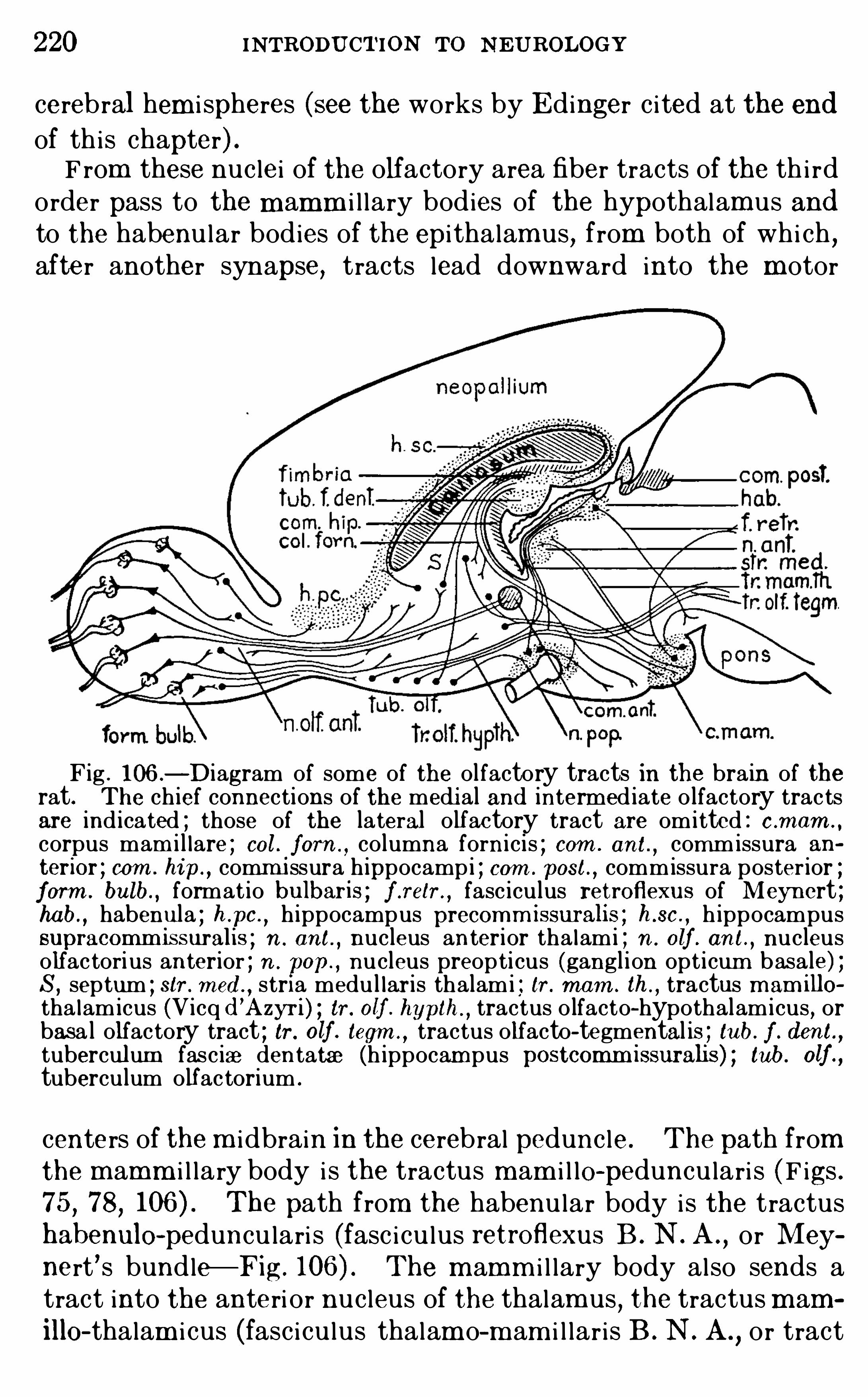
220 INTRODU CTION To N EU ROLOGY
cerebral hem i spheres (see the works by E dinger c i ted at the end
of th is chapter) .F rom these nuc lei of the olfactory area fiber tracts of the th i rd
order pass to the m amm i l lary bodies of the hypothalam us andto the habenu lar bodies of the epi thalam us, from both of whi ch ,after another synapse, tracts lead downward into the m otor
Fig . 106 .—Diagram of some of the olfactory tracts in the brain of the
rat. The ch ief connections of the medi al and interm ed iate o lfactory trac tsare ind icated ; those of the lateral o lfactory tract are omitted : c .mam . ,
corpus mam i l lare ; col . form ,co lumna forni cis ; com . ant.
,commissura an
terior ; com . hip ,comm i ssura h ippocampi ; com . post. , commissura poster ior ;
form . bulb. , format io bulbaris ; f .relr .,fasc iculus retroflexus of Meynert ;
hab. , habenula ; h .pc .
,h ippocampus precomm issuralis ; h .sc .
, h ippocampussupracomm issuralis ; n . ant. , nucleus anterior tha lami ; n . olf . ant.
,nucleus
olfac tori us anterior ; n . ep., nucleus preop ti cus (gangl ion opt i cum basale) ;
S, septum ; sir . med .,stria medul lar is thalami ; tr . mam . th . , tractus m am illo
tha lamicus (Vicq d ’
A zyr i ) ; tr . olf . hyplh .,tractus o lfacto-hypothalamicus , or
basa l olfactory tract ; tr . olf . tegm .,tractus olfacto-tegmental is ; tub. f . d ent ,
tuberculum fasciae dentatae (h ippocam pus postcomm issuralis) ; tub. olf .,
tuberculum o lfac torium .
centers of the m i dbrain in the cerebral pedunc le. The path fromthe mamm i l lary body i s the tractus m am i l le-
peduncu lari s (F igs .75 , 78, The path from the habenu lar body i s the tractushabenulo-
peduncu lari s (fasc icu lus retroflexus B . N . A .,or Mey
nert ’s bundle—Fig. The m amm i l lary body also sends atract into the anteri or nuc leus of the thalam us
,the tractus mam
illO - thalam i cus (fasc icu lus thalam o—m am i l lar i s B . N . A .,or tract
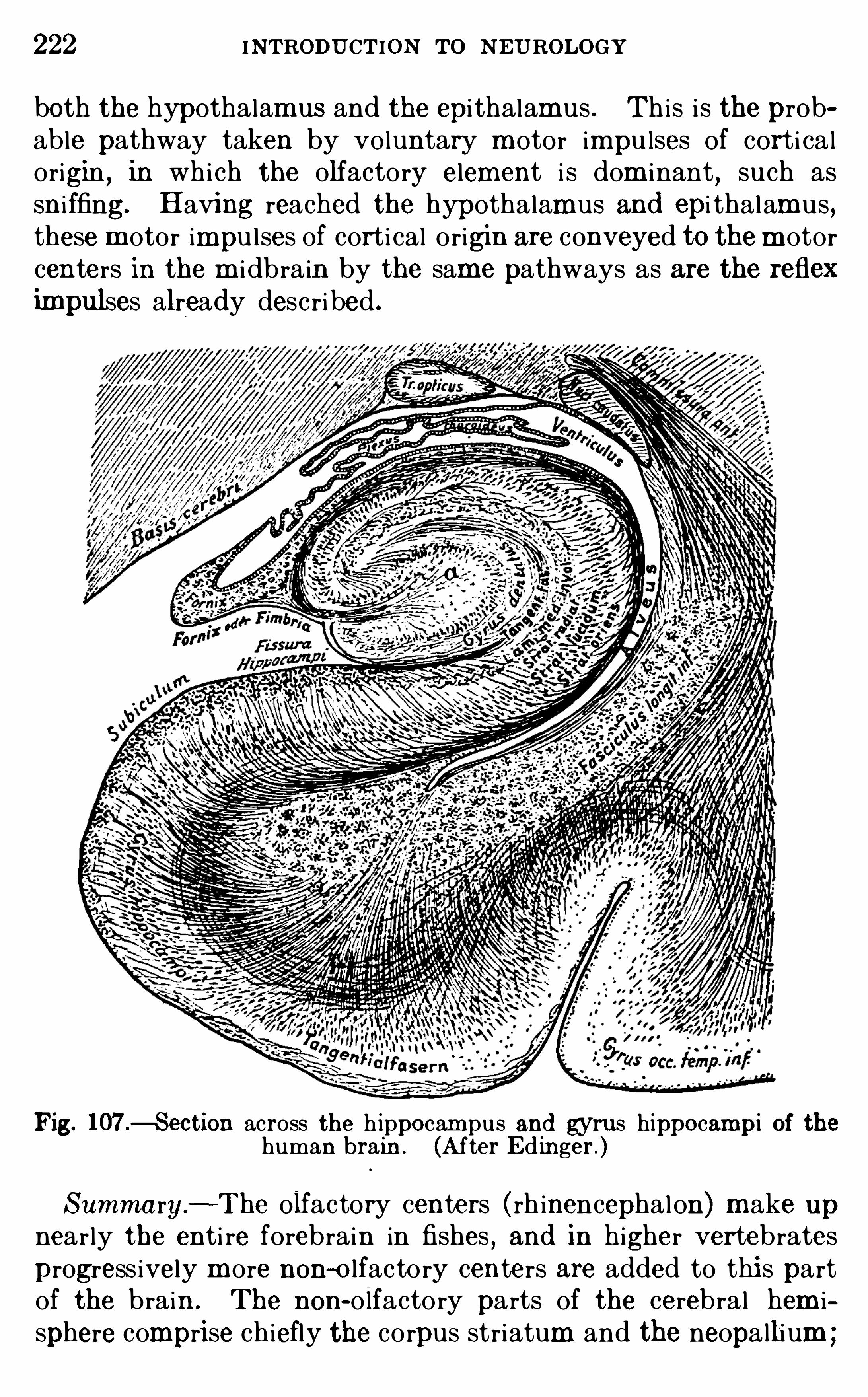
222 INTRODU CTION To N EU ROLOGY
both the hypothalam us and the epi thalamus . Th is i s the probable pathway taken by voluntary m otor impu lses of corti calorigin , in whi ch the olfactory element i s dom inant, such assni ffing. Having reached the hypothalam us and epi thalam us ,these m otor impu l ses of corti cal origin are conveyed to them otorcenters in the m i dbrain by the same pathways as are the refleximpulses already desc ribed .
Fig. 107 .4 ection across the h ippocam pus and gyrus hippocampi of thehuman brain . (After E d inger . )
Summary.
—The olfactory centers ( rh inencephalon) m ake upnearly the enti re forebrain in fishes
, and in higher vertebrates
progress ively m ore non-o lfactory centers are added to thi s partof the brain . The non-olfactory parts of the cerebral hem isphere comprise chiefly the corpus striatum and the neopallium ;

THE OLFACTORY APPARATU S 223
the latter m akes up by far the larger part of the human hem isphere. The rh inencephalon consi sts of a reflexpart in the brainstem and a cort i cal part in the archipal l ium . Sm el l and taste areboth chem i cal ly exc i ted senses , but the threshold of exc i tation i sm uch lower in the case of sm ell . Thi s i s brought about by thesuppressi on of a synapse in the per ipheral receptor organ and bya complex m echanism for the summ ation and reinf orcem ent ofst im uli in the pri m ary olfactory center in the olfactory - bu lb .
The secondary olfactory center i s the olfactory area , whi ch hasthree parts, each of which i s a reflex center of di stinct ive type.
The reflex path from the secondary center passes backward tothe epi thalam us and to the hypothalam us , from both of whi ch adescending path goes to the m otor centers in the cerebral
pedunc le. The secondary olfactory center also di scharges intothe olfactory cerebral cortex, whi ch i s chiefly contained withinthe hippocampus and from whi ch m ani fold assoc iat ion pathwaysconnect with al l other parts of the cerebral cortex.
LITE RATU REBAR KE R , L . F . 1901 . The Nervous System
,N ew York
, pp. 747- 781 .
EDIN GER , L . 1908 . VOrlesungen fi ber d en Bau d er nervescn Zentralorgane, Bd . 2 , Verglei chende An atomic d es Geh irns , Leipz ig.
1908 . The Relat ions of Comparat ive An atomy to ComparativePsycho logy, Jour . Com p . N euro logy, vo l . xvii i , pp . 437—457 .
1908 . Ueber d i e Orals inne d ienenden Apparate am Gehi rn d erSauger, Deutsch . Zeits . f l N ervenhcilkim d e, B d . 36 .
HE RRICK, C . JUDSON . 1908 . On the Phylogenet i c D ifferent iation of{1515
3
1 Oéfgians of Smel l and Taste, Jour . Comp . Neurology, vo l . xviii , pp .
19 10 . The E vo lution of Intell igence and I ts Organs , Science, N . S.,
vo l . xxxi,pp . 7—18.
19 10 . The Morpho logy of the Forebra in in Am ph ib ia and Reptilia,
Jour . Comp . N euro logy, vo l . xx , pp . 413—547 .
JOHNSTON , J . B . 1906 . The N ervous System of Vertebrates,Ph i la
delphia,pp . 176- 189
,292—337 .
1 9 09 . The Morpho logy of the Forebrain Ves i cle in Vertebrates,Jour . Comp . Neuro logy,
vo l . xix,pp . 457—539 .
.
19 13 . The Morpho logy of the Septum , Hippocampus and Pa lli alCommmsure in Reptiles and Mamm a ls , Jour . Comp . N euro logy, vol . xx ii i ,pp. 37 1—478 .
KAP PE RS,C . U . A . 1908 . D ie Phylogenese d es Rh inencephalons, d es
Corpus Striatum und d er Vord erhirnkomm issuren,Fo li a N eurob iologi ca,
B d . 1 , pp . 1 73—288 .
ZWAA RDEMAHE R,H. 1895 . Die Physiologic d eS Geruchs , Leipz ig .
1 900 . Revue ge’ nerale sur l’
olfaction,Année Psycho l . , vo l . vi .
1902 . Geruch . E rgebnisse d er Physiologic , B d . 1 .

CHAPTER XVI
THE SYMPATHETIC NERVOUS SYSTEM
BE FORE we can extend our analysi s of the conduction pathsinto the realm of the visceral activi ties of the body we m ust consider briefly the sympatheti c nervous system through whi ch theregu latory control of these activi ties i s effected . Most of thevisceral activi ties are pe rformed ei ther unconsc i ously or withvery imperfect awareness . The nervous m echan i sm s of m anyof them are sti l l obscu re. N evertheless the vi sceral functionsas a whole are of enorm ous importance, not on ly in the m aintenance of the physi cal welfare of the body, but also as the organi cbackground of the enti re consc i ous l ife (see p.
Many of the visceral functions can be perform ed qu i te apartfrom any nervous contro l whatever by the intr insi c m echan i sm sof the viscera them selves . The heart m uscu lature, for instance,beats autom ati cal ly with a characteri sti c rhythm , and m ost ofthe other vi sceral m usc les have the power of autom ati c rhythm i ccontraction . Som e of the glands of the body may be exc i tedto secretion by chem i cal substances di ssolved in the blood . Forinstance
,when food enters the sm al l intestine from the stom ach ,
the intestinal glands are di rectly exc i ted to activi ty by the presence of the food . Som e of thei r sec reti ons are pou red out intothe intestine to act as digestive j ui ces ; others are absorbed directly by the blood ( internal sec retions) . Am ong the latter i ssec retin, a substance whi ch i s carried by the blood- stream to the
pancreas and there exc i tes the secretory activi ty of th i s organ tothe form ation of panc reatic j u i ce, which i s , in turn, pou red intothe intestine. The very complex secretory activities involved inthe form ation of the intestinal and panc reati c j u ices under thestim u lus offered by the presence of food in the intestine, therefore, are not di rectly exc i ted by the nervous system ,
though theymay be brought under nervous contro l in a secondary way .
Most of the viscera are,however
,under imm ediate nervous
control of two sorts . Thi s control i s partly derived from the
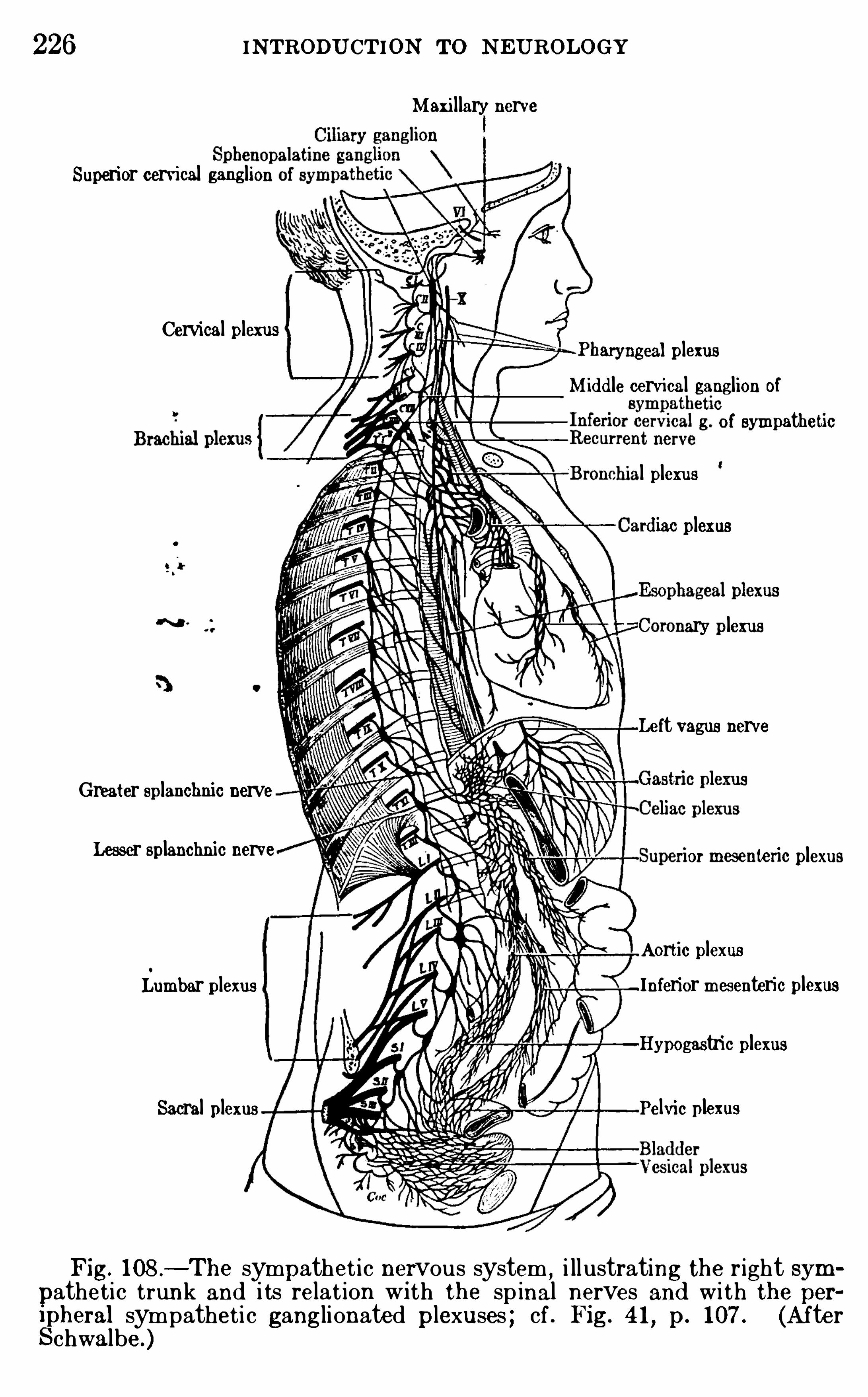
226 INTRODU CTION To N EUROLOGY
Superior cervical ganglion
Cervical plexus
Greater splanchnic nerveLesser splanchnic nerve
Lumbar plexus
Sacral plexus
Maxi l lary nerve
aryngeal plexusMid d le cervical ganglion of
of sympatheti c
l plexusrd iac plexus
nary plexus
Left vagus nerve
or mesenlcric plexus
Aortic plexusInferior mesenteric plexusHypogastric plexus
Pelvic plexus
Fig. 108 .—The sympathet i c nervous system ,
i ll ustrating the right sympatheti c trunk and i ts relat ion Wi th the sp inal nerves and wi th the peripheral sympatheti c ganglionated plexuses ; cf . Fig. 4 1
, p. 107 . (A fterSchwalbe.)

THE SYMPATHETIC NE RVOUS SYSTEM 227
sympatheti c system inc ludes those neurons whi ch put the peripheral autonom ous system into functional connection with thecentral nervous system
,thus providing a central regu latory con
trol over the autonom ous system . Thi s part of the sympatheti c nervous system inc ludes the per ipheral courses of theneurons involved in the general cerebro- spinal vi sceral reflex system s (see pp. 76
,89
,
The peripheral autonom ous nervous system appears to be adi rect su rvival of that di ffuse type of nervous system whi ch isfound in the l owest anim als which possess nerves at al l , such assom e jel ly-fishes and worm s . The central nervous system ofh igher an im als i s supposed to have devel oped by a concentrationof gangl ia in such a d iffuse system (see p. a portion of whi chrem ains as the per ipheral autonom ous sympatheti c system (Fig.
17, p. But dur ing evolution the central nervous system
increased in importance for integrating and regu lating the
functions of the body, the central control of the viscera assum ed
greater importance, and the general cerebro- spinal visceral system s were devel oped to serve th i s fun ction .
F igure 56 (p. 126) i l lustrates the typi cal arrangement of thevi sceral sensory and m otor fibers in the Spinal nerves , and thei rrelations to the sympatheti c gangl i a and nerves . These fibers
,
of cou rse,bel ong to the cerebro- spinal vi sceral system s ; the
peripheral autonom ous system i s not inc luded in the diagram .
The central control of the visceral apparatus i s effected (1 ) byafferent visceral nerve-fibers di str ibuted peripheral ly throughthe sympatheti c nerves and entering the spinal cord through thedorsal Spinal roots , and (2) by efferent vi sceral nerves whi chleave the spinal cord through the ventral roots and also enter thesym patheti c nerves . I n lower vertebrates (and possibly also inman) som e of these fibers leave by the dorsal roots also .
The cel l bod ies of the afferent neurons lie in part in the
spinal gangl i a and in part in the sympatheti c gangl ia . F igu re109 i llustrates the connections of these two types of afferentvi sceral neu rons . N euron 3 of th is figure may transm i t i tsimpu l se ei ther di rectly into the spinal cord through its central lydi rected process or by a col lateral branch to some other cel lbody of the spinal gangl i on (neu ron The fiber m arked 4ari ses from a cel l-body lying in som e sympatheti c ganglion and
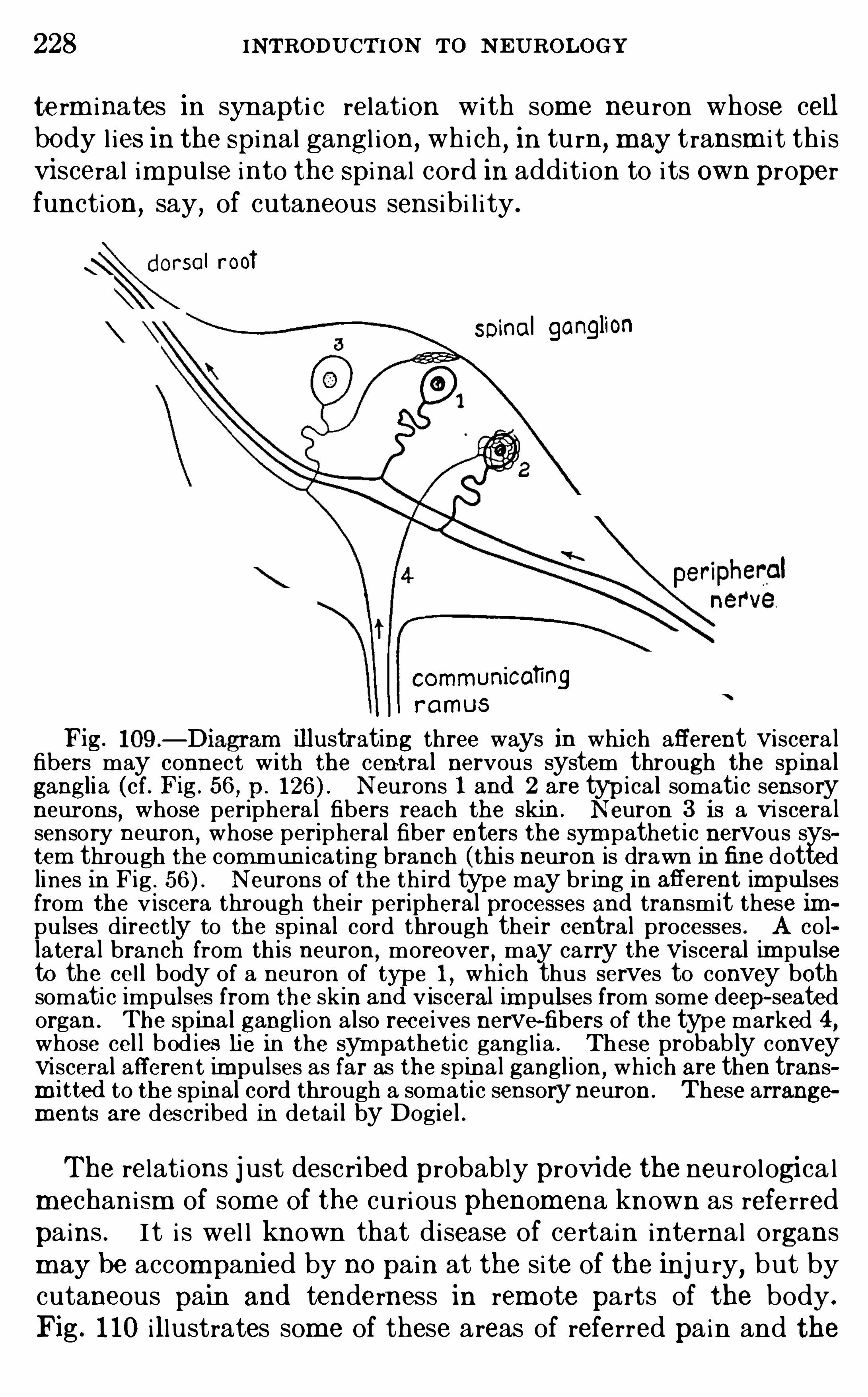
228 INTRODU CTION To N E U ROLOGY
term inates in synapti c relation with som e neu ron whose cellbody l ies in the Spinal gangl i on, whi ch , in tu rn ,
may transm i t th i svisceral impu l se into the Spinal cord in addit ion to i ts own properfunction
,say
,of cutaneous sensibi l i ty.
sol rooT
soinol ganglion
communicatingramus
Fig. illustrating three ways in whi ch afferent viscera lfibers may connect w i th the cent ra l nervous system through the spina lganglia (cf. Fig. 56 , p . Neurons 1 and 2 are typ i ca l somatic sensoryneurons
,whose periphera l fibers reach the skin. N euron 3 is a Viscera l
sensory neuron,whose peripheral fiber enters the sympathet i c nervous sys
tem through the commun i cat ing branch (th is neuron is drawn in fine dottedlines In Fig . N eurons of the th ird type may br ing in aflerent impulsesfrom the v iscera through their peripheral processes and transmit these 1m
pulses d irectly to the spinal cord through thei r central processes . A co llatera l branch from th is neuron
,moreover , may carry the visceral im pulse
to the ce l l body of a neuron of t e l,whi ch thus serves to convey both
somatic impulses from the skin an v isceral im pulses from some deep- seatedorgan . The sp ina l ganglion a lso receives nerve-fibers of the type marked 4,whose cel l bod ies lie in the sym pathet i c gang l ia . These probab ly conveyvisceral afferen t im pulses as far as the Spina l ganglion,
wh i ch are then transm itted to the sp inal cord through a somati c sensory neuron . These arrangements are described in deta i l by Dogiel .
The relations j ust desc ribed probably provide the neurologi ca lm echan ism of som e of the cu ri ous phenom ena known as referred
pains . I t i s wel l known that disease of certain internal organsm ay be accompanied by no pain at the si te of the inj u ry, but bycutaneous pain and tenderness in rem ote parts of the body .
Fig. 1 10 illustrates some of these areas of referred pain and the
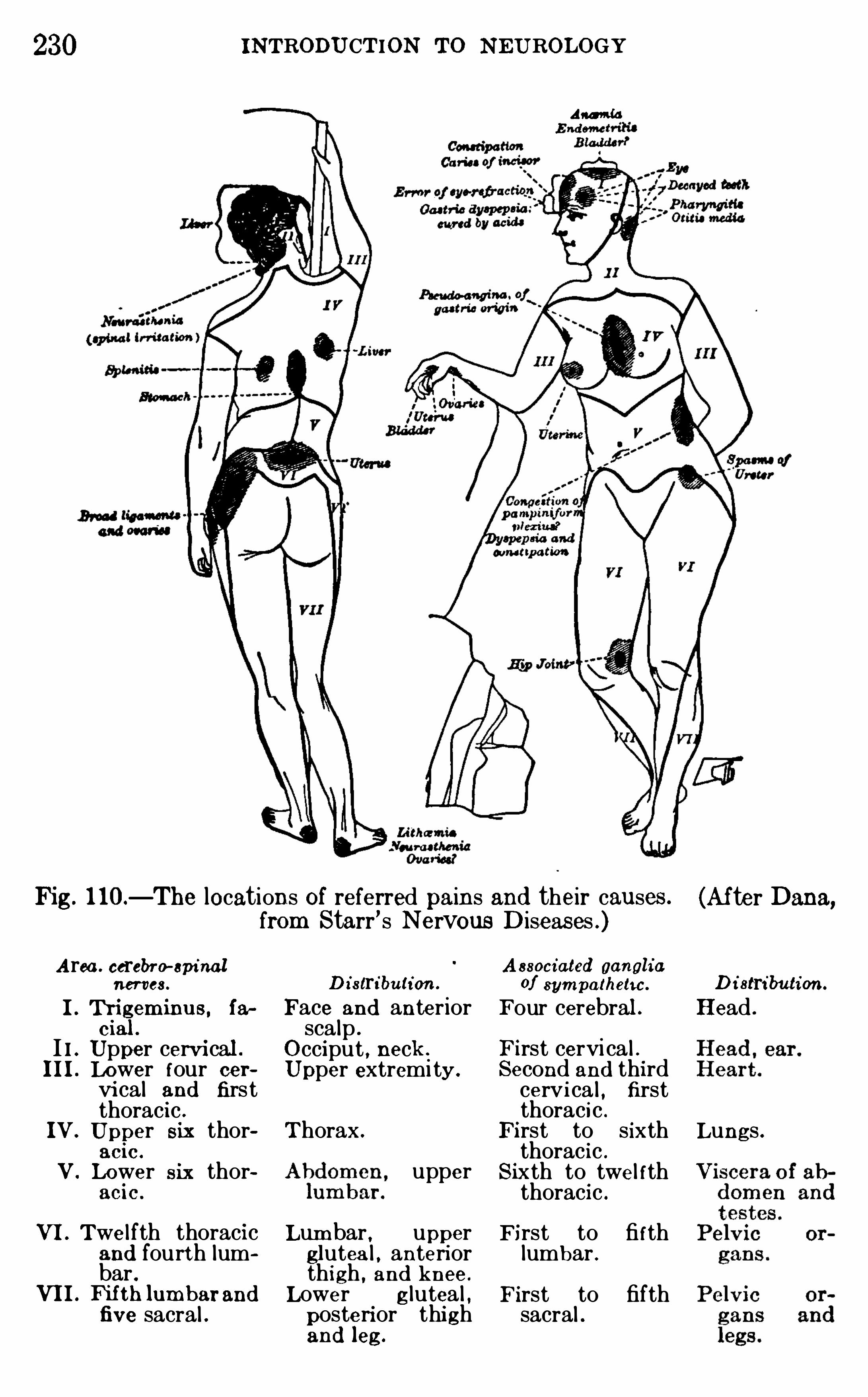
230
Fig. 1 10—The locations of referred pains and their causes .
from Starr ’ s Nervous Diseas es .)
A rea. cerebro-epinalnerves.
I. Thi gem inus , ia
c ial .
I I . Upper cervi cal .
III. Lower four cervi cal and firstthoracic .
IV. Upper Six thorac ic .
V. Lower six thoracic .
VI. Twe l fth thoracicand fourth lumbar .
VII. N th lum barandfive sacral .
D istribution .
Face and anteriorsca lp .
O cciput , neck .
Upper extremi ty .
Thorax .upper
Lum bar. upperglutea l , anteriorthigh , and knee .
Lower g lutea l ,posterior thi ghand leg.
INTRODU CTION TO NEU ROLOGY
Am i d
A ssociated gangliaof sym patheti c .
Four cerebral .First cervi ca l .Second and thirdcervi ca l , firstthoracic .
First to sixththoracic .
Sixth to twel fththoracic .
First to fi fthlumbar .First to fifthsacral .
(Af ter Dana,
D i stri bution .
Head .
Lungs .

THE SYMPATHETIC N ERVOUS SYSTEM 231
The port ions of the sympatheti c system related to these respective regi ons are as fol lows : ( 1 ) The m i dbrain sympatheti c , compr i sing chiefly the c i l iary gangl i on beh ind the eye and its nerves ,these being related to the brain through the I I I crani a l nerve.
(2) The bu lbar sympatheti c , related to the brain chiefly throughthe V I I , IX,
and X cranial nerves . (3) The thorac i c- lumbarsympatheti c , related to the spinal cord through the I thorac i cto I I or I I I lumbar nerves . (4) The sacral sym patheti c ,related to the spinal cord through the I I to IV sacral nerves .
E ach of these fou r regi ons has its own dist inctive physi ologi calcharacter i sti cs
,inc luding in some cases a spec ial type of reaction
to certain drugs . They al l exh ibit a comm on reaction to n i cotin in physiologi cal doses . The effect of this poison i s to paralyzethe synapses between the pregangl i on i c and the postgangl i on i cneurons and thus to i solate the peripheral sympatheti c neu rons
physi ologi cal ly from efferent impu lses ari s ing within the centralnervous system . Adrenal in (extract of the suprarenal glands)affects chiefly the thorac i c- lum bar sympatheti c system (see p.
On the other hand, poisons of a d ifferent group, inc luding
atropin ,m uscarin
,and pi l ocarpin , are said to act chiefly upon
the m i dbrain,bu lbar and sacral sympatheti c , but not upon the
thorac i c- lum bar system . There are other cases of very spec ificaction of drugs upon spec i al parts of the sympatheti c nervoussystem .
Summary—From the preceding considerations i t i s evi dentthat the sympatheti c nervous system cannot be sharply separated anatom i cal ly or physio logi cal ly from the cerebro- spinalsystem . The cel l bodies of the neurons of the cerebro- spinalvi sceral system lie part ly with in and partly wi thout the centralnervous axi s . A gangl i oni c sym patheti c trunk extends on eachside of the body along the spinal column, and the gangli a of th i strunk are connected with m ost of the spinal nerves by com
municating branches . The neu rons of th i s trunk of vertebralsympatheti c gangl ia bel ong chiefly to the cerebro—spinal Vi sceralsystem
,since they are concerned with the central regu latory
mechan i sm of the vi scera . All parts of the visceral nervous system whi ch lie peripheral ly of the comm un i cating branchesbetween the sympatheti c gangli onated trunks and the spinalroots, and can be anatom i cal ly separated from the peripheral
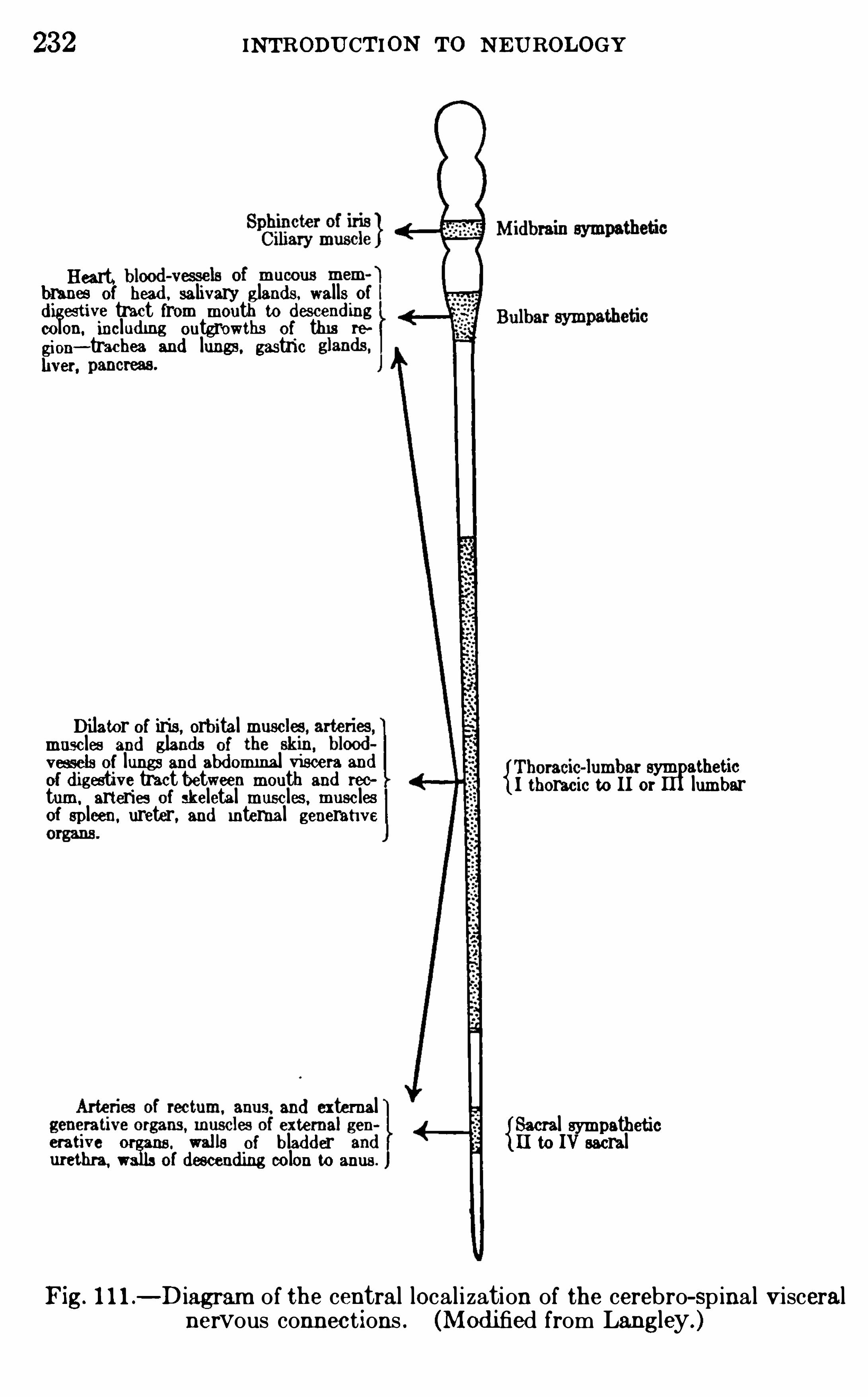
232 INTRODU CTION To NEUROLOGY
Heart.branes of head , salivary gland s walls of
d iges ti ve tract from mouth to d escend ingcolon. includ ing outgrowths of thi s re
glion—trachea and lungs, gastricver.
Di lator of iris, orbital musclmusc les and gland s of
zfess elsd ig
of lungs antee
abd ominal viseeh
r
sand
sym atheticestive tract tween mout an ree
turn. arteries of skeletal musc les, musc les I thoracic to II or lumbarof spleen. ureter, and internal generati ve
and external{Sacral sympathetic
erative
m eans “ of blad d er [ I to IV sacralof d escend ing colon to anus.
Fig. l l l .—Diagram of the central local ization of the cerebro-Spinal visceral
nervous connec t ions . (Mod i fied from Langley.)

CHAPTER XVII
THE VISCERAL AND GUSTATORY APPARATUS
OUR knowledge Of the functional local i zation with in the spinalcord of the general vi sceral reflex centers related to the spinalnerves i s sti l l rather indefin i te. Most Of the cerebro- spinalcontro l Of the visceral reactions of the body i s effected from the
bu lbar sympatheti c centers by way Of the vagus nerve. The
afferent fibers Of these system s al l enter the fasc i cu lus sol i tarius,a longi tudinal bundle O f fibers in the l ower part of the m edu l laOblongata, and they term inate in the nuc leus Of visceral sensoryneu rons whi ch accompanies th is fasc i cu lus (F igs . 71—74, 77,
The spec ial vi sceral fibers Of the nerves Of taste also term inate in th i s nuc leus . The efferent fibers of these system s arisechiefly from the dorsal m otor nuc leus O f the vagus, a c luster Ofneurons whi ch produces an em inence in the floor Of the fourthventri c le known as the ala c inerea or trigonum vagi (F igs . 7 174
,F rom th i s nuc leus ari se pregangl i on i c fibers for the
innervation of var ious system s of vi sceral m usc les Of bloodvessel s
,esophagus , stom ach , intestine, bronch i , and others .
Most vi scera possess a double innervation— from the thorac i clumbar sym patheti c system and from the m i dbrain,
bu lbar,or
sacral system (see p. For instance,the heart-beat is
accelerated by the thorac i c- lum bar system and inh ibi ted by thebu lbar system through the vagus ; and the i r i s i s contractedthrough the m i dbrain sympatheti c , but di lated through thethorac i c by way of the superi or cervi cal gangl i on (p.
O rgans of Ci rcu lati on - The nervous control of the heartand blood-vessel s i s far too complex for fu l l descripti on here. A
few general featu res on ly can be touched upon .
The rate of blood flow m ay be varied for the body as a wholeby changes in the rate and force Of the pu l sati ons of the heart ,and for part icu lar part s Of the body by changes in the cal iberOf i ts blood-vessels . The heart beats autom ati cal ly
,but its
234

THE VISCERAL AND GUSTATORY APPARATU S 35
rate i s regu lated th rough the cardiac nerves . The caliber Ofthe sm al ler blood-vessel s and hence the am ount Of blood whi chcan pass through them i s regu lated by vasom otor nerves . Boththe heart and the m uscu lar wal ls Of the vessels have a doubleinnervation . The heart has an accelerator nerve and an inbibitory nerve ; the sm al ler arter ies have vasodi lator and vasoconstri ctor nerves . The am ount Of blood pumped by the heartat any time w i ll depend upon the equ i l ibrium exi sting betweeni ts accelerator and its inh ibi tory fibers and upon the res i stance Offered by the peripheral vessel s ; that flowing throughany parti cu lar system Of blood-vessel s wi l l be affected also bythe equ i l ibr ium between the vasodi lator and the vas oconstr i ctornerves Of these vessels .There are sym patheti c gangl i a with in the heart . I ts extrinsi c
nerve supply inc ludes afferent fibe rs to the brain and efferentfibers Of two sorts, vi z . , the accelerator and inh ibitory fibersalready mentioned . The afferent fibers are represented in asm al l sym patheti c nerve, the nerve Of Cyon , whi ch i s also cal ledthe depressor nerve. They ari se from the wal l s Of the ventri c lesOf the heart and j oin the vagus
'
trunk,through whi ch they enter
themedu l la Oblongata . Stim u lat ion of th is nerve produces a fal lOf arterial pressure by di lat ing the vessel s throughout the body,espec ia l ly in the viscera . I t appears to act to reduce the laborOf the heart when intraventri cular pressure becom es excess ive.
The m edu l la Oblongata contains a center whose stim u lationcauses inhib ition Of the heart-beat . These efferent fibers gO outas pregangli oni c fibers Of the vagus nerve and term inate in thecardiac sympatheti c plexus (Fig. where thei r postgangl i on i cneurons are l ocated . There i s also a center in the m edu l la Oblongata (whi ch has not been prec i sely local ized) whose stimu lation causes acceleration Of the heart-beat . These acceleratornerve-fibers do not leave the brain through the vagus , but apparently they descend through the spinal cord to the l ower cervi calregi on and pass out into the sympatheti c nervous system at thislevel . The centers Of vasom otor contro l Of various regi ons Of
the body are indicated in Fig. 1 1 1 .
O rgans of Respi ration—Oxygen i s supplied to the ti ssues Ofthe body in a great variety Of ways in different anim als . In
some Of the s impler animals, as in plants generally, oxygen is

236 INTRODU CTION TO N E U ROLOGY
simply absorbed from the surrounding m edi um by the exposedsu rfaces . In al l but the l owest an im al s there i s a blood-vascu larsystem by m eans of whi ch the oxygen absorbed at the surface i stransferred to the deeper ti ssues . In insects
,however , th i s re
su l t i s Obtained chiefly by a different apparatus, nam ely, a system Of ai r tubes (tracheae) whi ch rami fy am ong the ti ssues andsupply oxygen di rectly to the functioning cel l s . In m ost waterbreathing an im als a portion Of the su rface Of the body i s lamellated and vascu lari zed to form gi l l s to fac i l i tate the absorption Of
oxygen by the blood- stream,and in ai r-breath ing vertebrates
lungs are developed to accompl i sh the sam e resu lt . The nervousm echani sm s Of respi rati on wi l l differ in al l Of the cases c i tedabove
, and i t i s on ly in m amm als that we shall here consider thedetai ls Of thi s m echani sm .
In ordinary breath ing, inspi ration i s effected by actively inc reasing the volume Of the thorac i c cavi ty and thus creating asuction through the trachea , whi le expi rat ion i s the resu l t Of the
passive return Of the organs involved to thei r form er posit ionsby reason of thei r own e last i c i ty . The m usc les involved ininspi rati on bel ong to two groups : ( 1 ) the internal apparatus, i . e.
,
the diaphragm ,and (2) the external apparatus , the intercostal
and other m usc les Of the body wal l . These are al l som ati cm usc les . In forced respi ration various other m usc les act in anaccessory way du ring both inspi ration and expi ration .
The diaphragm i s innervated by the phreni c nerve, whi chtakes i ts origin from the fou rth and fifth cervi cal spinal nerves ;and the intercostal m usc les are innervated by ventral spinalroots ari sing successively from al l thorac i c segm ents Of the spinalcord (Fig. The accessory m usc les are in part som ati cm usc les Of the abdomen and shou lder and in part spec ia l vi sceral m usc les Of the head
, particu lar ly those Of the glotti s
( innervated by the vagus) and Of the nostri l s ( innervated by theV I I c ranial nerve) .The anatom i cal relations j ust descr ibed imply that, althoughrespi ration i s a vi sceral function ,
in mammals the necessarym ovements for ordinary breathing are perfo rm ed by som ati cm usc les . Thi s i s not true in fishes . Here the organs Of respiration (gi l l s) are stri ctly visceral structures innervated by vi sceral components Of the c rania l nerves, whose cerebral center i s

238 INTRODU CTION TO N EUROLOGY
m ents Show that the spinal segm ents from which al l Of them otorrespi ratory nerves ari se cannot Of them selves effect the coord inations necessary in respi rati on . Thi s i s in m arked contrastwi th m any other reactions (both vi sceral and som atic) , whose
perform ance i s sti l l possible after the separation Of the spinalcord from the brain .
I f now,in a thi rd an im al
,the medu l la Oblongata i s cut across
at any point above the m i ddle Of i ts length , say at the l ower bord er Of the pons , the respi ratory processes are in no way disturbed .
Thi s shows that there i s a respi ratory correlation center in thel ower half Of the m edu l la Oblongata, that i s, somewhere i nthe regi on corresponding to the
“vi sceral area” Of the fishbrain .
The ai r tubes Of the lungs are provi ded with sm ooth m usc lefibers by whi ch their cal iber m ay be contracted . Thesem usc lesare innervated by the vagus, and the hyperexcitation Of thei rm otor nerves may impede respi ration,
th i s being one Of the factors whi ch cause asthm a . The cerebral center from whi ch thesei ntri ns i c musc les Of the lungs are innervated has been shown tolie in the m i ddle part Of the dorsal m otor vagus nuc leus (Fig. 73,nuc d orsali s vagi ) . These are pregangl i on i c neu rons, the corresponding postgangli on i c neurons lying in sympatheti c gangl iadi stri buted along thepu lm onary branches Of the vagus (Fig.
The apparatus described in the preceding paragraph i s, however, not responsible for the m aintenance Of the regu lar rhythmOf breathing. Physio logi cal experiments Show that there i s som e
where in the lower part of the m edu l la Oblongata a respi ratorycenter whi ch perform s th i s functi on . Th is center may apparently be exc i ted to activi ty di rectly by vari ations in the composition Of the blood whi ch reaches it
,espec ial ly ei ther by a d e
ficiency in oxygen or by an excess Of carbon dioxi d . Its activi tymay also be m odified by nervous influences reaching i t th roughthe peripheral afferent nerves , the vagus be ing the only nervewhich appears to be able to act di rect ly on the respi ratory center,though the strong exc i tation Of alm ost any sensory nerve Of thebody may under some c i rcum stances indi rectly affect the res
piratory rhythm . Coughing and sneez ing are spec ia l cases of
ibissort . The reflex mechani sm Of the cough i s i l lustrated in
ig. 1 13 .
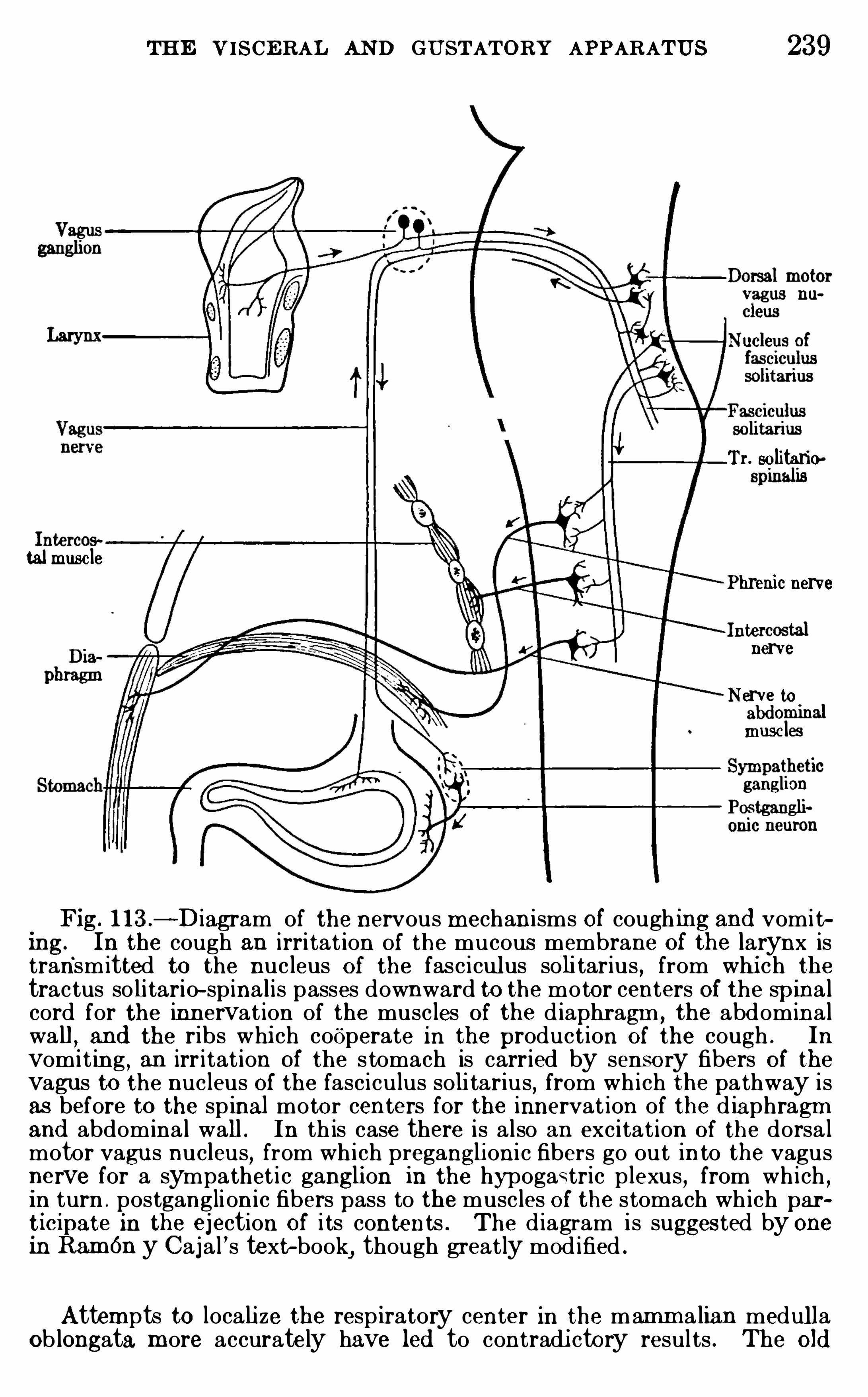
THE VISCERAL AND GUSTATORY APPARATUS 239
Tr. so litario
talmusc lePhrenic nerve
nerveNerve tomusc les
Fig. 1 13 .—Diagram of the nervous mechan isms Of cough ing and vomi t~
ing. In the cough an irri tat ion Of the mucous membrane Of the larynx istransmitted to the nucleus Of the fasciculus soli tarius , from whi ch thetractus so litario- spina l is passes downward to the motor centers Of the spina lcord for the innervat ion of the muscles Of the d iaphragm , the abdominalwa ll , and the ribs whi ch cooperate in the product ion Of the cough . In
vomi t ing , an irr itat ion Of the s tomach is carried by sensory fibers Of thevagus to the nucleus Of the fasciculus soli tarius, from wh i ch the pathway isas before to the spinal motor centers for the innervat ion of the d iaphragmand abdomina l wall . In th is case there is also an excitat ion Of the dorsa lmotor vagus nucleus, from wh i ch pregangli oni c fibers go out in to the vagusnerve for a sympathet i c gangli on in the hypogastric plexus , from wh i ch ,in turn . postganglion i c fibers pass to the muscles O f the stomach wh ich partic ipate in the ejection Of i ts contents . The d iagram is suggested by onein RamOn y Caja l ’s text-book
,though greatly mod ified .
Attempts to locali ze the respiratory center in the mamm a lian medullaOb longata m ore accurately have led to contradi ctory resul ts . The Old

240 INTRODU CTION TO N EUROLOGY
concep t ion Of Flourens that there is aminute vi ta l node under the lowestpo int Of the fourth ventri c le wh i ch is the respiratory center must be aband oned . Later the fasc iculus so l i tarius was identified as the “resp iratorytract
,
”and the nucleus associated w i th th is tract was regarded as the
respiratory center , but further experim ent has shown that thi s is not an
exact statem ent Of the case . Some phys iologi cal experim ents have sug
gested that the respiratory rhythm is ma inta ined by a center in the
reticular format ion Of the vagus region ventrally Of the fasciculus so l i tar ius .I t has recen tly been shown ,
as stated above, that afferent v iscera l fibersfrom the lungs whose ce ll bod ies lie in the vagus gangl ion enter the fasciculus soli tarius , and i t is kn own that from the nucleus Of th is tract a “tractusso li tario- sp ina li s ” (Fig. 1 12) descends into the motor centers Of the uppersegm ents Of the sp ina l cord . Th is descending v isceral sp inal trac t probab lyp lays some part in the regulat ion Of respirat ion,
though not the ch ief rOle.
RamOny Caj al and Kappers believe that , wh ile the upper part Of the nucleusOf the fascicu lus so l i tar ius has noth ing to do w i th respirat ion,
the lower endof this nucleus (comm issural nucleus O f Caj al , see Figs . 7 1 , 1 12, and 1 14) is atrue respiratory center . RamOn y Caj al
,in fact , th inks that thi s nucleus
serves both for reflexes exci ted by the sensory pulm onary nerves and a lso forthe norma l respiratory rhythm excited by carbon d ioxid in the b lood .
Th is hypothesis is not supported by d irect phys io logi ca l experiment , and forthe present we must content ourselves w ith the statement that the truerespiratory center has not been accurately located anatomically. Figure1 1 2 may be regarded as a true picture Of the essent ia l relations O f therespiratory nerves , w ith the reservation that the posi tion Of the resp iratorycenter is not precisely known .
There is a lso a reflex center for the regulation O f respirat ion in the med ialwal l Of the thalamus and others have been described in d ifferent par ts Of thebrain stem . The entire resp iratory mechanism is also under par tia l voluntary contro l from the cerebral cortex .
Whi le m any features Of the central respi rato ry mechani smrem ain Obscu re, i t seem s evident that the l ocation Of the ch iefrespi ratory center in the
“visceral area of the l ower part Of them edu l la Oblongata instead Of the portions Of the Spinal corddi rectly connected with the respi ratory m usc les i s a su rvival Ofthe ancestral condition found in fishes , where the enti re respi rato ry function i s carried on by a visceral apparatus (gi l ls) i nnervated from the vagus regi on .
O rgans of D igesti on.—Hunger seem s to be a complex in whi ch
at least three facto rs are present : ( 1 ) Spec ific hunger pangs d ueto waves Of m uscu lar contraction in the stom ach (Cannon ,Carl son) ; (2) appetite, or craving for food regardless Of the stateOf the stom ach ; (3) general m alaise from starvati on Of the ti ssuesand weakness . Appetite m ay pers ist after section Of the vagusnerves and i s probably a sensat ion d istinct from the hungerpangs.

242 INTRODU CTION TO N EU ROLOGY
no doubt play a very large part i n shaping the organi c background Of the enti re consc i ous l i fe (see p.
Students Of an imal behavior are in the habi t Of i nvestigatingthe abi l i ty of anim als to m ake simple assoc iations by train ingthem to perform parti cu lar acts under condit ions such that thenorm al stim u lus to the act i s always accompanied by a secondstim u lus of a different type. After m any repeti t i ons the re
sponse may be Obtained by presenting the second or col lateralstim u lus without the fi rst . For the nervous mechan i sm of“assoc iative m em ory” Of thi s sort see p. 64 . Pawlow hasfound that variations in the am ount Of sal iva secreted form anespec ia l ly good index Of assoc iations of th is type, and he has usedthis m ethod extens ively in analyz ing complex reactions, or cond itional reflexes, as he cal ls them . See the summ ary Of hi sresearches in the paper by Morgulis c i ted in the appended bibliography.
Tacti le sensib i l i ty is enti rely absent throughout the enti real imentary canal from the esophagus to the rectum ,
and thesam e holds true for m ost Of the other deep- seated vi scera Of thebody . E ven the substance Of the brain i s insens it ive to any
k ind Of mechani cal i rr i tati on . Sensibi l i ty to changes in tem
perature i s feebly developed or absent in m ost Of the viscera, theesophagus and anal canal being very sensi t ive to heat and cold,whi le the stom ach and colon are feebly sensi tive to these st im u l i .The enti re al imentary canal i s insens i tive to hydrochlor i c andorganic ac i ds in concentrations far in excess Of what ord inar i lyQCcu rs in ei ther norm al or pathologi cal conditions . The contact Ofalcohol wi th al l parts Of the m ucous m embrane Of the al im entarycanal gives r ise to a sensation Of warm th . Th is sensation i sd ifferent in charac ter from that caused by hot flu i ds and i s probably exc ited through the sym patheti c nerves , whi le the sensation Of warm th fel t in consequence of the passage Of hot flu i dthrough the esophagus i s exc i ted through the vagus .The dem onstrated absence of tacti le sensibi l i ty throughout
them ucous membrane Of the stom ach and intestine i s cons ideredby Hertz to indi cate that the sensations of fu lness ari sing fromthe distention of d ifferent parts Of the al im entary canal are d ueto the stretching Of the m uscu lar coat, and that, therefore, theseare to be regarded as varieties Of the m usc le sense. The same

THE VISCERAL AND GUSTATORY APPARATUS 243
may also be true of the bladder . The free nerve-endings (seeFig. 33 , p. 90) known to be present in these m ucous m embranes,part i cu lar ly in the bladder , may, however , share in exc i t ing thesesensations
,for these m embranes m ay wel l be sensi t ive to stretch
ing, even though qu i te insens i tive to simple pressu re. The onlyimmediate cause Of true visceral pain i s tensi on , and i t i s statedby Hertz that , SO far as the al im entary canal i s concerned , th istens ion is exerted on the m uscu lar coat
,not on the m ucous
l in ing. See the further discuss ion Of visceral pain, p. 250.
The vom i t ing reflex may be caused by exc i tations of sensoryterm ini Of the vagus nerve i n the stom ach , which are transm i ttedto the nuc leus Of the fasc i cu lus so l i tar ius in the m edu l la Oblon
gata, whence the nervous impu l ses are distr ibuted as shown i nFig. 1 13 to the appropr iate m otor centers .The Gustatory Apparatus—Taste , l ike smel l , i s a chem i cal
sense (see pp. 75 , 9 1 , Physi ological ly, i t i s c lassed bySher rington as an interoceptive or visceral sense, and i ts pr im arycerebral center i s intim ately j oined to the general v isceralsensory center in the-nuc leus Of the fasc i cu lus so l itar ius . Un l ikethe general v isceral sensory system ,
however , its per iphera lfibers have no connect ion with the sym patheti c nervous systemand the reactions m ay be vivid ly consc i ous . The end - organs ,or taste- buds (Fig. 35 , p. are present in the m ucous m em
brane Of the tongue, soft palate, and pharynx and are innervatedby theVII and IX c ran ia l nerves ; there are a few taste-buds alsoon the larynx and epigl ottis wh ich are probably suppl ied by thevagus (J . C . Wi lson) . All of these per iphera l gustatory fibers,upon entering the m edu l la Oblongata , term inate in the nuc leusOf the fasc i cu lus sol i tarius (F igs . 7 1 , 72, 73 , 1 14) along withthose Of genera l v isceral sensibi l i ty , those Of the gustatory system probably ending farther forward (toward the m outh) in th isnuc leus than thoseof the general vi sceral system s .There has been considerable controversy as to the exact course
taken by the per ipheral nerves of taste on thei r way to the brain,
m any c l in ical neuro logi sts bel ieving that al l of these fibers enterthe m edu l la Oblongata through the root Of the V c ran ia l nerve .
I t has now been c lear ly shown by the studies Of Cush ing and
others that the V nerve takes no part in the innervation Of
taste-buds . F igure 1 15 shows in continuous l ines the true

244 INTRODU CTION TO N EUROLOGY
Vmotor
Nuc. sensory V
VIImotorI pars. int.
IX sensory
Ala cinereaFasciculus so litariusNuc. commissuralis CajalNue. spinalis V
Fig. 1 14—Diagram the v iscera l afferent and efferent connections inthe medulla Ob longata , based on Fig . 7 1 ; compare a lso Figs . 77 and 86 .
The afferent roots and centers are ind i cated on the right side ; the efferent,on the left. Viscera l sensory fibers enter by the VI I nerve (pars intermed iaof Wrisberg
, VII pars . int. ) and by the IX and X nerves . Thesc root-fibersinclude both genera l v isceral sensory and gustatory fibers , a ll Of wh i ch enterthe fasciculus sol i tarius . (Fibers Of the IX and X nerves also enter thespina l V tract ; bu t since these are somati c sensory fibers from the auricularbranch they are not included in the d iagram . For further detai ls on thecompos it ion O f these cranial nerves see the tab le on pp . 146 ,On the left s ide O f the figure the genera l v iscera l efferent nuclei are ind i
cated by smal l dots and the Specia l v isceral nuclei by large dots . The
latter comprise the motor V nuc leus for the j aw musc les, the motor VI I
nucleus for the muscles related to the hyo id bone and the genera l facialmusculature, and the nucleus amb iguus supplying striated muscles of thepharynx and larynx by way Of the IX and X nerves . Three genera l visceralefferent nuclei are ind i cated—the dorsal motor nucleus Of the vagus underthe a la cinerea and the superior and inferior sa l ivatory nuclei . The su criornucleus (nue. sal . sup. ) supplies the sublingua l and submaxillary sa ivaryglands by way O f the VII nerve (pars intermed ia Of Wr isberg) , and the inferior nucleus (nuc . sal . i nf . ) supp l ies the arotid sal ivary gland by way Ofthe IX nerve. All Of the general visceraFcfferent fibers are preganglionicsym patheti c fibers (see p . 229 ) wh i ch end in sympatheti c gangl ia , whencepostgangl ioni c fibers carry the nervous im pulses onward to their respectivedestinat i ons .

246 INTRODU CTION TO NEU ROLOGY
part of the viscera l area (Fig. 43 , p . 1 1 1 ) O f fish bra ins , and in some speciesthes e centers are enormously enlarged , as in the carp (Fig. 136 p .
The pr im ary sensory center for the nerves of taste in the
nuc leus of the fasc i cu lus sol itar ius i s very intim ately connectedwi th al l of the m otor centers Of the m edu l la Oblongata for thereactions of mastication
'
and swal lowing, and also wi th the m otorcenters of the spinal cord . The ascend ing path from the pr imary gustatory nuc leus to the thalam us and cerebral cortex i swhol ly unknown in the hum an body . A gustato ry center i s
Fig . 1 16—The cutaneous gustatory branches arising from the geni culateganglion of the fac ia l nerve O f the catfish (Am ei urus m elas) , projected uponthe ri ght s ide of the body. Spinal cord and bra in stippled . The geni culateganglion
,its roots and cutaneous branches are drawn in b lack ; the branches
of th is nerve d istributed to the mucous l ining Of the mouth cavi ty are
omitted . Tas te- buds are found in a l l parts Of the outer skin to whi ch thesebranches are d istributed .
bel ieved to exi st in the cortex Of the gyrus h ippocampi near theanter ior end Of the temporal l obe. In fishes , where th is ascending gustato ry path is m uch larger , i t has been fol lowed to theroof Of the m idbrain and
,after a synapse here, to the regi on Of
the hypothalam us .Vi sceral E fferent Centers—The arrangem ent Of the viscera l
efferent nuc lei and nerve- roots Of the m edu l la Oblongata i s shownin Fig. 1 14 . There i s a l so a general vi sceral efferent componentOf the I I I c rania l nerve (Fig. 7 1
, p. 154,nuc . III . E whose
fibers pass out through thi s nerve to the c i l ia ry gangl i on in theorbit, which in turn connects with the intr ins i c m usc les Of the

THE VISCERAL AND GUSTATORY APPARATUS 247
eyebal l in the c ili ary process and i r i s . These fibers are i nvolvedin the m ovem ents Of accomm odat ion Of the eye for distance andin the regu lation of the diam eter Of the pupi l . The nuc leus Ofthe fasc i cu lus sol itar ius i s connected through the reticu lar form ation with all of the m otor centers of the m edu l la Oblongata forthe reactions Of m asti cation and swallow ing and for m any otherm ovem ents ; from th i s nuc leus there i s a descending tract to them otor centers Of the spinal cord , the tractus so litari o- spinal i s(F igs . 1 12 and There i s a lso a connection wi th the super ior and inferi or sal ivato ry nuc lei Of the V I I and IX nerves .The exc i tation Of the gustatory fibers Of these nerves by the
presence Of food in the m outh i s carr ied to the nuc leus Of thefasc icu lus soli tar ius and thence through the reti cu lar form at ionto the sal ivatory nuc lei , from whi ch the flow Of sal iva is exc i ted .
There are other connections with the m otor centers Of the spinalcord through the descending fibers Of the fasc icu lus so l i tar ius
,
som e Of these fibers crossing to the opposite s ide in the vi c ini tyOf the comm i ssural nuc leus Of Caj al (Fig.
Summary—The cerebro- spinal vi sceral system s fal l into ageneral group related per ipheral ly to the sympatheti c nerves anda Spec ia l group independent Of the sym patheti c .
‘
The secondgroup inc ludes the apparatus for taste and probably for sm el l .The centra l innervation Of the viscera i s partly from the spinaland m i dbrain regi ons, but ch iefly from the visceral area of themedull a Oblongata . The heart and blood-vessel s have a doubleinnervat ion derived from both the spinal and the bu lbar vi sceralcenters , and the nervous contro l Of the organs of c i rcu lat ion i svery complex. Respi ration in l ower vertebrates i s effected bystric tly visceral structures and is contro l led from the visceralarea of the medu l la Oblongata . In mamm als the m usc les ofordinary respi rat ion are al l of the som ati c type, but the centersOf contro l are retained in the visceral area Of the Oblongata . The
sensations related to the digestive tract are served ch iefly(though not exc lusively) by the vagus . There are spec ia l sal ivatory nuc lei related to the VII and IX crania l nerves . The
nerves Of taste are the V I I,IX
, and to a very l im i ted extent ( i nman) the X pai rs Of c ranial nerves . The prim ary cerebral gustatory center i s in the upper part of the nuc leus Of the fasc i cu lusso l i tarius, but the corti cal path i s unknown .

248 INTRODU CTION TO N E UROLOGY
L ITE RATU REAny Of the larger text-books Of phys io logy wi l l give further deta i ls Of the
visceral reactions . For a very brief and s im ple account of the circulatoryapparatus see the book by Sti les (pp. 1 18- 1 25 ) ci ted below . The experi
m ents Of Molhant have given us the most deta i led information regard ingthe viscera l functions O f the vagus and thei r centers in the m edulla Oblonata .gCAN NON ,
W . B . 1898 . The Movements Of the Stomach Stud ied byMeans Of the Rontgen Rays , Am er . Jour . Phys io l . , vo l . i , pp . 359—382 .
1902 . The Movements O f the l ntest ines Stud ied by Means Of theRontgen Rays , Amer . Jour . Phys iol . , vo l . vi , p . 25 1 .
19 12 . Perista ls is , Segmentat ion,and the Myenteric Reflex
, Amer.Jour . Phys io l . , vo l . xxx , pp . 1 14—128 .
CAN N ON ,W . B .
, and WASHBU RN , A . L . 19 12 . An E xp lanation ofHunger,Am er . Jour . Phys io l . , vo l . xxix , pp . 441—450 .
CARLSON , A . J . 19 12—19 15 . Contributions to the Physio logy of theStomach , Amer . Jour . Phys io l . , vols . xxxi—xxxv .
CUSHING , H. 1903 . The Taste Fibers and Their Independence Of the
N . Trigeminus , Johns Hopki ns Hospita l Bullet in ,vo l . xiv,
pp . 7 1—78 .
HE RRICK,C . J UDSO N . 1903 . The O rgan and Sense Of Taste in Fishes
,
Bul . U . S. Fish Comm iss ion for 1902 , pp . 237—272 .
1905 . The Centra l Gustatory Paths in the Brains of Bony Fishes,
Jour . Comp . N euro l . , vo l . xv ,pp . 375—456 .
1908 . On the Comm issura Infim a and I ts N uclei in the Brains O f
Fishes , Jour . Comp . N eurol .,vo l . xv i i i
,pp . 409—43 1 .
HE RTz , A . F . 19 1 1 . The Sensib i l i ty Of the A limentary Canal,London
,
Oxford Univers i ty Press .KA PPE RS, C . U . A . 19 14 . Der Geschm ack . porifer und central , zuglei ch
eine Ski zze d er phylogenet ischen Verand erungen in d er sensibelen VI I , IX ,
und X Wurzeln , Psychiat . en Neuro l . , Blad en ,pp . 1—57 .
MO LHANT, M . 19 10—19 13 . Le nerf vague : Etude anatomique et experimenta le, Le Névraxe , vo ls . xi ii—xv.
MO RG U L IS, S. 19 14 . Pawlow ’s Theory O f the Function Of the CentralN ervous System and a Digest of Some Of the More Recent Contr ibut ionsto this Subject from Pawlow ’s Laboratory
,Jour . Anima l Behav ior , vo l . iv,
pp . 362—379 .
PAWLOW , I . 19 13 . The Invest igation O f the Higher N ervous Funct ions , Br it . Med . Jour .
,vo l . i i for 1 9 13 , pp . 9 73—978 .
SHE LDON , R . E . 1909 . The Phylogeny O f the Facia l N erve and ChordaTym pan i , Anat . Record , vo l . i i i , pp . 593—6 17 .
1 909 . The Reactions O f the Dogfish to Chemica l St imuli , Jour.Comp . N euro l . , vo l . xix , pp . 273- 3 1 1 .
(1 IS
EIL ES
, P . G . 19 15 . The N ervous System and Its Conservation, Phi lae p 1a.
WILSON , J . G . 1905 . The Structure and Function of the Taste-buds ofthe Larynx, Bra in, vo l . xxviii , pp. 339—351 .

250 INTRODU CTION TO NE UR OLOGY
a sensation Of the qua l i ty typical for the organ in question , towhi ch there is added an agreeable or di sagreeable quali ty whi chmay be very pronounced , the disagreeable qual ity not beingpainfu l in the ordinary sense Of that term . Thi s m ixed qual i tyOf norm al sensations i s i l lustrated by certain Odors and savors,and on the agreeable side by certain sensat ions Of ti ckle and
warm th . F inal ly,som e i deational processes have an agreeable
or disagreeable qual i ty , and these, in turn,are very intim ately
related with the em otions and with estheti c and apprec iat ivefunctions of the m ost complex psych i c sort, as wel l as with questions Of habitual em otional atti tude and temperament .The superficial parts Of the body whi ch are m ore di rectly ex
posed to traum ati c inj u ry are, in general , m ore sens it ive to painthan are the deeper parts , and painfu l stim u l i here can be m oreaccurately local i zed . In som e parts , l ike the conj unctiva Of theeyebal l , where very sl ight i rri tation m ay ser i ously interferewi th the function , very gentle stim u l ation gives r i se to acute
pain ,and no other sensory quali ty may be present .
Su rgeons find that the brain mem branes are sensitive tomechan i cal inj ury, espec ial ly to stretch ing or pu l l ing. The brainsubstance i tself, however , i s qu i te insens i t ive to pain from ei thermechan ical or chem i cal stimulation . The deeper viscera Of thethorax and abdom en are insens itive to pinch ing, cutting with asharp instrum ent, or other m echani cal
,chem ical
,or thermal
stim u l i , though they are sensit ive to pains ar i sing from internald isorders, as in col i c (p. The vi sceral portions Of thepleural and per i toneal membranes are insens i t ive to pain , butthei r parietal port ions , form ing the i nnerm ost layer of the bodywal l , are Sens i t ive, and these pains can be accurately local i zed(Capps) .F rom these considerati ons i t appears that pain is an adaptive
function whi ch is present on ly where i t i s Of value to gi ve warning Of noxious influences liable to inj u re the body un less re
m oved . (See the excel lent discussion by Sherrington l
Schafer ’s Phys i ol ogy , vol . 11 , pp. 965
Pains of thi s sort are physiologi cal ly sim i lar to other exteroceptive sensations , that is , they have a defin i te local i zation andare external ly projected like other somati c sensations . But
other pains and discom forts (especially those related to the

PAIN AND PLE ASURE 51
visceral functions) and al l pleasu rable feel ings are devoid Of th isexternal proj ic ience and are exper ienced m erely as a non- l ocalized awareness Of m alaise or well-being (see p. They are
also m ore variable in relation to hab it,m ental attitude, fatigue,
and genera l health . Thi s latter group Of affective processes i s sodifferent from the ordinary sensations as to m ake i t desi rable tocons ider them separate ly, and , as wi l l appear beyond, they probably involve a qui te different ser ies Of nervous processes .There has been m uch controversy regarding the pathway
taken by painfu l impu lses through the spinal cord and brainstem
,and it is probable that this pathway is very complex.
All painfu l im pu lses carr ied by the spinal nerves , no m atterwhat the per ipheral sou rce, are d i scharged immedi ately uponenter ing the spinal cord into its gray m atter , and af ter a synapsehere the nerve-fibers Of the second order seem to take severalcourses . The m ost recent exper iments (Karplus and Kreid l
,
19 14) gO to Show that the as cending impu l ses Of painfu l sensibility i n the spina l cord Of cats fo llow a chain Of short neu rons,som e Of whose axons immediate ly c ross to the opposite side Of
the cord and som e ascend on the same side . These short fibersbe l ong to the fasc icu lus proprius system (p. and the nervousimpu l se i s at frequent intervals returned to the gray matter to
pass from one neu ron to another,and i t may cross the m idplane
repeatedly . This diffuse m ethod Of conduction appears to bethe prim i t ive arrangement . In the human spinal cord i t i s
probably present to a l im ited extent,but has been largely sup
planted by a m ore di rect pathway in the Spinal lemniscus , whose
prec i se l ocal i zation has been determ ined by the c l ini cal studies ofHenryHead and others (pp. 139 , Th is di rect path for fibersOf painfu l sensib i l i ty inc ludes axons Of neu rons Of the dorsal graycolumn , whi ch imm ediate ly c ross to the Opposite side of the cordand ascend di rectly to the thalam us . Inj u ry to th is path in thehum an body may cause complete insens i tivi ty to both superfic i a land deep pain on the opposi te side Of the body below the s i teOf the inj u ry , without loss Of general tacti le sens ibi l i ty . The
two m ethods of transm i ss ion Of impu lses Of painfu l sens ibi l i tyare shown diagramm atically in Fig. 1 17 .
I t may be assum ed that pain and an avo id ing react ion and pleasure and aseeking reaction have come to be ins tinctively assoc iated by natural selec
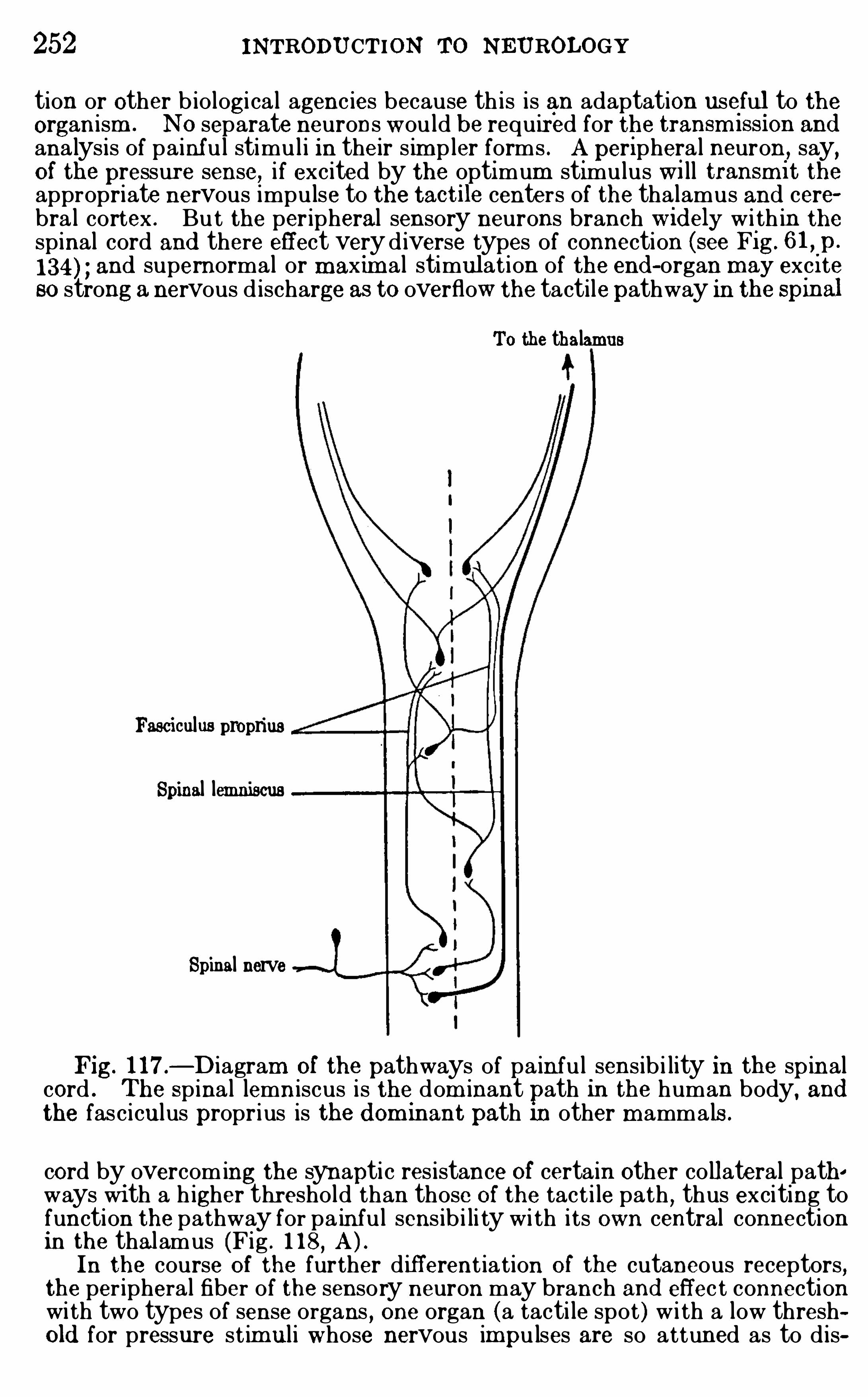
252 INTRODU CTION TO NEUROLOGY
tion or other b io logi ca l agenc ies because th is is an adaptation useful to theorganism . N O separate neurons would be required for the transm l ss1on and
ana lys is of painful stimul i in their s im pler forms . A periphera l neuron, say,Of the pressure sense, if excited by the optimum stimulus Wi ll transm l t the
appropriate nervous impulse to the tact1le centers of the thalamus and cerebra l cortex . Bu t the peripheral sensory neurons branch w 1d ely w i th in thespinal cord and there effect very d iverse types of connection (see F1g. 6 1 ,
.p.
and supernormal or maxim al stimul at ion of the end -organ may exc i teso strong anervous d ischarge as to overflow the tactilepathway in the spmal
To the thalamus
Fasciculus proprius
Spinal lemniscus
Spinal nerve
Fig. 1 17 .—D iagram Of the pathways of pa inful sens ib i lity in the spinal
cord . The Spina l lemn iscus is the.
dom inant path in the human body, andthe fasciculus proprius is the d ommant path 11) other m amma ls .
cord by overcoming the synaptic res istance of certain other co llateral pathways with a h igher thresho ld than those Of the tact i le path , thus exciting tofunction the pathway for pa inful sens ib i li ty wi th its own central connectionin the thalamus (Fig . 1 18, A ) .In the course Of the further d ifferent iation O f the cutaneous receptors,
the per ipheral fiber of the sensory neuron may branch and effec t connectionwi th two types of sense organs , one organ (a tacti le spot ) w i th a low threshold for pressure st im ul i whose nervous impulses are so attuned as to d is

254 INTRODU CTION TO NEU ROLOGY
ance of deep sensibili ty and the postu ral sensations, togetherw i th an exaggeration Of painfu l sens ib i li ty . The m od ificationsof pain and affective sens ibi l i ty are regarded by Head andHolm es as the m ost constant and character i sti c featu res of lesions of the lateral zone of the thalam us . A cute, pers istent,paroxysm al pains are always present , often into lerable and
yielding to no analgesic treatm ent . There i s a l so a tendency toreact excessively to unpleasant stim u li . Th is i s not necessa ri lyassoc iated with a lower ing Of the threshold Of stim u lation .
Deep pressure i s espec ial ly important here. The pain does notdevel op gradual ly out of the general sensation, but appearsexplosively . Th is pain has some factor to which the norm al halfOf the body i s not parti cu larly susceptib le. Therm al , visceral ,and other sense qual i ties are sim i larly affected . Ti ckl ing i s veryunpleasant on the affected side . The pleasu rable aspect Of
m oderate heat is accentuated on the affected side, yet thethreshold for heat i s never lowered . N ot on ly does the side ofthe body involved react m ore vigorously to an affective elementOf a stim u lus , but an overreaction can also be evoked by pu relymental states . The m ani festations Of th is increased suscepti
bili ty to states Of pleasu re and pain are stri ctly un i lateral .Assoc iated w i th th i s overreaction to painfu l stim u l i some l ossOf general sensation wi l l always be m ani fest on the affected sideof the body .
Pu re corti cal les i ons cause no change i n the thres hold to pain ,nor i s there the exaggerated affective qual i ty characteri sti c Ofthalam i c lesions . Head and Holm es assume that both the
thalam us and the cortex are concerned in consc i ous acti vity.
They say :
The most remarkab le feature in that group of tha lam i c cases w ithwhich we have dealt in this work is not the loss of sensation,
bu t an excess iveresponse to affect ive st im uli . Th is posi t ive effect, an actua l overload ing ofsensat ion wi th feel ing tone, was present in a ll our 24 cases Of thi s class .”Th is eff ec t is interpreted as d ue to the release of the inh ib itory or regu latoryinfluence O f the cortex ar is ing from the destruction of the as cending anddescend ing fibers between the thalamus and the cortex
,thus iso lating the
thalamus and a l lowing it to act to excess . These au thors add,s ince the
affect ive states can be increased when the thalamus is freed from corticalcontrol . we may conclude that the activi ty of the essent ia l thalam i c centeris mainly occupied wi th the affect ive s ide of sensation .
” “Th is conclus ion is
strengthened by the fact that stationary cort ical les ions, however extens ive,

PA IN AND PLEASUR E 255
whi ch cause no convuls ions or other s igns Of irr itation and shock , produceno efl
'
ect on sens ibi li ty to pa in . Destruction Of the cortex a lone does notdisturb the thresho ld for the painful or uncomfortab le aspec ts Of sensation .
”
Som e recent exper im ents by Cannon have revealed a veryintimate relat ion between em otion and som e Of the ductlessglands . The suprarenal (or adrenal ) glands , s ituated above thekidneys, secrete and pou r into the blood a rem arkable substanceknown as adrenalin or epinephr in . This substance exerts uponstructures wh i ch are innervated by sym patheti c nerves the sam e
effects as are produced by impulses pass ing along those nerves .The glands may them selves be exc i ted to activi ty by nervousimpu lses pass ing out through the sympatheti c nerves . Cannonhas shown that the em otions of fear
,rage, and pain exc i te these
glands to activ ity and cause the sec retion of adrenal in . The
blood Of a caged cat wh i ch has been torm ented by the barking Ofa d og will Show an i nc reased percentage Of adrena l in . The
addi tion Of adrenalin to the blood has the fu rther effect of causing l iberation Of sugar from the l iver into the blood to such anextent that sugar m ay appear in the ur ine (glycosu ria) and sugaris known to be the m ost avai lable form in whi ch energy can be
qu i ckly suppl ied to ti ssues wh ich have been exhausted by exer
cise. Adrenal in wi l l in th is and other ways act as an antidote tom uscu lar fatigue. I t also renders m ore rapi d the coagu lat i on Ofthe blood .
I f a m usc le i s fatigued , the threshold Of irr itab il i ty r i ses . I tmay r i se as m uch as 600 per cent . , but the average increase i sapproxim ately 2 00 per cent . I f the fatigued m usc le i s a l l owedto rest, the former i rr i tab i l i ty i s gradual ly regained , though twohours m ay pass before the recovery is complete . I f a sm al l doseof adrenalin i s adm in istered intravenously
,or the adrenal glands
are stim u lated to secrete,Cannon has found that the form er i r r i
tability Of the fatigued m usc le m ay be recovered wi th in th reem inutes . In th is way adrenal sec retion may largely restoreeffic iency after fatigue.
Fear and anger—as wel l as worry and distress—are attendedby cessation Of the contractions Of the stom ach and intestines .These mental states also reduce or temporar i ly abol ish the sec ret ion of gastr i c j u i ce. A d renalin inj ected into the body has thesame effect . Bes ides checking the funct ions Of the a l im entary

256 INTRODU CTION TO N EU ROLOGY
canal,adrenalin dr ives out the blood wh i ch , dur ing digestive
activity,floods the abdom inal viscera . Th is blood flows al l the
m ore rapi d ly and abundantly through the heart, the lungs, thecentral nervous system ,
and the l imbs .Cannon epitom i zes the account from whi ch the above has been
condensed in these words : “The em otional reactions above
descr ibed may each be interpreted, therefore, as m ak ing theorgan ism m ore effi c ient in the struggle whi ch fear or rage or
pain may involve. And that organ i sm whi ch , with the aid Of
adrenal secretion,best m obi l i zes i ts sugar , lessens i ts m uscu lar
fatigue, sends its blood to the vital ly important organs, and
provides against ser ious hem orrhage, wi l l stand the best chanceOf su rviving in the struggle for exi stence.
The preceding account inc ludes a summ ary Of some of them ost securely establ i shed facts regard ing the per ipheral andcentral nervous mechan i sm s of painfu l impressions and the
physi ology Of the em oti ons , together with a theoreti cal interpretation Of the apparently twofold nature of pain as a specificsensation and as a component Of the general affective state Of thebody as a whole. The m ore general questions concern ing thephysi ological processes related with pleasu rable and unpleasantexper ience and the affective l ife in general are sti l l m ore di fficu l tof analysis . I t seem s probable that pain ,
unpleasant and
pleasurable feel ings , em otion, and , in short , the enti re affectivel ife are very intim ate ly related on the neuro logical s ide.
Many physio logica l theories Of pleasure-pain have been e laborated , forthe most
fpa
rton very slender Observat ional grounds . I t has been suggested
that the exor movements Of the body are associated w i th pa in, the extensor
movements w i th pleasure ; that constructive metabo lism is pleasurab le,destructive metabo li sm d isagreeab le ; that heigh tened nervous d ischarge ispleasurab le, and the reverse (some form O f inh ibition or of antagonisticcontraction) is unpleasant . Some ho ld that pain an d unpleasantness ord isagreeab leness are d ifferent in degree only, not in kind . O thers regardpain as a true sensation , but d isagreeab leness and pleasure (affective ex
Eeri ence) as belonging to a d ifferent category wh ich is non- sensory . In thetter case the affec t ive experience may be neuro logica l ly related in someway Wi th the various sensat ions ( includ ing pa in) or the affect ive experienceand sensat i ons may be independent variab les w i th separate cerebralmechani sms . N one of these hypotheses , or many others wh i ch might bementi oned , are competent to exp la in satisfac tor ily a l l Of the known facts,though strong arguments can be adduced in support of each of them .
Our own view i s that pleasurab le and unpleasant experiences are not truesensations, that in the history of the psychogenes is Of prim i tive anima ls a

258 INTRODU CTION To N EUROLOGY
comes into consciousness at a ll (Of course, a large proportion of such reactionsare stri ct ly reflex and have no conscious s ign ificance) . Conversely, theim ped im ent to such d ischarge, no m atter what the occasmn, results in astasis in the nerve centers , the summation of st im uli and the developmentof a Si tuation Of unrel ieved nervous tens ion wh i ch is unpleasant unti l thetens ion is relieved by the appropriate adaptive reaction . Such a stas ismay be brought about by a confl i ct Of two sensory im pulses for the same
fina l common path (see p . by the di lemm a occasioned by the necessityfor d iscrim ination in an assoc iation center between two or m ore possib lefinal paths, by fatigue, auto- intoxi cation,
or other phys io logi ca l states wh i chlower the effi ciency Of the centra l m echanism , and by a variety Of othercauses . The unreli eved summation of st imuli in the nerve centers , invo lv ingstas is
,tens ion, and interference w ith free d ischarge Of nervous energy, gives
a feeling O f unpleasantness wh i ch in turn (in the h igher types of consciousreac tion at least) serves as a stimulus to other associated nerve centers toparticipate in the reaction unti l finally the appropriate avenue for an
ad apt ive response is Opened and the si tuation is relieved . W ith the releaseof the tens ion and free d ischarge, the feel ing tone changes to a d istinctlypleasurab le qua l ity (see C . L . Herrick
,
The fact that the pr im itive pain path in the spina l cord seems to fo llowa rather di ff usely arranged system of fibers in the fasciculus proprius, frequently interrup ted by synapses in the gray matter (Fig. 1 17) Wi th correspond ingly h igh res istance to nervous conduct ion ,
is perhaps correlated withth is general and d iffuse quali t of unpleasantness .Now , pain as a d ist inct an loca li zab le sensation has not been involved
in the si tuation described in the preced ing paragraphs . Pa in,cons idered as
a d ist inct sensation,was , however, born ou t Of this s ituation or d ifferentiated
from i t . Certa in sensat iona l elem ents whi ch have a h igh protective valuefor the organism are natura lly most often invo lved in such a s ituat ion .
These are warning ca lls , and usua l ly necess i tate an interruption of theord inary business Of l ife wh i ch may be in process at the time the dangerthreatens . The free flow O f ord inary sensori-motor activ i ty is abrup t lychecked , and the organ ism suddenly stops and makes the necessary readjustment as qui ckly as may be . In the interest Of increas ing the rapid ityof th is avo id ing reaction
,wh ich
,O f course, is frequently Of v i ta l importance,
the pathways of the exterocep tive pain reactions are well developed andsegregated from the more d iffuse and poorly organi zed affective apparatuswhi ch we have just been cons idering . Thus arose pain nerves (if such existseparate ly) and the pain tract of the spinal cord (whose anatomical d istinctness seems wel l estab l ished ) , and a lso perhaps a spec ia l mechanismfor pa inful react ions in the tha lam us . Sherrington has given a graph i cstatement Of the probab le h istory of th is process in the fo llow ing words(Schii fer
’
s Phys iology, vol . i i , p .
“The faci li ty Of path of these mo tor reflexes co l ligated to pain h ints at
their anti uity, or at thei r having been formed by some neural method particularly a le to , as i t were, make a good road . E ach reaction that employsa neural path seems to smooth i t by Sheer act of travel . Th is is true evenof s l igh t impulses—l ight traffi c—and more true O f heavy. Pa in reactionsare to be regarded as very heavy traffi c . Their im pressmns summate w i thpecul iar ease , take correspond ingly long periods to subside, and , to judgeby their inertia , move genera l ly masses of neural materia l relatively great .Such impress ions might wear a road w i th qui te especial Speed . Manysp inal reflexes imply, so to say, wel l-worn hab i ts based on ancient pa in

PA IN AND PLEASURE 259
reactions . One is almost emboldened to figuratively imagine them as con
nate mem ories Of the spina l cord . The m ajorit of them seem to be protective react ions that in organisms Of hi gh neura type are accompani ed bypm .
) 7 ’
But even in th is case the apparatus for pain is incapab le of acting asrapid ly as are those Of som e other sensat ions . If a sens i tive corn on the footis struck a sharp b low
,one wi ll Often feel a very d ist inct tact i le sensat ion an
appreciab le in terva l before the pa inful quahty is perceived , the latter, however, soon welling up into consciousness and obscuring the tacti le qua li tyenti rely. Th is is an i ll ustrat ion of the fact that even the h ighly protectiveexterocep t ive painful stimuli pass through a mechani sm of s lower reactiontime than the pr im ary exterocep tive sensations wi th whi ch they may beassociated .
We cannot here enter into a full d iscuss ion Of the larger quest ions centering about thephys io logi ca l correlates Of the h igher affective li fe, the emot ionsand esthet i cs . I t has O ften been po inted ou t that the conscious processesresul t ing from exterocep tive stim ulation tend to be d i rected outward , theattention being focussed on the externa l Objects giv ing rise to the st imuliwith a minim um Of personal reference . The deep sensat ions , both Of thepropr iocep tive and the interocept ive group , on the other hand , have a lessclearly defined loca l Sign and the menta l atti tude toward them is not one ofou tward ly d irected attent ion to the source Of the st imulus , but rather achange in the subject ive state and an a l terat ion.Of the genera l feeling toneOf the body as a who le. Under ord inary circum stances the visceral afferentand other deep nervous im pul ses do not come in to clear consciousness separately, but in the aggregate these complexes (Often termed as awhole comm on sensat ion) profound ly mod ify the genera l menta l attitude and equilibrium . The genera lized feelings of both the p leasurab le and the painful typeshare th is subjective reference with the common sensations . They are veryimportant factors in that sensory continuum wh i ch li es at the bas is O f them aintenance of persona l identi ty wh i ch the O lder psycho logists somet im esca lled the empir i ca l ego . Only the pains associated wi th the sharply localiz ed cutaneous sensat ion qua li t ies w ith a h igh adaptive va lue as warn ings igns Of externa l danger have a d ist inct periphera l reference, and even th is isless c learly defined than that of the accompanying sensat ions Of pressure,and SO forth . The deep pa ins are im perfec t ly loca li zed and havemore O f thegeneral subjective reference whi ch has j ust been mentioned , and all of thepleasurab le qua li t ies are Of th is type.
The s im pler affect ive types of exper ience, accord ingly, seem to be mostint imately associated wi th the “common sensation ” complex
,especia lly
wi th the vi scera l sensat ion componen ts O f th is complex . From th is i t hasbeen argued that the coarser emot ions , as well as the elementary feelings, arethe d irect express ion in consc iousness Of these visceral activi ties , that thewell-known viscera l changes associated w ith the emot ions are not the resul ts ,bu t the causes of the emot ions (Lange and Jam es ) . Th is hypothesis hasbeen attacked experim enta lly by Sherrington (see The Integrat ive A ct ionof the N ervous System , 1 906 , p . who found that cutt ing the afferentsym pathetic fibers from the abdomina l viscera in dogs made no apparentd ifference in the emot ional react ions Of the anim a ls ; but the experiments arenot very convincing, and the quest ion is probab ly too complex for so lut ionby so s im ple means as those here employed .
The probab i li ty is that we have here a circular type of react ion . Theinitial viscera l afferent im pulses , being heav i ly charged w ith affect ive qua li

260 INTRODU CTION TO N EU ROLOGY
ties and wi th a minim um Of Objec tive reference, excite w ith in the bra in,
probab ly in the med ia l tha lam i c nuclei , a genera l non- loca l ized pleasurab leor unpleasant feeling , a feeli ng Of well-being or ma la ise, as the case may be.
These thalamic receptive centers are in very int imate relation w i th thev iscera l efferent systems O f the hypotha lam us and a reflex response in thevi scera follows—a typica l or ani c Circuit. SO long as th is circuit involvesonly the v iscera and their tha amic centers the periphera l reference wi ll be ata minimum , and the fee ling remains an un local ized change in the affectiveconsciousness .The h igher emotional and esthet ic act ivi t ies are so charged w ith intellec
tua l content also as to requi re the participation Of the association centers ofthe cerebra l cortex . But no pleasure-pa in centers are known in the cortexand the evidence at present avai lab le seems to negat ive the presence Of suchcenters . The agreeab le or d isagreeab le components of the h igher emotiona lprocesses are very probab ly d ue to the colligat ion of tha lamic activ i tieswi th cortica l associationa l processes . In case these emot ional or estheti cprocesses are Of cort i cal origin,
that is, exci ted in the first instance by the
activ ity Of cort ica l as sociat iona l centers , the ir affect ive content may be d ueto the invo lvement Of the subcorti ca l p leasure-pa in apparatus in the associational process, and th is a paratus would , as above described , generateefferent im pulses from the re ated viscera l centers, thus caus ing the characteristic v iscera l m ovements , whi ch in turn would reinforce the viscera l activi ties Of the bra in centers
,and thus by a “back-stroke ”
action strengthenthe emotiona l content of the pr imary associationa l complex . Thus the comletion of the circular react ion may reinforce the affective cons ciousness soong as i t is Operat ive .
That pleas ure i s correlated w ith free d ischarge Of nervous energy is sug
gested further by the fact that in most Of the pleasurab le emotions and sent im ents there is present a large factor of reca ll Of previous experiences . Thees thetic enj oym ent O f a given s i tuation is in large measure proportiona l tothe wea lth of associated m emories incorporated with in i t , especially whenthese are recombined into new patterns . The pleasure experienced in listening to a complicated mus i ca l product ion like a symphony may be enhancedmany fo ld after one has become thorough ly fam i l iar w i th it, and sti ll m oreso if the l istener has h imself played it or parts of it .In conclud ing th is d iscussion of pleasure pain we note the fo llowing
paragraph from Sherrington ’s account of Cutaneous Sensations , alreadyreferred to (Schafer ’s Physiology, 1900 , vol . i i , p .
“Affec tive tone is an attribu te Of all sensation , and am ong the attribute
tones O f skin sensat ion is skin-pain . Affect ive tone inheres m ore intense lyin senses wh i ch refer to the body than in those wh i ch refer to the env ironment , that is , i t is strongest in the non-proj ic ient senses . It is , therefore,strongin the cu taneous senses , and . in them is inversely as their proj i cience,
there ore leas t in touch spots , m ore in therm a l spots , m ost in the SO—cal led‘pain-spots .
’
St im ul i evoking skin-
pa in are broad ly such as injureor threaten injury to the skin ; the skin may be sa id to have gone far towarddevelop ing a
espe
cia
lsense O f i ts own in jur ies . The centra l conducting
path concern w i th these skin fee l ings seems a s ide—path in to wh ich theim press10 ns from the various sk in spots embouch wi th various ease , thosefrom the ‘pa in spots
’
espec ially eas ily. The phys io logi ca l react ions connectedWi th th i s s ide-path are charac teri zed by tendency to ‘Summat ion ,
tendencyto
‘coo
llatera l irrad iation,
’ slow culm inat ion, and slow subs idence. TheyO ften invo lve wi th thei r own act ivity that of adjac ent sensory channels (as

262 INTRODU CTION TO N E UROLOGY
entiated , and pain takes i ts place am ong the other exteroceptivesenses . But even in man the thalam i c and visceral m echani sm sOf affective exper ience are preserved and gi ve a character i st i corgani c background to the ent ire consc i ous l i fe. I n the normalman these mechani sm s m ay function with a m in im um Of corti cal control
, giving the general feel ing tone Of wel l- being ormalaise
,or they may be tied up with the m ost complex cortical
processes , thus enter ing into the fabri c of the h igher sentim entsand affections and becom ing important factors in shaping hum anconduct .
LITE RATURECAN NON ,
W . B . 1914 . Recent Stud ies of Bod i ly Effects Of Fear,Rage,
and Pa in ,Jour . Phi los . Psych . Sci . Methods, vo l . xi , pp . 162—165 .
19 14 . The Interrelations of Emotions as Suggested by RecentPhys iologica l Researches , Am er . Jour . Psychol .
,vo l . xxv
,pp . 256—282 .
1 9 14 . The Emergency Funct ion Of the Adrenal Medulla in Pa in andthe Major Emotions , Am er . Jour . Physiol . , vo l . xxxi ii , pp. 356—372 .
19 15 . Bod ily Changes in Pain,Hunger, Fear, and Rage, N ew York
,
3 1 1 pages .CAPPS, J . A . 19 1 1 . An E xperimental Study of the Pain Sense in the
P leural Membranes , Arch . Internal Med ic ine, vo l . vi i i , pp. 7 17—733 .
HEAD , H.
,and HO LMES
,G . 19 1 1 . Sensory Disturbances from Cerebral
Les ions , Brain ,vo l . xxxiv
,pp . 109—254 .
HEAD , H., and THOMPSO N ,
T. 1906 . The Grouping of the Afferent Impulses W i th in the Spina l Cord , Brain,
vo l . xxix , p. 537 .
HE RRICK , C . L . 19 10 . The Summation- irrad iation Theory of Pleasureain. In The Metaphys i cs O f a N atural ist
,Bul . Deni son Univers i ty
c ient ific Laboratories,vo l . xv.
HO LME S, S. J . 19 10 . P leasure, Pa in, and the Beginn ings of Intell igence,Jou'
r . Comp . Neur . , vol . xx ,pp . 145—164 .
J AME S, W . 1890 . The Principles Of Psychology, New York, vo l . ii , pp .
442—485 .
1894 . The Physical Bas is of Em ot ions , Psych . R ev. ,vo l . i
, p. 5 16 .
KAR PLUS, J . P . , and KR E IDL , A . 19 14 . E in Bei trag zur Kenntnis d erSchmerz leitung iIn Rhekenm ark, nach glei chzei tigen Durchschneid ungenbeider R i ’i ckenm arkshalften in versch iedenen Hohen bei Katzen
,Pfluger
’
s
Arch iv , Bd . 158 , pp . 275—287 .
LAN GE ,C . 1887 . Ueber G em ii thsbewegungen . E ine Psycho-physio
logische Studi e,Leipz ig .
ME YE R, MAX . 1 908 . The Nervous Correlate O f P leasantness and Unpleasantness , Psych . Rev. , vo l . xv , pp . 201—2 16 . 292—322 .
SHE RRIN GTON , C . S. 1 900 . Cutaneous Sensations,in Schafer ’s Phys i
ology, vol . i i , p . 965 - 100 1 .
1906 . he Integrat ive Act ion O f the N ervous System, New York .
WATSO N , J . B . 19 13 . Im age and A ffect ion in Behavior,Jour . Phi los.
Psych . Sci . Methods,vo l . x . pp. 42 1—428.

CHAPTER XIX
THE STRUCTURE OF THE CEREBRAL CORTEX
THE preceding pages have inc luded a brief chapter on som e Of
the general b iologi cal princ iples under lying the differentiation Of
the structure and functions Of the nervous system,som e general
character isti cs Of the nervous t i ssues,a brief su rvey Of the
structure of the var ious great divi si ons Of the nervous system,
and final ly an analysi s Of the m ore important sensori -m oto rreflex c i rcu i ts . N early al l Of the mechani sm s h itherto consid
ered are concerned with the i nnate invariable types Of responserepresented in ,
the reflex and instinctive life of the organi sm(p. In the h igher m amm als , and espec ia lly in m an, the
individually acqu i red re lat ively variable types Of action, par
ticularly those whi ch are consc i ously performed,requ i re the
cooperation Of the cerebral cortex, and the followi ng chapterswi l l be devoted to a consideration Of the cortex
,i ts structure,
functions, evolution
,and biologi cal significance.
We have already comm ented (pp. 109,215 ) on the fact that the
cerebral cortex appeared later in vertebrate evolution than m ostOf the other parts O f the brain,
and that in general i t serves theindivi dual ly acqu i red and intell igent functions , in contrast withthe brain stem and cerebel lum ,
whi ch contain the apparatus forthe innate activities of the reflex type . The prim ary reflexcenters of the brain stem and cerebel lum ,
accordi ngly, are som e
tim es cal led the Old brain (palzeencephalon , see Fig. 45 , p.
whi le the cerebral cortex and those parts Of the brain stem whichdevel op as subsid iary to the cortex (such as the neothalam us, p.
163) are called the new brain (neéncephalon) . l
1 A review of the evolut ion of the brain and the phylogenetic origin Of
the cerebral cortex would lie beyond the limi ts Of th is work , for the li terature upon th is subjec t is very extens ive. The following papers may beconsu
éggd
)in the present context. (See also the b ib li ograph ies on pp .
1 59 ,
HE RRICK, C . JUDSON . 19 10 . The E volut ion Of Inte lligence and I tsOrgans , Science , N . S , vo l . xxxi , pp 7—18 .
SMITH, G . E LLIOT. The Arris and Gale Lectures on Some Prob
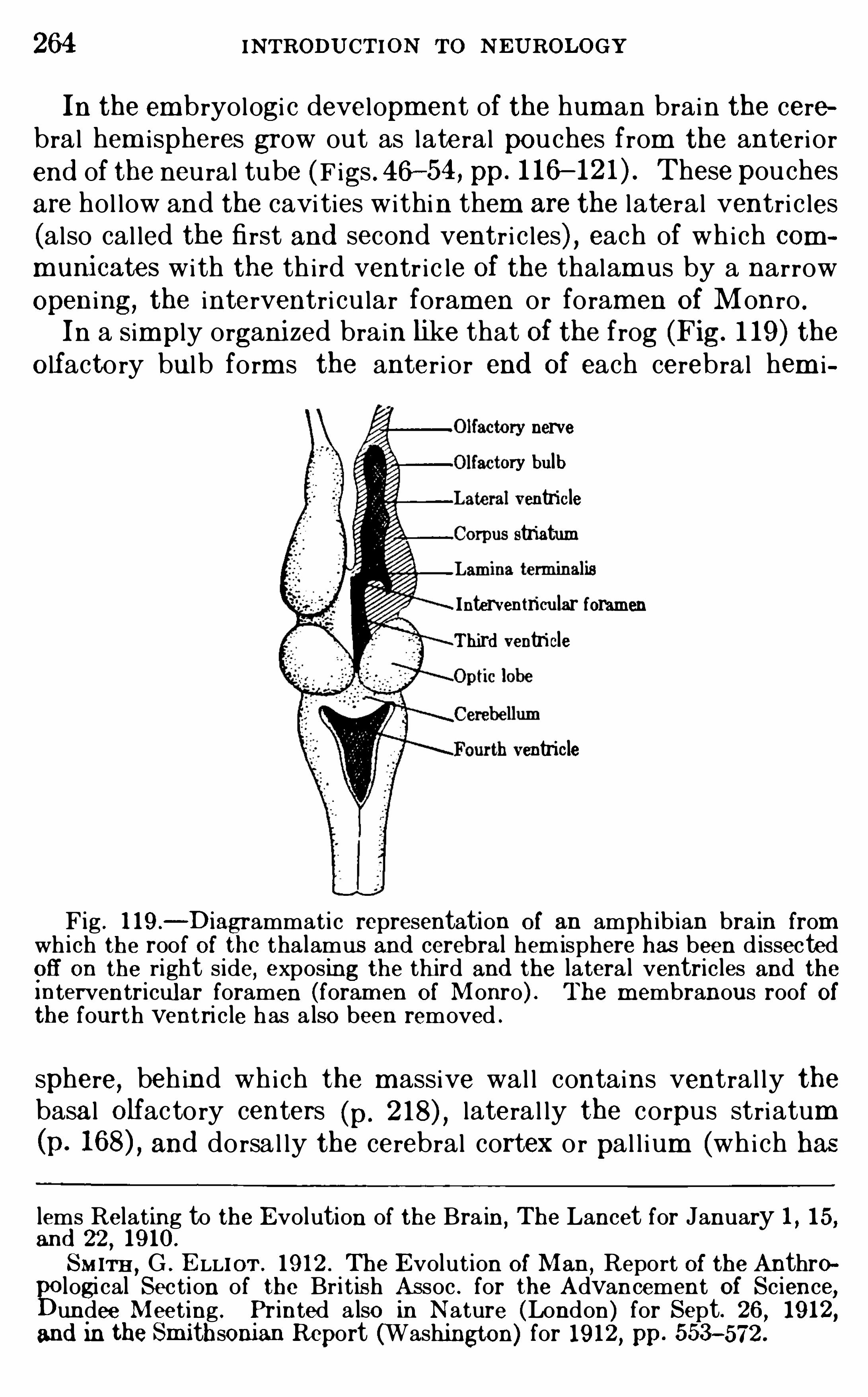
264 INTRODU CTION TO N EUROLOGY
In the em bryologi c developm ent of the hum an brain the cerebral hem i spheres grow out as lateral pouches from the anteri orend of the neural tube (F igs . 46—54 , pp. 1 16 These pouchesare hol low and the cavi ties within them are the lateral ventri c les
(also cal led the first and second ventr i c les) , each of wh ich communicates with the th i rd ventr ic le Of the thalam us by a narrowOpening, the interventri cu lar foram en or foram en Of Monro .
In a Simply organi zed brain like that Of the frog (Fig. 1 19 ) theO lfactory bulb form s the anter i or end Of each cerebral hem i
nervery bulbventriclestriatum
Interventricular foramenThird ventricle
ventricle
Fig . l l 9 .—D iagrammat i c representation O f an amph ib ian bra in from
wh i ch the roof Of the thalamus and cerebral hem isphere has been d issectedoff on the right s ide
,exposing the th ird and the latera l ventricles and the
interventricular foramen (foram en O f Monro) . The m embranous roof Ofthe fourth ventri cle has a lso been removed .
sphere, beh ind which the m assive wal l contains ventral ly thebasal O lfactory centers (p. lateral ly the corpus striatum(p. and dorsa l ly the cerebral cortex or pal l ium (whi ch has
lems Relating to the Evolution of the Bra in,The Lancet for January 1 , 1 5,
and 22, 19 10 .
SMITH, G . E LLIOT . 19 12 . The E volut ion O f Man ,Report Of the Anthro
logi cal Sec tion of the Br i t ish Assoc . for the Advancement Of Science,undee Meeting . Printed a lso in N ature (London) for Sept. 26 , 19 12,
Mi d in the Sm i thsonian Report (Washington) for 19 12 , pp. 553- 572 .

266 INTRODU CTION To N EU ROLOGY
primary factor in the higher m amm als has undoubtedly beenthe great inc rease in the superfic ia l area Of corti cal gray m atterwi thout a corresponding enlargement Of the sku l l .The hum an cerebral cortex i s som ewhat arbi trari ly divided
into frontal , temporal , parietal , and occ ipi tal l obes (Fig.
These lobes have no spec ial functi onal significance, but are di stinguished m erely for conven ience Of topographi c desc r iption .
Som e of the m ore important gyri and su lc i are nam ed on F igs .52 and 54 (pp. 1 19 and Between the temporal and frontallobes and under the lower end Of the lateral or Sylvian fissure i sa bu ried convoluti on , the i s land Of Rei l ( insu la) , whi ch i s seen insection in F igs . 79 and 80 (pp. 166 and The corti cal lobu leswhich cover the insu la are cal led Opercu la (Fig. 54, p.
Fig. 120—The lateral aspect Of the hum an brain,il lustrating the boundaries
of the lobes of the cerebral cor tex (of. Fig.
The wal ls Of the cerebral hem i spheres in the cort ical regi on are
very thi ck , the greate r part Of th i s th i ckness being occupied bywhi te matte r composed of nerve-fibers which effect vari ous typesOf connection with the neu rons Of the cerebral cortex. The
cortex i tsel f i s composed Of gray m atter and i s relatively th in ,
its inner border being m arked by a broken l ine in F igs . 79 and 80.
The subcort i cal whi te m atter contains three ch ief Classes offibers : ( 1 ) Corona radiata fibers wh ich connect the cortex withthe brain stem (Figs . 79 , Most of these fibers pass th roughthe internal capsu le and com pr ise the sensory and m otor projection fibers (pp. 165 (2) comm i ssu ral fibers Of the corpuscall osum and hippocampal comm i ssu re (F igs . 79 , 80) (3) assoc iation fibers , which connect different parts Of the cerebral cortex
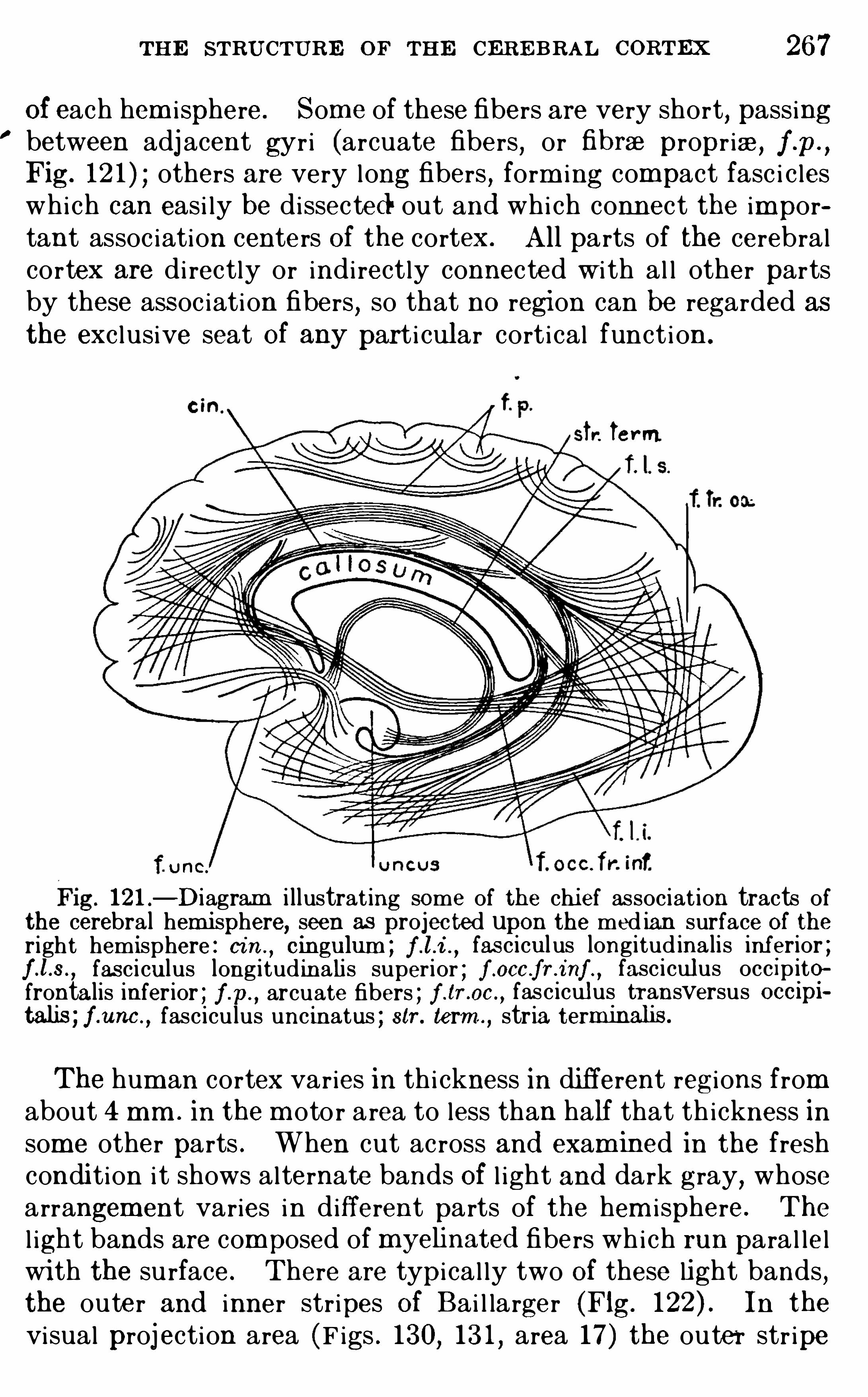
THE STRU CTURE OF THE CER E BRAL CORTEx 267
Of each hem i sphere. Som e of these fibers are very short, passingbetween adj acent gyri (arcuate fibers
,or fibrae propr iae, f .p.
,
Fig. 12 1 ) others are very long fibers , form ing compact fasc i c leswhich can easi ly be dissected out and whi ch connect the important assoc iation centers Of the cortex. All parts of the cerebralcortex are di rectly or indi rectly connected w i th al l other partsby these assoc iation fibers
,so that no region can be regarded as
the exc lusive seat Of any part i cular cort ical funct i on .
r. Term.
U HC.
121 —Diagram i llustrat ing some of the chi ef associat ion tracts ofthe cerebral hem isphere, seen as projected upon the med ian surface Of theright hemisphere . cin .
, cingulum ; f . fas cicul us longitud ina lis inferior ;fas ciculus longitud inalis super ior ; f mocc fr t
°
a fasciculus occ ipitofrontalis inferior ; f .p.
, arcuate fibers , f .tr . fasciculus transversus occipifasciculus uncinatus ; str . term ” stria ter
The hum an cortex varies in th i ckness in di fferent regi ons fromabout 4 mm . in the m oto r area to less than half that th ickness insome other parts . When cut ac ross and exam ined in the freshcondi tion i t shows alternate bands Of l ight and dark gray, whosearrangement varies in different parts of the hem i sphere. The
l ight bands are composed of myehnated fibers wh ich run para l le lwi th the su rface. There are typical ly two Of these light bands ,the outer and inner str ipes Of Bai l larger (Fig. In the
visual proj ection area (F igs . 130, 13 1 , area 17) the outer str ipe

268 INTRODU CTION TO N EUROLOGY
of Bai l larger i s greatly th i ckened by the opti c projecti on fibers,and here i t i s sometimes cal led the l ine Of Gennari . The por
tion of cortex exhibiting the fine of Gennar i i s cal led the areastriata .
The m ost character i sti c neurons of the cortex are pyram i dal i nshape, with the apex di rected toward the outer surface of thebrain and prolonged to form the pr inc ipal dendr i te . Sm al lerdendr i tes ar i se from other parts of the cel l body, and the axonar is ing from the base of the cel l body i s d i rected inward into thewhitem atter (F igs . 7 , 8, pp.
~42,
The cortex contains, m oreover, m any other types Of neurons, some of i rregu lar shape (polym orphi c or mu ltiform cel l s) and m any whose axons are short and
Fig . 122—Sect ions of the cerebral cortex, drawn nearly natural size andshowing the naked-eye appearance: 1 shows the layers as they appear inm any parts Of the cortex, and 2 shows the appearance Of a section from thev isua l cortex (area striata) from the neighborhood of the cal car ine fissure,wi th the conspicuous line of Gennari . (Af ter Bail larger .)
ram i fy c l ose to the cel l body without leaving the cortex i tself(Fig. 9 , p. These type I I neu rons probably assi st in thesumm ation and i rradiation Of stim uli (see p. Som e othertypes Of neurons are shown in Fig. 123 .
F igu re 124 illustrates a typ i cal arrangem ent Of the neu rons inthe postcentral gyrus (gyrus central i s poster i or Of Fig. 54 , p.
Most of the neu rons here shown send thei r axons inward topartic ipate in the form ation Of the white m atter and m ay discharge the i r nervous im pu l ses into rem ote parts Of the brain .
The endings of the afferent nerve-fibers which effect synapti cconnection with the neu rons here shown form a dense entanglement of fine unmyel inated fibers between the dendrites of theseneurons . These afferent fibers are not inc luded in Fig. 124 ; one
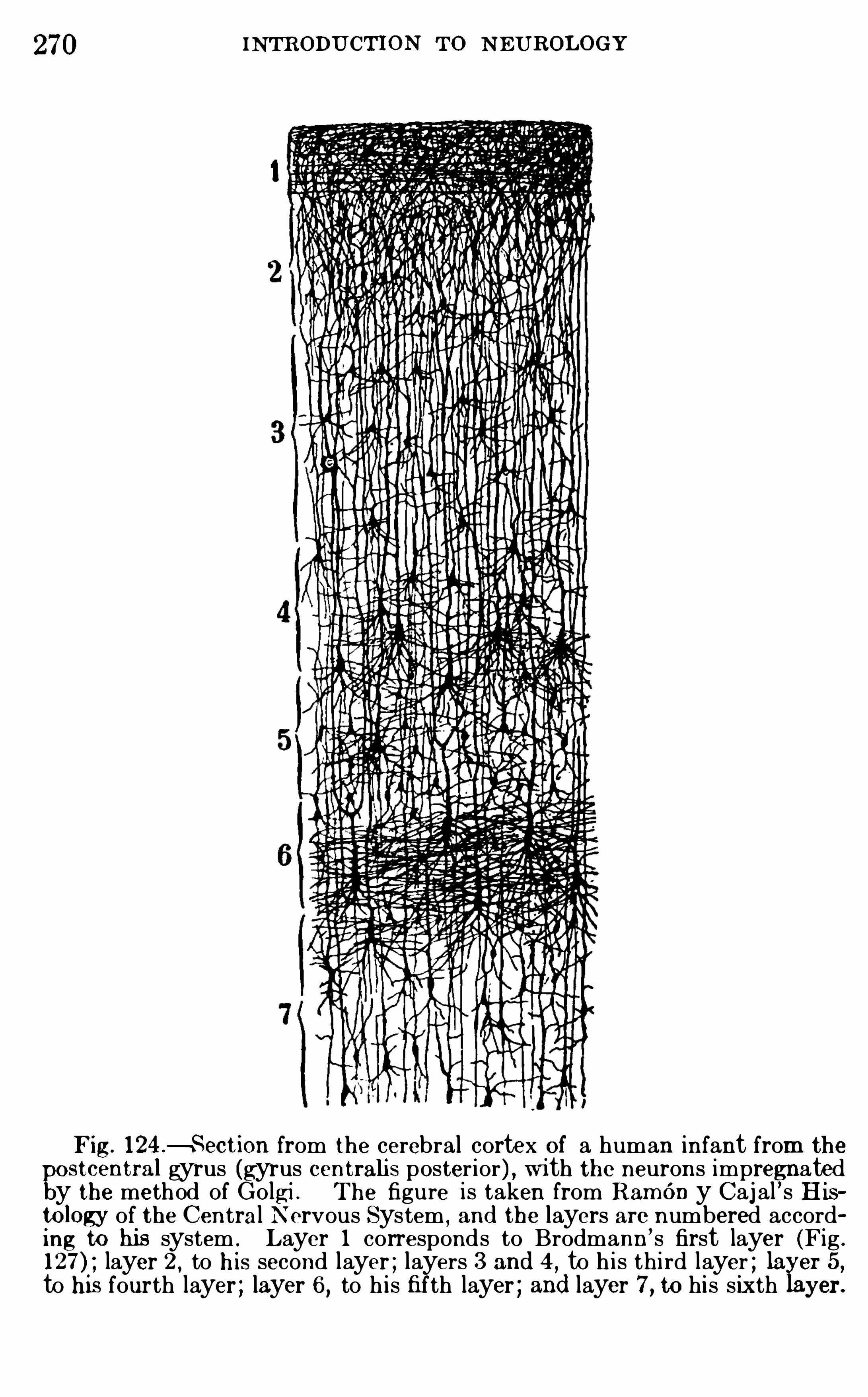
270 INTR ODU CTION TO N EU ROL OGY
F ig. 124 .fi Section from the cerebral cortex Of a human infant from the
stcentral gyrus (gyrus centrahs poster ior) , wi th the neurons impregnatedy the method of Go lgi . The figure is taken from RamOn y Caj al ’s Histology of. the Central N ervous System
, and the layers are numbered according to hi s system : L ayer 1 corresponds to Brodmann’ s first layer (Fig .
layer 2 , to h i s second layer ; layers 3 and 4 ,to h is third layer ; la er 5 ,
to his fourth layer ; layer 6 , to h is fifth layer ; and layer 7 , to his s ixth yer.

THE STRU CTURE OF THE CERE BRAL CoRTEx 27 1
pyram i dal cel l s wh ich are here widely expanded (see Fig. 8, p.
others end in dense arbor izations which c losely envelop
Fig . 125 .
-Section of the human cerebra l cortex from the precentralgyrus (gyrus centralis anterior) , i llustrat ing the free end ings Of the incom ingfibers . Th is regi on contains a large number Of cells Sim ilar to those shownin Fig . 1 24 ; bu t none O f the cells were sta ined in th is preparation, whi ch wasprepared by the method O f Golgi . At a and b are seen the termina l arborizat i on O f two indi v idual fibers . At 8 is a dense entanglement Of su ch terminal arborizat ions around the ce ll bod ies of the pyramidal neurons Of layer3 (Fig . C, D,
and E i l lustrate horizontal ly d irected nerve-fibers,from wh i ch the term ina l arbori zat ions shown in the upper part of thefigure ar ise. (After RamOn y Caj al .)
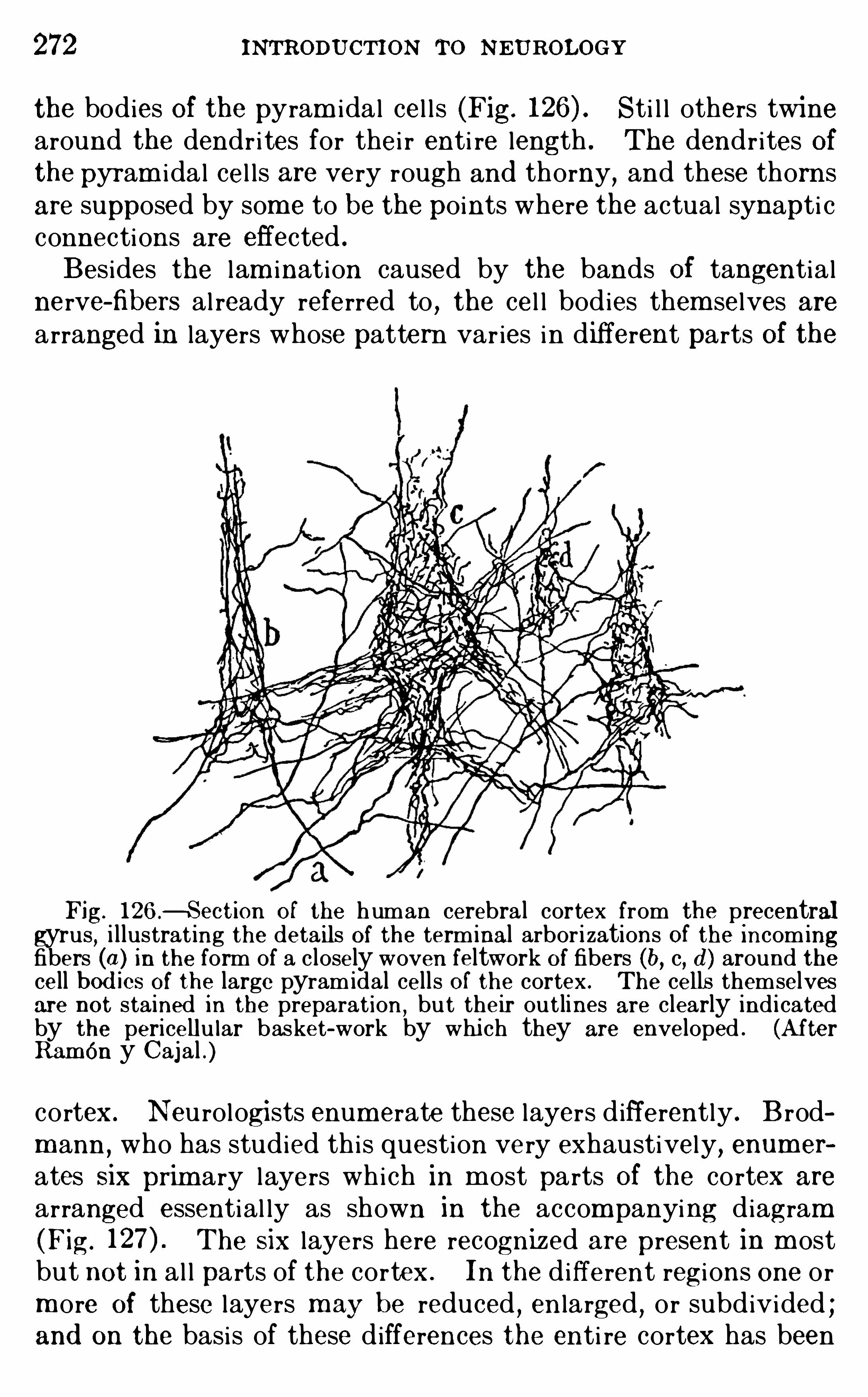
272 i NTRODUCTION To NEUROLOGY
the bodies Of the pyram idal cel l s (Fig. Sti l l others twinearound the dendr i tes for thei r enti re length . The dendr ites Ofthe pyram i dal cel ls are very rough and thorny, and these thornsare supposed by som e to be the points where the actual synapti cconnections are effected .
Besides the lam ination caused by the bands Of tangentialnerve-fibers al ready referred to
,the cel l bodies them selves are
arranged in layers whose pattern var ies in different parts Of the
Fig. 126 .fl Section O f the human cerebra l cortex from precentralgyrus, i l lustrat ing the details Of the terminal arbor i zations Of the incomingfibers in the form of a closely woven feltwork Of fibers (b, c, d ) around thecell bod ies Of the large pyramidal cells O f the cortex . The cells themselvesare not stained i n the preparat ion
,bu t their out lines are clearly ind i cated
by the pericellu lar basket-work by whi ch they are enveloped . (AfterRamOn y Cajal .
cortex. Neu rologi sts enum erate these layers differently . B rodm ann , who has studied this question very exhaustively, enumer
ates six prim ary layers which in m ost parts Of the cortex are
arranged essential ly as shown in the accompanying diagram(Fig. The six layers here recognized are present in m ostbut not in al l parts of the cortex. In the different regi ons one orm ore Of these layers m ay be reduced , en larged , or subdivided ;and on the basis of these differences the enti re cortex has been
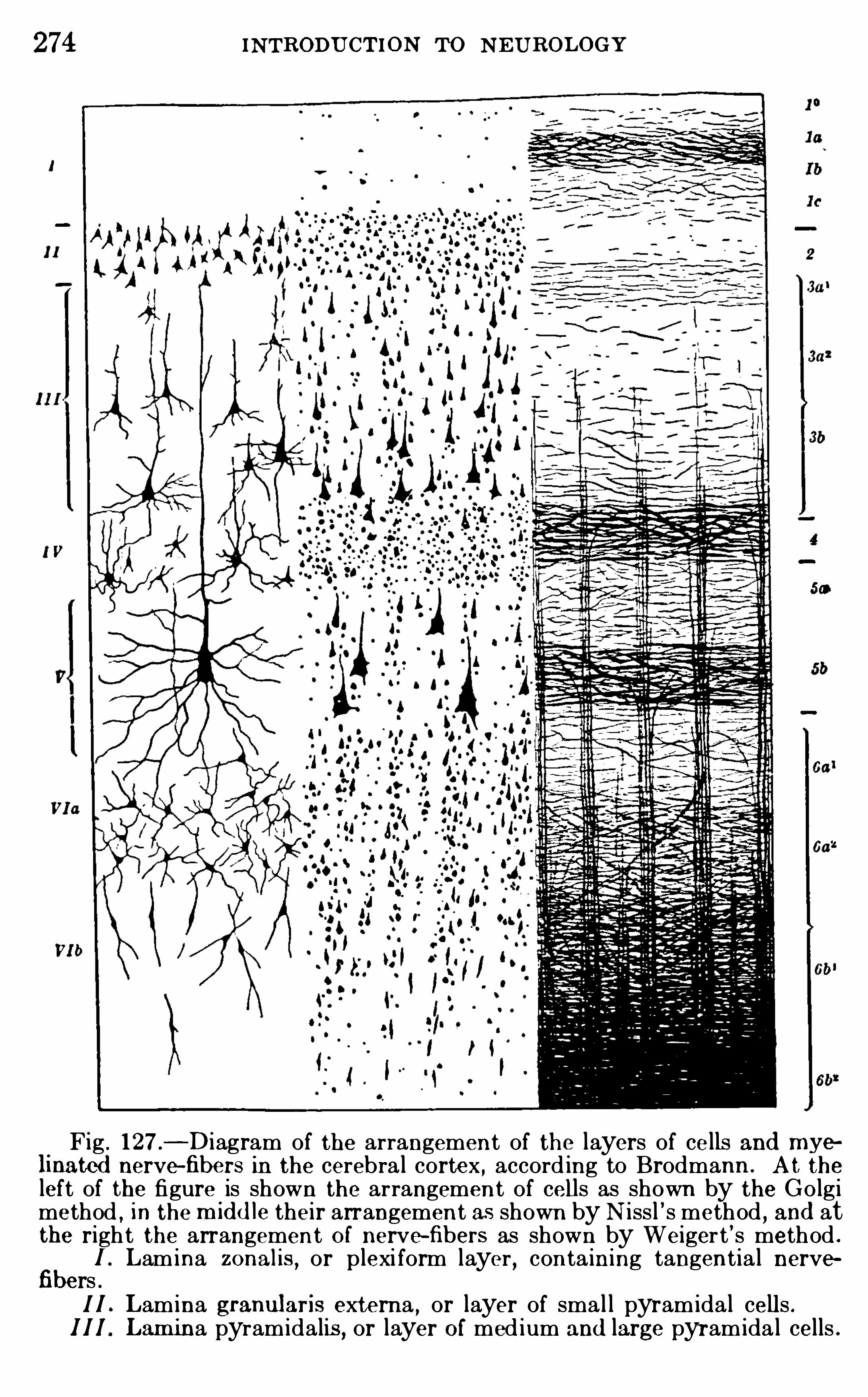
274 INTRODU CTION TO N E U ROL OGY
Fig . 127 ,
—Diagram of the arrangement of the layers of cells and myelinated nerve—fibers in the cerebra l cortex , accord ing to Brodmann . A t the
left of the figure is shown the arrangement of cells as shown by the Go lgimethod , in the m id d le their arrangement as shown by N iss l ’ s method , and atthe right the arrangement O f nerve-fibers as shown by Weiger t ’ s method .
fibe1 . Lamina zonalis , or plexi form layer , conta ining tangentia l nervers .1 I L amina granularis externa
,or layer of sma l l pyramidal cells .
1 1 1 . Lamina pyramidalis , or layer O f med ium and large pyramida l cells .

THE STRU CTURE OF THE CERE BRAL CoRTEx 275
whi ch i s character i sti c of the several cort ical regi ons in eachspec ies of m amm als , and that th is patte rn i s broadly sim i lar inal l of the higher m em bers of th is group of anim als .Data der ived from physio logical exper im ents m ade on dogs,
apes and other animals , and from the study of pathologi cal hum an brains have shown also that the difference in st ructural
pattern of the cort i cal areas is correlated wi th differences inthe functions perform ed by them . To these functional questions ou r attenti on wi l l next be d irected .
Summary—The cerebral cortex i s the organ Of the highestind ividual ly m odifiable functions, particu lar ly those of theintel lectual l ife. I t m atures late in both phylogeneti c and individual devel opm ent , and therefore has been cal led the neencephalon . In early developmental stages i t form s the roof of thelateral ventr i c le of each cerebral hem i sphere, but in the adu l thum an brain i t i s so en larged as to envelop m ost other parts ofthe hem i sphere. The cortex Of the two hem i spheres i s connected by comm i ssural fibers in the corpus cal losum and the hip
pocampal comm i ssure. The var ious regi ons of each hemi sphereare connected by a complex web Of assoc iat ion fibers, and som e
parts Of the cortex are connected wi th subcort ical regi ons byprojection fibers . The sensory proj ection fibers dischargeam ong the neu rons of the sensory projection centers, and them otor projection fibers arise from neu rons of the m otor projection cente rs . The interven ing assoc iati on centers are connectedwi th the projection centers and with each other by very intr icatesystem s of assoc iat ion fibers . The cortex i s lam inated by bandsof hor i zontal ly arranged nerve-fibers and by an arrangem ent ofi ts cel l s in layers . The pattern of th is lam ination var ies indifferent regions , and charts Of these structural ly defined regi onsare found to show a general correlation with the functionallydefined areas as physiological ly and pathological ly determ ined .
IV. Lam ina granularis interna , or inner granul ar layer , containing themedullated fibers of the external line of Baillarger ( in the Visua l area calledthe stripe of Gennari ) .
V. Lamina ganglionaris , or layer Of large cells, containing in the motorarea the giant pyramidal ce l ls or Betz cells , from wh ich the fibers of thepyram idal tract arise, and containing in m ost areas the medul lated fibers Ofthe internal l ine of Baillarger .
VI . Lam ina multiformi s, or layer Of polym orphi c cells .
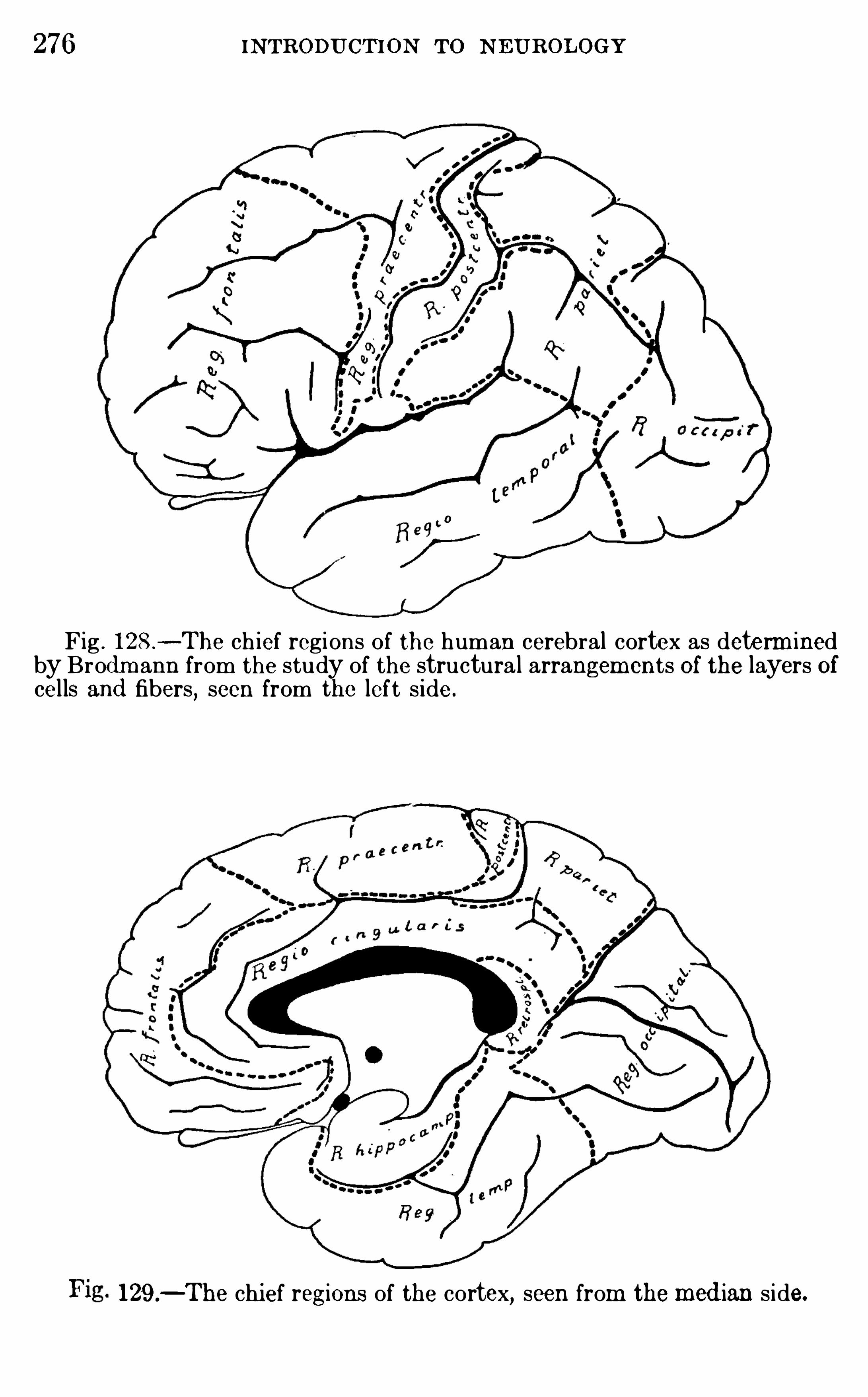
276 INTRODU CTION To N EUROLOGY
Fig . l 28 .
- The ch ief regions of the human cerebral cortex as determ inedby Brod m ann from the study of the structural arrangem ents Of the layers Ofcel ls and fibers, seen from the left s ide.
129 .—The chi ef regions of the cortex, seen from themed ian side.

278 INTRODU CTION TO NEUROLOGY
LITE RATUR EBO LTON ,
J . S. 19 10. A Contribution to the Local i zation of CerebralFunct ion,
Based on the Clini co-pathological Study Of Mental Disease,
Brain,vol . xxxii i , Part 129 , pp . 26—148 .
BOLTON ,J . S.
,and MOYES, J . M . 19 12 . The Cytoarch itecture of the
Cerebral Cortex of a Hum an Fetus of E ighteen Weeks, Brain, vol . xxxv.
BRODMANN ,K . 1907 . Die Kortexglied erung d es Menschen, Jour . f .
Psycho l . u . N euro l . , Bd . 10 .
1 909 . Verglei chende Lokalisationslehre d er G rossh irnrind e, Leipz ig .19 10 . Chapter entit led , Feinere Anatomie d eS Grossh irns, in Lewan
d owsky’
s Handbuch d er Neuro logie, Bd . 1, pp . 206- 307 .
CAMPBE LL , A .W . 1905 . Histo logical Stud ies on the Locali zation Of Cortical E mot ion, Cambr idge .
KAPPE RS,C . U . A . 19 13 . Cerebral Locali zation and the Si gnificance Of
Sulei , Proc . XVII Intern . Congress of Med ic ine, L ondon .
19 14 . Ueber das R in d enproblem und d ie Tendenz innerer Hirntei les ich (lurch Oberflachen-Verm ehrung statt Volum zunahm e zu vergrOsseren,
Fo lia Neuro-b iologi ca, Bd . 8,pp . 507—531 .
LRAMON Y CAJAL . 1900—1906 . Stud ien fi ber d ieHi rnr inde d es Menschen,
erpzrg.
SMITH,G . E LLIOT. 1907 . A New Topograph i cal Survey Of the Hum an
Cerebral Cortex , Jour . Anat. and Phys io l . , vo l . xl i .VOGT, O . 1903 . Zur anatomischen Gliederung d es Cortex Cerebri ,
Jour . f . Psych . u . Neuro l .,Bd . 2 .
1904 . Die Markreifung d es Kind ergehirns wahrend d er ersten vierLebensmonate un d ihre methodologische Bedeutung, Jena .

CHAPTER XX
THE FUNCTIONS OF THE CEREBRAL CORTEX
THE greatest divers i ty of V iew has prevai led and sti l l prevai l sregarding the m ethod of cort i cal function . That the cerebra lcortex i s concerned in som e way with the higher consc i ous functions i s c learly Shown by a large body Of exper im ental andc l in ical evidence .
The partia l or complete rem oval Of both cerebral hem i sphereshas been accom pl i shed in var ious Spec ies of an im als, from fishesto apes, and the changes in behavior carefu l ly studied . I nfishes and frogs the behavior i s but l i ttle m odified , save for thel oss Of the sense of sm el l , i f the thalam us i s left intact ; but if thethalam us also i s destroyed
,the anim al loses al l power Of Sponta
neousm ovement, Of feedingwhen hungry , etc .
,though i t wi ll sti l l
react to som e strong stim u l i in an apparently norm al m anner .The fundam ental reflexes of the spinal cord and brain stem are
but l i tt le m odified by th i s operation in frogs , save for the di sturbance Of the Olfactory and visual functions . The recentexper im ents of Bu rnett have, m oreover, Shown that frogs inwhi ch the cerebral hem i spheres alone have been rem oved are
somewhat m ore exc i table than norm al frogs (probably d ue to thel oss Of cortical inh ibitions) , and that Simple assoc iati ons easi lylearned by norm al frogs are in th is case impossible.
In the d og the loss of the cerebral hem i spheres alone leaves theanim al in a state of profound idiocy, though here also al l of the
pr im ary sensori -m otor reflexes (except the O l factory) rem ain ifthe thalam us i s un inj u red , and one such an im al operated on byGoltz l ived for eighteen m onths . During th is t im e, however, hehad to be artific ial ly fed
,for he had lost the abi l i ty to recogni ze
food when set before him,nor did he Show any of h is former signs
of intel l igence. (These experim ents are summ ar ized inSchafer ’sPhysiology, vol . i i , pp. 698 ff ,
to whi ch the reader is referred for279

280 INTRODU CTION TO N E U ROLOGY
references to the l i teratu re ; see also the papers by Goltz , E dinger ,and Holm es
,c i ted in the appended B i bl i ography .)
E dinger and F ischer report the case Of a boy who l ived threeyears and n ine m onths , whose brain when exam ined after deathshowed total lack of the cerebral cortex with no other importantdefects . In th i s boy there was practical ly no devel opment insensory or m otor power or in intel l igence from bi rth to the t im e
Of hi s death . The infant fed when put to the breast, but Showedno Signs of hunger , th i rst, or any other sensory process . I t lay ina profound stupor and du r ing the fi rst year of l ife m ade no
spontaneous m ovem ents Of the l im bs . Unti l the tim e Of deaththere was l i ttle change from th i s condition, save for continualc ryingfrom the second year on . Th i s case Shows that the reflexfunctions of the hum an brain stem are norm al ly under cort i calcontrol to a m uch greater extent than are those Of any Of the
lower anim als , and that the absence of the cortex accordinglyinvolves a m ore profound di sturbance of the subcortical apparatus (see p.
About a hundred years ago Gal l and Spurzheim exam i ned thebrain
,form of Sku l l
,and physiognom y of m any persons whose
m ental character i st ics were m ore or less fu l ly known,and
reached very defini te conc lus ions regarding the l ocal i zationwi th in the brain of parti cu lar m ental facu l t ies
,such as benevo
lence, wit, and destructiveness ; they c laimed,further , that the
si zes Of these spec ific parts Of the brain (and hence thei r relativephysi ological im portance) can be determ ined by study of theexternal configu rat ion of the Sku ll . Many valuable observationswere accum u lated by these m en and thei r fol lowers , but the datawere so uncr i tical ly used and the psychologi cal basi s Of thei r
genera li zations was so fau l ty that the alleged sc ience of phrenology which they founded i s now whol ly d isc redited and i s professed today only by ignorant Charlatans .The great popu lar ity of phrenology fifty years and m ore ago
grew out of the fact that it served to give a pseud oscientificcharacter to m ethods of reading character , and hence of forecasting the future form erly c laim ed by astro logers and nec rom ancers .Modern psychology recogni zes that the m ind cannot be subdivided into any such distinct
“facu lt ies” as the phrenologi stsused, and m odern neu rology finds no basi s for the sharply
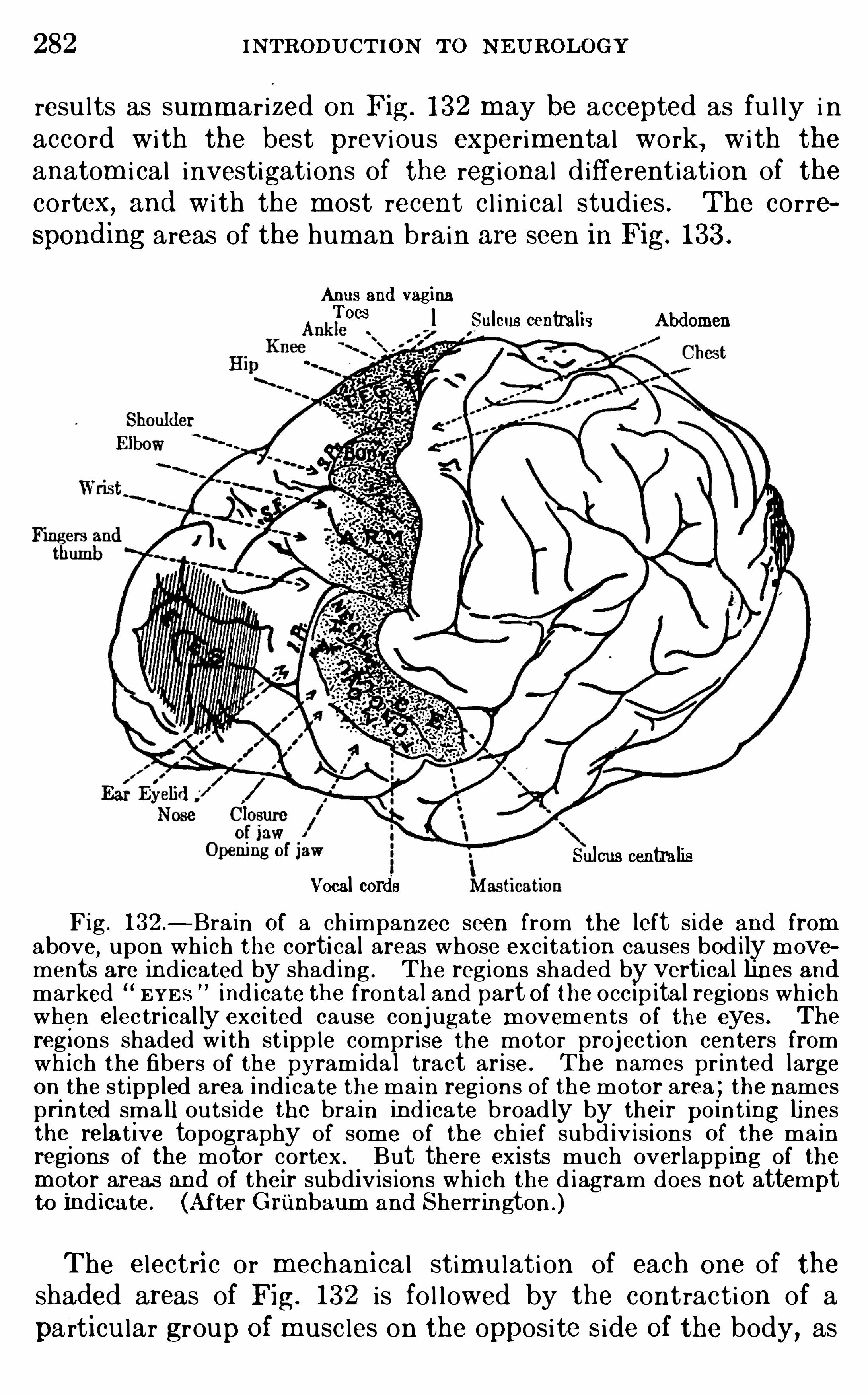
282 INTRODU CTION TO N E UROLOGY
resu l ts as summ ar ized on Fig . 132 m ay be accepted as fu l ly i naccord with the best previous exper im ental work, with the
anatom i cal investigations of the regi onal d ifferentiation of thecortex, and with the m ost recent c l ini cal studies . The correspond ing areas of the hum an brain are seen in Fig. 133 .
Anus and vaginaToes l Sulcus centralis Abd omen
Sulcus centralisVocal cords
Fig . 132—Bra in Of a ch impanzee seen from the left s ide and fromabove, upon wh ich the cortical areas whose excitat ion causes bod i ly m ovements are ind icated by shad ing . The regions shaded by vertica l lines andmarked “E YE S ” ind i cate the fronta l and part Of the occip ita l regions wh i chwhen elec trical ly exci ted cause conjugate m ovements of the eyes . The
regions shaded w i th st ipp le comprise the motor projec tion centers fromwh i ch the fibers of the pyramida l tract arise . The nam es pr inted largeon
.
the stippled area ind i cate the ma in regions of the motor area ; the namespri nted sma ll outs ide the bra in ind icate broad ly by thei r po int ing linesthe
.
re la t ive topography of some of the ch ief subd iv isions O f the ma inregi ons of the moto r cortex . But there exists much overlapp ing of them otor. areas and Of their subdiv is ions wh i ch the d iagram does not attemptto Ind i cate . (After G riinbaum and Sherrington .)
The electri c or mechani cal stim u lation of each one of theshad ed areas of Fig. 132 i s fo l lowed by the contracti on of apart i cu lar group Of m usc les on the opposi te Side Of the body, as
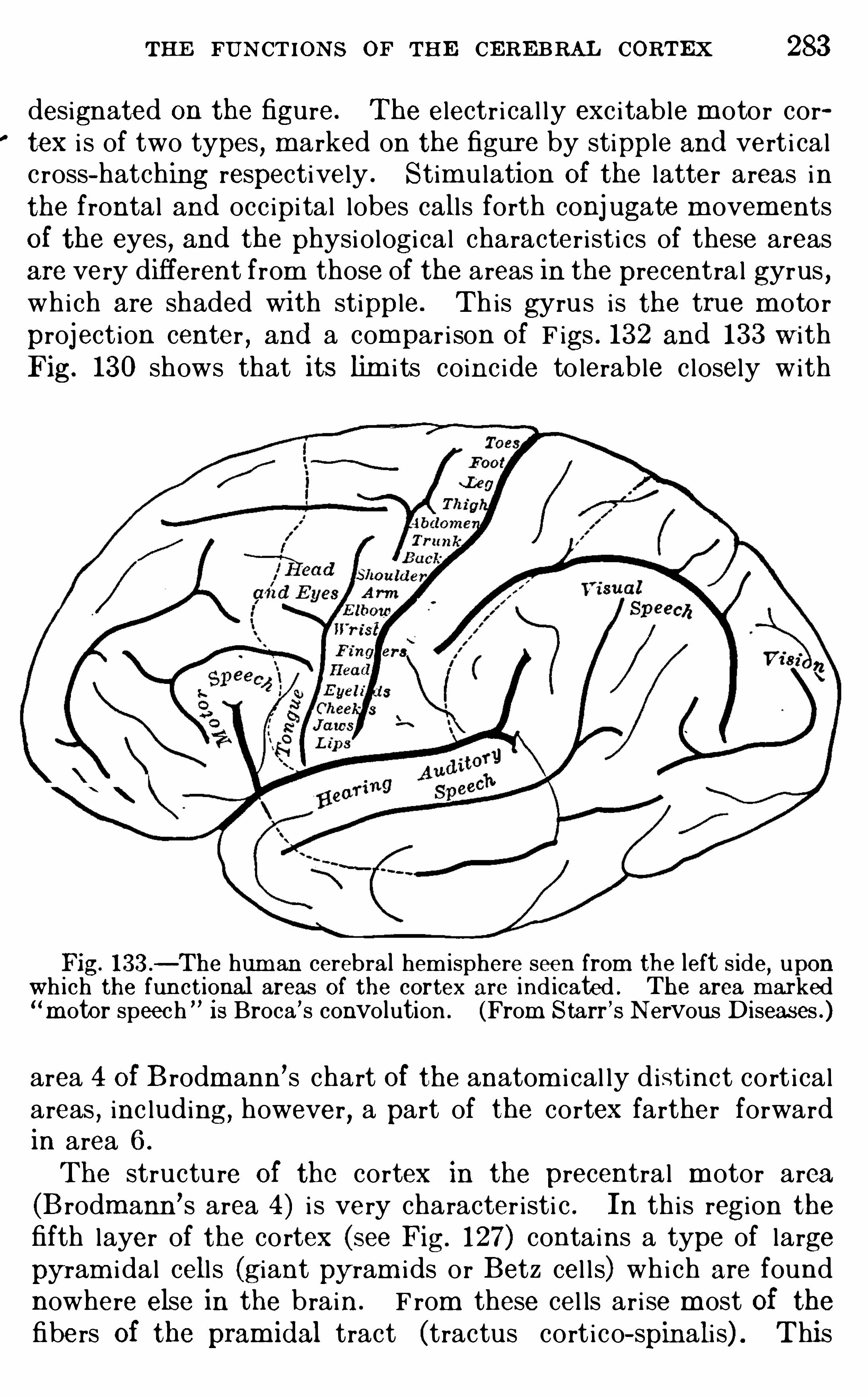
THE FU N CTIONS OF THE CEREBRAL CORTEX 283
designated on the figure. The electr i cal ly exc i table m otor cortex i s of two types, m arked on the figure by stipple and vert i calc ross- hatching respectively . Stim u lation Of the latter areas i nthe frontal and occ ipi tal lobes call s forth conj ugate m ovem entsOf the eyes
,and the physi ological character i st ics of these areas
are very different from those of the areas in the precentra l gyrus ,which are shaded wi th stipple. Th is gyrus i s the true m otor
project ion center , and a compar i son Of F igs . 132 and 133 w i thFig. 130 shows that its lim i ts coinc i de to lerable c losely with
Fig. 133 ._ The hum an cerebral hemisphere seen from the left s ide
,upon
wh ich the funct ional areas Of the cortex are ind icated . The area marked‘m otor speech ” i s Broca ’s convo lution . (From Starr ’SN ervous Diseases .)
area 4 Of B rodmann ’s chart Of the anatom i cal ly d istinct cort icalareas , inc luding, however, a part of the cortex farther forwardin area 6 .
The structu re Of the cortex in the precentral m otor area(B rodm ann
’s area 4) i s very character isti c . In th is region thefifth layer of the cortex (see Fig. 127) contains a type of largepyram i dal cell s (giant pyram i ds or Betz cel l s) whi ch are foundnowhere else in the brain . F rom these cel ls ar i se m ost Of thefibers Of the pram id al tract (tractus cort ico- spinali s) . This

284 INTRODU CTION To N EU ROLOGY
connection has been proved in several ways in addit ion to thedi rect physiologi cal exper im ents by electr i c stimu lation al readyreferred to . F i rst
,i f th is area Of the cortex (and a portion of
area 6 in front of i t) i s destroyed , the enti re pyram i dal tract wi l ldegenerate, a resu l t whi ch fol lows from the destruction of noother part of the cortex. Conversely, if the pyram i dal tract isinterrupted , the giant pyram i dal cel l s of thi s area are the on lyneurons of the cortex to give c lear pictu res of chrom atolysi s ofthei r chrom ophi l i c substance . In the th i rd place, these gi antcel ls of the hum an cortex have been counted, and a count of thenumber of fibers in the pyram idal tract Shows that the num bersare in to lerably c lose agreem ent (nearly on each side ofthe body) . F inal ly
,a case of sc leroti c degeneration involving
alm ost the enti re cortex has been descr ibed by Spielm eyer , i nwhi ch these giant cel ls and the fibers Of the pyram idal tract aloneescaped inj u ry .
The sensory proj ection centers of the cortex have al so beendeterm ined physio logical ly , though thei r l im i ts are less prec i selyknown than are those of the m otor cortex. The olfactoryreceptive area has al ready been m entioned as compr i sed with inthe arch ipal l ium (hippocampus and hippocampal gyrus , see
p. on ly a part Of which is exposed on the su rface of thebrain (the regi o hippocampica of Fig. 129 ; areas 27 , 28, 34 , 35 ofFig. The visual projection center , whi ch receives fibersfrom the thalam i c opti c centers in the pulvinar and lateral geni cnlate body (pp. 165 , i s in the occ ipi tal regi on (Fig.
A rea 17 (Fig. 13 1 ) appears to be the chief center for the reception of these visual proj ection fibers, though the adj acent area 18partic ipates in th is function ,
these areas together compr i singthe area striata of the cortex (p. The auditory proj ectioncenter is in the upper part of the temporal lobe (area 41 , andprobably to som e extent area 42 also
,of Fig. The tactual
project ion center l ies in the postcentral regi on (Fig. 128 ; areas 1 ,2, and 3 of Fig . The parts of the cerebral cortex whi ch liebetween the sensory and m otor projection centers whi ch havej ust)
been enum erated are the assoc iat ion centers (see pp. 287,290
Within each general sensory sphere there i s a focal area whi chis exc lusively receptive in function, such as area 17 (Fig. 131 ) i n

286 INTRODU CTION To N EU ROLOGY
the postcentral gyrus, to the m otor cortica l centers Of theprecentral gyrus,and to other w idely separated parts of the cortex . The SI nIficance of thi sfac t is sti ll Obscure. That the postcentral gyrus is Of erent functionaltype from the precentral gyrus is Shown by the fact that motor proj ecti onfibers arise from the latter and not from the former, by the d ifferences Inanatomica l structure Of these regions , by a large amount of exper im entaland clini cal evidence wh ich shows that tact i le sens ib i lity is not lost by thedestruction Of the precentral motor areas, and final ly by d irect phys io logicalexperiment upon human subjects .Dr . Harvey Cush ing in Operating upon brain tumors In 2 cases In
whi ch the use of an anesthetic was prohib i ted by the cond i tion of the patient,exposed the postcentral gyrus and , with the patient
’s consent, electrical lystim ulated i ts sur face . The patients , who were fully conscious during theOperation,
reported d ist inct cutaneous sensat ions wh i ch were subjectivelylocalized as if coming from the skin of the hand . There were no m otorresponses from th is and adj acent parts of the cortex beh ind the centralsulcus
,though in the same cases , upon stimulation of the precentral gyrus,
m otor responses were Obtained wh i ch were accompanied by no sensat ionssave those wh ich came from the muscles dur ing their contract ion. In aprevious s im ilar case Dr. Cush ing ( 1908) Obta ined typical motor responsesfrom stim ulat ion (w ith the pat ient
’s consent) of the precentral gyrus in an
Operation wi thout anesthesia, and these responses were unaccompanied bypainful sensations .A very extens ive series of experiments involving the stim ulation and
extirpation of these cortical areas in apes, dogs, and other an im a ls supportsthe conclus ion that the postcentral gyrus is the great receptive center forcutaneous react ions of the general cutaneous system . Wh at may be thefunct ions Of those thalamic fibers wh i ch pass to the motor centers in frontOf the central fissure is unsettled . Poss ibly these connections are concernedin cortical reflexes of the proprioceptive system or acquired automatism s .
The m yel inated fibers of the cerebral hem i sphere m ature,that i s , acqu i re thei r myel in sheaths , at various stages in the
developm ent of the brain,som e of these system s of fibers appear
ing before bi rth and some after bi rth . Much investigation hasbeen di rected to the determ inat ion of the exact facts regardingthe sequence of developm ent of these fibers , and m any interesting theor ies have been devel oped regarding the s ignificance ofthese facts .
.Flechsig in a long ser ies of researches made the first thorough study ofthi s prob lem ,
and h is conclus ions have exerted a profound influence uponall subsequent theor ies of the funct ions of the cerebral cortex . He proposeda series of laws of developmental sequence (myelogeny) of the cort i cal fibers ,among whi ch two may bementioned : ( 1 ) The myelinated fiber tracts of thebrain d o not all m ature at the same t ime, and fiber systems wh i ch are ofhke functi on, that is , wh i ch are so connected as to perform special movements In resp
onse to exci tat ion, tend to mature at the same tim e. Th is isFlechsi g’ s und am ental myelogeneti c law,” wh i ch may be stated in th isform ,
The myelination Of the nerve-fibers Of the developing brain fol lows

I
THE FUN CTIONS OF THE CERE BRAL CoRTEx 287
a definite sequence such that the fibers belonging to particular functionalsystems mature at the same time. (2 ) A second law states that in thecerebral cortex there are two great funct ional groups of fibers wh ich m atureat di fferent tim es . One of these groups contains the project ion fibers , wh i chm ature ear ly, ch iefly before b irth ; the other group contains the associationfibers, wh i ch mature after birth . These groups are fur ther subd iv ided intosubsidi ary funct iona l systems, each of wh ich connects wi th a definite regionof the cerebral cortex, so that i t is possible to map the corti ca l areas in ac
cord ance wi th the sequence of development of the related myelinated fibers .There are
, accord ingly, two groups of cortica l areas in thi s scheme ; theprojection centers whose fibers mature early and the association centerswhose fibers mature late .
Figures 134 and 135 i ll ustrate the arrangement of these areas , the primary areas (project ion centers) being marked by double cross-hatch ing andthe associat ion centers by s ingle cross- hatching or unshad ed areas . Thenumbers printed on the charts ind i cate the approxim ate order in wh i chthe correspond ing parts acquire their myelinated fibers . I t wi ll be noticedthat Flechsig
’ s projection areas do not correspond exactly wi th those determ ined by the phys iological method and by the h istologi ca l study Of the adultcortex (Figs . 130, 131 , 132 ,On the bas is of hi s stud ies, Flechs ig elaborated a h igh ly Speculative
theory of the si gnificance Of the associat ion centers,wh i ch has been cri ticized
as a return to the o ld attempt to local i ze particular mental funct ions indefinite cortical areas . These crit icisms are not wholly justified ; nevertheless it is even yet premature to attempt so detai led an analysis of thecorti cal mechani sms of psych i c processes as Flechsig has elaborated . HisObservations on the facts of myelogeny, m oreover , have not been confirmedby more recent students of the question (Monakow, Vogt, Dejer ine, andothers) , though i t seems to be estab l ished that the sensory and motorprojection centers in general acquire myelinated fibers earlier than otherarts Of the cerebra l cortex . (Th is entire quest ion is cr i ti cal ly reviewedy Brodmann in Lewandowsky’s Handbuch d er N eurologie, Band 2 , pp .
234 The only conclus ion at present poss ib le is that the factors wh i chOperate in determining the sequence of myelinat ion Of the nerve-fibers O fthe bra in are exceed ingly complex
,and it is impossib le from the facts at
pfies
gnt known to formulate the laws of the myelogenetic developm ent of
t e rain.
Attention shou l d be cal led here to the fact that there arem anydifferent k inds of projection fibers
,that is
,fibers connecting the
cerebral cortex with the under lying structu res of the brain stemand spinal cord . Most Of these projection fibers , except thoseOf the O lfactory system , pass through the corona radiata and
internal capsu le of the corpus striatum . The m ost importantO f these projection system s are the great sensory radiations whi chdischarge their nervous impu l ses into the cortical centers ofvision, hear ing, touch , and sm el l
,as al ready descr ibed (the
exact course of the gustatory projection fibers has not been d eterm ined ) , and the great m otor system Of the pyram i dal tract
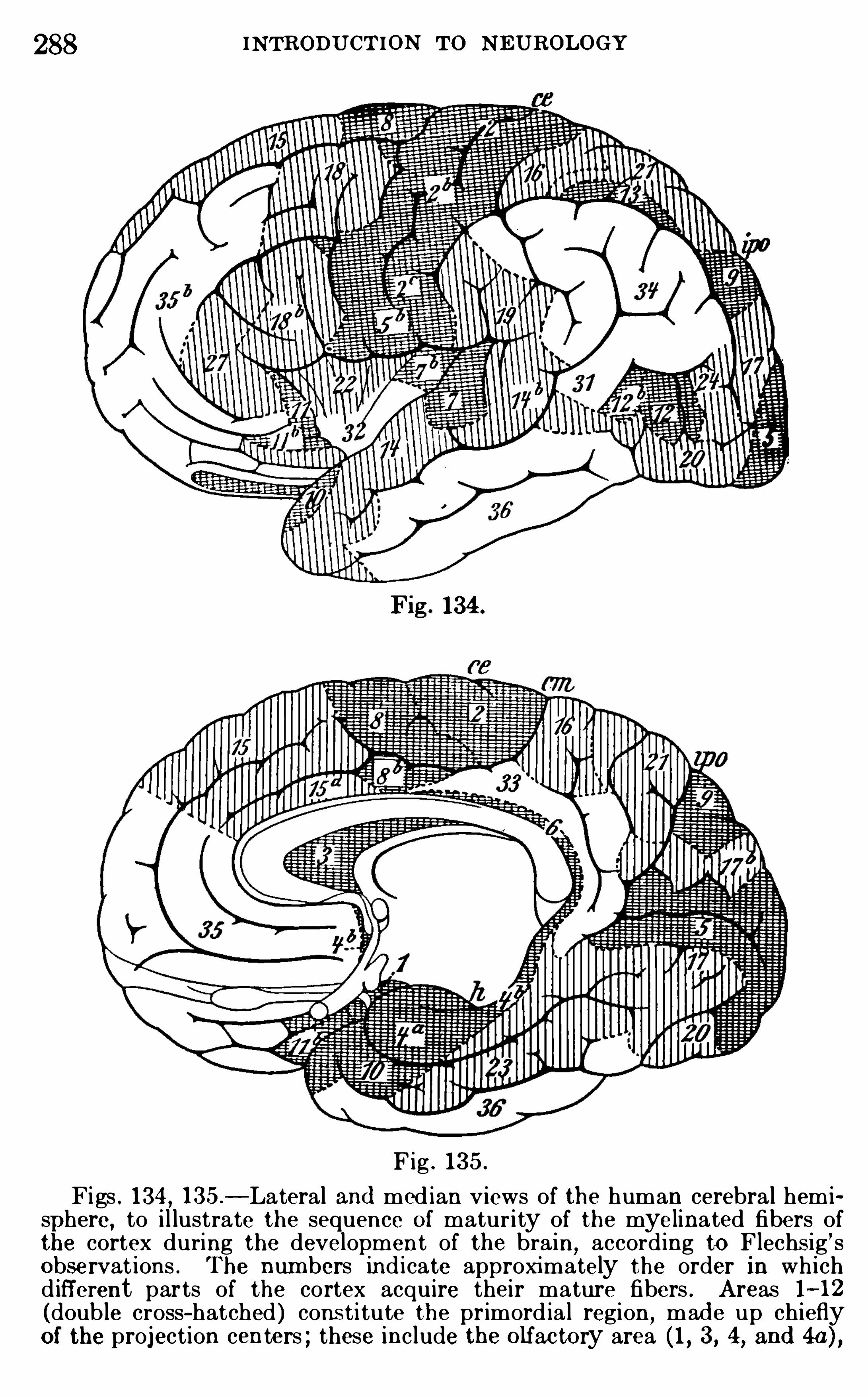
288 INTRODU CTION To NE U ROLOGY
Fig. 134 .
Fig . 135 .
Figs . 134 , 135—Lateral and med ian v iews of the human cerebral hemisphere, to i l lustrate the sequence O f matur ity of the myelinated fibers ofthe cortex during the development of the brain
,accord ing to Flechs ig ’s
Observat ions . The num bers ind i cate approximately the order in wh i chd ifferent parts of the cortex acquire thei r mature fibers . Areas 1—12(doub le cross-hatched ) cons ti tute the primord ia l region,
mad e up ch ieflyOf the projection centers ; these include the O lfac tory area ( 1 , 3, 4, and 4a) ,

290 INTRODU CTION To NEU ROLOGY
in general . Bolton believes that the granular layer ( layer IV of Fig. 127)marks an im portant boundary between fun ct ional ly d ifferent cort icalm echanisms . The infragranular portion Of the cortex is though t to beconcerned especially wi th the performance Of the s impler sensori-m otorreactions, part icular ly those Of the inst inctive type, wh i le the supragranularlayers serve the h igher associat ions m anifested by the capaci ty to learn byindividual experience and to develop the intellectual life.
The infragranular layers mature earlier in the development Of the brain ,
and they are the last to suffer degeneration in the destruct ion Of corti calcells in the acute dementias or insan i t ies . The supragranular layers(notab ly the pyram ida l neurons of Brodmann
’s th ird layer , Fig. 127 ) ma
ture later than any other layers . They are th inner in lower an imals and infeeb le-minded and imbeci le m en than in the norma l man ,
and they are the
first to Show degenerat ive changes in dement ia .
Th is doctrine is controverted by some other neurologists, but the evi
dence seem s to show that the supragranular pyram idal neurons are physiological ly the m ost imp ortant elements in the h igher associative processes ofthe cortex . In thi s connection i t is significant that the granular and infragranular layers are th i cker in the projection centers, wh i le in the associat ioncenters the supragranular layers of pyramidal cells are th i cker . But a ll ofthe layers in each region are very int im ately related , the processes Ofmost of the cells of the deeper layers extend ing throughout the th i ckness ofthe more superficial layers (see Figs . 123, 124 , 125 ) to reach the m ost superficial layer, and in the present state of our know ledge a fun ctional d iflerence between the layers cannot be said to have been estab l ished , save invery genera l terms .
$
I t m ust be borne in m i nd that the m ost signi ficant parts ofthe human cerebral cortex are the assoc iati on centers . Thesealone are greatly enlarged in the hum an brain as compared wi ththose of the higher apes . In the latter anim als the proj ecti oncenters are fu l ly as large as those Of man, the m uch sm al lerbrain weight be ing chiefly d ue to the relatively poor devel opment of the assoc iat ion centers .The data wh i ch we have summari zed in the preceding pages
have led to the m ost contrad i ctory theories as to the exactm ode of functioning of the assoc i ation centers . N eurologi stshave been prone, even up to the present tim e, to fal l into theer ror Of attem pting to find spec ific centers for parti cu lar m entalfunctions or facu l t ies . But the evidence at present avai lablegives sm al l prom i se of success in the search for such centers .I t i s , in fact , theoreti cal ly improbable that such discover ies wi l lever be m ade, for psychology today recogni zes no such m osai cof disc rete m ental facu lties as wou l d be impl ied in such a doctr ine.
The facts Of cerebral locali zation as c l ini cal ly and experi

THE FUN CTIONS OF THE CERE BRAL CORTEX 29 1
m ental ly dem onstrated,in them se lves and aside from any phi l o
sophi c theories based upon them ,contribute no evidence what
ever to a solution of the prob lem of a seat of consc i ousness or Ofpart icu lar mental “facu l ties .” That the proper functioning ofa given l ocus in the cortex i s essentia l to the executi on of a
given m otion or the experience of a given sensati on by no meansnecessari ly implies that the consc i ousness of the act i s locatedthere. The latter i s an enti rely independent problem whichm ust be separately investigated . I t i s not, then , the facts ofcerebral locali zation which can be cal led in question so m uch asthe interpretation Of these facts .The search for a single seat of consc i ousness, such as psychol
ogists and ph i losophers have so long sought , i s vain . The higherm ental processes undoubtedly requ i re the activ ity of assoc iation centers of the cerebra l cortex
, and the integr i ty of theassoc iat ional mechani sm as a whole i s essential for thei r ful lefii ciency. The cerebral cortex differs from the reflex centers ofthe brain stem ch iefly in that al l of its parts are interconnectedby inconceivably complex system s Of assoc iati ona l connecti ons,m any Of whi ch are probably acqu i red late i n l i fe under the influence of individual exper ience, and any combination Of whi chm ay,
iInd er appropr iate conditions Of external exc i tation and
internal physiologi cal state, becom e involved in any cerebral
process whatever .N evertheless , som e of these corti cal assoc iat ion paths are
structu ral ly m ore highly elaborated than others (Fig. 121 , p.
267 , illustrates the m ost di st inct Of these tracts) , and certaincom binations Of corti cal funct ions are, therefore, m ore l ikely tofo l l ow a given stim ulus than others . Thi s assoc iational patterni s doubtless part ly innate and part ly acqu ired . That there i s afair ly prec i se anatom i cal pattern of as soc iat ion tracts can be seenin any good di ssection of the cerebral hem i sphere, and that theelements of thi s pattern are related in defini te functional system swhi ch are spatial ly separate i s shown by num ber less c l ini cal observations in whi ch sharply c i rcum sc r ibed m ental defects arefound to be assoc iated with definite cerebral les ions . The
phenom ena Of aphas ia give the c learest il lustrations of theserelations .The term aphasia has comm only been applied to a variety of

292 INTRODU CTION TO NE UROLOGY
speech defects, but Hughlings J ackson extended the ConnotationOf the word to inc lude “a loss or defect in sym bo l i z ing relat ionsof things in any way . The lesion whi ch produces the defectaffects the assoc iation centers rather than the projecti on centers,for there i s no pr im ary sensory defect—no b l indness or deafnessor loss of general sensati on
—nor i s there any m otor paralysi s .The problem s connected with aphasia are very di fficult and
confused , and there i s by no means general agreem ent on ei therthe fundam ental physiologi cal m echani sm s involved in speech oron the nature of the les i ons wh ich produce the var i ous types ofObserved speech defects . The enorm ous l i teratu re relating tothis subj ect cannot be summ ari zed here ; see the text-books Of
physiology, physiological psychology , and c l in i cal neurology.
Les ions of the primary sensory or motor projection centers wil l not produce aphasia, for in these cases al l sensat ions or al l movements related tothe injure d parts are lost, whereas in aphasia only the correlations invo lvedin speech or other associat ional processes are impaired and all other sensorimotor correlations may be intact . Of course, the number of associationalpathways invo lved in the commun i cating of ideas by hear ing, read ing,speaking. and wri ting words is very large ; and the character O f the speechdefect w i l l depend in part upon the parti cular associational tracts affectedby the lesion and in part upon the effect of the les ion upon the general intelligence of the patient (di aschi s is effect, see p. The second factorseems to be exceed ingly variab le and has given rise to much controversy.
Distinct ive names have been given to the more im portant types Of speechdefect as clini cal ly Observed ; such as agraph ia or inab il ity to write correctly,aphemi a or inab i l i ty to u tter words , word-b lindness (alexia) or inab i l ity tocomprehend wri tten words, word-deafness or inab il ity to comprehendspoken words , and many others . Evidently an aphas ia may rasult frominjury to ( 1 ) a sensory associat ion area contiguous to the primary v isual oraud i tory projection centers (sensory types Of aphas ia) , or (2 ) to a motorassociation center contiguous to the motor projection centers for the speechmuscles (motor types) , or (3 ) to any Of the associational tracts connect ingthese associat ion centers .The second , or motor, type of aphas ia usually, though not invariab ly,
results from injury to the posterior part of the inferior frontal gyrus (seeFig . 54 , p . 12 1 ) O f the left hemisphere in right-handed persons and of theright hemisphere in left-handed persons . Th is relation was first di scoveredby Broca, and the area Of motor speech correlat ions (marked “motor SpeechIn Fig. 133 , p . 283 ) has s ince been term ed Broca ’s convo lut ion .
It should be rei terated that Broca’s convo lut ion does not lie in the excitable motor zone Of the cortex . Though the destruct ion of th is area may befo llowed by defects of speech
, the muscles Of the larynx , tongue, l ips, etc .,Involved in vocal i zat ion are not para lyzed . Th is case is typica l Of manyothermotor as sociation centers Of the cortex whose integrity ls essent ial forspeCIfic motor comb inat ions, though separate motor centers are present forall of the muscles invo lved In thos e m ovements .

294 INTRODU CTION To N EUROLOGY
impaired . In the course Of a few weeks or m onths a new equ ilibr ium m inus the lacking factors i s establ i shed and the patientvery rapi d ly improves . U l tim ately complete recovery m ay
occur,save for a perm anent residual defect whi ch resu l ts di rectly
from the l oss of the tissue destroyed .
The imm ediate shock- like interference wi th the act ivity Of
cerebral centers not di rectly affected by the lesion i s what vonMonakow means by diaschisi s . Upon the restoration of thenervous equilibr ium this transient diaschi si s effect i s whol ly o r
partially lost, and the residual symptom s Of defect give a fai r lyaccu rate pi ctu re of the intr insi c functions of the center d i rectlyattacked by the lesion . I t i s comm only assum ed that thereis also dur ing the process Of gradual recovery from such a cor t ical inj ury a certain capac ity for the compensatory devel opm entof other centers Of the sam e or the opposite cerebral hem i sphere,so that they learn to perform vi carious ly the functi ons of thel ost part .All functions Of the nervous system are fac i l i tated by repetition, and m any such repeti tions lead to an endur ing change inthe m ode Of response to stim u lation whi ch may be cal led physi ologi cal habit . Th is impl ies that the perform ance Of every reaction leaves som e sort of a residual change in the structure of theneuron system s involved . These acqu i red m odifications ofbehavior are mani fested in some degree by al l organi sm s (see
pp. 22, and th i s capac i ty lies at the basi s Of al l assoc iat ive
memory (whether consc i ously or unconsc i ously perform ed) andthe capac i ty of learn ing by exper ience. This m od ifiabilitythrough individual exper ience i s possessed by the cerebral cortexin h igher degree than by any other part Of the nervous system ;and the capac ity for reacting to stim u l i i n term s of past experience as wel l as Of the present si tuat ion l ies at the basi s of thatdoc il i ty and intelligent adaptation of m eans to ends which are
characteristi c of the higher m amm als . I t i s a fact Of comm onObservati on that those an imals whi ch possess the capac i ty for intelligent adj ustments of th is sort have larger assoc iati on centersin the cerebral cortex than do other spec ies whose behavior i scontro l led by m ore s imple reflex and instinctive factors , that i s,by inheri ted as contrasted with individual ly acqu ired organ i zation . This i s brought out wi th espec ial d i stinctness by a com

THE FU N CT IONS O F THE CE RE BRAL CORTEX 295
pari son of the brains of the higher apes with that of man (F igs .132, and of the lower races of men as contrasted with thehigher . In our own m ental l i fe we recogn i ze the pers istence Of
traces of previ ous exper ience subj ectively as m em ory , and m em
ory l ies at the bas i s of al l hum an cul ture. F rom thi s i t fo l lowsthat psychologi cal m em ory is probably a function of the assoc iation centers ; but i t m ust not be assumed that Spec ific m em oriesreside in parti cu lar corti cal areas , m uch less that they are pre
served as structu ral traces left in individual cortical cells , as hassometimes been done.
1
The simplest concrete m em ory that can appear in consc i ousness i s a very com plex process , and probably involves the activi tyOf an extensive system of assoc iation centers and tracts . Thatwhich persi sts in the cerebral cortex between the ini tia l experience and the recol lection of it is, therefore, in all probabili ty achange in the interneu ron i c res istance such as to alter the
physiologi cal equ i l ibr ium Of the component neu rons of som e
particu lar assoc iat ional system . What the nature of th is changemay be is unknown,
but it i s conceivable that i t m ight take theform of a perm anent m odification of the synapses between theneurons which were functional ly active during the ini t ial experience such as to fac i l i tate the active part ic ipation of the sam e
neu rons in the same physio logi cal pattern dur ing the reprod uc
tion .
That wh ich we know subjectively as the assoc iation of ideasmay, in a som ewhat sim i lar way, be pictured as involving neu rologically the discharge of nervous energy in the cortex betweentwo system s of neurons which have in som e previous exper iencebeen physiological ly united in some cort ical reacti on . I f
,for
instance, I heard a song of a m ocking bi rd for the fi rst tim e lastyear whi le walking in a rose garden,
upon revis iting the gard en I may recal l the song of the bi rd . Here the sight of the
garden (a h ighly complex apperceptive process involving m anyassoc iation tracts) actuates neuron system number one dom inated by present visual afferent impu lses , and the assoc iation
1 Thes e res idua of past cerebral act ivi ties form the bas is of those characteristi c ‘‘bra in di sposit ions ” wh ich are important fac tors In each personali ty. They have been term ed “
engram s” by Semon and “neurograms ’ ' by
Morton Pr ince (see Pr ince, The Unconsc ious , Chapter V, New York,

296 INTRODU CTION TO N E U ROLOGY
tract leading to neuron system number two (the auditory com
plex establ i shed last year when the songwas heard) has a l owered
physiologi cal res istance by vi rtue Of the previous col locationwith system number one, and I rem em ber the song (see p.
I t Shou l d be emphasi zed that the m echan i sm Of assoc iationhere suggested i s pu rely theoretical ; we have no sc ientific evidence regarding the detai ls of such physiological processes . Buti t can be confidently asserted that even the Simplest assoc iat ional
processes are at least as complex as this , and may involve the
partic ipation of thousands of neu rons in widely separate parts ofthe cortex ; and the consc i ousness m ust be regarded as a functionof the enti re process, not Of any detached center (cf . p.
In summariz ing th is dynamic conception of the nature of consciousness Iwi ll quote a few sentences from my brother
’s wr itings (see C . L . Herri ck,
19 10, pp . 13 ,“The theory of consciousness wh i ch seems best to conform to the cond i
tions O f bra in structure and i ts Observed unity is that each conscious state isan express ion Of the tota l equ il ibrium Of the conscious mechanism , and thatintercurrent stimuli are continual ly Sh ifting the equi librium from one toanother class of activ ities . In other words
, the sensat ion accompanyinga given co lor presentation is not d ue to the vibrations in the visua l centerin the occipita l lobe, but to the state of cortical equi l ibrium or the equat ionOf cort ical excitement when that co lor stimulus predominates . Previousvestigeal excitements and coord inations [associations, C . J . H . , see p. 35 ]w i th the data from other corti cal centers a ll enter into the conscious presentation . A s the wave O f exci tation passes from the v isua l center to otherparts , the proport iona l parti cipation Of other centers increases , producing acompos ite conta ining more d istantly related elements .”
“E very specific sense- content w ith i ts escort of reflexly produced associ
ated elements causes a more or less profound d isturbance of the psych i ca lequilibrium , and the nature of th is d isturbance depends not only on the
Intensi ty and state of concentrat ion,but very largely on the kind of equi
libr ium , a lready existing . The character of the conscious act (andthe elements of consciousness are a lways acts ) wi ll , of course, depend uponthe extent to wh i ch the severa l fac tors in the associat ional system partic ipate in
.
the equil ibrium . E ach d isturbance O f the e uilibrium Spreads fromthe poInt of impac t in such a way that progress ive y more O f the poss ib lereflex currents enter the complex, thus producing the extens ion from m eresensat ion to the h igher processes O f appercept ive associat ion . A consciousact 13 always a fluctuation of equ i l ibrium
,so that al l cognitive elements
a
ge a
‘fir
jakened In response to changes rather than invariab le or m onotonous
8 Im
The dynam i c V iew of consc i ousness here adopted m akes suchexpress10ns as
“the unconsc i ous m ind” impossible contrad ic
tIons . E i ther the menta l functions are in process or they are

298 INTRODU CTION TO N E U ROLOGY
cerned with the individual ly acqu i red and espec i al ly the intelli
gently perform ed activ ities as distingu i shed from the fund amental reflex and instinctive processes whose m echani sm s are i nnate.
There is a spec ific local i zation of function i n the cerebral cortex,in the sense that parti cular system s of sensory proj ection fibersterm inate in Spec ia l regi ons (the sensory proj ecti on centers) ,that from other spec ia l regi ons (the m otor projection centers)particu lar system s of eflerent fibers ari se for connection with thelower m otor centers related to groups of m usc les concerned withthe bodi ly m ovem ents
,and that between these projection centers
there are assoc iat ion centers , each of whi ch has fibrous connections of a m ore or less definite pattern with al l other parts of thecortex. The destruction of any part of the cortex or of thefiber tracts connected therewi th involves, fi rst
,a perm anent loss
Of the parti cu lar functions served by the neu rons affected, and ,in the second place, a trans itory di sturbance of the cort icalequ i libr ium as a whole (diasch i si s effect) . Spec ific m ental actsor facu l t ies are not resident in parti cu lar corti cal areas , but al lconsc i ous processes probably require the discharge Of nervousenergy throughout extensive regi ons Of the cortex, and the character of the consc i ousness wi l l depend in each case upon thedynam ic pattern of th i s d ischarge and the sequence of functi on Ofi ts component system s . Thi s pattern is inconceivably complexand only the grosser features are at present open to Observationby exper im ent and pathologi cal studies .N o cortical area can proper ly be desc ribed as the exc lusive
center of a particu lar function . Such “centers are m erelynodal points in an exceedingly complex system of neurons whichm ust act as a whole in order to perform any function whatsoever .Thei r relation to cerebra l functions i s analogous to that of therai lway stati ons of a big c i ty to traffic , each drawing from the
whole c i ty i ts appropri ate Share of passengers and freight ; andtheir great c lini cal value grows out of j ust th i s segregation offibers of l ike functional system s in a narrow space, and not to anymyster ious power of generating psych ic or any other spec i a lforces Of thei r own .
The essence of corti cal function i s correlation , and a corti calcenter for the perform ance Of a parti cu lar function i s a physi ological absu rdity , save in the restr i cted sense descri bed above, as

THE FUN CTIONS OF THE CE RE BRAL CORTEX 299
a nodal point in a very complex system of assoc iated conducti on
paths . Those reflexes whose simple functions can be l ocal i zedin a single center have thei r m echan i sm s abundantlyprovided forin the brain stem . The resting brain i s probably norm al lydu r ing l ife in a state of neural tension in m ore or less stableequ i l ibr ium . An effective stim u lus disturbs th is equ i l ibr iumand the prec i se effect wi l l depend upon variable synapti c resi stance or neuron thresholds whi ch change with different functionalstates Of the organi sm as a whole and of the brain in particu lar .I f th i s activi ty involves the cerebral cortex of a hum an brain, i tmay be a consc i ous activi ty, the k ind of consc i ousness dependingon the ki nd of discharge . But the consc i ousness m ust not bethought Of as locali zed in any cort ical area . The discharge i nquestion may reverberate to the extreme l im i ts of the nervoussystem and the per ipheral activities may be as essential in determ ining the consc i ous content as the cortical .
LITE RATU RE
SON BE CHTE REW
,W . 19 1 1 . Die Funktionen d er Nervencentra, vo l .
III, ena.
BU RNE TT,T. C . 19 12 . Some Observat ions on Decerebrate Frogs , w ith
Special Reference to the Format ion of A ssociations , Amer . J our . Physiol . ,vo l . xxx, pp. 80—87 .
CUSHIN G ,H. 1908 . Remova l Of a Subcorti cal Cystic Tum or at a Secondstage Operation W ithout Anesthes ia, J our . Amer . Med . Assoc ., 1908, vo l . i ,847
1909 . A N ote upon the Farad i c Stim ulation of the Postcentral Gyrusin Conscious Patients , Bra in , vo l . xxxii , pp . 44—54 .
E DIN GE R , L . 1893 . The Sign ificance of the Cortex Considered in Connection w ith a Report U pon a Dog from wh i ch the Who le Cerebrum hadbeen Removed by Professor Go ltz , Jour . Comp . N curol . , vo l . i i i , pp. 69- 77 .
1 908 . The Relations Of Com arative Anatomy to ComparativePsycho logy, J our . Comp . N eurol . , vo xv ii i , pp . 437—457 .
E DIN GE R, L ., and FISCHE R, B . 19 13 . E in Mensch Ohne Grosshirn ,
Arch .
f . ges . Phys io l . , Bd . 152 , pp . 1—27 .
FL E CHSIG , P . 1896 . Geh irn und Seele, Leipz ig.
1896 . D ie Lokal isation d er geistigen Vorgange, Leipz ig .
FRAN Z , S. I . 19 15 . Var iations in Distribution of the Motor Centers,Psycho logi ca l Monographs , Princeton, N . J vo l . xix, N O . 1
, pp . 80—1 62 .
FRITSCH, G ., and HITZ I G , E . 1870. Ueber d ie elektrische E rregbarkeitd es Grosshi rns , Arch . f . Anat . , Phys iol . u . W issen . Med , p . 300 .
GALL and SPU RZHE IM . 1810—19 . Anatom ic et Physiologic d u SystemeNerveux, Paris .G oLTz , F . 1869 . Beitrage zur Lehre von d en Functionen d er Nerven
centren d es Frosches, Berl in.

300 INTRODU CTION TO N EU ROLOGY
G oLTz , F . 1892 . Der Hund Ohne Grossh irn, Arch . f . ges . Phys io l . , Bd .
5 1 , . 570 .
(fRUNB AUM, A . S. F ., and SHE RRIN GTON , C . S. 1903 . Observations on the
Physio logy of the Cerebra l Cortex of the Anthropoid Apes, Proc . Roy. So‘
c . ,
v .o l lxxii , p . 1 52 .
HEAD ,H
, and HO LMES, G . 19 1 1 . Sensory Disturbances from CerebralLes ions , Bra in vo l . m xiv , pp . 109 - 254 .
HE RRICK, C. L . 19 10 . The E qu i libr ium Theory Of Consciousness , inThe Metaphys ics of a N atural ist, Bul . Sc i . Lab . Deni son Univers i ty
,vol .
xv,pp . 12—22 .
HITZ IG,E . 1 904 . Phys io logische und klinische Untersuchungen uber
das Geh irn, Berlin .
HO LMES, G . W . 1901 . The N ervous System of the Dog W ithout aForebrain,
J .0 ur Phys io l .,vo l . xxv i i .
L EWANDOWSKY , M . 1907 . Die Funk tionen d es zentralen Nervensys~tems , J ena .
MARIE ,P . 1906 . Rev is ion d e la Question d e l ’Aphasie, Sema ine Méd
icale, 23 May .
ME YE R,A . 19 10 . The Present Status of Aphasia an d Apraxia, The
Harvey Lectures for 1909—10, N ew York , pp. 228—250 .
VO N MO NAKOW, C . 1909 . Neue Ges i chtspunkte in d er Frage nach d erLoka li sation im Grossh irn , Zeits . f . Psycho logie, Bd . 54, pp. 16 1—182 .
19 10. Aufbau und Loka lisation d er Bewegungen beim Menschen .
Arbeiten a. d . hirnanatom , Institut in Zur i ch, Bd . 5 , pp . 1—37 ; also in Berichtiiber d en IV Kongress f . exp . Psycho logie In Innsbruck, 19 10.
19 13 . Theoretische Betrachtungen i ’Iber d i e Loka lisat ion In Zentra lnervensystem
,insbesondere Im Grossh irn,
E rgebnisse d erpp. 206—278 .
19 14 . Die Loka li sation im Grossh irn . G egenwartiger Stand d erFrage ‘
d er Lokalisat ion in d er G rossh irnr ind e, W iesbad en .
MU NK,H. 1890 . Ueber d ie Funktioncn d er G rosshirnrind e. Gesam
m elte Abhand l . , 2d cd .,Berli n .
1 902 . Zur Phys iologie d er Grosshirnrind e, Arch . f . Physiol ., 1902 .
PRINCE , M . 19 14 . The Uncons cious , New York .

302 INTRODU CTION TO N EUROLOGY
that i s, the heredi tary component of the comm only recogni zed
instincts (p. There i s no c lear boundary between reflexesand instinct- actions as j ust defined . These acti ons may be
exceedingly complex and thei r neu ro-m uscu lar m echani sm s m ay
be compli cated apparently without l im i t . The avai lable -
evi
dence suggests that they are always unconsc i ously performed .
Most Of ou r comm on act ivit ies inc lude al l three of these typesOf behavior in varying proporti ons , and accordingly they frequently have not been distingu i shed . The fi rst and th i rd typesare espec ia l ly l iab le to confus ion, for both are m an ifested asstereotyped , non- i nte l l igent behavi or . They can som etim es beseparated only by a study of thei r or igins ; nevertheless th i s d i stincti on i s of great importance, espec ial ly to educators .The nervous organs of the invar iable reactions are fai rly wel l
known and are character i zed in thei r m ore h ighly elaboratedform s by a c losely kni t system Of nerve- centers and distinct connecting fiber tracts SO organi zed that part i cu lar stim uli may cal lforth a response or a com bination of several responses selectedfrom a fixed num ber of possible actions . The range of poss iblereactions Of any gi ven functi onal system Of th i s type i s l im i tedby the structu ral complexi ty Of the nerve- centers involved .
Th i s complexi ty may be very great, wi th a cor respondingly greatnumber of m ovem ents necessary to complete the reaction , and i tmay inc lude the capac i ty for di sc rim inating between two or m orestructu ral ly possible m odes Of response by m eans of variableinternal functional states of the nerve- cente rs . But i n al l ofthese cases the response i s final ly determ ined with in rather narrow l im i ts by the natu re of the st im u l i and the innate structu ra lorgani zation not only of the nervous organs, but of the body as awhole.
In some cases an elaborate nervous reflex or instinctive actmay involve a m ore extens ive nervous apparatus than i s requ i redby an intel l igent act . I t i s not a m ere question of the si ze of thenervous m echan i sm s involved . For instance, a compari son Of
the brains Of the two spec ies of fishes shown in Fig. 136 Showsthat in the m edu l la Oblongata Of these rather c l osely relatedspec ies there is an astoni sh ing difference between the s i ze ofcertain reflex centers . The greater si ze of the m edu lla oblon
gata Of Carpiodes over that Of Hyod on i s d ue a lm ost enti rely to
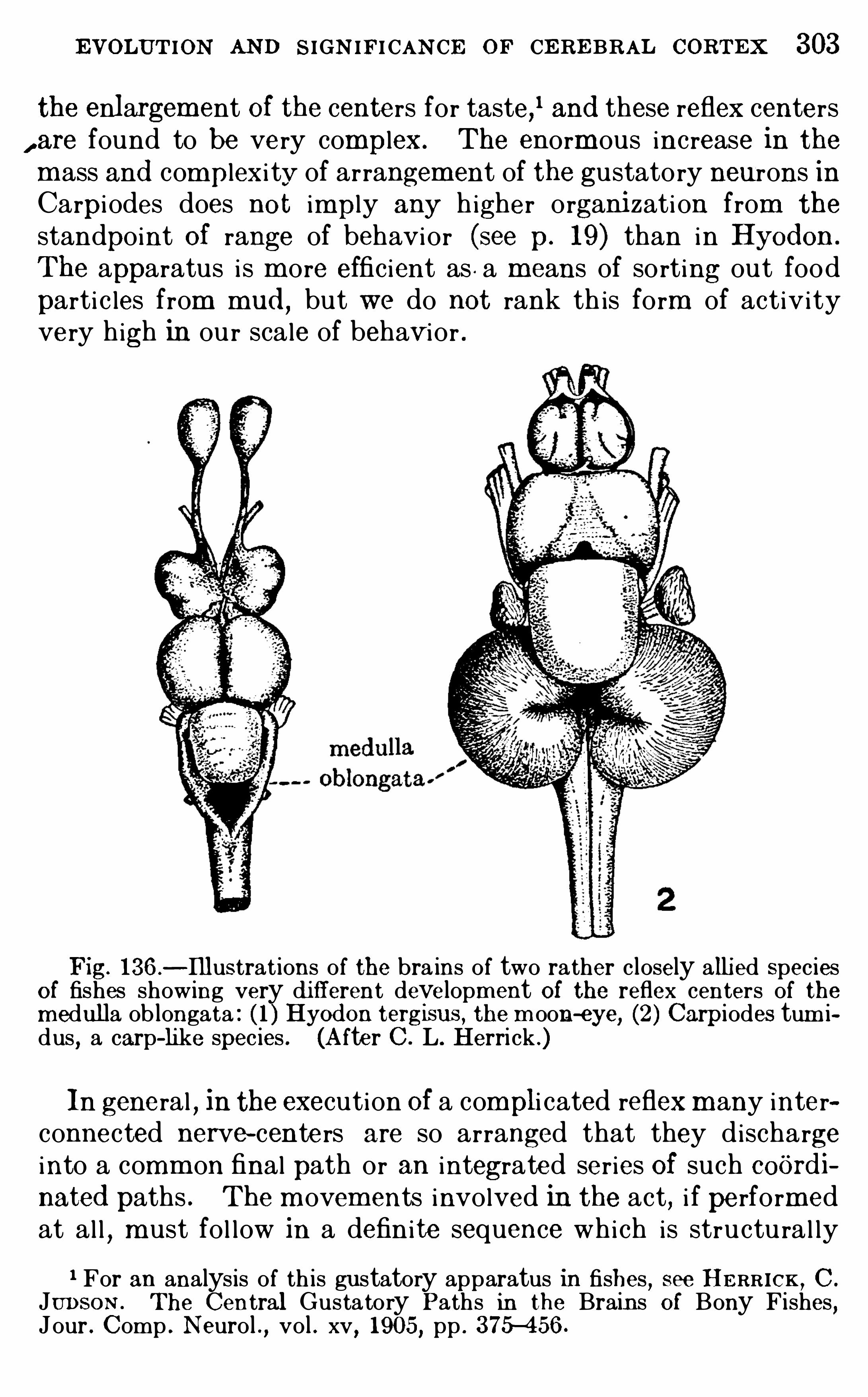
EVOLUTION AND SIGN IF ICAN CE OF CE RE BRAL CoRTEx 303
the enlargem ent Of the centers for taste,1 and these reflex centers, are found to be very complex. The enorm ous increase in them ass and complexi ty of arrangement of the gustatory neurons inCarpi odes does not imply any h igher organi zati on from the
standpoint Of range of behavior (see p. 19 ) than in Hyod on .
The apparatus i s m ore effic ient as . a m eans of sorting out foodparti c les from mud
,but we do not rank th is form of activ ity
very high in ou r scale of behavi or .
Fig. 136—Illustrat ions of the bra ins of two rather closely alli ed SpeciesO f fishes showing very di fferent development Of the reflex centers of themedulla ob longata : ( 1 ) Hyod on tergisus , the m oon-eye, (2 ) Carpiodes tum idus
,a carp-like species . (After C . L. Herri ck .)
In general , in the execution Of a compli cated reflexm any interconnected nerve- centers are so arranged that they dischargeinto a comm on final path or an integrated series Of such coOrd inated paths . The m ovem ents involved in the act, i f perform edat al l
,m ust fol low in a definite sequence which is structu ral ly
1 For an analys is of th is gustatory apparatus in fishes,see HE RR ICK , C .
JUDSON . The Central Gustatory Paths in the Brains Of Bony Fishes,
Jour . Comp . Neuro l . , vo l . xv, 1905, pp . 375—456 .

304 INTRODU CTION To N EU ROLOGY
predeterm ined in the inborn organ i zation of the nerve—centersconcerned . In the var iable type of response, on the other hand ,the assoc iat ion centers involved are so arranged that m any final
paths leading to different system s Of coordinated m otor centersdiverge from a Single center of correlat ion . Which of these pathswi l l be taken in a given reacti on, that is , whi ch of several possibledifferent (or even antagon i stic) m ovements wi l l resu l t, wi l l bedeterm ined by variable physio logi cal factors of internal res istance w i th in the correlating system (fatigue, habit, the i nfluenceof mem ory vestiges , accordingly , the response i s not predeterm ined by the inborn organ i zation of the apparatus .Defini te, wel l-establ ished reflexes general ly fo l low distinct
nervous pathways between Sharply l im i ted nerve- centers . Be
tween these centers there i s usual ly found,in addition to the
wel l insu lated tracts j ust m entioned,a m ore diffuse and loosely
organ i zed entanglement of nerve- cel l s and fibers,through whi ch
nervous impu l ses may be m ore Sl ow ly transm i tted in any di rcetion . Ti ssue of th is character i s found throughout the enti relength of the central nervous system , and in som e places i t occupies extensive regi ons (espec i al ly in the m edu l la Oblongata andupper part of the spinal cord) whi ch are term ed the reti cu lar form ation (see pp. 65
,127
,
The reti cu lar form ati on i s the parent tissue out of whi ch thehigher correlat ion centers have been differentiated . In the
Spinal cord and m edu l la oblongata , where i ts character i s m ostc learly seen, i t rece ives fibers from al l Of the sensory centersand may discharge m otor impu l ses into efferent centers of contiguous or very rem ote regi ons . I n the h igher parts Of the brainthe el aborate assoc iation centers Of the thalam us and cerebralhem i spheres have been devel oped from such a prim i t ive m atrix,and these centers are interconnected by Sim i lar undifferentiatednervous ti ssue .
The detai ls of the functional connections Of the reflex centersof the brain stem are m uch m ore prec i sely known than are thoseof the h igher corre lation centers of the thalam us and cerebra lcortex. And
,in fact, i t i s essential that these detai ls be fa i r ly
wel l understood before the functions of the h igher centers canbe investigated ; for al l nervous impu lses whi ch reach these highercente rs m ust first pass through the l ower centers and there be

306 INTRODU CTION To N E U ROLOGY
functional ly defined m otor areas, each group of m usc les whi chcooperate i n defini te reacti on complexes (termed synergi cm usc les, see p. 35) being exc i ted from a defin i te part of the m otorcorticalfield , whosem otor tract i s anatom i cal ly di st inct throughout i ts enti re further course from the cortex to the per iphery .
Between the sensory proj ection centers and the m otor areas areinterpolated the assoc iati on centers , and these are SO arrangedthat al l correlati on,
integration,and assim i lation of present
sensory impu lses with m em ory vestiges of past reactions arecompleted , and the nature of the response to be m ade i s determ ined before the resu l tant nervous impu l ses are discharged intothe m otor centers . Only such of the m otor areas wi l l be exc i tedto funct ion as are necessary for evok ing the parti cu lar reactionwhi ch i s the appropr iate (that is , adaptive) response to the totals ituation in whi ch the body finds i tself . This arrangem ent Ofassoc iation centers in relat ion to a ser ies of d istinct m otor areas
provides the flexib i l i ty necessary for complex delayed reactionswhose character i s not predeterm ined by the nature Of the con
gen i tal pattern of the nervous connections .1
The thalam us , as we have seen (p . has its own intrins i c systemof as sociation centers whi ch d ischarge downward into the cerebral pedumcles , and th is is the pr imary reflex apparatus Of this part Of the bra in The
thalamo—cortical connections arose to prom inence later in the evo lutionaryh istory, though feeb le rud iments Of these are present in lower brains .Paral lel with the enlargement of these corti cal connections a special part ofthe thalamus was set apart for them , and from theAm ph ib ia upward in theanim al scale th is dorsal part of the thalamus assum ed increasingly greaterim portance . Th is part is termed by E d inger the neothalamus , and makesup by far the larger part of the thalamus in the hum an and al l other mamma lian bra ins . It occupies the dorso—lateral part of the thalamus proper andcompri ses most of the great thalamic nuclei ( lateral and ventra l nuclei,pulvinar and lateral and med ial geni cul ate bod ies ) . The pr imitive ihtrinsic reflex thalamic apparatus in m an is a relatively un important areaof med ial gray matter and the subthalamic region (corpus Luysii, latti cenucleus , etc . , not to be confused w ith the hypothalamus whi ch lies fartherdown in the tuber cinereum and mammillary bod ies ) .The neothalamus , accord ingly, serves as a sort O f vestibule to the cortex,
every afferent impulse from the sensory centers (except the O lfactory system ) being here interrupted by a synapse and Opportuni ty offered for a w iderange of subcortical associat ions . The o lfactory cortex (h ippocampal formation) has a s im i lar relation to subcorti ca l correlation centers in the olfactory area in the anter ior perforated Space
,sep tum , etc .
1.The paragraphs wh i ch fol low (pp . 306—3 1 1 ) are reproduced w i th Snght
Egc
ad IficatIon from The Journal Of Animal Behav ior
,vo l . i ii , 1 9 13, pp . 228

EVOLUTION AND SIGN IF ICAN CE OF CE REBRAL CORTEX 307
From these anatom ical considerations i t fo l lows that no s im ple sensoryimpulse can ,
under ord inary circum stances, reach the cerebral cortexw i thout first being influenced by subcort ical correlat ion centers, w ith inwh i ch complex reflex combinat ions may be effected and various autom at ismsset O ff in accordance w i th their preformed structure. These subcort i calsystems are to some extent mod ifiab le by racial and ind ividual experience
,
but their reactions are chi efly Of the invar iab le or stereotyped charac ter,w ith a re lat ively limited range of poss ib le react ion types for any givenstimulus complex .
I t is shown by the lower vertebrates wh i ch lack the cerebral cortex thatthese subcort ica l mechani sms are adequate for all Of the ord inary sim pleprocesses of hfe
,inc luding some degree Of associat ive memory. But here,
when emergencies ar ise wh i ch invo lve s ituations too complex to be reso lvedby these mechanisms, the animal wi ll pay the inevitab le penalty Of failure;perhaps the loss of h is d inner, or even Of h is life .
In the higher mammals w i th well-developed cortex the au tomat isms ands imple associat ions are li kewise performed in the ma in by the subcorticalapparatus
,bu t the inad equacy of th is apparatus in anypart i cular s i tuation
presents not the certa in ty Of fai lure, but rather a di lemma . The rapidpreformed reflex mechan isms fa il to give rel ief, or perhaps the s i tuation prc~sents so many comp lex sensory exci tat ions as to cause mutual interferenceand inh ib it ion of a l l reaction . There is a stas is in the subcortical centers .Meanwh ile the h igher neural res istance of the cortical pathways has beenovercome by summat ion of st imul i and the cortex is exci ted to function.
Here is a mechanism adapted, not for a limi ted number of predeterminedand immed iate responses, but for a much greater range of combinat ion ofthe afferent i mpress ions w ith each other and with memory vest iges of previous reac tions and amuch larger range of possible modes of response to anygiven set of afferent impress ions . By a process of trial and error, perhaps,the e lements necessary to effect the adapt ive response may be assemb ledand the prob lem so lved .
It is evident here that the physiologi ca l factors in the d ilemma or prob lemas th is is presented to the cortex are by no means simple sensory im pressions , but defini tely organized systems Of neura l d ischarge, each of wh ich is aphys iologica l resultant O f the reflexes , automat isms , impulses, and inhibit ions characteristi c of i ts appropriate subcort ical centers . The prec ise formwh i ch these subcorti cal comb inat ions will assum e in response to any particular exci tat ion is in lar ge measure determined by the structural connections O f these centers i nter se. An d the pattern of these connect ions isto lerab ly uniform for a l l members Of any animal race or species . Th isimplies that i t is hered i tary and innate. Th is is the underlying bas is O finstinct .The connections between the cortical centers
,on the other hand , are
much less definitely la id down in the hered i tary pattern . The deta ils Ofthe defin i tive association pattern of any ind ividual are to a greater degreefixed by his part icular experience . Th is is the bas is Of doc ili ty and the
ind iv idual ly modifiab le or intell igent types of behavi or. The typica l cort ical activit ies, even when phys iologically considered , are far removed indeedfrom those of the bra in stem .
I t should be emphas ized , however, that the differences between the cortex and the lower centers of the brain stem ,
so far as these can be deducedfrom a study Of structure and from physio logica l experiment , are relat ive andnot abso lute . Indeed , the genera l pattern of the regional localizat ion of the

308 INTRODU CTION To N E U ROLOGY
cortex i tself is innate , and in ad ult life the cortex has acquired many m orecharacteris ti cs s imi lar to those of the bra in stem, w i th i ts own systems ofacquired automatisms and hab i tua lly fixed types of response. The largeras sociation centers retain their plasti ci ty longest , but ul tim ately these alsoceas e to exhib i t new types of correlation, and th is marks the onset of
The relations Of the cerebral cortex to the cerebellar cortex and thebrain stem have been compared (p . 192) to those of an en larged jud i cialbranch of the central governm ent charged with the duty of interpreting thedecrees Of the lower legis lat ive centers an d dominating the adm inistrat ivemachinery, and with the ad di tiona l power of shaping the general poli cy ofthe government.Dewey’s st im ulat ing analys is1 of the reflex arc concept or
,as he prefers
to say, the organi c circuit concept implies that the synthes is of the elementsO f a complex cha in reflex into an organi c uni ty is the es sential prerequi s iteof that appercept ive process whi ch w i l l make the total exper ience Of valuefor future discrim inative responses—for learning by experience. Thi s
,
wh ich is true in the ind ividual learning process, is also true phylogenetically.
The correlation centers (an d their capaci ty for the pres ervat ion Of ves tigesOf past reactions) are the organi c m echan ism for this synthes is . Theym akeit poss ib le that a new stimu lus may be reacted to , not as a detached element,but as a component of a complex series of past and pres ent adj ustments , towhi ch it is as s im i lated in the as sociat ion centers—appercept ion . Thisass im ilation or apperceptive process is an integral part of the receptor process in the h igher centers , giving the quale to the idea of the excit ing object.Cotemporaneously wi th th is st imulus-apperception process we have an
apperception- response—activi ty giving the Object or purpose- idea, so thatthe entire react ion is to be regarded as stimulus-appercept ion-response, as afunctional unitv rather than as a sequence : stim ulus> apperception> re
sponse .
Dewey’s organi c c ircui t concept is elaborated in term s of psycho logy.
Let us see how i t may be appli ed to b io logi cal behavior .The s imple reflex is commonly regarded as a causal sequence : given the
gun (a physio logical ly adap t ive structure) , load the gun (the constructivemetaboli c proces s) , aim ,
pull the trigger (appli cat ion of the stimulus) , d i scharge the projec t i le (phys io logica l response) , h it the mark (satisfaction ofthe organi c need ) . A ll of the fac tors may be related as members of a s implemechani cal causal sequence except the aim . For thi s in our illustrationa glance backward is necessary. An adaptive s imple reflex is adapt ivebecause of a pre-establ ished series Of functional sequences whi ch have beenb io logical ly determined by natura l select ion or some other evolutionaryprocess . Th is gives the react ion a defini te aim or objective purpose. In
short , the aim , hke the gun , is prov ided by b io logi ca l evo lut ion,and the
whole process is implic i t in the structure- function organization wh i ch ischaracteristic of the species and whose nature and origin we need not herefurther inquire into .
N ow, pass ing to the more complex ins tinct ive reactions , so far as theseare unconsc ious automatisms , they may be elaborations of cha in reflexesof the type di scussed above (p . But the aim (b io logica l purpose) is
The Reflex Arc Concept in Psycho logy , Psych . R ev.,vo l . iii , p. 357,
1 893 . See a lsoDewey’ s later statement in Jour . Ph i los .
,Psych , and Sc i .
Methods , vo l . ix, Nov., 1 9 12 , pp. 664—668, especially thej ootnote on p. 667.
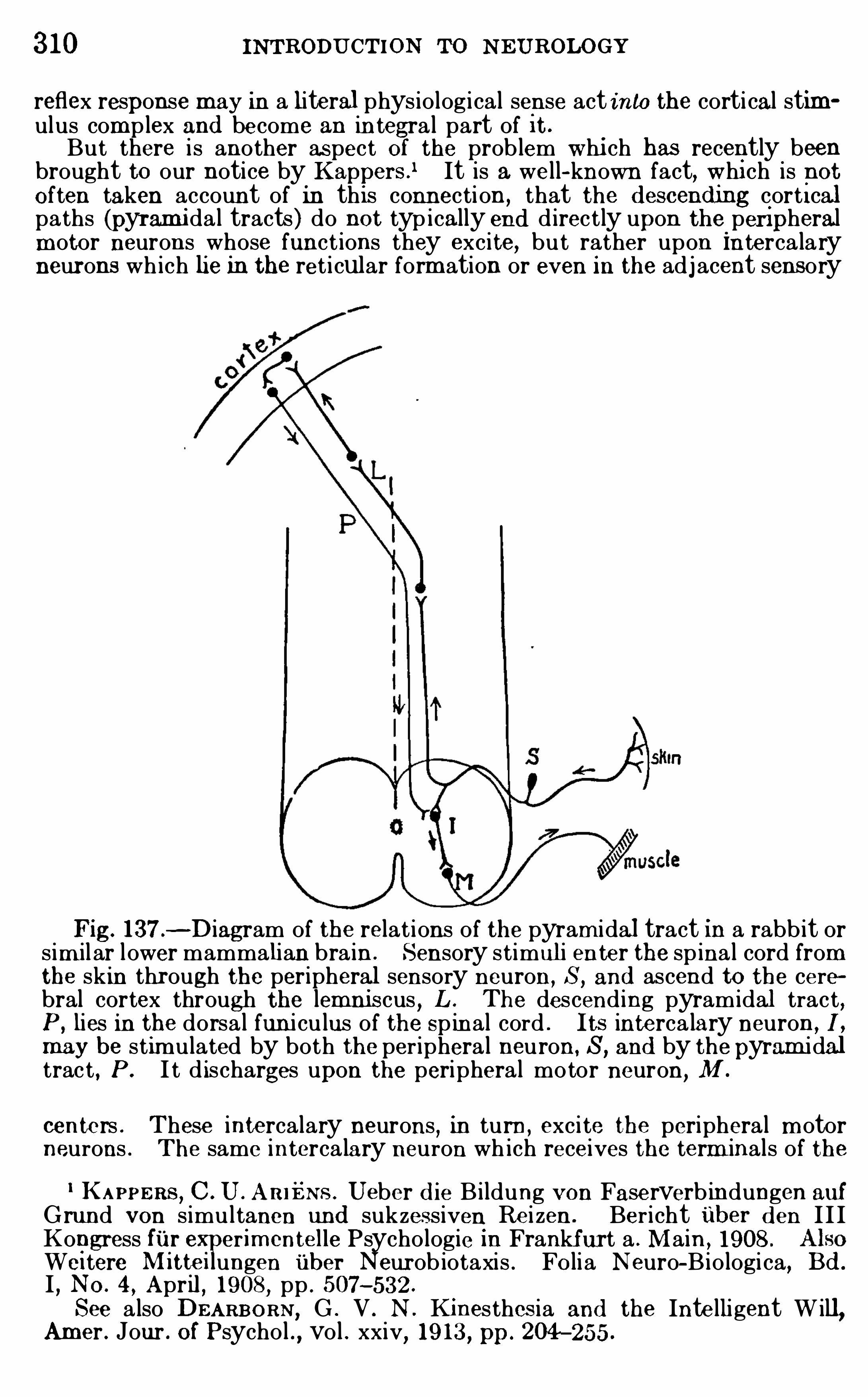
310 INTRODU CTION To N EUROLOGY
reflex response may in a literal phys io logical sense act into the corti cal stimulus complex and become an in tegra l part of i t .
But there is another aspect Of the prob lem whi ch has recently beenbrought to our notice by Kappers .
1 It is a well- known fact, whi ch is notO ften taken account of in this connection, that the d escend m g cort i calpaths (pyrami da l tracts) do not typ ically end d irectly upon the.
peripheralmotor neurons whose funct ions they excite, bu t rather upon Interca laryneurons wh ich lie in the ret i cular form ation or even in the adj acent sensory
Fig. l 37 .- Diagram of the relat ions of the pyram idal tract in a rabb it or
s im ilar lower mammalian bra in . Sensory stimuli enter the spina l cord fromthe skin through the peripheral sensory neuron, S, and ascend to the cerebral cortex through the lemniscus , L . The descend ing pyramidal tract,P , lies in the dorsa l fun i culus of the Spinal cord . Its interca lary neuron ,
I
may be stim ulated by both the per iphera l neuron, S,and by the pyram idal
trac t , P . I t d ischarges upon the peripheral motor neuron, M .
centers . These intercalary neurons , in turn ,exci te the peripheral m otor
neurons . The same interca lary neuron wh ich receives the term inals of the1 KAPPERS, C . U . A R IENs . Ueber d ie Bildung von Faserverb indungen auf
G rund von s imultanen und sukzes siven Re i zen . Ber i cht iiber d en I I IKongress fi 'Ir e erim cntelle Psycho logie in Frankfurt a . Main , 1908 .
Wei tere Mitte i ungen i'
Iber Neurob iotaxis . FO lia N euro—Bio logi ca, B d .
I , N O . 4 , April , 1908, pp . 507—532 .
See a lso DEARB O RN , G . V . N . Kinesthes ia and the Inte lli gent W ill,Amer . Jour . O f Psycho l ., vo l . xxiv, 19 13 , pp. 204—255 .

EVOLUTION AND SIGN IF ICAN CE OF CERE BRAL CORTEX 31 1
pyrami da l tract also receives collatera ls from the peripheral sensoryneuronsOf its own segment (Fig . Th is arrangement is the explanat i on of thefact that the pyramida l tract fibers descend through the human Spina lcord for the m ost part in the dorso- lateral region, not in the ventral funi cu lus hke most other motor tracts . In m ost lower mammals the pyram idaltract actual ly descends w i thin the dorsal funi culus in the c loses t poss ib leassociation w ith the per ipheral sensory fibers, and th is arrangement isc lear ly the prim i tive relat ion of the descending corti cal pathway.
A ccordingly, stimulat ion of the skin of the body exci tes a dorsal spina lroot fiber whi ch ascends toward the cortex with in the spina l cord and alsogives collatera l branches to intercalary neurons of the spina l cord i tself .The latter neurons may excite motor elements of the spina l cord to an immed iate reflex response wh i ch is we ll under way before the cort ica l returnmotor impulse gets back to the spina l cord and di scharges into these same
interca lary neurons wh ich ar e already under sensory stimulat ion di rec t lyfrom the periphery. The effec t Of thi s arrangement is that the centralmotor path dur ing function is under the influence of sensory stimulation atboth ends
,and is not, as common ly descr ibed , under simple sensory st imula
tion at the cortical end and purely emi ss ive in funct ion at the sp ina l end .
Viewed from the standpo in t of cerebral dynam i cs, the exact phys iologica leffect of the discharge of a central m otor bund le such as the pyram i dal tractw i ll be dependent upon the comb ined action Of the sensory stim ulation atthe cortical end and the state of sensory excitat ion at the spinal end , as wellas upon the res istance Of the motor apparatus i tself .We saw in a previous paragraph how the Simple reflexes of the sp inal
cord may become factors in the st imulus complex of the cortex . Here wefind , conversely, that the efferent cortical di scharge may become a factor inthe loca l reflex st im ulat ion of a motor spina l neuron . From both standpoints Dewey ’s conception Of the unitary nature of the organ i c c ircui t , ascontrasted wi th the class i ca l reflex arc concep t, receives strong support.The thalam i c correlat ion centers probab ly serve as the organs par excel
lence where are elaborated those organ i c c ircui ts wh i ch give to the hi gherappercept ive processes O f the cortex that quale to wh ich Dewey refers .The origin Of this qua le is to be sought partly in the subcorti cal assimi lat ionof a present st imulus complex to the pre—existing organic c ircuits structura lly laid down in the reflex mechani sm , and partly in an affect ive qual ityperta ining to the several organic c ircuits invo lved in the react ion . Th isaffective quality may be innate or i t may have been acquired by exper ienceof the results of prev ious reactions of the sort in question .
Head and Holmes have brought forward some very interesting evidencethat not only the affective qua le of sensations but a lso the em otiona l l ifein general is functionally related to the primit ive intrins i c nuclei of thethalamus, rather than to cortica l activi ty (see p . And certainlythere is m uch ev idence in the behav ior of lower an ima ls , especial ly b irds ,that a h igh degree of emotiona l act iv ity is poss ib le where the basa l centersar
e
e
Clh igh ly elaborated but the cerebra l cortex is sma ll and very Sim ply organ
i z
From al l of these cons iderations i t seems probable that the funct ions ofthe h igher associat ion centers of the cerebra l cortex do not cons ist of theelaboration O f crude sensory data or Of any s im ilar elements , but rather Ofthe assemb l ing and integrat ion Of h igh ly elaborated subcortica l organ ic circui ts wh ich in the aggregate make up the greater part of the reflex and instinctive l ife of the Species.

3 12 INTRODU CTION TO N EU ROL OGY
The norm al newborn chi ld brings into the world an inher i tedform of body and brain and a complex web of nerve- cel l s andnerve-fibers which provide a fixed m echanism , comm on exceptfo r m inor variati ons to al l m em bers of the race alike, for the
performance of the reflex and instinctive actions . The pat
tern of th is hereditary fabr i c can be changed on ly very slowlyby the agency of selective m atings and other str i ctly biologi calfactors or by degenerations of a d istinctly pathologi cal sort . I t i sthus m an i fest that the improvem ent Of the rac ia l stock Of norm alindividuals by the practi ce of eugeni cs m ust necessar i ly be veryslow
,though the improvem ent of defective or pathological strains
by selective m atings so as to breed out the objectionable characteristics i s fortunately in m ost cases m ore readi ly accompl i shed .
But in addition to th i s hereditary organ i zat ion the newbornch i ld possesses the large assoc iat ion centers of the brain withthei r vast and undeterm ined potenc ies , the exact form of whoseinternal organi zation is not whol ly la id down at bi rth , but i sin part shaped by each indivi dual separately dur ing the cou rse ofthe growth per i od by the processes of education to wh i ch he i ssubjected , that i s , by his exper ience. Th is capac i ty for individ uality in developm ent
,th i s abi l i ty to profit by exper ience,
th is doc ili ty,i s m an ’s m ost distinctive and valuable character
istic . And since the form whi ch th is m odifiable ti ssue wi l l takei s determ ined by the environ ing i nfluences to whi ch the ch i ldi s subjected , and since these influences are largely under soc ialcontro l , i t follows that hum an cu l tu re can advance by leaps andbounds wherever a h igh level of comm un i ty l ife and educationalideals is m aintained .
So wel l have we learned the lesson that the ch i ld br ings wi thhim into the world no mental endowm ents ready-m ade—noknowledge, no ideas , no m orals—but that these have to bedeveloped anew in each generation under the gu id ing hand ofeducation , that we devote one- thi rd of the expected span of l i feOf ou r m ost prom i sing youth to the educational training neces
sary to ensure the h ighest possible developm ent of the latentcu l tural capac i t ies of these assoc iation centers of the cerebralcortex.
But we have often been bl ind to the other side of the pi ctu re.
We have seen above that the adu l t cortex cannot function save

3 14 INTRODU CTION To N EU ROLOGY
com e into function in a suck i ng babe or a k indergarten pupi l .Ch i ldi sh cu r ios ity is ou r strongest al ly, if only we can use i twi sely , throughout the whole of the educational career from in
fancy to the graduate school . Anger i s a m ighty passion i n ch i ldhood . I t i s not wise to eradi cate i t a l together ; rather keep i t,though under curb , for there are tim es when real abuses ari sewhi ch requ i re that the m an know how to h it and to h it hard .
And so with the instincts of self-preservation, of fear , of sexthese al l have thei r parts to play in the nobler works of l ife andare by no m eans to be eradi cated . The asceti c i deal of m ortifica
tion of the flesh as a m eans of grace i s fundam ental ly wrong i n
pr inciple. Our case cal ls for no bl ind , indiscr im inate attackupon the world and the flesh , but rather the subj ugation and di scipline of these, so that wemay use them effectively in our attackupon the devi l .Confl i ct is inherent in the cosm i c process, at least in the bio
logical realm ,from beginn ing to end . There i s the struggle for
physi cal exi stence am ong the an im als . An d even in the l owerranks of li fe there ar ises also the struggle with in the i nd ivid ualbetween stereotyped innate tendenc ies or i nstincts and indivi dual ly acqu i red exper ience . Th i s i s c learly shown by exper imentson anim als as low down as the Protozoa . And out of th is innerconfl ict or di lemm a intel l igence was born . With the gradualem ergence Of self- consc i ousness in thi s process ari ses the eternalstruggle with sel f, that confl ict which leads to the bitter c ry ,“When I wou ld do good evi l i s present wi th m e.
” Confl i ct,
then, l ies at the basi s of al l evolution,and the factors of soc ia l
and even of m oral evolution can be traced downward throughoutthe cosm i c process .The soc ia l and eth i cal standards
,therefore, have not ar i sen
in opposition to the evolutionary process as seen in the brutecreation, but within that process . And ou r imm ediate educational problem i s the elaboration Of a practicable system of publ ic i nstruction whi ch can use to the full the enorm ous dynam i cenergy in the hereditary impu l s ive and instinctive endowm ent ofthe chi ld , and bu i ld upon th is , in the form best su i ted to the re
spective capac i ties of al l the separate individuals , a proper lyordered sequence of studies wh ich wil l devel op the latent capacities of each pupi l and ensure a vital balance between the strong

EVOLUTION AND SIGN IFICAN CE OF CERE BRAL CORTEX 3 15
bl ind impu l se of the i nnate nature and the acqu i red inte l lectual ,esthetic , and m oral contro l .And herein l ies the so lution of the prob lem of hum an freedom ,
so far as th i s rests with in ou r own contro l . The l im i ts of one ’s
powers and the range with in which h is freedom of action i s circum scribed are in part determ ined by hi s heredi tary endowm entsand by envi ronm ental influences over wh ich he has no control .These are decreed to him by h is fate
,and the innate organ i zation
Of the nervous system i s the chief instrument of this fate. But
m an differs from the brute creation chiefly in that he can m orecompletely contro l h i s own envi ronm ent and thereby to thatextent take h is fate into h is own hands ; in other words , he can
enr i ch h is own exper ience along l ines of h i s own selecti on . To
som e extent each individual can do thi s for h im sel f through selfcu l tu re ; but to ensure the best resu lts of such efforts there m ustbe a soc ia l contro l of the envi ronm ent as a whole by concertedcomm unity action . Individual freedom of action can ,
therefore,attain i ts h ighest effic iency only through a certain am ount ofvoluntary renunc i ation of the selfish interests where these con
flict with comm un ity welfare. E th i cal ideals and altru i sm are
as tru ly evoluti onary factors in hum an soc ieties as are the ele
mental laws of self-preservation and propagation of the spec ies .1
To return now to the devel oping nervous system , we note thatthe educational per iod i s l im i ted to the age dur ing whi ch theassoc iat ion centers , whose form i s not predeterm ined in heredity ,rem ain plastic and capable of m od ificatiOn under envi ronm entalinfluence . U l t im ately even the cerebral cortex m atu res and
l oses i ts power of reacting except in fixed m odes . I ts unspecialized ti ssue—original ly a diffuse and equ ipotentia l nervous meshwork— becom es differentiated along defin i te l ines and the fundam ental pattern becom es m ore or less r igi d . The doc i le per iod i s
past , and though the m an m ay continue to improve in the
techn i c of h is perform ance, he can no l onger do creative work .
He i s apt to say ,“The d og i s too old to learn new tri cks .
1 In th is connec tion reference may be made to two very interesting addresses recently del ivered before the American Society of N atural ists :J E NNINGS, H. S. 19 1 1 . Hered i ty and Persona li ty, Science, N . S. , vo l .
xxxiv , pp . 902—9 10.
CON KLIN ,E DWIN G . 19 13 . Hered ity and Responsib i l i ty, Science, N . S.
,
vol. xxxvii , pp . 46—54 .

3 16 INTRODU CTION To NEUROLOGY
Whether th is process occu rs at the age of twenty or eighty years ,i t i s the begi nning of sen ili ty . And , alas , that th i s coagu lat ionof the mental powers often takes place so ear ly $ Many a boy
’sbrains are curdled and squeezed into traditional artific ia l m oldsbefore he leaves the grades at school . His education i s completeand seni le sc lerosi s of the m ind has begun by the time he haslearned his trade. For how m any such disaste rs ou r br i ck-yardmethods in the publi c schools are responsib le i s a questi on oflively interest .We who seek to enter into the kingdom of knowledge and to
continue to advance therei n m ust not only become as l i ttlechi ldren, but we m ust learn to continue 80 . The problem ofSc ientific pedagogy , then , is essential ly th is : to prolong the plasticity of ch i ldhood , or otherwi se expressed , to reduce the interval between the first ch i ldhood and the second chi l dhood toas smal l d im ens i ons as possible.

3 18
Apoplexy, 293Apperception ,
249 , 296 , 308
Appet i te, 240Aqued uct of Sylviu s ( iter, Optoccele,mesocoele) , the ventri cle of themidbrain
,62 , 12 1 , 1 58, 160 , 16 1
Arachno id , the midd le brain m em
brane, 38Arbor vitae , the tree- l ike appearance O f the wh i te matter of thecerebellum in section , 1 90
Arch ipal l i um , the o lfactory cerebralcortex
,includ ing the Hippocam
pus and the Gyrus hippocam pi( in part ) , 2 17 , 22 1 , 222 , 273 , 284 ,
Area, acoustic . See Area, acousticolateral
,N ucleus , coch lear, and
N ucleus , vestibular .acoustico- lateral ( in fishes tuberculum acusticum ) , 1 1 1 , 1 12 ,
corti cal , as used in th is Index isa part of the cerebra l cortexwh i ch can be d ifferentiated fromi ts neighbors structurally by thearrangement O f i ts ce l ls and
fibers (sometimes termed field ) ;cf . Center, cortica l , 273 , 277
cutaneous,1 1 1 , 1 12 , 123 , 1 57
general somat i c sensory. See Area,
cutaneous .olfactoria, the region containingthe secondary olfactory centers
,
d ivided into anter ior, med ial ,interm ed iate and lateral olfactory Nuc lei , 165 , 167 , 2 15 , 2 1 7218
,2 1 9 , 22 1 , 306
paro lfactoria of Broca (gyrus olfactorius med ial is of RETZ IU S) ,a portion of the med ial Area 0 1factoria immed iately in front ofthe Gyrus subca llosu s
, 1 1 9
perforata . See Sub stantia perforata.
somati c , a smal l region in the fishbrain from wh i ch the N eopall ium and Corpus striatum weredeveloped , 1 1 1 , 1 1 2 , 123
striata , that part of the occipi tallobe of the cerebral cortex containing the Line of Gennari ; thevisual center, 268 , 284
INDEX AND GLOSSARY
See Center,
Back- stroke, the influence wh ich a
geripheral organ of response exertsack upon the center from wh ich
Area, visceral . See also Lobe, visceral , 1 1 1 , 1 12 , 123 , 148 , 149 , 152 ,
Arteries,nerves of . See Vasomotor
apparatus.Arti culates , behav ior of, 33A ssociation, correlation invo lving ah igh degree of m od ifiability and
also consciousness, 35 , 64 , 104,296
,
307
Association center .association .
fibers . See Fiber , association, andTract
,association.
of ideas,295
pattern,29 1
time of, 98A sthm a
,238
Ataxia , loss of the power of m uscularcoord ination ,
137, 138Atropin
,23 1
Attention,1 03 , 104
Aud itory apparatus, 60, 62 , 63 , 70,85 , 145 , 147 , 1 50, 1 57 , 160, 163—203
Aud itory reaction time, 98AU E RBACH,
plexus of (myentericplexus) , 241
Au la , the anterior end of the th irdventri cle where i t comm uni catesw ith the lateral ventri cles by wayO f the interventri cular Foram ina.
Auricle, of external car, 1 95Automatisms
,acquired , 35 , 57, 286 ,
Avalanche conduction . See Conduetion, avalanche.
A ids- cylinder, the centra l protoplasmic strand of a nerve-fiber ; partOf the Axon, 39
Axon (axi s-cylinder process , neurite,neuraxon
,neuraxi s ) , a process of
a N euron wh i ch conducts impulsesaway from the cel l body, 39 , 40,44
,4s, 47
Axon h illock, the po int Of origin of anaxon from the ce l l body, 40, 4 1 , 46
Axone. See Axon.

INDEX AND GLOSSARY 319
the response was exci ted ; a form ofcha in Reflex ; cf . Reflex circuit,260 , 309
BAILLARGE R,layer of
,stripe of . See
Line of BAILLARGE R .
BARKE R,L . F .
,36 , 40, 49 , 55 , 94,
BARTL E ME Z, G . W.
, 54 , 181
Bas is ped unculi (pes pedunculi ,crusta) , the ventral part of thecerebral Pedunc le, composed ofdescend ing fiber tracts
,1 14
Basle nomina anatomica (B . N .
BE CHTE REW , W.
,159 , 2 13 , 299
BE CHTE REW,vestibular nucleus of,
184 ,185
Behavior, invariab le, act iv i ties whosecharacter is determined by innate structure
,typified by reflex
and instinctive act ions,3 1
,67 ,
78, 1 15 , 1 81 , 363 , 294, 301—304 ,
3 1 2
range of, 19 , 303variab le, activi ties wh ich are mod ifiable by ind iv idua l experience,w i th or w i thou t consciousness,
67 , 78, 263,
290 , 294 , 301—304 ,
3 12 , 3 15 , 3 16 See Corpus restiBETHE
,A .
,47 , 55 , 90
Betweenbrain . See Diencephalon.
BETz , cel ls of . See Cel ls of BETz .
B IANCHI , A .,193
Birds,behav ior of, 6 1 , 309 , 3 1 1
nervous system of, 187 , 2 16Bladder, innervat ion of, 226 , 232 ,243
Blood,coagulation of, 255
Blood-pressure, 104 , 235Blood-vessels , nerves of . See C irculation of b lood , apparatus of,and Vasom otor apparatus .B . N . A . See Bas le nom ina ana
tomica .
Body of cell . See Cel l body.
chromoph i li c . See Substance,chrom ophili c .of fornix. See Forn i x body.
geni cu late, latera l (corpus gen ienlatum latera le, externa l geniculate body) , a v isua l centerin the Tha lamus , 1 14 , 1 50,163 , 1 64 , 167, 208, 2 10 , 2 12 ,284 , 306
Body, geni culate, m edia l (corpusgeni culatum med iale, interna lgeni culate body) , an aud i torycenter in the Thalamus
,1 14 ,
1 18 , 12 1 , 154 , 157 , 163 , 164 ,1 67 , 185 , 20 1 , 202, 306
habenular . See Habenu la.
of L U YS,1 67
,306
mamm i llary (corpus mam illare,corpus cand icans) , one of a pairof eminences at the poster i orend of the Tuber cinerium In
the Hypothalamus ; an olfac
tory center, 1 14 , 120, 163, 165 ,of N ISSL . See Substance, chromoph i l i c .
pinea l (corpus pinea le, pinea lg land , epiphysis
,conar ium ) , a
g landular outgrowth from the
Epithalam us ; in some lowervertebrates i t takes the form ofa med ian dorsa l eye. SeeParietal eye, 1 10 , 1 14 , 1 18, 1 19 ,1 62 , 1 64 , 167 , 2 12
pi tuitary. See Hypophys is.
quadr igemina l . See Corpora quadrigemina.
restiformform e.
striate . See Corpu s striatum .
tigroid . See Substance, chrome
ph i li c .trapezoi d (corpus trapezo ideum ) ,transverse decussating fibers inthe ventral par t of the medullaOb longata wh i ch connect the
aud itory nucle i of one Side wi ththe latera l L emni scus of theother s ide, 50, 201
BOLK , L .,1 93
BO LTON ,J . S. ,
273 , 277 , 289 , 290
BO N NET, R .
,8 1
Brach ium,of col l iculus infer ior . See
Brachi um quadrigeminum ia
ferius .
conjunctivum (prepeduncle) , the
superior or anter ior peduncle ofthe cerebel lum ; cf . Pedunc le,cerebe llar , 1 14 , 1 3 1 , 158 , 1 62 ,
ponti s (m ed iped uncle, pro cessuscerebel li ad pontem ) , the midd lepeduncle of the cerebellum ; cf .

320
Peduncle, cerebellar, 1 14 , 122,
Brach ium,quad rigeminum inferius
(brach ium of co l l iculus inferior) ,a ridge on the Corpora quadrigem ina formed by fibers from the
Co ll icu lus inferior to the m ed ialgeniculate Body, 1 1 4, 16 1 , 164 ,185
Brain (encephalon) , that portion ofthe centra l nervous system con
tained wi th in the Skull , 106development of . See N ervous system
,development of.
measurements of, 123new . See Ne
'
encephalon.
nomenclature of. See N ervoussystem
,nomenclature of .
old . See Palaae'
ncephalon.
stern, al l of the brain except thecerebel lum and the cerebralcortex , 8. e.
, the Segmentalapparatus, 1 13 , 1 14 , 1 15 , 123 ,1 64 , 181 , 185 , 186 , 192 , 266 ,280
reflexes of .bra in stem .
term ino logy of See Nervous system
,nomenclature of .
weight of, 123
Branch . See Ramus .Branch ial gangli a .
branch ia l .nerves . See G i l ls
,innervation of .
Bridge. See Pons .BROCA, P .
,292
BROCA’S area . See Area parolfactoria of Broca.Broca’ s convolution, the posteriorpart of the gyrus frontal is inferior ,supposed to func tion as a motorcorrelation center of Speech
,283 ,
292 , 293
BRO DMAN N , K .,269
,270
,272 , 273
Bronch ia l tubes , nerves of, 226 , 238BRO UWER
,B .
, 142 , 1 72
BRUCE,A .
, 142
BRU CE , A . N .,130 , 1 42 , 1 93
BUCHANAN ,FLORE NCE
, 98 , 105Bulb (bu lbus ) , any bu lb- l ike strueture ; specifically the Medu lla oblongata, as in bulbar para lysis
,
tractus bulbo- spinal is .
See Reflexes of
See Gangl ion,
INDEX AND GLOSSARY
Bulb , olfactory, a protuberance fromthe cerebra l hemisphere conta ining the primary Olfactory center
,
1 12 , 120, 1 65 , 2 15 , 2 16 ,
Bulbar formation .
bulbaris.
Bund le. See Tract and Fasciculus.
bas is,fundam enta l
,or ground .
See Fascicu lus proprius.
longitud inal med ia l . See Fascicu
lus longitud ina l is m ed iali s.
poster ior longi tud inal . See Fas
ciculus longi tu d ina l i s med ialis.
solitary. SeeFasciculus solitari us.
BURDACH,co lum n of . See Fascicu
lus cuneatus .
BU RNETT, T. C .,279 , 299
See Form atio
CAJAL . See RAMON Y CAJAL .
CAJAL, commissura l nucleus of. SecN ucleus, commissura l , of CAJAL .
Calcar avis (h ippocampus minor) , aprojec tion into the posterior hornof the latera l ventr icle formed bythe calcarine fissure .
CAMPBE LL , A . W.,273 , 278
Canal , central (canal is central is) , theventri cle Of the spina l cord
,1 26 ,
129
latera l line. See O rgans, lateral
neural , the lumen of the embryonic Neural tube ; also applied tothe spinal Canal Of the vertebralco lumn .
semicircular (ductus sem icircu
laris) . See also Vestibular apparatus
,1 1 1
,183 , 184 , 187,
Spina l , the canal in the vertebralco lum n containing the Spina lcord.
CANNO N , W . B .,240, 241 , 248, 255,
CAPPS, J . A . , 250, 252
Capsule, external (capsula externa) ,a th in band of nerve- fibers forming the outer border of theCorpus striatum , 1 66 , 169 , 1 70
interna l (capsula interna) , a strongband of nerve-fibers passingthrough the Corpus stri atum ,

322
Center for pain . See Thal amus,pain center in .
prIm ary,
projection,See Projection cen
ters .reflex . See also Reflex circui t,109 , 1 13 , 129 , 156
respiratory, 237 , 238, 239 , 240semi ova l (centrum semi ovale, centrum ovale) , the great mass ofwh ite matter in the center ofeach cerebral hemisphere .
sensory,1 17 , 1 20
tac ti le . See Area , cu taneous ,Touch , apparatus of, and Center, cortical , somestheti c .trophi c , a nerve—center
. wh ichregulates the nutri tion O f an
other part,109
Of trunk and l imb reflexes,1 29
vasomotor . See Vasomotor ap
paratus .visceral . See Area , visceral .v isual . See Visua l apparatus andCenter, cortical , v isual .
Central nervous system . See Ner
vous system,centra l .
pause, 98Centrifugal . See Efferent.Centripetal . See Afierent.Centrum . See Center.Ce hali c , pertaining to the head , or
ected toward the head end of
the body
,as Opposed to caudal ,
16
Cerebellum , the mas sive coord inat ion center dorsa lly of the upperend of the Medulla ob longata,1 10 , 1 1 1 , 1 12 , 1 1 5
,1 1 7—1 20 ,
120—122, 143 , 152 , 1 58, 186—194 ,264
cortex of . See Cortex , cerebellar .development of, 1 1 7 , 1 18 , 1 19 , 187fiber tracts of
,1 30 , 137 , 1 76 ,
187 ,functib ns of
, 186 , 189 , 192,les ions O f
,1 89
Ces
r
iz
bration
, unconscious , 297 , 309 ,
Cereb rum , that port ion of the brainlying above the Isthm us ; also usedas synonymous wi th Presencephalon and Cerebralspheres, 12 1 , 122, 143, 1 60
hemi
INDEX AND GL OSSARY
Chain, sympathetic . See Trunk,sympath eti c .
Chemical processes in nerve~ cells ,96 ,sensib ili ty
, 72 , 85
Chiasma, Opti c (ch iasm a Opticum ) ,the part ia l decussation of the
opti c Tracts on the ventral surfaceof the brain
,1 18, 1 1 9 , 120, 208,
CHILD , C . M ., 3 1 , 36 , 97, 105
Ch impanzee, cerebra l cortex of, 282Chorda tym pani ,Chorioid p lexus (choroid plexus) . SeeP lexu s , chorioid.
Chrionom us p lum osos, nervous system of, 30
Chrom atin, a nucleo—protein substance found in the cell nucleus, 99
Chrom ato lysi s , the solut ion and d isappearance of the chromophi l i cSubstance from a neuron,
48 , 49 ,136 , 284
Chromoph il i c bod ies, granules, orsubstance. See Substance, chrom ophi lic .
C i liary process . See Process , cili ary .
Cingulum , an association tract of thecerebra l hemisphere lying underthe Gyrus cinguli , 267
Circ le of W illi s , a po lygonal circuitof anastomos ing arteries on the
ventral surface of the brain,from
wh ich some of the ar ter ies of thebra in arise.
C ircuit,organi c . See Reflex circuit.
C irculation of the b lood , apparatusof . See also Vasomotor apparatu s ,89 ,
CLARKE,co lumn of , or dorsal nucleus
of . SeeN ucleus , dorsal , ofCLARKE .
CLAUDIUS, cel ls of, 197C lau strum , a th in band of gray matter between the externa l Capsuleand the cortex of the island ofRE IL
,or Insu la .
Clava , an eminence on the dorsalsurface of the lower end of themedulla Ob longata produced bythe nucleus of the Fasc iculusgracili s, 130 , 1 64 , 176 ,
Coch lea , the bony spira woundcanal conta in ing the aii
ld itory re
ceptor, 85 , 183 , 196 ,

INDEX AND GLOSSARY
CO-consciousness , 297l enterates , nervous system of, 27 ,227
COGHILL , G . E .
, 66 , 67, 85 , 94, 135
Co ld , sensat ions of . See Temperature, apparatus of.
CO LE ,L . J . , 2 13
Co l ic, 250Col lateral , a smal l side branch of anAxon, 4l) , 44
Colli culu s facialis (eminentia faciali s ,eminent ia abducent is , em inent iateres
,Em inentia m ed ial i s ) , an
eminence in the floor of thefourth ventric le produced bythe VII nucleus and the Genuof the facia l nerve, 1 54
inferior, one Of the lower pa i r OfCorpora quad rigemina , containing ch iefly reflex aud i tory cen
ters,1 14 , 154 , 157 , 160, 164,
1 74 , 1 76 , 1 85 , 201 , 202
superi or (optic lobe, op ti c tectum ,
nates) , one of the upper pair ofCorpora quadri gem ina , containing ch iefly reflex Opt ic centers ,
21 1 , 264
COLLINS, J 233
Co lon,242
Co lumn,anterior . See Funicul us
ventrali s.
of BU RDACH . See Fasciculuscuneatus.
of CLARKE . See Nucleus , dorsal ,of CLARKE .
dorsa l (co lumna dorsa li s grisea .See Co lumn, gray. This nameis also applied to the dorsa lFuni cu lus) , 1 26 , 127 , 128
—130,of Forni x . See Forn ix co lumn .
fund amenta l . See Fascicu lu s propri us.
of GOLL . See Fasciculus graci li s .
gray (co lumna gr isea ) , one of thelongitud inal columns of neuroneswh ich make up the gray matterof the spina l cord . There are
three co lum ns : ( 1 ) dorsal (posterior) , (2) ventra l (anter ior ) ,and (3 ) lateral (midd le or inter
323
med iate) . These columns wereformer ly called horns (cornua) ;cf. a lso Funi culu s , 126 , 127 , 1 28 ,
Co lumn , intermed io—lateral , of sp inalcord
,229
lateral (co lumna laterali s grisea ;see Co lumn, gray) , 126 , 128 ,150, 1 5 1 , 229
of medul la Ob longata , 15 1—156posterior . See Funiculus, dorsal .somat i c motor , 150—156sensory
,150—156
of TURCK , the ventra l corticospina l Tract .ventral ( co lumna ventralis grisea ;
see Co lumn , gray. Th is termis a lso applied to the ventralFun i culus ) , 1 26 , 127 , 128 , 129 ,
ves icular . See N ucleus , dorsal , ofCLARKE .
viscera l motor , 1 50—156sensory, 150—156
Co lumna . See Co lumn .
Coma , 297Comma tract of SCHU LTZE . See
Fasciculus interfas cicularis .
Comm i ssure (comm issura) , a bandof fibers connect ing corresponding parts of the centra l ner voussystem across the med ian plane ;many decu ssations are alsoca lled comm issures , 265
anterior (commissura anterior) ,fibers pass ing transverse lythrough the Lamina terminalisand connec ting the basal port ions of the two cerebra l hemispheres , 1 14 , 162 , 165 , 222 , 265dorsal , fibers wh i ch cross the midplane O f the Spina l cord dorsa l lyO f the ventricle , 1 27of forni x . See Comm issure of hippocampus .
of GUDDE N . See Comm i ssure,postopti c .habenu lar (super ior comm issure) ,a band O f fibers connecting thetwo Habenula immed iately infront of the pineal Body, 265of h ippocampus (commissura h ippocampi , commissura forn ic is ) ,fibers connecting the Hippo

324 INDEX AND GLOSSARY
campi of the two sides throughthe Fornix body, 1 70, 220, 265 ,
Comm i ssure, inferior . See Com
m i ssure, postopti c .of ME YNE RT. See Comm i ssure,postopti c .
mi dd le . See Massa interm ed ia.
mol l is . See Massa interm ed ia .posterior (comm issura posterior) ,
fibers pass ing transverselythrough the anterior end of theroof of the midbra in,
162 , 220
postopti c ( infer ior commissure) ,fibers pass ing transversely acrossthe floor of the hypotha lamusassociated w ith the opti c chiasma ; contains the commissures O fGUDDE N , ME YNE RT
,and other
fibers , 265soft . See Massa interm edia .
super ior . See Comm i ssure , habenular .
of tectum (commissura tecti ) ,fibers passing transversely acrossthe roof of the m idbrain
,con
tinuing backward the Com
m i ssura posterior, 16 1ventral , fibers wh i ch cross the
midplane of the nervous systemventra l ly of the ventricle, 1 27 ,
Compensation of func tion in cortex ,
Components of nerves . See System ,
functional .Conarium . See Body, pineal .Conduct , neuro logi cal basis of, 262,3 12—3 16
Cond uction, avalanche, the summ ation of nervous im pulses in acenter so as to increase the
intens i ty of d ischarge,101
,
192
nervous , 38 , 54 ,9 6
, 97 , 98, 101Coz
nes of retina,205 , 206 , 207 , 208 ,
1 1
Confli ct in evolution,3 14
Conjunctiva,84 , 85 , 250
Consciousness , d issociation of, 297evo lu tion of . See P sychogenesis .
of lower animals ; cf . Psychogenesi s , 32 , 257 , 305multiple, 297
See Corpora
Consciousness , neurologica l mechanism of
,104 ,
1 66 , 224 , 242 , 249 ,258, 259 , 260 , 26 1 , 280 ,
28 1 ,286 , 287 , 290
—297 , 305—3 16seat of, 29 1of self
, 3 14
Cont inui ty of consciousness,297
Convo lut ion . See Gyrus.
of BRO CA. See Broca ’s convolu
tion.
Coordination,the comb ination of
nervous impulses in motor cen
ters to ensure the cooperation ofthe appropriate musc les in a re
action,35 , 130 , 132 , 133 , 181
Cornea,84 , 85 , 249
Cornu . See Horn.
Corona ra d iata, the Projection fiberswh i ch rad iate from the internalCapsu le into the cerebra l hemisphere, 164 , 166 , 169 , 287
Corpora quadrigemina, the dorsalpart of the Mesencephalon, con
ta ining the super ior and inferiorConfinflL
Corpus cal losum , a large band ofcommissura l fibers connectingthe N eopal lium of the two cerebral hemispheres
,1 1 9
,1 62 , 1 65 ,
cand i cans . See Body, mamm il
ary.
dentatum . See N ucleus, dentate.
fom icis . See Fornix body.
geni culatum . See Body, geniculate.
mamil lare. See Body, mamm iltp inea le. See Body, pineal .ponto-bulbare
,1 14
quadrigem inum .
quad rigemina.
restiform e (restiform body) , theinferior peduncle of the cerebellum ; cf . Peduncle, cerebe llar,192 , 201
stri atum (striate body) , a subcort ical or basa l mass O f gray and
wh ite matter in each cerebralhemisphere, 1 16 , 1 17 , 1 18 , 1 23,1 62 , 165
,166 , 167 , 168, 169 ,
170 , 174 , 176 , 215, 264 , 287

326
Degeneration,sclerotic, of cortex ,
284
DE ITE RS, cel ls of, in spiral O rgan, 197vestibular nucleus of, 184, 185
DEJE RINE ,J . J .
,287
Dement ia, 290Dend rite,a process of aNeuron wh i chconducts toward the cel l body, 39 ,
44 , 96 , 103 , 104
Dependency, corti ca l , a par t of thebrain stem developed as a subsid iary of the cerebra l cortex , 2 12 ,263 , 305
Depression,101 , 102 , 103
Development of the nervous system .
St
ee Nervous system , development0
DEWEY, J . ,20
, 6 1 , 67 , 308, 309 , 3 1 1
DEWITT,LYDIA M . A .
, 87 , 90, 95
Diaphragm ,innervation O f
,236 ,
237—240Diasch i s i s , a transitory defect offunct ion d ue to d isturbance ofcortica l equi l ibr ium ,
292 , 293, 294
Diencephalon (betweenbra in, thalam encephalon) , the bra in regionlying between Mesencepha lonand Te lencephalon ; sometim escal led Tha lam u s or Opti c tha lamus , but properly d ivided intoTh alamus, Epithalam us
, and
Hypothalam us , 1 1 6—1 19 , 121 ,—167development O i
,l 16—1 19
Diffusion of nerve impulses , 104 , 192Digestion, apparatus of,240—243
, 256Di lemma
,58, 307
D iop tri c apparatus of eyebal l,205
Disagreeab le and agreeab le. SeeAffect ion.
Discrimination .
cr im inat ive .
Dissociat ion of consciousness,297
Distention,sensations of
,89
Dogéf
é
i
énctions of cortex of
,
scratch reflex of,134
Dogfish , nervous system of .Fishes , nervous system of .
DO O IE L, A . S. 83 , 228
,233
DO LLEY,D . H.
,
Dol
tph
z
i
o6absence of olfactory organs
0
See Reaction,d is
See
INDEX AND GLOSSARY
Dorsa l , on the back side of the body,termed posterior in the B . N . A .
l ists,1 16
Ductus coch learis, 1 95 , 1 96 , 1 97, 198endo lymphaticus, 195 , 1 96reuniens , 1 96semicircularis . See Canal , semic ircular
,and Vestibular appara
tus .utriculo- saccularis, 1 96
Dura m ater, the outer brain m em
brane, 38DU RU PT,
A .,1 93
Dynamic theory of consciousness,293, 296
E ar . See Aud itory apparatus andVestibular apparatus .brain
,1 12
, 123
evo lution of,1 99
,201
E arthworm,nervous system of, 28
E ctoderm (epib last) , the outer germlayer of the embryo , from wh i chthe epidermis and the Neural tubedevelop
,204
EDGEWO RTH, F . H.
, 180
EDINGE R,L .
, 36 , 1 15 , 124 , 1 59 , 2 19 ,
E DIN GE R—WE STPHAL,nucleus of (the
v iscera l efferent nucleus of the I I Inerve ; cf . N ucleus of oculomotornerve) .
E ducation, 32, 3 12
-3 16Efiector, an organ of response , 26 , 92
Efierent, conducting away from acenter
,25
,42
,108, 126 , 137, 145
150
E lectr i c excitab i l i ty of nervous tissues
,281 , 283 , 286
phenomena in nervous tissue,96
Embryology of nervous system , SeeN ervous system , embryology of .
Eminent ia abducentis . See Col
liculus facia li s .facial is . See Coll i culus faciali s .hypogloss i . See Trigonum hypogloss i .
m ed i a li s (em inentia teres) , amed ia l longitud ina l ridge in the
floor of the fourth ventr i cle ; anenlarged por tion is the Colliculus faciali s .

INDEX AN D GLOSSARY
Eminentia teres . See Em inentiamedia l i s .vagi . See Ala cinerea .
Emot ion . See Affection .
Empis stercorea, nervous system of30
Encephalon, the brain,120
E ndo lymph , 196 , 198End -organ , the peripheral apparatusrelated to a nerve ; a Receptor orEfiector, 25 , 40 , 70, 79 , 98
End -plate, motor , the terminal arborization of a motor axon upon amuscle-fiber, 40, 92 , 1 0 1
E ndyma . See Ependym a .Engram ,
295
Env ironment,17 , 18 , 69 , 3 12
Epencephalon, the cerebel lum .
Ependym a (endym a) , the liningmembrane of the ventricles of thebrain, derived from the or igina lepithel ium of the Neural tube , 38
E pib last . See E cto d erm .
Epicri ti c sens ib i l ity, a h igh ly refinedtype of cutaneous sensib i li ty
,espe
cially on hair less parts,84 , 85 , 132
Epiglott is,organs of taste upon ,
147 ,243
E pinephr in . See Adrenal in .
E piphys is . See Bo d y, pineal .Epi tha lam us, the dorsa l subd ivis ionof the Diencephalon, conta iningthe pineal Body and the Habenula, an important olfactory correlation cen ter, 1 1 1 , 1 12 , 1 18 , 1 19 ,1 21 , 122 , 1 62 , 1 65 , 167 , 220, 22 1
Epith eli um , a th in sheet O f cel ls , 24nerve end ings in , 90
O lfactory (Schneiderian m em
brane) , 2 17E qu il ibrium , apparatus of . See alsoVes tibular apparatus , 77 , 88 ,
nervous, 66 , 293 , 296
theory of consciousness , 296E sophagus
, 78 , 90 , 144 , 147 , 234 , 242
E stheti c experience. See Affection .
E th i cs . See Mora ls .E ugen ics , 3 12E ustachean tube (aud itory tube) ,1 9 5
E vo lution of mind . See P sychogenesi s .
See Tract,
E vo lu tion of N ervous system . SeeN ervous system , evo lution of.
EWALD, J . R . ,
184 , 203
E xci tab i li ty,e lectric . See E lectri c
excitab ili ty of nervous t issues .E xci tation
,fatigue of, 101 , 102 , 103
Experience, learning by, 34, 294 , 307,Exteroceptor, a sense organ exci tedby stimu l i aris ing ou ts ide the body,74 , 77 , 79
E xteroceptors , apparatus of, 132,
137 , 138, 1 39 , 14 1 , 145 , 163- 167 ,
172—174 , 250Extirpation of cortica l centers , 286Eye. See Visua l apparatus .accommodation of . See Aecomm od ation of v is ion .
bra in (ophthalm encephalon) , 1 12,123
conjugate movements of, 186 , 2 1 1 ,283
developmen t of, 1 16 , 1 17, 204evo lution of, 2 12muscles of, 92 , 1 10, 143, 146 , 148,
par ieta l . See Parietal eye.
p inea l . See Parietal eye.
Face brain,1 23
Facul ties , mental , 280, 285 , 290 , 29 1Falx cereb ri , a longi tud ina l fo ld ofDura m ater wh ich extends between the cerebra l hem ispheres inthe longitudinal fissure .
Fas cia dentata . See Gyrus dentatus .
Fascicu lus , a bund le O f nerve-fibersnot necessarily of s im ilar functional connections . The termis O ften use d , however, as asynonym for Tract, 128 .
antero- latera l is superfi cia l is (ofGOWE RS) . See Fasc iculus ven
tro- lateral is superficialis .
cerebel lo-spina l is . See Tract,spine-cerebellar .
cerebro- spina li s .corti co- spinal .circumolivaris pyramid is
,1 14
comm unis , a name former ly applied to the Fascicu lus so l i tarius in lower vertebrate brains .

328 INDEX A ND GLOSSARYFasci culu s cuneatus (column of BU R
DACH) , the lateral port ion ofthe dorsal fun icu lus of thespina l cord
,128, 130, 139 , 1 76 ,
1 77
nucleus of . See Tuberculumcuneatum .
dorso- latera lis (LISSAUE R ’S zone,L ISSA UE n
’
s trac t) , 1 30of G ows ns . See Fasciculusventro- lateral is superficialis .
grac i li s (co lumn of GO LL ) , themed ial portion of the dorsa lFuni culus of the sp inal cord ,
nucleus of . See Clava.inner spiral , of Spiral organ,
1 97
interfascicularis (comma tracttract of SCHU LTZE ) , 1 30, 13 1longitud inalis inferior of cerebralhemisphere
,222 , 267longi tud inalis m ed iali s (med ial
longitud ina l bund le,posterior
longi tud ina l bund le , fascicu luslongitud ina l is posterior or dorsalis) , a bund le of motor coord ination fibers running through thebra in stem ,
13 1 , 152 , 1 55 , 1 56 ,
longi tud ina lis superior of cerebralhemisphere
,267
marginalis ventral is , 13 1of ME YNE RT . Sec Tract
,haben
ule- peduncular .occipi to—fronta lis inferior of cerebra l hemisphere
, 267
proprius of cerebra l hemisphere .
See Fibers , arcuate ( l ) .propri us of spinal cord (fundamental columns
,bas is bund les
,
ground bund les) , that portion ofthe wh ite matter of the spina lcord wh ich borders the graymatter and conta ins correlation fibers ; arranged in dorsa l ,lateral , and ventra l subd ivisions , 127 , 130 , 13 1 , 133, 179 ,
retroflexus of ME YNE RT. SeeTract, habenulo—peduncular .
solitari us ( tractus so li tar ius , solitary bund le, in lower verte~brates often called fasc icu luscommuni s) , a longi tud inal
bund le of fibers in the medullaob longata conta ining the centra lcourses of the v iscera l sensoryroot-fibers of the crania l nerves
,
149 , 150,1 55 , 1 56 , 1 64 , 234 ,
237 , 239 , 240 , 24 1 , 244 , 247
Fasc icu lus sulco-margina l is, 130,
13 1
tha lamo-mamil laris . See Tract,mam illo- thalamic .transversus occipi tal is of cerebralhem isphere
,267
unc inatus of cerebra l hemi sphere,267
ventro- lateral is superfic ialis (antero- latera l fasciculus
,GOWE RS’
tract) , 128 , 130Fatigue
,101—103 , 255 , 256 ,
258, 304
Fear . See also Affect ion, 89 , 255 ,
250 .
Feeb le-mindedness . Sec Id iocy.
Fee d ing , reflexes of Sec Reflexes offeed ing .
Fee ling (affective) . See A ffection .
Feel ing tone . See also A ffection,249 ,
254 , 258—262
FE RRIE R,D .
, 194
Fiber , or fibers,fibrac. See N erve
fiber.arcuate, of the cerebral hemisphere, short associationfibers connecting neighboringgyri ; a lso ca l led fibra? propri-ccand fasciculi propr i i , 267 , 285of the medulla ob longata, decussating fibers lying in a superfic ial ser ies (externa l arcuatefibers ) and a deep ser ies ( interna l arcuate fibers ) , 155
assoc iation ; of. Tract , association,
206 ,267 , 269 , 285 , 287, 29 1
- 293
of MULLE R ,205
,206
postganglioni c . See Neuron, postganglionic .
preganglioni c . See Neuron, pregangli oni c .projection . See Pro
’
ection fibers .proprize (arcuate bers of the
cerebral hemisphere) , 267 , 285Field , aud i tory psych i c , 285corti cal , a term somet imes used asa synonym of Center, cortical , orof Area, corti ca l .visua l psychi c
,285

330 INDEX AND GLOSSARY
Fornix co lumns (columnar forni cis, Funi cu lu s , lateral (funiculus lateranterior pi llars of fornix ) , twoco lumnar masses of fibers d i verging from the anterior end of theFornix body to descend into thediencepha lon ,
165 , 170, 220, 221
comm issure. See Comm i s sure ofhippocampu s .
crus of (crus forn i cis, poster iorp il lar of fornix) , a band of fiberson each s ide of the brain eon
necting the posterior part of theFornix body wi th the Fimb ria.
longus of FORE L, fibers wh i ch perforate the Corpus callosum and
pass through the Septum pelluci
d um .
Fossa flocculi , 14 1laterali s (fossa of SYLVIU S) , adeeper part of the Fissuralateral is containing the Insula.rhomboi d al , the floor of the fourthventricle, 1 18
Fovea limbica (sulcus rh inalis, fissura rhini ca, fissura rh inal is, fissura ectorhinalis) , the sulcuswhi ch marks the lateral border ofthe latera l Area olfactoria and
Gym s hippocam pi or pyrifoLobe in the lower m amm a ls .FRAN Z
, S. I.,299
Fle ed om of act ion,3 15
h E Y , M . VON , 79 , 84 , 85 , 94Farrscn , G .
,281
,299
Frog,7
e
9
erebral cortex of, 2 16 , 264 ,
nerve end ings in,90 , 92
olfactory receptors in, 92
reaction tim e of , 98reactions of
, 63
veloci ty of nervous transm iss ionin
, 97Funi culus, one of the three principald ivis ions of wh i te matter on
each s ide of the spina l cord ;these funi culi were formerlyca lled Co lumns, 128
d orsal (fun iculus dorsa lis or posterior
, posterior columns ) , thewh i te matter of the spinal cordincluded between the dorsalfissure and the dorsal root
,1 28 ,
1 30, 134 , 138, 14 1 , 1 50, 1 5 1 ,175 , 176 , 177, 178, 1 79 , 3 10
a lis , lateral co lumns ) , the whitem atter of the spinal cord inc luded between the dorsal andventral roots
, 128
ventra l ( funiculus ventrali s or anterior, ventral , or anterior co lum us) , the wh i te matter of thespinal cord included between theventral fissure and the ventralroot
, 128
GALL,F . G .
,280, 281 , 300
Gangl ion, a collect ion of nerve-cells .
In vertebrates the term should beapplied only to periphera l cel lm asses
,though sometimes Nuclei
w i th in the brain are so designated ,108, 109
Gangl ion or ganglia, basal , a termsometim es applied to the Corpus striatum and other subcorti cal par ts of the cerebralhemi sphere.
branch ia l,of vagus , 149
cerebro- spinal,development of,
45 , 225
cerv i cal , inferior, 226m iddle, 226superior
,226 , 234
c i l iary, 143 , 146 , 149 , 226 , 231 ,246
of CO RTI . See Ganglion, spiral .of facial nerve. See Gangli on,
geni culate .
GASSE R ’S. See Ganglion,semi
lunar .geni culate (ganglion geni cul i , the
ganglion of the VII cranial orfacia l nerve) ,
habenulae. See Habenula.
of insects,29 , 30
interped unculare . See Nucleus ,interpeduncular.of invertebrates , 28, 29 , 30, 227jugular (gangli on jugulare) , 147,149
of lateral line nerves , 149nodosum
,147 , 237 , 239 , 240
opti cum basa le. See Nucleus,preoptic .ot i c
, 147 , 245

INDEX AND GLOSSARY
Ganglion, petrosa l (ganglion petrosum ) , 1 47, 245ofI
SCARPA . See Gangl ion vestibuar .
semi lunar (ganglion semilunare,GASSE R ’ s ganglion, the ganglioriof the V cran ial or trigem inalnerve) , 45 , 1 1 1 , 1 12 , 146 , 180,245
sphenopalatine,226 , 245
spina l , 25 , 43 , 109 , 1 25 , 126 , 134 ,
spira l (ganglion spirale, gangli on ofConn ) , 147submaxi l lary
,146
superior (ganglion superius of IXcranial nerve) , 147supra-esophageal
,29 , 30
sym pathet ic , 53 , 107 , 109 , 125 , 126 ,225 , 226 , 227 , 230, 237, 238,239
prevertebral , sympathet i c ganglia of the thorax and abdom en other than those oi thesympatheti c trunk .
vertebral , the ganglia of thesympath eti c Trunk .
of trigem inus . See Ganglion,
sem i lunar .of vagus . See Gangli on,
jugular ,and Ganglion nodosum .
of vertebrates , 108 .
vest ibular (ganglion of SCAR PA) ,l 7
GE HUCHTE N ,A . VAN
,25 , 45 , 86 , 194
Generative organs . See Sexual organs .
G enil
c
a
ulate body. See Body, genicute.
ganglion . See Gangli on, genien
late .
GE NN AR I , layer of st ripe of . SeeL ine of Gennari .Genu , a knee- shaped bend of an
organ, such as the genu of thecorpus ca llosum
,of the facial
nerve, etc .
of corpus callosum , 1 19
G i ll s. 236 , 240innervat ion of, 1 10, 1 1 1 , 1 12, 149 ,245
muscles of, 94 , 148G land
, adrenal . See G land , suprarenal .
331
G land , intes tinal , 224nerve—endings on, 94
pinea l . See Body, pineal .pi tui tary. See Hypophys i s .salivary
,innervati on of,
154 ,244
suprarenal , 231 , 255 , 256G li a . See Neurogl ia.
G lom eruli , o lfactory, sma l l globularm asses of dense Neuropil in the
o lfactory bulb containing the firstsynap se in the olfactory pathway,
G lycosur ia,255
GO LDSTE IN , K . , 13 1 , 194
G ()2
LGI,G .
,41
,43 , 44 , 49 , 55, 190,
74
GOLL, co lumn of . See Fasci culusgracil is .GO LTZ , F . ,
279 , 280, 281 , 300
GOWE RS, fasciculus of. See Fascieulus ventro—lateralis superficialis .
Grad ient , phys io logical , in nervefibers
, 97
Granules . See Cells , granule.
chromoph ili c,tigro id , of N ISSL .
See Sub stance , chromophi l i c .Gray, central , relat ively und ifferen
tiated gray Matter wh i ch reta in si ts primi tive pos ition near the
ventricles , 127 .
Groove,med ullary. See Neural
groove.
neura l . See Neural groove.
GR ij NB AU M,A . S. F .
,282 , 300
GUDDE N ,comm issure of See Com
m i ssure, postopti c .Gus tatory apparatus
, 72 , 74, 9 1 ,143
,144
, 146 , 147 , 148 , 149 , 1 50,157 , 163 , 2 18, 222 , 234, 243
—246 ,303
Gyrus , one of the convolutions orfo lds of the cerebra l cortexbounded by Sulci or Fissures ,265 , 266
angular is , 12 1central is an terior (precentral gyrus ) , 1 2 1 , 140, 181 , 269 , 27 1 ,272 , 282 , 283 , 285 , 286 , 288
posterior (postcentral gyrus) ,1 2 1 , 268, 270, 282 , 283 , 285 ,286
,288
cinguli , 1 19 , 170

332
Gyrus dentatus ( fascia dentata) , asubsid iary gyrus of the Hippocampus , 22 1 , 222forni caras (lim b i c lobe) , the marginal port ion of the cerebralcortex on the m ed ia l aspectof the hemi sphere, includ ingthe Gyrus cinguli , Gyrus hippocampi
,and others ; there is a
var iety of usage regarding i tslimi ts
,273
fronta l is inferior, 12 1 , 1 70 , 292med ius, 12 1superior
,1 19 , 12 1
h ippocampi , that part of the cerebra l cortex wh i ch borders theHippocampus . Part of i t ( theUncus ) is Arch ipal li um ; the re
m aind er is trans i tiona l to the
N eopa l lium . See Lobe, pyriform , 2 17 , 2 19 , 221 , 222 , 273 ,284
l ingual is , 1 19occipitalis lateral is
,12 1
o lfactorius latera lis . See Nucleusolfactorius lateralis.
medi alis . See Area parolfactoria of Broca
orb ital is,121
postcentral . See Gyrus centra l isposterior .precentral . See Gyrus central isanterior .
subcal losus (pedunculus corpor iscallosi ) , part of the Nuc leus olfactorius m ed iali s , 1 1 9 , 2 19
supram arginalis , 12 1
tempora li s inferior,12 1
med ius,12 1
superior, 12 1 , 1 70uncinatus . See Uncus.
Habenula (nucleus habenulae , ganghon habenular ) , an important oi
factory correlation center in theEpithalam us , 1 62 , 165 , 167 , 170 ,220
Hab i t, phys iological , 32 , 294 , 304Ha i r cel ls (cel ls of Con'
r i ) , 197 , 1 98 ,199
innervation of, 80 , 81
HARDESTY,I. , 1 98, 199 , 203
HARRIS, W.
,2 13
INDEX AND GLOSSARY
HEAD , H. , 7 9 , 84, 85, 94, 95, 132,142 , 166 , 171 , 172 , 173 , 1 75 , 1 79 ,
Hearing , organs of . See Aud i toryapparatus .
Hear t, innervation of, 144, 147, 232,234
Heat , sensations of .ture, apparatus of .
HE IDE NHAIN , M .,49 , 55
HE LD, H.
,50
HE LMHO LTZ , H. L . T. VON ,198,
203
HE LW IG , tract of . See Tract,o live
spinal .
Hemispheres, cerebellar , 120, 187cerebral , 63 , 64, 12 1
, 122 ,123 , 129 , 2 19 , 264 ,
comparative anatomy and evo
lut ien of,1 1 1 , 1 12 , 129 , 2 15 ,
264 , 265 , 279 , 280, 290, 294 ,301—306
Hemorrhage, cerebral , 293HE NSE N ,
cells of,19 7
stripe of , 197, 1 98HE RRICK , C . JUDSON , 36 , 6 1 , 66 , 67 ,95 , 124 , 135 , 142 , 1 59 , 17 1 , 182,194, 200 , 223 , 248, 263 , 303
HE RRICK , C . L ., 18 , 108, 1 94, 258,
262,296 , 300 , 303
HE RRICK , F . H .,01
, 68
HE RTZ , A . F .,95 , 242 , 243, 248
Hibernation,nerve cells in
,102
Hindbrai n, a term wh i ch has beenvarious ly app l ied to the corebellum
,the cerebel lum and pons, the
medulla ob longata, and the ent ire
rhombeneephalon .
Hippocampa l gyrus . See Gyrus hippocampi .
Hippocampus (hippocampus m aj or,Ammon ’s horn
,cornu Ammo
nis) , a submerged gyrus formingthe larger part of the Archipall ium , or olfactory cerebral cortex,
2 17 , 2 19 , 220 , 22 1 ,284 , 306
commissure of. See Comm i ssureof hippocampus .minor . See Ca lcar avi s .
HIs,WILLIAM
,49 , 55 , 1 15
—1 18 , 124Hi sto logy, the study of Tissues ,27
See Tempera

334
Jelly-fishes, nervous system of, 27,227JE NN INGS, H. S.
,37 , 68
1 24,
Jo ints, nerve-end ings in, 88
Kangaroo, cerebral cortex of, 2 17KAPPE RS, C . U . AR IE Ns , 203 , 223 ,
KARPLU S,J . P .
,25 1 , 262
o lasm , the
11protoplas
m of the
nuc eus of a ceKE IBE L , F.
,124
KOLL IKE R ,A . ,
44KRAU SE , W.,1 15 , 1 24
end -bulbs of, 84 , 85KR E IDL
,A .
,25 1 , 262
KR IE s,J . VON , 7 1
KUN '
rz,A .
,233
L ab ium vestibulare, 198Labyrinth of car, 1 95 , 196Lactic acid , 103LADD
,G . T.
, 98,Lageria,Lamina . See also Layer .affixa, a thin non-nervous part ofthe med ia l wall of the cerebralhemi sphere attached to the
tha lamus and bordered by thelateral chor io id P lexus .of neural tube. See P late.
term inal i s (termina l p late) , theanterior boundary of the thi rdventri cle
,1 18 , 165 , 2 15 , 264 , 265LAN CISI (LANCISIU S) , nerves of . See
Stri a longi tu d inal i s .striae of. See Stria longitu d ina li s .LANDACRE
,F . L .
, 35LA N GE , G .,259 , 262
LANG LE Y, J . N .
,
Laqueus . See Lemrii scus.
Larynx,239
Lateral line organs .lateral line.
Law, myelogenetic , of FLE CHSIG , 287
yer. See also Lamina .
of BAILLAR GE R . See L ine of Bai llarger.
oi cerebellar cortex,1 90
of cerebra l cortex,268—274 , 290
See O rgans ,
INDEX AND GLOSSARY
Layer of GE NNAR I . See Line of
Gennari .of retina,
Learning. See E xperience, learningy
Lemni scus (fillet, laqueus) , sensoryfibers of the second order terminating in the thalam us .
acoust i c . See Lemn iscu s , lateral .bulbar , ascend ing sensory fibers ofthe second order from the
medul la ob longata to the thal amus
,includ ing severa l di fferent
tracts , 157gustatory. See Lemni scus, vis
ceral .lateral , the acoust i c lemniscus,
fibers from the coch lear nucleito the co l liculus infer ior andthalamus
,1 14 , 1 57 , 16 1 , 163,
m ed ia l , ascend ing fibers of theproprioceptive system from the
sp ina l cord to the thalamus , 138,
2 10
optic , a term wh i ch m ight ap
propriately replace Opti c Tract,209
spinal , ascending fibers of touch ,temperature, and pain sens ib il ityfrom the spinal cord to the
thalamus . In the cord thesefibers form two tracts, the dorsal and ventral spine- thalam i ctracts
, 1 30, 13 1 , 134 , 138, 139 ,
trigeminal , ascend ing sensoryfibers of the second order fromthe sensory V nuclei to thethalamus
,139 , 14 1 , 1 57 , 1 6 1 ,
vi sceral , a name suggested for theascend ing secondary fibers fromthe nucleus of the fascicul us so litar ins to the hi gher cerebra l centers,
LE NHossEK, M . VON ,4 1
Lens,204—208 , 2 1 1 , 2 12
LEWANDOWSKY , M , 37, 1 94, 300Life, defini t ion of, 17L Igam ent
,spiral , of Cochlea, 1 97

IN DEX AND GLOSSARY
L imbus lam inae irali s, 1 97
Limen insulaa. ee Nuc leus olfac
torins laterali s, 2 19L ine of Bai llarger, a stripe of tan
gential wh i te fibers in the cerebral cortex ; there is an outer andan inner line, 267 , 268 , 274
of Gennari , a stripe of tangent ialwhi te fibers in the Area striataof the cerebral cortex ; i t is theouter Line of Bai llarger in thi sarea
,268, 274
L ingula cerebelli , a sma ll em inenceon the ventra l surface of the cerebellum where the anterior m ed nl
lary Ve lum j o ins the Vermi s ,1 62
LISSAUE R , tract of, zone of . SeeFasc iculus dorso- lateral is .Lizard , parietal eye of, 2 1 2Lobe
,frontal , 120, 266 , 283
of the lateral line ( lobus Iineaelateralis) , a hi ghly di fferent iatedpart of theacousticO—latera l Areaof fishes , 1 52limb ic . See Gyrus forni catus.
occip ital,266 , 283
olfactory ( lobus olfactorius) , theo lfactory Bulb , i ts Crus, and theanterior part of the Area olfactoria ; thi s is the B . N . A . usage ;the term is somet im es applied tothe olfactory Bulb al one and
sometimes to the Area olfac
toria a lone .
Op ti c . See Colli cu lus superior.parieta l , 266pyriform ( lobus p iriform i s) , thelateral exposed port ion of theolfacto ry cerebral cortex inlower mamm als, bounded dorsa lly by the Fovea limbica ; inman i t is represented by theUncus and part of the Gyrushippocampi , 217temporal , 120 , 201 , 202 ,
2 19 , 266
vaga l . See Lobe, visceral .visceral ( lobus v iscerali s , vagallobe, lobus vagi ) , the vi sceralsensory Area of fishes
,148, 149 ,
Lobulus paracentralis , 1 19par ietal is inferior, 12 1superior
,1 21
335
Maeula sacculi,196
utr icu li , 196MAGE ND IE , foramen of .m en of Magend ie .
MALL, F . P . , 124
Mamma ls,cortical regions of, 273
Mammillary body. See Bod y,mam
Mantle. See Cortex, cerebral .MARCH] , method of, 48, 135MAR IE , P .
, 300
Marsupia l anim als , cerebral cortexof
,2 17
Massa intermed ia (comm i ssura mollis
, soft commissure, midd le commissure) , a band of ay matterconnect ing the med ia surfaces ofthe two thalami across the th irdventricle ; i t is not a true comm issure, 1 1 9 , 1 62
MAST, S. O .
,2 13
Mast ication , apparatus of, 78, 143,146 ,
180 ,244 , 247
Matter, centra l gray. See Gray,central .
See Fora
Loca l si gn ; cf . Locali zation Of sensat ion
, 84 , 229 , 250, 259Locali zat ion of funct ions in centra lnervous system , 65 , 1 13, 230
232, 234 , 280
in cerebe llar cortex, 189 , 28 1 ,305
in cerebral cortex , 189 , 273, 280,281 , 282 , 283 , 284
—29 7 , 305 ,307
of sensation, 79 , 84, 85 , 228
—230,250 , 259 , 286Locomot ion , reflexes of, 134LO E B , J .
, 37, 6 1 , 68
LOWE NTHAL , tract of. See Tract,tecto—spina l .LUCIAN I , L .
,194
LU GAR O ,E .
, 104
Lumbricus, nervous system of, 28
Lungs , innervat ion of . See Respiratory apparatus .LU YS, body of . See body of LUYs .
Lyra . See Lyre of Davi d.Lyre of Davi d ( lyra David is, psalterium ) , the pos terior part of theFornix body, includ ing the Commi ssura hippocampi .

336
Matter , gray (substantia gr isea) ,gray nervous t issue composedch iefly of nerve- cel ls and un
mye l inated nerve-fibers, 108, 1 28whi te (substantia alba) , wh i te nervous tissue composed ch iefly ofmyelinated nerve-fibers
,108,
1 27 , 128 , 130
Meatus,external aud i tory
,1 9 5
Med ial (med ial is) , nearer the med ianplane ; Opposed to latera l .
Median (med ianus ) , lying in the axisor midd le plane of the body or oneof i ts members .
Medi us , intermed iate between twoother parts.
Medulla ob longata (bulb) , theMyelencephalon B . N . A . the o lderand better usage includes thewhole of the Rhombencephalon excep t the Cerebel lum and
P ons , 1 10, 1 1 1 , 1 12 , 1 16—120 ,1 2 1
,122
,143
,152 , 154 , 1 62 ,
232, 244 , 246 , 302 , 303
reflexes of , 1 43 , 148 , 18 1 , 234,
sp inal is . See Spinal cord .
Medullary sheath . See Myelinsh eath .
See N eural tube.
ME ISSNE R , corpuscle of, 82 , 83plexus of (submucous plexus) , 53 ,24 1
Membrane, basi lar, of spiral organ,
of the brain . See Meninges .l imi ting
,of retina (membrana
l imi tans externa and interna) ,207
mucous , nerves of, 90 , 1 26 , 146 ,
nuclear, 99
Schneiderian . See E pi thelium,O l
factory.
tector ia l , 197 , 198 , 199tympani c (drum membrane) , 85 ,19 5
, 196 , 24S, 249
vestibular (membrane of RE ISSN E R ) , 1 9 7
Memory,295
,297 , 304 , 307
assoc iative, 32, 65 , 242 , 294 , 29 5 ,296 , 307
Menid ia, nerves of , 148 , 149 , 200spinal cord of
,1 50
IN DE X AND GLOSSARY
Meninges, the membranes of thebrain and Spinal cord
,38, 146 , 250
ME RKE L,corpuscle of
,8 1 , 82 , 83
Mesencephalon (midbrain) , the Corpora qua d ri gemina and cerebralPedunc les , 62 , 63 , 1 16—1 19 , 121 ,122
,l 60
,16 l , 232
development of, 1 16 , 1 19 , 160Metabolism , chemical changes inprotoplasm , 96 , 97 , 99 , 163
Metathalamus, the posterior part ofthe Tha lam us , comprising the m e
d ial and lateral geni culate Bodies,
Metencephalon (hindbrain) , the an
terior part of the Rhom bencephalon, includ ing the Cerebe llum ,
Pens , and intervening part of theMed ulla ob longata, 1 1 7—1 1 9 , 12 1
ME YE R,A .
,49 , 55 , 293 , 300
ME YE R, MAX,
262
ME YNE RT,commiss ure of.
Comm issure, postopti c .decus sation of ( founta in decussat ion , dorsal tegmental decussation) , 16 1fasciculus retroflexus of .Tract
, habenulO-peduncular .
MICHE LSON ,A . A .
, 7 1
M idbrain . See Mesencephalon.
MILLIKAN,R . A .
, 7 1
M ind . See Consciousness .evolution of . See P sychogenes is.
unconscious,296
,297
Mo lecular substance, Molecularlayers, a name applied to the
N europi l .MO LHANT, M .
,248
MONAKOW,C . VO N ,
17 1 , 287 , 293 ,294 , 300trac t of. See Tract , rubro—spinal .
MONR O,foramen of . See Foram en
interventricu lare.
Moon-eye , brain of , 302 , 303Morals
, 3 13 , 3 14 , 3 15
MORGU L IS,S.
,242
,248
Motor apparatus , 62 , 63 , 1 17 , 120,1 54 , 163 , 180
—182,186 , 192, 234 ,
MO YE S, J . M .
,278
Mucous membrane, nerve end ings in.
See Membrane, mucous .Mii LL E R
,fibers of
,205 , 206
Mul tiple consciousness,297

338 INDEX AND GLOSSARY
N erve, auricular, 144 , 147branch ia l , 1 10, 1 1 1 , 1 12 , 149 , 245bucca l
,200
card iac . See Heart, innervationO f.
cerebral . See Nerve , crania l .cerebro- spinal , the peripheralnerves connected with the bra inand spinal cord , 107cervica l , 107 , 130 , 237chorda tympani
,146 , 245
ci liary,146
coccygea l,107
coch lear , 145 , 1 57, 183, 185 , 1 9 7 ,components
,tab le of
,146 , 147
crania l (cerebra l nerve) , a per ipheral nerve connected w i th thebrain ; these nerves are enu
m erated in 1 2 pa irs , 10 6, 1 1 0 ,1 1 1 , 1 1 2 , 143 , 144
—150,1 52 ,
of fishes,1 1 0
,1 1 1 , 1 1 2 , 148 , 149
cutaneous, 79—86
,125
,132 , 134 ,
143,145—150 ,
157 , 172—180,
of CYO N,
of deep sens ib i l i ty ; cf . Proprioceptors , apparatus of, 79 , 86 ,132 , 172
—180depressor
,of heart
,235
efferent. See Efierent.exci to- glandular
,108
facial (VII cran ial nerve, facialis) ,146
, 148,1 50
,23 1 , 236 , 243,
glossopharyngeal (IX crania lnerve) , 1 10 , Il l , 1 1 2 , 1 14 , 120 ,
gustatory, 243—246hyomand ibu lar
, 1 10—1 12 , 149
hypog lossus (XII crania l nerve) ,120 , 144 , 147 , 148
, 1 50,153 ,
156
inhibitory, a nerve wh ich checksor retards the action of theorgan in wh ich i t terminates
,
intercostal , 125 , 237 , 238 , 239intermed iate (nerve of WR ISR E RG
,
pars intermed ia facia l is,portio
intermed ia facial is, the sma ller
of the two roots of the VIIcra
é
n ial nerve) , 1 14 , 1 20,24
Nerve, intestina l , 1 1 1 , 149of JACO BSO N . See N erve
,tym
pani c .oi LAN CISI . See Stria longitud iua l i s .laryngeal , 147lateral (nervus lateral is, lateralline nerves) , branches O f theVII
,IX
, and X crania l nerveswh i ch supply the Lateral lineorgans , 1 10 , 1 1 1 , 1 12 , 145 ,148 , 149 ,accessory
”
(ramus lateral is ac
cesso1 1l i s facialis) ,cutaneous
,125
lingual,147 , 245
lu’mbar,107 , 130 , 23 1 , 232
mand ibular,1 10 , 1 1 1 , 146 , 200, 245
maxi l lary,1 10 , 1 1 1 ,
motor, a peripheral nerve wh ichconducts efierent impulses to amusc le
,108 , 120, 145
—150oculomotor ( l II cranial nerve) ,
246
olfactory (nervus o lfac torius, thefirst crania l nerve) , 9 1 , 1 1 0 , 1 1 1 ,
Ophthahnk; 1 1 0 , 1 1 1 , 146 , 149 ,
Opti c (nervus opticus , the secondcran ia l nerve) ; th is is not a truenerve
,but
,in real ity
,a cerebra l
tract ; cf . Tract, opti c , 1 1 1 , 1 20,
otic,149
of pain,249
,25 1
,252 , 253 , 254 ,
pa latine, a nerve of fishes correspond ing to the human greatsuperficia l petrosa l nerve
,1 10,
par ieta l (nerve of the Parieta leye) , 2 12phreni c
, 236 , 237 , 238 ,239
pneumogastric . See N erve, vagus .postganglioni c . See Neuron, postganglioni c .

INDEX AND GLOSSARY 339
N erve, pregangl ion ic . See N euron,preganglioni c .presp i racular (pretremati c branchof the facial ) , 1 1 1 , 149pretremat ic , of facia l, 1 1 1 , 149recurrent , 226sacra l
,1 07 , 130, 23 1 , 232
sciat ic,97
sensory, a periphera l nerve wh ichconducts afierent impulses froma sense organ to the spina l cordor brain
,108
,126
somati c,126 , 1 39 , 145 , 1 72
spinal , a periphera l nerve con
nected w i th the spina l cord,
centra l connections of,1 29—140
,
150 , 1 5 1 , 25 1 , 252 , 253 , 254
components of,145
,147 , 150,
15 1
splanchn ic,226
SUperfieial petrosal , 147, 245supratemporal , 149sympathet ic . See N ervous system ,sympathetic .
of taste . See Gustatory apparatus .term inal , a slender nerve associated w i th the O lfactory nerve,thoracic
,107 , 125 , 126 , 130, 23 1 ,
232,236
,237
trigeminal (trifacial nerve,V
crania l nerve) , 1 10 , 1 1 1 , 1 14 ,1 20 , 14 1 , 143 , 145
,146
,1 48 ,
149,1 50
,152 , 154 , 157 , 1 74 ,
1s0,243 , 244 , 245
troch lear (patheti cus , VI cranialnerve) , 1 10 , 1 14 , 120 , 143 , 145 ,
186
tympanic (nerve of JACOBSON ) ,vagus (pneumogastric nerve, Xcrania l nerve) , 1 10 , 1 1 1 , 1 12 ,
—245vasoconstr ictor , 235vasod i lator
,235
vasomotor . See Vasomotor ap
paratus .vestibular
,88 , 1 10 , 1 1 1 , 145 , 147 ,
Nerve, vid ian ,245
V iscera l,126
,144
,145—150, 259
of WRISBE RG . See N erve, intermed iate .
Nerve—cell . See Neuron.
Nerve-fiber, a Slender fibrous process of a N euron, 39afferent
,108
carbon d ioxid production in, 96 , 97
conduction in, 96 , 97
degeneration O f,46
eff erent ; cf. also Efierent, 108e lectric changes in
,96
fatigue of, 96 , 101
medu l lated . SeeNerve-fiber, myelinated .
myelinated, a fiber prov ided w i tha Mye lin sheath , 97 , 108, 286postganglioni c . See Neuron, postgangl ioni c .pregangli on i c . See Neuron, pre
gang li oni c .rate O f transmission in, 97 , 98regeneration of, 46unmyelinated or unm edul lated , a
fiber devo id of a Myelin sheath ,108
N ervous impulse, nature of, 96 , 97ve loci ty of
,97 , 98
N ervous system , the aggregat e of al lnervous tissues .autonomic ; cf . Nervous system ,
sympathetic , 225 , 229centra l
,28, 106 , 107
cerebro-Spinal , 76 , 225development of
,106 , 1 15 , 1 16 ,
1 17 , 1 18 , 120, 123 , 1 53 , 18 1 ,
d iffuse 27 , 53 , 66 , 227 , 25 1
embryon i c, See Nervoustem ,
development of.evo lution of ; see a lso Cortex ,cerebral , evolution Of, and
Hemisphere, cerebra l , comparative anatomy and evolu
t ion of,22
,24
,27 , 33 , 34 , 1 13 ,
1 15 , 129 , 180 , 181 , 182 , 2 12 ,2 15 , 2 19 , 227 , 25 1 , 252, 253 ,
general anatomy O f,106
cerebro- spina l V iscera l , 227invertebrate , 27 , 53 ,
227
nomenclature O f, 1 15 , 12 1 , 122.

340 INDEX AND GLOSSARY
Nervous system , peripheral , 106phylogeny of. Sec N ervous system , evolu tion O f.
phys io logy of, 96segmenta l . See Segmentationand Segmental apparatus .subd ivision of, 106 , 1 1 5—123sympathet ic , 53 , 65 , 76 , 89 , 93 ,
periphera l autonomous part,synapt i c, 53vertebrate, 29 , 106
Neural canal . See Cana l , neura l .groove (medullary groove) , the
trough- l ike for m assum ed by theN eural plate during i ts in
vagination to form the Neuraltube.
plate , a th i ckened plate of E ctoderm in ear ly vertebrate em
bryos from wh i ch the Neuraltube develops .tube, the embryoni c central ner
vous system when in the formof an epi thel ial tube
,106 , 1 16 ,
Neurasthen ia, 103N euraxi s , the central nervous system ; and also applied to the
Axon.
Neuraxon . See Axon.
N eurenteric canal , in the embryo,a
commun ication between the
caudal end of the N eural tube andthe d igest ive tract .
Neuri lemma, the ou ter sheath of aperiphera l nervek fiber
, 40 , 46Neur i te . See Axon.
Neurob last, an immature nerve cell,
39 , 45
Neurocyte . Sec Neuron.
Neurofibrils , del icate protoplasmicfibri ls wi thin the cytoplasm of theNeuron, 40 , 46 , 47 , 102
Neuroglia (glia) , a supporting fabri cO f cells and horny fibers pervading the central nervous system , 38,
Neurogram, 29 5
N euromasts . See O rgans , lateral
Neurom ere, one of the segm ents ofthe embryoni c N eural tube.
Neuron (neurocyte) , a nerve cell ;of . Cell , 49
, 56 , 96 ,
afl'
erent,42
b ipo lar,44 , 4S
correlation,133 , 134 , 158
efferent, 42fatigue of, 96 , 1 01 , 102Of fir st
,second
, etc .,order, 42
mul ti form , See N euron, poly
morph ic .po larizat ion of. See Po lari ty ofthe N euron .
po lymorph i c,268 , 269 , 274
postgangl ioni c , an efferent sympatheti c neuron wh ich is ex
ci ted by a preganglioni c Neuron,
238, 239 . 244
pregang l ioni c , an efl'
erent sym pathet ic neuron whose cell bodylies in the central nervous system , 93 , 126 , 146
—148,150 , 229 ,
pyramida l, Of cerebral cortex , 42 ,
retraction of, 103type I , 43 , 44 , 1 90type I I , 43 , 44 , 1 90 , 268, 269unipo lar
,45
N eurone. See N euron.
Neuropil (mo lecular substance, d otted substance) , an entanglementO f unmye linated fibers containingmany synapses , 65
Neuropore, in the embryon i c brainan Opening between the anteriorend Of the neural Canal and the
exterior , 1 16Nicotin
,23 1
N id ulus . See NucleusN idu s , a depression on the ventralsurface of the cerebellum ; a lsoused as a synonym for Nuc leus
108
N 1SSL , F .,42
, 46 , 55 , 274
bod ies of, granules of, substance
of . See Sub stance, chromophi li c .
Noci ceptor, a sense organ or Receptor wh i ch responds to injuriousinfl uences .

342 INDEX AND GLOSSARY
Nucleus, mesencepha l ic , of V nerve, Nucleus , ventral is thalami , 1 63 , 164 ,motor ius tegmenti , 181of oculomotor nerve, 62 , 63 , 64 ,
246
o lfactorius anterior, the anteriorund ifferentiated port ion ofthe Area olfactori a , 220intermed ius . See Tubercu lumolfactorium .
latera l i s , the lateral portion ofthe Area olfactoria, lying between the o lfactory Bu lb and
the Uncus , 2 1 9m ed ia l i s, the med ial portion ofthe Area olfactoria , containing the Septum and Gyrussubcal losus, 2 19
O l ivary . See O l ive .
of origin,108 , 1 28
ponti le (pont i le nuclei , nuclei pont is) , 1 58, 187 , 188 , 289poster ior tha lami
,163 , 1 64 , 1 65
167
preopt i c (gangl ion op ti cum basale) , 220reti cularis thalami ( lat ti ce nucleus ,G itterschicht) , 306roof , of cerebel lum (nuclei fast igii , globosus , and embolifor
mis) , 1 88 , 19 1rubcr (red nucleus) , 158 , 16 1 , 166 ,
sal ivatory,146
,147 , 1 54 , 156 , 241 ,
244 , 247
of SCHWALBE,vestibular , 184 , 185
of S'
r i LL ING . See N ucleus,dorsal
,
of CLARKE .
terminal,108 , 1 12
O f trigeminus,ch ief sensory
,149
,
motor , 146 , 1 50 , 1 54 , 180 , 244
spinal (nuc leus O f spinal Vtract ; old term ,
gelatinous substance of RO LANDO O f med ullaob longata) , 140 , 1 54 , 1 55 , 156
of troch lear nerve , 146 , 150, 154 ,
of vagus , dorsal , 1 47 , 150 , 1 54 ,
of ventra l gray colu mn of sp inalcord
,1 29 , 1 30
285, 306
ventro- latera l , of spinal cord, acollection O f neurones in theventral gray co lumn whi ch innervate the musc les of theIhnbs
,129 , 130
ventro-m ed ial , of th e spinal cord,a col lection of neurones in theven tral gray co lum n wh i ch innervate the muscles of thetrunk .
vest ibular , 143 , 150 , 154 , 155 , 156 ,
N U E L , J . P .,2 14
Space Of, 1 9 7
Number of BETZ cel ls,284
of fibers in human pyramidal tract,
284
of neurones in cerebra l cortex,26
OBE RSTE INE R, H. , 269
Ob longata . See Medulla ob longata.
O lfactory apparatus . See a lso Rh inencephalon, 72 , 74 , 9 2 , 1 10 ,1 1 1 , 1 12 , 122 , 146 , 148 , 160 , 162 ,163 , 2 15
—223,273 , 279
O l ive, accessory, 1 55inferior (ol iva , nucleus O l ivarisO l ivary body) , a large gray cen
ter in the med u l la Ob longatawh i ch produces an eminence on
its latera l surface,1 14 , 13 1 , 155 ,
superior, a nucleus in the secondary aud itory path embed dedin the medulla Oblongata dorsa lly O f the pens, 60 , 1 64 , 185 ,20 1
peduncle of,20 1
ON UF‘
,B .
,233
Opercu lum , the lobules of the frontal
,parietal , and tempora l cere
bra l cortex wh ich cover the Insula,Ophtha lm encephalon, the retina , opti c nerve
, and v isua l apparatus ofthe brain .
Opossum,cerebral cor tex of
,2 17
O ptic apparatus . See Visua l apparatus .ch iasma . See Chiasma, opti c .

INDEX AND GLOSSARY 343
Optic tectum , an Op tic reflex centerin the roof of the midbrain . SeeCo ll i cu lus , superior.thalamus . See D iencephalon.
vesi c le. See Ves i c le, Opti c .O ral , pertaining to the mouth , ord irected toward the mouth , asopposed to Cau d al .sense of EDIN GE R
,2 19
O rgan (organon) , a part of the bodywi th a part i cular function
,24
of CO RTI . See O rgan , spira l .generative. See sexua l organs .lateral line (neuromasts) , senseorgans in or under the skin offishes and amph ib ians of interm ed iate type between tact ile and aud itory organs
, 1 10
1 12 , 145, 1 48, 149 , 152, 199 ,200
parietal . See Parieta l eye.
pinea l . See Bod y, pineal .spira l (organon sp ira le) , the organof CO RTI or receptor for sound inthe Coch lea , 85 , 1 97 , 198 , 199
O ss i cles , aud itory, 1 95 , 196O xydat ion in neurones
,9 6, 97 , 99
O xy gen as respiratory st im ulus , 238
PACIN IAN corpuscle, 79 , 80 , 88
Pa in,apparatus of ; cf . A ffection,
85 ,
163—167 , 172—174 , 1 78—180, 228230 , 243 , 249
—262, 286conduction paths for
,249
,25 1 ,
referred , 228 , 229 , 230 , 26 1thalamic center for . See Thalamus
,pa in center in .
Palaee'
ncepha lon, the o ld bra in ,72. c .
,
all of the brain except the cerebralcortex and i ts dependencies, 1 15 ,363
Pale othalam us (old tha lamus) , thephylogenet i cally old part of theThalam us , present in animalswh ich lack the cerebra l cortex
,163
166
Palate , 243Pa l l ium . See Cortex, cerebral , 2 16Pancreas
,224
Paralys is from central les ion, 173 ,
Paraphys is , an evagination of themembranous roof of the telencepha lon in front Of the Ve lumtransversum in some vertebratebrains .
Parieta l eye (par ieta l organ ,pineal
eye, epiphysea l eye) , a mod ificat ion of the pinea l Body in somelower vertebrates to form a dorsalmed ian eye, 162 , 2 12
PAR KE R,G . H .
,37 , 75 , 9 5 , 199 , 203 ,
PAR ME LE E, M .
,37
Pars intermed ia of WRISBE RG . See
Nerve, interm ed iate.
Pars mam illaris hypothalm i , the
bo d ies and theirenvirons , 1 18
Optica hypothalam i , the opti cChiasma and i ts environs , 1 18 ,
Pause, central , 98PAWLOW ,
I.,242 , 248
Pedagogy. See Education.
Ped unc le (peduncu lus ) , a peduncleor stalk . See Crus .cerebellar , one of the fibrous stalksby wh i ch the cerebe llum is attached to the bra in stem . Thereare three ped uncles on each s ide :( 1 ) the superior peduncle (Brach i um conjunctivum ) , (2 ) the
midd le pedunc le (Brachi um pon
tis) , (3 ) the inferior pedunc le(Corpus restiform e) , 158, 187 ,188
cerebra l (pedunculus cerebri ) , theventra l part of the mesencephalon
,1 18 , 1 19 , 120 , 12 1
,
O f corpus cal losum . See Gyrussubca l losus .of superior O live, 201
Perikaryon, the protoplasm surround ing the nucleus in the
Cel l body O f a Neuron .
functions O f, 99Per i lymph , 196Perineureum , the connect ive- t issuesheath surround ing a peripheralnerve
Per istals is,24 1
Peritoneum , 80 , 250
P0 8 ped uncu li . See Bas is peduncu li .

344
Pharynx, innervation of, 144, 147,243
PHIL I’
PPSON , M .,133 , 135 , 142
Photoreceptors , nervous End -organssens i t ive to l ight , 2 12
Phrenology, 280, 281 , 285Phylogeny of nervous system . See
Nervous system , evo lut ion of .Phys iognomy, 280Pia mater, the inner bra inmembrane.
P igment, retina l . See Retina , pigment of.
P IKE ,F . H.
, 194
P i llar of CO RTI , 1 97, 198of fornix . See Fornix column and
Fornix crus.
P i locarp in 23 1
P ineal body. Sec Body, pinea l.eye . See Parietal eye.
P ituitary body. See Hypophys is.
Plants contrasted w ith an imals , 22P lasticity in behavior . See Behavior,variab le.
Plate ( lamina) , a general term ap
plied to any flat structure orayer ; specifica l ly to the sixlongi tud inal bands or zones in towh ich the Neura l tube is d iv idedas exp lained in the fo llow ingdefini tions, 1 17 .
d orsal (roof plate, Deckplatte) , theunpaired dorsal longi tud ina lepi thel ial zone of the Neuraltube ; i t is non-nervous and insome parts of the adul t brain isenlarged to form a Te la , 153 .
d orso- lateral (alar plate , wingplate, epencephal i c region
,Flu
gelplatte) , one of a pai r ofdorso—lateral longitud ina l zonesof the Neural tube ; it gives riseto the dorsal gray column of thespinal cord and to the sensorye
gnters of the brain
, 1 17 , 1 20,1 3
floor. See P late, ventral .neura l . See Neural plate.
roof. See P late, dorsal .ventral (floor plate, Bodenplatte) ,the unpa ired ventral longitud inal zone of the N eura l tube ;It IS original ly non-nervous , bu tm the adul t is invaded by theventral Comm issure, 153
INDEX AND GLOSSARY
P late, ventro- lateral (basal plate,hypencephali c region
,Boden
platte) , one of a a ir of ventrolatera l longitud ina zones O f theNeural tube ; i t gives rise to the
ventral gray co lum n of the cordand to the motor centers of thebrain, 1 17 , 120, 153
P lay,257
P leasantness,P leasure.
tion .
P leura, 1 25 , 250P lexus , chorioi d (choroid plexus ,
plexus chorioi d eus) , a th innon-nervous portion of thebra in wall to wh i ch h igh lyvascular Piamater is adherentand wh ich is crumpled and
thrust into the brain ven
tricles .
lateral , the chorio id plexuses ofthe lateral ventri cles of thecerebral hemispheres, 222
of fourth ventri c le (plexus chorioid eus ventricul i quart i ) , thechorioid plexu s whi ch fom i s
the roof of the fowt h ven
triele, 1 19 , 121 , 152 , 264of third ventri cle (plexus chori
oid eus ventricul i tert ii ) , thechorio id plexus wh i ch form sthe roof of the th ird ven
triele, 162 , 1 66 , 167gangl ioni c , of sympathetic nervoussystem , an entang lement ofsympathet i c nerves and gangl ioncells ; most of the nervous lex
uses enumerated in the fo ow
ing l ist are gangl ioni c plexusesO f th is type, 225 , 226 , 24 1
nervous , an interlacing of d ifferent kinds of nerve-fibers, 53aorti c , 226of AU E RR ACH (myenteric plexus) ,24 1
brach ial,226
bronch ial,226 , 238
cardi ac , 226 , 235cel iac
,226
cerv ical,226
coronary,226
esophageal,226
gastri c,226
hypogas tr i c,226 , 239
See Affec

346 INDEX AND GLOSSARY
Qual e, a quali ty pertaining to anyth ing ; specifically a quality of sonsation or other conscious process,
Rabb i t,cortico—spinal tract of
,3 10
development of eye of, 204spina l cord of, 133Radiations , sensory, the thalam ocortica l tracts . See Tract
,
tha lamo-corti cal,
and Coronara d iata, 287 , 289aud i tory
,1 70, 287
gustatory,287
o lfactory, the o lfacto-corti caltracts ; the term has also beenapplied to various subcort ica lo lfactory tracts
,287
opti c,1 70, 2 10, 287
somesthet i c (of tac tile and generalsensation) , 287Rad ix. See Root.
Rage . See Anger .RAMON Y CAJAL , S.
,42
,44
,48, 50
270- 273 , 278Ram u s comm unicans, a commun icating branch between the gangl iaof the sympatheti c Trunk and the
roots of the spina l nerves,125 , 126 ,
Range of behavior,19
,303
RANV IE R , node of. See Node of
Ranvier.Rat
,nervous system of
,220
Rate of nervous conduction , 97 , 98Reaction, a change in bod i ly statein response to stim ulat ion ; cf.Reflex, 66avo id ing . See Reflex
,avo id ing .
d iscr im inative,98
, 302, 308
tim e, the time required for re
sponse to st imulation,98
,258
,
259
Read ing , apparatus of . See Speech,
apparatus of.Receptor , a sense organ ,
25, 38 , 69
contact, a sense organ ad ap ted torespond to impress ions from oh
jects in contac t w i th the body ;opposed to d istance Receptor.
d istance, a sense organ ad apte d torespond to impress ions from oh
j ects remote from the body,23
Recess,epitympani c, 1 95
infund ibular,1 18 , 1 19
lateral , the w idest part of thefourth Ventric le under the corebell um .
opti c , the depress ion in the latera lwall of the d iencephalon formedby the evagination of the opticVes ic le, 1 16—1 19u tricular. See U tricle .
Reflex act, a simple form of invat iab le Behavior requir ing anervous system ,
25 , 32, 56 ,109 , 363
t ime of . See Reaction tim e.
a l l ied,57 , 58 , 59 , 6 1
antagon ist i c, 57 , 58 , 59 , 6 1
are. See Reflex circuit.avoid ing, 25 1 , 253 , 258of brain stem
,181 , 192, 279 , 280,
304 , 305
bu lbar,1 81
,279
cha in,57 , 58 , 60 , 6 1 , 308
circuit, a cha in of neurons wh i chfunct ion in a Reflex act, 25 , 42 ,56 , 58 , 60 , 62 , 63 , 66 , 109 , 1 13 ,132 , 133 , 134 , 260 , 308 , 309 ,3 1 1
cond i tional,242
corti cal,286
,290
cyclic,6 1 , 309
d iscrim inat ive . See Reaction,d is
crim inat ive .
of feed ing ; cf . O ral sense, 2 19 , 279locomotor
,134
of med ulla ob longata,181 , 279
myenteri c , 241pattern, 65 ,
2 19,305 , 307 , 3 12
propriocept ive,130, 175
- 193
of spina l cord,1 29 , 13 1
—135 , 174 ,
304 , 305
thalamic, 163 , 166 , 174 , 18 1 , 253 ,
Regenerat ion of nervous tissues, 46 ,132
Region, corti ca l , a group of relatedcort ica l Areas ,Regu lation,
the process of adaptat ion of form or behav ior of an or
ganism to changed cond it ions, 3 1RE IL , is land of . See Insula.Re inforcement
,59
,62 , 63 , 101 , 192,
2 18

INDEX AND GLOSSARY 347
RE ISSN E R , membrane of . See Mem
brane, vest ibular .Rept iles , cerebra l cortex of, 2 16Resistance,nervous
,104 , 252 , 258,
295 , 296 , 304, 307Reso lution,physio logica l
, 58 , 293,304
, 306 , 307Respiratory apparatus, 89 , 144 , 147 ,232 , 234
—210Restiforrn body. See Corpus restiform e.
Ret icular form at ion. See Formatioreti culari s .
pigment of, 205 , 207 , 208 , 2 1 1Retract ion of the neuron,103 , 104RETZ IUS, G .
, 85 , 88 ,1 24
,19 7
,20
2 19
Reverberat ion,cortical
,293 , 296 endo
Rhinencephalon (nose bra in ) , the
olfactory part of the bra in ,l l l
Rhodops in,207
Rhombencephalon, that part of thebrain below the Isthm us , including the Medulla ob longata and
Cerebellum , 1 16—1 1 9 , 12 1 , 122 ,
development of, 1 16—1 19RIVE RS, W . H. R ., 94 , 95 , 142
Rod of Com a (pillar of Conn ) , 197 ,1 98
of ret ina, 205 , 206 , 207 , 208 , 2 1 1RO LANDO,
fissure of. See Sulcuscentra l is .gelatinous substance of . See
Substantia ge latinosa Rolan d i .Root (rad ix) , a nerve root,or the
part of a nerve adjacent to thecenter w i th wh i ch i t is con
nected ; in the case of spina l andcranial nerves , the part lyingbetween the cells of or igin ortermination and the gangl ion .
anterior . See Root, ventral .dorsal (rad ix dorsal is , posteriorroot , rad ix poster ior) , the dorsa lor sensory Root of a spina l orcrania l nerve , 126 , 128 , 129 ,
poster ior . See Root, dorsal .spina l , composi tion of , 1 26 , 135 ,
Root, ventral (rad ix ventra lis , rad ixanterior) , the ventra l or motor rootof a sp ina l or cranial nerve, 126 ,128 , 129 , 130, 133 , 134 , 150, 1 5 1 ,182 , 227Rostral , perta ining to the beak orsnout
,or directed toward the
front end of the body as opposedto Caudal .Rostrum of corpus ca l losum
,1 1 9
RUSSE LL , J . S. R i ESE N,194
Sac , dorsal (saccus dorsal is) , a dorsalevagination of the Te la chorioi d ea of the th ird ventricle insome vertebrate brains .
endo lymphat i c (saccuslymphat icus) , 196
nasal , 1 10, 1 1 1Saccule (sacculus) , part of the mem
branous labyrinth of the car, 85 ,183 , 1 95 , 1 96 , 199 , 200 , 201
SACHS,E
,17 1
SALA,C . L .
, 80
Sa liva,secretion of . See a ls oG land ,
sa l ivary 146 , 147 , 24 1 , 242 , 247Sarcophaga carnaria
,nervous system
of,30
Sca la med ia . See Ductus coch learis ,
ty mpani , 197vestibuli , 1 97
SCAR PA, ganglion of . See Gangl ion,vestibular .
SCHAE FE R,E . A .
,2 14
SCRAPE R , A ., 194
ScorrNE MA NN,A .
,203
SCHU LTZE , tract of (comm a tract) .See Fasciculus interfasc icularis .
SCHWALBE , vestibular nucleus of,184 , 185
SCHWAN N , sheath of . See Neurilemma.
Scyll ium ,nervous system of , 1 10
Sea- rob in, nervous system of, 1 5 1 ,
153
Secretin,224
Secret ions , effect of fat igue and em o
tion on,103 ,
255
interna l,163 , 224 , 23 1 , 255 , 256
psych ic , 24 1 , 242Segment. mesodermal , or prim itive.
Sec Somi tes.

348
Segm ental apparatus , the Brainstem , 1 13 , 1 14 , 123
Segmentation of nervous system ,28 ,
Se l f- consciousness, 314Senil i ty, 3 16Serni circular cana ls , nerve end ings In,
SEMO N , R . 295
Sensation, a subjective experiencear is ing in response to stimula26 1
common,259
in lower an im als, 72
neurological mechan ism of,250,
257 , 26 1
v isceral , 77 , 9 1 , 148, 228, 234—246 ,250
,259
Sense,cri ter ia of
, 74
organ . See Receptor.Sentiments . See Affect ion .
Septum , the me d ial wal l of the cerebra l hemisphere between the
Lam ina term inali s and the ol
factory Bu lb ; in man its upperpart is th in and forms the Septum pe l lucidum , 2 19 , 220, 306
dorsa l med ian,of cord . SeeFissure,
dorsal .pel lucidum , a th in sheet of nervoustissue forming a portion of themed ial wal l of each cerebralhemisphere between the Corpuscal losum and the -Fornix, 1 62
Sexual organs, innervation of, 232sensat ions from
,89
SHAMBAU GH,G . E .
,198,
Shark , nervous system of . SeeFishes
,nervous system of .
Sheath med ullary . See Myelinsheath .
myel in . See Myelin sheath .primitive . See Neuri lemma.
of SCIrWANN . See Neurilemma.
SHE LDO N,R . S
, 95 , 248SHE RRE N , J .
, 94 , 142SHE RRINGTO N , C . S , 35 . 37 , 65 , 68 ,
194,243
,250
, 259 , 260 , 262 , 28 1 ,
Sight organs of . Sec Visual apparaSinus, infer ior , of labyrinth , 1 96
INDEX AN D GL OSSARY
Skin bra in,1 12, 123
nerves of See Nerves , cutaneous .nerve-end ings in
, 79 , 80, 81—83 , 84,
sens ib i lity of, 70, 72 , 79 , 80—86 ,132 , 172—180, 2 12 ,245
,249 , 250 , 252 , 253 , 260
Sleep , 103, 1 04 , 297Smell , organs of . See O lfactory apparatus .
SMITH, G . E LLIOT, 263 , 273 , 278
Sneeze,mechanism of
,238
Socia l evolution, 3 14 , 3 15
Somat i c area . See Area, somati c .cortex . See Neopal lium .
nerves . See N erve,somat ic .
organs , those concerned w ith theadjustment of the body to i tsenv ironment
, 76 , 79 , 92, 109 ,172
Som esthetic apparatus , the generalsomati c sensory systems, including cutaneous and deep sens ibil
ity,164 , 165 , 172
- 180
Som ites (myotom s,pr im i tive seg
ments, mesodermal segm ents ) ,
segm eiIted masses of mesoderm In
vertebrate embryos wh i ch giverise to the somatic muscles , 92
Sound , reaction time to , 98recepto rs for . See Aud i tory apparatus .
Space, d iscr iminat ion of, 130, 132,137 , 172 , 178
—180of N UE L, 1 97perforated . See Substantia per
forata.
Speech , apparatus of ( includ ing readm g and wri ting) , cf . Aphas ia, 283 ,29 1—293
SPE NCE R,HE RBE RT ,
17
Sphere,cort ical . See Center, corti
ca l .Spiders
,nervous system of
,29
SPIE LME YE R,W.
,284
Spinal cord (medulla spinal is) , thatportion of the central nervoussystem contained wi th in the
spinal Canal of the spinalcolumn
,106 , 1 07 , 1 10—1 12,
1 17 , 1 18 , 1 20, 122 , 125 , 126 ,127 , 1 28—130, 150, 15 1 , 182
cervi cal,128
,1 29 , 130
deve lopment of, 182

350
Substantia a lba . See Matter, whi te.
ge latinosa Rolan d i (gela tinous substance of RO LAN DO ) , an area of Summat ion , central .N europi l bordering the dorsa lgray co lumn of the Spina l cord ;somet im es a lso applIed to the
nucleus of the Spina l V tract inthe medul la ob longata, 1 29
grisea . See Matter, gray.
nigra (b lack substance) , an area ofgray matter immed iate ly dorsa l of the Basi s ped unculi ,funct ionally related to the
cortico—pon ti le tracts, 16 1 , 1 65 ,167 , 210
perforata, anterior (anter ior perforated substance or space) ,a region on the ventral surface of the brain in front ofthe opti c Chiasma wh ich ispierced by many small arteries
, 120 , 2 19 , 306
posterior (posterior perforatedsubstance or space) , a regionon the ventral surface of thebrain between the Bases ped unculi wh i ch is pierced bysmall arter ies, 120
Subthalam us , the ventral part ofthe Thalam us , 163 , 165 , 1 66 , 167 ,1 74 , 176 , 306
Sulcus , in the cerebra l cortex,a
superficia l fo ld not invo lv ingthe ent ire th ickness of the brainwall ; cf . Fissure, 266anter ior parolfactory
,1 19
centra l (fissure of RO LANDO ,cru
ciate sulcus) , 1 19 , 12 1 , 281 , 282cinguli , 1 19corporis callosi
,1 19
cruciate. See Sulcus,centra l .
frontalis, inferior, 1 2 1super ior
,1 2 1
interpar ietali s, 12 1
lim iting (sulcus lim itans) , a longitud inal groove on the ventricular
. surface of the embryoni cbra in separating the dorsolateral sensory P late from theventro—latera l motor P late, 36 ,occipi tal is transversus
,12 1
posterior paro l factory,1 19 , 2 19
precentralis,121
INDEX AND GL OSSARY
Sul cus rhinalis . See Fovea limbica.
sp ira li s,1 97, 198 , 199
See Cond uc
tion, avalanche, and Reinforcem ent.
of stim uli , the enhancement ofeffect by repeated stimulat ion ,
59 , 62 , 63, 192, 208, 2 18, 258,260, 268, 307
Suprasegm enta l apparatus , the cerebra l cortex and cerebellum w i ththeir immed iate dependencies
,
Suscep tib ili ty of neurones to poisons,97 , 231
Swal lowing, apparatus of, 78, 247SYLVIU S, aqued uc t of See Aqueductof Sylvi us.
fissure of . See Fi ssure, lateral .fossa of . See Fossa lateral is .
Symbo li z ing, defects of, 292Sympathet ic nervous system . SeeN ervous system
,sympatheti c .
Synapse, the place where the nervousim pulse is transmi tted from one
neuron to another,50, 5 1 , 52 ,
53 , 54 , 96 , 97 , 103 , 109 , 190,
fatigue of,102
,103
t im e of transm ission through,54 ,
98 , 99
Synergi c m uscles .synergic .
System , functional , all neurons ofcomm on phys io logica l type. Mostperiphera l nerves conta in severa lcomponents belonging to d ifferentsystems
,145—150
See Muscles,
Tabanus bovinus,nervous system of
Taenia, the l ine of attachment of amembranous part to a mass ivepart of the bra in wal l ; formerlyapplied a lso to some fibertracts
,as Ta3nia semic ircularis
Stri a terminali s , and Ta niathalami Stria medul laristha lami .
chorio idea , the line of attachm entof the lateral chorioid P lexus tothe med ia l wall of the cerebra l

INDEx AND GLOSSARY 351
hem isphere . (This port ion of themedi a l wall is adherent to the thalamus
,forming the Lam ina affixa.)
Taenia forni ci s , the line of attachment of the latera l ch orio idP lexus to the Fimbria of theFornix.
thalam i , the l ine of attachment ofthe Te la chorio idea of the thirdventric le to the dorsal marg inof the thalam us . Thi s nam e wasformerly applied to a band offibers , the Str ia m edullari sthalam i , wh ich borders th
ta-nia . 162 .
ventriculi quarti , the line of at
tachm ent of the membranousroof of the four th ventricle tothe medulla ob longata, 1 55 .
TASHIRO ,S.
,97 , 105
Taste, apparatus of . See Gustatoryapparatus .bud
,9 1 , 143 , 144 ,
2 18, 243 , 245 ,246
periphera l nerves of . See Nerves,gus tatory.
Tax is . See Tropi sm .
Tectum mesencephal i ,'
the roof ofthe midbra in
,compr is ing the
Co lli culus superior ( tectum opticum) and the Colli cu lus inferior , 1 6 1 , 246optic . See Colli cu lus , superior.
Teeth,85 , 146 , 249
Tegm en ventriculi quarti , the roof ofthe fourth ven tr icle ,
formed ch ieflyby the Ve lum medullare anterius ,the Ve lum m edu l lare posterius ,and the P lexus chorioi d eus ven
tri culi quarti .Tegm entum , the dorsa l part of thecerebral Pedunc le between the
Bas is pedunculi and the Aqueductof Sylvius ; often described as a lsoextend ing backward into the correspond ing part of the medul laob longata , 158, 181 , 1 82
Te la , any th in non- nervous part ofthe brain wal l .chorio idea , that port ion of thePia mater whi ch covers anythin non-nervous part of thebra in wall , includ ing the chorioi d P lexuses.
Telencephalon (endbrain) , the anterior end of the embryoni cNeural tube and its ad ult d erivatives, compris ing ch iefly thecerebral hemispheres and Lamina term inali s, 1 17—1 19 , 12 1
m ed ium , that portion of the em
bryonie Te lencephal on whi ch isnot evaginated to form the cerebral hemi spheres ; i t compriseschi efly the Lam ina termina li sand Pars Optica hypothalam i ,122
Te lodend ron, the terminal branchedend of a Dendrite ; somet im es app l ied also to that of an Axon ; cf .Terminal arborization.
Temperature, apparatus of, 7 1 , 84 ,13 1 , 132 , 137 , 138, 139 , 14 1 , 163
167, 172—174 , 242, 254, 260
Tendon,nerve end ings in
,87
sense, 77, 87 , 132Tentorium cerebell i , a transversefo ld of Dura mater between the
cerebellum and the cerebral hemispheres .
Terminal arborization, the bran chedend of an axon ; somet im es applied al so to that of a Dend rite,40
Terminology. See Nervous system ,
terminology of.Testes . See Co ll i culus, inferi or.Thalam encepha lon . See Dienceph
on.
Thalamus, the midd le and largersubd iv is ion of the Diencephalon, somet imes applied to theentire d iencepha lon and cal ledThalam us opti cus , 63 , 1 12 , 1 14 ,1 1 7 , 121—123
,14 1 , 1 62 , 163 ,
164—166 , 167 , 174 , 176 , 204,2 10, 279 , 31 1
les ions of, 253 , 254 ,279
new . See Neothalam us .o ld . See Palaeothalam us.
opticus . See Thalamus.
pain center in,167 , 253 , 254 ,
258,260
,26 1 , 3 1 1
respiratory center in,240
Th irst,apparatus of
,89
THO MPSON ,T .
,142
,175 , 1 79 , 262
Thorns of dendrites , 272

352 INDEx AND GLOSSARY
Threshold , the m in im a l st imuluswh ich w ill exci te an organ to ao
tivity, 72 , 80, 9 1 , 132, 172 , 2 18,252
,254 , 255
Tickle, 76 , 254
Tigroid bod ies,substance, or gran
ules . See Sub stance, ch rom o
Time,central . See Pause, central .
latent . See Pause, central .reaction . See Reaction tim e.
Tissue, the cellular fabri c of whi chthe body is composed , 24
TITCIIE NE R,E . B .
, 70
Tone, affect ive . See Feeling toneand A ffect ion .
analys is , 198, 199 , 202fee ling . See Feeling tone andAffection.
muscular, 77 , 89 , 189 , 192nervous
,101 , 192
Tongue, muscles of, 92 , 144 , 147 , 148nerves of
, 143, 144 , 1 46- 148, 243 ,
245
Touch,apparatus of
,62 , 70, 79 , 80
163
react ion time of, 98Toxines . See Poisons .TO ZE R . F . M .
, 180
Tract ( tractus) , a collection of nervefibers of like origin ,
termination,
and function ; cf . Fasc icu lu s , 27 ,association ; cf. Fibers , associat ion,
bulbo—spinal,1 57
central tegmental , 188cerebello- tegm ental , 188comma . See Fasc iculus interfascicularis .
corti co—bu lbar , 16 1 , 169 , 170 , 181cortico—cerebellar . See Tract
,cor
t ico-pont i le.
cortieo-mesencephal i c , 289corti co-oculomotor, 170cortico—pontile
,16 1 , 169 , 1 70, 187,
188 , 289
cort i co-rubral,170, 289
cortico- spinal (fasc iculus cerebrospinalis , B . N . A . , pyram idaltract) , the voluntary motorpath from the precentral gy
rus of the cerebral cortex to thespinal cord , where i t d iv ides intola tera l and ventral parts, 128, 130,131 , 140, 14 1 , 156 , 16 1 , 1 67 , 168,170, 181 , 188 , 1 92 , 282
—285 , 289 ,
Tract, corti co- spina l , lateral (fasciculus cerebro-spinalis lateral is , B . N . A ., lateral orcrossed pyram ida l tract) , 130,
ventral (fasciculus cerebrospina lis anter ior, B . N . A .,
ventra l or d irect pyramidaltract, column of Ti
’
JR CK) ,130, 13 1 , 14 1
corti co—thalam i c,289
d irect . cerebel lar . See Tract,
spino- cerebellar, dorsal .of FLE CHSIG . See Tract, spinocerebellar, dorsal .of GOWE RS. See Fasciculus ven
tro- latera lis superficialis.
habenulo-peduncu lar (fasciculusretroflexus, ME YN E RT
’
S bundle) ,165 , 220
of HE LW IG . See Tract, Olivo
spina l .intem uncial , a fiber tract connecting two nuclei or centers , 65of L ISSAU E R . See Fasciculus dorse- lateralis .of LOWE NTHAL . See Tract, tectospina l .
m am illo—ped uncular, 1 6 1 , 220mam illo-thalam i c (fasciculustha lamo-m am i l lar is B . N . A .
,
tract of V ICQ D’
AZYR) , 165 , 220mesencephal i c
,of V nerve, 16 1
of ME YNE RT. See Tract, habenulo-peduncular.
of MO NAKOW . See Tract, rubrospinal .
nomenclature of, 128olfactory (tractus o lfactorius ) , ol
factory fibers oi the secondorder passing from the olfac
tory Bu lb to the nuclei of theArea olfactoria. See Stria ol
factoria, 1 14 , 120, 165 , 2 17 , 2 18,
o lfacto-hypotha lam i c , 220olfacto-tegmenta l , 220, 221olive-cerebellar
,155 , 176 , 188

354
Tuberc le, anterior, of thalamus, aneminence on the dorsa l surfaceformed by the N ucleus anteriorthalami , 1 62
Tuberculum acusticum of fishes(part of the Area acusti colaterali s ) , 148, 149 , 1 52 , 200of mammals (the dorsa l coch learnucleus) , 20 1
cinereum , an eminence on the
lateral aspect of the medullaob longata produced ch iefly bythe spina l V tract and its nu
cleus .
cuneatum , an em inence on the
dorsa l surface of the lower endof the medulla ob longata laterally of the C lava produced bythe nucleus of the Fasc icul uscun eatus, 130, 139 , 14 1 , 156 ,
fascia: d entatze , 220olfactorium ( lobus paro lfactor iusof E DINGE R) , the intel m ed iate
o lfactory Nuc leus , lying in the
Substantia perforata anterior ; cf .Area olfactoria , 2 1 9 , 220, 273
Tunne l of Com a,19 7
TURCK , co lumn of, the ventra l cortico—spinal Trac t .
TU RNE R , W . A .
,194
Tympani c membrane, Ty mpanum .
See Membrane,tympanic .
Unconscious cerebration,297 , 309 ,
3 1 1
mind , 296 , 297 , 309Unconsciousness
,297
Uncus , the hook- shaped extremity ofthe Gyrus h ippocampi , part of theArchipa ll i um , 2 19 , 273
Unpleasantness . See Affection .
U tri c le (utriculus , recessus u triculi ) ,part of the membranous labyr inthof the inner ear, 85 , 183, 1 95 , 196 ,200 , 201
Valve of Vi E USSE Ns . See Velummedullare anterius.
Va lvula cerebell i . SeeVelum m ed ullare anterius .Var iab le behavior.variab le.
See Behavior,
INDEX AND GLOSSARY
Variation,negative, in nerve-fibers,
VARO LI (VARO LIUS) . See PonsVarolii .Vas spirale, 1 97Vasomotor apparatus, the neuromuscular mechan ism wh i ch con
tro ls the amount of b lood supplied to any part, 104, 1 14, 232 ,234
,235
Veins, nerves of . See Vasom otor apparatus.
Ve locity of nervous conduction .
See N ervous impulse , veloci ty of.Velum ant i cum . See Ve lum m ed ullare anterius.
interpos itum , the Tela chorioi d eaof the th ird ventricle.
m edul lare anterius , a thin portionof the brain wall containing afew myelinated fibers wh i chforms the roof of the four thventri cle in front of the cerebellum ,
posterius , a th in portion of thebrain wall conta in ing a fewmyelinated fibers wh ich formsa sma ll par t of the roof of thefourth ventr i cle immed i atelybeh ind the cerebell um .
superius . See Ve lum m ed ullareanteriu s .
transversum , a transverse fo ld oftheTe la chorio ideawh i ch marksthe boundary between the Di
encephalon and the Telencephal on in the embryon i c bra in .
Ventral , on the front or belly s ide ofthe body, terme d Anterior in the
B . N . A . lists, 1 15Ventri c le, a cav i ty Wi th in the bra in
and spinal cord derived fromthe lumen of the embryoni cNeural tube.
fi fth . See Cavum septi pellucidi .fi rst. See Ventri cle, lateral .fourth (metacm le) , the ventricle ofthe medulla ob longata, 1 19 , 12 1 ,152 , 264
lateral (paraccele) , the ventri cleof each cerebra l hemisphere ;these are a lso called first andsecond ventricles, 121 , 170, 2 10,

INDEX AND GLOSSARY
Ventricle, second . See Ventric le,lateral .third (d iacmle) , the ventr icle ofthe di encepha lon ,
121 , 1 62 , 1 70,264 , 265
VE RATTI, E ., 50
Vermi s cerebe ll i (worm) , the midd lelobe of the cerebel lum ,
1 19 , 187 ,19 1 , 201
Vertebrates, behavior O f, 33nervous sy stem of, 29
VE RWO RN ,M .
,37 , 101
Vesi c le, opti c , an outgrowth fromthe latera l wa l l of the d iencephalon wh ich forms the nervous partof the eyebal l . It first assumesthe form of a simple hollow sphere,the primary opt ic ves icle, wh ichlater co l lapses to form a two- layered opt ic cup , or secondary opti cves ic le, 1 1 6 , 1 17, 204Vest ibular apparatus
,88 , 89 , 1 10 ,
1 1 1 , 1 3 1 , 145 , 147 , 1 50 ,183—193 ,
Vest iges , memory, in cortex , 295297 , 304, 300
—308Vibrations. tab le of, 72Vibrissm,
innervation of, 80Vicar ious function in cortex , 294VICQ D
’
AZ YR , tract of. See Tract,m amillo—thalam i c .
Vl NCE NT, STE LLA B .
, 80 , 95 , 2 14
Vi scera, the internal organs , espec ially those concerned w i th the
interna l adjus tments of the bod y,76 , 89 , 93 , 109 , 144 , 234—243 , 250Viscera l apparatus , 224—247 , 250 , 259nerves . See Nerve, visceral .brain , 1 12 , 123
Vis ion,stereoscopic , 209 , 2 10
355
Visual apparatus , 62 , 63 , 7 1 , 86 , 1 101 12 , 123 , 145 , 140, 1 50, 160 , 163
1 67 , 1 70 , 180 , 204—2 13
,268 , 279
VO GT , O .,273 , 278 , 287
Vo luntary movement, apparatus of,78 , 100, 1 81 , 192 , 222 , 240 , 24 1 ,279 , 285 , 286
Vom i t ing,mechanism of, 239 , 243
WALDE YE R, W. , 49 , 55
Warm th,sensations of .
perature, apparatus of.WASHBUR N
, A . L .,248
WASHBU RN, NIARGARET F. , 37
WATSON,J . B .
, 37 , 95 , 203 , 2 14, 262
WE IGE RT, method of, 129 , 274WILLE MS, E .
,180
WILLIS, circle of .
See Tem
See Circle of
YE RKES, R . M ., 33 , 37 , 63 , 68
Zone, cortica l . See Center, cortical .of L ISSAUE R . See Fasciculusdorso- lateral is .of neura l tube. See Plate.
ZWAA RDE MAKE R , H. , 223
WILSO N , J . G ., 85 , 95 , 1 94 ,243
,248
Wooowon'
rn,R . S.
, 98, 105 , 2 13
Word-b lindness (A lexia) , 292Word-d eafness , 292Worm . See Vermi s cerebell i .Worm s
,nervous system of, 27 , 28 ,
227
WRISBE RG , nerve of . See N erve,ih
termed iate .
Wri ting, apparatus of. See Speech ,apparatus of .WUNDT, W. , 98


2 Saunders ' Col lege Text-Books
McFarlandp
s BiologyBiology: General an d Med ical . By J O SEPH MCFARLAND , M. D. ,
Pro fessor of Pathology and Bacteriology , Med ico-Chirurgical College. xzmo of 457 pages , i llustrated . C loth , net.
N ew (2 d ) E d ition.
This work is particularly adaptab le to the requirements of scientificcourses . There are chapters on the origin of l ife and i ts manifestations
, the cell and cell division, reproduction ,ontogenesis
,conformity
to type, d ivergence, structural and b lood relationsh ip , parasi tism , mu
tilation and regeneration ,grafting , senescence , etc .
Prof. W. R . Mcc onne l l, Penn sylvania State Co l lege : It has someadmirab le features, the most valuable of wh ich is the careful résumé ofthe subjects of heredi ty and evo lution.
”
Drew?s invertebrate Zoology
Invertebrate Zoology. By GILMAN A . DREW, Ph . D. , Assistant Director of the Marine B iological Laboratory , Wood s Hole. Mass .
x2mo of 2 1 3 pages . Cloth , net. N ew ( 2 d ) Ed ition.
Professor Drew’s work gives the student a working knowledge of comparative anatomyand leads h im to an appreciation of the adap tationof the anima ls to their environments. I t is a practi cal work , expressing the practical know ledge ga ined through experience. The typemethod of study has been fo l lowed .
Prof. J ohn M. Ty ler, Amherst Col lege : I t covers the ground wel l,
is clear and very compact . The tab le of definitions is excel lent .
Daugherty”s Econom icZoologyE conomic Zoology. By L . S. DAUGHERTY , M. S Ph . D Professorof Sc ience. Missouri Wesleyan Co l lege ; and M. C. DAUGHERTY.Part I—Piett and Laboratory Gui d e: rzmo of 23 7 pages , interleaved . C loth , Sr. 25 net . Part II—P rinciples: rzmo of 406
pages , il lustrated . Cloth , net.
Not only does thi s work give the sal ient facts of structural zoology andthe development of the various branches
,but also the natural h istory
—the li fe and habits. It emphas izes the economic phase throughout.Pro f. V. E. She lford , University of Ch icago : I t has many meri tsand is the best book of the kind on the market.

Saund ers' Col lege Text-Books 3
Stiles”Nutritional PhysiologyNutritional Phys iology. By PERCY G . STILES , Instructor in Physiology at Harvard University . rzmo of 2 7 1 pages, il lustrated .
C loth ,net .
Dr. Sti les ’ new work takes up each organ ,each secretion concerned in
the process of digestion,discussing the part each p lays in the physiol
ogy of nutrition—in the transformation of energy. In fact , the keynote oi the book throughout is energy - its source and its conservation . The illustrations and homely s imi les are noteworthy.
Pro f. M. E . J affa, University of Ca l ifornia : The presentation of thematter is excel lent and can be understoo d by all .”
Stiles9
Nervous SysternThe Nervous System an i Its Conservation. By PERCY GOLDTHWAITSTILES , Instructor in Physiology at Harvard University. rzmo
of 230 pages , il lustrated . C loth ,net .
Prof . Stiles ’ wonderful facul ty of putting scientific things in languagewith in the grasp of the non-medi cal reader is nowhere better i llustratedthan in this , h is newest book . He has a way of conveying facts accurately with rifle-bal l precision. Thi s new book is rea l ly a physio logyand anatomy of the nervous system ,
emphasiz ing the means of conserving nervous energy.
NeurologyIntrod uction to Neurology. By C J UDSON HERRICK, Ph . D. , Pro
fessor of Neurology in the University of Chicago. rzrno of 360
pages , w ith 1 37 i l lus trations . J ust Out.
Th is work wi l l help the student to organize h is know ledge and toappreciate the s ignificance of the nervous system as a working mechanism . It presents the actua l inner workings of the nervous mechanismsin terms that he can understand at the very beginning of h is course inpsychology, genera l zoo logy, comparative anatomy ,
and genera l medicine. Prof. Herri ck ’s many years of teaching experience has impressedupon h im the d ifficu l ties the student finds in grasping th is subject atthe beginning of h is course. He has aimed to remove these diffi cu l ties.
The text is wel l i llustrated.

4 Saunde rs Co l lege Text-Books
Had ley on the HorseThe Horse in Health on$ Disease. By FREDE RICK B. HADLEY ,D. V. M. , Assoc iate Professor of Veterinary Sc ience, Univers ityof W iscons in . xzmo of 2 60 pages, illustrated . C loth , net .
J ust Out.
This new work correlates the structure and function of each organ c fthe body, and shows how the h idden parts are related to the form ,
movements , and uti l ity of the animal . Then,in another part
,you get
a concise di scussion of the causes , methods of prevent ion ,and efi
'
ects
of d isease. The book is designed especia l ly as an introd uctory text tothe study of veterinary science in agricultural schoo ls and col leges .
Kaupo’s Poultry CulturePoultry Culture, Sanitation , and Hygiene. By B . F. KAUPP , M. S. ,
D. V. M Poultry Investigator and Patho logist, North Caro linaExperiment Station. rzmo of 2 50 pages, with 2 0 0 il lustrations .
J ust Out.
Th is work gives you the breeds and varieties of poultry, hygiene and
sanitation ,venti lation ,
poultry—house construction , equipment , riddingstock of vermin
,internal parasites , and other d iseases . You get the
gross anatomy and functions of the d igestive organs , food- stufi'
s , compounding rations, fattening , dressing , packing , sel ling, care of eggs ,handling feathers , value of droppings as ferti lizer, caponiz ing , etc .
, etc .
Lynch ’s Diseases of Sw ineDiseases of Swine. W ith Particular Reference to Hog-Cholera.
By CHARLES F . LYNCH, M. D., D. V. S. , Terre Haute Veterinary
Co llege. W ith a chapter on Castration and Spaying, by GEO RGER . WHITE , M. D D. V. S. , Tennessee. Octavo of 74 1 pages ,
i llustrated . C loth ,net .
You get first some 80 pages on the various breeds of hogs , with valu
able points in j ud ging swine. Then comes an extremely importantmonograph of over 40 0 pages on hog
-cholera, giving the h istory, causes,patho logy
,types
,and treatment . Then
,in add i tion, you get complete
chapters on al l other d iseases of swine.

6 Saunde rs ' Co l lege Text-Bo oks
Pyle 9s Personal HygieneP ersona l Hyg iene. E d ite d by WA LTER L . P \ L E
,M. D.
, Fel lowof the Am en can Aca d emy of Med ic ine. xzm o of 5 1 5 pages , i l lustrated . C loth , net . New (61h) Ed ition.
Dr. Pyle’s work sets forth the best means of preventing d isease- thebestmeans to perfect heal th . I t tel ls you how to care for the teeth , skin,comp lexion
,and ha ir. I t takes up mouth breathing , catch ing cold ,
care of the vocal cords , care of the eyes, school hygiene, body posture ,venti lation,
house- c leaning, etc . There are chapters on food adulteration (byDr. HarveyW Wiley) , domestic hygiene, and home gymnastics.Canad ian Tea che r Such a complete and authori tative treatiseshould be in the hands of every teacher .
Qalbraith ’s Exerc ise for Wom en
Per sona l Hyg iene a nd P /tysz'
ca l Tra in ing f or Wom en ByANNA M . G au nt /ur n
,M D . ,
Fe l low N ew York Acad emy of
Me d ic ine . 1 2 m o of 37 1 pages , i l lustrate d . C loth , 32 0 0 net.
Dr. Galbraith ’s book meets a need long existing—a need for a simp lemanual of personal hygiene and physical training for women along scientific l ines. There are chapters on hair , hands and feet , dress , development of the form ,
and the attainment of good carriage by dancing,walking, running , swimming, rowing, etc.
Dr. Harry B. Bo ice , Trenton State N ormal School : It is intenselyinteresting and is the finest work of the kind of wh ich I know.
”
McKenz ie on ExerciseE x er c ise in E d uca tion a nd Aled z
’
c ine. By R . Tan McKnm z rn,
M . D .,Professor of Physical E d ucation , U nivers ity of Pennsyl
van ia. Octavo of 585 pages , w ith 478 il lustrations. C loth ,
New (2 d ) E d ition.
Chapters of special value in college work are those on exercise by thed ifferent systems : play-
grounds, physical education in school,col lege,
and un iversity.
D. A ; Sargent, M. D Hemenway Gymnasium : I t should be in the
hands of every physical educator.

Saunders' Co l lege Text-Books 7
Hill”s Norm al HistologyNormal Histology and Organography. By CHARLES Hl LL , M. D
I zmo of 483 pages , 337 il lustrations . Flexible leather, net .
N ew ( 3d ) E d i tion.
Dr. Hil l ’s work is characterized by a brevity of style, yet a completeness of d iscussion
,rarely met in a book of this size . The entire field
is covered,beginning wi th the preparation of materia l
, the cel l , thevarious tissues
,on through the d ifferent organs and regions, and end
ing with fixing and staining so lutions.Dr. E. P . Po rterfie ld , St . Louis Universi ty : I am very much gratifiedto find so handy a work. I t is so fu l l and comp lete that i t meets allrequirements .
Bohm , David o lilf, Huber’s Histology
Histology. By A . A . BO HM,M . D .
,and M . VO N Davm or r ,
M . D. ,of Munich . E d ited by G . CA R L HU B ER
, M . D . ,Professor
o f Embryo logy at the W istar Inst itute , U nivers ity of Pennsyl
vania . O ctavo Of 52 8 pages , 377 i l lustrations . Flexib le c loth,
net . Second E d ition .
Th is work is conceded to be the most complete text—book on humanhisto logy pub l ished . Particu larly fu l l on microscopic technic and
staining , i t is especia l ly serviceab le in the laboratory. Every step intechnic is clearly and precisely detai led . It is a work you can dependupon always.New Yo rk Med ica l J ou rnal : There can be noth ing but praise forth is model text—book and laboratory guide.
”
Keefer‘s Military HygieneMi lita ry Hygiene and Sanitation. By LI E UT.
-COL . FRANK R .
KE EPER , Professor of Mi l itary Hygiene, United States M i litaryAcad emy , West Point. rzmo of 30 5 pages , i l lustrated . C loth ,
net. J ust Read y.
You get here chapters on the care of troops , recruits and recrui ting , personal hygiene, physica l training , preventable diseases , cloth ing , equipment, water—supp ly, foods and their preparati on , hygiene and sani tationof posts
,barracks
, the troopsh ip , marches , camps , and batt lefields ; d i sposa l of wastes , tropic and arcti c service. venereal diseases , alcohol , etc .

8 Saun d e rs' Co l lege Text-Books
J ord an”s General Bacterio logyG enera l Bacter iology By E Dw rN O j O R DA N , Ph . D ., Professorof Bacterio logy , University of Chicago. Octavo of 650 pages ,
il lustrated . Cloth , net . New (4ln) E d ition .
Th is work treats fu l ly of the bacterio logy of p lants , mi lk and milkproducts
,dairying, agricu l ture, water , food preservation ; of leather
tanning , vinegar making, tobacco curing ; of househo ld admin istrationand sanitary engineering . A chapter of prime importance to all students of botany, horticulture, and agric ulture is that on the bacteria ld iseases of p lants.Prof. T. J .
‘ Burri l l , University of I l l inois : I am using Jordan’s Bacteriology for class work and am convinced that i t is the best text inexistence.
”
Byre”s Bacteriologic TechnicBacteriologic Technic . By J . W . H. E YRE , M . D Bacteriologistto Guy ’
s Hospital , Lond on. Octavo of 5 2 5 pages, i l lustrate dC loth ,
net. Second E d ition.
Dr. Eyre gives c learly the technic for the bacterio logic examination ofwater
,sewage
,air
,soi l
,mi lk and i ts products
,meats
, etc . It is a workof much va lue in the laboratory. The i l lustrations are practica l andserve wel l to clarify the text . The book has been great ly enlarged .
The Lond on Lanc et : It is a work for al l technical students,whether
of brewing,dairying
,or agricu l ture.
”
Bliss” Ch em ical AnalysisQualitative Chem ical A nalysis . By A . R . BLISS, J R Ph . G M. D
Professor of Chem istry and Pharmacy , B irm ingham Med ical College. Octavo of 2 50 pages. Cloth , $2 0 0 net .
Th is work was prepared Special ly for laboratory workers in the fieldsof medicine
,dentistry, and pharmacy . I t gives you systematic pro
ced ures for the detection and separation of the most common bases andacids , and in such a manner that , in a short time, you.
wi l l be enab ledto gain a good practical know ledge of the theory and methods of qual itative chemi cal analysis .

IO Saun d e rs’ Co l lege Text-Bo oks
Morrowi
s Care of inj uredIm m ed iate Ca r e of t/ze Inj u r ed . By ALB ERT S. MO RROW
,M.D
Ad junct Professor of Surgery , New York Polyc linic . Octavo of
360 pages , 24 2 i l lustrations . Cloth , nct . Second E d i tion
Dr. Morrow’s book tel ls you fast what to d o in any emergency, and i tis i l lustrated in such a practica l way taat the idea is caught at once.
There is no book better adapted to first-a id class work .
Hea lth : Here is a book that should find a place in every workshopand factory and shou ld be made a text-book in our schoo ls .
Am erican lllllln strated l Dic tionaryAm er ican It/ustra tect tiled z
'
ea l D ictiona ry . By W. A . N BWM A N
DO R L A N I) , M. D . ,Mem ber of Comm ittee on N om enc lature and
C lass ification o f Diseases , Am erican Med ica l Assoc iation . O c tavo
of 1 1 0 7 pages , with 3 23 illustrations, 1 1 9 in co lors. Flexibleleather, $4 .50 net thumb ind exed , net. E ighth E d ition
If you want an unabridged med ical dictionary, th is is the one youwant . I t is down to the minute ; its defin itions are conc ise, yet accurate and clear ; it is extremely easy to consult ; i t defines al l the newestterms in med icine and the al l ied subjects ; i t is profusely i llustrated.John B . Mu rphy , M . D N orthwestern University : It is unquestionab ly the best lexicon on med ica l topics in the Eng l ish language, andwith al l that
,i t is so compact for ready reference .
”
Am erican Pocket DictionaryAmerican Pocket Med ical Dictionary. E d ited by W . A. N EW
MA N DO RLAND , M . D. 69 3 pages . Flexible leather, net ;
thumb ind ex, net . New (9 111 ) E d i tion.
A dictionary must be fu l l enough to give the student the informationhe seeks
,c learly and simp ly , yet i t must not confuse him wi th detail .
The editor has kept this in mind in compi l ing th is Pocket Dictionary.
1. V . S. Stanis lau s , M. D. , Med ico-Ch irurgical Co l lege : “We havebeen strong ly recommending th is l i tt le book as being the very best .”DESCR IPTIVE CIRCULARS OF ALL BOOKS SEN T FREE




































































































