Embed Size (px)
Citation preview

Contents lists available at ScienceDirect
Aquaculture
journal homepage: www.elsevier.com/locate/aquaculture
Physiological and histopathological impacts of increased carbon dioxide andtemperature on the scallops Argopecten purpuratus cultured under upwellinginfluences in northern Chile
Marco A. Lardiesa,b,⁎, Samanta Beniteza,b, Sebastian Osoresa,b, Cristian A. Vargasb,c,Cristián Duarted, Karin B. Lohrmanne, Nelson A. Lagosf
a Facultad de Artes Liberales, Universidad Adolfo Ibañez, Santiago, Chileb Center for the Study of Multiple-Drivers on Marine Socio-Ecological Systems (MUSELS), Universidad de Concepción, Concepción, Chilec Aquatic Ecosystem Functioning Lab (LAFE), Department of Aquatic Systems, Faculty of Environmental Sciences, Environmental Sciences Center EULA Chile, Universidadde Concepción, Chiled Facultad de Ecología y Recursos Naturales, Universidad Andrés Bello, Santiago, Chilee Facultad de Ciencias del Mar, Universidad Católica del Norte, Centro Innovación Acuícola Aquapacífico, Coquimbo, Chilef Centro de Investigación e Innovación para el Cambio Climático (CiiCC), Universidad Santo Tomás, Ejercito 146, Santiago, Chile
A R T I C L E I N F O
Keywords:Ocean acidificationHistopathologyMetabolismFood ingestionMultiple stressor impactsAquaculture
A B S T R A C T
In addition to the increase in temperature occurring in the world's oceans, new evidences suggest a tendency foran increase in upwelling-favorable winds, bringing to surface cold and high pCO2 corrosive waters. Changingtemperature and pCO2 conditions may have significant implications for the shellfish farming socio-ecologicalsystem. In order, to setup the basis for understand the impact of both environmental variables, in this study, weinvestigated the combined effects of changing temperature and pCO2 on the physiological rates and histo-pathology of scallops Argopecten purpuratus farmed in Tongoy Bay, an area permanently influenced by coastalupwelling. Juvenile scallops were reared at two pCO2 levels (400 and 1000 μatm) and two temperatures (14 and18 °C). After 18 d of experimental exposure, growth, metabolic and clearance rates increased significantly athigh temperature but independent of pCO2 level, indicating a positive effect of warming on the physiologicalprocesses associated with energy acquisition. However, ingestion rates of scallops showed a synergistic inter-active effect when exposed to both stressors. Increased pCO2 also impacts the health of A. purpuratus throughatrophy in the digestive gland. These results suggest that, the presence of trade-offs in energy allocation duringupwelling-induced stress (low temperature and high pCO2) can impact growth, metabolism, ingestion rates andhealth status of scallops cultured in Tongoy Bay. But, less severe response to high pCO2 levels, suggest thatnatural variability in upwelling areas may promote acclimation and adaptation potential in this farmed scallops.
1. Introduction
Coastal oceans are arguably one of the most important ecosystemsof the planet and impacted by human activities in the ocean. Globalenvironmental stressors, such as Ocean Warming (OW), OceanAcidification (OA) and Deoxygenation (DO) are undergoing pro-nounced change in the world's coastal oceans today. There is growingconcern with respect to the combined effects (thereby synergistically,additively or antagonistically) of these environmental stressors on thecoastal marine biota. During the last decades, there is growing interestabout the need to understand the interplay between environmental andsocio-economic drivers in marine resource management (Cooley and
Doney 2009; Perry et al. 2010). Essentially, humans' adaptation capa-cities based on a better understanding of the combined impacts ofmultiple global and local drivers on marine ecosystems services as foodprovision through fisheries and aquaculture activities will facilitate thesuccess of our societies in establishing sustainable pathways for thefuture (Gattuso et al., 2015).
In Chile, coastal shellfish aquaculture activities are focused on bi-valves (e.g. oysters, mussels, scallops), and gastropods (e.g. snails).From 1990, shellfish aquaculture has experienced an annual averagegrowth rate of ~18%. Shellfish farming accounted with ca.265,000 tons in 2013, or 14% of total biomass and Chile is the third andsecond largest producer of scallops and mussels in the world,
http://dx.doi.org/10.1016/j.aquaculture.2017.06.008Received 29 December 2016; Received in revised form 6 June 2017; Accepted 8 June 2017
⁎ Corresponding author.E-mail address: [email protected] (M.A. Lardies).
Aquaculture 479 (2017) 455–466
Available online 10 June 20170044-8486/ © 2017 Elsevier B.V. All rights reserved.
MARK
respectively (www.sernapesca.cl). Although the industry has thrived inthe past decade, global drivers such as climate, market variability, andanthropogenic activities provide major challenges in sustaining thefuture of the aquaculture sector in our coastal ocean. This is of parti-cular concern in the case of valuable commercially exploited speciessuch as the scallop Argopecten purpuratus. A recent crash in the scallopproduction has been recorded in Chile, where the number of scallopfarms became closed due to market and environmental drivers, withconcomitant reduction in national production from 20.000 tons in year2000 to 5000 tons in year 2013 (Lagos et al. 2016).
Eastern Boundary Upwelling Ecosystems, located off the mid-lati-tude western coasts of the U.S., South America, Iberia and Africa, arepredicted to be one of the most increasingly stressed marine ecosystemsunder multiple-stressors (e.g. ocean acidification, warming and deox-ygenation; Gruber, 2011). Upwelling ecosystems account for only 1.9%of the world's oceans, but provide an estimated 23% of global fisheries(Kudela et al. 2005; Chavez et al. 2008), being the Humboldt CurrentSystem the most biologically productive ecosystem of world' coastaloceans (Thiel et al. 2007). These areas are characterized by strongseasonal winds that promote the upwelling of cold and CO2 super-saturated subsurface waters. Climatic projections suggest an increase inthe intensification of upwelling events due to the change in the in-tensity and patterns of wind that favor the upwelling (Sydeman et al.2014). In northern Chile, upwelling dynamics occur year around, withfoci located at major point or headlands of the coastline. In particular, astrong latitudinal gradient in SST anomalies located at 30°S is asso-ciated with the coastal area of Punta Lengua de Vaca (PLV) (Figueroaand Moffat 2000). This coastal area at the PLV upwelling system mightresult in under-saturated subsurface waters with respect to aragonite(Ωarag < 1) and in lower pH in nearshore and bays areas (Torres et al.2011), characteristics previously expected to occur in the open oceanuntil the year 2100 in a scenario of ocean acidification (Orr et al. 2005;Gruber et al. 2012). Coastal waters with low levels of carbonate sa-turation state are corrosives, challenging calcification of shells andskeletons of marine organisms (Fabry et al. 2008; Beniash et al. 2010;Duarte et al. 2015). Furthermore, large upwelling areas are located
northward of 37°S [in northern (18–30°S) and central Chile (30°–37°S)]showing SST anomalies that varies between 4 and 7 °C, whereas in la-titudes above 37°S, SST varies between 2 °C to 4 °C (Lagos et al. 2008;Aravena et al. 2014). The Chilean scallop farming industry in northernChile is localized near to these upwelling centres and has been regularlyaffected by events of larval and adult mortality, potentially driven bythe effect of corrosive upwelling waters. The environmentally inducedphenotypic change, namely, phenotypic plasticity has been recognizedas an important strategy for maximizing or maintaining fitness in anorganism that inhabits variable environments (Bouvet et al. 2005;Pigliucci 2005; Schlichting and Pigliucci 1998; Via and Lande 1985).These notions, suggest that may be expected that populations of A.purpuratus that inhabit waters influenced by coastal upwelling, namely,subject to environmental heterogeneity in pH, temperature and oxygenconcentration may be “naturally” acclimated to this environmentalvariation in temperature and pCO2 (i.e., phenotypic plasticity) probablyas end product of local adaptation (see Lardies et al. 2014).
In this study we evaluate the combined effects of temperature andpCO2 on scallops Argopecten purpuratus cultured under the influence ofcoastal upwelling. A. purpuratus represents an important species foraquaculture, since it is the most cultivated mollusk species in the northof Chile (over 5000 tons in 2013) with its main production zone is lo-calized in Tongoy Bay (von Brandt et al. 2006; SERNAPESCA 2013). A.purpuratus is a relatively well studied species regarding the effect ofenvironmental variables on physiological processes (see Navarro andJaramillo, 1994, Navarro and González, 1998; Labarta et al. 1997;Uriarte et al. 2004; Martínez et al. 1995, 2000; Fernández-Reiriz et al.,2005; Soria et al., 2007, 2011; Ramajo et al. 2016a) and the impacts ofincreased pCO2 and temperature on shell properties of this species(Lagos et al. 2016). However, no studies have addressed how combinedeffects of these climate stressors will affect the physiological rates andhistopathology of this scallop species. Thus, in this study we evaluatetrough laboratory experiments the effect of cold temperatures and in-creased pCO2 conditions, which commonly occur during upwellingevents influencing the study area (i.e. Tongoy Bay) upon growth ratesand metabolic and feeding performance of A. purpuratus.
30ºS
71.4ºW
Tongoy Bay
Coquimbo
30ºS
72ºW
10 km
Fig. 1. Location of the study site at the Northern Chile coast (a);Circle indicates the location of the scallop farm inside Tongoy Bay,which is northward of the Punta Lengua de Vaca upwelling center(b).
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
456

2. Materials and methods
2.1. Study site and animal collection
Juvenile individuals of A. purpuratus (< 40 mm in size (LT)) werecollected from culture ropes (2 m of depth) in Tongoy Bay (30°12′S,71°34′O) located in northern Chile (Fig. 1). Thus, the juvenile scallopsexperienced during its early life stages the influence of Punta Lengua deVaca upwelling center which waters penetrate into the Tongoy Bayseasonally (i.e. spring and summer) when winds coming from the south,transport Equatorial subsurface waters (ESSW) with high salinity, highnutrients content, low temperature, low pH, and low oxygen content tothe same scallop farming area (Rutllant 1993; Uribe and Blanco 2001;Lagos et al. 2016).
These experimental scallops are produced in hatchery from brood-stock of the same Tongoy Bay. After collection, the scallops weretransported in chilled conditions to the Calfuco Coastal Laboratory ofthe Universidad Austral de Chile (LCC, ca. 39°S, Valdivia) and placed inplastic containers (30 cm in diameter and 40 cm in height). To accli-matize them to experimental conditions, the scallops were kept for5 days and fed daily with microalgae Tetraselmis spp. (~65 × 106 cell/ml) before the beginning of the experiments. In addition, during the dayof the scallop collection, 17 juvenile individuals (hereafter naturalcontrol) were immediately preserved for histological procedures (seebelow).
2.2. Seawater acidification system
After the acclimation period, the scallops were randomly assigned toone of four treatments: (Angilletta et al., 2004) 14 °C and 400 μatmpCO2 (ambient condition), (Angilletta, 2009) 14 °C and 1000 μatmpCO2 (upwelling condition), (Aravena et al., 2014) 18 °C and 400 μatmpCO2 (warming and upwelling relaxation), (Beniash et al., 2010) 18 °Cand 1000 μatm pCO2 (ocean acidification and warming) (Table 1), for aperiod 18 days which is larger time than two complete upwelling cyclesin northern Chile (see Strub et al. 1998; Garreaud et al. 2011). Eachtreatment was replicated five times and each replicate contained fourscallops (N = 80). The experimental animals were identified using beetags. We used a semi-automatic system for long-term seawater carbo-nate chemistry manipulation (Torres et al. 2013; Duarte et al. 2014;Navarro et al. 2013; Manríquez et al., 2013; Vargas et al. 2013; Duarteet al. 2015). The experimental temperatures represent the averagesurface value (14 °C) and alternatively the maximum surface value(18 °C) of the study area (Uribe and Blanco 2001; Pérez et al. 2012;Aravena et al. 2014). The pCO2 levels were defined considering themean range reported for this coastal embayment by Torres & Ampuero(2009) and Vargas et al. (2017). The temperatures were adjusted andmaintained using external chillers. To obtain the different CO2 con-centrations, the experimental containers were adjusted following themethodology described by Navarro et al. (2013). Briefly, for 400 ppm,pure atmospheric air was bubbled into experimental containers; for
1000 ppm, we blended dry air with pure CO2 to each target con-centration using mass flow controllers (MFCs, www.aalborg.com) forair and CO2; then this blend was bubbled into the corresponding con-tainer. Dry and clean air was generated by compressing atmospheric air(117 psi) using an oil-free air compressor. The experimental containerswere constantly bubbled with the corresponding CO2 concentration.During the experiment, total alkalinity (TA) of the water was monitoredevery 4 days (two replicates) and pH, temperature, salinity of the waterwas monitored every day in each aquaria (i.e. replicate) and eachheader tank (Navarro et al. 2013). Seawater was changed every day,with the corresponding pCO2 levels from the seawater acidificationhead-tank following the methodology proposed by Navarro et al.(2013). pHNBS and total alkalinity (AT) was monitored on day 2, 6, 10,14 and 18 for carbonate systems estimates. pH samples were collectedin 50 mL syringes and immediately transferred to a 25 mL thermo-statted cell at 25.0 ± 0.1 °C for standardization, with a pH meterMetrohm® using a glass combined double junction Ag/AgCl electrode(Metrohm model 6.0258.600) calibrated with standard calibrationbuffer Metrohm® 4 (6.2307.200), 7 (6.2307.210) y 9 (6.2307.220). pHvalues are reported on the NBS scale. Samples for AT were poisonedwith 50 μL of saturated HgCl2 solution and stored in 500 mL bor-osilicate BOD bottles with ground-glass stoppers lightly coated withApiezon L® grease and kept in darkness at room temperature. Tem-perature and salinity were monitored during incubations by using asalinometer Eco Sense EC 300, YSI. AT was determined using the open-cell titration method (Dickson et al. 2007), by using an automatic Al-kalinity Tritrator Model AS-ALK2 Apollo SciTech. The AS-ALK2 systemis equipped with a combination pH electrode (8102BNUWP, ThermoScientific, USA) and temperature probe for temperature control (StarATC probe, Thermo Scientific, USA) connected to a pH meter (OrionStar A211 pH meter, Thermo Scientific, USA). All samples were ana-lyzed at 25 °C (± 0.1 °C) with temperature regulation using a water-bath (Lab Companion CW-05G). The accuracy was controlled against acertified reference material (CRM, supplied by Andrew Dickson, ScrippsInstitution of Oceanography, San Diego, USA) and the AT repeatabilityaveraged 2–3 μmol kg−1. Temperature and salinity data were used tocalculate the rest of carbonate system parameters (e.g. pCO2, CO3
2−)and the saturation stage of Omega Aragonite (Ωarag). Analyses wereperformed using CO2SYS software for MS Excel (Pierrot et al. 2006) setwith Mehrbach solubility constants (Mehrbach et al. 1973) refitted byDickson and Millero (1987). The KHSO4 equilibrium constant de-termined by Dickson (1990) was used for all calculations.
2.3. Physiological rates
2.3.1. Growth rateGrowth rates of juvenile individuals of A. purpuratus were estimated
from changes in the Total Weight (TW) (to the nearest 0.01 mg) of thescallops (Palmer 1982). For all measurements, each scallop was care-fully removed from the water, immediately weighed under the water(buoyant weight = BW), then gently blotted, and weighed in air (TW).
Table 1Average (+SE) conditions of carbonate system parameters during experiments conducted with juvenile scallops: pH (NBS scale), Total Alkalinity (TA in μmol kg−1), partial pressure ofCO2 (levels of pCO2 in seawater in μatm), carbonate ion concentration (CO3
−2 in μmol kg−1), saturation state of the water with respect to aragonite minerals (Ωarag).
Low (400 μatm) High (1000 μatm)
14 °C 18 °C 14 °C 18 °C
Temperature (°C) 14.13 ± 0.19 18.13 ± 0.11 14.08 ± 0.14 18.14 ± 0.13Salinity (PSU) 34.48 ± 1.78 33.28 ± 1.97 35.86 ± 0.50 34.00 ± 0.68pHNBS 8.058 ± 0.017 8.032 ± 0.041 7.754 ± 0.027 7.720 ± 0.021Total alkalinity (TA) 1580.41 ± 206.40 1666.75 ± 117.55 1778.14 ± 127.82 1695.96 ± 133.49CO3
−2 86.54 ± 10.15 94.76 ± 3.92 53.76 ± 5.58 52.13 ± 5.07pCO2 367.77 ± 62.17 435.96 ± 74.43 891.26 ± 77.76 969.48 ± 74.27Ωaragonite 1.32 ± 0.15 1.48 ± 0.06 0.82 ± 0.09 0.81 ± 0.08Ωcalcite 2.07 ± 0.24 2.29 ± 0.09 1.28 ± 0.13 1.25 ± 0.13
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
457

Furthermore we estimated biomass increase (i.e. metabolically activetissue) as the change in the relation between (TW–BW) during the timeof the experiment.
2.3.2. Standard metabolic rate (SMR)Scallops were left in aquaria for 24 h with no food after exposition
time. Then, we measured oxygen consumption (mg O2 × h−1 g−1)using a Presens Mini Oxy-4 respirometer (see Gaitán-Espitía et al.2017). To quantify the SMR, animals of each replicate were used andplaced individually in respirometric chambers filled with 805 mL ofseawater with the corresponding pCO2 levels and temperature of eachtreatment (see above). In addition 33 psu salinity and oxygen-saturatedseawater through air bubbling were added before starting measuring.The measurements were performed at a controlled temperature of 14 °Cand 18 °C using an automated temperature chiller. In each chamber,dissolved oxygen was quantified every 15 s for 55 min. Sensors werepreviously calibrated in anoxic water, using a saturated solution ofNa2SO3 and in water 100% saturated with oxygen using bubbled air.The same respirometric chambers were used as controls, but withoutanimals inside, under the same experimental conditions (the controlnever had a decayment of the oxygen concentration higher than 3% ofthe measurements). Each oxygen decayment due to background noisewas deducted from the individual measurements performed in the ex-perimental chambers. The first and the last 5 min were discarded inorder to avoid possible disturbances when the scallops were placed inrespirometric chambers. Thus, oxygen estimations are the average ofthe remaining 45 min of measurements. Before and after each mea-surement, bouyant weight of scallops were recorded using an analyticalbalance (ADAM AFA-180LC with precision± 0.001 mg). The averageof both body mass measurements was used in the statistical analyses.Slopes were calculated by the decrease of oxygen in the chamber perhour and normalized to fresh mass to get respiration rates in units ofmg O2 L−1 h−1.
2.3.3. Ingestion rate (IR) and clearance rate (CR)Immediately after completion of the experiment period (i.e.
18 days), we determined clearance and ingestion rates. We used1000 mL polycarbonate bottles previously cleaned with HCl.Individuals belonging to each experimental group were individuallysorted into 1000 mL acid-washed polycarbonate bottles filled with fil-tered seawater (1 μm, salinity of 33‰) and equilibrated with the re-spective CO2 treatment. Three control bottles without A. purpuratus andthree experimental bottles with one individual for each four treatmentwere incubated for approximately 2 h, and periodically rotated by handto avoid particle sedimentation. Previous to the experiment juvenilebody mass (mb) was determined with an analytical balance(± 0.01 mg) in order to standardize our clearance and ingestion rateestimates in g−1 h−1. For these experiments, we added Tetraselmis spp.at a concentration of ~32,000 cell/mL−1, to supply food at saturationlevel. Bottles were immersed in a container with flow-through seawatersystem used for maintaining temperature fluctuation during a givenexperiment within one degree. After incubation we took a sub-sampleof 200 mL from each bottle, which was filtered through a GF/F filter of0.75 μm mesh-size. The filter was stored in an aluminum envelope at−20 °C until analysis of Chlorophyll (Chl-a) concentration. Chl-a wasdark extracted in acetone 95% before measurement on a TD 6700Turner Fluorometer (Strickland and Parsons 1968). Clearance (CR) andIngestion (IR) rates were estimated through the measurement of Chl-adepletion and cell removal, according to Frost (1972) modified byMarín et al. (1986). For all experiments, there were no significant dif-ferences in food availability among treatments. No mortality effectswere found on juveniles scallops in any of the experimental treatments,which indicates that the projected increase of pCO2 levels and tem-perature had chronic, but not lethal, effects on these organisms.
2.4. Histopathological measurements
2.4.1. Histopathological analysisTissues for histology analysis were excised from scallops exposed to
the different treatments of pCO2 and temperature (N = 40) and fromthose 17 scallops representing natural control conditions. The tissuesexcised included gills, digestive gland and mantle. The tissue sampleswere fixed for 24 h. in Davidson's fluid, transferred to 70% ethanol, andsent to the Histopathology Laboratory of the Universidad Católica delNorte (Coquimbo, Chile). They were routine processed for histology,5 μm sections were cut, and stained with Harris Hematoxilyn and Eosin.The slides were analyzed using a Zeiss Axiostar plus microscope.
2.4.2. Pathological conditionsThe prevalence, percentage of individuals presenting each parasite
and condition was assessed for A. purpuratus from the four experimentaltreatments and the natural controls. For the two most prevalent in-dicators observed (rickettsiales-like organisms, RLOs, and digestivegland atrophy), also the intensity of infection was assessed. For theRLOs this was accomplished counting all the inclusions of these para-sites from the whole 5 μm thick section. The mean intensity of infectionwas calculated dividing the total number of parasites through the totalnumber of infected hosts. The intensity of digestive gland atrophy wasassessed registering the percentage of this condition when present, andthe mean percentage was calculated. Prevalence, infection intensity andmean intensity of infection were calculated following Bush et al.(1997).
2.5. Statistical analyses
To avoid pseudo-replication problems, the variables measured wereaveraged for the four scallops of each replicate. Two-way ANOVA wereused to evaluate whether temperature and CO2 levels and the interac-tion between factors affected growth rate (increase in TW and Biomassincrease with time), ingestion and clearance rate (Zar, 1999). A two-way analysis of covariance (ANCOVA) was performed to determine theeffects of temperature and pCO2 level on mass-specific metabolism,using body mass as covariate. When the analysis showed significantinteractions, a one-way ANOVA was carried out for each factor sepa-rately in each level from the other factor, followed by Tukey's a pos-teriori HSD test. When the analysis did not show significant interac-tions, multiple comparisons were carried out using Tukey's a posterioriHSD test on each factor that showed significant differences(Underwood, 1997). Buoyant weight increases were expressed in g perscallop per day. All analyses were carried out using Statistica version7.0. Assumptions of normality and homoscedasticity of the one-wayANOVA were evaluated using the Kolmogorov–Smirnov and Burtletttests, respectively (Sokal and Rohlf, 1995). Finally, we used an exactbinomial calculation to estimate the 2-tail probability that the pre-valence of conditions evaluated in the histological analysis of scalloptissues was higher or lower than expected by chance (p= 0.5) in nindependent trials or scallops observations (Zar 1999).
3. Results
3.1. Physiological rates
3.1.1. Growth rateGrowth rate of the scallops increased significantly in treatment with
higher temperature compared with individuals maintained at lowtemperature, 14 °C (Two way ANOVA, F1,19 = 24.40; P < 0.0000),but were not influenced by increased pCO2 levels within each tem-perature treatment (F1,19 = 0.97; P = 0.340) (Fig. 2a). Although, in-creased growth rate was also recorded in those scallops reared at thetreatment with low temperature and high pCO2 levels, representing atypical ‘upwelling’ condition, these do not lead to a significant
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
458
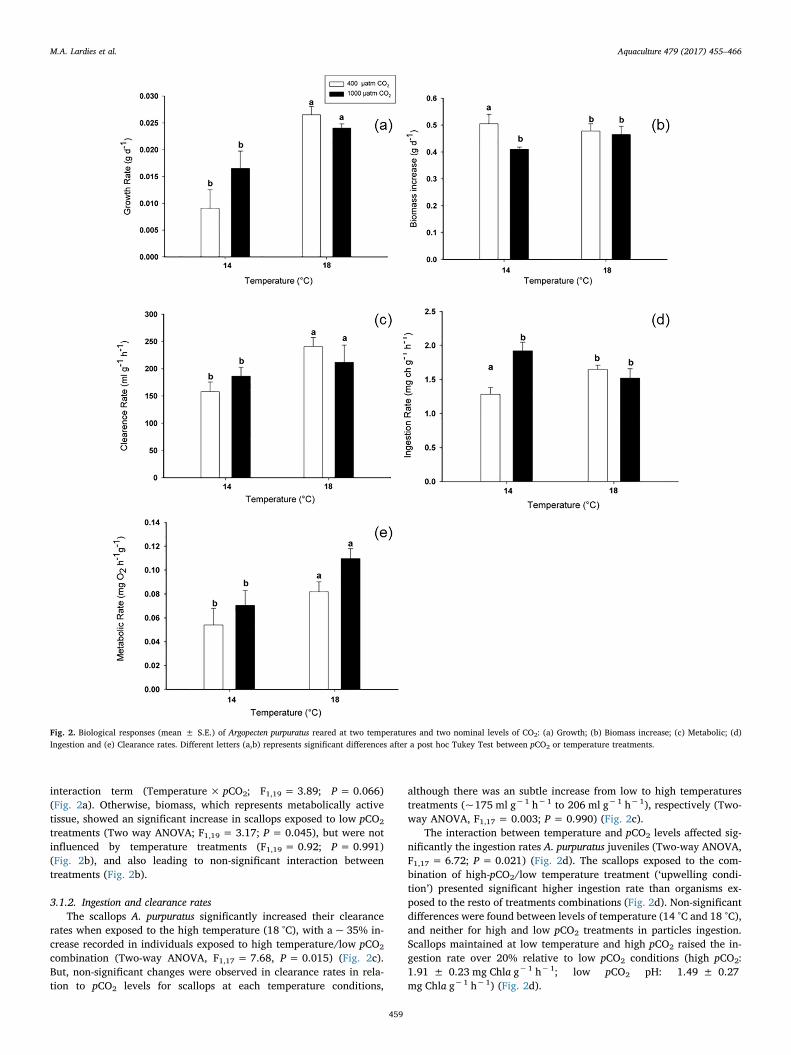
interaction term (Temperature × pCO2; F1,19 = 3.89; P= 0.066)(Fig. 2a). Otherwise, biomass, which represents metabolically activetissue, showed an significant increase in scallops exposed to low pCO2
treatments (Two way ANOVA; F1,19 = 3.17; P= 0.045), but were notinfluenced by temperature treatments (F1,19 = 0.92; P= 0.991)(Fig. 2b), and also leading to non-significant interaction betweentreatments (Fig. 2b).
3.1.2. Ingestion and clearance ratesThe scallops A. purpuratus significantly increased their clearance
rates when exposed to the high temperature (18 °C), with a ~ 35% in-crease recorded in individuals exposed to high temperature/low pCO2
combination (Two-way ANOVA, F1,17 = 7.68, P= 0.015) (Fig. 2c).But, non-significant changes were observed in clearance rates in rela-tion to pCO2 levels for scallops at each temperature conditions,
although there was an subtle increase from low to high temperaturestreatments (~175 ml g−1 h−1 to 206 ml g−1 h−1), respectively (Two-way ANOVA, F1,17 = 0.003; P = 0.990) (Fig. 2c).
The interaction between temperature and pCO2 levels affected sig-nificantly the ingestion rates A. purpuratus juveniles (Two-way ANOVA,F1,17 = 6.72; P = 0.021) (Fig. 2d). The scallops exposed to the com-bination of high-pCO2/low temperature treatment (‘upwelling condi-tion’) presented significant higher ingestion rate than organisms ex-posed to the resto of treatments combinations (Fig. 2d). Non-significantdifferences were found between levels of temperature (14 °C and 18 °C),and neither for high and low pCO2 treatments in particles ingestion.Scallops maintained at low temperature and high pCO2 raised the in-gestion rate over 20% relative to low pCO2 conditions (high pCO2:1.91 ± 0.23 mg Chla g−1 h−1; low pCO2 pH: 1.49 ± 0.27mg Chla g−1 h−1) (Fig. 2d).
Fig. 2. Biological responses (mean ± S.E.) of Argopecten purpuratus reared at two temperatures and two nominal levels of CO2: (a) Growth; (b) Biomass increase; (c) Metabolic; (d)Ingestion and (e) Clearance rates. Different letters (a,b) represents significant differences after a post hoc Tukey Test between pCO2 or temperature treatments.
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
459

3.1.3. Standard metabolic rate (SMR)Metabolic rate of A. purpuratus, measured as oxygen consumption
increased significantly at the high temperature condition (Two-wayANCOVA, F1,17 = 7.29; P = 0.017) (Fig. 2e). Despite that oxygenconsumption in scallops was higher in both high pCO2 treatments, thesedifferences were not significant (F1,17 = 3.35; P= 0.088). Further-more, a non-significant Temperature × pCO2 interaction was observedfor metabolic rate (Fig. 2e) (Two-way ANCOVA, F1,17 = 0.45;P = 0.515). Mass-specific metabolic rate in juveniles of A. purpuratuswas lower in the low temperature/low-pCO2 level treatment than in lowtemperature/high-pCO2 level combination (0.73 ± 0.19 and0.97 ± 0.37 mg O2 h−1 g−1, respectively). Metabolic rate of in-dividuals exposed to high temperature and high pCO2 level reached thehighest values observed in the experiment (1.60 ± 0.40mg O2 h−1 g−1), almost doubling oxygen consumption of scallops ex-posed to low temperature conditions (Fig. 2e).
3.2. Histopathological measurement
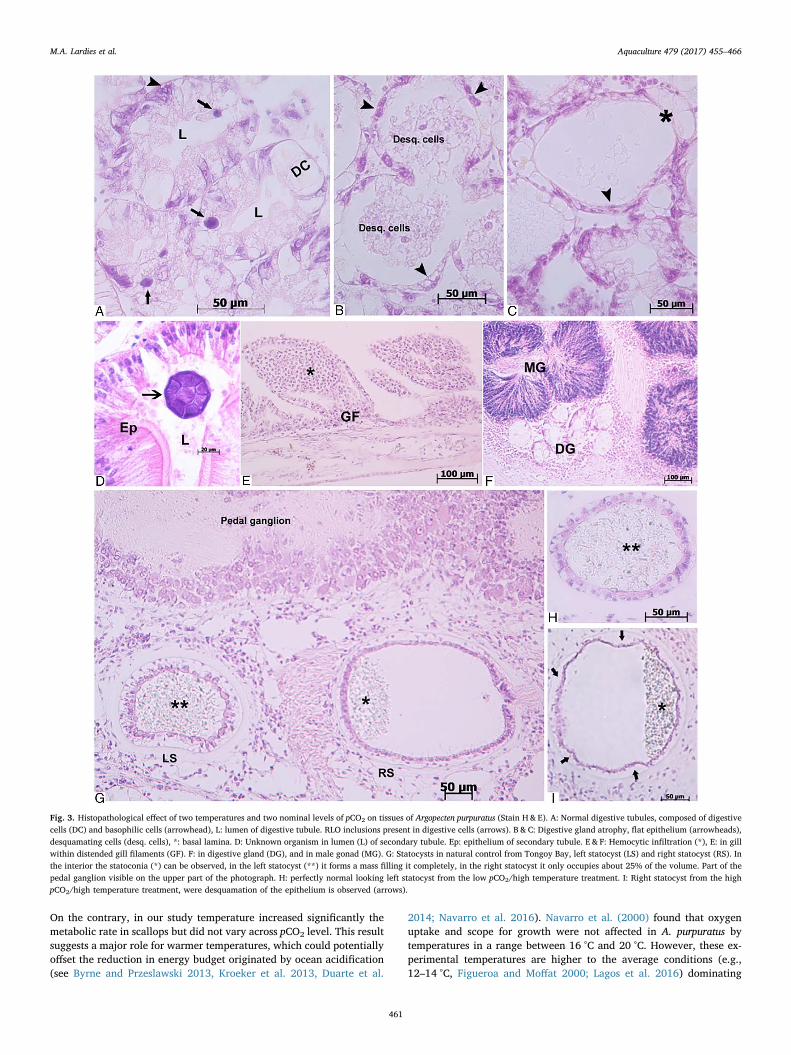
3.2.1. Histopathological analysisThe main affected tissue in juvenile scallops was the digestive gland,
showing degenerative alterations of the digestive tubules (Fig. 3anormal tubule) and secondary (transport) tubules across treatments(Fig. 3b,c). This alteration was evidenced as a desquamation of theepithelial cells (Fig. 3b), flattening the epithelium, and thus in someextreme cases leaving only the presence of the basal lamina (Fig. 3c).The secondary tubule atrophy, although present, showed a low in-tensity across all treatments. Although statocysts had not specificallybeen sampled for, they appeared located close to the pedal ganglia andthe digestive gland (Fig. 3g, right and left statocysts from naturalcontrol scallops). The left statocyst showed no alteration in the ex-perimental scallops (Fig. 3h), however, the right statocyst showed ir-regular outlines, due to epithelial desquamation in the high pCO2
condition at both levels of temperature (Fig. 3i).
3.2.2. Pathological conditionsThe prevalence of parasites and pathological conditions is shown in
Table 2. The rickettsiales–like organisms (RLOs) were detected as ba-sophilic, ovoid-spherical inclusions in epithelial cells of the digestivetubules (Fig. 3a), residing in otherwise completely normal digestivecells. RLOs showed low prevalence across the experimental treatmentsand higher, but non-significant prevalence increment in scallops naturalcontrol conditions. These RLOs were most prevalent in the naturalcontrol scallops (24%) and ranged from 0% to 10% in experimentaltreatments (Table 2). The intensity of infection by RLOs ranged from 1to 27 inclusions counted from one section, with a mean intensity of 8 inthe case of natural control scallops (Table 2). Another, unidentifiedmicroorganism, showed increased and significant prevalence in theLumina of digestive and secondary tubules, but also only in the naturalcontrol scallops (76%, Table 2); it had a strongly eosinophilic thickcover resembling a resting cyst, sometimes with cells inside (Fig. 3d).Hemocyte infiltration was also observed in gills (Fig. 3e) and digestivegland (Fig. 3f) mainly, and occasionally in gonad as well (Fig. 3f). Thishemocyte infiltration in gills was increased (70%) but not significantly,at the high-pCO2/low temperature combination (Table 2). At lowtemperature and irrespective of pCO2 level, the 100% of individualsshowed digestive tubule atrophy (Table 2). The approximate coveragepercentage (intensity) of this condition ranged from 5 to 30% in thenatural control scallops and varies across experimental treatments,showing the highest increment in scallops raised under the low tem-perature and high-pCO2 condition (39%; 20%–60%, Table 2).
4. Discussion
Our results indicate that under rise in pCO2 from 400 to 1000 μatmin a scenario of low temperature, which may resemble the influence of
upwelling conditions operating in Tongoy Bay, did not affect mostphysiological rates measured in juveniles of A. purpuratus, except for asignificant increase in about of 40% of the ingestion rate at the highpCO2 scenario. Nevertheless, physiological processes were sensitive tochanges in temperature, which induced a positive effect on the phy-siological processes associated with increased energy acquisition.Specifically, our factorial experiment showed that the environmentaldrivers studied, temperature and pCO2 operate in an additive way uponthe physiological performance of A. purpuratus from Tongoy Bay.Nevertheless, the current study also suggests that under experimentalconditions the combination of low temperature across both pCO2 con-ditions may reduce the health of A. purpuratus. Histopathological as-sessment of the soft tissues of juvenile scallops exposed to acidificationand temperature treatments, showed damage, particularly in digestivegland, but without mortality. In addition, though not fully assessed, italso suggests the role of high pCO2 on statocyst volume.
Temperature profoundly affects growth and its underlying processesin ectotherms (Angilletta 2009; Somero 2010), rising temperature leadsto increasing rates of biochemical-physiological processes (e.g. meta-bolic and heart rate) and life-history characteristics (Gillooly et al.2001; Willmer et al. 2005). Temperature increases the growth rate ofscallops, an expected pattern because temperature increases growth andbody size in marine invertebrates (see Sicard et al., 1999; Woods et al.2003; Angiletta et al. 2004). The increase of growth rate with tem-perature increment has been demonstrated previously in A. purpuratusfor populations from Chile and Perú (see Martínez and Pérez 2003;Thébault et al. 2008; Soria et al. 2011; Lagos et al. 2016). Water tem-perature is the environmental factor most often cited as influencingbivalve reproduction (see Sastry 1968; Wolff 1998; Navarro et al.2016). Nevertheless, we detected a significant increase in biomass ofthe scallops (i.e., metabolically active tissue) raised under low pCO2
levels. Thus, juveniles scallops experiencing high pCO2 conditions,probably due to higher energetic cost of homeostasis, decrease thetissue mass (see Lardies et al. 2014). Recently, we reported a reductionin shell length and growth in juveniles of A. pururatus from Tongoy Bayunder high pCO2 treatments (Lagos et al. 2016). Reduced growth underacidified conditions has been reported also for other scallops species(e.g. Talmage and Gobler 2009; White et al. 2013). A similar patternwas informed in the bivalve Mytilus galloprovincialis where high pCO2
significantly reduced growth rate at low temperature but this effectgradually decreased with warming (Kroeker et al. 2014). Negativeimpacts on biomass under low pH could be explained because energyallocation is being diverted away to other key physiological processessuch as calcification, acid–base balance, reproduction, and immunefunction (Wood et al. 2008; Findlay et al. 2009; Lagos et al. 2016).Nevertheless, previous studies report that juveniles of A. purpuratusmaintained between 10 °C and 18 °C showed no effect in growth rateand their scope for growth (Gonzalez et al. 2002). The origin of ex-perimental populations may explain this contradictory results, whereasthe juveniles scallops used in our experiment were obtained fromTongoy Bay (northern Chile, 30° 12′S), these authors collected scallopsfrom southern Chile (41° 54′S), thus suggesting the role of a possibleacclimatization and/or adaptation of local populations to environ-mental factors experienced in their native habitats. Temperature re-gimes vary widely along the Chilean coast and specifically in> 6 °Cbetween these two localities (Thiel et al. 2007; Lagos et al. 2008). Thus,geographical variation in the degree of plasticity and/or potential forlocal adaptation for these traits could be present within A. purpuratusspecies along the Chilean coast and partially may explain the differ-ences in biological responses observed between local population of A.purpuratus.
There are few studies on combined effects of temperature and pCO2
on metabolism and performance of marine invertebrates (Queirós et al.,2015). For instance, Navarro et al. (2016) found that the metabolic ratewas not affected by temperature but detected an increase in metabolismon the farmed mussels Mytilus chilensis when exposed to low pH level.
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
460
On the contrary, in our study temperature increased significantly themetabolic rate in scallops but did not vary across pCO2 level. This resultsuggests a major role for warmer temperatures, which could potentiallyoffset the reduction in energy budget originated by ocean acidification(see Byrne and Przeslawski 2013, Kroeker et al. 2013, Duarte et al.
2014; Navarro et al. 2016). Navarro et al. (2000) found that oxygenuptake and scope for growth were not affected in A. purpuratus bytemperatures in a range between 16 °C and 20 °C. However, these ex-perimental temperatures are higher to the average conditions (e.g.,12–14 °C, Figueroa and Moffat 2000; Lagos et al. 2016) dominating
Fig. 3. Histopathological effect of two temperatures and two nominal levels of pCO2 on tissues of Argopecten purpuratus (Stain H& E). A: Normal digestive tubules, composed of digestivecells (DC) and basophilic cells (arrowhead), L: lumen of digestive tubule. RLO inclusions present in digestive cells (arrows). B & C: Digestive gland atrophy, flat epithelium (arrowheads),desquamating cells (desq. cells), *: basal lamina. D: Unknown organism in lumen (L) of secondary tubule. Ep: epithelium of secondary tubule. E & F: Hemocytic infiltration (*), E: in gillwithin distended gill filaments (GF). F: in digestive gland (DG), and in male gonad (MG). G: Statocysts in natural control from Tongoy Bay, left statocyst (LS) and right statocyst (RS). Inthe interior the statoconia (*) can be observed, in the left statocyst (**) it forms a mass filling it completely, in the right statocyst it only occupies about 25% of the volume. Part of thepedal ganglion visible on the upper part of the photograph. H: perfectly normal looking left statocyst from the low pCO2/high temperature treatment. I: Right statocyst from the highpCO2/high temperature treatment, were desquamation of the epithelium is observed (arrows).
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
461

during upwelling processes influencing Tongoy Bay. Similar resultswere found in the scallop Pecten maximus where the respiration rate wasnot affected in juveniles displaying tolerance to levels of pH rangingbetween 8.2 and 7.6 (Sanders et al. 2013). It is important to emphasizethat in our experiment, as well as in the Sanders et al. (2013) study,food availability was not restricted. In addition to temperature, foodsupply seems to play a major role modulating organismal responses ofA. purpuratus by providing the energetic means to support the physio-logical cost imposed by ocean acidification stress (Ramajo et al. 2016a).
Our results can be discussed in the framework of natural pCO2
variability in coastal upwelling areas influencing scallop farming andthe potential for local adaptation in these regions (Vargas et al. 2017).This can be rationalized because the scallops were exposed to a pH andtemperature range that the animals experience in their natural en-vironment (see Torres and Ampuero 2009; Lagos et al. 2016; Vargaset al. 2017). Coastal upwelling modulates the physical and chemicalproperties of seawater over large areas of the coastal ocean. In relationto the current study, the Pt. Lengua de Vaca upwelling center increasesCO2-fluxes and promotes strong reductions in pH (pH ~ 7.6) in TongoyBay and the surrounding area (Torres and Ampuero 2009; Torres et al.2011) and adds also a high variability in sea surface temperature (seeAravena et al. 2014). That is, the moderate effect found on physiolo-gical performance of juvenile individuals of A. purpuratus from TongoyBay is likely due to the presence of phenotypic plasticity leading toincreased tolerances which is characteristic of organisms that inhabitheterogeneous environments (Via et al. 1995). Most upwelling zonesare acidic compared to the coastal and open ocean (Hofmann et al.2011). Furthermore, it has recently been demonstrated that most up-welling zones are indeed characterized by natural variations in sea-water chemistry, suggesting that in some particular habitats, residentorganisms are already experiencing pH regimes projected for the year2100 (Hofmann et al. 2011; Vargas et al. 2017). This can be the case ofthe Tongoy Bay due to the presence of a semi-permanent upwelling(Vargas et al. 2017). Our results evidenced that a less severe response tohigh pCO2 levels of the scallops that are naturally exposed to higher pHand temperature variability, suggesting the acclimatization and adap-tation potential for this species. This moderate effect of temperatureand high pCO2 has also been reported in other marine invertebrates thatinhabit highly variable environments on terms of pH and temperature(see Lardies et al. 2014; Collard et al. 2016; Vargas et al. 2017). Weobserved an increase of scallop metabolism in high pCO2 at both tem-peratures. This up-regulation observed in metabolic rate of scallopsunder high pCO2 scenarios has also been observed in others bivalvesspecies exposed to low pH conditions, and may be a requirement formaintaining intracellular pH, and cellular homeostasis (e.g., Cummingset al. 2011; Lardies et al. 2014; Ramajo et al. 2016a), which is con-sistent with the metabolic costs of mechanisms developed by calcifiers
to cope with acidic conditions (Ramajo et al. 2016b; Hendriks et al.2015). Metabolic up-regulation to face stressful environments is en-ergetically costly and may impose limits to the energy available tosustain other physiological activities like growth or ingestion rate,generating trade-offs between phenotypic traits (Wood et al. 2008;Deigweiher et al. 2010; Lardies et al. 2014; Navarro et al. 2016).
Studies analyzing the effect of high pCO2 on bivalve species haveshown negative effects on feeding activities, such as clearance and in-gestion rates, possibly due to deficiencies in the functioning of the di-gestive and filtering systems (Fernández-Reiriz et al. 2011; Navarroet al. 2013; Vargas et al. 2013; Vargas et al. 2015). Reduced feedingunder ocean acidification and warming conditions results in lowersurvival, size, and calcification (Dupont & Thorndyke, 2009;Byrne & Przeslawski 2013). In contrast, in our study juvenile scallopsshowed a significant increase of ingestion rate at high pCO2/low tem-perature treatment (i.e., upwelling conditions) and clearance rate wasincreased as expected in higher temperatures without effect of pH. It isevident from our results and the other available studies that high pCO2
increases juvenile feeding of scallop species. Our findings are consistentwith results reported for other scallops such as Chlamys nobilis andPecten maximus under low pH treatments (Wenguang &Maoxian, 2012;Sanders et al., 2013) and in the juveniles of A. purpuratus under dif-ferent scenarios of pH and food availability (Ramajo et al. 2016a).Navarro et al. (2000) show that A. purpuratus can actively regulateclearance rate and does not simply switch between feeding and non-feeding states. A different pattern have been reported in other molluskspecies for the Chilean coast, for instance in newly hatched larvae of thegastropod Concholepas concholepas and juvenile stages of the musselPerumytilus purpuratus, whose clearance rates decreased between 15 upto 70% under high pCO2 respect to control conditions (Vargas et al.2013; Vargas et al. 2015). On the other hand, the maintenance ofscallop biomass across treatments (see Fig. 2b) may require high foodingestion at the same time that more energy needs to be canalized to theshell calcification process (see Lagos et al. 2016). In upwelling areas theexposure of the organisms to low temperatures is concomitant withhigh-pCO2, which would depress the saturation state for carbonatebelow the level derived from high pCO2 alone (Feely et al. 2008). Theprevious is because the kinetics of dissolution/solubility of carbonateshells increases at low temperatures (Morse et al. 2007). These “up-welling conditions” experienced by scallops in our experimental treat-ments (i.e. low temperature and high pCO2) are already experienced byscallop farmed in Tongoy Bay, hence an adaptation to local environ-mental conditions trough phenotypic plasticity may be part of the re-sponses expressed by this scallop population. It is important to maintaina perspective about the several organismal responses as an integratedphenotype because low pH regimes can significantly affect calcificationprocess in A. purpuratus, which becomes enhanced under low
Table 2Prevalence (P) of pathological conditions evaluated in scallops collected from field (natural control) and under the four experimental CO2/temperature combinations. Mean infectionintensity (I) was also evaluated for Rickettsia-like organisms and digestive tubule atrophy. Significant prevalence evaluated using the 2-tail binomial probability test is shown in bold. Thenumber of scallops used in each evaluation (n) is also showed.
Condition evaluated Natural control Low (400 μatm) High (1000 μatm)
14 °C 18 °C 14 °C 18 °C
(n = 17) (n = 10) (n = 10) (n = 10) (n = 10)
Rickettsia-Like organisms (P) 24 10 10 0 10Rickettsia-Like organisms (I) 8 (1–27) 1 8 (8) 0 1Unidentified microorganism (P) 76 0 0 0 0Gill hemocyte infiltration (P) 18 40 40 70 50Digestive gland infiltration (P) 0 50 90 40 100Digestive tubule atrophy (P) 24 100 80 100 80Digestive tubule atrophy (I) 18% (5–30) 12% (5–30) 35% (10–40) 39% (20–60) 24% (10–40)Secondary tubule atrophy (P) 0 20 50 10 40Distended gill filaments (P) 0 10 10 30 30
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
462

temperatures such as those occurring during an active upwelling phase(Lagos et al. 2016; Vargas et al. 2017). Thus, the presence of trade-offsin energy allocation during stress induced by upwelling processes ispresent at least in terms of calcification, growth rate and metabolism.Therefore, studies that aim to improve our understanding about themechanisms that govern bivalve aquaculture species in terms of calci-fication, growth and physiological performance in upwelling zones areurgently needed.
For A. purpuratus the most striking histopathological effect was onthe digestive gland integrity, which consisted in disintegrating andsloughing of digestive cells. Many stressors can generate atrophied di-gestive tubules, such as pollution (Syasina et al. 1997), food avail-ability, saline and thermal stress (Carella et al. 2015). We measuredthat natural only a small proportion of scallops collected from the farm(natural control) showed reconstituting tubules morphology (Mathers1976, Henry et al. 1991, Mathers et al. 1979, Syasina et al. 1997),which was atrophied (60%) in scallops subjected to the experimentalconditions. This implies a reduction of digestive tubules available foraccomplishing intracellular digestion, which may alter an adequateingestion during the exposure time. Clearly, this pattern of tubulesatrophy in scallops across treatments may be associated with differ-ences between the natural diet from Tongoy Bay and the artificial foodused during the experiment. It has been reported for juvenile bivalvemolluscs that food availability can outweigh ocean acidification, asshown forMytilus edulis (Thomsen et al. 2013), Pecten maximus (Sanderset al. 2013) and Argopecten purpuratus (Ramajo et al., 2016a, b). Thislatter study was undertaken in a population of juvenile scallops fromthe same population as this study and consisted in a 30-day exposure todifferent pH and food concentrations (Ramajo et al. 2016a) similar tothose used in our incubations. Thus, an explanation of this positiveresponses could be the existence of a greater proportion of healthy di-gestive tubules, providing the capacity to digest the available food, andthe amelioration of ocean acidification effects trough increased tem-perature.
Hemocytic infiltration is an inflammatory response to pathogens,chemicals (such as pollutants) or physical cell injury (De Vico andCarella 2012; Carella et al. 2015). High prevalences but non-significantgill hemocytic infiltration were observed in the experimental treat-ments, respect to the natural control group. Gills had highest pre-valence in the high temperature treatment (i.e., independent of pCO2
level). Mytilus edulis showed highest hemocytes infiltration when ex-posed to high temperature conditions, but was absent under high pCO2
treatments (Mackenzie et al. 2014). In addition, two parasites or sym-bionts were found in this study, a prokaryote, rickettsiales-like or-ganism (RLO) and an unidentified protistan. Rickettsiales-like organ-isms (RLO) have been described as basophilic intracellular inclusionbodies in the gills or digestive gland of most bivalve molluscs (Boweret al. 1994), usually not causing any harm. They have also been de-scribed from digestive gland of adult cultivated A. purpuratus fromCaldera, and the bays of Guanaqueros and Tongoy (Lohrmann 2009). Inthis study RLOs were most prevalent in the natural control group thanin the experimental treatment, but with similar mean intensity. RLOsprevalence have though been linked to serious mortalities in some bi-valves such as in the scallop Pecten maximus (Legall et al. 1988), giantclams Hippopus hippopus (Norton et al. 1993) and clams Venerupisrhomboides (Villalba et al. 1999). Although, RLOs have always beendetected at low prevalence and intensity of infection in A. purpuratus,with no host hemocyte responses, is suggested that a harmless parasitecan become a pathogen under the influence of as climate stressors thatcan enhance the virulence and/or the parasite pathogenicity (Burgeet al. 2014).
In scallops, statocysts are sacs formed by a ciliated epithelium witha single statolith and a loose statoconia filling about 25% up to 100% ofthe sac, lie close to the pedal ganglion, are asymmetrical and functionas gravity receptor, which epithelium development, cell types and ciliaallow to differentiate the left and right statocysts (Cragg and Nott
1977). In our study the right statocyst observed in scallop exposed tothe high pCO2 treatment suggests that the statoconia have been par-tially lost due to increased pCO2 or acidification conditions. Observa-tion on scallops farmed at Tongoy bay (natural control), evidenced aright statocyst less developed and partially filled with statoconia, as hasbeen described for other pectinids (e.g., Von Buddenbrock 1915). Inaddition, this right statocyst of the high pCO2 treatment, regardless ofthe temperature, showed some desquamation of the epithelium. It is notclear if this loss of integrity of the right statocyst epithelium couldimpair the gravity perception of the scallops, as only removal of the leftone seemed to cause harm in other pectinids (Buddenbrock 1915).Recently, Schalkhausser et al. (2013) demonstrate that Pecten maximusreduce their clapping performance when exposed to low temperatureand increased pCO2 conditions suggesting potential link between theimpact on the escape behaviour of scallops and the reduced epithelialintegrity of the statocyst observed in our study.
There are several possible reasons to explain the moderate effect inA. purpuratus of ocean acidification and in scallops in general; firstly,the main mineral component of the shell is entirely of calcite a lesssoluble form of carbonates (Gazeau et al. 2013) and also a secretion of athicker periostracum has been reported involving higher levels ofpolysaccharides under OA conditions as shell protection mechanism injuveniles (Ramajo et al. 2016a). Secondly, seawater remained in alltreatments in our experiments above the calcite saturation state(Ωcalcite > 1). Lastly, Scanes et al. (2014) propose that swimming ca-pacity of scallops which produce elevations of CO2 in the haemolymphand alterations in acid-base status and is another possible reason for themoderate effects of ocean acidification on scallops when compared tothe negative physiological effects widely reported in oysters and mus-sels which are sessile mollusks. The previous is applicable for wildscallops in natural stocks; nevertheless, this mechanism is potentiallyless plausible in conditions of suspended culture where density in thelantern net is high (i.e., from 200 to 25 scallops per lantern level inTongoy Bay farm), which reduces the opportunities for swimming injuvenile scallops. However, the underlying mechanism by which highpCO2 impacts moderately scallops physiology remains unclear for us.
Although our treatment of low temperature and high pCO2 or “up-welling conditions” did not evidence strong effect on the studied phy-siological traits, it is still uncertain the potential effect of future oceanacidification upon biological responses measured. One of the mainproblems in order to evaluate the effect of ocean acidification in marineorganisms inhabiting these regions is the lack of information regardingto potential scenarios of ocean acidification for eastern boundary re-gions. There is no consensus of how the effects of warming and oceanacidification will interact (e.g. additive, synergistic, or antagonistic) toinfluence the physiology and performance and health of mollusks (e.g.Findlay et al. 2009, Gooding et al. 2009, Comeau et al. 2010, Byrne2011, Kroeker et al. 2013, Byrne and Przeslawski 2013; Duarte et al.2014; Lagos et al. 2016; Navarro et al. 2016). Although high pCO2
significantly reduced growth rate of scallops at 14 °C, this effect is notsignificant at temperatures of 18 °C, showing how moderate warmingcan ameliorate the effects of OA through temperature effects on bothphysiology and histopathology. However, these scenarios of progressiveacidification and warming may be not fully applicable to highly pro-ductive upwelling ecosystems, where the projected intensification ofupwelling events due to the change in the intensity and patterns of wind(Sydeman et al. 2014) forecast more extreme upwelling events withcold subsurface waters, which are supersatured with dissolved CO2
during longer periods. Therefore, the consideration of this kind of cli-matic scenarios in multistressor experiments is urgently needed forshellfish aquaculture species cultivated in upwelling zones in such as,for instance North America, Iberia and Africa.
Acknowledgements
We thank Pamela Tapia and Christian Tapia at Invertec–Ostimar
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
463

Hatchery and fieldwork facilities. We extend our acknowledgment toJorge López, and Paulina Contreras for their valuable assistance duringthe experiments and fieldwork. We thank two anonymous reviewers forinsightful criticisms to the manuscript. This study was supported by theMillennium Nucleus Center for the Study of Multiple drivers on MarineSocio-Ecological Systems (MUSELS) funded by MINECON NC120086and FONDECYT grant nos. 1140938 and 1140092 to N.A.L. and M.A.L.,and 1130254 to CAV. CAV was also funded by the Millennium Instituteof Oceanography (IMO), MINECON IC120019 during the preparation ofthis manuscript.
References
Angilletta Jr., M.J., 2009. Thermal Adaptation: A Theoretical and Empirical Synthesis.Oxford University Press, Oxford.
Angilletta Jr., M.J., Oufieroa, C.E., Searsb, M.W., 2004. Thermal adaptation of maternaland embryonic phenotypes in a geographically widespread ectotherm. Int. Congr.Ser. 1275, 258–266.
Aravena, G., Broitman, B., Stenseth, N.C., 2014. Twelve years of change in coastal up-welling along the central-northern coast of Chile: spatially heterogeneous responsesto climatic variability. PLoS One 9, e90276.
Beniash, E., Ivanina, A., Lieb, N.S., Kurochkin, I., Sokolova, I.M., 2010. Elevated level ofcarbon dioxide affects metabolism and shell formation in oysters Crassostrea virginica.Mar. Ecol. Prog. Ser. 419, 95–108.
Bouvet, J.M., Vigneron, P., Saya, A., 2005. Phenotypic plasticity of growth trajectory andontogenic allometry in response to density for Eucalyptus hybrid clones and families.Ann. Bot. 96, 811–821.
Bower, S., McGladdery, S., Price, I., 1994. Synopsis of infectious diseases and parasites ofcommercially exploited shellfish. Annu. Rev. Fish Dis. 4, 1–199.
von Brand, E., Merino, G., Abarca, A., Stotz, W., 2006. Scallop fishery and aquaculture inChile. In: Parsons, J. (Ed.), Scallop, Biology, Ecology and Aquaculture: Developmentsin Aquaculture and Fisheries Sciences. Elsevier, The Netherlands, pp. 1293–1311(1460).
Buddenbrock, W., 1915. Die Statocyste von Pecten, ihre Histologie und Physiologie. ZoolJb Abt Anat Physiol Bd 35, 302–358.
Burge, C.A., Eakin, C.M., Friedman, C.S., Froelich, B., Hershberger, P.K., Hofmann, E.E.,Petes, L.E., Prager, K.C., Weil, E., Willis, B.L., Ford, S.E., Harvell, C.D., 2014. ClimateChange Influences on Marine Infectious Diseases: Implications for Management andSociety. In: Carlson, C.A., Giovannoni, S.J. (Eds.), Ann. Rev. Mar. Sci. Vol 6 (Book 6).
Bush, A.O., Lafferty, K.D., Lotz, J.M., Shostak, A.W., 1997. Parasitology meets ecology onits own terms: Margolis et al. revisited. J. Parasitol. 83 (4), 575–583.
Byrne, M., 2011. Impact of ocean warming and ocean acidification on marine in-vertebrate life history stages: vulnerabilities and potential for persistence in achanging ocean. Oceanogr. Mar. Biol. Annu. Rev. 49, 1–42.
Byrne, M., Przeslawski, R., 2013. Multistressor impacts of warming and acidification ofthe ocean on marine Invertebrates' life histories. Integr. Comp. Biol. 53, 582–596.
Carella, F., Feist, S.W., Bignell, J.P., De Vico, G., 2015. Comparative pathology in bi-valves: aetiological agents and disease processes. J. Invertebr. Pathol. 131, 107–120.
Chavez, F.P., Bertrand, A., Guevara, R., Soler, P., Csirke, J., 2008. The northern HumboldtCurrent System: brief history, present status and a view towards the future. Prog.Oceanogr. 79, 95–105.
Collard, M., Rastrick, S.P.S., Calosi, P., Demolder, Y., Dille, J., Findlay, H.S., Hall-Spencer,J.M., Milazzo, M., Moulin, L., Widdicombe, S., Dehairs, F., Dubois, P., 2016. Theimpact of ocean acidification and warming on the skeletal mechanical properties ofthe sea urchin Paracentrotus lividus from laboratory and field observations. ICES J.Mar. Sci. 73 (3), 727–738.
Comeau, S., Gorsky, G., Alliouane, S., Gattuso, J.P., 2010. Larvae of the pteropodCavolinia inflexa exposed to aragonite undersaturation are viable but shell-less. Mar.Biol. 157, 2341–2345.
Cooley, S.R., Doney, S.C., 2009. Anticipating ocean acidification's economic consequencesfor commercial fisheries. Environ. Res. Lett. 4 (2), 024007.
Cragg, S.M., Nott, J.A., 1977. Ultrastructure of statocysts in pediveliger larvae of Pectenmaximus (l) (bivalvia). J. Exp. Mar. Biol. Ecol. 27, 23–36.
Cummings, V., Hewitt, J., Van Rooyen, A., Currie, K., Beard, S., Thrush, S., Norkko, J.,Barr, N., Heath, P., Halliday, N.J., Sedcole, R., Gomez, A., McGraw, C., Metcalf, V.,2011. Ocean Acidification at High Latitudes: Potential Effects on Functioning of theAntarctic Bivalve Laternula elliptica. PLoS ONE 6 (1) e16069.
De Vico, G., Carella, F., 2012. Morphological features of the inflammatory response inmolluscs. Res. Vet. Sci. 93, 1109–1115.
Deigweiher, K., Hirse, T., Bock, C., Lucassen, M., Pörtner, H.O., 2010. Hypercapnia in-duced shifts in gill energy budgets of Antarctic notothenioids. J. Comp. Physiol.180B, 347–359.
Dickson, A.G., 1990. Standard potential of the reaction AgCl(s)+1/2 H2(g) = Ag(s)+HCl(aq) and the standard acidity constant of the ion HSO4− in synthetic sea waterfrom 273.15 to 318.15 K. J. Chem. Thermodyn. 22, 113–127.
Dickson, A.G., Millero, F.J., 1987. A comparison of the equilibrium constants for thedissociation of carbonic acid in seawater media. Deep-Sea Res. 34 (10A), 1733–1743.
Dickson, A.G., Sabine, C.L., Christian, J.R. (Eds.), 2007. Guide to Best Practices for OceanCO2 Measurements. PICES Special Publication Vol. 3 North Pacific Marine SciencesOrganization, Sidney.
Duarte, C., Navarro, J.M., Acuña, K., Torres, R., Manríquez, P.H., Lardies, M.A., Vargas,C.A., Lagos, N.A., Aguilera, V., 2014. Combined effects of temperature and ocean
acidification on the juvenile individuals of the mussel Mytilus chilensis. J. Sea Res. 85,308–314.
Duarte, C., Navarro, J., Acuña, K., Torres, R., Manríquez, P.H., Lardies, M.A., Vargas,C.A., Lagos, N.A., Aguilera, V., 2015. Intraspecific variability in the response of theedible mussel Mytilus chilensis (Hupe) to ocean acidification. Estuar. Coasts 38,590–598.
Dupont, S., Thorndyke, M.C., 2009. Impact of CO2-driven ocean acidification on in-vertebrates early life-history-what we know, what we need to know and what we cando. Biogeosciences 6, 3109–3131.
Fabry, V.J., Seibel, B.A., Feely, R.A., Orr, J.C., 2008. Impacts of ocean acidification onmarine fauna and ecosystem processes. ICES J. Mar. Sci. 65, 414–432.
Feely, R.A., Sabine, C.L., Hernandez-Ayon, J.M., Ianson, D., Hales, B., 2008. Evidence forupwelling of corrosive ‘acidified’ water onto the continental shelf. Science 320,1490–1492.
Fernández-Reiriz, M., Navarro, J.M., Labarta, U., 2005. Enzymatic and feeding behaviourof Argopecten purpuratus under variation in salinity and food supply. Comp. Biochem.Physiol. 141A, 153–163.
Fernández-Reiriz, M.J., Range, P., Alvarez-Salgado, X.A., Labarta, U., 2011. Physiologicalenergetics of juvenile clams Ruditapes decussatus in a high CO2 coastal ocean. Mar.Ecol. Prog. Ser. 433, 97–105.
Figueroa, D., Moffat, C., 2000. On the influence of topography in the induction of coastalupwelling along the Chilean coast. Geophys. Res. Lett. 27, 3905–3908.
Findlay, H.S., Wood, H.L., Kendall, M.A., Spicer, J.I., Twitchett, R.J., Widdicombe, S.,2009. Calcification, a physiological process to be considered in the context of thewhole organism. Biogeosciences 6, 2267–2284.
Frost, B.W., 1972. Effects of size and concentration of food particles on the feeding be-havior of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 17,805–815.
Gaitán-Espitia, J.D., Bacigalupe, L.D., Opitz, T., Lagos, N.A., Osores, S., Lardies, M.A.,2017. Thermal physiological responses of an intertidal crab along a latitudinal cline.J. Theor. Biol. http://dx.doi.org/10.1016/j.jtherbio.2017.02.011.
Garreaud, R., Rutllant, J.A., Muñoz, R.C., Rahn, D.A., Ramos, M., Figueroa, D., 2011.VOCALS-CUpEx: the Chilean upwelling experiment. In: Atmospheric Chemistry andPhysics. VOCALS-REx Special Issue. 11. pp. 2015–2029.
Gattuso, J.P., Magnan, A., Billé, R., Cheung, W.W.L., Howes, E.L., Joos, F., Hoegh-Guldberg, O., 2015. Contrasting futures for ocean and society from different an-thropogenic CO2 emissions scenarios. Science 349 (6243), aac4722.
Gazeau, F., Parker, L.M., Comeau, S., Gattuso, J.P., O'Connor, W.A., Martin, S., Ross, P.M.,2013. Impacts of ocean acidification on marine shelled molluscs. Mar. Biol. 160 (8),2207–2245.
Gillooly, J.F., Brown, J.H., West, G.B., Savage, V.M., Charnov, E.L., 2001. Effects of sizeand temperature on metabolic rate. Science 293 (5538), 2248–2251.
González, M., López, D., Pérez, M., Castro, J.M., 2002. Effect of temperature on the scopefor growth in juvenile scallops Argopecten purpuratus (Lamark, 1819). Aquac. Int. 10,339–349.
Gooding, R.A., Harley, C.D.G., Tang, E., 2009. Elevated water temperature and carbondioxide concentration increase the growth of a keystone echinoderm. Proc. Natl.Acad. Sci. U. S. A. 106, 9316–9321.
Gruber, N., 2011. Warming up, turning sour, losing breath: ocean biogeochemistry underglobal change. Philos. Transact. A Math. Phys. Eng. Sci. 369, 1980–1996.
Gruber, N., Hauri, C., Lachkar, Z., Loher, D., Fröhlicher, T.L., Plattner, G.K., 2012. Rapidprogression of ocean acidification in the California current system. Science 337,220–223.
Hendriks, I.E., Duarte, C.M., Olsen, Y.S., Steckbauer, A., Ramajo, L., Moore, T.S., ...McCulloch, M., 2015. Biological mechanisms supporting adaptation to ocean acid-ification in coastal ecosystems. Estuar. Coast. Shelf Sci. 152, A1–A8.
Henry, M., Boucaud-Camou, E., Lefort, Y., 1991. Functional micro-anatomy of the di-gestive gland of the scallop Pecten maximus (L.). Aquat. Living Resour. 4, 191–202.
Hofmann, G.E., Smith, J.E., Johnson, K.S., Send, U., Levin, L.A., Micheli, F., Paytan, A.,Price, N.N., Peterson, B., Takeshita, Y., Matson, P.G., Crook, E.D., Kroeker, K.J.,Gambi, M.C., Rivest, E.B., Frieder, C.A., Yu, P.C., Martz, T.R., 2011. High-frequencydynamics of ocean pH: a multi-ecosystem comparison. PLoS One 6 (12), e28983.
Kroeker, K.J., Kordas, R.L., Crim, R., Hendriks, I.E., Ramajo, L., Singh, G.S., ... Gattuso,J.P., 2013. Impacts of ocean acidification on marine organisms: quantifying sensi-tivities and interaction with warming. Glob. Chang. Biol. 19 (6), 1884–1896.
Kroeker, K.J., Gaylord, B., Hill, T.M., Hosfelt, J.D., Miller, S.H., Sanford, E., 2014. Therole of temperature in determining Species' vulnerability to ocean acidification: acase study using Mytilus galloprovincialis. PLoS One 9 (7), e100353.
Kudela, R., Pitcher, G., Probyn, T., Figueiras, F., Moita, T., Trainer, V., 2005. Harmfulalgal blooms in coastal upwelling systems. Oceanography 18, 184–197.
Labarta, U., Fernández-Reiríz, M.J., Babarro, J.M.F., 1997. Differences in physiologicalenergetics between intertidal and raft cultivated mussels Mytilus galloprovincialis.Mar. Ecol. Prog. Ser. 152, 167–173.
Lagos, N., Castilla, J.C., Broitman, B.R., 2008. Spatial environmental correlates of inter-tidal recruitment: a test using barnacles in the northern Chile. Ecol. Monogr. 78 (2),245–261.
Lagos, N.A., Benítez, S., Duarte, C., Lardies, M.A., Broitman, B.R., Tapia, C., Tapia, P.,Widdicombe, S., Vargas, C.A., 2016. Effects of temperature and ocean acidification onshell characteristics of Argopecten purpuratus: implications for scallop aquaculture inan upwelling-influenced area. Aquac. Environ. Interact. 8, 357–370.
Lardies, M.A., Arias, M.B., Poupin, M.J., Manríquez, P.H., Torres, R., Vargas, C.A.,Navarro, J.M., Lagos, N.A., 2014. Differential response to ocean acidification inphysiological traits of Concholepas concholepas populations. J. Sea Res. 90, 127–134.
Legall, G., Chagot, D., Mialhe, E., Grizel, H., 1988. Branchial rickettsiales-like infectionassociated whit a mass mortality of sea scallop Pecten maximus. Dis. Aquat. Org. 4,229–232.
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
464

Lohrmann, K.B., 2009. How healthy are cultivated scallops (Argopecten purpuratus) fromChile? A histopathological survey. Rev. Biol. Mar. Oceanogr. 44, 35–47.
Mackenzie, C.L., Lynch, S.A., Culloty, S.C., Malham, S.K., 2014. Future oceanic warmingand acidification alter immune response and disease status in a commercial shellfishspecies, Mytilus edulis L. PLoS One 9 (6), e99712.
Manríquez, P.H., Jara, M.E., Mardones, M.L., Navarro, J.M., Torres, R., Lardies, M.A.,Vargas, C.A., Duarte, C., Widdicombe, S., Salisbury, J., Lagos, N.A., 2013. Oceanacidification disrupts prey responses to predator cues but not net prey shell growth inConcholepas concholepas (loco). PLoS One 8 (7), e68643.
Marin, V., Huntley, M.E., Frost, B., 1986. Measuring feeding rate of pelagic herbivores:analysis of experimental design and methods. Mar. Biol. 93 (1), 49–58.
Martínez, G., Pérez, H., 2003. Effect of different temperature regimes on reproductiveconditioning in the scallop Argopecten purpuratus. Aquaculture 228, 153–167.
Martínez, G., Caceres, L.A., Uribe, E., Díaz, M.A., 1995. Effects of different feeding re-gimens on larval growth and the energy budget of juvenile Chilean scallops,Argopecten purpuratus Lamarck. Aquaculture 132 (3–4), 313–323.
Martínez, G., Brokordt, K., Aguilera, C., Soto, V., Guderley, H., 2000. Effect of diet andtemperatura upon muscle metabolic capacities and biochemical composition ofgonad and muscle in Argopecten purpuratus Lamarck, 1819. J. Exp. Biol. Ecol. 247,29–49.
Mathers, N.F., 1976. Effects of tidal currents on rhythm of feeding and digestion inpecten-maximus l. J. Exp. Mar. Biol. Ecol. 24, 271–283.
Mathers, N.F., Smith, T., Colins, N., 1979. Monophasic and diphasic digestive cycles inVenerupis decussata and Chlamys varia. J. Molluscan Stud. 45, 68–81.
Mehrbach, C., Culberson, C.H., Hawley, J.E., Pytkowicz, R.N., 1973. Measurement of theapparent dissociation constants of carbonic acid in seawater at atmospheric pressure.Limnol. Oceanogr. 18, 897–907.
Morse, J.W., Arvidson, R.S., Lüttge, A., 2007. Calcium carbonate formation and dis-solution. Chem. Rev. 107, 342–381.
Navarro, J.M., González, C.M., 1998. Physiological responses of the Chilean scallopArgopecten purpuratus to decreasing salinities. Aquaculture 167, 315–327.
Navarro, J.M., Jaramillo, R., 1994. Evaluación de la oferta alimentaria natural disponiblea organismos filtradores en la Bahía de Yaldad (43–08; 73–44), sur de Chile. Rev.Biol. Mar. 29, 57–75.
Navarro, J.M., Leiva, G.E., Martínez, G., Aguilera, C., 2000. Interactive effects of diet andtemperature on the scope for growth of the scallops Argopecten purpuratus duringreproductive conditioning. J. Exp. Mar. Biol. Ecol. 247, 67–83.
Navarro, J.M., Torres, R., Acuña, K., Duarte, C., Manríquez, P.H., Lardies, M., Lagos, N.A.,Vargas, C., Aguilera, V., 2013. Impact of medium-term exposure to elevated pCO2levels on the physiological energetics of the musselMytilus chilensis. Chemosphere 90,1242–1248.
Navarro, J.M., Duarte, C., Manríquez, P.H., Lardies, M.A., Torres, R., Acuña, K., Vargas,C.A., Lagos, N.A., 2016. Ocean warming and elevated carbon dioxide: multiplestressor impacts on juvenile mussels from southern Chile. ICES J. Mar. Sci. 73,764–771.
Norton, J.H., Shepherd, M.A., Abdonnaguit, M.R., Lindsay, S., 1993. Mortalities in thegiant clam Hippopus hippopus associated with rickettsiales-like organisms. J.Invertebr. Pathol. 62, 207–209.
Orr, J.C., Fabry, V.J., Aumont, O., Bopp, L., Doney, S.C., Feely, R.A., Key, R.M., 2005.Anthropogenic ocean acidification over the twenty-first century and its impact oncalcifying organisms. Nature 437 (7059), 681–686.
Palmer, A.R., 1982. Growth in marine gastropods: a non-destructive technique for in-dependently measuring shell and body weight. Malacologia 23, 63–73.
Pérez, E.P., Azócar, C., Araya, A., Astudillo, O., Ramos, M., 2012. Comparación del cre-cimiento de Argopecten purpuratus entre cohortes obtenidas de captación de larvas enambiente natural y de hatchery. Lat. Am. J. Aquat. Res. 40 (4), 1026–1038.
Perry, R.I., Ommer, R.E., Barange, M., Werner, F., 2010. The challenge of adaptingmarine social–ecological systems to the additional stress of climate change. Curr.Opin. Environ. Sustain. 2, 356–363.
Pierrot, D., Lewis, E., Wallace, D.W.R., 2006. MS Excel Program Developed for CO2System Calculations, ORNL/CDIAC- 105. Carbon Dioxide Information AnalysisCenter. Oak Ridge National Laboratory, US Department of Energy, Oak Ridge, TN.
Pigliucci, M., 2005. Evolution of phenotypic plasticity: where are we going now? TrendsEcol. Evol. 20, 481–486.
Queirós, A.M., Fernandes, J.A., Faulwetter, S., Nunes, J., Rastrick, S.P.S., Mieszkowska,N., Artioli, Y., Yool, A., Calosi, P., Arvanitidis, C., Findlay, H.S., Barange, M., Cheung,W.W.L., Widdicombe, S., 2015. Scaling up experimental ocean acidification andwarming research: from individuals to the ecosystem. Glob. Chang. Biol. 21 (1),130–143.
Ramajo, L., Marba, N., Prado, L., Peron, S., Lardies, M.A., Rodriguez-Navarro, A.B.,Vargas, C.A., Lagos, N.A., Duarte, C.M., 2016a. Biomineralization changes with foodsupply confer juvenile scallops (Argopecten purpuratus) resistance to ocean acidifica-tion. Glob. Chang. Biol. 22, 2025–2037.
Ramajo, L., Prado, L., Rodriguez-Navarro, A.B., Lardies, M.A., Duarte, C.M., Lagos, N.A.,2016b. Plasticity and trade-offs in physiological traits of intertidal mussels subjectedto freshwater-induced environmental variation. Mar. Ecol. Prog. Ser. 553, 93–109.
Rutllant, J., 1993. Coastal Lows and Associated Southerly Winds Events in North-CentralChile. Proceeding Confer. Southern Hemisphere Meteorology and Oceanography,Australia (2 pp).
Sanders, M.B., Bean, T.P., Hutchinson, T.H., Le Quesne, W.J., 2013. Juvenile king scallop,Pecten maximus, is potentially tolerant to low levels of ocean acidification when foodis unrestricted. PLoS One 8 (9), e74118.
Sastry, A.N., 1968. The relationships among food, temperature, and gonad developmentof the bay scallop Aequipecten irradians Lamarck. Physiol. Zool. 41, 44–53.
Scanes, E., Parker, L.M., O'Connor, W.A., Ross, P.M., 2014. Mixed effects of elevated pCO2
on fertilisation, larval and juvenile development and adult responses in the mobile
subtidal scallop Mimachlamys asperrima (Lamarck, 1819). PLoS One 9 (4), e93649.Schalkhausser, B., Bock, C., Stemmer, K., Brey, T., Pörtner, H.O., Lannig, G., 2013. Impact
of ocean acidification on escape performance of the king scallop, Pecten maximus,from Norway. Mar. Biol. 160, 1995–2006.
Schlichting, C.D., Pigliucci, M., 1998. Phenotypic Evolution: A Reaction NormPerspective. Sinauer Associates, Sunderland, MA.
SERNAPESCA (Servicio Nacional de Pesca y Acuicultura), 2013. Anuarios estadísticos delServicio Nacional de Pesca y Acuicultura. www.sernapesca.cl.
Sicard, M.T., Maeda-Martinez, A.N., Omart, P., Reynoso-Granados, T., Carvalho, L., 1999.Optimum temperature for growth in the catarina scallop (Argopecten ventricosus cir-cularis, Sowerby II, 1842). J. Shellfish Res. 18, 385–392.
Sokal, R.R., Rohlf, F.J., 1995. Biometry: The Principles and Practice of Statistics inBiological Research. Freeman, W.H., New York.
Somero, G.N., 2010. The physiology of climate change: how potentials for acclimatizationand genetic adaptation will determine “winners” and “losers”. J. Exp. Biol. 213,912–920.
Soria, G., Merino, G., von Brand, E., 2007. Effect of increasing salinity on physiologicalresponse in juvenile scallops Argopecten purpuratus at two rearing temperatures.Aquaculture 270, 451–463.
Soria, G., Merino, G., Uribe, E., von Brand, E., 2011. Effect of increasing salinity onweight-specific filtration rate of juvenile scallop Argopecten purpuratus reared at twotemperatures: is any effect related to ammonia buildup? J. Shellfish Res. 30 (2),279–286.
Strickland, J.D.H., Parsons, T.H., 1968. A Practical Handbook of Seawater Analysis,Fisheries Research Board of Canada, Bulletin 167. Ottawa, 293 pp.
Strub, P.T., Mesías, J.M., Montecino, V., Rutllant, J., Salinas, S., 1998. Coastal oceancirculation off western South America. In: Robinson, A.R., Brink, K.H. (Eds.), The Sea.John Wiley and Sons, New York, pp. 273–314.
Syasina, I.G., Vaschenko, M.A., Zhadan, P.M., 1997. Morphological alterations in thedigestive diverticula of Mizuhopecten yessoensis (Bivalvia: Pectinidae) from pollutedareas of Peter the Great Bay, Sea of Japan. Mar. Environ. Res. 44, 85–98.
Sydeman, W.J., García-Reyes, M., Schoeman, D.S., Rykaczewski, R.R., Thompson, S.A.,Black, B.A., Bograd, S.J., 2014. Climate change and wind intensification in coastalupwelling ecosystems. Science 345, 77–80.
Talmage, S., Gobler, C., 2009. The effects of elevated carbon dioxide concentrations onthe metamorphosis, size, and survival of larval hard clams (Mercenaria mercenaria),bay scallops (Argopecten irradians), and eastern oysters (Crassostrea virginica). Limnol.Oceanogr. 54, 2072–2080.
Thébault, J., Thouzeau, G., Chauvaud, L., Cantillánez, M., Avendaño, M., 2008. Growth ofArgopecten purpuratus (Mollusca: Bivalvia) on natural bank in northern Chile:sclerochronological record and environmental controls. Aquat. Living Resour. 21,44–45.
Thiel, M., Macaya, E.C., Acuña, E., Arntz, W.E., et al., 2007. The Humboldt current systemof northern and central Chile. Oceanographic processes, ecological interactions andsocioeconomic feedback. Oceanogr. Mar. Biol. Annu. Rev. 45, 195–344.
Thomsen, J., Casties, I., Pansch, C., Kortzinger, A., Melzner, F., 2013. Food availabilityoutweighs ocean acidification effects in juvenile Mytilus edulis: laboratory and fieldexperiments. Glob. Chang. Biol. 19, 1017–1027.
Torres, R., Ampuero, P., 2009. Strong CO2 outgassing from high nutrient low chlorophyllcoastal waters off central Chile (30° S): the role of dissolved iron. Estuar. Coast. ShelfSci. 83, 126–132.
Torres, R., Pantoja, S., Harada, N., González, H.E., Daneri, G., Frangopulos, M., Fukasawa,M., 2011. Air-sea CO2 fluxes along the coast of Chile: From CO2 outgassing in centralnorthern upwelling waters to CO2 uptake in southern Patagonian fjords. J. Geophys.Res. Oceans Atmos. 116 (C9).
Torres, R., Manriquez, P.H., Duarte, C., Navarro, J.M., Lagos, N.A., Vargas, C.A., Lardies,M.A., 2013. Evaluation of a semi-automatic system for long-term seawater carbo-natechemistry manipulation. Rev. Chil. Hist. Nat. 86, 443–451.
Underwood, A.J., 1997. Experiments in Ecology: Their Logical Design and InterpretationUsing Analysis of Variance. Cambridge University Press, Cambridge.
Uriarte, I., Farías, A., Hernández, J., Schäfer, C., Sorgeloos, P., 2004. Reproductive con-ditioning of Chilean scallop (Argopecten purpuratus) and the Pacific oyster (Crassostreagigas): effects of enriched diets. Aquaculture 230, 349–357.
Uribe, E., Blanco, J.L., 2001. Capacidad de los sistemas acuáticos para el sostenimiento delcultivo de pectínidos: El caso de Argopecten purpuratus en la Bahía de Tongoy, Chile.Los Moluscos Pectínidos de Iberoamérica: Ciencia y Acuicultura A. N. Maeda-Martínez (ed.) 2001. Cap. 12, 000.
Vargas, C.A., de la Hoz, M., Aguilera, V., San Martín, V., Manríquez, P.H., Navarro, J.M.,Torres, R., Lardies, N.A., Lagos, N.A., 2013. CO2-driven ocean acidification reduceslarval feeding efficiency and changes food selectivity in the mollusk Concholepasconcholepas. J. Plankton Res. 33 (5), 1059–1068.
Vargas, C.A., Aguilera, V., Martín, V., Manríquez, P., Navarro, J., Duarte, C., Torres, R.,Lardies, M., Lagos, N., 2015. CO2-driven ocean acidification disrupts the filter feedingbehavior in Chilean gastropod and bivalve species from different geographic lo-calities. Estuar. Coasts 38, 1163–1177.
Vargas, C.A., Lagos, N.A., Lardies, M.A., Duarte, C., Manríquez, P.H., Aguilera, V.H.,Broitman, B., Widdicombe, S., Dupont, S., 2017. Species specific responses to oceanacidification should account for local adaptation and adaptive plasticity. Nature Ecol.Evol. 1, 00.
Via, S., Lande, R., 1985. Genotype-environment interaction and the evolution of pheno-typic plasticity. Evolution 505–522.
Via, S., Gomulkiewicz, R., De Jong, G., Scheiner, S.M., Schlichting, C.D., Van Tienderen,P.H., 1995. Adaptive phenotypic plasticity: consensus and controversy. Trends Ecol.Evol. 10 (5), 212–217.
Villalba, A., Carballal, M.J., Lopez, C., Cabada, A., Corral, L., Azevedo, C., 1999.Branchial rickettsia-like infection associated with clam Venerupis rhomboides
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
465

mortality. Dis. Aquat. Org. 36, 53–60.Von Buddenbrock, W., 1915. Die statocysten von Pecten, ihre histologie und physiologie.
Zool Jb.(abt. allg. Zool. Physiol.) 35, 301–356.Wenguang, L., Maoxian, H., 2012. Effects of ocean acidification on the metabolic rates of
three species of bivalve from southern coast of China. Chin. J. Oceanol. Limnol. 30(2), 206–211.
White, M.M., McCorkle, D.C., Mullineaux, L.S., Cohen, A.L., 2013. Early exposure of bayscallops (Argopecten irradians) to high CO2 causes a decrease in larval shell growth.PLoS One 8 (4), e61065.
Willmer, P., Stone, G., Johnston, I., 2005. Environmental Physiology of Animals, 2nd edn.
Blackwell, Carlton.Wolff, M., 1998. Spawning and recruitment in the Peruvian scallop Argopecten
Purpuratus. Mar. Ecol. Prog. Ser. 42 (1998), 213–217.Wood, H.L., Spicer, J.I., Widdicombe, S., 2008. Ocean acidification may increase calci-
fication rates, but at a cost. Proc. R. Soc. Lond. B Biol. Sci. 275, 1767–1773.Woods, H.A., Makino, W., Cotner, J.B., Hobbie, S.E., Harrison, J.F., Acharya, K., Elser,
J.J., 2003. Temperature and the chemical composition of poikilothermic organisms.Funct. Ecol. 17 (2), 237–245.
Zar, J.H., 1999. Biostatistical Analysis, 4th ed. Prentice Hall, Upper Saddle River.
M.A. Lardies et al. Aquaculture 479 (2017) 455–466
466





























