Embed Size (px)
Citation preview

lable at ScienceDirect
Journal of Archaeological Science 36 (2009) 513–519
Contents lists avai
Journal of Archaeological Science
journal homepage: ht tp: / /www.elsevier .com/locate/ jas
Ancient DNA supports lineage replacement in European dog gene pool:insight into Neolithic southeast France
M.F. Deguilloux a,*, J. Moquel b, M.H. Pemonge a, G. Colombeau b
a Universite Bordeaux 1, UMR 5199 PACEA, Laboratoire d’Anthropologie des Populations du Passe, bat. B8, av. des Facultes, 33405 Talence cedex, Franceb Universite Bordeaux 1, UMR 5199 PACEA, Institut de Prehistoire et Geologie du Quaternaire, bat. B18, av. des Facultes, 33405 Talence cedex, France
a r t i c l e i n f o
Article history:Received 28 July 2008Received in revised form 1 October 2008Accepted 2 October 2008
Keywords:Ancient DNAMitochondrialDomesticationFranceChassean cultureCanis familiaris
* Corresponding author. Tel.: þ33 (0)5 40 00 37 38E-mail address: [email protected]
0305-4403/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.jas.2008.10.011
a b s t r a c t
We report palaeogenetic analysis of domesticated dog (Canis familiaris) remains excavated from threearchaeological sites from southeast France and dating from Middle Neolithic. Ancient DNA analysis wasattempted on teeth and bone samples taken from 11 dogs. Three 266-base-pair fragments of themitochondrial genome Hypervariable Region I (HVR-I) could be retrieved and revealed two haplotypesbelonging to HVR-I lineage C. These three sequences were compared to the sequences of Swedish andItalian Neolithic dogs and permitted to confirm that clade C was largely represented all over WesternEurope during this period. One haplotype defined in Neolithic French dog was observed for the first timein Canis mtDNA, underlining the loss of mitochondrial diversity in Europe since the Neolithic. Finally,these results point out mitochondrial lineage replacement in Europe, since lineage C represents only 5%of extant European dogs. Altogether, these results support the proposition that palaeogenetic studies areessential for the reconstruction of the past demographic history and the domestication process of dogs.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
The domesticated dog Canis familiaris L. is descended from thegrey wolf Canis lupus L. (Clutton-Brock, 1995). While this theory hasbeen well studied and established from genetics as well asmorphological data (Clutton-Brock, 1995; Vila et al., 1997), thelocation, time and mode of dog domestication are still muchdebated. For instance, the origin of domestic dogs from a singlewolf population or from multiple populations at different timesremains unclear. The first dog attested fossils have been found inSpain (w17,000 14C YA, Altuna et al., 1985; Vigne, 2005), Germany(w14,000 14C YA, Nobis, 1979, 1986) and Russia (13,000–17,000 14CYA, Sablin and Khlopachev, 2002) and consisted in robust wolf-sized dogs. At the opposite, smaller dog remains were discovered inMiddle East, in Natoufian sites (w12,000 14C YA, Tchernov andValla, 1997). These findings would favour independent domestica-tion events. This would also indicate that founders were recruitedfrom a large and varied wolf population (Clutton-Brock, 1995),explaining why living dogs present high phenotypic and geneticvariability. This extreme phenotypic diversity of dogs could alsoimply occasional interbreeding with wild wolf populations,following founding events (Vila et al., 1997).
; fax: þ33 (0)5 40 00 25 45.rdeaux1.fr (M.F. Deguilloux).
All rights reserved.
Archaeological evidence suggests an origin of the domestic dogduring the last glaciations. Recent genetic studies focusing on themitochondrial genome Hypervariable Region 1 (HVR-1) of extantdog have attempted to test this hypothesis. They point towards theexistence of several dog clades (Tsuda et al., 1997; Vila et al., 1997;Leonard et al., 2002; Savolainen et al., 2002), which are interpretedas indicative of multiple and independent origins from multiplewolf matrilines. The molecular time estimates of the dog–wolfdivergence vary from 76,000–130,000 YA for Vila et al. (1997) to15,000–40,000 YA for Savolainen et al. (2002). The analysis of intra-clade genetic diversity and the distribution of unique haplotypesled Savolainen et al. (2002) to favour an East Asian origin for thethree main dog clades, defined as haplogroups (Hg) A, B and C, allemerging about 15,000 YA.
Humans have significantly influenced the genetics of wolvesand dogs during the last centuries. Wolves were highly persecuted,leading to the extinction or at least the bottlenecking of mostpopulations (Vila et al., 1999; Randi et al., 2000; Flagstad et al.,2003). Dogs breeding and distribution were highly influenced byhuman, and the elaboration of the main dog breeds over the lastcentury had important consequences for the make-up of the doggene pool. Genetic studies have suggested that most of the currentbreeds may represent a recent radiation from a common stock(Parker et al., 2004). Moreover, the combined analyses of paternallyinherited Y chromosome markers with maternally inheritedmitochondrial DNA and biparentally inherited autosomal micro-satellite markers in domestic dogs have demonstrated a sex bias in

M.F. Deguilloux et al. / Journal of Archaeological Science 36 (2009) 513–519514
the origin of breeds, with fewer males than females contributinggenetically (Sundqvist et al., 2006). Therefore, these historic humaninfluences must have strongly changed genetic signatures. Onlystudies based on ancient DNA (aDNA) can give direct insight intothe composition of the ancient dog and wolf gene pool, before theconfounding factor of artificial selection and persecution.
This palaeogenetic approach has already been successfullyapplied to canid remains. In 2005, Verginelli et al. succeeded in thegenetic analysis of five ancient Italian canid remains (ages rangingfrom 15,000 to 3,000 YA) and found that mitochondrial sequenceswere highly diverse. Indeed, the five ancient mitochondrialsequences encompassed the three major clades of extant dogsequences (i.e. clades A/B/C and I/II/IV according to Savolainenet al., 2002 and Verginelli et al., 2005, respectively). Phylogeneticanalysis pointed out relationships between the ancient Italiansequences and geographically widespread extant dog matrilinesand extant wolf matrilines of mainly East European origin. Theauthors concluded that European wolves played a role in the originof the three major clades and that multiple independent domesti-cation events took place. This conclusion contrasts with the moresimplistic scenario proposing an East Asian origin for the threemajor clades (Savolainen et al., 2002). More recently, Malmstromet al. (2005, 2008) carried out a palaeogenetic analysis on Neolithicand Medieval dog remains from Scandinavia. Surprisingly, theyidentified sequences corresponding to the Hg A and C which aresupposed to originate from Asia (Savolainen et al., 2002), instead oflineage D, that is restricted to Europe and consequently presentedas the potential result of central or northern Europe domesticationevent. This led to the conclusion that modern data are essential toresolve the number and the antiquity of domestication events,however, they are inadequate to uncover their geographic origin.
Altogether, both studies highlighted the need to use aDNA tobuild a better understanding of past canid population genetics.Specifically, aDNA analyses give a direct measurement of theimpact of human activities on the haplogroup frequencies amongdomesticates. The present study contributes to this promisingapproach, by describing novel aDNA sequences obtained fromMiddle Neolithic dogs originating from a site in southeast of Franceand belonging to the Chassean culture. The three French Neolithic
Table 1Samples analysed in this study, their excavated location, age, and amplification success (
Sample name Code Archaeological site Individual information
Berriac dog A1 BerA1 Berriac Adult 1 of sepulchre A
Berriac dog A2 BerA2 Berriac Adult 2 of sepulchre A
Berriac dog A3 BerA3 Berriac Young individual of sepulchre A
Berriac dog B BerB Berriac Individual 1 of deposit B
Villeneuve-Tolosane tooth1
VTC1 Villeneuve-Tolosane
Tooth in pit (2.5–3 m depth)
Villeneuve-Tolosane tooth2
VTC2 Villeneuve-Tolosane
Tooth in pit (3.5–4 m depth)
Villeneuve-Tolosane tooth3
VTC3 Villeneuve-Tolosane
Tooth in pit (4.5–5 m depth)
Villeneuve-Tolosane tooth4
VTC4 Villeneuve-Tolosane
Tooth in pit (1.5–2 m depth)
Villeneuve-Tolosane tooth5
VTC5 Villeneuve-Tolosane
Tooth in pit (6–6.5 m depth)
Le Cres dog 1 LC1 Le Cres Adult individual associated with hburial
Le Cres dog 2 LC2 Le Cres Adult in dog dual sepulchre
HVR-I sequences obtained were compared with sequences of 541modern dogs (from Verginelli et al., 2005) and 22 ancient canids(Verginelli et al., 2005; Malmstrom et al., 2008) sequences toprovide novel insight into the ancient gene pool of European dogs.
2. Materials and methods
2.1. Archaeological material
Dog specimens were provided by the Institut National deRecherches Archeologiques Preventives (Table 1). Remains werecollected from three Chassean culture sites in the southeast ofFrance: Berriac (Vaquer, 1998), Villeneuve-Tolosane (Vaquer et al.,1980) and Le Cres (Loison et al., 2004). For reason of biomoleculepreservation, teeth are the sample of choice for palaeogeneticstudies (thought to be the place of better DNA conservation andprotection against contaminations, Ricaud et al., 2004). As far aspossible our analyses were conducted on this type of material.
Remains from the site of Les Plots - Berriac consisted in teethand bones recovered from dogs deposited in two different features(Table 1). The first structure contained the nearly complete remainsof two adults and one young dog and was interpreted as thevoluntary and simultaneous deposit of the three individuals. Thesecond structure contained one complete dog skeleton. The pres-ence of ash and burn marks on the skull, as well as the lack ofcervical vertebrae, both tend to exclude the hypothesis of a funeraldeposit. Teeth or tibias were collected from all four individuals ofBerriac and were submitted to genetic analysis (Table 1).
The sample of ancient dog remains from Villeneuve-Tolosaneconsisted in numerous teeth found in a pit 1.5 m wide and 7.5 mdeep. This feature was interpreted as a rubbish pit and was full ofsediment, ash and faunal remains. Teeth were collected from each0.5 m level, in order to minimize the risk that they belong to thesame individual. Five teeth corresponding to five different levelswere analysed (Table 1).
The last archaeological site, Le Cres of Bezier, consisted of a largeburial site with more than 200 structures encompassing 44 pits (33of which contain human burials). A total of 7 dog individuals wasfound in four large circular pits, four of them presenting clear
7/9¼ 7 positive amplifications over 9 tested).
Sample analysed Age Amplification success
HVR-I B HVR-I D
PositivePCR
PositivePCR
Inferior canine (right) 4200–4000BC
0/6 0/6
Distal epiphysis tibia (right) 4200–4000BC
0/6 0/6
First superior premolar (right) 4200–4000BC
0/6 0/6
Tibia (left) 4200–4000BC
0/6 0/6
Superior canine (right) 4000–3800BC
7/9 6/10
Second superior premolar (right) 4000–3800BC
6/8 5/10
Deciduous superior premolar(left)
4000–3800BC
1/12 6/12
Second inferior premolar (right) 4000–3800BC
0/8 0/8
Inferior canine (right) 4000–3800BC
0/8 0/8
uman Inferior canine (left) 4200–4000BC
0/8 0/8
First superior molar (right) 4200–4000BC
0/8 0/8
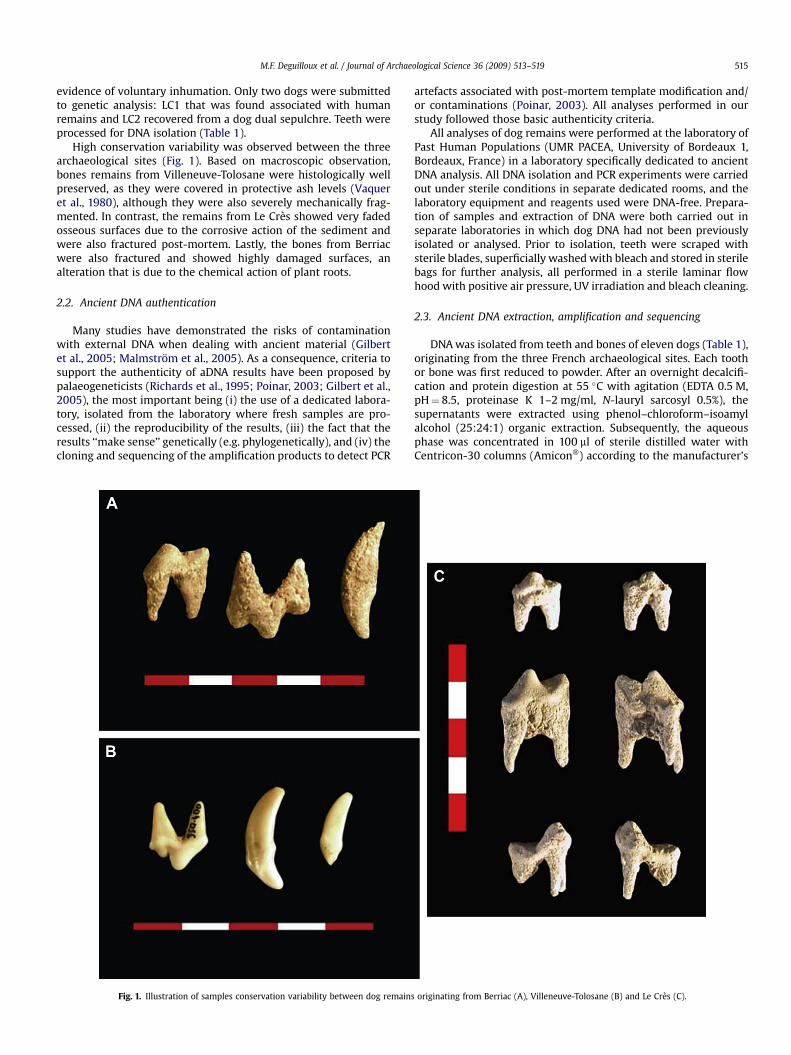
M.F. Deguilloux et al. / Journal of Archaeological Science 36 (2009) 513–519 515
evidence of voluntary inhumation. Only two dogs were submittedto genetic analysis: LC1 that was found associated with humanremains and LC2 recovered from a dog dual sepulchre. Teeth wereprocessed for DNA isolation (Table 1).
High conservation variability was observed between the threearchaeological sites (Fig. 1). Based on macroscopic observation,bones remains from Villeneuve-Tolosane were histologically wellpreserved, as they were covered in protective ash levels (Vaqueret al., 1980), although they were also severely mechanically frag-mented. In contrast, the remains from Le Cres showed very fadedosseous surfaces due to the corrosive action of the sediment andwere also fractured post-mortem. Lastly, the bones from Berriacwere also fractured and showed highly damaged surfaces, analteration that is due to the chemical action of plant roots.
2.2. Ancient DNA authentication
Many studies have demonstrated the risks of contaminationwith external DNA when dealing with ancient material (Gilbertet al., 2005; Malmstrom et al., 2005). As a consequence, criteria tosupport the authenticity of aDNA results have been proposed bypalaeogeneticists (Richards et al., 1995; Poinar, 2003; Gilbert et al.,2005), the most important being (i) the use of a dedicated labora-tory, isolated from the laboratory where fresh samples are pro-cessed, (ii) the reproducibility of the results, (iii) the fact that theresults ‘‘make sense’’ genetically (e.g. phylogenetically), and (iv) thecloning and sequencing of the amplification products to detect PCR
Fig. 1. Illustration of samples conservation variability between dog remains
artefacts associated with post-mortem template modification and/or contaminations (Poinar, 2003). All analyses performed in ourstudy followed those basic authenticity criteria.
All analyses of dog remains were performed at the laboratory ofPast Human Populations (UMR PACEA, University of Bordeaux 1,Bordeaux, France) in a laboratory specifically dedicated to ancientDNA analysis. All DNA isolation and PCR experiments were carriedout under sterile conditions in separate dedicated rooms, and thelaboratory equipment and reagents used were DNA-free. Prepara-tion of samples and extraction of DNA were both carried out inseparate laboratories in which dog DNA had not been previouslyisolated or analysed. Prior to isolation, teeth were scraped withsterile blades, superficially washed with bleach and stored in sterilebags for further analysis, all performed in a sterile laminar flowhood with positive air pressure, UV irradiation and bleach cleaning.
2.3. Ancient DNA extraction, amplification and sequencing
DNA was isolated from teeth and bones of eleven dogs (Table 1),originating from the three French archaeological sites. Each toothor bone was first reduced to powder. After an overnight decalcifi-cation and protein digestion at 55 �C with agitation (EDTA 0.5 M,pH¼ 8.5, proteinase K 1–2 mg/ml, N-lauryl sarcosyl 0.5%), thesupernatants were extracted using phenol–chloroform–isoamylalcohol (25:24:1) organic extraction. Subsequently, the aqueousphase was concentrated in 100 ml of sterile distilled water withCentricon-30 columns (Amicon�) according to the manufacturer’s
originating from Berriac (A), Villeneuve-Tolosane (B) and Le Cres (C).

M.F. Deguilloux et al. / Journal of Archaeological Science 36 (2009) 513–519516
instructions. An extraction blank was systematically co-extractedwith the ancient dog samples during each extraction session, totrace potential cross-contamination between samples. No morethan two ancient samples were co-extracted at the same time.
PCR amplifications were performed on the HypervariableRegion 1 (HVR-I) of the mitochondrial genome. HVR-I region of themtDNA control region (position 15,426–15,692, including primers)was obtained through two overlapping fragments obtained witha set of two primers’ pairs (Verginelli et al., 2005): fragment HVR-IB obtained with primers L15426int and H15555int, fragment HVR-ID with L15529int and H15692int.
The pre-PCR mix was prepared in a DNA-free room in theancient DNA laboratory. PCR amplifications were performed ina total reaction volume of 25 ml containing 6.5 mM MgCl2, 0.4 mMdNTPs, 0.66 mg/ml BSA, 1 mM of each primer, 1�GeneAmp PCRBuffer (Perkin–Elmer), 0.25 ml pure DNA extract and 1.25 UAmpliTaq Gold� (Applied Biosystems). PCR was run for 55 cycles at94 �C for 45 s, 55 �C for 45 s, and 72 �C for 45 s. Three independentblanks were carried out for each set of PCR experiment.
Sequences of each mitochondrial fragment were confirmed byat least two separate amplifications on each DNA extract. As theproducts of ancient DNA amplification generally contain a largenumber of mutations generated by induced damage and Taqpolymerase errors (Handt et al., 1996), the PCR products weresubcloned into bacterial vectors using the Topo TA cloning kit(Invitrogen) according to the manufacturer’s instructions.Screening of colonies was accomplished by PCR, transferring thecolonies into a 40 ml reaction mix made of 67 mM Tris–HCl, 2 mMMgCl2, 1 mM of each M13 forward and reverse universal primer,0.125 mM of each dNTP and 0.75 U of Taq polymerase (Perkin–Elmer). PCR fragments were separated by agarose gel electropho-resis and clones with the expected insert size were selected forsequencing (Genome Express). Sequences from multiple clones ofthe same PCR product were aligned in order to resolve HVR-Iambiguities. Artefacts caused by the amplification of damagedtemplates from a low copy number were carefully and systemati-cally eliminated. The authentic sequences were always deducedfrom the consensus of at least 10 clones, corresponding to at leasttwo different amplification products (except for VTC3 - HVR-I Bfragment). Extractions and PCR amplification blanks were used asnegative controls, in order to detect possible contamination byexternal DNA.
2.4. Phylogenetic analysis
All sequences were aligned using the MEGA3.1 software (Kumaret al., 2004) and all detected sequence variations were verified byvisual assessment in the electrophoregrams. The HVR-I sequencesobtained from ancient French dogs were compared with mtDNAsequences from modern dog, using the data compiled by Verginelliet al. (2005). The database consists of 541 sequences of extant dogsfrom all continents, including 248, 232, 31 and 30 dogs from Asia,Europe, Africa and America, respectively (Okumura et al., 1996; Vilaet al., 1997; Savolainen et al., 2002, 2004). Five mitochondrialsequences from ancient Italian canids (Verginelli et al., 2005) wereincluded, as well as 17 sequences from ancient Scandinavian dogs(Malmstrom et al., 2008). A 230 bp region encompassing position15,458–16,687 was chosen as it was available from all data sources.The sequence NC_002008, corresponding to the first completemtDNA dog sequence published (Kim et al., 1998) was used asa reference for the numbering of polymorphisms, as proposed byPereira et al. (2004).
DNA variants were identified in the 230 bp sequence usingDnaSP v.3 software (http://www.ub.es/dnasp/) and Median-Joining (MJ) networks connecting the ancient and extant dogssequences were constructed using the NETWORK 4.201 software
(www.fluxus-engineering.com) and the median-joining algorithm(using default weights).
3. Results
We attempted to isolate DNA from eleven dog remains fromFrench archaeological sites belonging to the Neolithic Chasseanculture (from 4,200 to 3,800 BC). Two separate mtDNA segmentspermitting the identification of partial HVR-I sequence (266 bp)were successfully amplified and sequenced for three of the 11ancient samples tested, originating from the site of Villeneuve-Tolosane and dated from 4000 to 3800 BC (samples VTC 1, 2 and 3,Table 1). Owing to poor preservation of biomolecules, no PCRproduct was obtained from the remaining samples, despite severalattempts. With regards to the samples originating from the Ville-neuve-Tolosane site, no correlation between depth of samples andDNA conservation was found in our analysis, since teeth found inboth greater and lesser depth yielded no results. This reinforces theidea that different soil microenvironments inside archaeologicalsite lead to variable DNA conservation.
All Villeneuve-Tolosane sequences were deduced from multipleclone alignment, from 10 to 25 clones for each mitochondrialfragment. Sequence ambiguities were resolved by analysis ofmultiple clones and deaminations observed in several clones weresystematically eliminated. All ancient sequences retrieved belongto dog clade C (Fig. 2). Similar sequences were obtained for VTC2and VTC3 and present the Hg C characteristic mutations: 15508T,15526T, 15611C, 15639G and 15650C (Savolainen et al., 2002; Per-eira et al., 2004). This sequence clusters to the central node of thelineage, as shown in the MJ network of clade C (Fig. 3). VTC2 andVTC3 share their HVR-I sequence with numerous individuals fromall continents except America (i.e. 64%, 33% and 3% of Asian,European and African dogs, respectively). This sequence is similarto one Neolithic Italian dog aged of 4,100 BP (Casal Del Dolce, PIC4,Verginelli et al., 2005), and with nine Neolithic dogs from Scandi-navia (Malmstrom et al., 2008). Morphological observationsuggests that VTC2 and VTC3 remains correspond to two distinctindividuals. Indeed, VTC2 premolar shows usury indices corre-sponding to a young individual (at least 1 year old), and VTC3deciduous premolar is cutting suggesting it belonged to a fewmonths old individual. Nevertheless, we cannot exclude that thosedogs were related, a fact that could introduce bias in geneticdiversity calculations.
The sequence obtained for VTC1 presents the same poly-morphisms as VTC2 and VTC3, as well as two additional mutationsat positions 15508 and 15625. The mutation at position 15508 isa feature shared with one ancient Italian wolf (PIC1, aged 15,000BP) from the Late Glacial levels of the Palidoro Upper Palaeolithicrock shelter (Verginelli et al., 2005). As sample VTC1 presents anadditional mutation at position 15625, the sequence appears noveland occupies a derived position.
The star-shaped topology of clade C network suggests an originfrom a single haplotype (Fig. 3). Briefly, clade C encompasses doghaplotypes from Eurasia, America and Africa, as well as numerousancient European dog sequences: one sequence from an ancientItalian dog dated of 15,000 BP, and a total of 15 sequences fromEuropean Neolithic dogs (11 from Scandinavia, 3 from France and 1from Italy). Interestingly, Asian sequences are represented by onlytwo haplotypes, whereas six different haplotypes are encounteredin modern and ancient European samples.
A total of 95.4% of dog sequences compiled in our worldwidedataset belong to haplogroups A, B and C (with 67.8% and 10%belonging to Hg A and Hg C, respectively). Despite the low reso-lution obtained with our short mitochondrial sequence (230 bp)the MJ network constructed for all clades shows the same patternalready described by Savolainen et al. (2002), with Asian
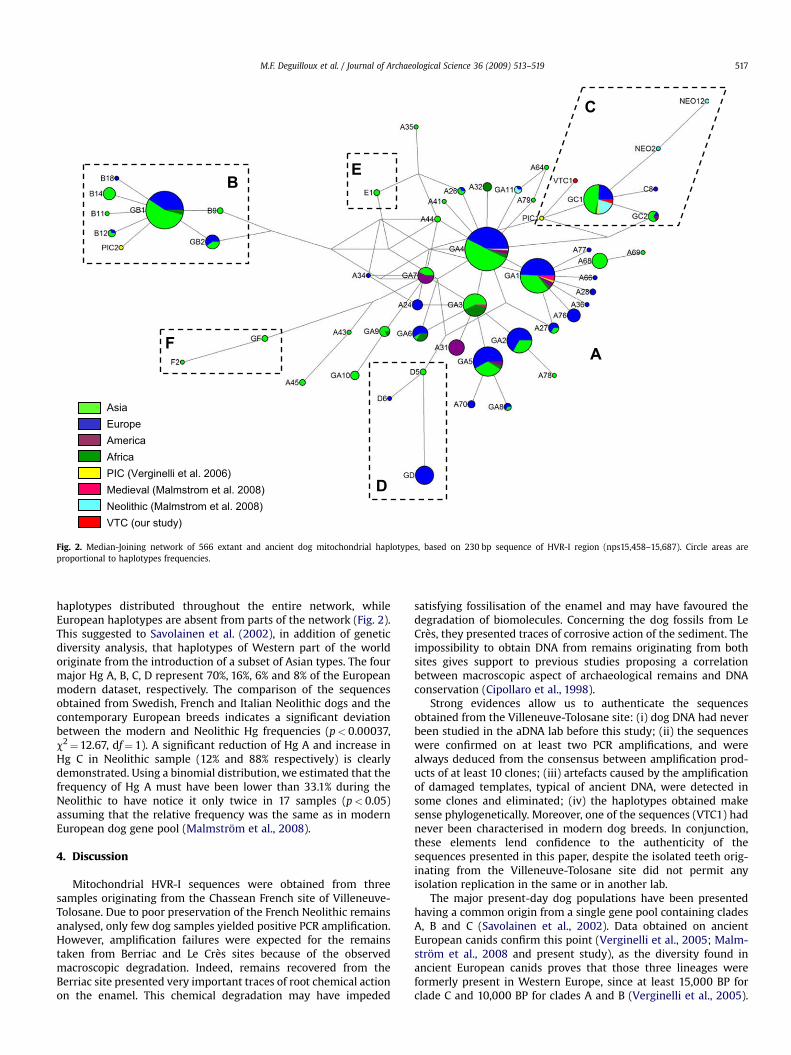
AsiaEuropeAmericaAfricaPIC (Verginelli et al. 2006)Medieval (Malmstrom et al. 2008)Neolithic (Malmstrom et al. 2008)VTC (our study)
F
B
E
D
C
A
Fig. 2. Median-Joining network of 566 extant and ancient dog mitochondrial haplotypes, based on 230 bp sequence of HVR-I region (nps15,458–15,687). Circle areas areproportional to haplotypes frequencies.
M.F. Deguilloux et al. / Journal of Archaeological Science 36 (2009) 513–519 517
haplotypes distributed throughout the entire network, whileEuropean haplotypes are absent from parts of the network (Fig. 2).This suggested to Savolainen et al. (2002), in addition of geneticdiversity analysis, that haplotypes of Western part of the worldoriginate from the introduction of a subset of Asian types. The fourmajor Hg A, B, C, D represent 70%, 16%, 6% and 8% of the Europeanmodern dataset, respectively. The comparison of the sequencesobtained from Swedish, French and Italian Neolithic dogs and thecontemporary European breeds indicates a significant deviationbetween the modern and Neolithic Hg frequencies (p< 0.00037,c2¼12.67, df¼ 1). A significant reduction of Hg A and increase inHg C in Neolithic sample (12% and 88% respectively) is clearlydemonstrated. Using a binomial distribution, we estimated that thefrequency of Hg A must have been lower than 33.1% during theNeolithic to have notice it only twice in 17 samples (p< 0.05)assuming that the relative frequency was the same as in modernEuropean dog gene pool (Malmstrom et al., 2008).
4. Discussion
Mitochondrial HVR-I sequences were obtained from threesamples originating from the Chassean French site of Villeneuve-Tolosane. Due to poor preservation of the French Neolithic remainsanalysed, only few dog samples yielded positive PCR amplification.However, amplification failures were expected for the remainstaken from Berriac and Le Cres sites because of the observedmacroscopic degradation. Indeed, remains recovered from theBerriac site presented very important traces of root chemical actionon the enamel. This chemical degradation may have impeded
satisfying fossilisation of the enamel and may have favoured thedegradation of biomolecules. Concerning the dog fossils from LeCres, they presented traces of corrosive action of the sediment. Theimpossibility to obtain DNA from remains originating from bothsites gives support to previous studies proposing a correlationbetween macroscopic aspect of archaeological remains and DNAconservation (Cipollaro et al., 1998).
Strong evidences allow us to authenticate the sequencesobtained from the Villeneuve-Tolosane site: (i) dog DNA had neverbeen studied in the aDNA lab before this study; (ii) the sequenceswere confirmed on at least two PCR amplifications, and werealways deduced from the consensus between amplification prod-ucts of at least 10 clones; (iii) artefacts caused by the amplificationof damaged templates, typical of ancient DNA, were detected insome clones and eliminated; (iv) the haplotypes obtained makesense phylogenetically. Moreover, one of the sequences (VTC1) hadnever been characterised in modern dog breeds. In conjunction,these elements lend confidence to the authenticity of thesequences presented in this paper, despite the isolated teeth orig-inating from the Villeneuve-Tolosane site did not permit anyisolation replication in the same or in another lab.
The major present-day dog populations have been presentedhaving a common origin from a single gene pool containing cladesA, B and C (Savolainen et al., 2002). Data obtained on ancientEuropean canids confirm this point (Verginelli et al., 2005; Malm-strom et al., 2008 and present study), as the diversity found inancient European canids proves that those three lineages wereformerly present in Western Europe, since at least 15,000 BP forclade C and 10,000 BP for clades A and B (Verginelli et al., 2005).
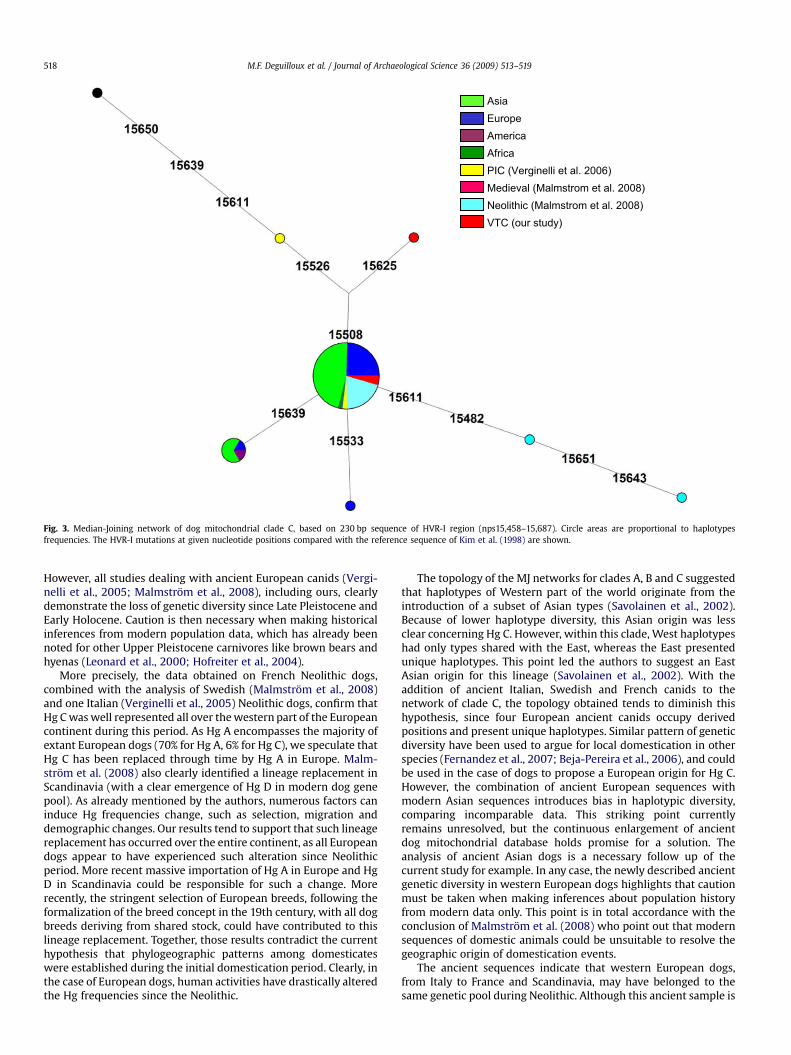
AsiaEuropeAmericaAfricaPIC (Verginelli et al. 2006)Medieval (Malmstrom et al. 2008)Neolithic (Malmstrom et al. 2008)VTC (our study)
Fig. 3. Median-Joining network of dog mitochondrial clade C, based on 230 bp sequence of HVR-I region (nps15,458–15,687). Circle areas are proportional to haplotypesfrequencies. The HVR-I mutations at given nucleotide positions compared with the reference sequence of Kim et al. (1998) are shown.
M.F. Deguilloux et al. / Journal of Archaeological Science 36 (2009) 513–519518
However, all studies dealing with ancient European canids (Vergi-nelli et al., 2005; Malmstrom et al., 2008), including ours, clearlydemonstrate the loss of genetic diversity since Late Pleistocene andEarly Holocene. Caution is then necessary when making historicalinferences from modern population data, which has already beennoted for other Upper Pleistocene carnivores like brown bears andhyenas (Leonard et al., 2000; Hofreiter et al., 2004).
More precisely, the data obtained on French Neolithic dogs,combined with the analysis of Swedish (Malmstrom et al., 2008)and one Italian (Verginelli et al., 2005) Neolithic dogs, confirm thatHg C was well represented all over the western part of the Europeancontinent during this period. As Hg A encompasses the majority ofextant European dogs (70% for Hg A, 6% for Hg C), we speculate thatHg C has been replaced through time by Hg A in Europe. Malm-strom et al. (2008) also clearly identified a lineage replacement inScandinavia (with a clear emergence of Hg D in modern dog genepool). As already mentioned by the authors, numerous factors caninduce Hg frequencies change, such as selection, migration anddemographic changes. Our results tend to support that such lineagereplacement has occurred over the entire continent, as all Europeandogs appear to have experienced such alteration since Neolithicperiod. More recent massive importation of Hg A in Europe and HgD in Scandinavia could be responsible for such a change. Morerecently, the stringent selection of European breeds, following theformalization of the breed concept in the 19th century, with all dogbreeds deriving from shared stock, could have contributed to thislineage replacement. Together, those results contradict the currenthypothesis that phylogeographic patterns among domesticateswere established during the initial domestication period. Clearly, inthe case of European dogs, human activities have drastically alteredthe Hg frequencies since the Neolithic.
The topology of the MJ networks for clades A, B and C suggestedthat haplotypes of Western part of the world originate from theintroduction of a subset of Asian types (Savolainen et al., 2002).Because of lower haplotype diversity, this Asian origin was lessclear concerning Hg C. However, within this clade, West haplotypeshad only types shared with the East, whereas the East presentedunique haplotypes. This point led the authors to suggest an EastAsian origin for this lineage (Savolainen et al., 2002). With theaddition of ancient Italian, Swedish and French canids to thenetwork of clade C, the topology obtained tends to diminish thishypothesis, since four European ancient canids occupy derivedpositions and present unique haplotypes. Similar pattern of geneticdiversity have been used to argue for local domestication in otherspecies (Fernandez et al., 2007; Beja-Pereira et al., 2006), and couldbe used in the case of dogs to propose a European origin for Hg C.However, the combination of ancient European sequences withmodern Asian sequences introduces bias in haplotypic diversity,comparing incomparable data. This striking point currentlyremains unresolved, but the continuous enlargement of ancientdog mitochondrial database holds promise for a solution. Theanalysis of ancient Asian dogs is a necessary follow up of thecurrent study for example. In any case, the newly described ancientgenetic diversity in western European dogs highlights that cautionmust be taken when making inferences about population historyfrom modern data only. This point is in total accordance with theconclusion of Malmstrom et al. (2008) who point out that modernsequences of domestic animals could be unsuitable to resolve thegeographic origin of domestication events.
The ancient sequences indicate that western European dogs,from Italy to France and Scandinavia, may have belonged to thesame genetic pool during Neolithic. Although this ancient sample is

M.F. Deguilloux et al. / Journal of Archaeological Science 36 (2009) 513–519 519
not representative of ancient European population, it is remarkableto encounter the same haplogroup, and even the same sequences,during the Neolithic period, in these distant regions. This could beaccounted by distant domestic species exchanges betweenNeolithic human populations and/or shared breeding practicessince Neolithic times. Numerous French sites from the Chasseanculture show strong evidence of special attention paid to dogburials (Arbogast et al., 1989) and the voluntary burial of dogsappears like a common practice of those populations. This care isoften interpreted as special relationship between Neolithic humanpopulations and domesticated dogs. Since this human–dog rela-tionship appears important during the Neolithic, more compre-hensive palaeogenetic analysis of ancient dog remains belonging tothis period could demonstrate, on the one hand, the extensiveconnection between dog populations and, on the other, the extantlinks and exchanges between Neolithic human populations.
In conclusion, the sequences obtained on ancient European dogshighlight the relevance of palaeogenetic data to: (i) shade light onthe loss of genetic diversity during the domestication process, (ii)reveal lineage(s) replacement(s) through time that can be causedby various factors such as selection, migration and extinction, and(iii) resolve more accurately the origin of the domestication events.The accumulation of genetic data on ancient European dog remainsbegins to bring new elements on dog domestication. The obser-vation of shared Hg C sequences between Italian, French andSwedish Neolithic dogs permits to speculate on a mitochondriallineage replacement since Neolithic times, and suggests sharedbreeding practices since this period. We believe that furtherpalaeogenetic analysis of European and Asian dog remains will givecritical insight into the domestication process of Canis.
The sequences reported in this paper have been deposited intoGenBank (accession numbers EU287460, EU287461, and EU287462).
Acknowledgements
The authors would like to acknowledge H. Duday (UMR PACEA)for providing contacts with archaeologists and zooarchaeologistsand for proofreading the manuscript. We are grateful to J. Vaquer, G.Loison and V. Forrest who supplied dog samples from Berriac,Villeneuve-Tolosane and Le Cres-Beziers and for informations onarchaeological sites, and to D. Chagne for his help in the preparationof the manuscript. Funding for this study was provided by theUniversity of Bordeaux 1 (BQR).
References
Altuna, J., Baldeon, A., Mariezkurrena, K., 1985. Cazadores magdalenienses en laCueva de Errala (Cestona, Pais Vasco). Munibe. Antropologia-Arkeologia 37.
Arbogast, R.M., Blouet, V., Desloges, J., Guillaume, C., 1989. Le cerf et le chien dansles pratiques funeraires de la seconde moitie du Neolithique du Nord de laFrance. Anthropozoologica 3, 37–42.
Beja-Pereira, A., Caramelli, D., Laluza-Fox, C., Vernesi, C., Ferrand, N., Casoli, A.,Goyache, F., Royo, L.J., Conti, S., Lari, M., Martini, A., Ouragh, L., Magid, A.,Atash, A., Zsolnai, A., Boscato, P., Triantaphylidis, C., Ploumi, K., Sineo, L.,Mallegni, F., Taberlet, P., Erhardt, G., Sampietro, L., Bertranpetit, J., Barbujani, G.,Luikart, G., Bertorelle, G., 2006. The origin of European cattle: evidence frommodern and ancient DNA. Proceedings of the National Academy of Sciences 103,8113–8118.
Cipollaro, N., Di Bernardo, G., Galano, G., Galderisi, U., Guarino, F., Angelini, F.,Cascino, A., 1998. Ancient DNA in human bone remains from Pompeii archaeo-logical site. Biochemical and Biophysical Research Communications 247, 901–904.
Clutton-Brock, J., 1995. Origins of the dog: domestication and early history. In:Serpell, J. (Ed.), The Domestic Dog: Its Evolution, Behaviour and Interaction withPeople. Cambridge University Press, Cambridge, pp. 8–20.
Fernandez, H., Hughes, S., Vigne, J.D., Helmer, D., Hodgins, G., Miquel, C., Hanni, C.,Luikart, G., Taberlet, P., 2007. Divergent mtDNA lineages of goats in an earlyNeolithic site, far from the initial domestication areas. Proceedings of theNational Academy of Sciences 103, 15375–15379.
Flagstad, Ø., Walker, C.W., Vila, C., Sundqvist, A.K., Fernholm, B., Hufthammer, A.K.,Wiig, Ø., Koyola, I., Ellegren, H., 2003. Two centuries of the Scandinavian wolfpopulation: patterns of genetic variability and migration during an era ofdramatic decline. Molecular Ecology 12, 869–880.
Gilbert, M.T.P., Bandelt, H.J., Hofreiter, M., Barnes, I., 2005. Assessing ancient DNAstudies. Trends in Ecology and Evolution 20, 541–544.
Handt, O., Krings, M., Ward, R.H., Paabo, S., 1996. The retrieval of ancient humanDNA sequences. American Journal of Human Genetics 59, 368–376.
Hofreiter, M., Serre, D., Rohland, N., Rabeder, G., Nagel, D., Conard, N., Munzel, S.,Paabo, S., 2004. Lack of phylogeography in European mammals before the lastglaciation. Proceedings of the National Academy of Sciences USA 101, 12963–12968.
Kim, K.S., Lee, S.E., Jeong, H.W., Ha, J.H., 1998. The complete nucleotide sequence ofthe domestic dog (Canis familiaris) mitochondrial genome. Molecular Phylo-genetics and Evolution 10 (2), 210–220.
Kumar, S., Tamura, K., Nei, M., 2004. MEGA3: integrated software for molecularevolutionary genetics analysis and sequence alignment. Briefings in bio-informatics 5, 150–163.
Leonard, J.A., Wayne, R.K., Cooper, A., 2000. Population genetics of Ice Age brownbears. Proceedings of the National Academy of Sciences USA 97 (4), 1651–1654.
Leonard, J.A., Wayne, R.K., Wheeler, J., Valadez, R., Guillen, S., Vila, C., 2002. AncientDNA evidence for old world origin of new world dogs. Science 298, 1613–1616.
Loison, G., Fabre, V., Villemeur, I., 2004. Le Cres. Habitats prehistoriques en bordurede l’Orb. Structures domestiques et sepultures du Chasseen ancien. Rapportfinal d’operation. INRAP, Montpellier, p. 304.
Malmstrom, H., Stora, J., Dalen, L., Holmlund, G., Gotherstrom, A., 2005. Extensivehuman DNA contamination in extracts from ancient dog bones and teeth.Molecular Biology and Evolution 22 (10), 2040–2047.
Malmstrom, H., Vila, C., Gilbert, M.T.P., Stora, J., Willerslev, E., Holmlund, G.,Gotherstrom, A., 2008. Barking up the wrong tree: modern northern Europeandogs fail to explain their origin. BMC Evolutionary Biology 8 (71).
Nobis, G., 1979. Der alteste Haushund lebte vor 14,000 Jahren. Umschau 19, 610.Nobis, G., 1986. Die Wildsaugetiere in der Umwelt des Menschen von Oberkassel bei
Bonn und das Domestikationsproblem bei Wolfen im Jungpalaolithikum. Bon-ner Jahrbucher 186, 367–376.
Okumura, N., Ishiguro, N., Nakano, M., Matsui, A., Sahara, M., 1996. Intra- andinterbreed genetic variations of mitochondrial DNA major non-coding regions inJapanese native dog breeds (Canis familiaris). Animal Genetics 27 (6), 397–405.
Parker, H.G., Kim, L.V., Sutter, N.B., Carlson, S., Lorentzen, T.D., 2004. Geneticstructure of the purebred domestic dog. Science 304, 1160–1166.
Pereira, L., Van Asch, B., Amorim, A., 2004. Standardisation of nomenclature for dogmtDNA D-loop: a prerequisite for launching a Canis familiaris database. ForensicScience International 141, 99–108.
Poinar, H.N., 2003. The top 10 list: criteria of authenticity for DNA from ancient andforensic samples. International Congress Series 1239, 575–579.
Randi, E.V., Lucchini, V., Christensen, M.F., Mucci, N., Funk, S.M., Dolf, G.,Loeschcke, V., 2000. Mitochondrial DNA analysis in Italian and East Europeanwolves: detecting the conservation of small population size and hybridization.Conservation Biology 14, 464–473.
Ricaud, F.-X., Keyser-Tracqui, C., Crubezy, E., Ludes, B., 2004. STR-genotyping from humanmedieval tooth and bone samples. Forensic Science International 151 (1), 31–35.
Richards, M.B., Sykes, B.C., Hedges, R.E.M., 1995. Authenticating DNA extracted fromancient skeletal remains. Journal of Archaeological Science 22, 291–299.
Sablin, M.V., Khlopachev, G.A., 2002. The earliest ice age dogs: evidence from Eli-seevichi 1. Current Anthropology 43, 795–799.
Savolainen, P., Zhang, Y., Luo, J., Lundeberg, J., Leitner, T., 2002. Genetic evidence foran east Asian origin of domestic dogs. Science 298, 1610–1613.
Savolainen, P., Leitner, T., Wilton, A.N., Matisoo-Smith, E., Lundeberg, J., 2004. Adetailed picture of the origin of the Australian dingo, obtained from the study ofmitochondrial DNA. Proceedings of the National Academy of Sciences USA 101(33), 12387–12390.
Sundqvist, A.K., Bjornerfeldt, S., Leonard, J.A., Hailer, F., Hedhammar, A., Ellegren, H.,Vila, C., 2006. Unequal contribution of sexes in the origin of dog breeds.Genetics 172, 1121–1128.
Tchernov, E., Valla, F.R., 1997. Two new dogs, and other Natufian dogs, from thesouthern Levant. Journal of Archaeological Science 24, 65–95.
Tsuda, K., Kikkiwa, Y., Yonekawa, H., Tanabe, Y., 1997. Extensive interbreedingoccurred among multiple matriarchal ancestors during the domestication ofdogs: evidence from inter- and intra-species polymorphisms in the D-loopregion of mitochondrial DNA between dogs and wolves. Genes and GeneticSystems 72, 229–238.
Vaquer, J., Clottes, J., Coulardou, J., Gireau, J.P., Rouzaud, F. 1980. Villeneuve-Tolosane,Haute Garonne. Rapport de fouilles.
Vaquer, J., 1998. Les sepultures du Neolithique moyen en France mediterraneenne.In: Guilaine, J. (Ed.), Sepultures d’Occident et genese des megalithismes.Editions Errance, Paris, pp. 167–186.
Verginelli, F., Capelli, C., Coia, V., Musiani, M., Falchetti, M., Ottini, L., Palmirotta, R.,Tagliacozzo, A., De Grossi Mazzorin, I., Mariani-Costantini, R., 2005. Mito-chondrial DNA from Prehistoric canids highlights relationships between dogsand South-East European wolves. Molecular Biology and Evolution 22 (12),2541–2551.
Vigne, J.D., 2005. L’humerus de chien magdalenien de Errala (Gipuzkoa, Espagne) etla domestication tardiglaciaire du loup en Europe. Munibe. Antropologia-Arkeologia 57.
Vila, C., Savolainen, P., Maldonado, J.E., Amorin, I.R., Rice, J.E., Honeycutt, R.L.,Crandall, K.A., Lundeberg, J., Wayne, R.K., 1997. Multiple and ancient origins ofthe domestic dog. Science 276, 1687–1689.
Vila, C., Maldonado, J.E., Wayne, R.K., 1999. Phylogenetic relationships, evolution,and genetic diversity of the domestic dog. The American Genetic Association90, 71–77.





























