Embed Size (px)
Citation preview

30
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Anatomia foliar de Psychotria viridi...
ANATOMIA FOLIAR DE Psychotria viridisRUIZ & PAV. (RUBIACEAE)
MARIANA MARTINS DA COSTA QUINTEIRO1
DANIEL CABRAL TEIXEIRA1
MOEMY GOMES DE MORAES1,2
JANIE GARCIA DA SILVA1
1. Departamento de Biologia Geral, Instituto de Biologia, Universidade Federal Fluminense.2. Caixa Postal 100.436, CEP24001-970, Niterói, RJ, Brasil ([email protected]).
RESUMO: Quinteiro, M.M.C.; Teixeira, D.C.; Moraes, M. G.; Silva, J. G. Anatomia foliar de Psychotriaviridis Ruiz & Pav. (Rubiaceae). Revista Universidade Rural: Série Ciências da Vida, Seropédica, RJ:EDUR, v. 26, n2 p. 30-41, jul-dez,2007. Psychotria viridis Ruiz & Pav. é uma Rubiaceae nativa da regiãoAmazônica que vem sendo cultivada em outras regiões por ser constituinte da Ayahuasca, uma bebidaritualística e com propriedades medicinais. Neste trabalho foi analisada a anatomia das folhas de P. viridis,de plantas cultivadas na zona de amortecimento do Parque Estadual dos Três Picos, município de NovaFriburgo (RJ). Foram observadas as seguintes características: pecíolo circular, passando a plano-convexona porção distal, apresenta colênquima angular, parênquima com idioblastos contendo ráfides e feixe vascularcolateral em arco com extremidades fletidas para o interior. A lâmina foliar é dorsiventral, hipostomática, comestômatos paracíticos e paralelocíticos. Tricomas tectores foram encontrados na face abaxial. Foram obser-vadas domácias na superfície abaxial da nervura central. Substâncias fenólicas, lipídeos e alcalóides foramdetectados nas células parenquimáticas do mesofilo, nas células ao redor da domácia e na epiderme doscoléteres. As estruturas morfológica e anatômica das folhas de P. viridis correspondem a de plantas mesófitas,estão em concordância com as características gerais do gênero Psychotria e da família Rubiaceae. Distin-guem-se, entretanto, por apresentar, na face adaxial, células epidérmicas contendo uma drusa por célula edomácias infundibuliformes diferentes das descritas até o momento para a família.Palavras-chave: Ayahuasca, domácia, Rio de Janeiro
ABSTRACT: Quinteiro, M.M.C.; Teixeira, D.C.; Moraes, M. G.; Silva, J. G. Leaf anatomy of Psychotriaviridis Ruiz & Pav. (Rubiaceae). Revista Universidade Rural: Série Ciências da Vida, Seropédica, RJ:EDUR, v. 26, n2 p. 30-41, jul-dez,2007. Psychotria viridis Ruiz & Pav. is a native Rubiaceae from AmazonRegion which has been cultivated abroad due its use as a component of Ayahuasca, a ritualistic drink withmedicinal properties. This work was analyzed the leaf anatomy of P. viridis plants growing in the region ofParque Estadual dos Três Picos, Nova Friburgo, Rio de Janeiro State. The following anatomical features werefound: round petiole to plan-convex on distal portion, with angular collenchyma, parenchyma with raphideidioblasts, colateral vascular bundle, arch-shaped whose borders are internally curved. Dorsiventral,hypostomatic leaves, with paracytic and parallelocytic stomata. Trichomes were found on the lower epidermis.Domatia were found on midvein from lower epidermis. Phenolic compounds, lipids and alkaloids were presentalong parenchyma mesophyll cells, around domatia and colleters epidermis. P. viridis leaf morphology andanatomy match with mesophytic plants features and are in agreement to general attributes of the genusPsychotria and Rubiaceae family. However it distinguishes by showing upper epidermis with one druse ineach cell and infundibuliformis domatia, which are different from the ones already described for other Rubiaceaespecies.Key words: Ayahuasca, domatia, Rio de Janeiro.
INTRODUÇÃO
Psychotria viridis Ruiz & Pav.,pertencente à família Rubiaceae, é umaplanta nativa da Amazônia que vem sendoutilizada e cultivada em diferentes regiõesdo Brasil e do mundo, por ser constituintede uma bebida ritualística e medicinal, a
Ayahuasca. O chá é utilizado em rituaisrel igiosos, inicialmente dos povosindígenas da Região Amazônica e,posteriormente, em diversos outros, comoSanto Daime, União do Vegetal eBarquinha. A bebida é feita da decocçãodo caule do cipó Banisteriopsis caapi(Spruce) Morton. ex. Briesb.

31
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Quinteiro, M.M.C.; et al.
(Malpighiaceae) com as folhas do arbustoPsychotria viridis (Rubiaceae), sendocaracter izada como alucinógena(MCKEENA, 1996).
Algumas variações no fei tioconvencional da bebida são comumenteencontradas. P. viridis é a espécie maiscomumente utilizada, produzindo um tipode experiência mais alucinógena e do tipovisionária. Outras espécies utilizadas sãoPsychotr ia carthagenensis Jacq. ePsychotria leiocarpa Mart. (LABATE,2004). Os alucinógenos são uma classede agentes psicofarmacognósticoscapazes de causar profundas mudançasno pensamento, no humor, na emoção ena percepção. Os estados experienciaisproduzidos por sua ingestão sãosemelhantes aos sonhos, estadosmeditativos da mente e aos estadospsicóticos, embora não possam seridentificados exatamente como nenhumdestes estados (CALLAWAY et al., 1999).A bebida, portanto, é considerada comoenteógena, o que significa farmaco-logicamente uma ou mais substânciasatuando em conjunto, capazes de alterara consciência sem provocar alucinaçõese que despertam aspectos religiosos nosindivíduos (RUCK et al., 1992).
As folhas de P. viridis contêm umpotente alcalóide alucinógeno de açãorápida, a N, N-dimetiltriptamina (DMT),enquanto o caule de B. caapi contém osderivados beta-carbolínicos: harmina,tetrahidroarmina e harmalina comoprincipais alcalóides. Esses são potentesinibidores seletivos da enzima monoaminaoxidase-A (MAO-A), para a qual aserotonina e, provavelmente, outrastriptaminas, incluindo a DMT, são ossubstratos preferenciais. A DMT não éativa quando ingerida oralmente, maspode se apresentar oralmente ativoquando na presença do inibidor periféricoda MAO e essa interação é a base da açãoalucinógena do chá. Enquanto a harminae a harmalina inativam reversivelmente a
MAO-A, a tetrahidroharmina inibefracamente a recaptação de serotoninaem sítios pré-sinápticos. Juntas, ambasas ações aumentam as at iv idadesserotoninérgicas central e periférica, alémde facilitar a psicoatividade da DMT(CALLAWAY et al., 1999).
A maioria dos trabalhos sobre P.viridis são relacionados aos seus aspectosfarmacológicos, como os de Mckeena et al.(1984), Callaway et al. (1994) e Mckeena(1996), o que deixa lacunas na literatura,principalmente no que se refere àtaxonomia, anatomia e conservação destaespécie. Estudos sobre os efeitos dabebida sugerem remissão de desordenspsíquicas prévias, incluindo ansiedade,depressão e dependências alcoólica equímica, como a da cocaína e do “crack”(GROB et al ., 1996). Por suaspropriedades, P. viridis e B. caapidespertam o interesse de pessoas dediversas partes do mundo, fato que poderepresentar um risco para as plantas,devido à exploração indiscriminada desterecurso.
A família Rubiaceae apresenta 637gêneros e aproximadamente 10.700espécies sendo, portanto, um dos maioresgrupos entre as Angiospermas(ROBBRECHT, 1988). A estrutura foliar desuas espécies caracteriza-se pelapresença de hipoderme, estômatos dotipo paracítico e paralelocítico, epidermeadaxial papilosa, mesofilo geralmentedorsiventral e ocorrência freqüente dedomácias (METCALFE & CHALK, 1979).Outra característica típica é a presençahabitual de coléteres nas estípulas(LERSTEN, 1974). Várias espécies derubiáceas vêm sendo estudadas quantoa seus aspectos anatômicos, inclusive asdo gênero Psychotria, que possui cercade 1000 espécies (STEYERMARK, 1974).
Dentre os estudos anatômicos nogênero, destacam-se os de anatomia foliarde Psychotria nuda (Cham. & Schltdl)Wawra e Psychotria leiocarpa Cham. &

32
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Anatomia foliar de Psychotria viridi...
Schltdl (VIEIRA et al., 1992), Psychotriastenocalyx Mül l . Arg. e Psychotriatenuinervis Müll. Arg. (VIEIRA & GOMES,1995, GOMES et al., 1995), Psychotriavelloziana Benth. (CUNHA & VIEIRA,1993-1997) e Psychotria suterella Müll.Arg. (BARROS et al. , 1997) todasespécies ocorrentes na Mata Atlântica.
O conhecimento sobre a biologiadas plantas e as variedades usadasconstitui uma etapa importante para omelhor entendimento de característicaspeculiares da bebida. Pesquisas sobre aestrutura anatômica de folhas destaespécie podem servir como subsídiospara estudos taxonômicos, propiciar seudiagnóstico como matéria prima paraelaboração de medicamentos, além decontribuir para o conhecimento de suarelação com o ambiente no qual sedesenvolve. Neste trabalho foi descrita aanatomia foliar de P. viridis, visando obterdados para o melhor manejo econhecimento desta espécie.
MATERIAL E MÉTODOS
Folhas completamente desenvol-vidas, provenientes do terceiro nó caulinar,foram coletadas de plantas que sedesenvolvem na zona de amortecimentodo Parque Estadual dos Três Picos,município de Nova Friburgo (RJ), emaltitude de 700 m, clima mesotérmicosempre-úmido, com temperatura médiade 17,8o C (LIMA & GUEDES-BRUNI,1994). Estípulas presentes nos ápicescaulinares foram também coletadas. Umramo fértil foi herborizado e a exsicatadepositada no Herbário do JardimBotânico do Rio de Janeiro, sob o númerode registro RB 401.374.
O material foi fixado em FAA50(JOHANSEN, 1940). Os cortes foramfeitos à mão livre e corados com Azul deAstra (1%) e Safranina (0,5%)(BUKATSCH, 1972). Cortes feitos em
material recém coletado foramsubmetidos aos testes histoquímicosSudam III (SASS, 1951) para detecção delipídeos e Dragendorff para alcalóides(YODER & MAHLBERG, 1976).Substâncias fenólicas foram detectadascom sulfato ferroso em formol(JOHANSEN, 1940). Cristais de oxalatode cálcio foram identi f icados pelasolubil idade em ácido clorídrico einsolubi l idade em ácido acét ico(STRASBURGER, 1986).
Fragmentos da porção medianadas folhas foram dissociados pelo métodode Jeffrey (JOHANSEN, 1940) e coradoscom safranina (1%) para visualização daepiderme. Todos os procedimentos foramfeitos de acordo com a metodologiadescrita em Kraus & Arduim (1997).
As estruturas anatômicas foramanalisadas através de secções trans-versais, longitudinais e das epider-mesdissociadas da lâmina foliar, pecíolo eestípulas em microscópio óptico Olympus,modelo CX40 e fotografadas em fotomi-croscópio Olympus CX30. Os desenhosesquemáticos foram realizados comauxílio de câmara clara acoplada aomicroscópio estereoscópico, modeloOlympus SZX12, seguindo o diagrama deMetcalfe & Chalk (1950).
A classificação dos estômatos foifeita de acordo com Metcalfe & Chalk(1979), a do sistema vascular de acordocom LEAF ARCHITECTURE WORKINGGROUP (1999) e a descr ição dasdomácias seguiu os modelos analisadospor Barros (1959; 1961; 1962).
RESULTADOS E DISCUSSÃO
As folhas de Psychotria viridispossuem pecíolo circular em sua parteproximal e plano-convexo em direção àparte distal, tendo a parte adax ialprojeções laterais na região distal (Figura1B). Seu limbo é do tipo lanceolado, combase cuneada e ápice agudo (Figura 1A),
33
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Quinteiro, M.M.C.; et al.
é flexível e apresenta coloração verde-escura. Estas últimas características sãotípicas de espécies encontradas emambientes úmidos e sombreados(DICKINSON, 2000). O padrão devenação é do tipo broquidódromo,caracterizado pela união das nervuraslaterais, formando arcos proeminentespróximos à margem (Figura 1A). Asnervuras secundárias são levementeascendentes, algumas opostas, outrasalternas, formando arcos que não atingemas margens (Figura 1A). Estascaracterísticas são similares as de P. nudae P. leiocarpa (VIEIRA et al., 1992).
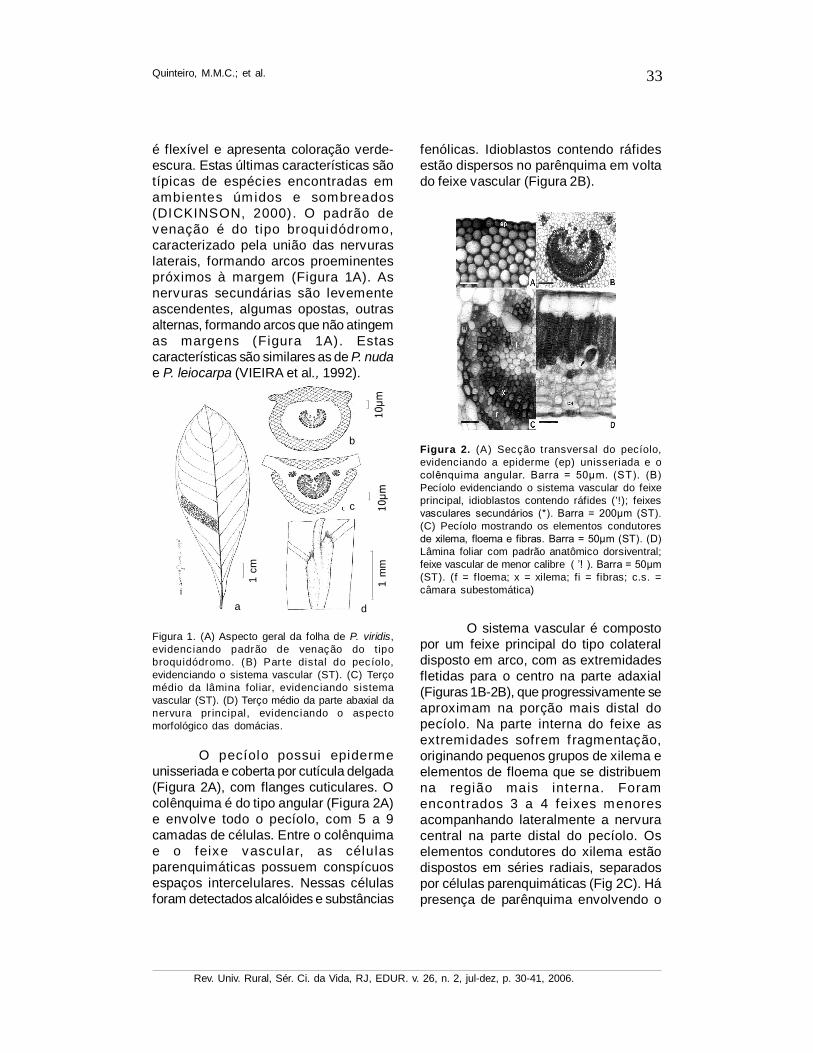
O pecíolo possui epidermeunisseriada e coberta por cutícula delgada(Figura 2A), com flanges cuticulares. Ocolênquima é do tipo angular (Figura 2A)e envolve todo o pecíolo, com 5 a 9camadas de células. Entre o colênquimae o feixe vascular, as célulasparenquimáticas possuem conspícuosespaços intercelulares. Nessas célulasforam detectados alcalóides e substâncias
Figura 1. (A) Aspecto geral da folha de P. viridis,evidenciando padrão de venação do t ipobroquidódromo. (B) Parte dis tal do pecíolo,evidenciando o sistema vascular (ST). (C) Terçomédio da lâmina foliar, evidenciando sistemavascular (ST). (D) Terço médio da parte abaxial danervura principal, evidenciando o aspectomorfológico das domácias.
1 cm
a
10µm
b
10µm
c
1 m
m
d
fenólicas. Idioblastos contendo ráfidesestão dispersos no parênquima em voltado feixe vascular (Figura 2B).
O sistema vascular é compostopor um feixe principal do tipo colateraldisposto em arco, com as extremidadesfletidas para o centro na parte adaxial(Figuras 1B-2B), que progressivamente seaproximam na porção mais distal dopecíolo. Na parte interna do feixe asextremidades sofrem fragmentação,originando pequenos grupos de xilema eelementos de floema que se distribuemna região mais interna. Foramencontrados 3 a 4 feixes menoresacompanhando lateralmente a nervuracentral na parte distal do pecíolo. Oselementos condutores do xilema estãodispostos em séries radiais, separadospor células parenquimáticas (Fig 2C). Hápresença de parênquima envolvendo o
Figura 2. (A) Secção transversal do pecíolo,evidenciando a epiderme (ep) unisseriada e ocolênquima angular. Barra = 50µm. (ST). (B)Pecíolo evidenciando o sistema vascular do feixeprincipal, idioblastos contendo ráfides (’!); feixesvasculares secundários (*). Barra = 200µm (ST).(C) Pecíolo mostrando os elementos condutoresde xilema, floema e fibras. Barra = 50µm (ST). (D)Lâmina foliar com padrão anatômico dorsiventral;feixe vascular de menor calibre. ( ’! ). Barra = 50µm(ST). (f = f loema; x = xilema; fi = fibras; c.s. =câmara subestomática)

34
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Anatomia foliar de Psychotria viridi...
feixe central. Em direção à parte distaldo pecíolo, encontram-se fibras queprogressivamente envolvem o feixesvascular principal e os de pequeno porte.Estes refletem a mesma organização dofeixe principal, com exceção dasextremidades fletidas para o centro.
A forma do feixe principal dopecíolo observada é comum em outrasrubiáceas. P. nuda (VIEIRA, 1986-1988),Coussarea graciliflora Benth. & Hook(TAVARES & VIEIRA, 1994), Rudgeamacrophylla Benth. (MANTOVANI et al.,1995) e Tocoyena bullata (Vell.) Mart.(VIEIRA, 1986-1988) igualmente possuemfeixe central em forma de arco, comextremidades fortemente encurvadas. EmT. bullata (VIEIRA, 1986-1988) ocorreainda o mesmo padrão de aproximaçãoprogressiva de suas extremidades. Em R.macrophylla (MANTOVANI et al., 1995) foitambém observado que as extremidadesdo feixe principal sofrem fragmentação naregião da medula, originando pequenosgrupos de xi lema e f loema semorganização definida.
A lâmina foliar de P. viridis possuimesofilo dorsiventral (Figura 2D). Aepiderme é unisseriada, contendo paredepericlinal externa convexa. A paredepericlinal interna é reta, assim como asparedes antic l inais. As célulasepidérmicas, em v ista f rontal, sãopoligonais (Figuras 3A-B). Na face adaxial,as células apresentam 4-7 lados edistingue-se por apresentar uma drusa porcélula (Figura 3A), além de cristaisprismáticos. Na face abaxial, as célulaspossuem 3-6 lados, são menores,possuem formato mais variado (Figura3B). A cutícula aparece estriada emambas as faces, sendo mais espessa naface adaxial.
A folha é hipostomática comestômatos são do tipo paracítico eparalelocítico (Figura 3B). De acordo comMetcalfe & Chalk (1979) estas sãocaracterísticas gerais comuns ao gênero
e família em questão. Os estômatosencontram-se no mesmo nível oulevemente acima das demais célulasepidérmicas (Figura 3C). Tricomastectores unisseriados com 1 a 11 célulasestão presentes (Figura 3D),principalmente na região das nervurasprincipal e secundárias, onde ocorremdomácias.
Figura 3. (A) Vista frontal da face adaxial daepiderme da lâmina foliar dissociada evidenciandouma drusa (’!) por célula. Barra = 50µm. (B) Vistafrontal da face abaxial da epiderme da lâmina foliardissociada demonstrando estômatos paracíticos eparalelocíticos. Barra = 50µm. (C) Lâmina foliar comestômatos levemente ac ima das célulasepidérmicas (*). Barra = 50µm (ST). (D) Nervuraprincipal evidenciando tricomas tectores. Barra =50µm (ST). (E) Estípulas mostrando coléteres dotipo padrão; idioblastos contendo ráfides (*). Barra= 50µm (ST). (F) Detalhe do coléter. Barra = 200µm(ST). (t = tricomas tectores; fv = feixe vascular dasestípulas)
As características encontradas naepiderme de P. viridis são comuns emRubiaceaase, iniciando pelos tios deestômatos, sendo o paracítico o maiscomum na família (KOCSIS et al., 2004).Os tricomas presentes apresentampequena diversidade estrutural,constituindo outro atributo comum em

35
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Quinteiro, M.M.C.; et al.
Rubiaceae (ROBBRECHT, 1988). A formapoligonal das células epidérmicas,estruturada por paredes anticlinais retasa levemente curvas, foi verificada emoutras espécies do mesmo gênero quevivem no interior da mata, como P. nudae P. leipocarpa (VIEIRA et al., 1992), P.velloziana (CUNHA & VIEIRA, 1993/1997), ou em espécies de outros gênerose habitats semelhantes, como Rudgeadecipens Müll. Arg e Rudgea macrophyllaBenth. (MANTOVANI et al., 1995) eespécies de Bathysa (GOMES et al.,2000).
O mesofilo possui uma camadade parênquima paliçádico composto porcélulas justapostas (Figura 2D),portadoras de lipídeos, substânciasfenólicas e alcalóides. O parênquimalacunoso possui 3-5 estratos de células(Figura 2D). Os feixes vasculares depequeno porte possuem bainhaparenquimática sem projeções. Foramobservados idioblastos com feixes deráfides no mesofilo. O arranjo do mesofiloé típico de mesófitas, encontrado emoutras espécies de Rubiaceae (VIEIRA etal., 1992, CUNHA & VIEIRA, 1993/1997,GOMES et al., 2000, KOCSIS et al., 2004).Este arranjo com uma proporção deparênquima paliçádico signif icativapossibilita resulta em maior condutividadedo dióxido de carbono, principalmente emplantas de sombra (IVANOVA &P’YANKOV, 2002).
A organização do sistemavascular da nervura principal reflete a dopecíolo nos terços basal, mediano e apicalda folha. O mesmo padrão de aproxima-ção progressiva das extremidades do fei-xe principal pode ser observado (Figura 1C).
As três categorias de cristais deoxalato de cálcio encontrados em P. viridis(cristais prismáticos, ráfides e drusas) sãofreqüentes em outras Rubiaceae. Ráfidese drusas são mais comumente referidasno gênero Psychotria. Lersten (1974) citaque a ocorrência de ráfides é típica da
subfamília Rubioideae, a qual pertence ogênero Psychotria e que a presença dedrusas é freqüente em espécies Sul-americanas. Entretanto, não foramencontrados relatos sobre a ocorrência deuma drusa por célula epidérmica nogênero Psychotria, como observado nestaespécie. O papel da ocorrência de cristaisde oxalato de cálcio nos v egetaispermanece por ser esclarecido. Narevisão feita por Nakata (2003) foi relatadoque, enquanto a formação de feixes deráfides tem duas funções prováveis,regulação de cálcio intracelular e defesada planta contra herbivoria, a formação dedrusas está estritamente envolvida naprimeira função. Webb (1999) discute quepouco se sabe a respeito dos mecanismosem que as células controlam a morfologiados cristais, sendo consenso, entretanto,que estas formas estão sobre controlegenético, sendo os tipos e padrõessuficientemente estáveis para seremempregados com fins taxonômicos, o quecorrobora a afirmação de Lersten (1974)sobre a utilização da distribuição doscristais com tal f inalidade no gêneroPsychotria.
As estípulas de P. viridis sãointerpeciolares, o que é uma característicada família Rubiaceae e caem depois daemergência da nova folha. Seu mesofiloé homogêneo, possui epiderme comtricomas uni e pluricelulares, portadoresde lipídeos e papilas na face abaxial.
Na superf ície adaxial dasestípulas, em meio aos tricomas ocorremcoléteres com formato cônico a deltóide(Figuras 3E-F), compostos por um eixocelular parenquimático rodeado por umacamada de células epidérmicas em formade paliçada (Figura 3F). Foi verificada umaleve constrição na base dos coléteres.Foram detectados alcalóides, lipídeos esubstâncias fenól icas nas célulasepidérmicas e nas células parenqui-máticas do eixo, sendo nestas, verificada

36
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Anatomia foliar de Psychotria viridi...
a presença de idioblastos contendo ráfides(Figura 3F).
A estrutura anatômica doscoléteres está de acordo com a descritapor Lersten (1974), podendo serclassificados como padrão, sendo este otipo mais comum, segundo Thomas(1991). A ocorrência de coléteres éfreqüente em Rubiaceae, como discuteVieira (1986). Metcalf & Chalk (1979)descrevem essas estruturas comomulticelulares, glandulares, compostaspor eixo de suporte ovalado/alongado ecélulas epidérmicas glandulares muitoalongadas, que secretam mucilagementre as estípulas fundidas e o primórdiofol iar. Além do tipo padrão, sãoencontrados também coléteres den-dróides e do tipo pincel (LERSTEN, 1974;THOMAS, 1991), sendo estes encon-trados em espécies de Rubiaceae quepossuem nódulos bacterianos nas folhas.Em P. viridis, que possui coléteres do tipopadrão, não foi encontrado nenhumnódulo foliar.
Coléteres padrão também foramencontrados na superfície adaxial dasestípulas de Simira glazovii, Simira pikiae Simira rubra (Rubiaceae), todasapresentando constrição na base docoléter (KLEIN et al., 2004). Em P. viridis,os coléteres ocorrem em meio aoindumento formado por tricomaspluricelulares, assemelhando-se aoscoléteres de Oxypetalum sp.(Apocynaceae) analisados por Swartz &Furlan (2002).
Além de lubrificarem as gemas, amucilagem secretada pelos coléteres temsido relacionada ao desenvolvimento demicrorganismos associados aos vegetais.Lersten & Horner (1967) relataram apresença de bactérias na mucilagemsecretada pelos coléteres de Psychotriabacteriophila Val. que, posteriormente,causariam nódulos na região dosestômatos das folhas adultas. Miller et al.1983 enfatizaram que mucilagem
secretada é essencial ao desenvolvimentodas colônias de bactérias para posteriorinfecção e formação dos nódulos emPsychotria kirkii Hiern. Posteriormente,Van Oevelen et al. (2003) ressaltaram paraa mesma espécie, que a manutenção dascolônias de bactérias, garantida pelamucilagem secretada pelos coléteres, éessencial para o desenvolv imentoadequado e, conseqüentemente para asobrevivência de P. kirkii. Klein et al.(2004) relataram a presença demicrorganismos embebidos em secreçãopróximo à parede exterior das células doscoléteres em Simira glaziovii (K. Schum.)Steyerm, e Simira rubra (Mart.) Steyerm.(Rubiaceae).
Além de otimizarem as condiçõespara o desenvolvimento das gemascaulinares, as substâncias produzidaspelos coléteres são importantes paragarantir a interação com microrganismos,que por sua vez, podem induzir e/ouotimizar a produção de metabólitossecundários (BENNETT &WALLSGROVE, 1994; SCHULZ et al.,2002), o que pode influenciar na qualidadeda Ayahuasca.
Outra característica comum emRubiaceae é a presença de domáciasfol iares (BARROS, 1959, 1962;METCALFE & CHALK, 1979). Em P. viridisForam observ adas domácias nasuperfície abaxial da folha (Figura 1A).Não foram encontradas informaçõesprévias sobre sua observação em P.viridis, mesmo em trabalhos como os deBarros (1959, 1962) que relatam aocorrência e classificação das domáciasem mais de 600 espécies da famíliaRubiaceae. Em P. viridis, estas estruturassão popularmente conhecidas como“velas”, sendo uti l izadas em seureconhecimento no campo. As domáciasocorrem, principalmente, na metade distaldas folhas e seu tamanho varia com adimensão da folha. Foi observado,entretanto, que em indivíduos muito

37
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Quinteiro, M.M.C.; et al.
jovens elas não estão presentes ouaparecem em pequena quantidade.Martinez-Solis et al. (1993), relataram quedomácias de espécies de Rhamnaceaeocorrem independente-mente do tamanhoou idade da folha.
As domácias encontradas em P.viridis apresentam-se como pequenassaliências alongadas, distribuídas aolongo da nervura principal sendo, portanto,nervais (Figuras 1D-4). Assemelham-se,parcialmente, às domácias em forma “debolsa” descritas em Barros (1961). Essassão relatadas como estruturas que seinsinuam sob a nervura principal, najunção com as nervuras secundárias, sobforma de pequenas bolsas de reentrânciano tecido da folha, podendo ser pilosa ounão (BARROS, 1961). Em P. viridis, noentanto, estas estruturas aparecemelevadas e não como reentrâncias (Figura4), assemelhando-se às encontradas emum gênero de Rhamnaceae por Martinez-Solis et al. (1993), que descreveram-nascomo estruturas infundibuli formes,abertas, com nume-rosos tricomas,maiores do que os encontrados em outraspartes da folha. Algumas espécies, comoCinnamomum camphora (L.) J. Presl,podem apresentar até quatro tiposdiferentes de domácia (NISHIDA et al.,2006). Estes autores discutem quedomácias com cara-cterísticas inter-mediárias ou mistas têm sido relatadas,entretanto são escassos os estudos quecontemplam a anatomia e ontogênesedessas estruturas.
As domácias de P. viridisapresentam células epiteliais justapostas,não lignificadas, cobertas por cutícula, querevestem a parte interna da cavidade (Fig4). Foram detectados alcalóides esubstâncias fenólicas nas camadas decélulas parenquimáticas ao redor destacavidade. C. camphora, Nishida et al.(2006), verificaram que a diferença entreas diferentes formas de domácias estavaassociada às diferenças na estrutura do
mesofilo e ao número e tamanho dascélulas epiteliais da cavidade. Estesautores verificaram também a associaçãoentre a ocupação dos diferentes tipos dedomácia e o hábito alimentar de diferentestaxa de ácaros.
A presença de domácias emplantas tem sido objeto de inúmerosestudos com enfoque ecológico, pois suapresença nos órgãos dos vegetais temsido associada às relações comartrópodes, principalmente ácaros eformigas (WALTER, 1996; AGRAWAL &KARBAN, 1997). Folhas que possuemdomácias têm maior concentração deácaros predadores e, portanto, estasestruturas atuam como defesa constitutivacontra outros artrópodes herbívoros(WALTER, 1996). Agrawal & Karban(1997) relataram que, quando domáciasartificiais são acrescentadas em plantasde algodão, diversas espécies de ácarospredadores aumentaram em abundância,enquanto a população de ácarosherbívoros decresceu e a produtividadedas plantas aumentou, quandocomparados aos controles. As domáciassão essenciais ao mirmecofitismo porproverem, além do alimento, espaço paraoviposição (FIALA & MASCHWITZ, 1992;
Figura 4. Terço médio da nervura centralevidenciando o padrão anatômico da regiãomediana da domácia; células epiteliais (’! ) ;cavidade da domácia (*). Barra = 200µm (ST).

38
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Anatomia foliar de Psychotria viridi...
ROMERO & BENSON, 2004; TILLBERG,2004; TEPE et al., 2007).
Não foram observados ácaros ouqualquer outro animal no interior dasdomácias de P. viridis para se estabeleceralgum vínculo entre estes organismos,contudo seria recomendado um estudocom este objetivo. Entretanto, foiverificada a visita freqüente de formigasnos indivíduos P. viridis. Metcalfe & Chalk(1979), ressaltam que a maioria dosautores acredita que as domácias sãoestruturas que surgem independente-mente da presença de hóspedes, sendoutil izadas por eles quando já estãoformadas. Essas características asdiferenciam das galhas, que possuem umnúmero muito grande de células, sendoestas maiores que as adjacentes, e sãodesenvolv idas após contato com ohospedeiro.
P. viridis apresenta característicasanatômicas relacionadas às espéciesmesófitas, como: mesofilo dorsiventralcom uma camada de parênquimapaliçádico, estômatos no mesmo nível oulevemente acima das demais célulasepidérmicas. As estruturas morfológica eanatômica estão em concordância com ascaracterísticas gerais do gêneroPsychotria e da famíl ia Rubiaceae.Entretanto, distingue-se de outrasespécies de Psychotria por apresentarepiderme da face adaxial contendo umadrusa por célula e domácias infun-dibuliformes. Essas características sãoúteis para a determinação da espécie,auxiliando na análise farmacognóstica,garantindo a obtenção de drogas paramedicamentos e a qualidade da bebida.Não são encontradas, até o momento,referências sobre a anatomia dessaespécie em seu bioma original.Observações de indivíduos crescentes naFloresta Amazônica podem ser úteis paraverificação de modificações anatômicasem plantas ocorrentes na Mata Atlântica.
AGRADECIMENTOS
Agradecemos aos dirigentes das igrejasdo Santo Daime, Caparelli e Carlos, pelofornecimento de material necessário paraesse trabalho.
REFERÊNCIAS BIBLIOGRÁFICAS
AGRAWAL, A.A.; KARBAN R. Domatiamediate plant-arthropod mutualism.Nature, v.387, p.562-563, 1997.
BARROS, M.A.A. Ocorrência dasDomácias na Família Rubiaceae. Anais daEscola Superior de Agricultura Luiz deQueiroz, v.16, p.331-337, 1959.
BARROS, M.A.A. Ocorrência dasdomácias nas Angiospermas. Anais daEscola Superior de Agricultura Luiz deQueiroz, v.18, p.113 –146, 1961.
BARROS, M.A.A. Ocorrência dasdomácias nas Rubiáceas. Anais da EscolaSuperior de Agricultura Luiz de Queiroz,v.19 p.1- 21, 1962.
BARROS, C.F.; CALLADO, C.H.; CUNHA,M.; COSTA, C.G.; PUGIALLI, H.R.L.;MARQUETE, O.; MACHADO, R.D.Anatomia ecológica e micromorfologiafoliar de espécies de floresta montana nareserva ecológica de Macaé de cima. In:Serra de Macaé de Cima: DiversidadeFlorística e Conservação em MataAtlântica. LIMA, H.C.; GUEDES-BRUNI,R.R. (eds.). Rio de Janeiro : Editora JardimBotânico do Rio de Janeiro, 1997. p.226 -236.
BENNETT, R.N.; WALLSGROVE, R.M.Secondary metabolites in plant defencemechanisms. New Phytologist, v.127,p.617-633, 1994.
BUKATSCH, F. Bemerkungen zurDoppelfärbung Astrablau-Safranin.

39
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Quinteiro, M.M.C.; et al.
Mikrokosmos, v.61, n.8, p.225, 1972.CALLAWAY, J.C.; AIRAKSINEN, M.M.;MCKEENA, D.J.; BRITO, G.S.; GROB,C.S. Platelet serotonin uptake sitesincreased in drinkers of ayahuasca.Psychopharmacology, v.116, p.385-387,1994.
CALLAWAY, J.C.; MCKENNA, D.J.;BRITO, G.S.; GROB, G.S.; RAYMON, L.P.;POLAND, R.E.; ANDRADE, E.N.; MASH,D.C. Pharmacokinet ics of Hoascaalkaloids in healthy humans. JournalEthnopharmacology, v.65, n.3, p.243-246,1999.
CUNHA, M.; VIEIRA; R.C. Anatomia foliarde Psychotr ia vel loziana Benth.Rodriguesia, v.45-49, n.71-75, p.39-50,1993-1997.
DICKINSON, W. C.. Integrative PlantAnatomy. San Diego : Hartcourt AcademicPress, 2000, 533 p.
FIALA, B.; MASCHWITZ, U. Domatia asmost important adaptations in the evolutionof myrmecophytes in the paleotropical treegenus Maracanga (Euphorbiaceae). PlantSystematics and Evolution, v.180, p.53-64,1992.
GOMES, D.M.S.; MANTOVANI, A.;VIEIRA, R. C. Anatomia fol iar dePsychotria tenuinervis Muell. Arg. ePsychotr ia stenocalix Muell . Arg.(Rubiaceae). Arquivos de Biologia eTecnologia, v.38, n.1, p.15–33, 1995.
GROB, C.S.; MCKEENA, D.J.;CALLAWAY, J.C.; BRITO, G.S.; NEVES,E.S.; OBERLANDER, G.; SAIDE, O.L.;LABIGALINI, E.; TALCA, C.; MIRANDA,C.T.; STRASSMAN, R.J.; BOONE, K.B.Human psychopharmacology of hoasca,a plant hallucinogen used in ritual context
in Brazil. The Journal of Nervous & MentalDisease, v.184, p.86-94, 1996.
IVANOVA, L.A., P´YANKOV, V.I. Structuraladaptation of the leaf mesophyll toshading. Russian Journal of PlantPhysiology, v.49, n.3, p.419–431, 2002.
JOHANSEN, D.A. Plant Microtechnique.New York : Mac Graw-Hill Book Inc, 1940,523p.
KLEIN, D.E.; GOMES, V.M.; SILVA NETOS.J.; CUNHA, M. The structure of colletersin several species of Simira (Rubiacee).Annals of Botany, v.94, p.733-740, 2004.
KOCSIS, M.; DARÓK J.; BORHIDI, A.Comparative leaf anatomy andmorphology of some neotropicalRondeletia (Rubiaceae) species. PlantSystematics and Evolution, v.248, p.205–218, 2004.
KRAUS, J.E.; ARDUIM, M. Manual básicode métodos em morfologia vegetal.Seropédica: Editora Universidade Rural,1997, 198p.
LABATE, B.C. A reinvenção do uso daayahuasca nos centros urbanos, SãoPaulo: Mercado de Letras/Fapesp, 2004.533 p.
LEAF ARCHITECTURE WORKINGGROUP. Manual of Leaf Architecture –Morphological descript ion andcategorization of dicotyledonous and net-veined monocotyledonous angiosperms.Washington DC: Smithsonian Institution,1999, 65p.
LERSTEN, N.R.; HORNER, H.T.Development and structure of bacterialleaf nodules in Psychotria bacteriophila

40
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Anatomia foliar de Psychotria viridi...
Val. (Rubiaceae). Journal of Bacteriology,v.94, n.6, p.2027-2036, 1967.
LERSTEN, N.R. Morphology anddistribution of colleters and crystals inrelation to the taxonomy and bacterial leafnodule symbiosis of Psychotria(Rubiaceae). American Journal of Botany,v.61, n.9, p.973-981, 1974.
LIMA, M.P.M.; GUEDES-BRUNI, R. R.Reserva Ecológica de Macé de Cima,Nova Friburgo, RJ: Aspectos florísticosdas espécies vasculares, Editora JardimBotânico do Rio de Janeiro: Rio deJaneiro, 1994. v.1, p.17-27.
MANTOVANI A.; GOMES, M.; GOMES,D.M.S.; VIEIRA, R.C. Anatomia foliar deRudgea decipiens Müll .Arg. e R.Macrophylla Benth. (Rubiaceae). ActaBotanica Brasilica, v.9, n.2, p.247-261,1995.
MARTINEZ–SOLIS, I.; IRANZO, J.;ESTRELLES, E.; IBARS, A. M. Leafdomatia in the section Alaternus (Miller)DC. of the genus Rhamnus(Rhamnaceae). Botanical Journal of theLinnean Society, v.112, p.311-318, 1993.
MCKEENA, D.J.; TOWERS, G.H.N.;ABBOT, F. Monoamine oxidase inhibitorsin South American hallucinogenic plants:tryptamine and carboline constituents ofayahuasca. Journal ofEthnopharmacology, v.11, p.189-206,1984.
MCKEENA, D.J. Plant hallucinogens:springboards for psychotherapeutic drugdiscovery. Behavioural Brain Research,v.73, p.109-116, 1996.
METCALF, C.R.; CHALK, L. Anatomy ofDicotyledons. Oxford : Clarendon Press,1950, 1500p.
METCALF, C.R.; CHALK, L. Anatomy ofDicotyledons, 2nd ed., Oxford: OxfordUniversity Press, v. 1, 1979. 288p.
NAKATA, P.A. Adv ances in ourunderstanding of calcium oxalate crystalformation and function in plants. PlantScience, v.164, p.901-909, 2003.
NISHIDA, S.; TSUKAYA, H.; NAGAMASU,H.; NOZAKI; M. A comparative study onthe anatomy and development of differentshapes of domatia in Cinnamomumcamphora (Lauraceae). Annals of Botany,v.97, p.601-610, 2006.
ROBBRECHT, E. Tropical woodyRubiaceae. Opera Botanica Belgica, v.1,p.272-277, 1988.
ROMERO, G.Q.; BENSON, W.W. Leafdomatia mediate mutualism betweenmites and a tropical tree. Oecologia, v.140,p.609–616, 2004.
RUCK, C.; WASSON, R.G.; KRAMRISCH,S.; OTT, J. Persephone’s Quest. New Haven: Yale University Press, 1992, 257 p.
SASS, J.E. Botanical Microtechnique. 2nded., Ames : Iowa State College Press,1951, 391 p.
SCHULZ, B.; BOYLE, C.; DRAEGER, S.;RÖMMERT, K.; KROHN, K. Endophyticfungi: a novel source of novel biologicallyactive secondary metabolites. MycologicalResearch, v.106, p.996-1004, 2002.
STEYERMARK, J.A. Rubiaceae. In: Florade Venezuela. LASSER, T. (ed). Caracas:Edition Especial del Instituto Botânico,1974, 2070p.
STRASBURGER, E. Tratado de botânica.7th ed. Barcelona: Marin, 1986, 1098p.

41
Rev. Univ. Rural, Sér. Ci. da Vida, RJ, EDUR. v. 26, n. 2, jul-dez, p. 30-41, 2006.
Quinteiro, M.M.C.; et al.
SWARTZ, E.A.; FURLAN, A. Coléteresfol iares de Oxypetalum R. Br.(Asclepioideae, Apocynaceae) – aspectosultraestruturais e anatômicos úteis àtaxonomia das espécies do Paraná(Brasil). Acta Biologica Paraaense, v.31,n.1,2,3,4, p.79-97, 2002.
TAVARES, E.S.; VIEIRA, R.C. Anatomiafoliar de Coussarea meridionalis (Vell.)Muell. Arg. e Coussarea graciliflora Benth.& Hook (Rubiaceae). Bradea, v.6, n.39,p.320-330, 1994.
TEPE, E, J.; VINCENT, M.A.; WATSON,L. E. Stem diversity, cauline domatia, andthe evolution of ant–plant associations inPiper sect. Macrostachys (Piperaceae).American Journal of Botany, v.94, p.1-11,2007.
THOMAS, V. Structural, functional andphylogenetic aspects of the colleter.Annals of Botany, v.68, p.287-305, 1991.
TILLBERG, C.T. Cordia gerascanthus(Boraginaceae) produces stem domatia.Jounal of Tropical Ecology, v.20, p.355-357, 2004.
VAN OEVELEN, S.; DE WACHTER, R.;ROBBRECHT, E.; PRINSEN, E. Inductionof a crippled phenotype in Psychotria(Rubiaceae) upon loss of the bacterialendophyte. Bulgarian Journal of PlantPhysiology, Special issue, p.242–247,2003.
VIEIRA, R.C. Tocoyena bullata (Vell.) Mart.(Rubiaceae). Anatomia foliar. Rodriguesia,v.64/66, n.38/40, p.33-39, 1986/1988.
VIEIRA, R.C.; GOMES, D.M.S. Superfícieda lâmina foliar de Psychotria nuda(Cham. & Schltdl.) Wawra, P. leiocarpaCham. & Schldtl., P. stenocalyx müll. Arg.
e P. tenuinervis Müll Arg. (Rubiaceae).Acta Botanica Brasilica, v.92, n.2, p.263-270, 1995.
VIEIRA, R.C.; GOMES, D.M.S.; FERRAZ,C.L.A. Anatomia foliar de Psychotria nudaWawra e Psychotria leiocarpa Mart.(Rubiaceae). Hoehnea, v.19, n.1/2, p.185-195, 1992.
WALTER, D.E. Living on leaves: mites,tomenta, and leaf domatia. Annual Reviewof Entomology, v.41, p.101-114, 1996.
WEBB, M.A. Cell-Mediated Crystallizationof Calcium Oxalate in Plants. The PlantCell, v.11, p.751-761, 1999.
YODER, L.R.; MAHLBERG, P.G.Reactions of Alkaloid and histochemicalindicators in Laticifers and specializedParenchyma cells of Catharanthus roseus(Apocynaceae). American Journal ofBotany, v.63, n.9, p. 1167-1173, 1976.
























![Clase 19-20-21-22-23-24 PAV RIGIDOS[resumen]](https://img.pdfslide.net/doc/110x75/634868ad09e11fdd740abfe0/clase-19-20-21-22-23-24-pav-rigidosresumen.jpg)




