Embed Size (px)
Citation preview

Journal of Steroid Biochemistry & Molecular Biology 110 (2008) 284–294
Contents lists available at ScienceDirect
Journal of Steroid Biochemistry and Molecular Biology
journa l homepage: www.e lsev ier .com/ locate / j sbmb
Androgenic profile and genotoxicity evaluation of testosterone propionateand novel synthesized heterocyclic steroids
Hanaa Y. Hanaa, Wagdy K.B. Khalil b, Aida I. Elmakawyb, Gamal A. Elmegeeda,∗
a Hormones Department, National Research Center, 12622 Dokki, Cairo, Egypt
b Cell Biology Department, National Research Center, Dokki, Cairo, Egypta r t i c l e i n f o a b s t r a c t
e andith t
esizeomalred w
rivatited to sig
celnd pyifican≤ 0.0
testede melogicarone p
ever
Article history:Received 2 April 2007Accepted 13 November 2007
Keywords:TestosteroneHeterocyclesGenotoxicityChromosomal aberrationsRAPD–PCRSperm analysis
In this study, we tested thcyclic steroids compared weffects of the novel synthmale mice using chromosMale mice were administepyrimidinoandrostene defor 2 weeks. Results indicapropionate. There were nin both somatic and germtestosterone propionate aderivative 9a caused signsomatic and germ cells (Pthe control and the othersignificant decrease in thdecreased sperm morphodata indicate that testosteno genotoxic activity. How
to a modulation of the differenin the nearly future.1. Introduction
Androgens are a class of steroids that are responsible for theprimary and secondary sex characteristics of the male. In addi-tion, these steroids possess potent anabolic or growth-promotingproperties. Testosterone is the androgenic, anabolic and naturallyoccurring steroid that controls sperm production, sexual drive, andsexual performance throughout a man’s life, as well as influencingmuscle and bone development [1]. It is produced in men by the Ley-dic cells, which are very small species in the testes. Testosteronedeficiency causes sexual dysfunction, infertility, small testes, anddecreased muscle mass and strength [2,3]. Testosterone replace-ment may provide short-term increased sexual drive and enhancedsexual performance but the longer-term effects are not clearlyknown and the risk factors should be considered [4–6].
∗ Corresponding author. Tel.: +20 2 35682070; fax: +20 2 33370931.E-mail address: [email protected] (G.A. Elmegeed).
0960-0760/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.jsbmb.2007.11.006
rogenic activity of three structurally promising novel synthesized hetero-estosterone propionate in male mice. Additionally, the possible genotoxicd heterocyclic steroids in comparison with testosterone propionate on
analysis of somatic and germ cells as well as RAPD–PCR were investigated.ith two doses of testosterone propionate, pyridoandrostene derivative 4b,
ve 9a and thienoandrostene derivative 12 (200 and 400 mg/kg b.w.) dailyhat compounds 4b and 12 have androgenic activity as well as testosteronenificant differences in the frequencies of total chromosomal aberrationsls as well as no alteration in the DNA bands patterns between control,ridoandrostene 4b treated animals. However, the pyrimidinoandrostenet increase in the mean value of total chromosomal aberrations of both1) as well as enhanced the polymorphic bands patterns as compared tocompounds. On the other hand, thienoandrostene derivative 12 induced
an values of chromosomal aberrations in both somatic and germ cells,l abnormalities, increased the sperm count and motility than control. Ourropionate; pyridoandrostene 4b and thienoandrostene derivative 12 have
, pyrimidinoandrostene derivative 9a has genotoxic activity possibly duet expression of the catalyzing enzyme systems which will be investigated
© 2008 Elsevier Ltd. All rights reserved.
Traditionally, sex steroids are recognized as non-genotoxic car-cinogens [7]. To date, few studies have been reported on any aspectof DNA damage caused by testosterone treatment in organs of intactanimals [8,9]. However, controversial results have been reported.Ho and Roy [8] reported that testosterone combined with estro-gen induced a dramatic increase in DNA strand breaks. In contrary,when female rats, converted to ‘male type’ by ovariectomy, treatedwith testosterone for one week, DNA damage caused by hepatocar-cinogen (DL-ZAMI 1305) was completely abolished [9].
The general chemical structure of androgens is based uponthe androstane steroids. The largest number of contributions toandrostane chemistry has dealt with synthesis of compounds towhich an extra or fused heterocyclic ring had been added, thesederivatives have frequently showed hormonal activity of phar-macological interest [10,11]. Since Clinton et al. [12] synthesizedthe pyrazoloandrostane derivative (stanazolol) in 1959 as anabolicandrogenic heterocyclic steroid, a large number of heterocyclicsteroid systems have been synthesized and their androgenic and/orantiandrogenic properties have been examined [13–15]. It has been

istry
H.Y. Hana et al. / Journal of Steroid Biochemreported that the fusion of heterocyclic ring onto a steroid usuallyproduced changes in the endocrinological activity, the most note-worthy effects were apparent in the altered anabolic/androgenicratios and in the development of new types of activity depending onthe position of heterocyclic ring on the steroid nucleus [14]. How-ever, to our knowledge, the genotoxic effects of such compoundshave not been studied.
The main goals of the present study were to elucidate the activ-ity of some novel synthesized heterocyclic steroids as androgenicagents. This can be achieved through investigating the effects ofthe selected newly synthesized steroid derivatives on the weightsof accessory sex organs and on the sperm morphology and counts incomparison with testosterone propionate. Furthermore, this studywas undertaken to verify the possible genotoxic effects of testos-terone and its novel heterocyclic derivatives using chromosomeanalysis of bone marrow and germ cells as well as RAPD–PCR.
2. Materials and methods
2.1. Synthetic methods and spectral data
Testosterone was purchased from Sigma Company, USA. Sol-vents were purified by distillation prior to using. All melting pointswere measured using an electrothermal apparatus and were uncor-rected. The IR spectra were recorded (KBr discs) on a ShimadzuFT-IR 8201 PC spectrophotometer and expressed in cm−1. 1H NMRand 13C NMR spectra were recorded with Jeol instrument (Japan),at 270 and 125 MHz, respectively, in DMSO-d6 as solvent and thechemical shifts were recorded in ppm relative to TMS. Mass spectrawere recorded on a GCMS-QP 1000 Ex spectra mass spectrom-eter operating at 70 eV. Elemental analyses were carried out byMicroanalytical Data Unit at National Research Centre, Giza, Egypt.Reactions were monitored on Merck aluminum thin layer chro-matography (TLC) plates and visualized by UV light (254 nm). Forthe nomenclature of steroid derivatives, we used the definitive rulesfor the nomenclature of steroids published by the Joint Commis-sion on the Biochemical Nomenclature (JCBN) of IUPAC [16,17]. Thestarting compounds 1 (17�-acetoxyandrost-4-en-3-one), 2a and b(2-arylmethylen-17�-acetoxyandrost-4-en-3-one) were preparedaccording to the published procedure [18,19].
2.1.1. General procedure for the preparation of compounds (4aand b) and (5a and b)
To a solution of compound 2a (0.83 g, 2 mmol) or 2b (0.89 g,
2 mmol) in absolute ethanol (30 ml) containing a catalytic amountof piperidine (0.5 ml), either cyanoacetamide 3a (0.16 g, 2 mmol) orcyanothioacetamide 3b (0.20 g, 2 mmol) was added. The reactionmixture, in each case, was refluxed for 5–7 h until all starting mate-rials had disappeared as indicated by TLC. The reaction mixturewas left to cool at room temperature, poured over ice/water mix-ture, neutralized with dilute hydrochloric acid and extracted withdiethyl ether (3× 20 ml). The organic layer, in each case, was driedover calcium chloride. Removal of the solvent in vacuo afforded thecorresponding product, which was crystallized from the appropri-ate solvent.2.1.1.1. 17ˇ-Acetoxy-3′-cyano-2′-oxo-6′-phenyl-1′H-pyrido[4′,5′:3,2]androst-4-ene (4a). Brown powder from1,4-dioxan, yield 0.69 g (72%), mp 154–155 ◦C; IR (�/cm−1):3325 (NH), 3030 (CH-aromatic), 2895, 2854 (CH3, CH2), 2225 (CN),1730 (acetate C O), 1695 (C O), 1590 (C C); 1H NMR (ı ppm):0.95 (s, 3H, 19-CH3), 1.12 (s, 3H, 18-CH3), 2.15 (s, 3H, CH3CO2),3.76–3.84 (m, 1H, C17–�H), 4.32 (s, 1H, NH, D2O-exchangeable),5.73 (s, 1H, C4–H), 7.24–7.43 (m, 5H, Ar–H); MS (EI): m/z (%): 483(M•++1, 58), 422 (M•+-CH3COOH, 23), 253 (C19H25, 100). Calc. for
& Molecular Biology 110 (2008) 284–294 285
C31H34N2O3 (482.62): C, 77.15; H, 7.10; N, 5.80; Found: C, 77.43; H,7.28; N, 6.04%.
2.1.1.2. 17ˇ-Acetoxy-3′-cyano-2′-oxo-6′-(4-methoxyphenyl)-1′H-pyrido[4′,5′:3,2]androst-4-ene (4b). Pale brown crystals fromethanol, yield 0.75 g (74%), mp 192–194 ◦C; IR (�/cm−1): 3332(NH), 3035 (CH-aromatic), 2897, 2850 (CH3, CH2), 2220 (CN),1730 (acetate C O), 1697 (C O), 1635 (C C); 1H NMR (ı ppm):0.93 (s, 3H, 19-CH3), 1.16 (s, 3H, 18-CH3), 2.05 (s, 3H, CH3CO2),3.72 (s, 3H, OCH3), 3.82–3.97 (m, 1H, C17–�H), 4.24 (s, 1H, NH,D2O-exchangeable), 5.97 (s, 1H, C4–H), 6.72 (dd, 2H, J = 9 Hz, Ar–H),7.19 (dd, 2H, J = 9 Hz, Ar–H); 13C NMR: ı = 16.7 (Me-19), 17.4 (C-15),17.8 (CH3CO), 20.6 (Me-18), 20.8 (C-11), 23.3 (C-16), 27.5 (C-12),29.9 (C-8), 31.8 (C-7), 33.8 (C-6), 34.2 (C-1), 36.8 (C-10), 42.6(C-13), 43.9 (C-14), 45.6 (C-9), 80.7 (C-17), 97.7 (C-3′), 117.6 [C-2(C-5′)], 118.4 (C-4), 128.3 (C-6′), 150.5 (C-5), 163.1 (C-2′), 170.9 [C-3(C-4′)], 171.8 (CH3CO), 128.4, 114.0, 161.6 (Ph); MS (EI): m/z (%):512 (M•+, 45), 452 (M•+-CH3COOH, 17), 405 (M•+-C7H7O, 25), 118(C7H4NO,100). Calc. for C32H36N2O4 (512.65): C, 74.97; H, 7.07; N,5.46; Found: C, 75.13; H, 7.29; N, 5.24%.
2.1.1.3. 17ˇ-Acetoxy-3′-cyano-2′-thioxo-6′-phenyl-1′H-pyrido[4′,5′:3,2]androst-4-ene (5a). Yellow crystals from ethanol,yield 0.65 g (67%), mp 212–213 ◦C; IR (�/cm−1): 3334 (NH), 3028(CH-aromatic), 2895, 2850 (CH3, CH2), 2220 (CN), 1732 (acetateC O), 1632 (C C), 1295 (C S); 1H NMR (ı ppm): 0.96 (s, 3H,19-CH3), 1.13 (s, 3H, 18-CH3), 2.07 (s, 3H, CH3CO2), 3.69–3.75 (m,1H, C17–�H), 4.30 (s, 1H, NH, D2O-exchangeable), 5.92 (s, 1H,C4–H), 7.16–7.32 (m, 5H, Ar–H); MS (EI): m/z (%): 498 (M•+, 45),438 (M•+-CH3COOH, 28), 253 (C19H25, 100). Calc. for C31H34N2O2S(498.69): C, 74.66; H, 6.87; N, 5.61; S, 6.43; Found: C, 74.28; H,7.02; N, 5.44; S, 6.65%.
2.1.1.4. 17ˇ-Acetoxy-3′-cyano-2′-thioxo-6′-(4-methoxyphenyl)-1′H-pyrido[4′,5′:3,2]androst-4-ene (5b). Pale brown crystals frommethanol, yield 0.72 g (68%), mp 187–188 ◦C; IR (�/cm−1): 3340(NH), 3030 (CH-aromatic), 2897, 2856 (CH3, CH2), 2220 (CN),1738 (acetate C O), 1630 (C C), 1295 (C S); 1H NMR (ı ppm):0.89 (s, 3H, 19-CH3), 1.02 (s, 3H, 18-CH3), 2.17 (s, 3H, CH3CO2),3.60–3.72 (m, 1H, C17–�H), 3.80 (s, 3H, OCH3), 4.25 (s, 1H, NH,D2O-exchangeable), 5.78 (s, 1H, C4–H), 6.80 (dd, 2H, J = 9 Hz, Ar–H),7.24 (dd, 2H, J = 9 Hz, Ar–H); MS (EI): m/z (%): 528 (M•+, 62), 468(M•+-CH3COOH, 37), 421 (M•+-C7H7O, 25), 107 (C7H7O, 100).Calc. for C32H36N2O3S (528.72): C, 72.69; H, 6.89; N, 5.29; S, 6.06;
Found: C, 72.84; H, 7.16; N, 5.49; S, 5.83%.2.1.2. General procedure for the preparation of compounds (7aand b) and (8a and b)
Either urea 6a (0.12 g, 2 mmol) or thiourea 6b (0.15 g, 2 mmol)was added to a suspension of compound 2a or 2b (2 mmol) insodium ethoxide [0.46 g, 20 mmol sodium metal in 25 ml absoluteethanol]. The reaction mixture, in each case, was heated in a boil-ing water bath for 7 h until all starting materials had disappearedas indicated by TLC, and then evaporated to dryness under reducedpressure. The obtained residue was washed with 10% hydrochlo-ric acid and finally solidified with cold water. The obtained solidproduct was collected by filtration, dried and crystallized from theproper solvent.
2.1.2.1. 17ˇ-Acetoxy-2′-oxo-6′-phenyl-1′H-pyrimidino[4′,5′:3,2]androst-4-ene (7a). Pale brown crystalsfrom ethanol, yield 0.56 g (62%), mp 190–191 ◦C; IR (�/cm−1): 3348(NH), 3010 (CH-aromatic), 2897, 2856 (CH3, CH2), 1735 (acetateC O), 1692 (C O), 1668 (C N), 1630 (C C); 1H NMR (ı ppm): 0.93(s, 3H, 19-CH3), 1.12 (s, 3H, 18-CH3), 2.04 (s, 3H, CH3CO2), 3.83–3.92

istry
containing a catalytic amount of triethylamine (0.5 ml), elemen-
286 H.Y. Hana et al. / Journal of Steroid Biochem
(m, 1H, C17–�H), 4.22 (s, 1H, NH, D2O-exchangeable), 4.87 (s, 1H,C4–H), 7.14–7.36 (m, 5H, Ar–H); MS (EI): m/z (%): 458 (M•+, 42), 398(M•+-CH3COOH, 35), 421 (M•+-C6H5, 36), 198 (C12H10N2O, 100).Calc. for C29H34N2O3 (458.60): C, 75.05; H, 7.47; N, 6.10; Found: C,75.30; H, 7.32; N, 5.94%.
2.1.2.2. 17ˇ-Acetoxy-2′-oxo-6′-(4-methoxyphenyl)-1′H-pyrimidino[4′,5′:3,2]androst-4-ene (7b). Yellow powder frommethanol, yield 0.67 g (69%), mp 173–174 ◦C; IR (�/cm−1): 3353(NH), 3038 (CH-aromatic), 2875, 2856 (CH3, CH2), 1738 (acetateC O), 1703 (C O), 1668 (C N), 1635 (C C); 1H NMR (ı ppm):0.84 (s, 3H, 19-CH3), 1.02 (s, 3H, 18-CH3), 2.22 (s, 3H, CH3CO2),3.68–3.75 (m, 1H, C17–�H), 3.85 (s, 3H, OCH3), 4.27 (s, 1H, NH,D2O-exchangeable), 4.97 (s, 1H, C4–H), 6.75 (dd, 2H, J = 9 Hz, Ar–H),7.16 (dd, 2H, J = 9 Hz, Ar–H); MS (EI): m/z (%): 488 (M•+, 72), 428(M•+-CH3COOH, 47), 381 (M•+-C7H7O, 53), 107 (C7H7O, 100). Calc.for C30H36N2O4 (488.63): C, 73.74; H, 7.42; N, 5.73; Found: C,73.95; H, 7.30; N, 5.52%.
2.1.2.3. 17ˇ-Acetoxy-2′-thioxo-6′-phenyl-1′H-pyrimidino[4′,5′:3,2]androst-4-ene (8a). Brown crystals fromDMF, yield 0.56 g (60%), mp 218–219 ◦C; IR (�/cm−1): 3355 (NH),3035 (CH-aromatic), 2894, 2850 (CH3, CH2), 1727 (acetate C O),1668 (C N), 1630 (C C), 1290 (C S); 1H NMR (ı ppm): 0.98 (s,3H, 19-CH3), 1.16 (s, 3H, 18-CH3), 2.01 (s, 3H, CH3CO2), 3.73–3.82(m, 1H, C17–�H), 4.18 (s, 1H, NH, D2O-exchangeable), 4.90 (s, 1H,C4–H), 7.15–7.32 (m, 5H, Ar–H); MS (EI): m/z (%): 474 (M•+, 32), 414(M•+-CH3COOH, 62), 397 (M•+-C6H5, 36), 214 (C12H10N2S, 100).Calc. for C29H34N2O2S (474.67): C, 73.38; H, 7.21; N, 5.90; S, 6.75;Found: C, 73.54; H, 7.04; N, 6.07; S, 6.93%.
2.1.2.4. 17ˇ-Acetoxy-2′-thioxo-6′-(4-methoxyphenyl)-1′H-pyrimidino[4′,5′:3,2]androst-4-ene (8b). Yellow crystals frommethanol, yield 0.64 g (64%), mp 245–255 ◦C; IR (�/cm−1): 3357(NH), 3042 (CH-aromatic), 2889, 2858 (CH3, CH2), 1723 (acetateC O), 1664 (C N), 1629 (C C), 1293 (C S); 1H NMR (ı ppm):0.80 (s, 3H, 19-CH3), 0.97 (s, 3H, 18-CH3), 2.02 (s, 3H, CH3CO2),3.88–3.95 (m, 1H, C17–�H), 3.76 (s, 3H, OCH3), 4.17 (s, 1H, NH,D2O-exchangeable), 4.88 (s, 1H, C4–H), 6.78 (dd, 2H, J = 9 Hz, Ar–H),7.18 (dd, 2H, J = 9 Hz, Ar–H); MS (EI): m/z (%): 504 (M•+, 27), 444(M•+-CH3COOH, 63), 397 (M•+-C7H7O, 43), 107 (C7H7O,100). Calc.for C30H36N2O3S (504.69): C, 71.39; H, 7.18; N, 5.55; S, 6.35; Found:C, 71.54; H, 6.96; N, 5.49; S, 6.60%.
2.1.3. General procedure for the synthesis of compounds (9a andb)
A mixture of compounds 2a or 2b (2 mmol), guanidinehydrochloride 6c (0.12 g, 2 mmol) and sodium acetate (0.16 g,2 mmol) in absolute ethanol (30 ml) was refluxed for 4 h. The reac-tion mixture was concentrated in vacuo and then triturated withice/water mixture and neutralized with dilute hydrochloric acid.The formed solid product, in each case, was collected by filtration,dried and crystallized from the appropriate solvent.
2.1.3.1. 17ˇ-Acetoxy-2′-imino-6′-phenyl-1′H-pyrimidino[4′,5′:3,2]androst-4-ene (9a). Brown crystals fromethanol, yield 0.63 g (69%), mp 218–217 ◦C; IR (�/cm−1): 3460–3324(2NH), 3042 (CH-aromatic), 2985, 2864 (CH3, CH2), 1728 (acetateC O), 1670, 1645 (2C N), 1618 (C C); 1H NMR (ı ppm): 0.93 (s,3H, 19-CH3), 1.02 (s, 3H, 18-CH3), 2.10 (s, 3H, CH3CO2), 3.82–3.92(m, 1H, C17–�H), 4.28 (s, 1H, NH, D2O-exchangeable), 4.98 (s, 1H,C4–H), 7.18–7.32 (m, 5H, Ar–H), 8.67 (s, 1H, NH, D2O-exchangeable);13C NMR: ı = 16.2 (Me-19), 17.4 (C-15), 17.6 (CH3CO), 20.7 (Me-18),20.9 (C-11), 23.6 (C-16), 27.7 (C-12), 29.5 (C-8), 30.4 (C-1), 30.8(C-7), 33.5 (C-6), 37.8 (C-10), 42.5 (C-13), 43.0 (C-14), 45.4 (C-9),
& Molecular Biology 110 (2008) 284–294
81.2 (C-17), 103.6 [C-2 (C-5′)], 106.4 (C-4), 148.3 (C-6′), 156.5 (C-5),163.4 (C-2′), 165.9 [C-3 (C-4′)], 171.0 (CH3CO), 126.0, 127,7, 128.4,134.6 (Ph); MS (EI): m/z (%): 458 (M•+-1, 28), 399 (M•+-CH3COOH,45), 120 (C6H6N3, 100). Calc. for C29H35N3O2 (459.61): C, 75.78; H,7.67; N, 6.09; Found: C, 75.93; H, 7.48; N, 5.88%.
2.1.3.2. 17ˇ-Acetoxy-2′-imino-6′-(4-methoxyphenyl)-1′H-pyrimidino[4′,5′:3,2]androst-4-ene (9b). Pale brown powderfrom 1,4-dioxane, yield 0.68 g (70%), mp 155–156 ◦C; IR (�/cm−1):3475–3335 (2NH), 3032 (CH-aromatic), 2980, 2846 (CH3, CH2),1720 (acetate C O), 1672, 1640 (2C N), 1615 (C C); 1H NMR(ı ppm): 0.98 (s, 3H, 19-CH3), 1.08 (s, 3H, 18-CH3), 2.13 (s, 3H,CH3CO2), 3.73 (s, 3H, OCH3), 3.85–3.94 (m, 1H, C17–�H), 4.45(s, 1H, NH, D2O-exchangeable), 4.90 (s, 1H, C4–H), 6.76 (dd,2H, J = 9 Hz, Ar–H), 7.14 (dd, 2H, J = 9 Hz, Ar–H), 8.79 (s, 1H, NH,D2O-exchangeable); MS (EI): m/z (%): 486 (M•+-1, 28), 427 (M•+-CH3COOH, 45), 370 (M•+-C7H7O, 62), 120 (C6H6N3, 100). Calc. forC30H37N3O3 (487.64): C, 73.89; H, 7.64; N, 8.61; Found: C, 74.08;H, 7.78; N, 8.42%.
2.1.4. Synthesis of17ˇ-acetoxy-3-(3′-iminobutyronitrile-2′-ylideno)androst-4-ene(11)
To a mixture of 1 (1.65 g, 5 mmol) and ammonium acetate (1.0 g),equimolar amount of 3-imino-butyronitrile (acetonitrile dimmer)10 (0.4 g, 5 mmol) was added, the reaction mixture was heated in anoil bath at 120 ◦C for 30 min. The solid product formed upon cool-ing at room temperature was triturated with ethanol, collected byfiltration dried, and crystallized from DMF to yield 1.42 g (72%) ofcompound 11, yellow crystals, mp 106–107 ◦C; IR (�/cm−1): 3425(NH), 2968, 2855 (CH3, CH2), 1725 (acetate C O), 2220 (CN), 1670(C N), 1625 (C C); 1H NMR (ı ppm): 0.90 (s, 3H, CH3), 1.00 (s, 3H,19-CH3), 1.16 (s, 3H, 18-CH3), 2.08 (s, 3H, CH3), 3.84–3.95 (m, 1H,C17–�H), 5.87 (s, 1H, C4–H), 8.95 (s, 1H, NH, D2O-exchangeable); MS(EI): m/z (%): 394 (M•+, 76), 334 (M•+-CH3COOH, 45), 255 (C19H27,100). Calc. for C25H34N2O2 (394.56): C, 76.10; H, 8.68; N, 7.10;Found: C, 76.35; H, 8.76; N, 7.29%.
2.1.5. Synthesis of 17ˇ-acetoxy-2′-amino-3′-ethanimidoylthieno[4′,5′-3,2]androst-4-ene(12)
To a solution of compound 11 (0.78 g, 2 mmol) in ethanol (30 ml)
tal sulfur (0.064 g, 2 mmol) was added. The reaction mixture washeated under reflux for 4 h until all starting materials had disap-peared as indicated by TLC, The reaction mixture was concentratedunder vacuum and then left to cool at room temperature, pouredover ice/water mixture and neutralized with dilute hydrochloricacid. The formed solid product was filtered off, dried and crystal-lized from ethanol to yield 0.60 g (71%) of compound 12, yellowcrystals, mp 142–143 ◦C. IR (�/cm−1): 3452–3340 (NH, NH2), 2975,2858 (CH3, CH2), 1730 (acetate C O), 1676 (C N), 1630 (C C); 1HNMR (ı ppm): 0.87 (s, 3H, CH3), 0.98 (s, 3H, 19-CH3), 1.13 (s, 3H,18-CH3), 2.15 (s, 3H, CH3CO2), 3.85–3.94 (m, 1H, C17–�H), 5.42(s, 2H, NH2, D2O-exchangeable), 5.92 (s, 1H, C4–H), 8.68 (s, 1H,NH, D2O-exchangeable); 13C NMR: ı = 16.5 (Me-19), 17.4 (C-15),17.6 (CH3CO), 20.6 (Me-18), 20.8 (C-11), 23.5 (C-16), 27.7 (C-12),29.5 (C-8), 31.3 (C-7), 32.4 (C-1), 33.7 (C-6), 39.8 (C-10), 42.5 (C-13), 42.8 (C-14), 43.4 (C-9), 81.7 (C-17), 118.4 (C-4), 123.4 (C-2′),125.3 (C-3′), 132.9 [C-3 (C-4′)], 139.6 [C-2 (C-5′)], 152.5 (C-5), 170.6(CH3CO); MS (EI): m/z (%): 426 (M•+, 76), 366 (M•+-CH3COOH,45), 150 (C7H6N2S, 100). Calc. for C25H34N2O2S (426.62): C, 70.38;H, 8.03; N, 6.56; S, 7.51; Found: C, 70.18; H, 7.96; N, 6.40; S,7.78%.

istry
H.Y. Hana et al. / Journal of Steroid BiochemTable 1Experimental design for the androgenic profile and genotoxicity evaluation of testos-terone and novel synthesized heterocyclic steroids in male mice
Groups Treatment Concentration(mg/kg b.w.)
Time of sampling afterlast day of treatmenta
1 3% Tween
200
3 and 35 days
2 TP4 Compound 4b6 Compound 9a8 Compound 121 3% Tween
4003 TP5 Compound 4b7 Compound 9a9 Compound 12
a After the last treatment five animals of each group were used to assess the chro-mosome analysis of the bone-marrow cells as well as RAPD–PCR assay. After 35 daysof the last treatment the other five animals of each group were used to assess theandrogenicity assay. TP: Testosterone propionate.
2.2. Bioassay
2.2.1. ChemicalsTestosterone propionate was purchased from Misr Co. for
Pharmaceutical Industries, Cairo, Egypt. Solutions of each of testos-terone propionate and the heterocyclic steroid derivatives 4b, 9a,and 12 were prepared by dissolving in 3% Tween solution.
2.2.2. AnimalsNinety male mice with body weight ranging from 20 to 25 g,
purchased from the Animal House Laboratory, National ResearchCentre, Cairo, Egypt, were used in this study. The animals werehoused in plastic cages, ten per cage, and maintained on stan-dard laboratory diet (protein, 16.04%; fat, 3.63%; fiber, 4.1%; andmetabolic energy, 0.012 MJ) and water ad libitum at the AnimalHouse Laboratory, National Research Center, Cairo, Egypt. All ani-mals left to acclimatize to the new place for 1 week before theexperiment and received humane care in compliance with theguidelines of the Animal Care and Use Committee of NationalResearch Center, Egypt.
2.2.3. Experimental designTable 1 is intended to demonstrate the experimental design.
Animals were randomly divided into nine groups (10 mice/group).The first group was injected with 3% Tween and used as control
group. The second and third groups were injected with testosteronepropionate (TP). The fourth and fifth groups were injected withpyridoandrostene derivative 4b. The sixth and seventh groups wereinjected with pyrimidinoandrostene derivative 9a. The eighth andninth groups were injected with thienoandrostene derivative 12. Allanimals were receiving subcutaneous injection daily for a period of14 days. The doses of all drugs (200 or 400 mg/kg b.w.) were chosenbased on previous studies [20].2.2.4. Androgenicity assayAfter 35 days of the last dose (duration of spermatogenesis)
[21], half number of the animals of each treated group (five ani-mals per group) was sacrificed by neck vertebra luxation. Theepididymides and testes from each mouse were removed andweighed. Sperm were collected as quickly as possible when eachmouse was dissected. To release sperm, the cauda epididymideswere cut in a pre-warmed Petri dish containing 1 ml of saline solu-tion at 37 ◦C. After mincing with scalpels, the suspension was stirredand dropped on grease-free clean slide to determine the motility ofsperm using microscope. Spermatozoa were counted using heamo-cytometer and a drop of a homogenate smeared on a cleaned slide
& Molecular Biology 110 (2008) 284–294 287
allowed to air dry and stained with approximately 0.05% aqueouseosin Y. The slides were coded and used for the examination ofsperm head and tails abnormalities. For each animal 500 spermwere examined for morphological abnormalities according to thecriteria of Jeong et al. [22].
2.2.5. Genotoxicity assays2.2.5.1. Chromosome aberration test. At the end of the treatment,five animals of all treated groups were injected intraperitoneallywith colchicine to arrest cell division at metaphase. Two hours afterinjection, animals were sacrificed for preparation of the chromo-somes of bone-marrow cells and spermatocyte cells. Chromosomesof bone-marrow cells were prepared by using the methodology ofYosida and Amano [23]. Spermatocytes were prepared according toBrewen and Preston [24]. After staining the slides with 10% Giemsadye, 100 good metaphase spreads of each cell type for all animalswere examined microscopically to analyze the different types ofchromosomal aberrations.
2.2.5.2. Molecular biology analysis (RAPD–PCR analysis). Thegenomic DNA was isolated from mice liver using phe-nol/chloroform extraction and ethanol precipitation methodwith minor modifications [25]. RAPD–PCR profiles from miceDNA were generated using 10 oligodecamers (10-mer randomprimers: D01: 5′-ACCGCGAAGG-3′, D02: 5′-GGACCCAACC-3′, D03: 5′-GTCGCCGTCA-3′, B07: 5′-GGTGACGCAG-3′,B09: 5′-TGGGGGACTC-3′, B10: 5′-CTGCTGGGAC-3′, B12: 5′-CCTTGACGCA-3′, B14: 5′-TCCGCTCTGG-3′, B18: 5′-CCACAGCAGT-3′
and B20: 5′-GGACCCTTAC-3′, Operon Technologies, USA). DNAamplification reactions were performed under conditions reportedby Luceri et al. [26]. PCR amplification was conducted in 50 �lreaction volume containing 100 ng genomic DNA; 100 �M dNTPs;40 nM primer (Operon, Almeda, CA, USA); 2.5 units of Taq DNApolymerase and 5 �l promega 10× Taq DNA polymerase buffer.The reactions were carried out in Thermocycler (Perkin-Elmer9700) programmed with a first denaturation of 5 min at 94 ◦C,followed by 45 cycles of 0.5 min at 94 ◦C, 1 min at 36 ◦C and 2 minat 72 ◦C and finally, 1 cycle at 72 ◦C for 5 min. The PCR product wasanalyzed by electrophoresing 10 �l of the amplified mixture onagarose gel. The Gel-Pro Analyzer (Media Cybernetics) was used todocument ethidium bromide DNA gels.
2.2.6. Statistical analysesStatistical analyses were performed with SPSS software. Data
were analyzed using one-way analysis of variance (ANOVA) fol-lowed by Duncan’s post hoc test for multiple comparisons betweenpairs. Results are reported as mean values ± S.D. and deference’swere considered as significant when P ≤ 0.05.
3. Results and discussion
3.1. Chemistry
Steroids with thiophene, pyrimidine and pyridine ring fusedto different positions of steroid skeleton are endowed withuseful biological activities [10,27,28]. Hence, it was thought ofinterest to undertake the synthesis of these derivatives start-ing with 17�-acetoxy-androst-4-en-3-one (1) which can easilybe obtained via simple acetylation of testosterone with acetylchloride. 2-Arylmethylen-17�-acetoxyandrost-4-en-3-one 2a andb was prepared via aldol condensation of compound (1) with ben-zaldhyde or p-methoxy benzaldhyde in refluxing sodium ethoxidesolution [18,19] (Scheme 1). The structure of compounds 2a andb was confirmed by elemental analysis, spectral data and chem-ical transportation. Condensation of compounds 2a and b with
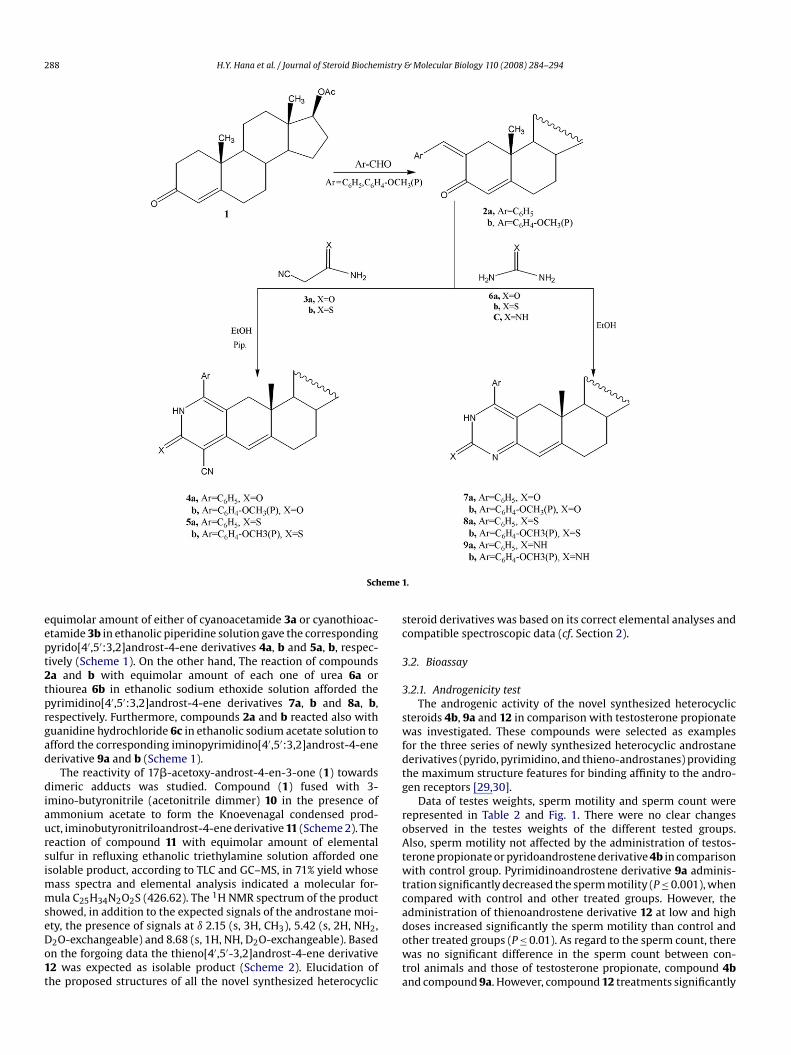
288 H.Y. Hana et al. / Journal of Steroid Biochemistry & Molecular Biology 110 (2008) 284–294
eme
Schequimolar amount of either of cyanoacetamide 3a or cyanothioac-etamide 3b in ethanolic piperidine solution gave the correspondingpyrido[4′,5′:3,2]androst-4-ene derivatives 4a, b and 5a, b, respec-tively (Scheme 1). On the other hand, The reaction of compounds2a and b with equimolar amount of each one of urea 6a orthiourea 6b in ethanolic sodium ethoxide solution afforded the
pyrimidino[4′,5′:3,2]androst-4-ene derivatives 7a, b and 8a, b,respectively. Furthermore, compounds 2a and b reacted also withguanidine hydrochloride 6c in ethanolic sodium acetate solution toafford the corresponding iminopyrimidino[4′,5′:3,2]androst-4-enederivative 9a and b (Scheme 1).The reactivity of 17�-acetoxy-androst-4-en-3-one (1) towardsdimeric adducts was studied. Compound (1) fused with 3-imino-butyronitrile (acetonitrile dimmer) 10 in the presence ofammonium acetate to form the Knoevenagal condensed prod-uct, iminobutyronitriloandrost-4-ene derivative 11 (Scheme 2). Thereaction of compound 11 with equimolar amount of elementalsulfur in refluxing ethanolic triethylamine solution afforded oneisolable product, according to TLC and GC–MS, in 71% yield whosemass spectra and elemental analysis indicated a molecular for-mula C25H34N2O2S (426.62). The 1H NMR spectrum of the productshowed, in addition to the expected signals of the androstane moi-ety, the presence of signals at ı 2.15 (s, 3H, CH3), 5.42 (s, 2H, NH2,D2O-exchangeable) and 8.68 (s, 1H, NH, D2O-exchangeable). Basedon the forgoing data the thieno[4′,5′-3,2]androst-4-ene derivative12 was expected as isolable product (Scheme 2). Elucidation ofthe proposed structures of all the novel synthesized heterocyclic
1.
steroid derivatives was based on its correct elemental analyses andcompatible spectroscopic data (cf. Section 2).
3.2. Bioassay
3.2.1. Androgenicity test
The androgenic activity of the novel synthesized heterocyclicsteroids 4b, 9a and 12 in comparison with testosterone propionatewas investigated. These compounds were selected as examplesfor the three series of newly synthesized heterocyclic androstanederivatives (pyrido, pyrimidino, and thieno-androstanes) providingthe maximum structure features for binding affinity to the andro-gen receptors [29,30].
Data of testes weights, sperm motility and sperm count wererepresented in Table 2 and Fig. 1. There were no clear changesobserved in the testes weights of the different tested groups.Also, sperm motility not affected by the administration of testos-terone propionate or pyridoandrostene derivative 4b in comparisonwith control group. Pyrimidinoandrostene derivative 9a adminis-tration significantly decreased the sperm motility (P ≤ 0.001), whencompared with control and other treated groups. However, theadministration of thienoandrostene derivative 12 at low and highdoses increased significantly the sperm motility than control andother treated groups (P ≤ 0.01). As regard to the sperm count, therewas no significant difference in the sperm count between con-trol animals and those of testosterone propionate, compound 4band compound 9a. However, compound 12 treatments significantly

H.Y. Hana et al. / Journal of Steroid Biochemistry & Molecular Biology 110 (2008) 284–294 289
Scheme 2
Table 2Androgenic activity of testosterone propionate and the tested compounds in testisof male mice
Experimental groups Testes weightsa Motilityb Sperm countc
Control 0.3 ± 0.02 88.6 ± 0.8c 546.6 ± 40.3cd
TP-L 0.3 ± 0.02 88.8 ± 0.6c 580.0 ± 33.1cd
Compound 4b-L 0.3 ± 0.02 88.4 ± 0.9c 471.4 ± 14.6d
Compound 9a-L 0.3 ± 0.03 72.2 ± 1.0d 671.4 ± 36.9bc
Compound 12-L 0.4 ± 0.02 95.4 ± 0.8b 454.4 ± 28.9bcd
TP-H 0.3 ± 0.02 90.2 ± 0.6c 677.0 ± 2.2bc
Compound 4b-H 0.3 ± 0.02 90.6 ± 0.5c 601.4 ± 10.7bcd
Compound 9a-H 0.3 ± 0.02 73.4 ± 1.1d 601.8 ± 76.2bcd
Compound 12-H 0.4 ± 0.02 99.8 ± 0.2a 900.4 ± 15.4a
Significant difference level Ns P ≤ 0.01 P ≤ 0.01
a Data shown as mean testis weights ±S.D.b Data show as percent motile sperm ±S.D.c Data shown as mean number of sperm (×106)/ml ±S.D.; values with differ-
ent superscripts letters within columns represent significant statistical differences(P < 0.05, Duncan’s post hoc test); L: low dose of treatment; H: high dose of treat-ment; TP: testosterone propionate.
increased sperm count (P ≤ 0.01) as compared with testosteronepropionate and compounds 4b and 9a.
The mean values of different types of the abnormalities ofsperm morphology were shown in Table 3 and Fig. 2. The most
Fig. 1. Androgenic activity of testosterone propionate and the tested compounds ondifferent reproductive parameters in male mice. Different superscript letters repre-sent significant statistical differences (P < 0.05, Duncan’s post hoc test); L: low doseof treatment; H: high dose of treatment; TP: testosterone propionate.
.
frequently observed abnormality was coiled tail sperm. There wasno difference in the frequencies of the morphologically abnormalsperm of testosterone propionate at low dose and control animals.Additionally, testosterone propionate at high dose as well as com-pounds 4b and 12 at low dose caused non-significant decreasein the frequencies of the sperm abnormalities. Meanwhile, com-pound 12 at high dose treatment induced significant decreasein the frequencies of these abnormalities. In contrary, compound9a treatment showed significant increase in the frequencies ofsperm abnormalities (P ≤ 0.01) when compared with all testedcompounds.
These results on the androgenic activity indicated that the novelsynthesized heterocyclic steroids, pyridoandrostene derivative 4band thienoandrostene derivative 12, have androgenic activity aswell as testosterone propionate. Since, the results show that com-pound 4b did have negative effect on the sperm motility, count andabnormalities. In addition, compound 12 increased sperm motil-ity, count and decreased the mean value of sperm abnormalities.This finding confirms that these compounds possess androgenic-like activity which is thought to be derived from the fusion ofpyridine or thiophene rings to androstane skeleton. To our knowl-
edge, we demonstrate herein for the first time the androgeniceffect of such pyrido- and thieno-androstane derivatives. Directedby the structure of compounds 4b and 12, it is probably that thesecompounds have a dual mechanism for their androgenic activity.Stimulation of androgen receptors which activate specific genes toproduce proteins which go to another organ as feed back mecha-nism. They also may affect the activation rate of enzyme systemsinvolved in protein metabolism, thus enhancing protein synthesisand inhibiting protein degradation. These actions are responsi-ble for changes in primary and secondary sexual characteristics[31]. Compound 9a induced toxic impact on the spermatogene-sis. Where the male mice treated with compound 9a showed signsof reproductive toxicity since sperm morphological abnormali-ties were significantly high than in control, and therefore reducesthe fertility. It is known that the induction of sperm abnormal-ities indicate point mutations in germ cells which should havetriggered structural changes in cell organelles involved in headand tail formation, leading to sperm abnormalities [32,33]. Thisfinding may shed light on the antiandrogenic activity of pyrim-idinoandrostane derivative 9a which may resemble that of otherpyrimidine derivatives which exhibit their antiandrogenic affect via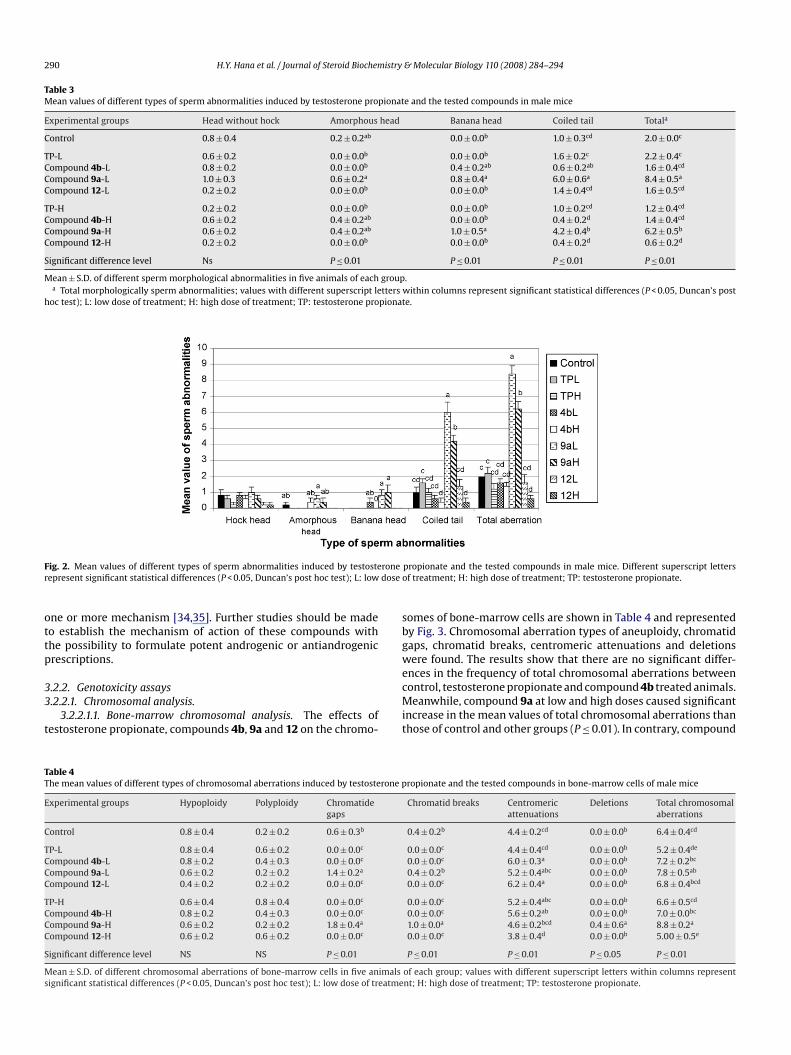
290 H.Y. Hana et al. / Journal of Steroid Biochemistry & Molecular Biology 110 (2008) 284–294
Table 3Mean values of different types of sperm abnormalities induced by testosterone propionate and the tested compounds in male mice
Experimental groups Head without hock Amorphous head Banana head Coiled tail Totala
Control 0.8 ± 0.4 0.2 ± 0.2ab 0.0 ± 0.0b 1.0 ± 0.3cd 2.0 ± 0.0c
TP-L 0.6 ± 0.2 0.0 ± 0.0b 0.0 ± 0.0b 1.6 ± 0.2c 2.2 ± 0.4c
Compound 4b-L 0.8 ± 0.2 0.0 ± 0.0b 0.4 ± 0.2ab 0.6 ± 0.2ab 1.6 ± 0.4cd
Compound 9a-L 1.0 ± 0.3 0.6 ± 0.2a 0.8 ± 0.4a 6.0 ± 0.6a 8.4 ± 0.5a
Compound 12-L 0.2 ± 0.2 0.0 ± 0.0b 0.0 ± 0.0b 1.4 ± 0.4cd 1.6 ± 0.5cd
TP-H 0.2 ± 0.2 0.0 ± 0.0b 0.0 ± 0.0b 1.0 ± 0.2cd 1.2 ± 0.4cd
Compound 4b-H 0.6 ± 0.2 0.4 ± 0.2ab 0.0 ± 0.0b 0.4 ± 0.2d 1.4 ± 0.4cd
Compound 9a-H 0.6 ± 0.2 0.4 ± 0.2ab 1.0 ± 0.5a 4.2 ± 0.4b 6.2 ± 0.5b
Compound 12-H 0.2 ± 0.2 0.0 ± 0.0b 0.0 ± 0.0b 0.4 ± 0.2d 0.6 ± 0.2d
Significant difference level Ns P ≤ 0.01 P ≤ 0.01 P ≤ 0.01 P ≤ 0.01
Mean ± S.D. of different sperm morphological abnormalities in five animals of each group.a Total morphologically sperm abnormalities; values with different superscript letters within columns represent significant statistical differences (P < 0.05, Duncan’s post
hoc test); L: low dose of treatment; H: high dose of treatment; TP: testosterone propionate.
Fig. 2. Mean values of different types of sperm abnormalities induced by testosteronerepresent significant statistical differences (P < 0.05, Duncan’s post hoc test); L: low dose
one or more mechanism [34,35]. Further studies should be madeto establish the mechanism of action of these compounds withthe possibility to formulate potent androgenic or antiandrogenicprescriptions.
3.2.2. Genotoxicity assays3.2.2.1. Chromosomal analysis.
3.2.2.1.1. Bone-marrow chromosomal analysis. The effects oftestosterone propionate, compounds 4b, 9a and 12 on the chromo-
Table 4The mean values of different types of chromosomal aberrations induced by testosterone p
Experimental groups Hypoploidy Polyploidy Chromatidegaps
Control 0.8 ± 0.4 0.2 ± 0.2 0.6 ± 0.3b
TP-L 0.8 ± 0.4 0.6 ± 0.2 0.0 ± 0.0c
Compound 4b-L 0.8 ± 0.2 0.4 ± 0.3 0.0 ± 0.0c
Compound 9a-L 0.6 ± 0.2 0.2 ± 0.2 1.4 ± 0.2a
Compound 12-L 0.4 ± 0.2 0.2 ± 0.2 0.0 ± 0.0c
TP-H 0.6 ± 0.4 0.8 ± 0.4 0.0 ± 0.0c
Compound 4b-H 0.8 ± 0.2 0.4 ± 0.3 0.0 ± 0.0c
Compound 9a-H 0.6 ± 0.2 0.2 ± 0.2 1.8 ± 0.4a
Compound 12-H 0.6 ± 0.2 0.6 ± 0.2 0.0 ± 0.0c
Significant difference level NS NS P ≤ 0.01
Mean ± S.D. of different chromosomal aberrations of bone-marrow cells in five animalssignificant statistical differences (P < 0.05, Duncan’s post hoc test); L: low dose of treatme
propionate and the tested compounds in male mice. Different superscript lettersof treatment; H: high dose of treatment; TP: testosterone propionate.
somes of bone-marrow cells are shown in Table 4 and representedby Fig. 3. Chromosomal aberration types of aneuploidy, chromatidgaps, chromatid breaks, centromeric attenuations and deletionswere found. The results show that there are no significant differ-ences in the frequency of total chromosomal aberrations betweencontrol, testosterone propionate and compound 4b treated animals.Meanwhile, compound 9a at low and high doses caused significantincrease in the mean values of total chromosomal aberrations thanthose of control and other groups (P ≤ 0.01). In contrary, compound
ropionate and the tested compounds in bone-marrow cells of male mice
Chromatid breaks Centromericattenuations
Deletions Total chromosomalaberrations
0.4 ± 0.2b 4.4 ± 0.2cd 0.0 ± 0.0b 6.4 ± 0.4cd
0.0 ± 0.0c 4.4 ± 0.4cd 0.0 ± 0.0b 5.2 ± 0.4de
0.0 ± 0.0c 6.0 ± 0.3a 0.0 ± 0.0b 7.2 ± 0.2bc
0.4 ± 0.2b 5.2 ± 0.4abc 0.0 ± 0.0b 7.8 ± 0.5ab
0.0 ± 0.0c 6.2 ± 0.4a 0.0 ± 0.0b 6.8 ± 0.4bcd
0.0 ± 0.0c 5.2 ± 0.4abc 0.0 ± 0.0b 6.6 ± 0.5cd
0.0 ± 0.0c 5.6 ± 0.2ab 0.0 ± 0.0b 7.0 ± 0.0bc
1.0 ± 0.0a 4.6 ± 0.2bcd 0.4 ± 0.6a 8.8 ± 0.2a
0.0 ± 0.0c 3.8 ± 0.4d 0.0 ± 0.0b 5.00 ± 0.5e
P ≤ 0.01 P ≤ 0.01 P ≤ 0.05 P ≤ 0.01
of each group; values with different superscript letters within columns representnt; H: high dose of treatment; TP: testosterone propionate.
H.Y. Hana et al. / Journal of Steroid Biochemistry & Molecular Biology 110 (2008) 284–294 291
Fig. 3. Frequencies of different types of chromosomal aberrations induced by testosterone propionate and the tested compounds in bone-marrow cells of male mice.can’s
ions in
Different superscript letters represent significant statistical differences (P < 0.05, Dunpropionate.
Table 5The mean values and standard deviations of different types of chromosomal aberratof male mice
Experimental groups Hypoploidy Polyploidy
Control 1.6 ± 0.2 1.2 ± 0.2c
TP-L 0.8 ± 0.4 1.8 ± 0.2bc
Compound 4b-L 0.8 ± 0.4 2.4 ± 0.2ab
Compound 9a-L 1.2 ± 0.4 2.8 ± 0.2a
Compound 12-L 0.6 ± 0.2 1.2 ± 0.2c
TP-H 1.2 ± 0.2 2.0 ± 0.5bc
Compound 4b-H 1.0 ± 0.3 2.2 ± 0.2ab
Compound 9a-H 1.6 ± 0.2 2.4 ± 0.2ab
Compound 12-H 0.8 ± 0.4 1.2 ± 0.2c
Significant difference level NS P ≤ 0.01
Mean ± S.D. of different chromosomal aberrations of spermatocytes in five animals of eachstatistical differences (P < 0.05, Duncan’s post hoc test); L: low dose of treatment; H: high
12 at high dose showed significant decrease in the frequencies oftotal chromosomal aberrations (P ≤ 0.01) than those of control andother compounds.
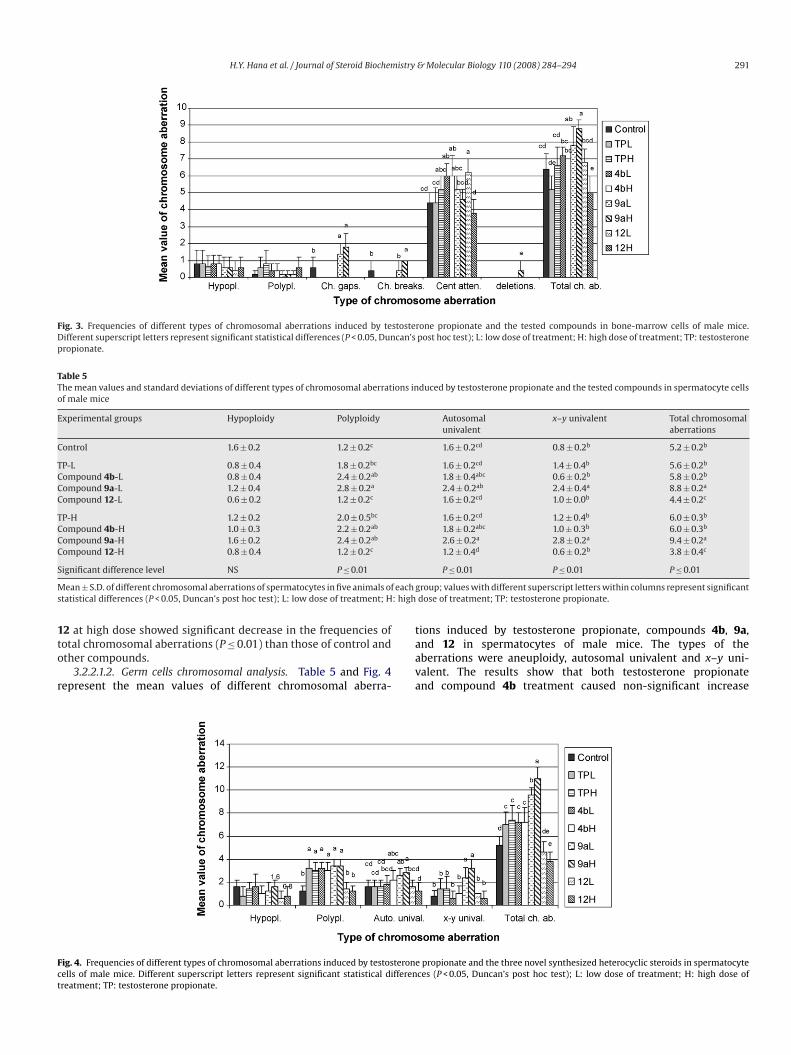
3.2.2.1.2. Germ cells chromosomal analysis. Table 5 and Fig. 4represent the mean values of different chromosomal aberra-
Fig. 4. Frequencies of different types of chromosomal aberrations induced by testosteroncells of male mice. Different superscript letters represent significant statistical differentreatment; TP: testosterone propionate.
post hoc test); L: low dose of treatment; H: high dose of treatment; TP: testosterone
duced by testosterone propionate and the tested compounds in spermatocyte cells
Autosomalunivalent
x–y univalent Total chromosomalaberrations
1.6 ± 0.2cd 0.8 ± 0.2b 5.2 ± 0.2b
1.6 ± 0.2cd 1.4 ± 0.4b 5.6 ± 0.2b
1.8 ± 0.4abc 0.6 ± 0.2b 5.8 ± 0.2b
2.4 ± 0.2ab 2.4 ± 0.4a 8.8 ± 0.2a
1.6 ± 0.2cd 1.0 ± 0.0b 4.4 ± 0.2c
1.6 ± 0.2cd 1.2 ± 0.4b 6.0 ± 0.3b
1.8 ± 0.2abc 1.0 ± 0.3b 6.0 ± 0.3b
2.6 ± 0.2a 2.8 ± 0.2a 9.4 ± 0.2a
1.2 ± 0.4d 0.6 ± 0.2b 3.8 ± 0.4c
P ≤ 0.01 P ≤ 0.01 P ≤ 0.01
group; values with different superscript letters within columns represent significantdose of treatment; TP: testosterone propionate.
tions induced by testosterone propionate, compounds 4b, 9a,and 12 in spermatocytes of male mice. The types of theaberrations were aneuploidy, autosomal univalent and x–y uni-valent. The results show that both testosterone propionateand compound 4b treatment caused non-significant increase
e propionate and the three novel synthesized heterocyclic steroids in spermatocyteces (P < 0.05, Duncan’s post hoc test); L: low dose of treatment; H: high dose of
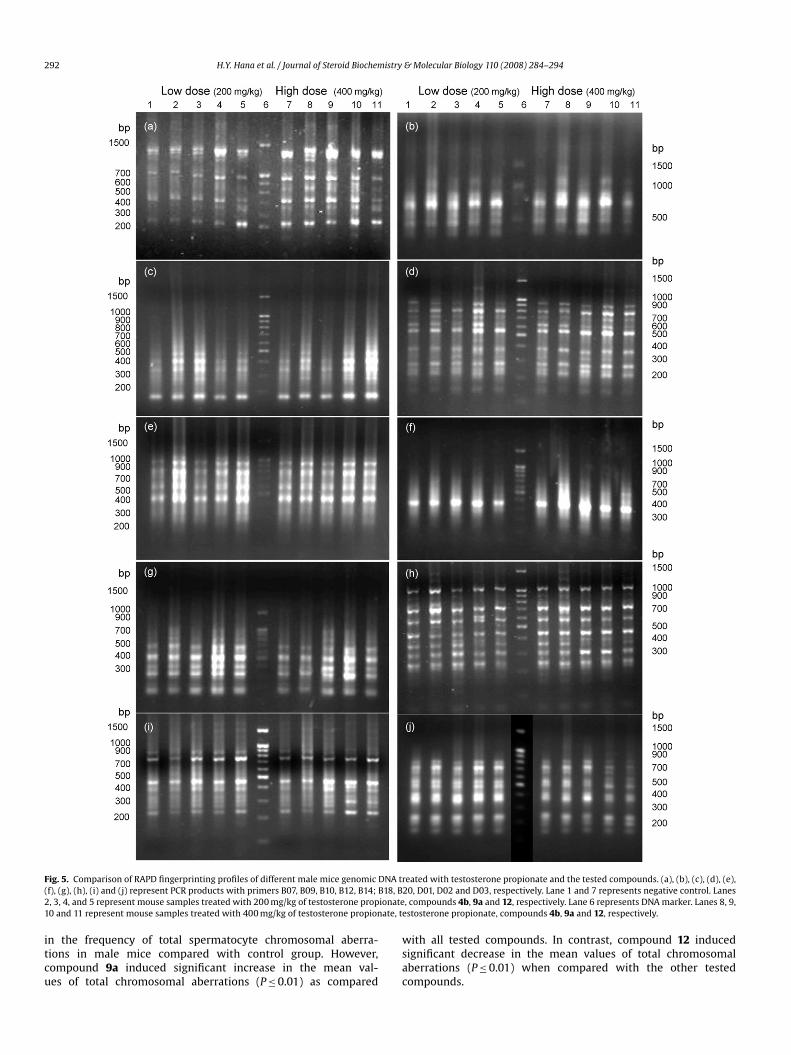
292 H.Y. Hana et al. / Journal of Steroid Biochemistry & Molecular Biology 110 (2008) 284–294
Fig. 5. Comparison of RAPD fingerprinting profiles of different male mice genomic DNA t(f), (g), (h), (i) and (j) represent PCR products with primers B07, B09, B10, B12, B14; B18, B2, 3, 4, and 5 represent mouse samples treated with 200 mg/kg of testosterone propionate10 and 11 represent mouse samples treated with 400 mg/kg of testosterone propionate, t
in the frequency of total spermatocyte chromosomal aberra-tions in male mice compared with control group. However,compound 9a induced significant increase in the mean val-ues of total chromosomal aberrations (P ≤ 0.01) as compared
reated with testosterone propionate and the tested compounds. (a), (b), (c), (d), (e),20, D01, D02 and D03, respectively. Lane 1 and 7 represents negative control. Lanes, compounds 4b, 9a and 12, respectively. Lane 6 represents DNA marker. Lanes 8, 9,
estosterone propionate, compounds 4b, 9a and 12, respectively.
with all tested compounds. In contrast, compound 12 inducedsignificant decrease in the mean values of total chromosomalaberrations (P ≤ 0.01) when compared with the other testedcompounds.

istry
H.Y. Hana et al. / Journal of Steroid Biochem3.2.2.2. RAPD fingerprinting pattern. The molecular genetic vari-ability among the treated mice genomes and their control wereevaluated using 10 oligodecamers (10-mer random primers). Allof these primers gave positive and detectable bands (Fig. 5).They amplified a total of 164 different bands, with an average of16.4 ± 1.8 bands per primer. All of the oligodecamers used revealedmonomorphic bands in the control samples and those treated withtestosterone propionate at 200 or 400 mg/kg b.w. (Fig. 5). Thebands resulted from compound 4b were relatively similar to thescorable bands resulted from control or with testosterone propi-onate (Fig. 5). Where, all of bands at the low dose of compound 4bwere monomorphic with those resulted from control and testos-terone propionate. In addition, at the high dose of compound 4b,most (60%) primers (D03, B07, B09, B10, B14 and B18) used revealed100% monomorphic band patterns with those resulted from controland testosterone propionate samples (Fig. 5). On the other hands,compound 9a revealed high polymorphic bands patterns. Despite,at the low dose of compounds 9a and 12, 60% of the primers (D02,D03, B09, B14, B18 and D20) used revealed monomorphic band pat-terns with other groups (Fig. 5), at the high dose of compounds9a and 12, 80% of the primers (D01, D02, D03, B07, B10, B12, B18and B20) used revealed polymorphic bands. These polymorphicbands, which did not appear in the DNA samples of other groups,were observed more in compound 9a (32 bands) than in compound12 (20 bands) (Fig. 5). These new bands could be considered as“genus diagnostic” markers which attributed to the treatment bycompound 9a.
These findings of the genotoxicity study show that testosteronepropionate and pyridoandrostene derivative 4b did not inducesignificant differences in either chromosomal aberration of bothsomatic and germ cells, or the DNA bands patterns. In agreementwith these results, Marzin [36] studied the mutagenicity of somesynthetic androgen steroids, using number of genotoxicity tests invitro and in vivo systems for gene mutations, chromosomal muta-tions and primary DNA damage demonstration. The results of thisstudy showed no genotoxic activity attributed to these steroids.It is also found that 17�-alkylated steroids are directly toxic tohepatocytes, whereas the non-alkylated steroids show no effectsat tested doses [37]. Tsutsui et al. [38] reported that testosteronedid not induce gene mutations at the hprt or Na+/K+ ATPase locus.When testosterone was added at a final concentration of 2 mmol/lto DNA obtained from human surgical resections, rat liver, HepG2cells, and calf thymus, did not form adducts with naked DNA.Furthermore, no adducts were observed in DNA isolated from
HepG2 cells incubated with 10–100 �mol/l testosterone for 24 h[39].Despite testosterone and its esters having negative effects inall clastogenicity tests [40–42], our results indicated that thetestosterone derivative pyrimidinoandrostene 9a caused signifi-cant increase in total chromosomal aberrations of both somaticand germ cells as well as enhanced the polymorphic bands pat-terns compared to the control. Martelli et al. [43] mentioned thatsex steroids, due to the generally negative responses observedin routinely employed standard genotoxicity assays, are consid-ered epigenetic carcinogens. Sex steroids differ for their ability toinduce DNA repair, and their genotoxicity may be different in ratand human hepatocytes, dependent on the sex of the donor, andaffected by inter-individual variability [44]. This explains that com-pound 9a induced sperm abnormalities may be due to the increasedincidence of chromosomal aberrations as well as enhanced thepolymorphic bands pattern. Many studies reported that variouschromosomal abnormalities, both structural and numerical anddeletions of Y chromosome could cause spermatogenic breakdownat various points, consequently resulting in chromosomally derivedinfertility [45,46].
[
[
[
[
[
& Molecular Biology 110 (2008) 284–294 293
Interest in the present study that thienoandrostene derivative 12treatments induced significant decrease in the mean values of chro-mosomal aberrations in both somatic and germ cells, and decreasedsperm morphological abnormalities as well as decreased the alter-ation in the DNA bands patterns compared to control group. Mossuzet al. [47] reported that sex steroids have the ability to enhancethe cell cycle associated with the differentiation. We have foundin the present study that compound 12 was able to enhance thecell cycle of the sperm, where it increased the count and motilityof the sperms. To our knowledge, this is the first study that sug-gests significant stimulating effects of thienosteroid derivative onandrogenic anti-mutagenic activities in mice.
3.3. Conclusion
In this study we have described efficient synthesis of novel het-erocyclic steroids. Compound 4b has androgenic activity as well astestosterone propionate. There were no significant differences inthe frequencies of total chromosomal aberrations in both somaticand germ cells as well as no alteration in the DNA bands patternsbetween control, testosterone propionate and pyridoandrostene 4btreated animals. On the other hand, thienoandrostene derivative 12induced significant decrease in the mean values of chromosomalaberrations in both somatic and germ cells, decreased sperm mor-phological abnormalities, increased the sperm count and motilityas compared with testosterone propionate. However, the pyrim-idinoandrostene derivative 9a has antiandrogenic and genotoxicactivities as compared to the control. Compound 12 showed appar-ent hormone agonist properties with no genotoxic activity, thatmay play a role in male sexual function in general and erectilephysiology in particular. Thus compound 12 may be helpful in thetreatment and monitoring the male hypogonadism.
References
[1] W.M.D. Regelson, C. Colman, The Super-Hormone Promise, Simon & Schuster,New York, 1996, ISBN 0-684-83011-6, pp. 115–152.
[2] H.W.G. Baker, Testicular dysfunction in systematic disease, in: K.L. Becker (Ed.),Principles and Practice of Endocrinology and Metabolism, 2nd edition, J.B. Lip-pincott Co., Philadelphia, 1995, pp. 1083–1089.
[3] M.D. Hendler, Ph.D. Sheldon Saul, The Doctor’s Vitamin and Mineral Encyclo-pedia, 201, Fireside, New York, 1990, ISBN 0-671-66784-X, pp. 385–387.
[4] S. Bhasin, T. Storer, N. Berman, C. Callegari, B. Clevenger, J. Phillips, T.J. Bun-nell, R. Tricker, A. Shirazi, R. Casaburi, The effects of supraphysiologic doses oftestosterone on muscle size and strength in normal men, N. Engl. J. Med. 335
(1) (1996) 1–7.[5] A. Gray, J.A. Berlin, J.B. McKinley, C. Longcope, An examination of research designeffects on the association of testosterone and male aging: results of a meta-analysis, J. Clin. Epidemiol. 44 (1991) 671–684.
[6] D. Simon, P. Preziosi, E. Barrett-Connor, et al., The influence of aging on plasmasex hormones in men: the Telecom Study, Am. J. Epidemiol. 135 (1992) 783–791.
[7] S.M. Ho, M. Yu, Selective increase in type II estrogen binding sites in the dys-plastic dorsolateral prostates of Noble rats, Cancer Res. 53 (1993) 528–532.
[8] S.M. Ho, D. Roy, Sex hormone-induced nuclear DNA damage and lipid per-oxidation in the dorsolaterel prostates of Noble rats, Cancer Lett. 84 (1994)155–162.
[9] G. Ragnotti, M. Presta, J.A.M. Maier, M. Rusnati, G. Mazzoleni, F. Legati, R. Chiesa,M. Braga, D. Calovini, Critical role of gonadal hormones on the genotoxic activityof the hepatocarcinogen DL-ZAMI 1305, Cancer Lett. 36 (1987) 253–261.
10] K. Penov-Gasi, M. Sakac, E. Djurendic, R. Kovacevic, S. Andric, Rearrangement ofsome steroidal �,�-unsaturated pyridine-N-oxides and antiandrogenic activityof obtained compounds, J. Serb. Chem. Soc. 63 (1998) 189–192.
11] F. Sparatore, R. Cerri, G. Delrio, R.K. Rastogi, 3,4-Dihydro-1,2,4-benzotriazin-3,3′-spirosteriod derivatives of pharmacological interest, Farmaco [Science] 35(1980) 725–734.
12] R.O. Clinton, A.J. Manson, F.W. Stonner, A.L. Beyler, G.O. Potts, A. Arnold,Steroidal[3,2-c]pyrazoles, J. Am. Chem. Soc. 81 (1959) 1513–1514.
13] G. Zanati, M.E. Wolff, Heterocyclic steroids 4: synthesis and androgenic activityof A-ring homosteroids, J. Med. Chem. 15 (1972) 368–369.
14] R.O. Clinton, A.J. Manson, F.W. Stonner, H.C. Neumann, R.G. Christiansen, R.L.Clarke, J.H. Ackerman, D.F. Page, J.W. Dean, W.B. Dickinson, C. Carabateas,Steroidal[3,2-c]pyrazoles. II. Androstanes, 19-norandrostanes and their satu-rated analogs, J. Am. Chem. Soc. 83 (1961) 1478–1491.

istry
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
294 H.Y. Hana et al. / Journal of Steroid Biochem
15] R.G. Christiansen, M.R. Bell, T.E. D’Ambra, J.P. Mallamo, J.L. Herrmann, J.H. Ack-erman, C.J. Opalka, R.K. Kullnig, R.C. Winneker, B.W. Snyder, F.H. Batzold, H.P.Schane, Antiandrogenic steroidal sulfonylpyrazoles, J. Med. Chem. 33 (1990)2094–2100.
16] IUPAC, Joint Commission on Biochemical Nomenclature (JCBN), Definitive rulesfor nomenclature of steroids, J. Am. Chem. Soc. 82 (1960) 5577–5581.
17] IUPAC, Joint Commission on Biochemical Nomenclature (JCBN), The nomen-clature of steroids. Recommendations 1989, Eur. J. Biochem. 186 (1989)429–458.
18] D.H.R. Barton, Kekule Symposium “Theoretical Organic Chemistry”, Messrs,Buttor, Worth Scientific Publications, London, 1959, p. 127.
19] H.M. Safwat, M.H. El-Gamal, M.M. Hussein, M.M. Abdullah, Functionalizationat ring D of some steroids: aldol condensation of aromatic aldehydes with 3-hydroxyandrostane-17-one, Bull. Fac. Pharm. Cairo Univ. 40 (2002) 47–56.
20] S.B. Nagelberg, L. Laue, D.L. Loriaux, L. Liu, R.J. Sherins, Cerebrovascular accidentassociated with testosterone therapy in a 21-year-old hypogonadal man, N.Engl. J. Med. 314 (10) (1986) 449–450.
21] V. Monesi, Autoradiographic study of DNA synthesis and the cell cycle in sper-matogonia and spermatocytes of mouse testis using tritiated thymidine, J. CellBiol. 14 (1962) 1–18.
22] S.H. Jeong, B.Y. Kim, H.G. Kang, H.O. Ku, J.H. Cho, Effects of butylated hydrox-yanisole on the development and functions of reproductive system in rats,Toxicology 208 (2005) 49–62.
23] T.H. Yosida, K. Amano, Autosomal polymorphism in laboratory bred and
wild Norway rats, Rattus norvegius, found in Misima, Chromosoma 16 (1965)658.24] G.J. Brewen, V.R. Preston, Analysis of chromosome aberrations in mammaliancells, Chem. Mutagen. 5 (1987) 127–150.
25] L. Sambrook, E.F. Fritsch, T. Manitatis, Molecular Cloning: A Laboratory Manual,Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989.
26] C. Luceri, C. De Filippo, G. Caderni, L. Gambacciani, M. Salvadori, A. Giannini, P.Dolara, Detection of somatic DNA alterations in azoxymethane-induced F344rat colon tumors by random amplified polymorphic DNA analysis, Carcinogen-esis 21 (9) (2000) 1753–1756.
27] G.A. Elmegeed, W.W. Wardakhan, A.R. Baiuomy, Synthesis of thiazolyl andthieno cholestane derivatives: a novel class of potent antiinflammatorysteroids, Pharmazie 60 (2005) 328–333.
28] T. Matsumoto, M. Watanabe, S. Mataka, T. Thiemann, Estrano[17,16-e]pyrimidine-peptide conjugates, Steroids 68 (9) (2003) 751–757.
29] D. Yin, Y. He, M.A. Perera, S.S. Hong, C. Marhefka, N. Stourman, L. Kirkovsky,D.D. Miller, J.T. Dalton, Key structural features of nonsteroidal ligands forbinding and activation of the androgen receptors, Mol. Pharmacol. 63 (2003)211–223.
30] A.T. Glen, L.R. Hughes, J.J. Morris, P.J. Taylor, Structure–activity relationshipsamong nonsteroidal antiandrogens, in: R.W. Lambert (Ed.), Proceeding of theThird Sci-RSC Medicinal Chemistry Symposium, Royal Society of Chemistry,London, 1986, pp. 345–361.
31] T.D. Fahey, Anabolic–androgenic steroids: mechanism of action and effects onperformance, in: T.D. Fahey (Ed.), Encyclopedia of Sports Medicine and Science,Internet Society for Sport Science: http://sportsci.org, 1998.
32] K. Narayana, N. Prashanthi, A. Nayanatara, H.H.C. Kumar, K. Abhilash, K.L. Bairy,Effects of methyl parathion (o,o-dimethyl o-4-nitrophenyl phosphorothioate)
[
[
[
[
[
[
[
& Molecular Biology 110 (2008) 284–294
on rat sperm morphology and sperm count, but not fertility, are associated withdecreased ascorbic acid level in the testis, Mutat. Res. 588 (2005) 28–34.
33] K.P.S. Rao, K. Narayana, In vivo chromosome damaging effects of an inosinemonophosphate dehydrogenase inhibitor: ribavirin in mice, Indian J. Pharma-col. 37 (2) (2005) 90–95.
34] WHO, Pesticide Residues in Food-1995 Evaluations. Part II. Toxicology.WHO/PCS/96.48, International Programme on Chemical Safety, 1995, WorldHealth Organization, Geneva.
35] A.M. Vinggaard, H. Jacobsen, S.B. Metzdorff, H.R. Andersen, C. Nellemann,Antiandrogenic effect in short-term in vivo studies of the fungicide fenarimol,Toxicology 207 (2005) 21–34.
36] D. Marzin, Trenbolone: application of the Ames test. Recent data, Ann. Rech.Vet. 22 (3) (1991) 257–262.
37] A.A. Welder, J.W. Robertson, R.B. Melchert, Toxic effects of anabolic–androgenicsteroids in primary rat hepatic cell cultures, J. Pharmacol. Toxicol. Methods 33(4) (1995) 187–195.
38] T. Tsutsui, A. Konine, J. Huff, J.C. Barrett, Effects of testosterone, testos-terone propionate, 17beta-trenbolone and progesterone on cell transformationand mutagenesis in Syrian hamster embryo cells, Carcinogenesis 16 (1995)1329–1333.
39] M.J. Seraj, A. Umemoto, M. Tanaka, A. Kajikawa, K. Hamada, Y. Monden, DNAadduct formation by hormonal steroids in vitro, Mutat. Res. 370 (1) (1996)49–59.
40] M. Richold, The genotoxicity of trenbolone, a synthetic steroid, Arch. Toxicol.
61 (1988) 249–258.41] T. Morita, N. Asano, T. Awogi, Y.F. Sasaki, S. Sato, H. Shimada, S. Sutou, T. Suzuki,A. Wakata, T. Sofuni, M. Hayashi, Evaluation of the rodent micronucleus assay inthe screening of IARC carcinogens (groups 1, 2A and 2B), Mutat. Res. 389 (1997)3–122.
42] H.E. Holden, D. Studwell, J.B. Majeska, Oxymetholone. I. Evaluation in a com-prehensive battery of genetic toxicology and in vitro transformation assays,Toxicol. Pathol. 27 (1999) 501–506.
43] A. Martelli, F. Mattioli, M. Angiola, R. Reimann, G. Brambilla, Species, sex andinter-individual differences in DNA repair induced by nine sex steroids in pri-mary cultures of rat and human hepatocytes, Mutat. Res. 536 (1–2) (2003)69–78.
44] R. Reimann, S. Kalweit, R. Lang, Studies for a genotoxic potential of someendogenous and exogenous sex steroids. II. Communication: examination forthe induction of cytogenetic damage using the chromosomal aberration assayon human lymphocytes in vitro and the mouse bone marrow micronucleus testin vivo, Environ. Mol. Mutagen. 28 (1996) 133–144.
45] A. Vicdan, K. Vicdan, S. Gunalp, A. Kence, C. Akarsu, A. Zeki, E. Sozen, Geneticaspects of human male infertility: the frequency of chromosomal abnormali-ties and Y chromosome microdeletions in severe male factor infertility, Eur. J.Obstet. Gynecol. Reprod. Biol. 117 (1) (2004) 49–54.
46] M.D. Mudry, A.M. Palermo, M.S. Merani, M.A. Carballo, Metronidazole-inducedalterations in murine spermatozoa morphology, Reprod. Toxicol. 23 (2) (2007)246–252.
47] P. Mossuz, F. Cousin, A. Castinel, M. Chauvet, M.F. Sotto, B. Polack, J.J. Sotto,L. Kolodie, Effects of two sex steroids (17beta estradiol and testosterone) onproliferation and clonal growth of the human monoblastic leukemia cell line,U937, Leuk. Res. 22 (11) (1998) 1063–1072.





























