Embed Size (px)
Citation preview

Accepted Manuscript
Antigenotoxic effect, composition and antioxidant activity of Dendrobium spe‐
ciosum
Massimo Moretti, Lina Cossignani, Federica Messina, Luca Dominici, Milena
Villarini, Massimo Curini, Maria Carla Marcotullio
PII: S0308-8146(12)01533-6
DOI: http://dx.doi.org/10.1016/j.foodchem.2012.10.022
Reference: FOCH 13160
To appear in: Food Chemistry
Please cite this article as: Moretti, M., Cossignani, L., Messina, F., Dominici, L., Villarini, M., Curini, M.,
Marcotullio, M.C., Antigenotoxic effect, composition and antioxidant activity of Dendrobium speciosum, Food
Chemistry (2012), doi: http://dx.doi.org/10.1016/j.foodchem.2012.10.022
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers
we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and
review of the resulting proof before it is published in its final form. Please note that during the production process
errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

1
Antigenotoxic effect, composition and antioxidant activity of 1
Dendrobium speciosum. 2
3
Massimo Morettia, Lina Cossignani,b Federica Messinac, Luca Dominicia, 4
Milena Villarinia, Massimo Curini
c, Maria Carla Marcotullio
c,*. 5
6
aDipartimento di Specialità Medico-Chirurgiche e Sanità Pubblic -Sez. di 7
Sanità Pubblica- Università degli Studi di Perugia- Via del Giochetto- 8
06122 Perugia-Italy 9
cDipartimento di Chimica e Tecnologia del Farmaco-Sez. Chimica 10
Organica - Università degli Studi di Perugia - Via del Liceo, 1-06123 11
Perugia-Italy. 12
13
14
*Corresponding author: tel.: +39-075-5855107; fax: +39-075-5855116; 15
email: [email protected] 16
17

2
Abstract 1
The chemical composition, the antiradical properties of D. speciosum 2
(Orchidaceae) leaves and stem extracts have been studied. Furthermore, in 3
view of the use of this orchid as “bush foods”, the genotoxic/antigenotoxic 4
effects of the extracts have been evaluated. 5
6
Keywords 7
Dendrobium speciosum (Orchideaceae), genotoxicity, antigenotoxicity, 8
lipids, polyphenols, flavonoids, antioxidant, DPPH, bush foods. 9
10
11
1. Introduction 12
The relationship between diet and health has been stated throughout the 13
centuries, but only in the last decades the importance of diet in the cancer 14
onset has been pointed out. As a consequence, diet in general is considered 15
to be one of the largest source of mutagenic and carcinogenic substances for 16
human beings. Several compounds in food such as mycotoxins, 17
pyrrolizidine alkaloids, polycyclic aromatic hydrocarbons, N-nitrosamines 18
and ethanol are suspected as mutagens (Goldman & Shields, 2003). On the 19
other hand, food can contain components that decrease cancer risk (de Kok, 20
de Waard, Wilms & van Breda, 2010). 21
Dendrobium (Orchidaceae) is a large genus of orchids that contains more 22
than 1000 species. Dendrobium species are either epiphytic or occasionally 23
lithophytic. They have adapted to a wide variety of habitats, from the high 24
altitudes in the Himalayan mountains to lowland tropical forests and even to 25
the dry climate of the Australian desert. Many Dendrobium spp. have been 26

3
used for centuries as medical plants. For example D. nobile is used in China 1
for the treatment of anorexia and gastrointestinal disorders (Miyazawa, 2
Shimamura, Nakamura & Kameoka, 1997), D. chrysotoxum is used in loss 3
of appetite and fever (Yang, Chou, Wang, Guo, Hu & Xu, 2004) and D. 4
moniliforme is used as sialogogue (Zhao, Liu, Halaweish, Shao, Ye & Zhao, 5
2003). 6
Dendrobium species are known to produce alkaloids (Suzuki, Hayakawa & 7
Aoki, 1973), bibenzyl derivatives (Li et al., 2009), phenanthrenes and 8
stilbenes (Ito et al., 2010). Many secondary metabolites isolated from 9
different Dendrobium spp. show interesting biological activities, such as 10
moscatilin that has antimutagenic and antiplatelet aggregation activities 11
(Miyazawa, Shimamura, Nakamura, Sugiura, Kosaka & Kameoka, 1999; 12
Chen, Wu, Ko & Teng, 1994), or dendrobine, an antagonist of α-alanine, 13
taurine, and presynaptic inhibitor of the frog’s spinal cord (Kubo, Tanaka & 14
Yamada, 1983). 15
Dendrobium speciosum is widely distributed in Australia where it is known 16
as Wara-gal-darra or Sidney Rock Lily. The Cadigal people used to eat the 17
starchy stems raw or after roasting on hot stones (Royal Botanic Gardens & 18
Domain Trust, 2012). 19
Due to the lack of informations about D. speciosum genotoxicity, it is 20
important to evaluate the effects of this plant on genetic alterations in view 21
of an increasing interest in “bush foods” (Australian Bush Foods Magazine, 22
2012). 23
Among short-term genotoxicity tests, the comet assay is a simple, rapid and 24
very sensitive test for the quantification of DNA damage thus providing 25
direct determination of DNA single- and doublestrand breakage (Tice et al. 26

4
2000). Many authors have used the comet assay to evaluate the in vitro 1
and/or in vivo genotoxicity/antigenotoxicity of several chemicals in various 2
cell lines (CHO, V79, HepG2) (Valentin-Severin, Le Hegarat, Lhuguenot, 3
Le Bon & Chagnon, 2003). HepG2 cells are known for their phase I and II 4
biotransformation enzyme activities that influence processes related to 5
chemical carcinogenesis (Knasmüller et al., 2004). 6
The aim of the present work was to evaluate the safety of D. speciosum 7
leaves and stems (genotoxicity/antigenotoxicity) along with the 8
determination of the chemical composition, fatty acid profiles, total 9
polyphenols, total flavonoids and antiradical activity. 10
11
2. Material and Methods 12
2.1 Reagents 13
Dimethyl sulfoxide (DMSO), ethanol, ethylenediamine tetraacetic acid 14
disodium salt (Na2EDTA), hydrochloric acid (HCl), sodium chloride (NaCl) 15
and sodium hydroxide (NaOH) were purchased from Carlo Erba Reagenti 16
Srl, Milan, Italy. Ethidium bromide (EB), fluorescein diacetate (FDA), low- 17
and normal-melting-point agarose (LMPA and NMPA, respectively), 4-18
nitroquinoline N-oxide (4NQO), tris(hydroxymethyl)aminomethane (Tris), 19
and Triton X-100 were obtained from Sigma-Aldrich Srl, Milan, Italy. 20
Gibco® Minimum Essential Medium with Earle’s salts and L-glutamine 21
(MEM), antibiotics (i.e. penicillin and streptomycin), fetal bovine serum 22
(FBS), sodium pyruvate, Dulbecco’s phosphate-buffered saline, pH 7.4 23
(PBS) and trypsin were purchased from Invitrogen Srl, Milan, Italy. 24
Conventional microscope slides and coverslips were supplied by Knittel-25
Glaser, Braunschweig, Germany. Distilled water was used throughout the 26

5
experiments. Anthrone (Fluka), Folin-Ciocalteu, AlCl3, Trolox, Methyl 1
nonadecanoate (Fluka) were purchased by Sigma-Aldrich. All the solvents 2
were of analytical grade and were obtained from Carlo Erba Reagenti. 3
4
2.2 Plant Material 5
D. speciosum stems and leaves of ca. 10 years old plants, were collected on 6
June 2007 in the Botanical Garden of South Wales University (Sidney, 7
Australia). Voucher specimens of the dried leaves (152.6 g) (# MCM-2) and 8
stems (168.7 g) (# MCM-3) are deposited at the Dipartimento di Chimica e 9
Tecnologia del Farmaco-Sez. Chimica Organica of the University of 10
Perugia (Italy). 11
12
2.3 Chemical composition 13
Samples of dried leaves and stems were analysed for chemical composition 14
(protein, fat, ash and carbohydrates). The crude protein content (N x 6.25) 15
was calculated by Kjeldahl method (Chang, 2003) using the UDK 130A 16
Distillation Unit (Velp Scientifica, Usmate Milan, Italy). Total lipids were 17
determined by extraction of 5 g of grounded leaves or stems with 100 ml of 18
hexane:acetone (1:1). The resulting suspension was sonicated for 1 h. After 19
this period the suspension was filtered, evaporated under vacuum and the 20
residue weighed. The ash content was determined by incineration of 5 g of 21
sample at 550 °C for 12 h in a muffle furnace (Mod. 10-D1418/A, Controls, 22
Milan, Italy). Soluble sugars were determined spectroscopically using the 23
anthrone method (UV/VIS V530 JASCO instrument) (Marcotullio, Oball-24
Mond Mwankie, Cossignani, Tirillini & Pagiotti, 2008). The soluble sugar 25

6
content was determined and expressed as g /100 g dry weight. Sucrose was 1
used as standard. Total polysaccharides were calculated by difference. 2
All the results are expressed as g/100 g of dried weight and represent the 3
mean of three determinations ± standard deviation (SD). 4
5
2.4 Analysis of lipid fraction 6
Lipid fraction was subjected to transesterification to prepare the fatty acid 7
methyl esters (FAME) of acyl components and to evaluate the fatty acid 8
(FA) composition by successive high resolution gas chromatography 9
(HRGC) analysis. 10
Total lipids have been also subjected to thin layer chromatography on silica 11
gel plates (SIL G-25, 20x20cm, 0.25mm, Macherey-Nagel, Germany), using 12
the mixture petroleum ether/diethyl ether/formic acid (70:30:1, v/v) as 13
developing solvent. The free fatty acid (FFA) fraction (Rf≅0.45) was 14
extracted from silica with diethyl ether and subjected to esterification 15
reaction to prepare the respective FAME and to evaluate the FA 16
composition by HRGC analysis. 17
18
2.4.1 Preparation of FAME and HRGC analysis 19
Acyl lipids were transesterified using 2N KOH in methanol while free fatty 20
acids (FFA) fraction was esterified by adding an ethereal solution of 21
diazomethane. 22
HRGC analysis of FAME was performed on a Dani 1000 (Norwalk, CT, 23
USA) instrument equipped with a split-splitless injector and a flame 24
ionization detector (FID). A fused silica WCOT capillary column CP-Select 25
CB for FAME (50 m x 0.25 mm i.d., 0.25 µm f.t.; Varian, Superchrom, 26

7
Milan, Italy) was used. The injector and detector temperature was 250 °C; 1
the oven temperature was initially held at 180 °C for a 6 min, then increased 2
to 225 °C at a rate 3 °C/min; the final temperature was held for 10 min. The 3
carrier gas (He) flow rate was 1 ml/min and the split ratio was 1:70. 4
For the acquisition and integration of chromatograms Clarity (DataApex 5
Ltd., Prague, Czech Repuplic) software was used. The HRGC analyses were 6
carried out in triplicate. To confirm the identity of FAME a GC-MS QP-7
2010 (Shimadzu, Japan) was used with the same capillary column and the 8
same chromatographic conditions as those described above for HRGC-FID 9
analysis. 10
11
2.5 Determination of total phenolics and flavonoids content 12
The amount of total phenolics of D. speciosum leaves and stem extracts was 13
determined by the Folin-Ciocalteu method (Marcotullio, Oball-Mond 14
Mwankie, Cossignani, Tirillini & Pagiotti, 2008). Briefly, 1 g of grounded 15
leaves or stems were extracted with 100 ml of methanol (MeOH) at room 16
temperature for 24 h. The suspension was then filtered and diluted to 100 ml 17
with MeOH. To 1 ml of this solution, 10 ml of Folin reagent and 8 ml of a 18
7.5% Na2CO3 solution and 1 ml of H2O were added. The mixture was kept 19
for 20 min in the dark before the absorbance was measured at 765 nm 20
(UV/VIS V530 JASCO instrument) against a blank containing all the 21
reagents, except the polyphenol solution, which had been substituted with 22
distilled water. The average of three measurements was used to calculate the 23
polyphenol content. Gallic acid was used as a standard and the total 24
phenolic content was expressed as g of gallic acid equivalents per 100 g of 25
dried material. 26

8
The flavonoid content was determined spectrophotometrically using a 1
method based on the formation of a flavonoid-aluminum complex 2
(Lamaison & Carnat, 1990). Absorbance at 449 nm was read using a 3
UV/VIS V530 JASCO instrument. Total flavonoid content was expressed 4
as g of quercetin equivalents per 100 g of dried material. 5
6
2.6 DPPH test 7
Total methanolic extract of leaves or stems was prepared by extraction of 1 8
g of dry material with 100 ml of MeOH at r.t. for 24 h. After this time the 9
suspension was filtered and the filtrate evaporated under reduced pressure. 10
Fresh DPPH stock solution at a concentration of 9 mg/100 ml MeOH was 11
prepared. Methanol was used instead of the sample for blank measurements. 12
An aliquot of 1 ml of different concentrations of either standard antioxidant 13
or D. speciosum leaves or stems methanolic solution was added to 1 ml of 14
DPPH solution (Marcotullio, Oball-Mond Mwankie, Cossignani, Tirillini & 15
Pagiotti, 2008). The mixture was vortexed for a few seconds and left to 16
stand in the dark for 20 min at room temperature. The absorbance of the 17
resulting solutions was measured UV/VIS V530 JASCO spectrophotometer 18
at 517 nm against blank (without DPPH). All tests were run in triplicate and 19
the mean values calculated. Trolox (6-Hydroxy-2,5,7,8-20
tetramethylchroman-2-carboxylic acid) was used as reference 21
compound.The IC50 (concentration in mg/ml required for 50% inhibition of 22
DPPH radical) was evaluated. 23
24
2.7 Testing Dendrobium speciosum extracts for genotoxicity/ 25
antigenotoxicity 26

9
1
2.7.1 Cell culture 2
HepG2 cells (ATCC HB 8065) were obtained from Istituto Zooprofilattico 3
Sperimentale della Lombardia e dell’Emilia Romagna “Bruno Ubertini”, 4
Brescia, Italy. 5
The cells were grown as monolayer cultures in MEM supplemented with 6
10% (v/v) FBS, 1 mM sodium pyruvate, 100 U/ml penicillin and 0.1 mg/ml 7
streptomycin at 37 ºC in a humidified atmosphere containing 5% CO2. 8
HepG2 cells were subcultured by dispersal with 0.05% trypsin in 0.02% 9
Na4EDTA for a contact time of 5 min and replated at a 1:2 dilution, which 10
maintained cells in the exponential growth phase. All experiments were 11
performed on HepG2 cells at passages between 101 and 108. Cell stocks 12
were routinely frozen and stored in liquid N2. 13
14
2.7.2 Samples preparation 15
The extracts were prepared by maceration of 10 g of dry, chopped stems or 16
leaves in 500 ml of MeOH for 24 h. After this period the suspensions were 17
filtered and the filtrates were evaporated obtaining 785.2 and 412.4 mg of 18
stem and leaves extracts, respectively. 19
The solutions used in the following experimental approach were obtained by 20
dissolving the D. speciosum stem or leaves extracts in DMSO. At each 21
solvent addition, to ensure optimal solubilization of the D. speciosum 22
extracts, the solutions were subjected to mixing procedures (i.e. vortex 23
mixing and sonication). The addition of DMSO stopped when, upon visual 24
observation, the procedure resulted in a clear solution with no cloudiness or 25
precipitate. 26

10
The obtained concentrations for the D. speciosum extracts were: 200 µg/ml 1
for the steams extract and 3.4 mg/mL for the leaves extract. 2
3
2.7.3 Lactate dehydrogenase (LDH) leakage assay 4
The D. speciosum stem or leaves extracts concentrations to be tested for 5
genotoxicity/antigenotoxicity were selected from preliminary cytotoxicity 6
experiments. Cytotoxicity induced by the extracts was assessed by lactate 7
dehydrogenase (LDH) release into the culture medium. For cytotoxicity 8
testing, HepG2 cells (1.25×105/well) were seeded in 96-well tissue culture 9
plates (Orange Scientific, Braine-l’Alleud, Belgium) and were allowed to 10
attach for 24 h before treatment with D. speciosum extracts. The medium 11
was then removed and replaced by fresh MEM containing a range of stems 12
(0.078 to 10 µg/ml) or leaves (13.3 to 1.700 µg/ml) extracts concentrations 13
obtained by twofold serial dilutions. The highest concentrations considered 14
in the LDH leakage assay corresponded to the maximum possible volumes 15
of extracts in DMSO that could be possible to test. The cells were then 16
treated for 4 h. Following the treatment the plates were centrifuged at 250×g 17
for 10 min in order to obtain a cell free supernatant and the culture medium 18
was aspirated. The activity of LDH in the medium was determined using 19
Takara's LDH Cytotoxicity Detection Kit according to manufacturer's 20
instructions. LDH leakage assay is based on the conversion of lactate to 21
pyruvate in the presence of LDH with parallel reduction of NAD+ on a 22
coupled reaction which converts a yellow tetrazolium salt into a red, 23
formazan-class dye. The formation of formazan from the above reaction 24
results in a change in absorbance at 492 nm. Absorbance was recorded using 25
a Tecan Sunrise microplate reader (Tecan Italia Srl, Milan, Italy). All tests 26

11
were performed at least in triplicate and repeated twice to calculate the 1
amount of LDH released. The results of LDH leakage assay determined the 2
choice of three concentrations (i.e. 2.5, 5 and 10 µg/ml for the stems extract 3
and 25, 50 and 100 µg/ml for the leaves extract) of D. speciosum extracts to 4
be evaluated afterwards, in the genotoxicity/antigenotoxicity protocols. 5
6
2.7.4 Genotoxicity/antigenotoxicity testing 7
Treatments for genotoxicity/antigenotoxicity testing were carried out in 8
HepG2 cells subcultured in 6-well tissue culture plates (Orange Scientific, 9
Belgium) inoculated with 5 ml of complete MEM containing 5×105 cells per 10
well. The overall culture length was 52 h throughout the experiments. After 11
seeding for 48 h, the culture medium was replaced by fresh complete MEM 12
containing different concentrations of stem or leaves extracts (2.5 to 10 13
µg/ml for the stem extract and 25 to 100 µg/ml for the leave extract). The 14
cells were then incubated further for 4 h according to the following scheme: 15
(a) Dendrobium speciosum stems (2.5 to 10 µg/ml) or leaves (25 to 100 16
µg/ml) extract, to check the absence of genotoxicity; (b) direct acting 17
mutagen 4NQO (3 µM); and (c) simultaneous exposure to 4NQO and 18
Dendrobium speciosum stem or leaves extract. Negative controls (1% 19
DMSO) were also included. 20
At the end of treatments, the cells were washed twice with 5 ml ice-cold 21
PBS, pH 7.4, and detached with 300 µl of 0.05% trypsin in 0.02% 22
Na4EDTA. After 3 min, trypsinization was stopped by adding 700 µl 23
complete culture medium. Cells were then collected by centrifugation (70×g, 24
8 min, 4 °C). Each treatment protocol was carried out in duplicate. 25
26

12
2.7.5 Fluorochrome-mediated viability test 1
At the end of the treatments, cytotoxic effects were evaluated in culture 2
aliquots by simultaneous staining with FDA and EB (Jones & Senft, 1985). 3
Dye working solutions were prepared immediately prior to use by adding 20 4
µl of 5 mg/ml FDA (in acetone) to 5 ml of PBS, or 20 µl of 20 µg/ml EB to 5
1 ml of PBS. Subsequently, 100 µl of 20 µg/ml FDA and 30 µl of 0.4 µg/ml 6
EB were added directly to 200 µl of each cell suspension. The cells were 7
then stained for 3 min at room temperature. Cell counts were performed 8
using ten-chamber disposable microscope slides provided with a 9
haemocytometer-like counting grid (Kova, Hycor Biomedical, USA). After 10
the counting chamber was loaded, the cells were examined with a standard 11
fluorescence microscope (Dialux 20, Leitz, Germany) equipped with epi-12
illumination provided by a 50 W high-pressure mercury lamp (HBO 50, 13
Osram, Germany). Viable cells fluoresced green, whereas dead cells were 14
indicated by orange-stained nuclei. 15
16
2.7.6 Alkaline single-cell microgel-electrophoresis (comet) assay 17
Immediately after the exposure, HepG2 cells were processed in the comet 18
assay under alkaline conditions (alkaline unwinding/alkaline electrophoresis, 19
pH > 13), basically following the original procedure (Singh, McCoy, Tice & 20
Schneider, 1988; Tice et al., 2000), with minor modifications (Moretti et al., 21
2005; Zampini, Villarini, Moretti, Dominici & Isla, 2008). Briefly, cell 22
pellets were gently resuspended in LMPA (0.7% in Ca++/Mg++-free PBS, 23
w/v) maintained at 37 °C. Then, 65 µl of cell suspension in agarose were 24
rapidly layered onto pre-coated (1% NMA in Ca++
/Mg++
-free PBS) 25
conventional 26×76 mm microscope slides and covered with a coverslip. 26

13
After brief agarose solidification at 4 °C, the coverslips were removed and 1
the cell containing microgels covered with a top layer (75 µl) of 0.7% 2
LMPA. The coverslips were further removed and the slides immersed in 3
cold, freshly prepared lysing solution (2.5 M NaCl, 100 mM Na2EDTA, 10 4
mM Tris–HCl and NaOH to pH 10; 1% Triton X-100 added just before use) 5
for at least 60 min at 4 ºC to obtain the lysis of cellular and nuclear 6
membranes of the cells embedded in agarose microgels. 7
The slides were then drained and placed in a horizontal electrophoresis box 8
(HU20, Scie-Plas, Cambridge, UK) filled with a freshly prepared 9
electrophoresis solution (10 mM Na2EDTA, 300 mM NaOH; pH>13). After 10
20 min of pre-electrophoresis to allow DNA unwinding and expression of 11
alkali-labile damage, electrophoresis runs were performed in an ice bath for 12
20 min by applying an electric field of 25V (1 V/cm) and adjusting the 13
current to 300 mA (Power Supply PS250, Hybaid, Chesterfield, MO, USA). 14
The microgels were then neutralized with 0.4 M Tris-HCl buffer (pH 7.5). 15
For preservation, the slides were dehydrated in 70% ethanol (10 min), 16
allowed to air-dry and store in slide boxes at room temperature until ready 17
to stain and analyze. All the steps of the comet assay were conducted in 18
yellow light to prevent the occurrence of additional DNA damage. 19
Immediately before scoring, the air-dried slides were stained with 65 µl of 20
EB (20 µg/ml) and were covered with a coverslip. The comets in each 21
microgel were analyzed (blind), at 500×g magnification, with an 22
epifluorescent microscope (BX41, Olympus, Japan) under a 100 W high-23
pressure mercury lamp (HSH-1030-L, Ushio, Japan), using appropriate 24
optical filters (excitation filter 510–550 nm and emission filter 590 nm). The 25
microscope, equipped with a high sensitivity black and white CCD camera 26

14
(PE2020, Pulnix, UK), was connected to a computerized analysis system 1
(“Comet Assay III”, Perceptive Instruments, UK) that acquires images, 2
computes the integrated intensity profile for each cell, estimates the comet 3
cell components, head and tail, and evaluates a range of derived parameters. 4
These include: tail length (measured from the head centre, expressed in µm), 5
tail intensity (percent of fluorescence in the comet tail), and tail moment, a 6
composite parameter in which the migration distance and the amount of 7
migrated DNA (by analogy with the mechanical term) are expressed as a 8
single value. A total of 100 randomly selected comets (50 cells/replicate 9
slides) were evaluated for each experimental point. 10
11
2.7.7 Statistical analysis 12
Cytotoxicity and genotoxicity/antigenotoxicity data are presented as the 13
mean (± SEM) of triplicate tests. Statistical significance was tested by one-14
way analysis of variance (ANOVA). If a significant F-value (p < 0.05) was 15
obtained, to examine where the differences actually occurred multiple 16
comparisons were conducted by Bonferroni post hoc analysis. Dose-17
response relationships were established by applying linear regression 18
analysis to the experimental data. 19
Lastly, the inhibition rate (IR%) of Dendrobium speciosum extracts against 20
4NQO-mediated genotoxicity was calculated according to the following 21
formula: 22
IR%= [1-(A-C/B-C)] x 100 23
where A corresponds to the extent of DNA damage observed in cells 24
subjected to the antigenotoxic treatment (co-exposure to Dendrobium 25
speciosum extracts plus the known mutagen), B corresponds to DNA 26

15
damage observed in 4NQO exposed cells and C corresponds to DNA 1
damage extent in the negative control. 2
3
3. Results and Discussion 4
5
3.1 Composition of D. speciosum leaves and stems. 6
The results of the chemical composition for Dendrobium speciosum leaves 7
and stems are reported in Table 1. 8
It is evident that leaves were richer in lipids, ash and soluble carbohydrates 9
than stems, which had higher polysaccharide content. Protein amount was 10
almost equivalent in leaves and stems. The FA composition of lipid 11
fractions was determined by HRGC analysis. Table 2 shows the FA 12
percentage composition of acyl lipid and FFA fraction, respectively, 13
analysed as methyl esters. 14
The FA composition in the two fractions (free and esterified) was extremely 15
different. In the acyl lipid fraction a small percentage of capric (10:0) and 16
lauric (12:0) acids was present. In the FFA fraction, SFA were present in 17
higher percentage both in leaves and stems (61.8 and 87.7 %, respectively) 18
than in the esterified fraction (47.5 and 43.8 %, respectively). The latter 19
accounted for a higher amount of UFA (52.5 and 56.2 % in leaves and stems, 20
respectively). In particular, stems acyl lipid fraction showed higher 21
percentage of oleic acid (26.2%) than the other fractions while linoleic acid 22
content was generally high (25.5-27.7%) with the exception of stems FFA 23
fraction (8.9%). α-Linolenic acid content was higher in acyl lipids of leaves 24
than in the other fractions. D. speciosum leaves lipids contain higher 25
amounts of essential FA than stems lipids. 26

16
By spectrophotometric analysis of the methanolic extract of leaves and 1
stems we determined a good percentage of polyphenols (1.15±0.10 and 2
1.06±0.12, respectively) and flavonoids resulted to be 0.21±0.08 % and 3
0.12±0.07%, respectively. 4
The DPPH scavenging activities of leaves and stem methanolic extract 5
expressed as IC50 were 0.026±0.004 mg/ml and 1.054±0.047 mg/ml, 6
respectively. Trolox was used as reference compound (IC50 0.020±0.003 and 7
0.014±0.001 mg/ml for leaves and stems, respectively). According these 8
data, D. speciosum leaves extract was a more efficient free radical scavenger 9
by the lower IC50 value than stems extract and its activity is comparable 10
with Trolox activity. The greater radical scavenging activity of D. 11
speciosum leaves can be ascribed to the higher content of polyphenols. 12
13
3.2 Cytotoxicity (LDH leakage) 14
LDH leakage in the medium was determined in order to evaluate cell 15
injuries influenced by D. speciosum extracts (data not shown). The results of 16
LDH leakage assay determined the choice of three concentrations (i.e. 2.5, 5 17
and 10 µg/ml for the stem extract and 25, 50 and 100 µg/ml for the leaves 18
extract) of D. speciosum extracts to be evaluated afterwards, in the 19
genotoxicity/antigenotoxicity protocols. 20
21
3.3 Genotoxicity/antigenotoxicity (Comet assay) 22
Table 3 shows the extent of DNA damage in HepG2 cells exposed 23
according to protocols for genotoxicity (i.e. stem or leaves extract) or 24
antigenotoxicity (i.e. simultaneous exposure to 4NQO and stems or leaves 25
extract) testing. 26

17
D. speciosum stems extract did not induce DNA strand breaks in the range 1
of concentration tested, with mean tail intensity values obtained being 2
similar to those for the negative control. Whereas, at the higher 3
concentration tested (i.e. 100 µg/ml) the leaves extract caused a significant 4
increase in the extent of DNA damage when compared with the negative 5
control, thus indicating a genotoxic effect. 4NQO used as known mutagen 6
(positive control) demonstrated the sensitivity of the comet assay and 7
yielded a clear positive response at the concentration used. 8
As regard antigenotoxicity testing, any significant reduction of 4NQO-9
induced DNA damage was observed for the leaves extract at the 10
concentration range tested. Conversely, simultaneous treatment of cells 11
showed a reduction in the extent of DNA damage for cultures treated with 12
the stems extract. The antigenotoxic effect toward the extent of 4NQO-13
induced DNA damage increased with an inverse dose-response correlation 14
(IR% = 10.11, 22.09 an 44.34 for the concentrations of 10.0, 5.0 and 2.5 15
µg/mL, respectively; r2 = 0.963), being the lowest concentration the most 16
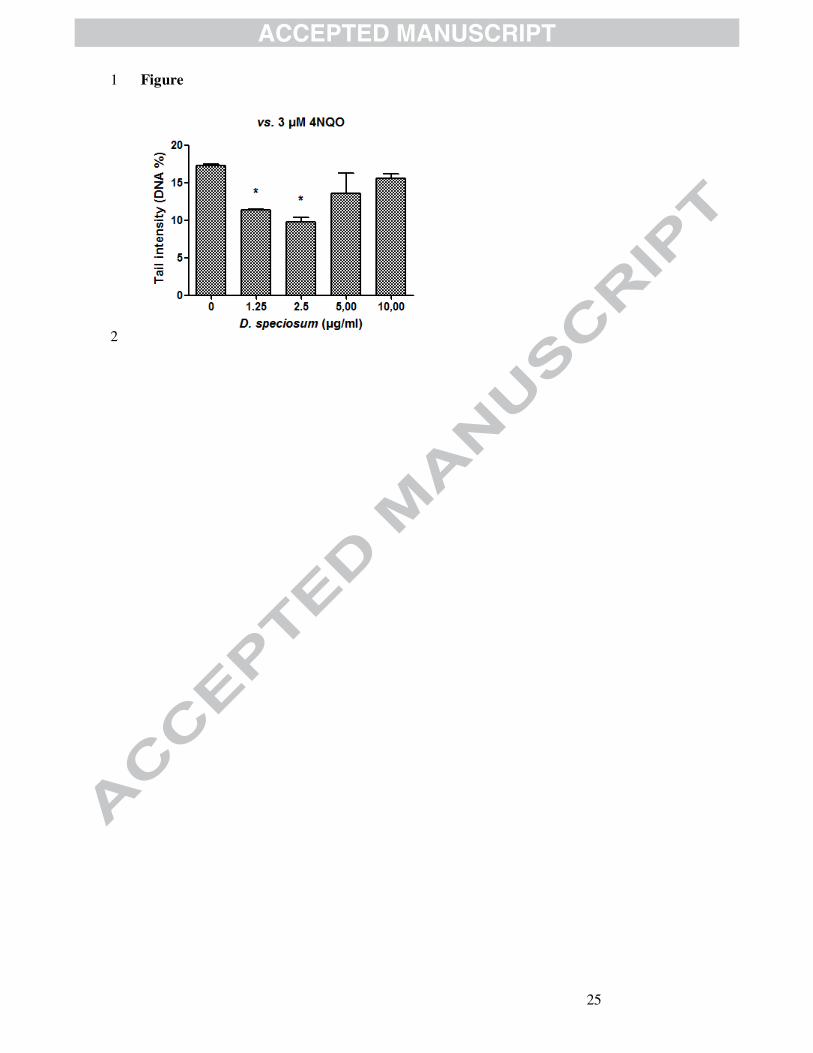
effective. To better evaluate the tendency of the effect, the stem extract was 17
further evaluated at 1.25 µg/ml (Figure 1). We observed a J-shaped 18
concentration-response curve of residual DNA damage with the highest 19
effect at the dose of 2.5 µg/ml. 20
In the present approach, genotoxicity/antigenotoxicity testing has been 21
performed on the HepG2 cell line. HepG2 cells retain many of the 22
morphological characteristics of the liver parenchymal cells from which 23
they originate (Knowles, Howe & Aden, 1980) and present endogenous 24
bioactivation capacity expressing phase I and phase II enzymes involved in 25
the activation and/or detoxification of xenobiotics (Diamond, Kruszewski, 26

18
Aden, Knowles & Baird, 1980; Sassa, Sugita, Galbraith & Kappas, 1987). 1
Thus, the metabolically competent HepG2 cell line is considered to be an 2
excellent model to investigate in vitro the toxicity of drugs, being this model 3
closer to the in vivo situation as the addition of an exogenous metabolic 4
activation system (i.e. S9-mix) (Knasmüller et al. 1998; Knasmüller et al. 5
2004). 6
DNA-protective substances can be classified as desmutagens or 7
bioantimutagens (Kada, Inoue & Namiki, 1982), being desmutagenic agents 8
compounds that interact directly with mutagens, or their precursors, in order 9
to inactivate them. These agents interact with a mutagenic compound in an 10
irreversible way, inactivating it chemically, usually through a direct link. By 11
applying the simultaneous protocol, the Dendrobium speciosum stems 12
extract showed an antigenotoxic activity toward the model mutagen 4NQO, 13
even though at low concentration, likely acting as a desmutagenic agent. 14
When testing the Dendrobium speciosum stems extract for antigenotoxiciy, 15
we observed an hormetic-like, J-shaped dose-response curve (Calabrese & 16
Baldwin, 2002), where low doses result in enhanced activity, whereas 17
higher doses result in no effect (Figure 1). 18
This study evaluated crude extract of stems of D. speciosum in view to state 19
if their use as bush food is safe. Traditional use interests only starchy stems 20
that showed to be not genotoxic and to posses an antimutagenic activity. On 21
the other hand, the leaves extract showed to be slightly genotoxic, even if at 22
very high concentrations. From a nutritional point of view both stems and 23
leaves are characterized by low protein and lipid content. Stem acyl lipids 24
showed similar percentage of palmitic, oleic and linoleic acid with a 25
balanced ratio of linoleic to α-linolenic acid. 26

19
The stem extract showed a small amount of polyphenols and flavonoids that 1
accounts for the little antioxidant activity of the extract. 2
From these results we can conclude that the use of D. speciosum stems as 3
“bush foods” is safe. 4
5
Acknowledgements The authors acknowledge Dr. R. Mammucari and Dr. 6
H. Wildman for providing the plant material and Prof. B. Tirillini for the 7
DPPH test experiments. 8
9
10
References 11
12
Australian Bush Foods Magazine. http://www.ausbushfoods.com/. Accessed 13
05.24.12. 14
Calabrese, E. J., & Baldwin, L.A. (2002). Defining hormesis. Human & 15
Experimental Toxicology, 21, 91-97. 16
Chang, S. K. C. (2003). Protein analysis. In: Nielsen S. S., Food Analysis 17
3rd
ed., Kluwer Academic, New York, 131-142. 18
Chen, C.-C., Wu, L.-G., Ko, F.-N., & Teng, C.-M. (1994). Antiplatelet 19
aggregarion principles of Dendrobirm loddigesii. Journal of Natural 20
Products, 57, 1271-1274. 21
de Kok, T. M. C. M., de Waard, P., Wilms, L. C., & van Breda, S. G. J. 22
(2010). Antioxidative and antigenotoxic properties of vegetables and dietary 23
phytochemicals: the value of genomics biomarkers in molecular 24
epidemiology. Molecular Nutrition & Food Research, 54, 208-217. 25

20
Diamond, L., Kruszewski, F., Aden, D. P., Knowles, B. B., Baird, W. M. 1
(1980). Metabolic activation of benzo[a]pyrene by a human hepatoma cell 2
line. Carcinogenesis, 1, 871-875. 3
Goldman, R., & Shields, P. G. (2003). Food mutagens. Journal of Nutrition, 4
133, 965S–973S. 5
Ito, M., Matsuzaki, K., Wang, J., Daikonya, A., Wang, N.-L., Yao, X.-S., & 6
Kitanaka, S. (2010). New phenanthrenes and stilbenes from Dendrobium 7
loddigesii. Chemical & Pharmaceutical Bulletin, 58, 628-633. 8
Jones, K. H., & Senft, J. A. (1985). An improved method to determine cell 9
viability by simultaneous staining with fluorescein diacetate-propidium 10
iodide. Journal of Histochemistry & Cytochemistry, 33, 77-79. 11
Kada, T., Inoue, T., & Namiki, N. (1982). Environmental desmutagens and 12
antimutagens. In: Klekowski EJ (ed) Environmental Mutagenesis and Plant 13
Biology. Praeger, New York, pp 137-151. 14
Knasmüller, S., Mersch-Sundermann, V., Kevekordes, S., Darroudi, F., 15
Huber, W. W., Hoelzl, C., Bichler, J., & Majer, B. J. (2004). Use of human-16
derived liver cell lines for the detection of environmental and dietary 17
genotoxicants; current state of knowledge. Toxicology, 198, 315–328. 18
Knasmüller, S., Parzefall, W., Sanyal, R,. Ecker, S., Schwab, C., Uhl, M., 19
Mersch-Sundermann, V., Williamson, G., Hietsch, G., Langer, T., Darroudi, 20
F., & Natarajan, A. T. (1998). Use of metabolically competent human 21
hepatoma cells for the detection of mutagens and antimutagens. Mutation 22
Research, 402, 185-202. 23
Knowles, B.B., Howe, C. C., & Aden, D. P. (1980). Human hepatocellular 24
carcinoma cell lines secrete the major plasma proteins and hepatitis B 25
surface antigen. Science, 209, 497-499. 26

21
Kubo, Y., Tanaka, A., & Yamada, K. (1983). Dendrobine, an antagonist of 1
α-alanine, taurine and of presynaptic inhibition in the frog spinal cord. 2
British Journal of Pharmacology, 78, 709-715. 3
Lamaison, J.L., & Carnat, A. (1990). The amount of main flavonoids in 4
flowers and leaves of Crataegus monogyna Jacq. and Crataegus laevigata 5
(Poiret) DC. (Rosaceae). Pharmaceutica Acta Helvetiae, 65, 315-320. 6
Li, Y., Wang, C. L., Wang, Y. J., Wang, F. F., Guo, S. X., Yang, J. S., & 7
Xiao, P. G. (2009). Four new bibenzyl derivatives from Dendrobium 8
candidum. Chemical & Pharmaceutical Bulletin, 57, 997-999. 9
Marcotullio, M.C., Oball-Mond Mwankie, G.N., Cossignani, L., Tirillini, B., 10
& Pagiotti, R. (2008). Phytochemical Analysis and Antiradical Properties of 11
Sarcodon imbricatus (L.:Fr) Karsten. Natural Product Communications, 3, 12
1907-1910. 13
Miyazawa, M., Shimamura, H., Nakamura, S., & Kameoka, H. (1997). 14
Antimutagenic activity of gigantol from Dendrobium nobile. Journal of 15
Agricultural and Food Chemistry, 45, 2849-2853. 16
Miyazawa, M., Shimamura, H., Nakamura, S., Sugiura, W., Kosaka, H., & 17
Kameoka, H. (1999). Moscatilin from Dendrobium nobile, a naturally 18
occurring bibenzyl compound with potential antimutagenic activity. Journal 19
of Agricultural and Food Chemistry, 47, 2163-2167. 20
Moretti, M., Villarini, M., Simonucci, S., Fatigoni, C., Scassellati-Sforzolini, 21
G,. Monarca, S., Pasquini, R., Angelucci, M., & Strappini, M. (2005). 22
Effects of co-exposure to extremely low frequency (ELF) magnetic fields 23
and benzene or benzene metabolites determined in vitro by the alkaline 24
comet assay. Toxicology Letters, 157, 119-128. 25

22
RBGSYD (The Royal Botanic Gardens & Domain Trust, Sydney) 1
http://www.rbgsyd.nsw.gov.au/education/Resources/bush_foods/Dendrobiu2
m_speciosum. Accessed 05.24.12. 3
Sassa, S., Sugita, O., Galbraith, R. A., & Kappas, A. (1987). Drug 4
metabolism by the human hepatoma cell, Hep G2. Biochemical and 5
Biophysical Research Communications, 143, 52-57. 6
Singh, N. P., McCoy, M. T., Tice, R. R., & Schneider, E.L. (1988). A 7
simple technique for quantitation of low levels of DNA damage in 8
individual cells. Experimental Cell Research, 175, 184-191. 9
Suzuki, M., Hayakawa, Y., & Aoki, K. (1973). Stereochemistry of 10
intermediates in the syntheses of Dendrobium alkaloids. Tetrahedron 11
Letters, 4, 331-334. 12
Tice, R.R., Agurell, E., Anderson, D., Burlinson, B., Hartmann, A, 13
Kobayashi, H., Miyamae, Y., Rojas, E., Ryu, J. C., & Sasaki, Y.F. (2000). 14
Single cell gel/comet assay: guidelines for in vitro and in vivo genetic 15
toxicology testing. Environmental and Molecular Mutagenesis, 35, 206-221. 16
Valentin-Severin, I., Le Hegarat, L., Lhuguenot, J. C., Le Bon, A. M., & 17
Chagnon, M.C. (2003). Use of HepG2 cell line for direct or indirect 18
mutagens screening: comparative investigation between comet and 19
micronucleus assays. Mutation Research, 536, 79–90. 20
Yang, H., Chou, G.-X., Wang, Z.-T., Guo, Y.-W., Hu, Z.-B., & Xu, L.-S. 21
(2004). Two new compounds from Dendrobium chrysotoxum. Helvetica 22
Chimica Acta, 87, 394–399. 23
Zampini, I. C., Villarini, M., Moretti, M., Dominici, L., & Isla, M.I. (2008). 24
Evaluation of genotoxic and antigenotoxic effects of hydroalcoholic extracts 25
of Zuccagnia punctata Cav. Journal of Ethnopharmacology, 115, 330-335. 26

23
Zhao, C., Liu, Q., Halaweish, F., Shao, B., Ye, Y., & Zhao, W. (2003). 1
Copacamphane, picrotoxane, and alloaromadendrane sesquiterpene 2
glycosides and phenolic glycosides from Dendrobium moniliforme. Journal 3
of Natural Products, 66, 1140–1143. 4

24
Figure captions. 1
2
Figure 1. Antigenotoxic effects of Dendrobium speciosum stem extract on 3
4NQO-induced DNA damage in HepG2 cells. Each result is expressed as 4
the mean ± standard error (SEM) of three independent experiments. 5
Statistical significance (ANOVA, followed by Bonferroni post hoc analysis: 6
* p < 0.05. 7
25
Figure 1
2

26
Tables 1
Table 1: Chemical composition of D. speciosum leaves and
stemsa
Leaves Stems
Crude protein 6.90±0.30 6.10±0.25
Total lipids 3.60±0.28 2.20±0.19
Ash 8.55±0.21 5.00±0.42
Soluble carbohydrates 0.80±0.11 0.45±0.09
Total polysaccharidesb 80.15±0.29 86.25±0.34
aData are given as mean ± SD (n=3) and are expressed as g/100g dry weight.
bThe value is obtained by difference and includes lignin content.
2
Table 2: Fatty acid compositiona of lipid components
b of
D. speciosum leaves and stems
Acyl lipids FFA fraction
Leaves Stems Leaves Stems
C 10:0 1.3±0.2 tr tr tr
C 12:0 8.5±0.6 3.8±0.2 tr tr
C 14:0 4.1±0.2 1.5±0.1 0.5±0.0 0.9±0.1
C 15:0 2.4±0.1 1.8±0.2 3.2±0.3 7.3±0.2
C 16:0 20.8±0.7 25.4±1.2 37.0±0.9 59.1±2.5
C 17:0 3.1±0.2 2.5±0.3 7.0±0.2 9.4±0.8
C 18:0 7.3±0.2 8.8±0.4 14.1±0.5 11.0±0.6
C 18:1n-9 8.9±0.3 26.2±0.9 6.5±0.3 1.3±0.1
C 18:2n-6 27.7±1.1 25.5±1.1 25.6±1.3 8.9±0.4
C 18:3n-3 15.9±0.4 4.5±0.2 6.1±0.3 2.1±0.2
SFA 47.5 43.8 61.8 87.7
MUFA 8.9 26.2 6.5 1.3
PUFA 43.6 30.0 31.7 11.0
amean ± SD (n=3).
bAnalysed as methyl esters C10:0 (capric acid);
C12:0 (lauric acid); C14:0 (myristic acid); C15:0 (pentadecanoic
acid); C16:0 (palmitic acid); C17:0 (heptadecanoic acid); C18:0
(stearic acid); C18:1n-9 (oleic acid); C18:2n-6 (linoleic acid);
C18:3n-3 (α-linolenic acid); saturated fatty acids (SFA);
monounsaturated fatty acids (MUFA); polyunsaturated fatty acids
(PUFA); tr <0.1%.
3

27
Table 3: DNA damage in HepG2 liver cells treated in vitro with D. speciosum extracts
alone (genotoxicity testing) or subjected to simultaneous exposure to test compounds and
model mutagens 4NQO (3µM) (antigenotoxicity testing).a
Tested
doses
(µg/ml)
Leaves Tested
doses
(µg/ml)
Stems
Controlb 2.33 ± 0.50
0.42 ± 0.32
4NQOc 14.26 ± 1.67 17.24 ± 0.24
Genotoxicity assessment
D. speciosum extract 25 4.11 ± 0.67 2.5 0.53 ± 0.08
50 3.82 ± 0.09 5.0 0.14 ± 0.01
100 5.57 ± 0.26* 10.0 1.92 ± 1.09
Antigenotoxicity assessment
D. speciosum extract + 4NQOc 25 14.88 ± 0.94 2.5 9.78 ± 0.60
50 14.50 ± 0.82 5.0 13.52 ± 2.70
100 15.35 ± 0.43 10.0 15.54 ± 0.65
a Each result is expressed as the mean ± standard error (SEM) of three independent experiments.
b 1% DMSO.
c 4-nitroquinoline N-oxide (3 µM), positive control.
1
2
3

28
Highlights 1
1. Dendrobium speciosum (Orchidaceae) as bush food 2
2. Evaluation of genotoxicity and antigenotoxicity of the leaves and stems 3
extracts 4
3. Chemical composition to evaluate nutritional value (proteins, lipids and 5
saccharides) 6
4. Polyphenols and flavonoids and their antiradical activity 7
8
9
10





























