Embed Size (px)
Citation preview

Apoptosis-like death in Leishmania donovanipromastigotes induced by eugenol-rich oil ofSyzygium aromaticum
Mohammad Islamuddin,1 Dinkar Sahal2 and Farhat Afrin1
Correspondence
Farhat Afrin
Received 25 June 2013
Accepted 24 October 2013
1Parasite Immunology Laboratory, Department of Biotechnology, Jamia Hamdard (HamdardUniversity), New Delhi 110 062, India
2International Centre for Genetic Engineering and Biotechnology, Aruna Asaf Ali Marg,New Delhi 110 067, India
Leishmaniasis consists of a complex spectrum of infectious diseases with worldwide distribution
of which visceral leishmaniasis or kala-azar caused by Leishmania donovani is the most
devastating. In the absence of vaccines, chemotherapy remains the mainstay for the control of
leishmaniasis. The drugs of choice are expensive and associated with multiple adverse side
effects. Because of these limitations, the development of new antileishmanial compounds is
imperative and plants offer prospects in this regard. The present work was conducted to study the
antileishmanial potential of oil from Syzygium aromaticum flower buds (clove). The S. aromaticum
oil was characterized by gas chromatography and GC-MS and eugenol as well as eugenyl
acetate were found to be the most abundant compounds, composing 59.75 % and 29.24 %,
respectively of the oil. Our findings have shown that eugenol-rich essential oil from S. aromaticum
(EROSA) possesses significant activity against L. donovani, with 50 % inhibitory concentration of
21±0.16 mg ml”1 and 15.24±0.14 mg ml”1, respectively, against promastigotes and intracellular
amastigotes. Alterations in cellular morphology and growth reversibility assay substantiated the
leishmanicidal activity of EROSA. The leishmanicidal effect was mediated via apoptosis as
confirmed by externalization of phosphatidylserine, DNA nicking by TdT-mediated dUTP nick-end
labelling (TUNEL) assay, dyskinetoplastidy, cell cycle arrest at sub-G0–G1 phase, loss of
mitochondrial membrane potential and reactive oxygen species generation. EROSA presented no
adverse cytotoxic effects against murine macrophages even at 200 mg ml”1. Our studies
authenticate the promising antileishmanial activity of EROSA, which is mediated by programmed
cell death, and, accordingly, EROSA may be a source of novel agents for the treatment of
leishmaniasis.
INTRODUCTION
Leishmaniasis is a parasitic disease caused by at least 17protozoan species of the Leishmania genus (Croft &Coombs, 2003). The forms of the disease are related tothe type of parasite and differ in geographical distribution,the host and vector involved, incidence rate and mortality(Ashford et al., 1992). Visceral leishmaniasis, or kala-azar,caused by Leishmania donovani is the most deadly form ofleishmaniasis, affecting millions of people around theworld. The toxicity of the present drugs and the emergenceof strains that are not responsive to drug therapy have
made the discovery of novel therapeutic agents a priority(Iwu et al., 1994). In this regard, plants are very importantsources of diverse types of bioactive organic compounds,including the complex group of essential-oil-related com-pounds. Essential-oil constituents contribute to beneficialor adverse effects based on their characteristic biologicalactivity. Essential oils are known to possess antibacterialand antifungal properties against human pathogenicmicro-organisms (Fyfe et al., 1998). Antiprotozoal activ-ities of essential oils have also been extensively reviewed(Perez et al., 2012). Essential oils obtained from Achilleamillefolium and Ocimum basilicum (Santoro et al., 2007)exhibited toxic effects against trypanosomes, which areprotozoal species closely related to Leishmania. A plethoraof studies indicate promising leishmanicidal effects ofessential oils from various plants, including Croton cajucara(Rosa et al., 2003), Ocimum gratissimum (Ueda-Nakamura
Abbreviations: EROSA, eugenol-rich essential oil from S. aromaticum;FBS, fetal bovine serum; GI50, 50 % growth inhibitory concentration;IC50, 50 % inhibitory concentration; PI, propidium iodide; PS,phosphatidylserine; ROS, reactive oxygen species; TUNEL, TdT-mediated dUTP nick-end labelling.
Journal of Medical Microbiology (2014), 63, 74–85 DOI 10.1099/jmm.0.064709-0
74 064709 G 2014 SGM Printed in Great Britain

et al., 2006), Artemisia herba-alba (Mohamed et al., 2010),Eugenia uniflora (Rodrigues et al., 2013) and Origanumvulgare (Sanchez-Suarez et al., 2013).
The Syzygium genus is one of 75 genera (about 3000species) belonging to the Myrtaceae family. Syzygiumaromaticum, commonly known as clove, is native totropical America and Australia (Mishra & Singh, 2008).Clove oil has been widely investigated due to its popularity,availability and high essential-oil content. The clove oil ofS. aromaticum (syn. Eugenia caryophyllata) has long beenconsidered to have medicinal properties (Chaieb et al.,2007). It is reported to have antibacterial (Burt & Reinders,2003) and antitrypanosomal activities (Santoro et al.,2007). The flower bud extract of S. aromaticum displayedantimalarial potency even in its crude form (Bagavan et al.,2011). Based on the reported biological activities, in thepresent study we sought to determine the in vitroleishmanicidal potential of essential oil obtained from S.aromaticum L. against L. donovani.
METHODS
Materials. Fetal bovine serum (FBS) was from Gibco-BRL, DMSOfrom SRL, methanol from Merck, and annexin V-FITC and the Apo-Direct kits from Roche. All the other chemicals were from Sigma-Aldrich unless stated.
Parasite and cell culture. L. donovani parasites, strain MHOM/IN/83/AG83, were maintained in vivo in BALB/c mice. Promastigoteswere cultured in M199 medium supplemented with 10 % heat-inactivated FBS, penicillin G sodium (100 U ml21) and streptomycinsulfate (100 mg ml21) at 22 uC and subcultured every 72 h in thesame medium at a mean density of 26106 cells ml21 (Afrin et al.,2001).
The RAW264.7 cell line was grown at 37 uC in RPMI 1640 medium(pH 7.4) supplemented with 10 % heat-inactivated FBS for 48–72 hin 5 % CO2 and subcultured in fresh RPMI 1640 medium at a meandensity of 26105 cells ml21.
Plant material. S. aromaticum L. (Myrtaceae) flower buds werepurchased from the local market of Khanpur, New Delhi. The plantwas authenticated by Dr H. B. Singh, taxonomist, NISCAIR, CSIR,New Delhi (voucher no. 2012-2013/2065/73).
Extraction of oil from S. aromaticum. The essential oil of the S.aromaticum flower buds was extracted with a modified Clevenger-type apparatus (borosilicate), using the steam distillation technique(Gottlieb & Guimaraes, 1960). After extraction, the essential oilobtained was carefully separated and stored in opaque glass vials withrubber lids, covered with aluminium foil to protect the contents fromlight, in a refrigerator (220 uC) prior to analysis and biologicalassays. Stock solutions of 1 mg ml21 were prepared aseptically inDMSO (cell culture grade) and diluted further in culture medium toachieve a final DMSO concentration of not more than 0.2 %.
GC and GC-MS analysis of oil from S. aromaticum. The chemicalcomposition of the active compounds from S. aromaticum flower budoil was analysed by GC and GC-MS using a Shimadzu QP2010 with aDB-5 column (30 m, film 0.25 mm, internal diameter 0.25 mm). Thetemperature of the column was programmed from 45 uC to 270 uC at5 uC min21, the injector and detector temperature for the analysiswas 250 uC. Helium was used as the carrier gas at a flow rate of 1.5 ml
min21. The mass spectrometer was operated in electron impactionization mode at 70 eV. The chemical constituents of the oil wereidentified after correlation of their recorded mass spectra with thoseobtained from the WILEY8.LIB and NIST08.LIB library spectra providedby the software of the GC-MS system (Rosa et al., 2003), and eugenolwas found to be the major constituent.
Antileishmanial activity of eugenol-rich oil of S. aromaticum
(EROSA) in a growth kinetics assay. Promastigotes of L. donovani(26106 cells ml21) were incubated at 22 uC in the presence of 100 mgml21 EROSA in M199 containing 10 % FBS (complete medium).Promastigotes (26106 cells ml21) treated with reference drugpentamidine (100 mg ml21) served as positive control; the equivalentnumber of promastigotes in the medium alone served as negativecontrol, while parasite with 0.2 % DMSO (representing the highestconcentration of diluent in the test samples) was taken as solventcontrol to ensure that the solvent did not have any effect on parasiteviability. An aliquot of the treated and untreated parasites was dilutedin 16 PBS (pH 7.2) and counted daily for 7 days in a haemocy-tometer using a phase-contrast microscope under a 640 objective(Afrin et al. 2001).
Leishmanicidal effect of EROSA in promastigote growth
reversibility assay. To confirm the leishmanicidal effect of EROSA,treated and untreated parasites after 7 days of incubation were washedtwice with fresh M199 and then resuspended in complete M199 andallowed to grow further at 22 uC. The viable parasites were enumeratedmicroscopically after 4 days (Afrin et al. 2001).
Determination of 50 % promastigote growth inhibitory concen-
tration (GI50) of EROSA. To determine the GI50 (the concentrationof drugs that inhibits 50 % growth of promastigotes), the parasites ata density of 26106 cells ml21 were incubated without or with EROSAat serial threefold dilutions in triplicates, starting from 100 mg ml21,for 4 days at 22 uC. Pentamidine was used as a reference drug. Themean percentage viability was calculated as: (mean cell number oftreated promastigotes ml21)6100/(mean cell number of untreatedpromastigotes ml21). Accordingly, GI50 was determined by graphicalextrapolation after plotting the mean percentage viability versus theconcentration of the drug.
Analysis of cellular morphology of EROSA-treated promasti-
gotes. Promastigotes (26106 cells ml21) were incubated in thepresence of EROSA in M199 containing 10 % FBS (completemedium) at a concentration of 100 mg ml21. Pentamidine (100 mgml21)-treated promastigotes served as positive control while parasiteswithout drug were taken as negative control. After 4 days oftreatment, an aliquot was placed in a haemocytometer and cellularmorphology such as elongated, slender, shrunken, oval or roundparasites, and loss of flagella were checked under a phase-contrastmicroscope with a 640 objective. At least 20 microscopic fields wereobserved for each sample. Data were recorded by using NIS-Elementsimaging software.
Antileishmanial activity of EROSA in an ex-vivo macrophage–
amastigote model. To evaluate the effects of EROSA on intracellularL. donovani amastigote forms, murine macrophages (RAW264.7,26106 cells ml21) were allowed to adhere to round glass coverslips in24-well tissue culture plates in RPMI 1640 medium supplementedwith penicillin G sodium (100 U ml21), streptomycin sulfate (100 mgml21) and 10 % FBS, and cultured at 37 uC in 5 % CO2. After 12 h,adherent macrophages were infected with stationary-phase promas-tigotes using a cell to parasite ratio of 1 : 10 and incubated at 37 uC fora further 12 h. The non-phagocytosed parasites were removed bygentle washing, and the infected macrophages were incubated for anadditional 48 h with EROSA or pentamidine at serial fourfolddilutions in triplicates, starting from 100 mg ml21. The coverslips
Apoptosis-like death in L. donovani promastigotes
http://jmm.sgmjournals.org 75

were fixed in methanol and Giemsa-stained for microscopic
evaluation. At least 200 macrophages per coverslip were counted
for calculation of mean percentage infection as (number of
amastigotes per 200 macrophages in treated group)6100/(number
of amastigotes per 200 macrophages in untreated group). The 50 %
inhibitory concentration for amastigotes (IC50) was determined by
graphical extrapolation (Dutta et al., 2008).
Visualization of phosphatidylserine (PS) exposure in EROSA-
treated promastigotes by annexin V-FITC/propidium iodide
(PI) binding. Externalization of PS in the outer membrane of
EROSA-treated promastigotes was measured by double staining for
annexin V and PI according to Mukherjee et al. (2009) with minor
modifications. Briefly, exponential-phase promastigotes (26106 cells
ml21) were incubated with EROSA at a concentration of 100 mg ml21
for 72 h. Pentamidine, which has been studied by others for the
apoptotic killing of the Leishmania parasites (Nguewa et al., 2005;
Singh & Dey 2007), served as the reference antileishmanial drug.
Parasites without any treatment were taken as control. Untreated and
treated parasites were centrifuged at 3000 g for 10 min and washed
twice in cold PBS, and the pellet was resuspended in annexin V-
FLUOS labelling solution (100 ml, containing both annexin V and PI),
as per the manufacturer’s instructions (Roche). After 15 min of
incubation in the dark at 26 uC, 400 ml of incubation buffer was
added and data were acquired using a BD LSR II flow cytometer and
analysed using Deva software.
Terminal deoxynucleotidyltransferase (TdT)-mediated dUTP
nick-end labelling (TUNEL) assay in EROSA-treated promasti-
gotes. DNA fragmentation was detected by catalytically incorporat-
ing FITC-dUTP at the 39-hydroxyl end of the fragmented DNA using
an in situ cell death detection kit (Roche) as per the manufacturer’s
instructions. In brief, exponential-phase promastigotes (26106 cells
ml21) were incubated with EROSA (100 mg ml21), with pentamidine
as positive control or with medium alone. After 72 h, the cells were
washed, fixed with 4 % paraformaldehyde on ice for 1 h, washed with
PBS and incubated with 3 % H2O2 in methanol for 10 min at 25 uC.
This was followed by washing with PBS and the cells were then
permeabilized with freshly prepared chilled 0.1 % Triton X-100 for
5 min on ice. The cells were washed twice with PBS, after which 50 ml
of reaction mixture containing TdT and FLUOS-labelled dUTP was
added for 1 h at 37 uC. The cells were washed and finally resuspended
in PBS for data acquisition in a BD LSR II flow cytometer and
analysed using Deva software. Histogram analysis of FL-1H (x-axis;
FITC fluorescence) was recorded to show the shift in fluorescence
intensity compared with the control (Dutta et al., 2007).
Study of dyskinetoplastidy in EROSA-treated parasites by
fluorescence microscopy. Exponential-phase promastigotes (26106
cells ml21) were treated with EROSA (100 mg ml21) for 72 h at
22 uC, washed twice with PBS, fixed with 80 % chilled ethanol and
kept at 4 uC for 24 h. The cells were then washed with PBS and the
pellet was resuspended in 500 ml DNase-free RNase (200 mg ml21)
for 1 h at 37 uC. The cells were stained with PI (50 mg ml21) and
incubated in the dark for 20 min at 4 uC. An aliquot of 20 ml was
taken from each sample, placed on a glass microscope slide
and observed directly (without fixation) under a high-resolution
fluorescence microscope. Images were processed (Singh et al., 2009)
using NIS-Elements imaging software. At least 20 microscopic fields
were observed for each sample.
Cell cycle analysis in EROSA-treated promastigotes. The
potency of EROSA for damaging DNA was detected through flow
cytometry by PI staining. Exponential-phase promastigotes (26106
cells ml21) were treated with EROSA or pentamidine (100 mg ml21)
for 72 h at 22 uC, washed twice with PBS, fixed with 80 % chilled
ethanol and kept at 4 uC for 24 h. Cells were then washed with PBS
and the pellet was resuspended in 500 ml DNase-free RNase A (200 mg
ml21) for 1 h at 37 uC. The cells were stained with PI (50 mg ml21)
and incubated in the dark for 20 min at 4 uC. Data acquisition was
carried out using a BD FACS Calibur flow cytometer and analysed
using CellQuest software (Dutta et al., 2007).
Determination of mitochondrial membrane potential (DYm).Mitochondrial damage was assessed using a cell-permeable dye, JC-1
(Misra et al., 2009). Briefly, exponential-phase promastigotes after
exposure to EROSA or pentamidine (100 mg ml21) for 72 h were
centrifuged and loaded with JC-1 (10 mM) in PBS at 37 uC for 10 min.
Cells were then analysed by a BD LSR II flow cytometer using DEVA
software. The ratio of the red to green fluorescence intensities denoted
the relative change in mitochondrial membrane potential (DYm).
Measurement of intracellular reactive oxygen species (ROS)
generation. To monitor the level of ROS generated, the cell-
permeant probe, H2DCFDA (Misra et al., 2009) was used with minor
modifications. In brief, promastigotes were treated with EROSA or
pentamidine at 22 uC for 72 h. The cells were washed twice with PBS
and finally resuspended in 500 ml PBS, loaded with cell-permeant
probe H2DCFDA (10 mM) and incubated for 30 min at 25 uC. The
cells were analysed for intracellular ROS by a BD LSR II flow
cytometer and DEVA software.
Cytotoxicity of EROSA to mammalian cells. A murine macro-
phage cell line (RAW264.7) was cultured in RPMI 1640 medium
supplemented with penicillin G sodium (100 U ml21), streptomycin
sulfate (100 mg ml21) and 10 % FBS. Macrophages at a density of
16106 cells ml21 were incubated with different concentrations of
EROSA (0–200 mg ml21) for 48 h in a CO2 incubator (5 % CO2,
37 uC). Pentamidine served as reference drug and 0.2 % DMSO was
used as solvent control. Macrophages without any treatment were
taken as control. The cells were observed under a phase-contrast
microscope and viability was ascertained by trypan blue dye exclusion
(Afrin et al., 2001; Dutta et al., 2008).
Statistical analysis. In vitro antileishmanial activity was expressed
as GI50 by linear regression analysis. Values are mean±SE of samples
in triplicate from at least three independent experiments. Statistical
analysis was performed using ANOVA followed by Dunnett’s test.
*P,0.05, **P,0.01 and ***P,0.001 compared with the control
group were considered significant.
RESULTS
GC and GC-MS analysis of S. aromaticum oil
GC and GC-MS analysis revealed eugenol (59.75 %) andeugenyl acetate (29.24 %) as the major chemical constitu-ents of S. aromaticum oil (Fig. 1), which together compose88.99 % of EROSA. The other 16 components weredetected in minor amounts (0.11–3.75 %), including b-caryophyllene (3.75 %), nonacosane (1.39 %) and caryo-phyllene oxide (1.05 %), as presented in Table 1.
EROSA inhibits growth of L. donovanipromastigotes
The effect of EROSA on the proliferation of exponentiallygrowing L. donovani promastigotes was studied by monitor-ing the kinetics of cell growth. A time-dependent reduction
M. Islamuddin, D. Sahal and F. Afrin
76 Journal of Medical Microbiology 63

175
150
125
100
75
50
25
200
10–4× Intensity
0
40302010
HO
H3CO
CH2
H3C
CH2O
O
OCH3
29
.02
5
11
.48
61
2.2
91
14
.21
9
16
.11
4
18
.34
51
9.0
56
20
.39
5
22
.54
6
24
.28
8
33
.05
8
34
.85
1
36
.14
2
39
.58
7
31
.94
5
47
.81
1
49
.79
5
55
.52
9
Eug
enol
Eug
enyl
ace
tate
50
Time (min)
Fig. 1. Chromatogram obtained by high-resolution GC-MS analysis of S. aromaticum oil. Insets show eugenol (59.75 %) andeugenyl acetate (29.24 %).
Table 1. Constituents of S. aromaticum oil
Peak Retention time (min) Relative area Area (%) Compounds
1 10.809 9 449 0.11 Cubebene
2 11.424 15 732 0.18 L-Camphor
3 13.664 335 619 3.75 b-Caryophyllene
4 15.588 43 140 0.48 a-Humulene
5 17.933 60 725 0.68 Cadina-1(10),4-diene
6 18.596 14 042 0.16 Copaene
7 19.935 23 972 0.27 L-Calamenene
8 23.768 94 101 1.05 Caryophyllene oxide
9 25.169 15 725 0.18 cis-Limonene oxide
10 28.305 5 345 203 59.75 Eugenol
11 31.045 2 615 654 29.24 Eugenyl acetate
12 32.112 34 488 0.39 Farnesene
13 33.561 36 521 0.41 Viridiflorol
14 33.744 38 605 0.43 Chavicol
15 34.911 42 577 0.48 29,39,4’-Trimethoxyacetophenone
16 38.563 50 510 0.56 2,4-Diacetylphloroglucinol
17 47.020 45 555 0.51 Henicosane
18 54.108 124 237 1.39 Nonacosane
Total 8 945 855 100.00
Apoptosis-like death in L. donovani promastigotes
http://jmm.sgmjournals.org 77
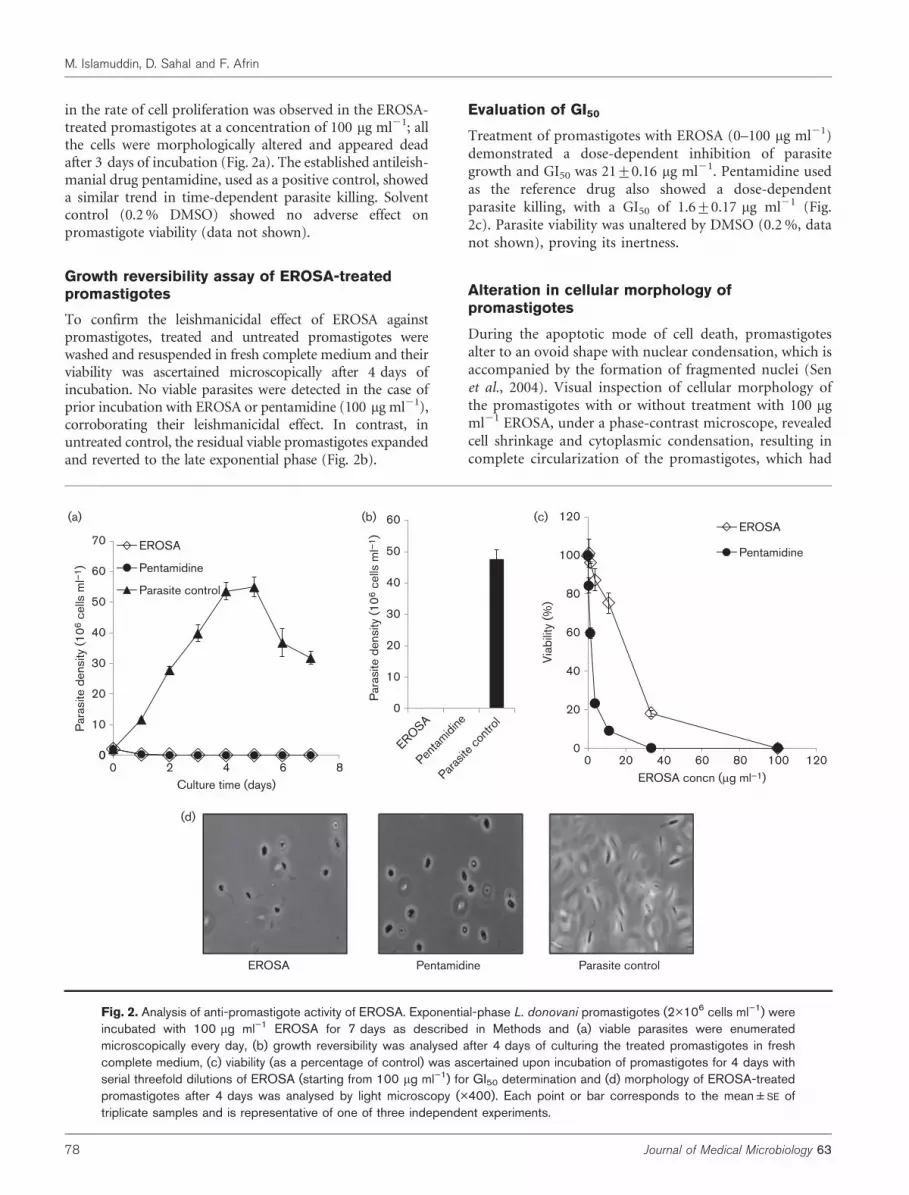
in the rate of cell proliferation was observed in the EROSA-treated promastigotes at a concentration of 100 mg ml21; allthe cells were morphologically altered and appeared deadafter 3 days of incubation (Fig. 2a). The established antileish-manial drug pentamidine, used as a positive control, showeda similar trend in time-dependent parasite killing. Solventcontrol (0.2 % DMSO) showed no adverse effect onpromastigote viability (data not shown).
Growth reversibility assay of EROSA-treatedpromastigotes
To confirm the leishmanicidal effect of EROSA againstpromastigotes, treated and untreated promastigotes werewashed and resuspended in fresh complete medium and theirviability was ascertained microscopically after 4 days ofincubation. No viable parasites were detected in the case ofprior incubation with EROSA or pentamidine (100 mg ml21),corroborating their leishmanicidal effect. In contrast, inuntreated control, the residual viable promastigotes expandedand reverted to the late exponential phase (Fig. 2b).
Evaluation of GI50
Treatment of promastigotes with EROSA (0–100 mg ml21)demonstrated a dose-dependent inhibition of parasitegrowth and GI50 was 21±0.16 mg ml21. Pentamidine usedas the reference drug also showed a dose-dependentparasite killing, with a GI50 of 1.6±0.17 mg ml21 (Fig.2c). Parasite viability was unaltered by DMSO (0.2 %, datanot shown), proving its inertness.
Alteration in cellular morphology ofpromastigotes
During the apoptotic mode of cell death, promastigotesalter to an ovoid shape with nuclear condensation, which isaccompanied by the formation of fragmented nuclei (Senet al., 2004). Visual inspection of cellular morphology ofthe promastigotes with or without treatment with 100 mgml21 EROSA, under a phase-contrast microscope, revealedcell shrinkage and cytoplasmic condensation, resulting incomplete circularization of the promastigotes, which had
EROSA
12060
60
70
(a)
(d)
(b) (c)
Pentamidine
Parasite control 80
100
EROSA
Pentamidine
40
50
30
40
50
40
60
10
20
30
0
10
20
Para
site d
ensity
(10
6 c
ells
ml–
1)
Para
site d
ensity
(10
6 c
ells
ml–
1)
0100806040200 120
20
Via
bili
ty (
%)
0
Penta
mid
ine
EROSA
Paras
ite c
ontro
l
06420 8
Culture time (days)
EROSA Pentamidine Parasite control
EROSA concn (μg ml–1)
Fig. 2. Analysis of anti-promastigote activity of EROSA. Exponential-phase L. donovani promastigotes (2�106 cells ml”1) wereincubated with 100 mg ml”1 EROSA for 7 days as described in Methods and (a) viable parasites were enumeratedmicroscopically every day, (b) growth reversibility was analysed after 4 days of culturing the treated promastigotes in freshcomplete medium, (c) viability (as a percentage of control) was ascertained upon incubation of promastigotes for 4 days withserial threefold dilutions of EROSA (starting from 100 mg ml”1) for GI50 determination and (d) morphology of EROSA-treatedpromastigotes after 4 days was analysed by light microscopy (�400). Each point or bar corresponds to the mean±SE oftriplicate samples and is representative of one of three independent experiments.
M. Islamuddin, D. Sahal and F. Afrin
78 Journal of Medical Microbiology 63

mostly become aflagellated, coupled with substantialreduction in size compared with that of untreated parasites(Fig. 2d).
Anti-amastigote activity of EROSA
In Leishmania infection, the promastigotes transform intoamastigotes within the phagolysosomal vacuoles of macro-phages. Accordingly, the antileishmanial activity of EROSAwas tested against intracellular amastigotes in L. donovani-infected RAW264.7 macrophages. Giemsa-stained cover-slips were analysed microscopically for the presence ofphagocytosed amastigotes within the macrophages. The IC50
of EROSA and pentamidine for L. donovani amastigotes wasfound to be 15.25±0.14 and 1.1±0.04 mg ml21, respectively(Fig. 3).
EROSA-treated promastigotes exhibit bothannexin V and PI binding
Translocation of phospholipids from the inner side to theouter layer of the plasma membrane is an attribute of cellsin the early phase of apoptosis. Annexin V, a Ca2+-dependent phospholipid-binding protein with a highaffinity for PS, is routinely used to show externalizationof PS. Since annexin V can also label necrotic cells, apropidium iodide (PI) stain was used to differentiatebetween the apoptotic cells (annexin V +ve and PI 2ve),necrotic cells (annexin V 2ve and PI +ve), late apoptoticcells (annexin V +ve and PI +ve) and live cells (annexinV –ve and PI –ve). Promastigotes treated with EROSA weredouble-stained with annexin V and PI, and analysed byflow cytometry. As shown in Fig. 4, a significant percentageof EROSA-treated promastigotes stained positive for bothannexin V and PI (55.60 %, upper right quadrant) ascompared with 79.13 % with pentamidine, after 72 h ofincubation at 100 mg ml21. In contrast, only 2.20 %
double-positivity was observed in untreated controls.This observation suggests that EROSA induced cell deathby loss of membrane integrity, as shown by the increased PIincorporation and annexin V binding, indicating lateapoptotic phase.
EROSA induces in situ DNA nicking in L. donovanipromastigotes
Fragmentation of nuclear DNA into oligonucleosomalunits is one of the biochemical hallmarks of eukaryoticapoptosis. DNA nicking resulting from exposure to EROSAwas detected using a TUNEL assay in which the proportionof DNA nicks was quantified by measuring the incorpora-tion of dUTP-FLUOS into the nicked end via TdT. Thus theproportion of DNA nicks was directly proportional to thefluorescence obtained from dUTP-FLUOS. Promastigotestreated with 100 mg ml21 EROSA for 72 h showed anincrease in nuclear DNA fragmentation as evidenced byenhanced dUTP-FLUOS incorporation. The mean fluores-cence intensity of untreated promastigotes and promasti-gotes incubated with EROSA and with pentamidine wasfound to be 522, 4694 and 3876, respectively (Fig. 5a).
EROSA-induced cytotoxicity causesdyskinetoplastidy in promastigotes
The kinetoplast is a specialized portion of a mitochondrioncomprising a network of circular DNA (kDNA), char-acteristic of flagellates of the order Kinetoplastida. ThekDNA has been shown to be susceptible to elimination(dyskinetoplastidy) by certain DNA-intercalating drugs, forexample, acriflavine and berenil. Having observed EROSA-induced genomic DNA damage in L. donovani promasti-gotes, we next investigated whether EROSA causes anydamage to kDNA. Cells were treated with EROSA (100 mgml21), permeabilized and stained with PI. In untreatedpromastigotes, the cells appeared with two discrete redspots representing intact kDNA of mitochondria (K) andnucleus (N) respectively. In EROSA- as well as pentami-dine-treated promastigotes (72 h), the spots representingthe mitochondria were abolished, indicating dyskineto-plastidy (Fig. 5b).
EROSA-induced apoptosis causes an increase inthe sub-G0–G1 population
Flow cytometric analysis after cell permeabilization andlabelling with PI is used to quantify the percentage ofpseudohypodiploid cells. In a given cell, the amount of PIcorrelates with the DNA content, and, accordingly, DNAfragmentation in apoptotic cells translates into a sub-G0–G1 peak. Promastigotes treated with EROSA (100 mg ml21)for 72 h caused a hypodiploid peak at sub-G0–G1, asanalysed by flow cytometry (Fig. 6). The proportion ofuntreated control parasites in the sub-G0–G1 region of themonoparametric plot of parasites with reduced DNAcontent was 1.20 %, which increased to 47.58 % and
0100806040200 120
20
40
60
80
100
120
Infe
ction (
%)
EROSA concn (μg ml–1)
EROSA
Pentamidine
Fig. 3. Effects of EROSA on L. donovani intracellular amastigoteforms. After infection with promastigotes, RAW264.7 macro-phages were incubated with serial fourfold dilutions of EROSA(starting at 100 mg ml”1) for 48 h and infection (as a percentage ofcontrol) was determined microscopically, as described in Methods.Error bars indicate ±SE.
Apoptosis-like death in L. donovani promastigotes
http://jmm.sgmjournals.org 79

2.20 % 1.68 %
1.68 %
103102
FL1-H
FL2
-H
101100
103
102
101
100
104
FL2
-H
103
102
101
100
104
FL2
-H
103
102
101
100
104
104103102
FL1-H
101100 104 103102
FL1-H
101100 104
90.44 %
Annexin V-FITC
PI
Parasite control EROSA Pentamidine
79.13 % 4.75 %
2.11 % 14.03 %
4.99 % 55.60 %
3.27 % 36.08 %
Fig. 4. Externalization of phosphatidylserine in EROSA-treated promastigotes. Parasites were incubated with EROSA orpentamidine (100 mg ml”1) for 72 h, co-stained with PI and annexin V-FITC and analysed by flow cytometry as described inMethods. The percentage of positive cells is indicated in the different quadrants. This is a representative profile of threeindependent experiments.
dUTP-FITC
100
50
150
(a)
(b)
0102101100 103
Count
A Parasite control
B Pentamidine
C EROSA
A C
B
(i)
(iii) N
N
N
N N
N
N
K
K
K
N
N N
(ii)
Fig. 5. Analysis of TUNEL positivity and dyskinetoplastidy in EROSA-treated promastigotes. Exponential-phase promastigoteswere incubated with EROSA or pentamidine (100 mg ml”1) for 72 h. Cells were fixed and (a) stained with dUTP-FITC in thepresence of TdT and analysed by flow cytometry or (b) probed with PI and images were taken at �200 magnification under aNikon fluorescence microscope as described in Methods. N, nucleus; K, mitochondrion.
M. Islamuddin, D. Sahal and F. Afrin
80 Journal of Medical Microbiology 63

18.26 % upon treatment with EROSA and pentamidine,respectively.
Induction of sustained depolarization ofmitochondrial membrane
Loss of mitochondrial membrane potential (DYm), acharacteristic feature of metazoan apoptosis, is also evidentin protozoans (Sen et al., 2007). JC-1, a lipophilicfluorescent cation, was used to measure the mitochondrialmembrane potential. This dye is known to freely permeatethe mitochondrial membrane and form J-aggregates whichfluoresce red; accordingly, the viable cells stained with JC-1exhibit a pronounced red fluorescence. An apoptoticstimulus induces a decrease in the DYm, resulting infailure of JC-1 to enter mitochondria, and JC-1 remains ascytosolic monomers that emit a green fluorescence.Therefore the ratio of J-aggregates to monomers serves asan effective indicator of the mitochondrial energy state of theparasites, allowing apoptotic cells to be easily distinguishedfrom their non-apoptotic counterparts. In untreated pro-mastigotes, the red/green fluorescence ratio was 3.063;treatment with 100 mg ml21 EROSA or pentamidine for72 h induced a significant decrease in DYm, resulting inpredominance of JC-1 monomers which fluoresce green,thereby translating into a decrease in the mean fluorescenceintensity ratio to 0.755 and 0.741, respectively (Fig. 7).
Pro-oxidant activity of promastigotes exhibited byEROSA
To evaluate the effect of EROSA on the oxidative status ofpromastigotes, H2DCFDA, a lipid-soluble membrane-permeable compound, was used based on the evidencethat, following cleavage by cellular non-specific esterases,impermeable H2DCF is formed which is subsequently
oxidized by intracellular ROS to produce a fluorescentcompound DCF; therefore, the resultant fluorescence(green) is directly proportional to the quantity of ROSgenerated. EROSA (100 mg ml21) treatment of promasti-gotes for 72 h led to a significant increase in ROS levels incomparison with control, as also observed with pentami-dine (Fig. 8).
Cytotoxicity of EROSA to mammalianmacrophages
Murine macrophage cell line RAW264.7 was used to checkthe adverse side effects of EROSA, using pentamidine asreference drug. The cytotoxicity assay revealed that therewas no adverse toxicity of EROSA to RAW264.7, even at200 mg ml21 (Fig. 9). Similar results were obtained withmouse peritoneal macrophages (data not shown).
DISCUSSION
Many traditional medicinal plants have been shown topossess antileishmanial activities validating their use in folkmedicine (Rocha et al., 2005). Plant essential oils and activecomponents can be used as alternatives or additions tocurrent antiparasitic therapies (Perez et al., 2012). Leish-manicidal activity of essential oils has been recentlydocumented (Sanchez-Suarez et al., 2013; Rodrigues et al.2013). Linalool-rich essential oil from leaves of C. cajucara(Rosa et al., 2003), eugenol-rich essential oil from O.gratissimum (Ueda-Nakamura et al., 2006) and sesquiter-penes-rich essential oil from E. uniflora (Rodrigues et al.,2013) have been demonstrated to possess cytotoxicityagainst Leishmania amazonensis parasites. In this study, wehave shown, we believe for the first time, that eugenol-richoil from S. aromaticum (EROSA) induced a profoundantileishmanial effect against L. donovani, with IC50 of
G0/G1: 1.20
160
120
80
40
Count
200
0
800600
FL2-A
4002000 1000
G1: 62.67 S: 3.85 G2/M: 17.57
Parasite control
EROSA Pentamidine
G0/G1: 18.26 G1: 54.40 S: 9.77 G2/M: 11.38
G0/G1: 47.58 %
G1: 30.40 %
S: 12.07 %
G2/M: 6.49 %
160
120
80
40
Count
200
0
160
120
80
40
Count
200
0
800600
FL2-A
4002000 1000 800600
FL2-A
4002000 1000
Fig. 6. Analysis of the cell cycle status of EROSA-treated promastigotes. Exponential-phase promastigotes were incubatedwith EROSA and pentamidine (100 mg ml”1) for 72 h, fixed in chilled methanol, probed with PI and analysed using an LSR IIflow cytometer for subsequent analysis using Cell Quest software as described in Methods.
Apoptosis-like death in L. donovani promastigotes
http://jmm.sgmjournals.org 81

21 mg ml21 and 15.24 mg ml21, respectively, againstpromastigotes and intracellular amastigotes. The effectwas leishmanicidal as also revealed in the promastigotegrowth reversibility assay and was more pronounced onintracellular amastigotes, suggesting that cellular eventsmight be important for the inhibitory effect of EROSA.
Having established the leishmanicidal effect on L. donovanipromastigotes and amastigotes, we sought to delineate themode of cell death caused by EROSA. In PI-impermeantlive cells, PS is predominantly on the annexin V-inaccessible inner leaflet of the cell membrane bilayer.Hence live cells are stained neither by FITC-annexin V
FITC-A green fluorescence intensity, FL–1
High Δψm Low Δψm
EROSA
Parasite control
Pentamidine0
104103102
100
50
150
100
75
50
25
125
0
100
75
50
25
125
0
P2
P2
P2
0105
Paras
ite c
ontro
l
EROSA
Penta
mid
ine
0.5
1
1.5
2
2.5
3
3.5
(a) (b)
MFI ra
tio (
PE
/FIT
C) Δψ
m
Count
Count
Count
Fig. 7. Effect of EROSA on mitochondrial membrane potential in L. donovani promastigotes. Exponential-phase promastigoteswere incubated with EROSA and pentamidine (100 mg ml”1) for 72 h and probed with JC-1 as described in Methods. (a) FL-1channel indicates green fluorescence intensity. (b) Bar diagram represents ratio of red/green fluorescence intensity. P2 lineindicates the interval gate for FL-1 positive cells.
FITC-A fluorescence
104103102
100
75
50
25
125
100
50
150
0
0
100
75
50
25
125
Count
Count
Count
0
105
Parasite control
EROSA
Pentamidine Fig. 8. EROSA induces the generation ofROS. Increased production of ROS wasobserved in promastigotes labelled withH2DCFDA (10 mM) after treatment withEROSA and pentamidine (100 mg ml”1) for72 h as described in Methods.
M. Islamuddin, D. Sahal and F. Afrin
82 Journal of Medical Microbiology 63

(green) nor by PI (red). Apoptosis in metazoan andunicellular cells results in externalization of PS from theinner leaflet to the outer leaflet of the bilayer, renderingapoptotic cells annexinV-positive (green). However theseannexinV-positive cells remain PI-negative since they areimpermeant to PI. The necrotic (dead) cells become PI-permeable even though they remain negative to annexin V.However, in late apoptotic/secondary apoptotic cells(apoptosis followed by necrosis), cells become positivefor both annexin V and PI (Dutta et al., 2007; Saha et al.,2009). In our studies, EROSA-treated L. donovani promas-tigotes became positive for both annexin V and PI. Hence,EROSA-induced cell death may be due to loss ofmembrane integrity, as evidenced by the increase in PIincorporation and annexin V binding, which is indicativeof late apoptosis. Apoptosis-like changes that shared severalphenotypic features with metazoan apoptosis (Debrabantet al., 2003) have been reported to mediate the leishma-nicidal action of camptothecin (Sen et al., 2004) andracemoside A (Dutta et al., 2007). Further, in situ TUNELof nicked DNA was observed, thus strongly substantiatingapoptosis in L. donovani promastigotes. This has also beenevidenced with racemoside A (Dutta et al., 2007). Apopto-tic cells generally feature active endonucleases thatpreferentially cleave DNA, translating into an increasedcell population located proximal to the G0–G1 peak on theleft side on a DNA frequency histogram. In promastigotestreated with other antileishmanial drugs such as curcumin,artemisinin and berberine chloride, a substantial propor-tion of cells have been identified in the sub-G0–G1 region(Das et al., 2008; Sen et al., 2007; Saha et al., 2009; Duttaet al., 2007; Li et al., 2009; Mukherjee et al., 2006). Ourstudy is in agreement with the previous reports and
conclusively shows that EROSA-treated L. donovani promas-tigotes exhibit a hypodiploid peak at sub-G0–G1, containingparasites with reduced DNA content, again substantiatingDNA fragmentation and induction of apoptosis.
In eukaryotes, mitochondria produce superoxide anionsand hydrogen peroxide, which, by changes in electron flowthrough the electron transport chain, generate reactiveintermediates like hydroxyl and other free radicals whichsubsequently oxidize and damage lipids, proteins and DNA(Jimenez Del Rıo & Velez-Pardo, 2004; Azmi et al., 2006).Damage to mitochondrial DNA causes inhibition of theexpression of electron transport proteins, leading to theaccumulation of ROS (Van Houten et al., 2006). Sustainedhypopolarization of mitochondrial membrane and genera-tion of ROS have also been established with promastigotestreated with the essential oil of Piper betle (Misra et al.,2009). We demonstrated that EROSA-induced cell death inL. donovani promastigotes induced loss of DYm and ROSgeneration, corroborating that the effect was mediatedthrough programmed cell death. The induction ofmitochondrial dysfunction in the apoptotic process wasapparent in the PI staining results. In the present study, wefound that EROSA-induced cell death was accompanied bydyskinetoplastidy, i.e. loss of kDNA network, as has alsobeen affirmed by others (Singh et al., 2009).
Essential oils have been reported to provoke depolarizationof the mitochondrial membranes by decreasing themembrane potential, influencing Ca2+ cycling (Richter &Schlegel, 1993) and other ionic channels and therebyreducing the pH gradient, and affecting (as in bacteria) theproton pump and the ATP pool. The essential oils changethe fluidity of the membranes, which become abnormallypermeable, resulting in leakage of free radicals, cytochromec, calcium ions and proteins, as in the case of oxidativestress and bioenergetic failure. Permeabilization of outerand inner mitochondrial membranes can lead to cell deathby apoptosis or necrosis (Yoon et al., 2000). Pro-oxidantaction of eugenol has also been reported to causecytotoxicity (Fujisawa et al., 2002). This prompted us toevaluate whether EROSA could induce oxidative stress in L.donovani promastigotes. It is well documented that ROSgeneration in cells following drug treatment can direct thecells towards apoptosis (Chipuk & Green, 2005). Theincrease in ROS after exposure to EROSA followed bydeath of the parasites suggested that EROSA-mediatedgeneration of ROS by the promastigotes was responsiblefor their apoptotic death. Earlier studies have establishedthat mitochondria are a possible target of ROS-inducedapoptosis in promastigotes which occurs via a decrease inDYm (Sudhandiran & Shaha, 2003). Further studies wereconducted to understand the changes occurring inpolarization of mitochondrial membrane after oxidativestress that were responsible for the apoptotic cell death. Inour study, the observation of a sharp lowering of DYm
evidently indicates that mitochondrial dysfunction occurs,giving evidence of apoptosis in promastigotes upontreatment with EROSA. In line with this, the following
80
60
40
20
100
0150100
EROSA
Via
bili
ty (
%)
Pentamidine
Concentration (µg ml–1)
50 200
Fig. 9. Adverse toxicity of EROSA to mammalian macrophages.Macrophages from the peritoneal cavity of mice were incubated for72 h at 37 6C in a CO2 incubator with increasing concentrationsof EROSA or pentamidine (0–200 mg ml”1) and viability wasascertained. Each bar represents mean±SE of triplicate samplesand data are representative of one of three independentexperiments.
Apoptosis-like death in L. donovani promastigotes
http://jmm.sgmjournals.org 83

reaction mechanism for EROSA can be envisaged: theessential oils of S. aromaticum, by penetrating thecytoplasmic membrane, disrupt and permeabilize it andespecially damage the mitochondrial membranes.
GC and MS analysis of EROSA revealed significantly highercontent of eugenol and eugenyl acetate, which togethercompose 88.99 % of S. aromaticum oil. Misra et al. (2009)attributed the pro-apoptotic effect of landrace BanglaMahoba of Piper betle on L. donovani to the high content ofeugenol (73.01 %), which also exhibited potent antileish-manial activity, whereas no activity was observed with Piperbetle landrace Kapoori Vellaikodi (27.15 % eugenol). So thefindings of Misra et al. also suggest that the antileishmanialactivity of EROSA observed by us may be due predomi-nantly to a higher content of eugenol.
Effectiveness of eugenol in stimulating apoptosis is evidentin human melanoma cells (Kim et al., 2006). Moreover, itis known that eugenol at lower concentrations acts as ananti-oxidant and anti-inflammatory agent, whereas athigher concentrations it can work as a pro-oxidant,resulting in enhanced production of tissue-damaging freeradicals (Suzuki et al., 1985). In the present study,apoptosis induced by EROSA was evidenced to bemediated via ROS generation. This finding, along withthe fact that eugenol at higher concentrations enhances thegeneration of ROS and induces apoptosis, validates theantileishmanial efficacy of EROSA, which is likely attrib-utable to the higher eugenol content. The leishmanicidalactivity of the eugenol-rich essential oil derived from O.gratissimum further substantiates our observation (Ueda-Nakamura et al., 2006).
Taken together with the combined use of differentapoptotic markers such as PS externalization, in situDNA nicking, dyskinetoplastidy, cell cycle arrest at sub-G0–G1 phase, reduction of DYm and ROS generation, we haveconclusively established that EROSA induced apoptosis-like programmed cell death in L. donovani promastigoteswith no adverse effect on mammalian cells. Our findingsvalidate eugenol-rich essential oil as a source of a new leadcompound for novel antileishmanial drugs.
ACKNOWLEDGEMENTS
We are thankful to Dr Ajai Kumar, Senior Technical Assistant, AIRF,JNU (New Delhi) for GC and GC-MS analysis. We are also thankfulto Fateh Mohammad, Laboratory Technician, Parasite ImmunologyLaboratory, Department of Biotechnology, Jamia Hamdard, NewDelhi, for his assistance throughout in this work. This work wassupported by research grants to F. A. from the Department ofBiotechnology and Department of Science and TechnologyGovernment of India, New Delhi, India. M. I. is the recipient ofSRF from the Department of Biotechnology, New Delhi, India.
REFERENCES
Afrin, F., Dey, T., Anam, K. & Ali, N. (2001). Leishmanicidal activity ofstearylamine-bearing liposomes in vitro. J Parasitol 87, 188–193.
Ashford, R. W., Desjeux, P. & Deraadt, P. (1992). Estimation of
population at risk of infection and number of cases of leishmaniasis.Parasitol Today 8, 104–105.
Azmi, A. S., Bhat, S. H., Hanif, S. & Hadi, S. M. (2006). Plantpolyphenols mobilize endogenous copper in human peripheral
lymphocytes leading to oxidative DNA breakage: a putative
mechanism for anticancer properties. FEBS Lett 580, 533–538.
Bagavan, A., Rahuman, A. A., Kaushik, N. K. & Sahal, D. (2011). In
vitro antimalarial activity of medicinal plant extracts against
Plasmodium falciparum. Parasitol Res 108, 15–22.
Burt, S. A. & Reinders, R. D. (2003). Antibacterial activity of selected
plant essential oils against Escherichia coli O157 : H7. Lett ApplMicrobiol 36, 162–167.
Chaieb, K., Hajlaoui, H., Zmantar, T., Kahla-Nakbi, A. B., Rouabhia,M., Mahdouani, K. & Bakhrouf, A. (2007). The chemical compositionand biological activity of clove essential oil, Eugenia caryophyllata
(Syzigium aromaticum L. Myrtaceae): a short review. Phytother Res 21,
501–506.
Chipuk, J. E. & Green, D. R. (2005). Do inducers of apoptosis trigger
caspase-independent cell death? Nat Rev Mol Cell Biol 6, 268–
275.
Croft, S. L. & Coombs, G. H. (2003). Leishmaniasis – current
chemotherapy and recent advances in the search for novel drugs.Trends Parasitol 19, 502–508.
Das, R., Roy, A., Dutta, N. & Majumder, H. K. (2008). Reactive oxygen
species and imbalance of calcium homeostasis contributes tocurcumin induced programmed cell death in Leishmania donovani.
Apoptosis 13, 867–882.
Debrabant, A., Lee, N., Bertholet, S., Duncan, R. & Nakhasi, H. L.(2003). Programmed cell death in trypanosomatids and other
unicellular organisms. Int J Parasitol 33, 257–267.
Dutta, A., Ghoshal, A., Mandal, D., Mondal, N. B., Banerjee, S., Sahu,N. P. & Mandal, C. (2007). Racemoside A, an anti-leishmanial, water-
soluble, natural steroidal saponin, induces programmed cell death inLeishmania donovani. J Med Microbiol 56, 1196–1204.
Dutta, A., Sarkar, D., Gurib-Fakim, A., Mandal, C. & Chatterjee, M.(2008). In vitro and in vivo activity of Aloe vera leaf exudate in
experimental visceral leishmaniasis. Parasitol Res 102, 1235–1242.
Fujisawa, S., Atsumi, T., Kadoma, Y. & Sakagami, H. (2002).Antioxidant and prooxidant action of eugenol-related compounds
and their cytotoxicity. Toxicology 177, 39–54.
Fyfe, L., Armstrong, F. & Stewart, J. (1998). Inhibition of Listeria
monocytogenes and Salmonella enteriditis by combinations of plant
oils and derivatives of benzoic acid: the development of synergisticantimicrobial combinations. Int J Antimicrob Agents 9, 195–
199.
Gottlieb, O. R. & Guimaraes, M. T. (1960). Modified distillation trap.Chemist Analyst 49, 114–116.
Iwu, M. M., Jackson, J. E. & Schuster, B. G. (1994). Medicinal plantsin the fight against leishmaniasis. Parasitol Today 10, 65–68.
Jimenez Del Rıo, M. & Velez-Pardo, C. (2004). Transition metal-
induced apoptosis in lymphocytes via hydroxyl radical generation,mitochondria dysfunction, and caspase-3 activation: an in vitro model
for neurodegeneration. Arch Med Res 35, 185–193.
Kim, G. C., Choi, D. S., Lim, J. S., Jeong, H. C., Kim, I, . R., Lee, M. H. &Park, B. S. (2006). Caspases-dependent apoptosis in human
melanoma cell by eugenol. Korean J Anat 39, 245–253.
Li, Q., Zhou, Y., Yao, C., Ma, X., Wang, L., Xu, W., Wang, Z. & Qiao, Z.(2009). Apoptosis caused by Hsp90 inhibitor geldanamycin in
Leishmania donovani during promastigote-to-amastigote transforma-tion stage. Parasitol Res 105, 1539–1548.
M. Islamuddin, D. Sahal and F. Afrin
84 Journal of Medical Microbiology 63

Mishra, R. K. & Singh, S. K. (2008). Safety assessment of Syzygiumaromaticum flower bud (clove) extract with respect to testicularfunction in mice. Food Chem Toxicol 46, 3333–3338.
Misra, P., Kumar, A., Gupta, S., Khare, P., Kumar, N. & Dube, A.(2009). Pro-apoptotic effect of landrace Bangla Mahoma of Piperbetle- on Leishmania donovani may be due to high content of eugenol.J Med Microbiol 58, 1058–1066.
Mohamed, A. H. H., El-Sayed, M. A., Hegazy, M. E., Helaly, S. E.,Esmail, A. M. & Mohamed, N. S. (2010). Chemical constituents andbiological activities of Artemisia herba-alba. Rec Nat Products 4, 1–25.
Mukherjee, P., Sen, P. C. & Ghose, A. C. (2006). Lymph node cellsfrom BALB/c mice with chronic visceral leishmaniasis exhibitingcellular anergy and apoptosis: involvement of Ser/Thr phosphatase.Apoptosis 11, 2013–2029.
Mukherjee, P., Majee, S. B., Ghosh, S. & Hazra, B. (2009). Apoptosis-like death in Leishmania donovani promastigotes induced bydiospyrin and its ethanolamine derivative. Int J Antimicrob Agents34, 596–601.
Nguewa, P. A., Fuertes, M. A., Cepeda, V., Iborra, S., Carrion, J.,Valladares, B., Alonso, C. & Perez, J. M. (2005). Pentamidine is anantiparasitic and apoptotic drug that selectively modifies ubiquitin.Chem Biodivers 2, 1387–1400.
Perez, S. G., Ramos-Lopez, M. A., Sanchez-miranda, E., Fresan-orozco, M. C. & Perez-Ramos, J. (2012). Antiprotozoa activity ofsome essential oils. J Medicinal Plants Res 6, 2901–2908.
Richter, C. & Schlegel, J. (1993). Mitochondrial calcium releaseinduced by prooxidants. Toxicol Lett 67, 119–127.
Rocha, L. G., Almeida, J. R., Macedo, R. O. & Barbosa-Filho, J. M.(2005). A review of natural products with antileishmanial activity.Phytomedicine 12, 514–535.
Rodrigues, K. A. F., Amorim, L. V., de Oliveira, J. M., Dias, C. N.,Moraes, D. F. C., Andrade, E. H. A., Maia, J. G. S., Carneiro, S. M. P. &Carvalho, F. A. A. (2013). Eugenia uniflora L. essential oil as apotential anti-Leishmania agent: effects on Leishmania amazonensisand possible mechanisms of action. Evid Based Complement AlternatMed 2013, 279726.
Rosa, M. S. S., Mendonca-Filho, R. R., Bizzo, H. R., Rodrigues, I. A,Soares, R. M. A., Souto-Padron, T., Alviano, C. S. & Lopes, A. H. C. S.(2003). Antileishmanial activity of a linalool-rich essential oil fromCroton cajucara. Antimicrob Agents Chemother 47, 1895–1901.
Saha, P., Sen, R., Hariharan, C., Kumar, D., Das, P. & Chatterjee, M.(2009). Berberine chloride causes a caspase-independent, apoptotic-like death in Leishmania donovani promastigotes. Free Radic Res 43,1101–1110.
Sanchez-Suarez, J. F., Riveros, I. & Delgado, G. (2013). Evaluationof the leishmanicidal and cytotoxic potential of essential oils derivedfrom ten Colombian plants. Iran J Parasitol 8, 129–136.
Santoro, G. F., Cardoso, M. G., Guimaraes, L. G., Mendonca, L. Z. &Soares, M. J. (2007). Trypanosoma cruzi: activity of essential oils fromAchillea millefolium L., Syzygium aromaticum L. and Ocimumbasilicum L. on epimastigotes and trypomastigotes. Exp Parasitol116, 283–290.
Sen, N., Das, B. B., Ganguly, A., Mukherjee, T., Tripathi, G.,Bandyopadhyay, S., Rakshit, S., Sen, T. & Majumder, H. K. (2004).Camptothecin induced mitochondrial dysfunction leading to pro-grammed cell death in unicellular hemoflagellate Leishmaniadonovani. Cell Death Differ 11, 924–936.
Sen, R., Bandyopadhyay, S., Dutta, A., Mandal, G., Ganguly, S., Saha,P. & Chatterjee, M. (2007). Artemisinin triggers induction of cell-cycle arrest and apoptosis in Leishmania donovani promastigotes.J Med Microbiol 56, 1213–1218.
Singh, G. & Dey, C. S. (2007). Induction of apoptosis-like cell deathby pentamidine and doxorubicin through differential inhibition oftopoisomerase II in arsenite-resistant L. donovani. Acta Trop 103,172–185.
Singh, N., Kaur, J., Kumar, P., Gupta, S., Singh, N., Ghosal, A., Dutta,A., Kumar, A., Tripathi, R. & other authors (2009). An orally effectivedihydropyrimidone (DHPM) analogue induces apoptosis-like celldeath in clinical isolates of Leishmania donovani overexpressingpteridine reductase 1. Parasitol Res 105, 1317–1325.
Sudhandiran, G. & Shaha, C. (2003). Antimonial-induced increase inintracellular Ca2+ through non-selective cation channels in the hostand the parasite is responsible for apoptosis of intracellularLeishmania donovani amastigotes. J Biol Chem 278, 25120–25132.
Suzuki, Y., Sugiyama, K. & Furuta, H. (1985). Eugenol-mediatedsuperoxide generation and cytotoxicity in guinea pig neutrophils. JpnJ Pharmacol 39, 381–386.
Ueda-Nakamura, T. R., Mendonca-Filho, R. R., Morgado-Dıaz, J. A.,Korehisa Maza, P., Prado Dias Filho, B., Aparıcio Garcia Cortez, D.,Alviano, D. S., Rosa, M. S., Lopes, A. H. & other authors (2006).Antileishmanial activity of eugenol-rich essential oil from Ocimumgratissimum. Parasitol Int 55, 99–105.
Van Houten, B., Woshner, V. & Santos, J. H. (2006). Role ofmitochondrial DNA in toxic responses to oxidative stress. DNARepair (Amst) 5, 145–152.
Yoon, H. S., Moon, S. C., Kim, N. D., Park, B. S., Jeong, M. H. & Yoo,Y. H. (2000). Genistein induces apoptosis of RPE-J cells by openingmitochondrial PTP. Biochem Biophys Res Commun 276, 151–156.
Apoptosis-like death in L. donovani promastigotes
http://jmm.sgmjournals.org 85





























