Embed Size (px)
Citation preview

Neuroscience Vol. 37, No. 2, pp. Sll-$22, 1990 Printed in Great Britain
03~4522/90 $3.00 + 0.00 Pergamon Press plc
c 1990 IBRO
ANGIOTENSINOGEN IN THE DEVELOPING RAT FETAL HINDBRAIN AND SPINAL CORD FROM 18TH
TO 20TH DAY OF GESTATION: AN IMMUNOCYTOCHEMICAL STUDY
P. P. SOOD,*~ J. P. RICHOUX,$ M. PANIGEL,~ 3. BOUHNIK// and R. WECMANN$
*Laboratory of Neurobiology, Department of Biosciences, Saurashtra University, Rajkot 360 005, India JLaboratoire d’Histologie, Faculte de Medecine, 9, avenue de la Foret de Haye,
54505 Vandoeuvre, France SLaboratoire de Biologie de la Reproduction et Institut d’Histochimie Medicale, 3at A. 7,
Quai Saint-Bernard, Universite Pierre et Marie Curie, 75005 Paris, France IlInstitut National de la Recherche Medicale, U 36, 17 rue du Fer-a-Moulin, 75005 Paris, France
Abstract-We have detected angiotensinogen immunoactivity in the hindbrain and in the spinal cord of rat fetuses during the 18th to 20th day of gestation. In the Ilth-day fetus, a few immunor~ctive angiotensinogen cells are localized in precise brain areas. Their number sharply increase during the 19th and 20th day gestation period when there is an active cell differentiation and cell growth.
These observations suggest a role of the renin-angiotensinogen system during cell growth and cell differentiation.
In an earlier report we have shown renin-like immunoreactivity in the 19th-day fetal brain7B9 and given an indication that the fetal brain maintains a distinct renin-angiotensinogen system during late gestation period. As further verification of this we have endeavoured to demonstrate the presence of the renin substrate, angiotensinogen, in the hindbrain and spinal cord of rat fetuses during the 18th to 20th day of gestation.
EXPERIMENTAL PROCEDURES
The age of Wistar rat fetuses was dated from the observa- tion of spermatozoa in the vaginal smears of the mated femaie (day 0 of gestation). The pregnant rats were killed from the 18th to 20th day of gestation. The brains of four fetuses of each day were fixed in situ or after dissection in Bouin-Hollande sublimate. Tissues were embedded in paraffin; 556pm serial sections were obtained in frontal and sagittal planes. Every 20th section of each series was deparaffinized, rehydrated and equilibrated with phosphate buffer (0.1 M, pH_= 7.4) containing 0.5% Triton x-100, 1 mM EDTA. 0.05% NaN, and 1% lamb serum.
Angiotensinogen antisera were prepared using pure rat angiotensinogen.’ The usual indirect immuno~roxida~ techniques were used. (1) Specific angiotensinogen anti- serum was diluted 1/2~1~4~ for 2 h at room temperature; (2) peroxidase-labelled sheep anti-rabbit IgG diluted l/l00 for 30min at room temperature was employed.
Peroxidase activity was detected and intensified with diaminobenzidine (O.Ol%), Ni (NH,), (SO,), (0.02%) and H,Oz (0.003%) dissolved in 0.1 M phosphate buffer (pH = 7.4). Low nuclear counter-staining was performed using Kernechtrot.
The specificity of immunostaining was established by inhibition test using homologous or heterologous antigens
IVisiting Professor at Universite Pierre et Marie Curie, Paris, France.
Ahhreviution: EDTA, ethylenediaminetetra-acetate.
covalently linked to acrylamide agarose beads. Angioten- sinogen gel (0.3 x IO-‘ymol/ml) and human serum albumin gel were prepared. The specificity of the staining was also established by the conventional methods of verification for immunolo~cal studies, i.e. replacement of the specific serum by buffer or preimmune rabbit serum and staining inhibition tests with sequential dilutions of pure rat antigens. In addition, angiotensin I and II were tested to verify that they did not cross-react with the rat antiogensinogen antisera used.
RESULTS
The schematic distribution of angiotensinogen- immunopositive cells in the hindbrain and spinal cord
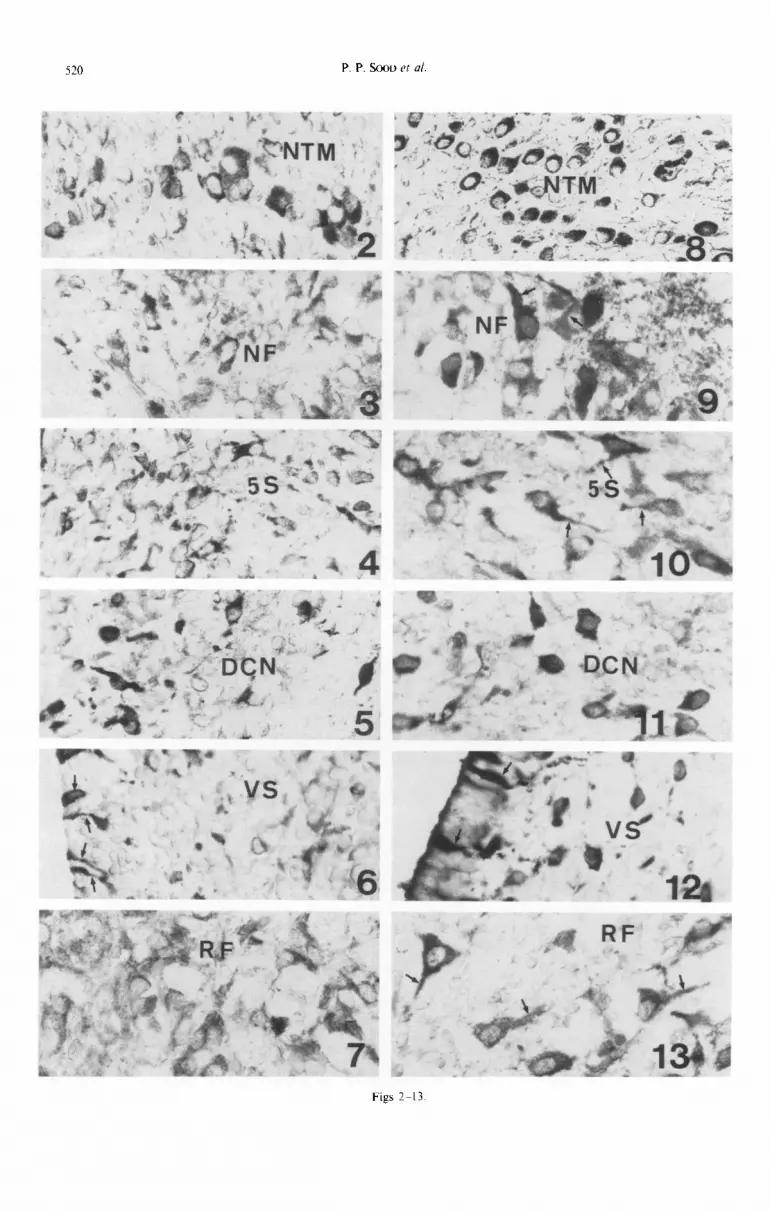
of fetuses during the 18th (A,-A& 19th (B,-Bs) and 20th (C&J day of gestation is shown in Fig. 1. The black dots are the areas where the angiotensinogen- positive cells are localized. In the 18th-day fetal brain the immunoa~tivity is quite weak, although the scat- tered positive cells are seen distributed all over the medulla oblongata and pons (A,-A,; Fig. 1). The number of positive cells in different nuclei is sum- marized in Table 1 and some of these areas are shown in Figs 2-7. It is evident from all these illustrations that the number of positive cells in the various nuclei of hindbrain on the 18th day of gestation is quite limited.
On the 19th day of gestation, the number of immunoreactive cells increases in all parts of the hindbrain (B,-B,; Fig. I). The increase in the number of positive cells can be seen, in particular, in the mesencephalic trigeminal nucleus (Fig. 8) nucleus n. facialis (Fig. 9) fifth motor trigeminal nucleus, prin- cipal sensory trigeminal nucleus {Fig. lo), deep cere- bellar nuclei (Fig. 1 I), nucleus vestibularis superior (Fig. 12) and reticular formation (Fig. 13).
517

18 DAY
P. P. So00 el al.
19 DAY 20 DAY
Fig. 1. Schematic drawings of serial sections of the hindbmin and spinal cord of fetuses on the 18th (A,-A,), 19th (B,--B,) and 20th (C,--C,) day of gestation. (A,, B,, C,) Sections passing through the pons; (A:, B,, Cz) cross-sections through fifth motor nucleus; (A,, B,. C,) middle region of medulla oblongata: (A,, B,, C,) sections through nucicus tractus solitarius; (A,, B,, C,) sections through caudal region of medulla oblongata; (Ah. B,, C,) cross-sections through cervical spinal cord. The black dots in differ- ent areas represent the angiotensinogen-immunopositive cells. On the 18th day of gestation there is a limited number of positive cells distributed all over the hindbrain. The number of positive cells increase in the 19th- and 20th-day fetal brain, thus representing a parallel increase of this prohormone with increasing cell differentiation and cell growth from the 18th day onwards. (Abbreviations used are listed
in Table 1.)

Angiotensinogen in developing fetal brain
Table I. Localization of immunoreactive angiotensinogen cells in the different nuclei of hindbrain of fetuses during the 18th to 20th day of gestation
18th-day 19th-day 20th-day Nwlei Abbreviations fetus fetus fetus
519
Cerebellum CB Central canal cc Colliculus inferior CI Dorsal horn DH Lemniscus nuclei LN Nucleus n. hy~glossi NH Nucleus ambiguus NA Nucleus cuneatus NC Nucleus dorsalis n. vagi ND Nucleus n. facialis NF Nucleus olivaris inferior NO1 Nucleus pontis NP Nucleus raphe magnus NRM Nucleus raphe obscurus NRO Principal sensory trigeminal nucleus 5s Nucleus tractus solitarii NTS Nucleus parabrachialis PB Nucleus prepositus hypoglossi PH Reticular formation RF Nucleus reticularis lateralis RL Substantia grisea centralis SGC Ventral horn VH Nucleus vestibularis lateralis VL Nucleus vestibularis superior vs Mesencephalic trigeminal nucleus NTM Fifth motor trigeminal nucleus 5M Deep cerebellar nuclei DCN
++
ff i-
++ + + + +
-l-f
++
it
N ++
+
zi + ;: -t ++ + ++ iG
+++
+++ ++
+++ + + + +
+++ +++
+4 N f
++ ++ ++
t ++ ++
+ +
++ ++
+++ f-t ++
-t--f-+
+++-I- ++
++++ +
i-+ ++ c
+++ ++++ +++
+ +
+++ +++ ++-t-
+ +++ +++ +i- ++
+++ +++
++++ +++ ++
The results are presented in relative number of positive cells per field (x 40): + + + + , > 15; + t f, 10-15; + f, 5-10; f, ~5; N, no staining.
On the 20th day of gestation the number of positive cells further increases in all the nuclei of hindbrain (C-C,; Fig. 1). The increase of positive cells is most prominent in colhculus inferior (Fig. 14), nucleus ohvaris inferior (Fig. 1 S), principal sensory trigeminal nucleus (Fig. 16), nucleus pontis {Fig. IS), reticular formation (Figs 18, 22), iemniscus nuclei (Fig. 20), and nucleus vestibularis superior (Fig. 21). Contrary to these, the nucleus dorsalis n. vagi and nucleus n. hypoglossi (Figs 17, 23) in the 20th-day fetal hind- brain do not show much difference in i~unoactivity as compared with 19th-day fetal brain. However, nucleus raphe magnus shows a few positive cells on the 20th-day of gestation. Spinat cord of 18th-day fetus exhibits a few angiotensinogen-positive cells
which are restricted to the dorsal and ventral horns (Fig. 1). Their number is slightly increased during the 19th and 20th day of gestation (B6, C,) (Fig. 1). The overall increase in the number of immuno~active cells in the hindbrain and spinal cord is summarized in Table I.
DlSCUSSION
First of ali it may be mentioned that the number of angiotensinogen-immunoreactive cells in the hind- brain and spinal cord of the 18th-day fetus is quite limited as compared with that of the forebrain8 Nevertheless, all the brain areas showed an increase in the immunopositive cells from the 18th day on- wards, and on the 20th day there is a maximum
Figs 2-7. Angiotensinogen immunoactivity in the fetal hindbrain and spinal cord on the 18th day of gestation. Note positive cells in the mesencephalic trigeminal nucleus (Pig. 2), nucleus n. facilitis (Fig. 3), principal sensory trigeminal nucleus (Fig. 4), deep cerebellar nuclei (Fig. 5), nucleus vestibularis superior (Fig. 6) and reticular formation (Fig. 7). Also note positive neuroepithelia cells at the border of fourth
ventricle (arrows; Fig. 6).
Figs 8-13. Areas of the 19th-day fetal brain. Note the increase of an~otens~nogen-positive cells in all the nuclei mentioned in the l&h-day fetal brain. (Compare Figs 2-7 with S-13.)
Figs 14-23. Transverse sections of the 2Oth-day fetal hindbrain and spinal cord demonstrating an increase of an~otensinogen-positive cells in colliculus inferior (Fig. 14), nucleus olivaris inferior (Fig. Is), principal sensory trigeminal nucleus (Fig. 16) nucleus reticularis lateralis (Fig. 18), nucleus pontis (Fig. 19), nucleus lemniscus (Fig. 20), nucleus vestibularis superior (Fig. 21) and reticular formation (Fig. 22). In nuclei dorsalis n. vagi (Fig. 17) and nucleus n. hypoglossi (Fig. 23) there are a few large cells positive for
angiotensinogen.
520 P. P. Scm et al.
-
Figs Z-13

Angiotensinogen in d
-_-. -- ^...__ - ” Figs 14-23.

522 P. P. S00D et al
number of immunoreactive cells in all the nuciei of hindbrain as well as in the spinal cord.
The precise localization of angiotensinogen in the fetal brainstem with increasing immunoreactivity from the 18th to 20th day of gestation, clearly shows a local synthesis of this prohormone and not diffusion from blood plasma. 3-4 In a recent study, we have also found renin-like immunoactivity in the 19th-day fetal brain including cerebellum and medulla oblongata.’ These observations, together with the fact that angiotensin II has already been reported in the tissue culture of the 20th.day fetal brain6 indicate that all the components of renin-angiotensinogen system are present in the fetal brain during late gestation period.
It is known that during late gestation period, there is a progressive neuronal cell differentiation and cell growth which continues during postnatal develop- ment. A progressive increase of angiotensinogen in the hindbrain and spinal cord of developing fetus
certainly show an increase in angiotensin II for- mation as angiotensinogen degrading enzyme, renin, and angiotensin II are already reported in the fetal brain.6,7 The participation of renin-angiotensinogen system in cell differentiation and cell growth which are associated with increased DNA, RNA and hor- mone synthesis, as well as release of memory and the regulation of blood pressue are well documented in the literature.2*s”~10 In view of the fact that the fetus maintains its own independant renin-angiotensino- gen system this shows that it performs all these functions during intrauterine life.
Acknowledgements-The study was supported by a research grant from the C.I.E.S. (France), which is gratefully acknowledged. Our thanks are due to Prof. G. Grignon (Faculte de Medecine, Vandoeuvre, France) for providing research facilities. Authors are aiso grateful to Mme Bema- dette Cunin for her excellent technical assistance and to Mme Robin for hand drawings.
REFERENCES
1. Bouhnik J., Clauser E., Gardes J., Menard J. and Corvol P. (1982) Direct radioimmunoassay of rat angiotensinogen and its appIi~tion to rats in various endocrine states. Clin. Sci. 62, 335-360.
2. Morris B. J. and Reid I. A. (1978) The distribution of angiotensinogen in dog brain studied by cell fractionation. Endocrinology 103, 492-500.
3. Printz M. P., Lewicki J. A. and Wallis C. J. (1980) Enzymatic Release of Vusoactive Peptides (eds Gross F. and Vogel H. G.), pp. 193-207. Raven Press, New York.
4. Printz M. P. and Gregory T. J. (1981) Cenfral ~er~!o~ System ~ec~~isrn in ~yperze~iun (eds Buckley J. P. and Ferrario C. M.), pp. 31 l-326. Raven Press, New York.
5. Peach M. (1977) Renin-angiotensin system: biochemistry and mechanism of action. Physiol. Rev. 57, 313-370. 6. Raizada M. K., Philipps M. 1. and Grendt J. S. (1983) Primary culture from fetal brain incorporate (3H)-isoleucine
and (3H)-valine into immunorecipitable angiotensin II. Neuroendocrinology 36, 64-67. 7. Sood P. P., Panigel M. and Wegmann R. (1987) The existence of renin-an~otensin system in the rat fetal brain: I.
Immunocytochemical localization of renin-like activity at the 19th day of gestation. Cell. molec. Biol. 33, 675680. 8. Sood P. P., Richoux J. P., Panigel M., Bouhnik J. and Wegmann R. (1987) The existence of renin-angiotensinogen
system in rat fetal brain: II. Immunocytochemical localization of angiotensinogen in the telencephalon and diencephalon. CeN. molec. Biol. 33, 681490.
9. Sood P. P., Panigel M. and Wegmann R. (1989) Co-existence of renin-like immunoactivity in the rat maternal and fetal neocortex. Neurochem. Res. 14,49!-502.
10. Vecsei P., Hackenthal E. and Ganten D. (1978) The renin-angiotensin-aldosterone system. Past, present and future. Min. Wschr. 56, 5-21.
(Accepted 15 November 1989)





























