Embed Size (px)
Citation preview

Título: Análisis del efecto de suplementación crónica con pectinas en los niveles de proteína UCP1, ATGL
y PGC1α en tejido adiposo blanco epididimal de ratas
AUTOR: María Gabriela Cuyún Prado
Memoria del Trabajo de Fin de Máster
Máster Universitario en Nutrigenómica y Nutrición Personalizada (Especialidad/Itinerario Investigación)
de la
UNIVERSITAT DE LES ILLES BALEARS
Curso Académico 2012- 2013
Fecha: 27 Septiembre 2013 Firma del autor_________________
Nombre Tutor del Trabajo: Ana María Rodríguez Guerrero Firma Tutor____________________
Nombre Cotutor: Teresa Priego Cuadra Firma Cotutor__________________
Aceptado por el Director del Máster Universitario en Nutrigenómica y Nutrición Personalizada Firma ___________________

2
Abreviaciones AGCC – Ácido Graso de Cadena Corta
AMPK – Cinasa Activada por AMP
AMPc – Adenosin monofosfato cícico
ATGL- Adipose triglyceride lipase
ATP – Adenosin Trifosfato
AX - Arabinoxylans
DAG - diacilglicerol
GE – Grado de Esterificación
GLP - glucagon-like peptide
GRP43 - G-coupled receptor protein 43
HEP – Pectinas de alto grado de esterificación
ITF – inulina tipo fructano
LPS – lipopolisacárido
MI – Microbiota Intestinal
PPARα – Peroxisome Proliferator Activated Receptor α
PGC1α – PPAR coactivador-1 α
PYY - Péptido YY
SNS – Sistema Nervioso Simpático
SRBP – sterol regulatory element binding protein
TAB – Tejido Adiposo Blanco
TABe – Tejido Adiposo Blanco epididimal
TAG –triacilglicerol
TAM – Tejido Adiposo Marrón
TGI – Tracto Gastrointestinal
UCP1 – Proteína desacoplante 1
WB – Western Blot

3
Resumen
Los efectos fisiológicos de los prebióticos están relacionados con la mejora de la
composición de la microbiota intestinal, funciones de la barrera intestinal, reducción del
riesgo de algunas enfermedades metabólicas, entre otros.
En este estudio se utilizaron ratas suplementadas con pectinas de alto grado de
metoxilación (HEP) como prebióticos hasta los seis meses de edad, momento en el
que fueron sacrificados y se recuperaron muestras de los tejidos para realizar el
análisis de marcadores bioquímicos (no realizado por la autora). Posteriormente, por la
técnica de Western Blot se midieron tres proteínas relacionadas con el metabolismo
lipídico y oxidativo (UCP1, ATGL y PGC1α) para analizar el efecto de la
suplementación crónica con pectinas.
Las ratas que recibieron la suplementación con pectinas presentaron una disminución
de la adiposidad y peso corporal, asimismo presentaron niveles significativamente
menores de insulina circulante, índice HOMA y leptina comparado con los grupos
control. También se observó un incremento significativo en la expresión a nivel de
proteína de PGC1α, lo cual sugiere un incremento en el metabolismo oxidativo de los
animales suplementados. Respecto a la expresión a nivel de proteína de UCP1, no
hubo hallazgos y de ATGL los hallazgos no fueron significativos.
En conclusión, el estado de salud de los animales que recibieron suplementación con
pectinas es mejor respecto a los grupos control, de acuerdo a parámetros de
adiposidad, peso corporal, niveles circulantes de insulina, índice HOMA, niveles de
leptina y el aumento del metabolismo oxidativo relacionado con el incremento de la
expresión a nivel de proteína de PGC1α.
Palabras clave: Prebióticos; Pectinas; Tejido Adiposo Blanco; PGC1

Índice
Abreviaciones __________________________________________________________________________ 2 Introducción ____________________________________________________________________________ 5
Obesidad y microbiota intestinal _________________________________________________________ 5 Disbiosis y prebióticos _____________________________________________________________________ 6 Prebióticos y Pectinas ______________________________________________________________________ 9 Papel del Tejido Adiposo Blanco _______________________________________________________ 10
Objetivos _______________________________________________________________________________ 12 Diseño experimental y metodología utilizada __________________________________ 12
Diseño experimental _______________________________________________________________________ 12 Metodología _________________________________________________________________________________ 14 Detección de proteínas específicas por WB __________________________________________ 14
1. Homogenización de tejido adiposo blanco _______________________________________ 15 2. Método de Bradford _________________________________________________________________ 16 3. Electrotransferencia e inmuno-detección ________________________________________ 16
Estadística empleada ______________________________________________________________________ 18 Resultados y discusión _____________________________________________________________ 18
Peso corporal en respuesta a tratamiento y dieta ___________________________________ 18 Niveles de glucosa en sangre en respuesta a tratamiento y dieta _______________ 20 Niveles circulantes de insulina en respuesta a tratamiento y dieta _____________ 21 Índice HOMA en respuesta a tratamiento y dieta ______________________________________ 23 Niveles circulantes de leptina en respuesta a tratamiento y dieta _______________ 24 Nivel de proteína UCP1 en respuesta a tratamiento y dieta _______________________ 25 Niveles de proteína ATGL en respuesta a tratamiento y dieta ____________________ 26 Niveles de proteína PGC1α en respuesta a tratamiento y dieta __________________ 27
Conclusiones __________________________________________________________________________ 28 Comentarios ___________________________________________________________________________ 29 Anexos __________________________________________________________________________________ 30
Anexo 1 - Protocolo para determinación de la cantidad de proteína ____________ 30 1. Fundamento __________________________________________________________________________ 30 2. Reactivos _____________________________________________________________________________ 30 3. Procedimiento ________________________________________________________________________ 30
Anexo 2 - Protocolo para Western Blot ________________________________________________ 31 1. Preparación geles de acrilamida __________________________________________________ 31 2. Electroforesis en gel de poliacrilamida en presencia de SDS (SDS-PAGE) _ 32 3. Electrotransferencia (semi-dry electroblotter) ____________________________________ 33 4. Revelado de la membrana _________________________________________________________ 33
Bibliografía _____________________________________________________________________________ 35

5
Introducción
Obesidad y microbiota intestinal
La epidemia mundial de la obesidad representa un serio problema para salud
pública, la causa primordial está básicamente relacionada con un desequilibrio
nutricional durante un período largo de tiempo (revisado en1,2). El desarrollo de la
obesidad asociado a una ingesta elevada en grasas está relacionado entre otros
factores, con el sistema inmune innato (revisado en3). De hecho, una dieta
hiperlipídica desencadena el desarrollo de obesidad, inflamación, resistencia a la
insulina, diabetes tipo 2, entre otros (revisado en3,4).
El tracto gastrointestinal (TGI) es uno de los ecosistemas de la naturaleza más
poblados e investigados.5 En los últimos años, se ha estudiado acerca de la microbiota
intestinal (MI), la cual en condiciones ideales vive en simbiosis con el hospedador.
Representa en tamaño más de 100 veces el genoma humano y hoy en día se
considera como órgano microbiano.5 Además de la función del intestino en la digestión
y absorción de nutrientes, etc., el TGI humano contiene una diversa recolección de
microorganismos, que habitan principalmente en el colon. (revisado en1,3,5).
Este nuevo panorama abre el camino para comprender cómo la MI es
fundamental en el control del metabolismo energético. La MI se encuentra involucrada
en muchas funciones biológicas intestinales como: defensa contra patógenos,
inmunidad, desarrollo de microvellosidades, degradación de polisacáridos no
digeribles, entre otras.6-8 Recientes evidencias indican que la MI es esencial en el
control de la homeostasis energética del hospedador. De hecho, se cree que la MI
almacena energía para el hospedador a partir de compuestos dietéticos, ingeridos
pero no digeridos por éste mismo (revisado en1,3).
Coexistimos en simbiosis con nuestra MI, pero en algunos casos, esta relación
se torna patológica como es el caso de la obesidad, diabetes, etc.; factores incluyendo
la edad, genética y patrones alimenticios se ven influenciados en la composición de la
MI.6
Se ha observado que la composición de la MI puede ser distinta entre
individuos sanos e individuos que presenten obesidad y diabetes tipo 2. Esto ha
conducido a la investigación de este factor medioambiental como una pieza clave

6
entre la fisiopatología de estas enfermedades metabólicas y su relación con la MI (revisado en3,7). Ley, et al. (revisado en3) demuestra en modelos animales que la
obesidad puede estar asociada a una microbiota alterada. Otras intervenciones en
humanos, han demostrado cambios en la MI detectables con 24 horas de iniciación de
dieta hiperlipídica/baja en fibra o hipolipídica/alta en fibra.6
La MI puede estar involucrada en el desarrollo de inflamación leve, asociada
con desórdenes metabólicos relacionados con la obesidad (revisado en 1). Por
ejemplo, un producto bacteriano como el nivel sérico de lipopolisacárido (LPS), se ha
observado que es aproximadamente el doble en individuos obesos, diabéticos o
individuos con una dieta hiperlipídica, en comparación a individuos sanos.7,8
Otro producto bacteriano son los ácidos grasos de cadena corta (AGCC) que
provienen de la degradación de la matriz y otros polisacáridos dietéticos (pectinas,
almidón, xilanos e inulina)8. Como se menciona anteriormente, la MI es capaz de
obtener energía a través de la dieta y puede favorecer el incremento de la lipogénesis
(revisado en3,8).
Disbiosis y prebióticos
Uno de los enfoques actuales se centra en la investigación de los beneficios
que aporta la administración de oligosacáridos prebióticos. Éstos estimulan el
crecimiento y/o nivel de proteína metabólica de las bacterias intestinales, por lo cual
resulta interesante evaluar los efectos de prebióticos en la microbiota intestinal como
alimentos funcionales y suplementos.8
El concepto de prebiótico dietético es un ingrediente fermentado
selectivamente que da como resultado cambios específicos en la composición y/o nivel
de proteína de la microbiota intestinal, confiriendo de esta manera beneficio(s) al
hospedador9 (Figura 1).

7
Figura No.1
Recientemente muchos artículos y revisiones concuerdan con la idea de
normobiosis9 y disbiosis (Figura 2), que es el cambio inadecuado de microbiota
intestinal y composición y/o nivel de proteínas relacionadas con la enfermedades del
hospedador y caracteriza a individuos con sobrepeso u obesidad.1,7 Esto da lugar a la
pregunta de si la “disbiosis” es real, además ¿se puede considerar que hay alguna
manera de modificar favorablemente el ambiente microbiano y mejorar la salud y
bienestar del hospedador? (revisado en1,3).
Por otro lado, se ha observado que la suplementación o tratamiento con
prebióticos modifica el patrón de expresión génica en tejido adiposo blanco en
modelos animales, produciendo un incremento de la lipólisis, descenso de
adipogénesis e incremento en la respuesta metabólica a hormonas como la leptina e
insulina; todos estos fenómenos en conjunto contribuyen a una reducción de la
adiposidad (revisado en1).
Mejora • y/o estabilización de la composición de la microbiota intestinal • de funciones intestinales • absorción mineral y salud ósea • de las funciones de la barrera intestinal (reducción de endotoxemia metabólica)
Modulación • de la producción de péptidos gastro-‐intestinales • metabolismo energético • saciedad
Iniciación, regulación y modulación • (post natal) de funciones inmunes
Reducción del riesgo de • infecciones intestinales • obesidad, diabetes tipo 2, síndrome metabólico, etc. • cáncer de colon
Resumen de los principales efectos de los prebióticos a nivel fisiológico y
fisiopatológico.9

8
Figura No.2
Efecto de los carbohidratos dietéticos con propiedades prebióticas en la
fisiopatología del hospedador relacionado a la obesidad. Se ha observado
que los carbohidratos dietéticos con propiedades prebióticas son capaces
de modificar la composición de la microbiota intestinal, favoreciendo a las
bacterias involucradas en el control de la función de la barrera intestinal y la
inmunidad del hospedador. En el tracto intestinal, los prebióticos ayudan a
reforzar la barrera intestinal y las hormonas gastrointestinales que controlan
el apetito, homeostasis de la glucosa y la inflamación sistémica. El enfoque
del uso de prebióticos es también contrarrestar la esteatosis hepática,
resistencia a la insulina a nivel hepático y la adiposidad, modificando la
expresión genética a nivel tisular. SRBP – sterol-regulatory-element-binding
protein; GRP43, G-coupled receptors protein 43; GLP, glucagon-like
peptide; PYY, peptide YY. ITF, insulin-type fructans; AX, arabinoxylans
(imagen adaptada1).

9
Prebióticos y Pectinas
La pectina es una fibra soluble que se encuentra en las paredes celulares de
muchas plantas.10 En los últimos años su investigación ha adquirido gran interés
debido a que sus aplicaciones pueden ser muy diversas en base a sus propiedades
físico-químicas y a su biodegradabilidad.11 Las pectinas son probablemente los
polisacáridos más complejos que se pueden encontrar en las paredes celulares de
ciertas plantas; éstas consisten casi exclusivamente de homogalacturanos
(polisacárido formado a partir de residuos de ácido galacturánico) renombrados por su
habilidad formadora de geles.12 El interés en el tamaño y su distribución de las
partículas de las pectinas a lo largo del tracto gastrointestinal se debe a estas
características físicas que le confieren un rol importante en el proceso de digestión,
tránsito intestinal, fermentación y excreción fecal.12
La capacidad formadora de gel de las pectinas se encuentra relacionada con el
grado de metoxilación.13 Las pectinas se pueden clasificar en dos grupos principales
en función de su grado de esterificación: pectinas de alto/bajo grado de
metoxilación.11,12 El grado de metoxilación está definido como el número de moles de
ésteres metil por 100 moles de residuo de ácido galacturónico.12 Éste está expresado
en los grados de esterificaciones (GE); se considera alto grado de metoxilación (HEP)
cuando es >50% GE y forma geles viscosos en el estómago a un pH de 2,2-3,5.12 Esta
propiedad, influye directamente en la capacidad formadora de geles de cada una. Las
pectinas HEP forman geles con un elevado contenido de azúcar (>60%) a un pH
bajo.12 Se ha demostrado que las pectinas HEP tienen un mayor efecto inhibitorio en
cuanto al efecto y la captación de glucosa a nivel intestinal respecto a las pectinas de
bajo grado de metoxilación.9,14
La pectina al ser ingerida, es capaz de atravesar de forma inalterada la mayor
parte del tracto digestivo, por lo tanto esta permanece intacta en la parte alta del tracto
gastrointestinal.11 Esto es debido a la estructura tridimensional que forman las paredes
celulares, que confieren sus características físico-químicas: el tamaño de la partícula,
el área de superficie, hidratación, viscosidad, gelificación, capacidad de intercambio
catiónico, entre otros.12
Altas concentraciones de azúcar crean condiciones de baja nivel de proteína de
agua lo cual promueve interacciones en cadena en lugar de interacciones cadena-
solvente y el ácido asegura que los grupos carboxilos no sean disociados,

10
disminuyendo su repulsión cadena-electrostática.12 Por ello las pectinas son solamente
degradadas por la microflora del colon por biodegradación específica por las enzimas
producidas por las bacterias que forman parte de la flora colónica.11 Estas enzimas
fermentan la pectina originando gases como hidrógeno, dióxido de carbono, metano y
AGCC, ejerciendo así un efecto protector frente a desórdenes intestinales agudos o
crónicos.11 Otra propiedad importante de las pectinas es su capacidad de provocar
sensación de saciedad debido a su alta capacidad para absorber líquidos, de manera
que el consumo de alimentos puede verse disminuido.11 En concordancia con todo
esto, se ha observado que las HEP pueden modular parámetros como el peso
corporal, insulinemia, glucemia y lípidos sanguíneos, entre otros.14
No obstante por el momento pocos ensayos preclínicos y clínicos han
demostrado los efectos de los microbios intestinales específicos y prebióticos sobre
marcadores biológicos de desórdenes metabólicos, aunque los resultados obtenidos
hasta el momento indican que los avances en este campo podrían ser de utilidad en la
lucha contra la obesidad y desórdenes metabólicos asociados.8
Papel del Tejido Adiposo Blanco
El tejido adiposo juega un papel crítico en el control del balance energético.15 El
tejido adiposo blanco (TAB) tiene como función almacenar energía en forma de
triacilglicerol (TAG) para hidrolizar y proporcionar ácidos grasos para otros órganos en
escasez de energía.15,16 En el TAB se expresan diferentes genes claves para el control
del metabolismo energético. La función de la ATGL (Adipose triglyceride lipase)
conocida también como desnutrin17 es hidrolizar los TAG a diacilglicerol (DAG)
proporcionando sustratos para la lipasa sensible a hormonas (HSL).15,16 Según
Ahmadian et. al. la ATGL es fosforilada por AMPK para aumentar la lipólisis y esto es
acompañado de un incremento en la oxidación de ácidos grasos libres y de la
inducción de la UCP1 para termogénesis.15 Sin embargo, el mecanismo que regula la
nivel de proteína de la ATGL no está totalmente comprendido.16 Aunque también se
sabe que está regulada por la activación de la cascada del AMPc.18
Otra proteína importante es la UCP1 tiene un rol esencial en la termogénesis,
ésta es expresada en la membrana interna mitocondrial de los adipocitos marrones
particularmente.19 La regulación transcripcional del gen UCP1 tiene dos caras difíciles
de separar una de la otra, por un lado se encuentra la activación transcripcional por
señales vía endocrina, paracrina e intracrina y por otro lado, su expresión en los

11
adipocitos marrones y BRITE.20 Cuando es activada, promueve el desacoplamiento y
oxidación de sustratos, como combustible a partir de la producción de ATP, generando
calor.20
Cabe mencionar, que modificaciones en la ingesta dietética pueden estar
asociados con remodelación del tejido adiposo blanco (TAB), como la malnutrición
perinatal materna.19 Se ha observado que esta condición incrementa la aparición de
adipocitos BRITE (brown-in-white; adipocitos con características similares a los
marrones, que aparecen en los depósitos de TAB) en el TAB gonadal de ratas
destetadas, posiblemente por efectos indirectos en la nivel de proteína del sistema
nervioso simpático (SNS).19
El desarrollo multiocular propio de los adipocitos marrones en los depósitos
TAB en etapa post-natal desaparece luego del destete y se especula que la presencia
de un elevado número de adipocitos BRITE en crías neonatas de madres con
restricción energética se debe a que la ingesta de energía probablemente es
insuficiente para promover la adipogénesis y se redirige hacia la termogénesis para
prevenir la hipotermia.19 En definitiva, existen diferentes fuentes que muestran que
determinadas características ambientales (en particular dietéticas) pueden favorecer el
aparecimiento de adipocitos BRITE a los depósitos de TAB y la expresión de UCP1.
Entre los reguladores maestros cabe destacar los factores de transcripción de
Peroxisome Proliferator Activated Receptor-α (PPARα) que participan en la
diferenciación de adipocitos marrones, en la inducción y expresión génica de UCP1 en
adipocitos marrones y en la inducción del PPAR coactivador-1 α (PGC1α), entre
otras.20 Estudios genéticos y bioquímicos sugieren que un efecto biológico de la
activación de PPARα es promover la remodelación del TAB, actuando a través de la
inducción del regulador PGC1α.19 El PGC1α es un coactivador transcripcional que
regula el metabolismo energético.21,22 El PGC1α interactúa con un alto número de
factores de transcripción involucrados en múltiples respuestas biológicas incluyendo la
termogénesis adaptativa, biogénesis mitocondrial y metabolismo de carbohidratos y
grasas.13 El PGC1α es altamente expresado en tejidos con abundantes mitocondrias
donde el metabolismo oxidativo está activo como el TAM, corazón y el músculo
esquelético.23 Sin embargo, se ha observado su expresión en TAB pero de manera
reducida.23 Alteraciones en la expresión del PGC1α están altamente involucradas en
desórdenes metabólicos como la obesidad, diabetes y cardiopatías.22

12
Objetivos
Dados los antecedentes aquí presentados, el objetivo principal fue analizar el
efecto de la suplementación crónica con pectinas en los niveles de proteínas clave
relacionadas con el metabolismo lipídico y energético en tejido adiposo blanco
epididimal (TABe) en un modelo experimental de propensión a obesidad (crías de
madres sometidas a una restricción calórica moderada durante los primeros 12 días
del embarazo).
Diseño experimental y metodología utilizada
Diseño experimental
Para el presente trabajo se utilizaron las muestras TABe obtenidas del
experimento realizado en el laboratorio anteriormente (no realizado por la autora) cuya
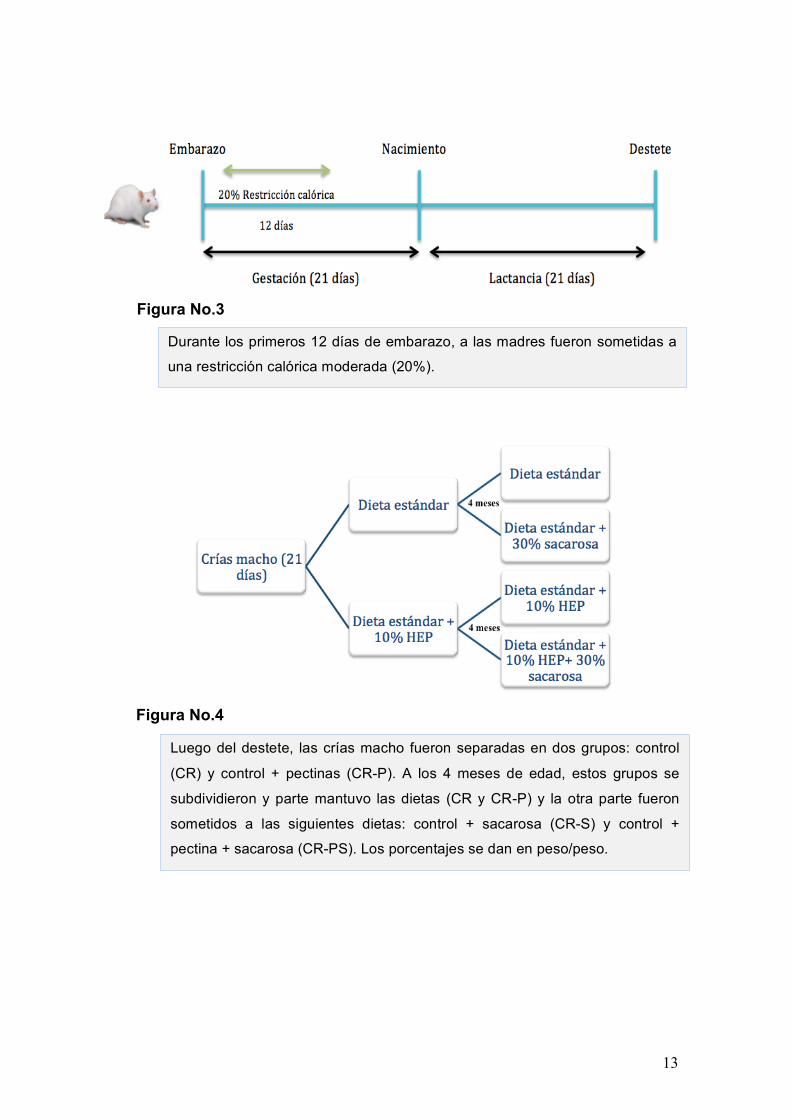
metodología es la expuesta a continuación. Durante los primeros 12 días de embarazo
a las madres se les realizó una restricción calórica moderada de un 20%24 (Figura 3).
Luego del destete se dividieron a las crías macho en dos grupos hasta los 4
meses, parte de las crías fueron alimentadas con una dieta estándar (se denominó
grupo CR, ya que son animales descendientes de madres con restricción calórica,
pero alimentados con dieta control) y otra parte fueron suplementadas con 10% de
pectinas de alto grado de esterificación (HEP) (grupo CR-P).
Posteriormente se subdividieron los grupos, al primer subgrupo se les mantuvo
las dietas (CR y CR-P), y a los otros dos subgrupos se les alimentó con una dieta rica
en sacarosa (30%) hasta los 6 meses de vida, momento en el que se sacrificaron los
animales y se recuperaron muestras de los tejidos, se realizó un seguimiento del peso
y composición corporal de los animales (grupo CR-S/ grupo CR-PS) (Figura 4).
13
Figura No.3
Figura No.4
Durante los primeros 12 días de embarazo, a las madres fueron sometidas a
una restricción calórica moderada (20%).
Luego del destete, las crías macho fueron separadas en dos grupos: control
(CR) y control + pectinas (CR-P). A los 4 meses de edad, estos grupos se
subdividieron y parte mantuvo las dietas (CR y CR-P) y la otra parte fueron
sometidos a las siguientes dietas: control + sacarosa (CR-S) y control +
pectina + sacarosa (CR-PS). Los porcentajes se dan en peso/peso.

14
Metodología
Se utilizó la técnica de Western Blot (WB) para determinar los niveles de
proteínas relacionadas con el metabolismo lipídico y energético en concreto en el
TABe: Proteína desacoplante 1 (UCP1), Adipose triglyceride lipase (ATGL) y Receptor
Activado por Proliferador Peroxisomales coactivador-1 α (PGC1α). Parte experimental
en la cual participé.
Detección de proteínas específicas por WB
El análisis de los niveles de proteína UCP1, ATGL y PGC1α de TABe, fueron
realizados por medio de WB (n=28). Las muestras de TABe almacenadas a -80°C
fueron colocadas en nitrógeno a -200°C, se pesaron 0,3 mg de TABe
aproximadamente y se añadió el peso x3 de PBS celular con pH de 7,4.
Posteriormente se homogenizó el tejido y luego se centrifugó a 600g durante 10
minutos a 4°C y se utilizó el sobrenadante para el posterior análisis de los niveles de
las proteínas. Para determinar la cantidad de proteína se utilizó el método de Bradford.
La carga proteica utilizada fue de 20µg, seguidamente se sometió a electroforesis en
gel de poliacrilamida (SDS-PAGE; 10% poliacrilamida) y luego fue electrotransferida a
las respectivas membranas de nitrocelulosa (Bio-Rad, España). Posteriormente las
membranas fueron incubadas con tampón de bloqueo durante toda la noche, al día
siguiente se lavaron 3 veces con PBS-tween (los primeros dos durante 15 minutos y el
último durante 5 minutos). Luego cada membrana fue incubada durante 1 hora con el
anticuerpo primario (anticuerpo primario, anti-UCP1, anti-ATGL y anti-PGC1α) luego
se realizaron tres lavados nuevamente, se incuba cada membrana durante 1 hora con
el tampón de bloqueo. Para reciclar cada membrana y poder analizar las tres proteínas
de interés, se realizó un procedimiento de stripping entre los diferentes análisis de
inmuno-detección. Por último las membranas fueron incubadas durante 30 minutos
con el anticuerpo secundario (anti Ig-G de conejo) marcado con fluorescencia. (Tabla
No.1)
La membranas fueron escaneadas en el canal 700 u 800 nm (correspondiente
a la fluorescencia de marcaje de los anticuerpos utilizados), reveladas, y cuantificadas
(Odyssey, Li-Cor, España). La incubación con alfa tubulina se realizó como control de
la carga; el resultado de la cuantificación, se normalizó con los datos de alfa tubulina
para su posterior análisis.25 A continuación, en la tabla 1 se resumen los principales
anticuerpos primarios y secundarios utilizados.

15
Tabla No. 1 Incubación de membrana con anticuerpo primario y secundario
Incubado (1 hora) Anticuerpo primario (dilución y casa comercial)
Anticuerpo secundario
UCP1 Anti-UCP1 1:2000 GeneTex Inc. (GTX10983) 60 min
Anti-rabbit 1:10000 Odyssey, Li-Cor 30 min
ATGL Anti-ATGL Dilución Cayman Chemical 60 min
PGC1α Anti-PGC1 Dilución Santa Cruz, Biotechnology Inc. 60 min
αTUBULINA Alfa tubulina TU-02 mouse monoclonal IgM 1:2000 Santa Cruz, Biotechnology Inc. 60 min
Anti-mouse 1:10000 Odyssey, Li-Cor 30 min
A continuación, se expone brevemente las bases de la técnica comenzando
desde la homogenización del tejido adiposo blanco epididimal y la cuantificación de
proteínas por el método de Bradford (parte experimental en la cual participé).
1. Homogenización de tejido adiposo blanco El objetivo de la homogenización es romper la membrana plasmática y conseguir el
trasvase al medio de todo el contenido material. Dependiendo de la naturaleza del
material biológico a homogenizar y el objetivo del experimento se debe elegir la
técnica, en este caso se utilizó el mecanismo de fuerzas de cizalladura producidas por
líquido con el dispersor como homogeneizador. Es importante tomar en cuenta que el
material biológico es termolábil y degradable por las propias proteasas de degradación
intracelular. Por lo tanto es indispensable en el protocolo la presencia de aditivos en el
medio de homogenización para preservar las propiedades funcionales y estructurales
de la fracción de interés, controlar la temperatura y el tiempo en cada etapa.25

16
2. Método de Bradford Se utilizó el método de Bradford26 para determinar la concentración de proteínas. El
procedimiento se basa en la formación de un complejo entre el colorante Comassie
Blue G-250 y la proteína en solución. El complejo proteína-colorante provoca un
cambio en el máximo de absorción del colorante (465-595 nm). De hecho, la cantidad
de absorción es proporcional a la proteína presente.27 El rango de concentración linear
va desde 0.1 a 1.4 mg/ml de proteína utilizando BSA (albúmina de suero bovino) como
proteína estándar. El reactivo de Bradford es compatible con agentes reductores
(utilizados para estabilizar proteínas en solución) como ventaja ante otros
procedimientos similares que no son compatibles (Lowry y BCA).27 El reactivo de
Bradford es compatible únicamente con bajas concentraciones de detergentes. Las
ventajas que ofrece es que el reactivo está listo para su uso (no requiere de mezcla o
diluciones), con una incubación rápida de cinco minutos se puede proceder a la lectura
de la placa, la presencia de azúcares y sustancias reductoras no interfiere con este
reactivo27, entre otros. (Protocolo - Anexo 1).
3. Electrotransferencia e inmuno-detección La técnica de Western Blot (WB) permite al investigador determinar, con un anticuerpo
primario específico, la cantidad relativa de proteína presente en las distintas muestras.
La transferencia de proteínas por medio del WB se realiza mediante
electrotransferencia.
Brevemente, las muestras de los tejidos homogenizadas son preparadas con un
tampón que protege a la proteína de interés de ser degradada; la muestras son
separadas utilizando SDS-PAGE en base a su peso molecular (los principios teóricos
de la técnica de SDS-PAGE se explican en el siguiente sub-apartado)
El gel y la membrana se sitúan a modo de sándwich entre un conjunto de hojas
de papel de filtro y almohadillas de esponja en un marco de plástico. Se utilizan
membranas de naturaleza hidrofóbica, preferentemente (ya que ofrecen una gran
resistencia química a los reactivos y son estables mecánicamente). Para poder buscar
la proteína de interés con anticuerpos específicos contra ella, para esto la membrana
es incubada en el anticuerpo primario. Las proteínas se unen a la membrana de forma
no covalente quedando accesibles para interaccionar con las moléculas que permitirán
su reconocimiento. Por último, se incuba la membrana con un anticuerpo secundario
marcado con fluorocromo, el cual reconozca al anticuerpo primario y se adhiere a las
localizaciones del anticuerpo primario para su revelado posterior.25 (Anexo 2.1-2.2)

17
Electroforesis SDS-PAGE El nombre de esta técnica proviene de sus siglas en inglés SDS-PAGE (electroforesis
de gel de poliacrilamida - Sulfato dodecil de Sodio). Es un método utilizado para
separar las proteínas de acuerdo a su tamaño.28
La metodología consta de dos etapas: (1) desnaturalización de las proteínas, el
detergente SDS interacciona con la parte hidrofóbica de la macromolécula disociando
y desnaturalizando sus subunidades. El detergente proporciona carga negativa que
enmascara la carga intrínseca de la unidad peptídica, de forma que la densidad de
carga es constante.25 Esta propiedad anula el efecto del parámetro de la carga sobre
la movilidad electroforética que pasa a ser anódica y depende solamente del tamaño
molecular. Es importante tomar en cuenta que para que la proteína se desnaturalice es
necesario la acción conjunta de un detergente y un agente reductor para romper los
puentes disulfuro. La muestra debe prepararse en tampón de carga de la solución
proteica que contiene glicerol o sacarosa al 10% para poder aumentar la densidad de
la solución, también contiene SDS y presencia de un agente reductor.25 (2) La
separación electroforética por SDS-PAGE, el gel de poliacrilamida actúa de tamiz
molecular y la subunidades proteicas se separan en función de su tamaño molecular.
Las moléculas de menor tamaño tienen una mayor movilidad electroforética anódica.
La electroforesis habitualmente se desarrolla en el sistema de tampón discontinuo y en
presencia del detergente SDS mencionado anteriormente, para mantener condiciones
desnaturalizantes durante la separación.25 Esta técnica permite determinar el número
de subunidades con distinta movilidad electroforética que componen una
macromolécula y la estimación de su peso molecular. Es importante recalcar que
proporciona información acerca de las propiedades de las subunidades proteicas en
estado desnaturalizado.25 (Protocolo - Anexo 2.3)
Revelado de membranas – inmunofluorescencia La inmunofluorescencia es una técnica cuyo fundamento se basa en los conceptos
básicos de microscopía y fluorescencia. Permite la detección y visualización directa del
material biológico para obtener información sobre la presencia o ausencia de un
fluoróforo y su posición en la partícula biológica analizada.25 A través de esta técnica
es posible la visualización de proteínas específicos o antígenos en secciones de
células o tejidos a través de la unión de un anticuerpo específico químicamente
conjugado con tinción fluorescente.25

18
Los métodos de detección más habituales son con anticuerpos monoclonales o
policlonales específicos a través del procedimiento denominado inmunoblotting en una
solución de anticuerpos primarios que reconocen de forma específica a la proteína de
interés. En este caso, la membrana se preparó como ya lo he comentado
anteriormente. A su vez, este anticuerpo primario es reconocido por el anticuerpo
secundario el cual está conjugado con compuestos fluorescentes, estas propiedades
de fluorescencia de estas moléculas permiten la visualización de la posición de las
proteínas de interés en la membrana durante el posterior revelado.25 (Protocolo –
Anexo 2.4)
Estadística empleada
El análisis estadístico por ANOVA de 2 factores fue realizado con el programa,
Graph Pad Prisim (Versión 6.00, Mac) y Excel 2010. Se realizaron también las pruebas
de T-student sin embargo, no se encontraron resultados significativos, por lo que no se
presentan. El nivel de significancia se fijó en p < 0.05.
Resultados y discusión
De acuerdo a los resultados ya obtenidos del presente experimento realizado
en el Laboratorio de Biología Molecular, Nutrición y Biotecnología -LBNB- de la UIB se
utilizaron los datos de peso corporal, glucosa, insulina, leptina circulantes e índice
HOMA (no obtenidos por la autora). Posteriormente con el análisis estadístico se
compararon los datos experimentales con los resultados obtenidos de los niveles de
las proteínas analizadas: UCP1, ATGL Y PGC1α.
Peso corporal en respuesta a tratamiento y dieta
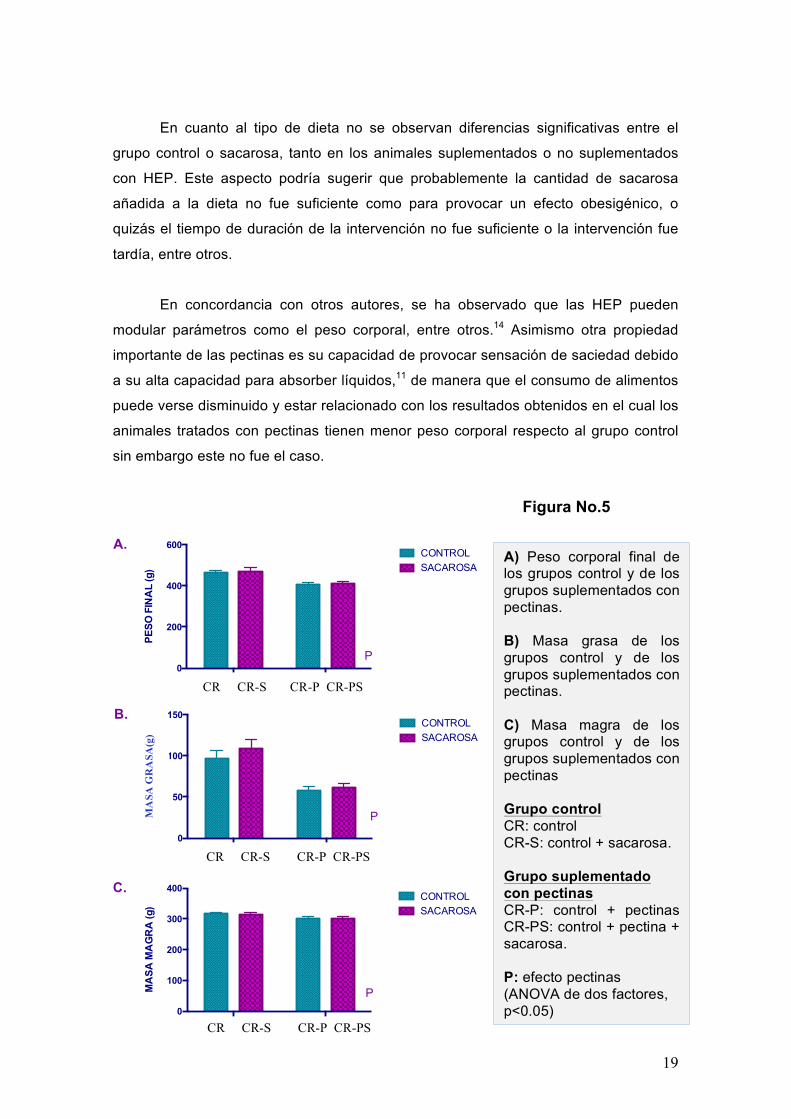
Respecto al peso corporal final de los animales (Figura 5A) se observa que los
animales suplementados con pectinas tuvieron menor peso estadísticamente
significativo respecto al grupo control (p<0.0001). Observándose un claro efecto P
(efecto pectinas) por el análisis de ANOVA de 2 factores. En cuanto a los resultados
obtenidos del peso corporal graso (Figura 5B) se observa que los animales
suplementados con pectinas tenían valores significativamente menores (p<0.0001)
respecto al grupo control. En los resultados obtenidos se observa que la composición
de masa magra (Figura 5C) no hay diferencia significativa entre los animales
suplementados con pectinas y el grupo control.
19
En cuanto al tipo de dieta no se observan diferencias significativas entre el
grupo control o sacarosa, tanto en los animales suplementados o no suplementados
con HEP. Este aspecto podría sugerir que probablemente la cantidad de sacarosa
añadida a la dieta no fue suficiente como para provocar un efecto obesigénico, o
quizás el tiempo de duración de la intervención no fue suficiente o la intervención fue
tardía, entre otros.
En concordancia con otros autores, se ha observado que las HEP pueden
modular parámetros como el peso corporal, entre otros.14 Asimismo otra propiedad
importante de las pectinas es su capacidad de provocar sensación de saciedad debido
a su alta capacidad para absorber líquidos,11 de manera que el consumo de alimentos
puede verse disminuido y estar relacionado con los resultados obtenidos en el cual los
animales tratados con pectinas tienen menor peso corporal respecto al grupo control
sin embargo este no fue el caso.
Figura No.5
CR
PECTINAS
0
200
400
600
TRATAMIENTO
PESO
FIN
AL
(g)
CONTROLSACAROSA
A.
P
CR
PECTINAS
0
100
200
300
400
TRATAMIENTO
MA
SA M
AG
RA
(g)
CONTROLSACAROSA
C.
P
CR CR-S CR-P CR-PS
CR CR-S CR-P CR-PS
CR CR-S CR-P CR-PS
CR
PECTINAS
0
50
100
150
TRATAMIENTO
PESO
GR
ASA
(g)
CONTROLSACAROSA
B.
PMA
SA G
RA
SA(g
)
A) Peso corporal final de los grupos control y de los grupos suplementados con pectinas. B) Masa grasa de los grupos control y de los grupos suplementados con pectinas. C) Masa magra de los grupos control y de los grupos suplementados con pectinas Grupo control CR: control CR-S: control + sacarosa. Grupo suplementado con pectinas CR-P: control + pectinas CR-PS: control + pectina + sacarosa. P: efecto pectinas (ANOVA de dos factores, p<0.05)

20
Niveles de glucosa en sangre en respuesta a tratamiento y dieta
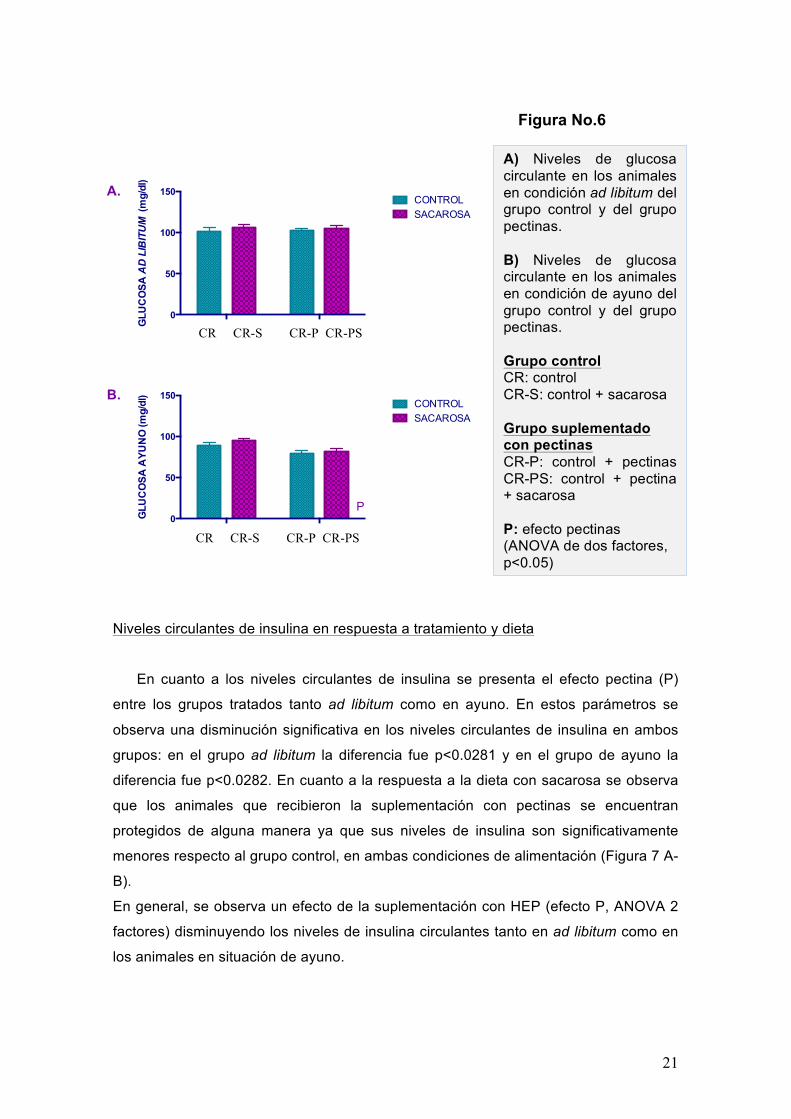
Aunque no existe una diferencia significativa entre grupos en la glucosa circulante
ad libitum (Figura 6A), sí se observa diferencia significativa (p<0.0016) en los niveles
de glucosa circulante en ayuno en los animales que han sido tratados con pectinas,
independientemente si recibieron dieta con o sin sacarosa (Figura 6B).
Ante este estrés metabólico al que se someten a los animales con la
suministración de sacarosa en la dieta se observa que tanto los que fueron
alimentados con dieta control y con la suplementación crónica de pectinas no
presentan diferencias significativas entre grupos ante este estímulo obesigénico, lo
cual podría indicar que probablemente el porcentaje que se les dio no fue suficiente
para provocar algún cambio más drástico.
Considero relevante los hallazgos en los niveles de glucosa circulante en ayunas
ya que todos los animales se encuentran en la misma condición, a diferencia de los
que estuvieron ad libitum. Otros autores han descrito beneficios de la glucemia en
condiciones de ayuno y/o postprandial luego de la administración de prebióticos (Ej.:
inulina) en modelos animales (revisado en1). Entre los mecanismos que podrían estar
implicados destaca: la secreción de péptidos intestinales con la función de las
incretinas, tales como péptido similar al glucagón 1, que participa en la mejora de la
resistencia a la insulina hepática (revisado en1).
Algunos estudios en humanos manifiestan igualmente una mejora en la glucemia
postprandial luego de la intervención de suplementación con prebióticos (revisado en 1). Los beneficios que puede llegar a aportar a la salud estas intervenciones dietéticas
y nutricionales que puedan lleguen a mejorar este marcador bioquímico, son
asombrosos. Considero interesante en este modelo analizar las modificaciones en la
composición de la microbiota intestinal y en los enterotipos predominantes antes y
después de la intervención para poder evaluar como estos cambios en la glucosa
circulante y demás marcadores bioquímicos están ligados.
21
Figura No.6
Niveles circulantes de insulina en respuesta a tratamiento y dieta
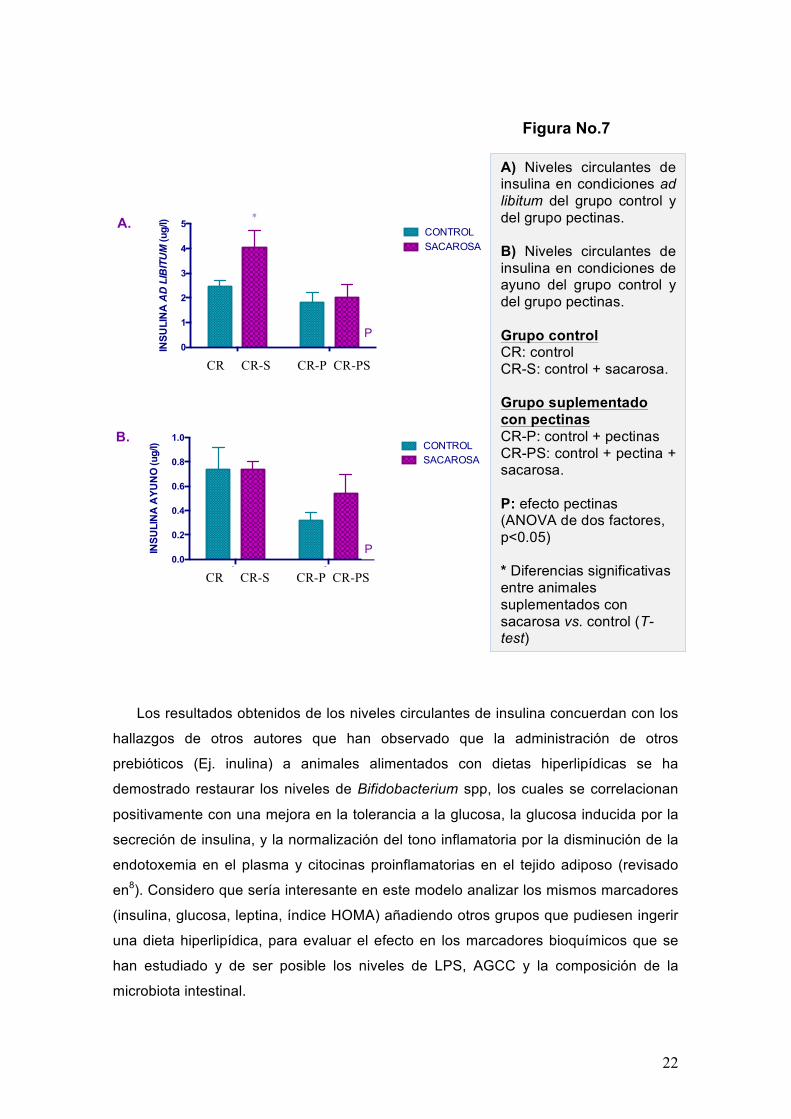
En cuanto a los niveles circulantes de insulina se presenta el efecto pectina (P)
entre los grupos tratados tanto ad libitum como en ayuno. En estos parámetros se
observa una disminución significativa en los niveles circulantes de insulina en ambos
grupos: en el grupo ad libitum la diferencia fue p<0.0281 y en el grupo de ayuno la
diferencia fue p<0.0282. En cuanto a la respuesta a la dieta con sacarosa se observa
que los animales que recibieron la suplementación con pectinas se encuentran
protegidos de alguna manera ya que sus niveles de insulina son significativamente
menores respecto al grupo control, en ambas condiciones de alimentación (Figura 7 A-
B).
En general, se observa un efecto de la suplementación con HEP (efecto P, ANOVA 2
factores) disminuyendo los niveles de insulina circulantes tanto en ad libitum como en
los animales en situación de ayuno.
CR
PECTINAS
0
50
100
150
TRATAMIENTO
GLU
CO
SA A
D L
IBIT
UM
(m
g/dl
)
CONTROLSACAROSA
A.
CR
PECTINAS
0
50
100
150
TRATAMIENTO
GLU
CO
SA A
YUN
O (m
g/dl
)
CONTROLSACAROSA
B.
P
CR CR-S CR-P CR-PS
CR CR-S CR-P CR-PS
A) Niveles de glucosa circulante en los animales en condición ad libitum del grupo control y del grupo pectinas. B) Niveles de glucosa circulante en los animales en condición de ayuno del grupo control y del grupo pectinas. Grupo control CR: control CR-S: control + sacarosa Grupo suplementado con pectinas CR-P: control + pectinas CR-PS: control + pectina + sacarosa P: efecto pectinas (ANOVA de dos factores, p<0.05)
22
Figura No.7
Los resultados obtenidos de los niveles circulantes de insulina concuerdan con los
hallazgos de otros autores que han observado que la administración de otros
prebióticos (Ej. inulina) a animales alimentados con dietas hiperlipídicas se ha
demostrado restaurar los niveles de Bifidobacterium spp, los cuales se correlacionan
positivamente con una mejora en la tolerancia a la glucosa, la glucosa inducida por la
secreción de insulina, y la normalización del tono inflamatoria por la disminución de la
endotoxemia en el plasma y citocinas proinflamatorias en el tejido adiposo (revisado
en8). Considero que sería interesante en este modelo analizar los mismos marcadores
(insulina, glucosa, leptina, índice HOMA) añadiendo otros grupos que pudiesen ingerir
una dieta hiperlipídica, para evaluar el efecto en los marcadores bioquímicos que se
han estudiado y de ser posible los niveles de LPS, AGCC y la composición de la
microbiota intestinal.
CR
PECTINAS
0
1
2
3
4
5
TRATAMIENTO
INSU
LIN
A A
D L
IBIT
UM
(ug/
l)
CONTROLSACAROSA
A.
P
CR
PECTINAS
0.0
0.2
0.4
0.6
0.8
1.0
TRATAMIENTO
INSU
LIN
A A
YUN
O (u
g/l) CONTROL
SACAROSA
B.
P
CR CR-S CR-P CR-PS
CR CR-S CR-P CR-PS
*
A) Niveles circulantes de insulina en condiciones ad libitum del grupo control y del grupo pectinas. B) Niveles circulantes de insulina en condiciones de ayuno del grupo control y del grupo pectinas. Grupo control CR: control CR-S: control + sacarosa. Grupo suplementado con pectinas CR-P: control + pectinas CR-PS: control + pectina + sacarosa. P: efecto pectinas (ANOVA de dos factores, p<0.05) * Diferencias significativas entre animales suplementados con sacarosa vs. control (T-test)
23
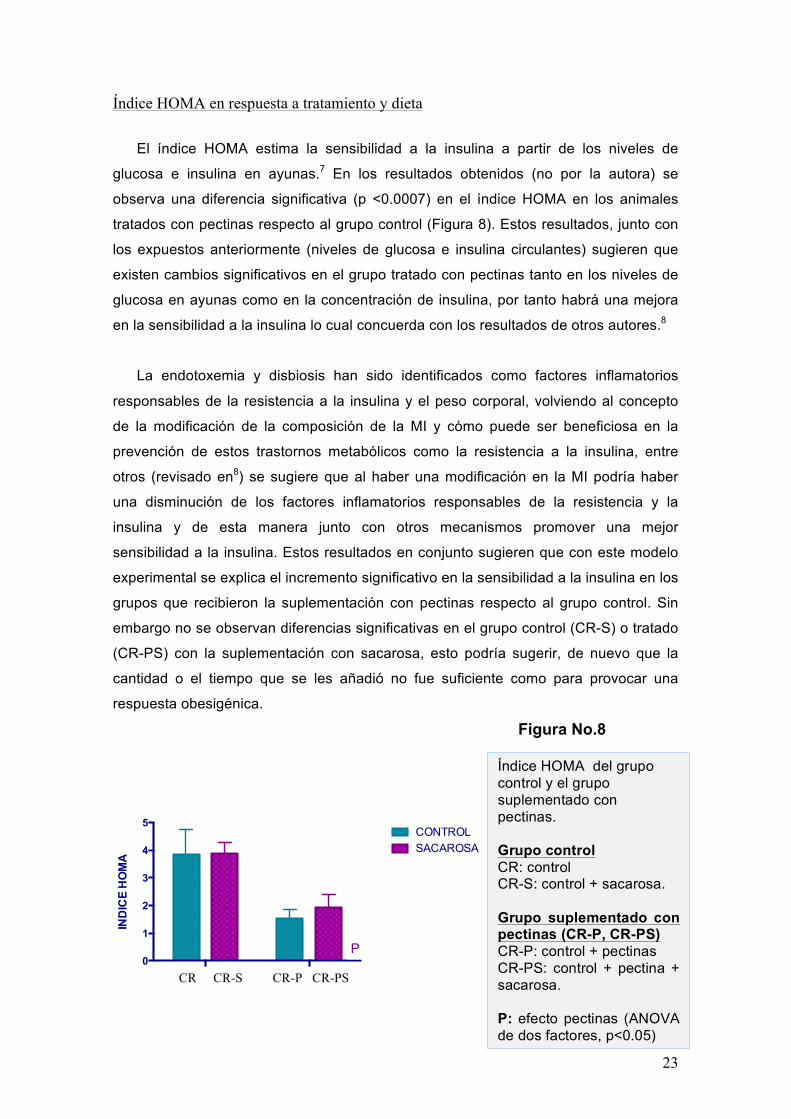
Índice HOMA en respuesta a tratamiento y dieta
El índice HOMA estima la sensibilidad a la insulina a partir de los niveles de
glucosa e insulina en ayunas.7 En los resultados obtenidos (no por la autora) se
observa una diferencia significativa (p <0.0007) en el índice HOMA en los animales
tratados con pectinas respecto al grupo control (Figura 8). Estos resultados, junto con
los expuestos anteriormente (niveles de glucosa e insulina circulantes) sugieren que
existen cambios significativos en el grupo tratado con pectinas tanto en los niveles de
glucosa en ayunas como en la concentración de insulina, por tanto habrá una mejora
en la sensibilidad a la insulina lo cual concuerda con los resultados de otros autores.8
La endotoxemia y disbiosis han sido identificados como factores inflamatorios
responsables de la resistencia a la insulina y el peso corporal, volviendo al concepto
de la modificación de la composición de la MI y cómo puede ser beneficiosa en la
prevención de estos trastornos metabólicos como la resistencia a la insulina, entre
otros (revisado en8) se sugiere que al haber una modificación en la MI podría haber
una disminución de los factores inflamatorios responsables de la resistencia y la
insulina y de esta manera junto con otros mecanismos promover una mejor
sensibilidad a la insulina. Estos resultados en conjunto sugieren que con este modelo
experimental se explica el incremento significativo en la sensibilidad a la insulina en los
grupos que recibieron la suplementación con pectinas respecto al grupo control. Sin
embargo no se observan diferencias significativas en el grupo control (CR-S) o tratado
(CR-PS) con la suplementación con sacarosa, esto podría sugerir, de nuevo que la
cantidad o el tiempo que se les añadió no fue suficiente como para provocar una
respuesta obesigénica.
Figura No.8
CR
PECTINAS
0
1
2
3
4
5
TRATAMIENTO
IND
ICE
HO
MA
CONTROLSACAROSA
P
CR CR-S CR-P CR-PS
Índice HOMA del grupo control y el grupo suplementado con pectinas. Grupo control CR: control CR-S: control + sacarosa. Grupo suplementado con pectinas (CR-P, CR-PS) CR-P: control + pectinas CR-PS: control + pectina + sacarosa.
P: efecto pectinas (ANOVA de dos factores, p<0.05)
24
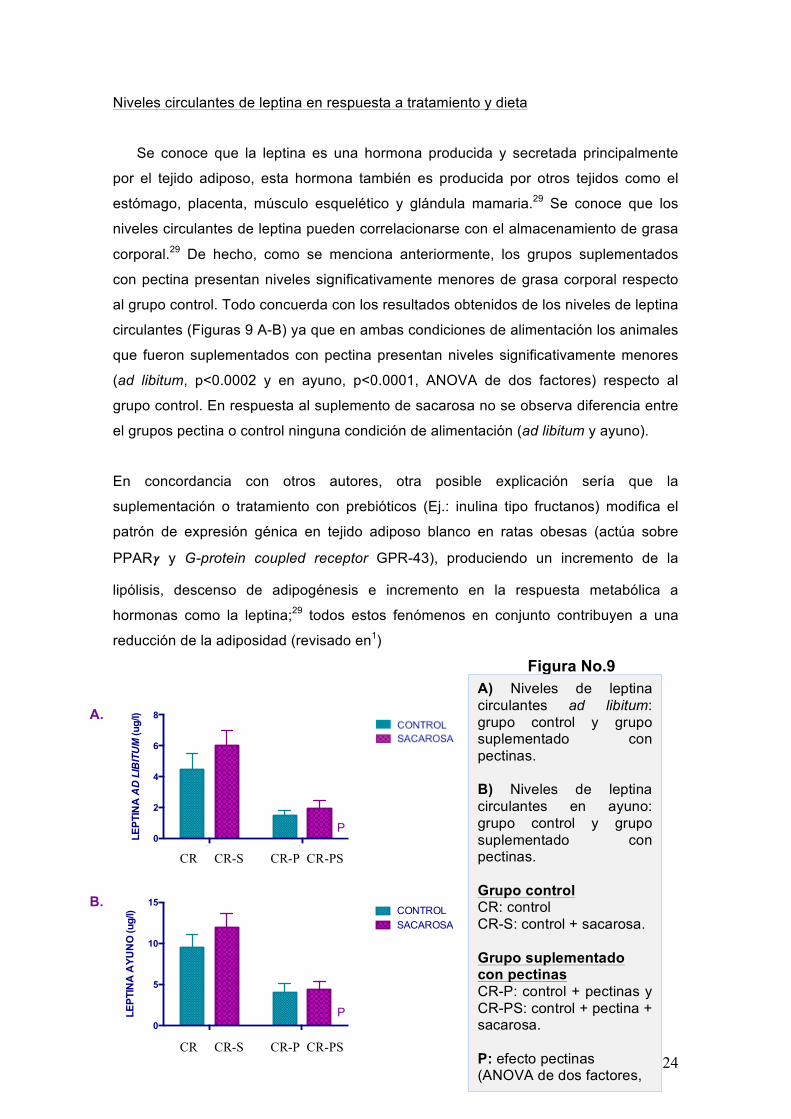
Niveles circulantes de leptina en respuesta a tratamiento y dieta
Se conoce que la leptina es una hormona producida y secretada principalmente
por el tejido adiposo, esta hormona también es producida por otros tejidos como el
estómago, placenta, músculo esquelético y glándula mamaria.29 Se conoce que los
niveles circulantes de leptina pueden correlacionarse con el almacenamiento de grasa
corporal.29 De hecho, como se menciona anteriormente, los grupos suplementados
con pectina presentan niveles significativamente menores de grasa corporal respecto
al grupo control. Todo concuerda con los resultados obtenidos de los niveles de leptina
circulantes (Figuras 9 A-B) ya que en ambas condiciones de alimentación los animales
que fueron suplementados con pectina presentan niveles significativamente menores
(ad libitum, p<0.0002 y en ayuno, p<0.0001, ANOVA de dos factores) respecto al
grupo control. En respuesta al suplemento de sacarosa no se observa diferencia entre
el grupos pectina o control ninguna condición de alimentación (ad libitum y ayuno).
En concordancia con otros autores, otra posible explicación sería que la
suplementación o tratamiento con prebióticos (Ej.: inulina tipo fructanos) modifica el
patrón de expresión génica en tejido adiposo blanco en ratas obesas (actúa sobre
PPAR𝜸 y G-protein coupled receptor GPR-43), produciendo un incremento de la
lipólisis, descenso de adipogénesis e incremento en la respuesta metabólica a
hormonas como la leptina;29 todos estos fenómenos en conjunto contribuyen a una
reducción de la adiposidad (revisado en1)
Figura No.9
CR
PECTINAS
0
2
4
6
8
TRATAMIENTO
LEPT
INA
AD
LIB
ITU
M (u
g/l)
CONTSACAROSA
A.
P
CR
PECTINAS
0
5
10
15
TRATAMIENTO
LEPT
INA
AYU
NO
(ug/
l) CONTROLSACAROSA
B.
P
CR CR-S CR-P CR-PS
CR CR-S CR-P CR-PS
A) Niveles de leptina circulantes ad libitum: grupo control y grupo suplementado con pectinas. B) Niveles de leptina circulantes en ayuno: grupo control y grupo suplementado con pectinas. Grupo control CR: control CR-S: control + sacarosa. Grupo suplementado con pectinas CR-P: control + pectinas y CR-PS: control + pectina + sacarosa. P: efecto pectinas (ANOVA de dos factores, p<0.05)

25
Con todos los resultados anteriores, se puede observar que la suplementación con
pectinas promueve una menor glucemia en ayunas, un menor índice HOMA y por
tanto, suponen una mejor sensibilidad a la insulina respecto al grupo control. También
se puede notar claramente que los animales suplementados con pectinas tienen una
menor ganancia de peso corporal. De acuerdo con lo anterior, los resultados en
general suponen un posible mejor estado de salud, al menos de los parámetros
estudiados en los animales que fueron suplementados con pectinas respecto al grupo
control.
Nivel de proteína UCP1 en respuesta a tratamiento y dieta
Se realizó el WB para medir la expresión a nivel de proteína de UCP1 sin embargo el
resultado fue confuso y poco convincente ya que el resultado no salía precisamente en
el lugar correspondiente al marcador de peso molecular ni al control positivo (TAM)
pero aún así no confirmaba con certeza que no era expresión de UCP1 lo que se veía.
Por lo tanto, se procedió a realizar de nuevo la incubación con UCP1 en la membrana
de confirmación (Figura 10) debido a la incertidumbre comentada en los primeros
geles. Enseguida, se realizó de nuevo la prueba con control positivo (TAM) y como
control negativo (hígado). Se obtuvo el resultado y se comprobó que no era UCP1 lo
que se observaba en los geles anteriores. Por tanto, no hubo hallazgos de UCP1 en
TABe.
Figura No.10
Membrana de confirmación UCP1. Prueba con TAM como control positivo e hígado como control negativo.
26
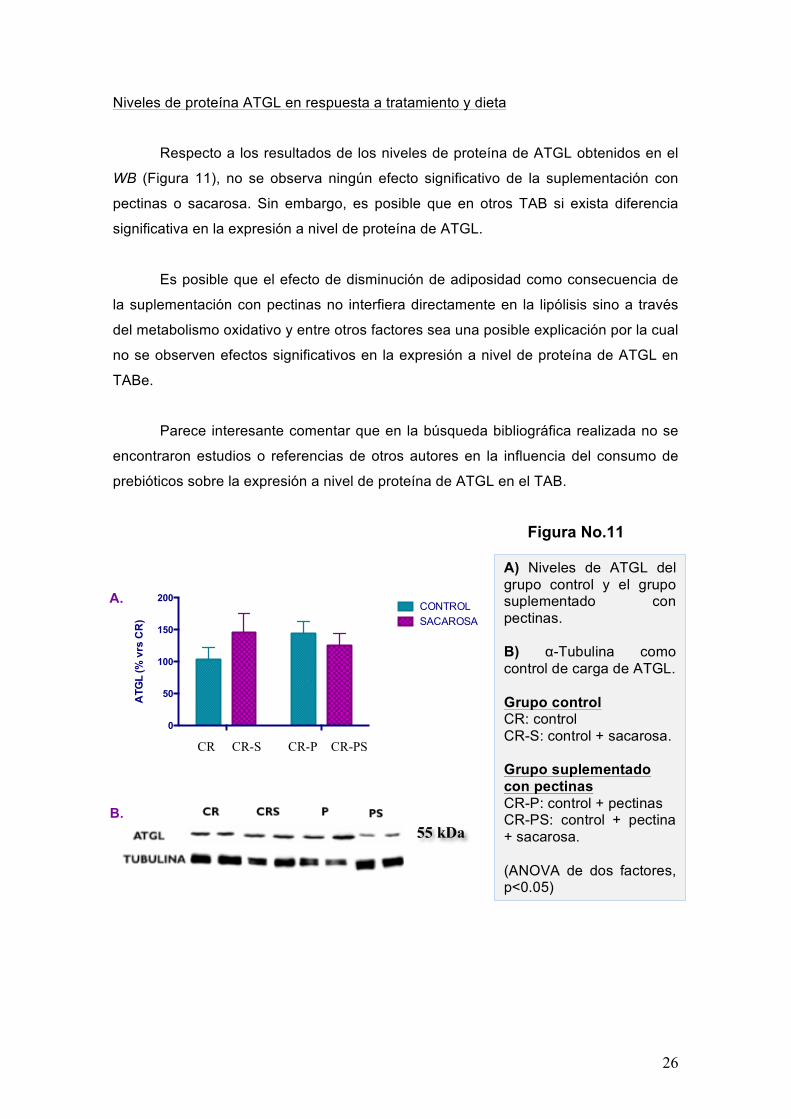
Niveles de proteína ATGL en respuesta a tratamiento y dieta
Respecto a los resultados de los niveles de proteína de ATGL obtenidos en el
WB (Figura 11), no se observa ningún efecto significativo de la suplementación con
pectinas o sacarosa. Sin embargo, es posible que en otros TAB si exista diferencia
significativa en la expresión a nivel de proteína de ATGL.
Es posible que el efecto de disminución de adiposidad como consecuencia de
la suplementación con pectinas no interfiera directamente en la lipólisis sino a través
del metabolismo oxidativo y entre otros factores sea una posible explicación por la cual
no se observen efectos significativos en la expresión a nivel de proteína de ATGL en
TABe.
Parece interesante comentar que en la búsqueda bibliográfica realizada no se
encontraron estudios o referencias de otros autores en la influencia del consumo de
prebióticos sobre la expresión a nivel de proteína de ATGL en el TAB.
Figura No.11
CR
PECTINAS
0
50
100
150
200
TRATAMIENTO
ATG
L (%
vrs
CR
)
CONTROLSACAROSA
CR CR-S CR-P CR-PS CR
PECTINAS
0
2
4
6
8
TRATAMIENTO
LEPT
INA
AD
LIB
ITU
M (u
g/l)
CONTSACAROSA
A.
P
55 kDa
CR
PECTINAS
0
5
10
15
TRATAMIENTO
LEPT
INA
AYU
NO
(ug/
l) CONTROLSACAROSA
B.
P
A) Niveles de ATGL del grupo control y el grupo suplementado con pectinas. B) α-Tubulina como control de carga de ATGL. Grupo control CR: control CR-S: control + sacarosa. Grupo suplementado con pectinas CR-P: control + pectinas CR-PS: control + pectina + sacarosa. (ANOVA de dos factores, p<0.05)

27
Niveles de proteína PGC1α en respuesta a tratamiento y dieta
Se ha descrito que el PGC1α es un coactivador transcripcional que regula
muchas rutas biológicas relacionadas con el metabolismo energético. Originalmente
fue descrito como un coactivador de PPAR que modula la expresión UCP1 y la
termogénesis del TAM, también se ha demostrado que su función en el control de la
biogénesis mitocondrial y el metabolismo oxidativo en muchos tipos de células.30
En la Figura 12 se presentan los resultados obtenidos en el WB de la nivel de
proteína de PGC1α en TABe, se observa una diferencia significativa en el aumento en
la expresión de los niveles de proteína de PGC1α (ANOVA de 2 factores) p<0.02,
entre los animales que fueron suplementados con pectinas (CR-P) respecto al grupo
control (CR) y también se observa una tendencia en el aumento en los niveles de
PGC1α en los animales suplementados o no con pectina que también recibieron
sacarosa (CR-PS, CR-S).
Por tanto, el efecto pecina observado en los niveles de expresión de PGC1α
tras la suplementación de pectinas y sacarosa, sugiere un aumento en el metabolismo
oxidativo de los animales que recibieron suplemento con pectinas. Resultaría
interesante continuar investigando otros marcadores ya que PGC1 actúa como
coactivador de otros factores de transcripción relacionados con la función mitocondrial
y de biogénesis en TAM, músculo esquelético y posiblemente en otros tejidos.
Figura No.12
CR
PECTINAS
0
50
100
150
200
250
TRATAMIENTO
PGC
1 (%
vrs
CR
)
CONTROLSACAROSA
P
CR CR-S CR-P CR-PS
CR
PECTINAS
0
2
4
6
8
TRATAMIENTO
LEPT
INA
AD
LIB
ITU
M (u
g/l)
CONTSACAROSA
A.
P
91 kDa
CR
PECTINAS
0
5
10
15
TRATAMIENTO
LEPT
INA
AYU
NO
(ug/
l) CONTROLSACAROSA
B.
P
A) Niveles de PGC1α del grupo control y del grupo suplementado con pectinas. B) α-Tubulina como control de carga de PGC1α. Grupo control CR: control CR-S: control + sacarosa Grupo suplementado con pectinas CR-P: control + pectinas CR-PS: control + pectina + sacarosa. P: efecto pectinas. (ANOVA de dos factores, p<0.05)

28
En resumen, los resultados presentados anteriormente muestran que la
suplementación con prebióticos como las pectinas promueven: una disminución de la
adiposidad, por tanto del peso corporal, mejor respuesta a la insulina confiriendo
protección a través de mecanismos que aún están por estudiar, mejor sensibilidad a la
insulina, niveles significativamente menores de leptina respecto a sus controles lo cual
concuerda con la reducción de la adiposidad y peso corporal. Según el aumento
significativo de los niveles de proteína de PGC1α, también se sugiere un aumento del
metabolismo oxidativo en los animales que recibieron la suplementación con pectinas.
Por otro lado, se sugieren una posible modificación de la microbiota intestinal,
mecanismos por investigar aún, continúa entre otros mecanismos involucrados que
promueven una beneficios a la salud. Recapitulando de esta manera, procedo a las
siguientes conclusiones.
Conclusiones
En general el estado de salud de los animales que recibieron suplementación
con pectinas es mejor respecto al grupo control de acuerdo a parámetros de
adiposidad, peso corporal, niveles circulantes de insulina y sensibilidad a la insulina,
niveles de leptina y según el incremento del nivel de proteína PGC1α se sugiere un
aumento del metabolismo oxidativo en estos animales.
La suplementación con pectinas de alto grado de metoxilación promueven una
disminución de la adiposidad y del peso corporal, este resultado se asocia con las
propiedades físico-químicas de estas y así mismo se sugiere la probabilidad de
mecanismos que involucran una modificación de la microbiota intestinal y otros
factores que están aún por estudiar.
El incremento significativo de la sensibilidad a la insulina observado en los
animales que recibieron la suplementación con pectinas observados así mismo en los
parámetros del metabolismo de glucosa e insulina, sugiere un posible uso terapéutico
de los prebióticos como las pectinas para promover una mejor respuesta a la insulina.
Respecto a los niveles de proteína desacoplante UCP1 obtenidos en este WB, no
hubo hallazgos en TABe y en cuanto a ATGL no se observaron efectos significativos,
sin embargo es posible que haya expresión a nivel de proteína en otros TAB en este
modelo experimental.

29
Los hallazgos en cuanto a los niveles de proteína de PGC1α en TABe en los
animales que recibieron la suplementación con pectinas se observa un aumento
significativo, lo cual sugiere un incremento en el metabolismo oxidativo de estos
animales. Sería interesante continuar con el estudio de otros marcadores ya que
PGC1 actúa como coactivador de otros factores de transcripción relacionados con la
función mitocondrial y de biogénesis en TAM, músculo esquelético y posiblemente en
otros tejidos.
Comentarios La frase “Where there´s a will there´s a way” me sorprende cada día más. Las
prácticas de laboratorio me han enseñado que no hay límites, que vale soñar. Ahora
me da ternura recordar el primer día del curso de técnicas de laboratorio, ya que
durante la clase la Profesora hablaba y yo no tenía idea de lo que decía, al salir fue un
sentimiento muy desagradable de frustración; ya que no entendí de lo que habían
hablado en clase, no tenía experiencia en laboratorio y era próximo a comenzar las
prácticas de laboratorio. Pero fue sorprendente como con disciplina de leer e
informarme y estudiar fuerte para las clases de técnicas de laboratorio para
prepararme porque sabía que el laboratorio iba a ser duro. Me sorprendió que fui
capaz con la ayuda y paciencia de mi tutora Teresa y compañera de laboratorio de
alcanzar los objetivos propuestos de las prácticas. Siempre me había llamado la
atención la investigación, me intrigaba mucho, sin embargo en mi país no se fomenta.
Pero ahora creo con más fuerza que el éxito está en la disciplina, ahora continúo con
el sueño y aspiración de continuar en el doctorado. En cuanto a la repercusión del
trabajo de laboratorio en mi formación como alumna, puedo agradecerles el hecho de
permitirme ver que puedo ser lo que quiero, que a través de disciplina, esfuerzo y
dedicación todo es posible.

30
Anexos
Anexo 1 - Protocolo para determinación de la cantidad de proteína
1. Fundamento El método de Bradford (1976) se basa en que el azul brillante Coomassie G-250 se
combina con las proteínas originando un complejo de color azul con un elevado
coeficiente de extinción, lo que permite cuantificar concentraciones de proteína desde
2µg/mL.
2. Reactivos - Patrón de proteínas: 1mg/mL de seroalbúmina bovina (BSA) 0.0100 g/10 mL
(BSA diluída en el tampón en que estén diluidas las muestras: PBS, Lysis
Buffer, etc.)
- Reactivo de Bradford (para 400 muestras o para 4 placas): Se disuelven 10 mg
de azul brillante Coomassie G-250 en 5 mL de etanol al 96%. Sin dejar de
agitar, se añaden 60 mL de agua y después 10 mL de H3PO4 al 85%. Se
enrasa a 100 mL con agua y se almacena en frasco topacio a temperatura
ambiente.
3. Procedimiento
- Se prepara la recta patrón pipetando, por duplicado, en la primera fila de la
placa 0, 2’5, 5, 7’5 y 10 µL de patrón.
- Si es necesario, se diluye la muestra con tampón de homogenado de modo
que su concentración de proteínas esté comprendida entre las de los tubos
patrones (el homogenado de hígado conviene diluirlo 1:10, el de tejido adiposo
no suele hacer falta)
- Se toman 5 µL de muestra
- Se les añade a todos los pocillos 250 µg/mL de reactivo de Bradford, se agita la
placa y se introduce en el lector de placa.
- El lector agita la placa, la incuba 2 minutos a temperatura ambiente (en
oscuridad) y la lee a 595 nm
- El resultado del lector se obtiene en mg/mL, pero hay que tener en cuenta las
diluciones realizadas
Bradford MM (1976). A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal biochem 72:248-
254

31
Anexo 2 - Protocolo para Western Blot
1. Preparación geles de acrilamida a. Reactivos
Resolving gel (10%)
i. 6,6 ml de solución stock de acrilamida
ii. 5 ml de tampón Tris-HCl pH 8,8
iii. 8,8 de agua bidestilada
iv. 200 µl SDS 10%
v. 20 µl TEDMED (añadirlo en campana)
vi. 100 µl de persulfato amónico al 10%
Stacking gel
i. 1,3 ml de solución stock de acrilamida
ii. 2,5 de tampón Tris-HCl pH 6,8
iii. 6 ml de agua bidestilada
iv. 100 µl SDS 10%
v. 20 µl de TEDMED (añadirlo en campana)
vi. 50 µl persulfato amónico al 10%
Observaciones: El TEDMED y el persulfato amónico actúan como catalizadores de la
polimerización de la acrilamida; deben añadirse en último lugar, justo antes de formar
los geles.
b. Procedimiento
El sistema de electroforesis de Laemmi incluye un gel en el que tiene lugar la
separación de las proteínas (resolving gel, con acrilamida al 10%) y un gel espaciador
(stacking gel, con acrilamida al 3%) que se polimeriza por encima del primero y que
sirve para concentrar las muestras.
- Ensamblar las placas de vidrio que delimitarán los geles y disponerlas
convenientemente
- Mezclar todos los ingredientes de la solución resolving gel, excepto el TEMED
y el persulfato, en un vaso de precipitado
- Añadir el TEMED y el persulfato y agitar suavemente haciendo girar el vaso,
procurando no formar burbujas de aire.
- Inmediatamente, pipetear la solución de resolving gel en las placas de
electroforesis hasta una altura de 5cm
- Con una pipeta y con cuidado, añadir 1 ml de agua destilada. El agua se añade
para conseguir que el borde del gel polimerice y quede plano (en contacto con

32
el aire, la acrilamida no polimeriza o lo hace mal). Dejar que el gel polimerice
durante un mínimo de 30 minutos, a temperatura ambiente
- Una vez polimerizando el resolving gel, eliminar el agua y absorber el agua
residual con un trozo de papel de filtro, con cuidado. Es importante que la
superficie del resolving gel quede seca, a fin de conseguir una buena
polimerización en la interfase de los geles
- Mezclar todos los ingredientes de la solución del stacking gel excepto el
TEMED y el persulfato. Añadir el temed y el persulfato, agitar suavemente
- Añadir la solución del stacking gel e inmediatamente introducir un peine entre
las dos placas de vidrio evitando la formación de burbujas de aire. Dejar que
polimerice el gel durante 30 minutos a temperatura ambiente. Quitar el peine y
lavar con tampón Tris-Glicina los pocillos.
- El gel está listo.
2. Electroforesis en gel de poliacrilamida en presencia de SDS (SDS-PAGE) a. Reactivos
i. Tampón Tris-Glicina (Tris 0,0025 M; Glicina 0,1912M) 1. 15,14 g Tris base 2. 72 g Glicina 3. 5 g SDS 4. Ajustar a pH 8,3 con HCl diluído 5. Enrasar a 5 litros con agua destilada 6. Guardar a temperatura ambiente
ii. Tampón Tris-HCl, pH 6,8 (Tris 0,5 M) 1. 6 g Tris Base 2. Ajustar a pH 6,8 con HCl diluído 3. Enrasar a 100 ml con agua destilada 4. Guardar en nevera
iii. Tampón Tris-HCl, pH 8,8 (Tris 0,5 M) 1. 18,15 g Tris Base 2. Ajustar a pH 8,8 con HCl diluído 3. Enrasar a 100 ml con agua destilada 4. Guardar en nevera
iv. Tampón para las muestras (sample buffer, Tris 0,0625 M) 1. 6,25 ml de Tris-HCl 0,5M, ph 6,8 2. 2,5 g SDS 3. 5 ml glicerol 4. 2,5 ml 2-mercaptoetanol 5. 0,125 ml de azul de bromofenol al 1% 6. Enrasar a 50 ml con agua destilada 7. Guardar en nevera
b. Procedimiento
- Disponer las muestras problema en tubos eppendorf y añadir el volumen
pertinente de tampón para las muestras. Se añade un mínimo de 1 µl de
tampón para muestras por cada 10 µg de proteína, de modo que la proporción

33
de proteína/SDS quede 1:4. Conviene aplicar entre 5-10 µg deproteína por
pocillo, en un volumen que no exceda los 15 µl.
- Con una aguja hipodérmica, practicar unos orificios en la tapa de los
eppendorf.
- Calentar las muestras en un baño de agua hirviendo durante 2 minutos. En
este momento las muestras están listas para ser palicadas a los geles.
- Ensamblar el equipo de electroforesis y correr la electroforesis en tampón Tris-
Glicina a 80 V durante aproximadamente 3 horas.
3. Electrotransferencia (semi-dry electroblotter) a. Reactivos v. Cathode buffer (pH 9,4)
1. 0,445 g β-Alanina 2. 2,301 g Tris-base 3. 100 ml Metanol absoluto 4. Enrasar a 500 ml con agua bidestilada
vi. Buffer 0,025 M Tris (pH 10,4) 1. 1,51 g Tris-base 2. 100 ml Metanol absoluto 3. Enrasar a 500 ml con agua bidestilada
vii. Buffer 0,3M Tris (pH 10,4) 1. 18, 17 g Tirs-base 2. 100 ml metanol absoluto 3. enrasar a 500 ml con agua bidestilada
b. Procedimiento
- Mojar con agua destilada las placas de grafito del electroblotter
- Colocar las diferentes hojas de papel Whatman sobre una de las placas de
grafito, de la siguiente manera y evitando la formación de burbujas y arrugas:
o 6 hojas modas en el buffer 0,3 M Tris
o 3 hojas mojadas en el buffer 0,025M Tris
o Membrana de nitrocelulosa mojada con el buffer 0,025M Tris
o El gel de poliacrilamida de la electroforesis realizada
o 9 hojas mojadas en el cathode buffer
- Finalmente se coloca la segunda placa de grafito y se conecta a una intensidad
constante de 300 mA durante 30 min.
4. Revelado de la membrana a. Reactivos
i. Tampón PBS-tween (PBS-T)
1. 1 L PBS (pH 7,5)
2. 1 g tween 20

34
ii. Tampón de bloqueo (Odyssey)
b. Procedimiento
Todo el procedimiento de revelado de la membrana debe realizarse en agitación a
temperatura ambiente.
- Bloquear la membrana con 10 ml de tampón de bloqueo durante toda la noche
- Lavar 3 veces con PBS-T: 1 lavado - 15 minutos, 2 lavados - 5 minutos
- Incubar 1 hora con 10 ml de PBS-T con 0,1% de BSA y con 5 µl del primer
anticuerpo (dilución 1:2000). El anticuerpo puede reciclarse, debiendo ser
guardado con azida sódica 0,1% y a 4°C.
- Lavar 3 veces con PBS-T: 1 lavado - 15 minutos, 2 lavados - 5 minutos
- Incubar 1 hora con 10 ml de tampón de bloqueo y con 1µl del segundo
anticuerpo marcado con fluorescencia (dilución1:1000)
- IMPORTANTE: a partir de este momento la membrana se debe proteger de la
luz
- Lavar 3 veces con PBS-T: 1 lavado - 15 minutos, 2 lavados - 5 minutos
- La membrana está preparada para la detección del marcaje:
o Secar la membrana antes de ponerla en el escáner
o Escanear la membrana escogiendo los canales adecuados según el
marcaje fluorescente del anticuerpo (799 o 800 nm)
o Si la señal de la membrana es muy débil o muy fuerte, se puede re-
escanear subiendo o bajando la intensidad del escáner.

35
Bibliografía
1 Delzenne, N. M., Neyrinck, A. M. & Cani, P. D. Gut microbiota and metabolic disorders: How prebiotic can work? Br J Nutr 109 Suppl 2, S81-85, doi:10.1017/S0007114512004047 (2013).
2 Hotamisligil, G. S. Inflammation and endoplasmic reticulum stress in obesity and diabetes. Int J Obes (Lond) 32 Suppl 7, S52-54, doi:10.1038/ijo.2008.238 (2008).
3 Cani, P. D. & Delzenne, N. M. The role of the gut microbiota in energy metabolism and metabolic disease. Curr Pharm Des 15, 1546-1558 (2009).
4 Gil, A., Ortega, R. M. & Maldonado, J. Wholegrain cereals and bread: a duet of the Mediterranean diet for the prevention of chronic diseases. Public Health Nutr 14, 2316-2322, doi:10.1017/S1368980011002576 (2011).
5 Xu, J. et al. Evolution of symbiotic bacteria in the distal human intestine. PLoS Biol 5, e156, doi:10.1371/journal.pbio.0050156 (2007).
6 Wu, G. D. et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 334, 105-108, doi:10.1126/science.1208344 (2011).
7 Dewulf, E. M. et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut, doi:10.1136/gutjnl-2012-303304 (2012).
8 Sanz, Y., Rastmanesh, R. & Agostonic, C. Understanding the role of gut microbes and probiotics in obesity: how far are we? Pharmacol Res 69, 144-155, doi:10.1016/j.phrs.2012.10.021 (2013).
9 Roberfroid, M. et al. Prebiotic effects: metabolic and health benefits. Br J Nutr 104 Suppl 2, S1-63, doi:10.1017/S0007114510003363 (2010).
10 Sánchez, D. et al. Highly methoxylated pectin improves insulin resistance and other cardiometabolic risk factors in Zucker fatty rats. J Agric Food Chem 56, 3574-3581, doi:10.1021/jf703598j (2008).
11 Pseidy Luz Mamani Crispin, R. R. C., Ma Dolores Veiga. Pectina: Usos Farmacéuticos y Aplicaciones Terapéuticas. (31 Octubre 2011).
12 Ralet, J.-F. T. a. M.-C. Pectins, their origin, structure and function. 32 (2008). 13 Judd, P. A. & Truswell, A. S. The hypocholesterolaemic effects of pectins in
rats. Br J Nutr 53, 409-425 (1985). 14 Kim, M. High-methoxyl pectin has greater enhancing effect on glucose uptake
in intestinal perfused rats. Nutrition 21, 372-377, doi:10.1016/j.nut.2004.07.006 (2005).
15 Ahmadian, M. et al. Desnutrin/ATGL is regulated by AMPK and is required for a brown adipose phenotype. Cell Metab 13, 739-748, doi:10.1016/j.cmet.2011.05.002 (2011).
16 Caimari, A., Oliver, P. & Palou, A. Adipose triglyceride lipase expression and fasting regulation are differently affected by cold exposure in adipose tissues of lean and obese Zucker rats. J Nutr Biochem 23, 1041-1050, doi:10.1016/j.jnutbio.2011.05.008 (2012).
17 Villena, J. A., Roy, S., Sarkadi-Nagy, E., Kim, K. H. & Sul, H. S. Desnutrin, an adipocyte gene encoding a novel patatin domain-containing protein, is induced by fasting and glucocorticoids: ectopic expression of desnutrin increases triglyceride hydrolysis. J Biol Chem 279, 47066-47075, doi:10.1074/jbc.M403855200 (2004).
18 Lampidonis, A. D., Rogdakis, E., Voutsinas, G. E. & Stravopodis, D. J. The resurgence of Hormone-Sensitive Lipase (HSL) in mammalian lipolysis. Gene 477, 1-11, doi:10.1016/j.gene.2011.01.007 (2011).

36
19 Bonet, M. L., Oliver, P. & Palou, A. Pharmacological and nutritional agents promoting browning of white adipose tissue. Biochim Biophys Acta, doi:10.1016/j.bbalip.2012.12.002 (2012).
20 Palou, A., Picó, C., Bonet, M. L. & Oliver, P. The uncoupling protein, thermogenin. Int J Biochem Cell Biol 30, 7-11 (1998).
21 Froy, O. The circadian clock and metabolism. Clin Sci (Lond) 120, 65-72, doi:10.1042/CS20100327 (2011).
22 Liang, H. & Ward, W. F. PGC-1alpha: a key regulator of energy metabolism. Adv Physiol Educ 30, 145-151, doi:10.1152/advan.00052.2006 (2006).
23 Puigserver, P. et al. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 92, 829-839 (1998).
24 Picó, C., Palou, M., Priego, T., Sánchez, J. & Palou, A. Metabolic programming of obesity by energy restriction during the perinatal period: different outcomes depending on gender and period, type and severity of restriction. Front Physiol 3, 436, doi:10.3389/fphys.2012.00436 (2012).
25 Mairata, F. B. Técnicas instrumentales en bioquímica y biología. Universitat de les Illes Balears Colección de materiales didácticos ,105. edn, (Palma, Marzo 2003).
26 online, P. PROTOCOLS, 2013). 27 SIGMA-ALDRICH. Reactivo Bradford, 2013). 28 online, P. SDS-PAGE, 2013). 29 Picó, C. et al. The intake of physiological doses of leptin during lactation in rats
prevents obesity in later life. Int J Obes (Lond) 31, 1199-1209, doi:10.1038/sj.ijo.0803585 (2007).
30 Boström, P. et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 481, 463-468, doi:10.1038/nature10777 (2012).




























