Embed Size (px)
Citation preview

ration 92 (2007) 43–54www.elsevier.com/locate/jgeoexp
Journal of Geochemical Explo
Arsenic remobilization from sediments contaminated with minetailings near the Adak mine in Västerbotten district
(northern Sweden)
Joyanto Routh a,⁎, Aparajita Bhattacharya a, Ambujom Saraswathy a,Gunnar Jacks b, Prosun Bhattacharya b
a Department of Geology and Geochemistry, Stockholm University, SE-10691 Stockholm, Swedenb Department of Land and Water Resources Engineering, KTH, SE-10044 Stockholm, Sweden
Received 17 January 2006; accepted 6 July 2006Available online 22 August 2006
Abstract
Weathering of mine tailings have resulted in high As concentrations in water (up to 2900 μg l−1) and sediment (up to 900 mgkg−1) samples around the Adak mine. Notably, As occurs as As(III) species (15–85%) in the oxic surface and ground watersamples, which is not common. Time-series based sediment incubations were set up in the laboratory with contaminated sedimentsto study the microbial processes involved in transformation and remobilization of As across the sediment–water interface. Themicrocosm experiments indicate that microorganisms are capable of surviving in As-rich sediments and reduce As(V) to As(III). Adecrease in total As concentration in sediments is coupled to an increase in As(III) concentration in the aqueous media. In contrast,the controls (treated with HgCl2 and formaldehyde) did not show growth, and As(V) concentrations increased steadily in thesediments and aqueous medium. The results imply that active metabolism is necessary for As(V) reduction. These microorganismspossess reduction mechanisms that are not necessarily coupled to respiration, but most likely impart resistance to As toxicity.© 2006 Elsevier B.V. All rights reserved.
Keywords: Arsenic; Microbes; Microcosm; Reduction; Speciation; Mobilization
1. Introduction
Arsenic (As) ranks 20th in abundance on the Earth'scrust and is commonly associated with chalcophilicelements (e.g., Cu, Pb, and Au). Arsenic exists in naturein four oxidation states: As(III) arsenite under reducingconditions, As(V) arsenate under oxidizing conditions,(As3−) arsine as traces in gases emanating from anoxicenvironments, and (As0) elemental As (Cullen andReimer, 1989). Arsenic toxicity, mobility, and bioavail-
⁎ Corresponding author. Tel.: +4686747824; fax: +4686747855.E-mail address: [email protected] (J. Routh).
0375-6742/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.gexplo.2006.07.003
ability vary depending on the oxidation state. As(III)exhibits greater hydrological mobility in the environ-ment and is 25–60 times more toxic than As(V) (Korteand Fernando, 1991). This difference in mobility resultsfrom the high affinity of As(V) for hydrous ferric,aluminum, and manganese oxides. Arsenic compoundspose a serious health hazard because of increased oxi-dative stress, and disruption of oxidative phosphoryla-tion in living tissues (Oremland and Stolz, 2003).
Microbial processes involved in As cycling includeoxidation, reduction, and methylation reactions (forreview see Oremland and Stolz, 2003). Microbial pro-cesses are many times faster than chemical reactions

44 J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
(Sohrin et al., 1997; Jones et al., 2000), and play animportant role in transforming and mobilizing As(Ahmann et al., 1997; Macur et al., 2001; Routh et al.,2005). Direct reduction of As(V) as a process of inter-cellular detoxification by using the ars operon has beenwidely documented in several species of bacteria, algae,and fungi under aerobic and anaerobic conditions (e.g.,Cai et al., 1998; Macur et al., 2001; Mukhopadhyayet al., 2002; Macur et al., 2004). The ars operon lowersthe intracellular As concentration, and permits microbialsurvival in high As environments. Microbial reductionof As(V) also occurs under anoxic conditions due todissimilatory reduction, whereby As(V) is used as theterminal electron acceptor by anaerobic microorganisms(Ahmann et al., 1997; Newman et al., 1998). Thesemicroorganisms gain energy by coupling As(V) reduc-tion with organic matter oxidation.
It is estimated that mining and ore processing innorthern Sweden have resulted in N600,000 tons of As inmine tailings exposed to weathering (Jacks et al., 2003).Several As contaminated sites occur in the region, andthese sites pose a serious environmental challenge forclean up and remediation (Carlsson et al., 2003; Jacks etal., 2003). The present study, builds on an environmentalassessment of the Adak mine site in Västerbotten districtby Bhattacharya et al. (2006). In Adak, water, sediment,and soil samples are contaminated with mine tailings,and trace metal concentrations were high, particularlyAs. In addition, the results of As speciation in this studywas rather unusual. Arsenic primarily occurred as As(III)species (15–85%) in the oxic surface and ground watersamples. Few other studies have also reported of similarobservations in natural systems (e.g., Kuhn and Sigg,1993; Aurillo et al., 1994); the authors related Asspeciation to microbial transformation of As(V). Thesituation is far from thermodynamic equilibrium, andfurther investigations are needed to develop a betterunderstanding and predictability of As transformation innature.
The Adak mine site provides an opportunity to studythe unusual phenomena associated with speciation andsubsequent remobilization of As from sediments due toin situ microbial processes. Our investigations arebased on field measurements and microcosm experi-ments conducted in sediments contaminated with minetailings. Surprisingly very few studies have investigatedthe biogeochemical processes associated with As cyclingaroundmine sites even though As is a major contaminantin sulfidic tailings. The present study provides insightsinto such processes, and several aspects of this studycould be extended to other areas that have experiencedanalogous problems with As contamination. We believe
that these results could influence surface and ground-water treatment plans to reduce the high As concentra-tions in several mining towns in the Västerbotten district.Finally, to the best of our knowledge, this is the firstreport on microbial transformation and remobilization ofAs from mine tailings in northern Sweden.
2. Materials and methods
2.1. Study area
The Västerbotten district of northern Sweden hasbeen extensively mined for Au, Ag, Cu, Zn, and Pbsince the early 20th century. The Adak mine is a Cu–Zn–Pb sulfide deposit intercalated with mafic volcanicsand redeposited volcano-clastic rocks (Willden, 1986).The massive sulfide ore bodies are composed of pyrite,arsenopyrite, chalcopyrite, sphalerite, and pyrrhotite(Wagner et al., 2004). Similar volcanic hosted massivesulfide deposits of the Kuroko-type ores occur world-wide e.g., British Columbia (Canada), Ural Mountains(Russia), the Iberian Peninsula (Spain), and Australia(Vivallo, 1987). The enriched Adak and Brännmyran(Cu 1.7–4.5%, As 1%) and Rudtjebäcken (Cu 0.8%, Zn3.0%) deposits were discovered in the 1930s, and min-ing began around 1945. Mineralization around Adakoccurs in mica and graphite schists, dacites, and mafictuff (Frietsch et al., 1995). During mining from 1945to 1975, over 6.3 million tons of ores were excavatedfrom these mines, which included 123,000 tons of Cu,100,000 tons of Zn, 42 tons of Ag, and 1.4 tons of Au.
An ore-processing unit located near the mine receivedraw material to extract the sulfidic minerals of Cu, Zn,Au, and Pb. Archival records from the area indicate that anearby peat bog was initially used for the disposal ofmilled tailings. Later, the tailings were discarded into theeastern part of Lake Ruttjejaure, before the mining com-pany began storing them at the present location. Wesuspect that the change may have to do with stricterregulations for disposal of tailings, or deteriorating waterquality in the lake.
Once mining slowed down around 1975, the inflowof tailings decreased. Before the site was finally aban-doned, the metal-rich tailings extending over1500 m×1000 m×5 m were covered with a 1–2 mthick layer of glacial till to prevent weathering. Theactual aerial extent of the tailings has however increasedbeyond the original limits of the tailings pile due toweathering (Fig. 1). Presently, weathered tailings extendclose to the shores of Lake Ruttjejaure situated about700 m away. Two shallow streams run adjacent to thetailings impoundment and drain into Lake Ruttjejaure.
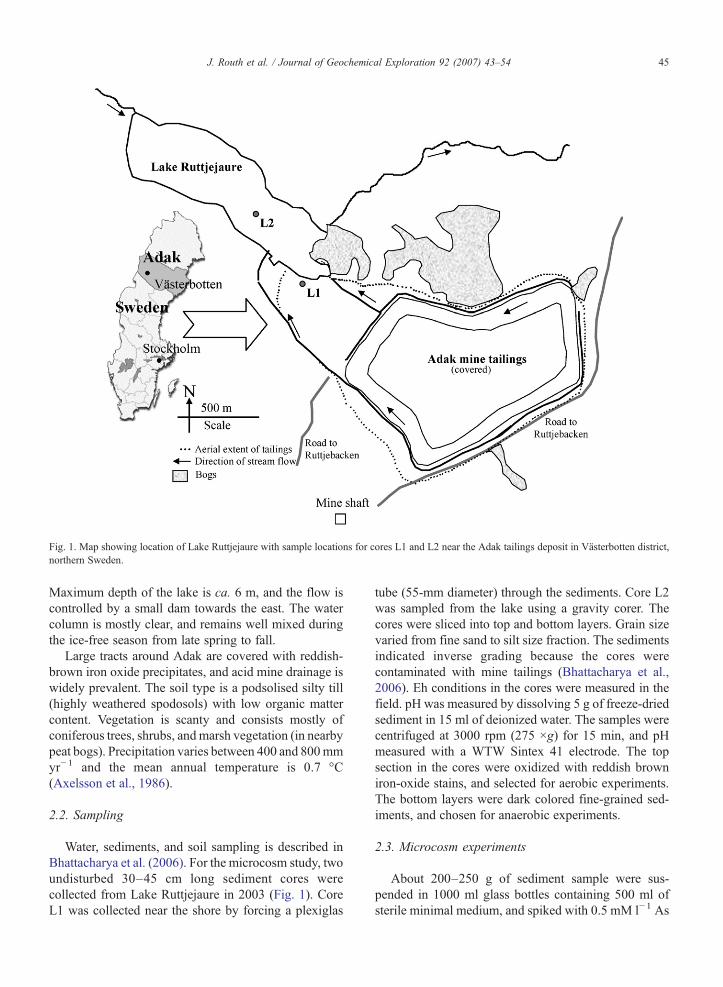
Fig. 1. Map showing location of Lake Ruttjejaure with sample locations for cores L1 and L2 near the Adak tailings deposit in Västerbotten district,northern Sweden.
45J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
Maximum depth of the lake is ca. 6 m, and the flow iscontrolled by a small dam towards the east. The watercolumn is mostly clear, and remains well mixed duringthe ice-free season from late spring to fall.
Large tracts around Adak are covered with reddish-brown iron oxide precipitates, and acid mine drainage iswidely prevalent. The soil type is a podsolised silty till(highly weathered spodosols) with low organic mattercontent. Vegetation is scanty and consists mostly ofconiferous trees, shrubs, andmarsh vegetation (in nearbypeat bogs). Precipitation varies between 400 and 800mmyr−1 and the mean annual temperature is 0.7 °C(Axelsson et al., 1986).
2.2. Sampling
Water, sediments, and soil sampling is described inBhattacharya et al. (2006). For the microcosm study, twoundisturbed 30–45 cm long sediment cores werecollected from Lake Ruttjejaure in 2003 (Fig. 1). CoreL1 was collected near the shore by forcing a plexiglas
tube (55-mm diameter) through the sediments. Core L2was sampled from the lake using a gravity corer. Thecores were sliced into top and bottom layers. Grain sizevaried from fine sand to silt size fraction. The sedimentsindicated inverse grading because the cores werecontaminated with mine tailings (Bhattacharya et al.,2006). Eh conditions in the cores were measured in thefield. pH was measured by dissolving 5 g of freeze-driedsediment in 15 ml of deionized water. The samples werecentrifuged at 3000 rpm (275 ×g) for 15 min, and pHmeasured with a WTW Sintex 41 electrode. The topsection in the cores were oxidized with reddish browniron-oxide stains, and selected for aerobic experiments.The bottom layers were dark colored fine-grained sed-iments, and chosen for anaerobic experiments.
2.3. Microcosm experiments
About 200–250 g of sediment sample were sus-pended in 1000 ml glass bottles containing 500 ml ofsterile minimal medium, and spiked with 0.5 mM l−1 As

46 J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
(As2O5). Lactate (0.9 mM l−1) was added as the solecarbon source for heterotrophic microorganisms. Theminimal medium was prepared after modifying themethod of Turpeinen et al. (1999). Three solutions weremixed: 1) solution 1 containing 1.4 g of KH2PO4, 2.5 gof NH4Cl, 5 g of KCl, 1.5 g of CaCl2·2H2O, 5 g of NaCl,and 6.2 g of MgCl2 in 1000 ml of H2O, 2) solution 2containing 0.5mg of para-amino benzoic acid in 1000mlof H2O, and 3) solution 3 containing mixture of 1 mgMnCl2·4H2O, 1.2 mg of Co(NO3)2·6H2O, 0.7 mg ofZnCl2, 0.6 mg of H3BO3, 0.25 mg of Ni(NO3)2·6H2O,0.15 mg of CuCl2·2H2O, 0.25 mg of Na2MoO4·2H2O,and 15 mg of FeCl2 in 1000 ml of H2O. All chemicalsused for this study were reagent grade or better, and wereused, without further purification.
The aerobic samples were incubated on a rotaryshaker (175 rpm) at room temperature. The anaerobicsamples were sealed with butyl rubber-stoppers. Nitro-gen was bubbled through the medium for 30 min and thesamples were incubated in darkness in a N2-flow throughglove box at 22 °C. The controls were treated with7.4 mM l−1 HgCl2 (25 wt.% vol−1 of sediment) and0.04% of formaldehyde prior to incubation. Additionalcontrols were set up without lactate in the minimalmedium. Initially growth was inferred in the serumbottles by visually inspecting increase in turbiditycompared to the controls. Subsequently, the microcosmswere sampled four times over 28 days, and 75 ml of eachsample was sacrificed periodically. In the anaerobicchambers, nitrogen was flushed through the system for30 min after each sampling event. Fresh Anaerocult Astrips (Merck EuroLab AB, Sweden) were placed in thebottles, and change in color was monitored to see if thesystem was leaking. Orion electrodes were used tomeasure Eh (Model 91-03AP), pH (Model 96-78), andoxygen (Model 97-08) in the incubation bottles duringsampling. The sediment slurries extracted under asepticconditions through the rubber septa were first centri-fuged at 3000 rpm (907 ×g). The aqueous and sedimentphases obtained were separately analyzed for As(III) andAs(V) species, and compared to the heterotrophic platecounts of the corresponding day.
The heterotrophic plate counts were performed byserially diluting the samples using a phosphate bufferedsaline solution (8 g of NaCl, 1.21 g of K2HPO4, and 0.3 gof KH2PO4 in 1000 ml of H2O at pH=7.2). Aliquots(0.1 ml) of appropriate dilutions were plated in triplicateon tryptic soy agar (TSA) medium spiked with0.435 mM l−1 As. The TSA plates are suitable forgrowing fastidious microbes including anaerobes andfacultativemicrobes. The plates for aerobic cultures wereincubated for 7 days at room temperature. The plates for
anaerobic cultures were placed in bags with AnaerocultA strips and incubated in darkness for 7 days at roomtemperature.
2.4. Analytical methods
2.4.1. Water and sediment samplesArsenic speciation in water was performed in the field
using disposable cartridges (Meng and Wang, 1998). As(V) was obtained as a difference between total As (Astot)and As(III) species. Sediment samples were dried in N2-filled glove bags to prevent oxidation. About 0.5 g ofsample was weighed and extracted with 7 M HNO3 at130 °C for 1 h in an autoclave (Swedish StandardsInstitute, 1993). In a limited number of samples oxalicacid extraction was done. 1 g of dry sediment wasextracted with 100 ml of oxalic acid and shaken for 4 h indark. The samples were later centrifuged (4000 ×g for15 min) and filtered (0.2 μm polycarbonate filters).Major and trace elements in the water and sedimentsample extracts were analyzed on a Varian Vista Pro AxICP-OES. The detection limits were based on three timesthe standard deviation of a reagent blank that wasanalyzed ten times. For most trace elements, the detec-tion limit was from 2 to 5 μg l−1. An external standard(SRM 1640) was run in between the samples, and stan-dard deviation was b10% for the trace elements of in-terest. An internal standard (a marine sediment) was runin between the samples (treated the same way as thesamples), and the relative error was b1% for the ana-lyses, compared to previous sample runs.
2.4.2. Speciation studies in microcosm phasesArsenic speciation in sediments was adapted after
modifying the method of Chappell et al. (1995). 5 g ofdry sediment was digested with 20 ml of 10 M HCl. Thesamples were shaken for 30 min and centrifuged at3000 rpm (907 ×g) for 5 min. The supernatant wasgravity filtered (Whatman® 44 filter paper). The processwas repeated twice and the extract volume was made upto 100 ml. 10 ml of this acid extract was treated with80ml of 10MHCl followed by extraction of As(III) withCHCl3 (4×10 ml aliquots). Arsenic was back-extractedfrom the organic fraction into aqueous phase (2×20 mlaliquots), and the extract diluted to 100 ml. The stepinvolving CHCl3 extraction was repeated four times, andback-extraction with water was repeated two times toobtain themaximum amount of As(III). Another 10ml ofthe acid extract was treated with 10 ml of 50% KIsolution, and heated in a water bath at 60 °C for 30 min.The sample was extracted with CHCl3 and water todetermine the total As (Astot) concentration. The extracts

47J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
were analyzed on a Varian Spectra AA graphite furnace.The As(V) values were obtained by subtracting As(III)fromAstot concentrations. The instrument detection limitfor As analyses was b2 μg l−1. The relative standarddeviation for the reference standard SRM 1640was b3%.
Arsenic species in the medium were measured aftermodifying the methods of Johnson and Pilson (1972)and Cummings et al. (1999). The aqueous phase(nutrient media) was filtered through a 0.45 μm Milli-pore filter. The sample was divided into unoxidized,oxidized, and reduced fractions by adding 100 μl of HCl,KI, or the cold reducing agent (mixture of sodium metathiosulfate, sodium meta bisulphate, and H2SO4) to1200 μl of sample. Finally, 600 μl of mixed reagent(containing H2SO4, ascorbic acid, ammonium molyb-date, and potassium antimonyl tartarate) was added tothe samples. The samples were incubated for 10 min at78 °C and cooled for 5 min at −20 °C. The blue coloredarseno–molybdenum complex was analyzed spectro-photometrically at 865 nm (Hitachi U-2001) to deter-mine the As concentration. As(III) was obtained asa difference of the unoxidized and oxidized samples, andAs(V) was obtained as the difference of the oxidized andreduced samples. The instrument detection limit for Aswas ∼ 10 μM.
3. Results and discussion
3.1. Water, sediment, and soil chemistry
We have briefly discussed the salient characteristicsof the Adak water, sediment, and soil chemistry in orderto highlight the important factors affecting the distribu-tion of trace elements. Proximity to the tailings, pH, grainsize, and distribution of Al and S affect the distribution oftrace elements in solid and aqueous phases. Moreover,the direct disposal of milled tailings into the lake, andmixing of weathered tailings with nearby sediment andsoil have contaminated the site within a 2 km (or greater)radius. Specific details regarding the distribution of traceelements in solid and aqueous phases are discussed inJacks et al. (2003) and Bhattacharya et al. (2006).
Weathering of sulfide-rich tailings at the Adak minesite results in acid mine drainage. The low pH keeps thetrace elements dissolved, and results in greater remobi-lization and spatial distribution. Thewater table in the areais shallow and occurs few meters below the surface.Metal-enriched leachates from the tailings pile contam-inate the surface and ground water supply. In fact, there islittle difference between the surface and ground waterchemistry. The water samples indicate low pH (2.7–4.1),
and high sulfate concentrations (up to 800 mg l−1;Table 1). Trace element concentrations in the watersamples are high compared to unimpacted surface andground water samples. Particularly, As concentrations arevery high, and occur significantly above the drinkingwater standard. Astot in surface water is up to 1200 μg l
−1,and in ground water the concentration is up to 2900 μgl−1. Arsenic speciation in the water samples indicates thatAs(III) is the predominant species (up to 84%).
The surface sediments in Lake Ruttjejaure are highlyoxidized, particularly the top 5–10 cm, and they arecharacterized by reddish-brown iron oxide coatings.Organic carbon content in these sediments is low (b2%).While the surface sediments are oxidized, sulfatereduction occurs in deeper sediments. The sedimentsbecome grayish in color (beyond 10 cm or so), and theFe/S ratio decreases with depth. Likewise, the totalreducible sulfide fraction increases with depth. Thesechanges suggest a switch from oxidizing conditions to amore reducing environment at depth. High concentra-tions of Fe, Mn, and S occur in the stream and lakesediments (Table 1). Amongst the different trace ele-ments analyzed in this study, concentrations of As, Cu,and Zn are high compared to background values, and theaverage crustal composition. Moreover, the analysesindicate that∼ 40% ofAs occur in the oxalate extractablefraction, implying As is mainly sorbed to Fe or Al oxidesin these sediments. This is not surprising because thesediments are oxidized, and high Al and Fe concentra-tions are prevalent (Table 1).
The soil profile (65-cm in depth and collected near thelake) indicates higher trace element concentrations, butlow Fe and S concentrations compared to the streambedand lake sediments. Trace element concentrations of Al,As, Co, Cu, Pb, and Zn decrease with depth. Arsenicconcentrations in the soil zones are high (700–900 mgkg−1) compared to streambed and lake sediments. Thereis sharp decrease in organic content (6.4–0.1%) withdepth. The total reducible sulfide fraction in the soil islow (0.1–1.8% of total S), but increases with depthimplying more reducing conditions.
3.2. Microcosm studies
3.2.1. Sediment phase characteristics in microcosmsArsenic concentrations in the sediment and aqueous
phases, and corresponding heterotrophic plate counts inthe microcosms are plotted in Figs. 2 and 3. Analyses ofsediments from Lake Ruttjejaure indicate higher con-centrations of As(V) compared to As(III) on 0 day. Thisprobably arises from the oxidized condition particularly,the top few centimeters, where iron-oxide stains are

Table1
Geochem
ical
trendin
water
andsedimentsamples
from
Adak,
Västerbottendistrict,northern
Sweden
(from
Bhattacharya
etal.,2006)
Eh(m
V)
pHCond
HCO3−
CI−
S‡
NO3−
Na
KCa
Mg
Al
As tot
%As(III)
Cu
Fe
Mn
Zn
Surface
water
146to
423
2.7–
4.1
230–
2980
bdl–0.20
0.13–1.8
1.9–
800
bdl–0.23
1.1–
220.80–48
2.2–
500
1.1–
100
0.14–9.2
0.007–
1.2
15–85
0.01–0.50
2.4–
600
0.01
–5.4
0.02
–2.2
Groundwater
138to
158
3.1–
3.9
1003–2530
0.0
0.07–1.2
15–700
0.0–
0.32
0.70–21
0.70–71
7.2–
500
1.0–
200
0.13–4.7
0.004–
2.9
42–84
0.01–1.3
0.50
–400
0.10
–8.3
0.1–
8.8
Unimpacted
surfacewater
5.6
74.3
2.0
0.6
220.0
1.2
0.5
7.1
1.6
0.4
141.3
10–
65.1
48Unimpacted
ground
water
5.0–
6.0
1.0–
130.4–
2.3
1.0–
5.9
0.1–
0.2
0.9–
5.9
0.2–
1.5
0.7–
110.2–
3.4
0.1–
0.7
––
3.7–
890.0–
1.3
10–110
30–347
Stream
sediments
2.5–
3.6
100–
5200
–31–2100
51–1700
60–2100
300–
3800
18–400
1.1–
100
2000
–42600
2.4–
621.6–
19Lakesediments
55to
−90
3.7–
7.2
100–
3300
–70–400
380–
1200
400–
3000
1100–4900
100–
800
11–700
2200
–32700
17–74
8.3–
400
Soil
3.6–
7.1
400–
1600
–200–
230
1300–4000
1300
–3500
2300
–5100
700–
900
16–400
6100
–8700
22–33
8.5–
33Unimpacted
sediments
3.9▪
2.7▪
3.7▪
2.1▪
14▪
0.9
356.6▪
0.1▪
45
Conductivity
μScm
−1 ;traceelem
entconcentrations
inwater
(mgl−1 );traceelem
entconcentrations
insediment/soil(m
gkg
−1 );▪concentrations
reported
aspercentoxide.
S‡assumed
assulfatein
water
samples;–notanalyzed.
48 J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
visible in the sediments. As the experiment progressed,Astot, As(III), and As(V) concentrations start to declinein the live samples (samples not treated with HgCl2 andformaldehyde and indicate an increase in turbidityimplying growth). Consequently, there is an overalldecrease in the Astot budget in live sediment samples(27–51% within 28 days). The decrease in Astotconcentrations in sediments with almost an equivalentor greater increase in Astot concentrations in the aqueousphase implies that As is mobilized from sediments. Thisinvolves dissolution and microbial reduction of As(V) tothe more mobile and toxic As(III) species, whichdiffuses into the water column (see discussion below).
The sediment controls indicate an overall increase inAs(V) concentrations as the experiment progressed. Thecontrols also indicate a drop in the Astot budget, but this issignificantly less (ca. 6–8%) compared to the livesamples. The steady decline in the viable microbialpopulation in the controls suggests that our treatment withHgCl2 and formaldehyde was not adequate to immedi-ately kill all the in situmicroorganisms in these sediments.More importantly, the decrease in Astot budget in thecontrols imply that abiotic processes such as chemicaldissolution or geochemical leaching (Pantsar-Kallio andManninen, 1997), and desorption from iron oxides(Manning and Goldberg, 1997) occur in these samples.These abiotic processes may result from the presence ofexchangeable ions in the medium such as phosphate,chloride, or carbonate ions (Pantsar-Kallio andManninen,1997).Nevertheless, it is difficult to distinguish accuratelythe different As fractions remobilized by abiotic versusmicrobial processes without using labeled compounds todetect microbial uptake (e.g., [73As] sodium arsenate;half-life of 80.3 days; Gao et al., 2004).
Inference of biogeochemical processes in microcosmexperiments in this study are based on sediments con-taminated with mine tailings that were collected from inand around the lake. It is possible that similar processescould be happening in the tailings pile itself. This can beproved by conducting similar experiments in samplesdirectly procured from the tailings pile. In addition, therecould be other processes active in mobilizing As from thetailings pile. For example, when the glacial till cover wasinstalled it cut-off the oxygen supply making the en-vironmentmore reducing. In this case, it is possible that As(V) is reduced toAs(III),which ismoremobile and leachesout from sediments. These abiotic reductive-dissolutionreactions could be catalyzed by in situ microorganisms.
3.2.2. Aqueous phase characteristics in microcosmsThe aqueous phase indicates higher concentrations of
As(V) over As(III) in all the samples on day 0 (Figs. 2
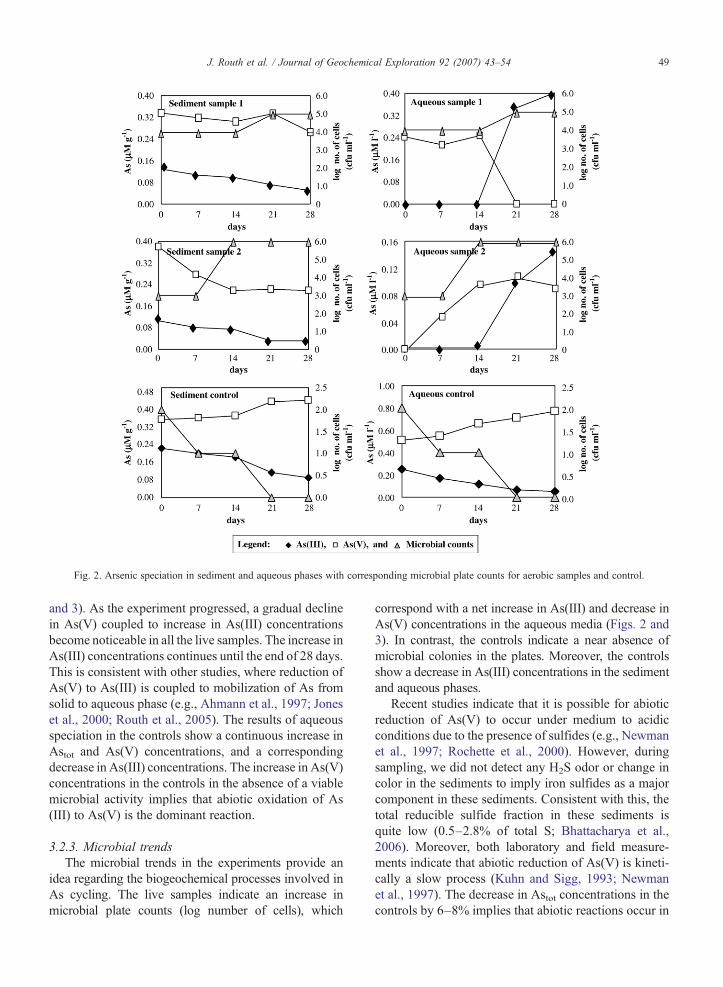
Fig. 2. Arsenic speciation in sediment and aqueous phases with corresponding microbial plate counts for aerobic samples and control.
49J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
and 3). As the experiment progressed, a gradual declinein As(V) coupled to increase in As(III) concentrationsbecome noticeable in all the live samples. The increase inAs(III) concentrations continues until the end of 28 days.This is consistent with other studies, where reduction ofAs(V) to As(III) is coupled to mobilization of As fromsolid to aqueous phase (e.g., Ahmann et al., 1997; Joneset al., 2000; Routh et al., 2005). The results of aqueousspeciation in the controls show a continuous increase inAstot and As(V) concentrations, and a correspondingdecrease in As(III) concentrations. The increase in As(V)concentrations in the controls in the absence of a viablemicrobial activity implies that abiotic oxidation of As(III) to As(V) is the dominant reaction.
3.2.3. Microbial trendsThe microbial trends in the experiments provide an
idea regarding the biogeochemical processes involved inAs cycling. The live samples indicate an increase inmicrobial plate counts (log number of cells), which
correspond with a net increase in As(III) and decrease inAs(V) concentrations in the aqueous media (Figs. 2 and3). In contrast, the controls indicate a near absence ofmicrobial colonies in the plates. Moreover, the controlsshow a decrease in As(III) concentrations in the sedimentand aqueous phases.
Recent studies indicate that it is possible for abioticreduction of As(V) to occur under medium to acidicconditions due to the presence of sulfides (e.g., Newmanet al., 1997; Rochette et al., 2000). However, duringsampling, we did not detect any H2S odor or change incolor in the sediments to imply iron sulfides as a majorcomponent in these sediments. Consistent with this, thetotal reducible sulfide fraction in these sediments isquite low (0.5–2.8% of total S; Bhattacharya et al.,2006). Moreover, both laboratory and field measure-ments indicate that abiotic reduction of As(V) is kineti-cally a slow process (Kuhn and Sigg, 1993; Newmanet al., 1997). The decrease in Astot concentrations in thecontrols by 6–8% implies that abiotic reactions occur in
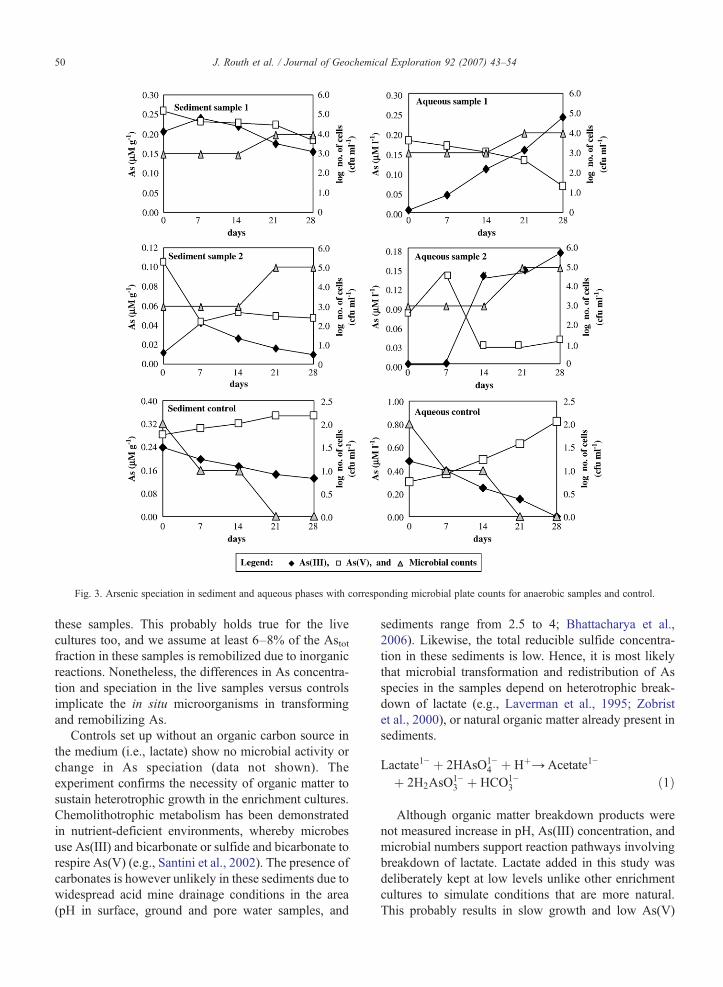
Fig. 3. Arsenic speciation in sediment and aqueous phases with corresponding microbial plate counts for anaerobic samples and control.
50 J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
these samples. This probably holds true for the livecultures too, and we assume at least 6–8% of the Astotfraction in these samples is remobilized due to inorganicreactions. Nonetheless, the differences in As concentra-tion and speciation in the live samples versus controlsimplicate the in situ microorganisms in transformingand remobilizing As.
Controls set up without an organic carbon source inthe medium (i.e., lactate) show no microbial activity orchange in As speciation (data not shown). Theexperiment confirms the necessity of organic matter tosustain heterotrophic growth in the enrichment cultures.Chemolithotrophic metabolism has been demonstratedin nutrient-deficient environments, whereby microbesuse As(III) and bicarbonate or sulfide and bicarbonate torespire As(V) (e.g., Santini et al., 2002). The presence ofcarbonates is however unlikely in these sediments due towidespread acid mine drainage conditions in the area(pH in surface, ground and pore water samples, and
sediments range from 2.5 to 4; Bhattacharya et al.,2006). Likewise, the total reducible sulfide concentra-tion in these sediments is low. Hence, it is most likelythat microbial transformation and redistribution of Asspecies in the samples depend on heterotrophic break-down of lactate (e.g., Laverman et al., 1995; Zobristet al., 2000), or natural organic matter already present insediments.
Lactate1− þ 2HAsO1−4 þ Hþ→Acetate1−
þ 2H2AsO1−3 þ HCO1−
3 ð1Þ
Although organic matter breakdown products werenot measured increase in pH, As(III) concentration, andmicrobial numbers support reaction pathways involvingbreakdown of lactate. Lactate added in this study wasdeliberately kept at low levels unlike other enrichmentcultures to simulate conditions that are more natural.This probably results in slow growth and low As(V)

51J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
reduction rates in the enrichment cultures. Moreover, aswith any microbial cultivation method, the microorgan-isms selected under specific enrichment conditions donot necessarily represent all the populations. Instead, thecultivable organisms (as in the case of enrichment mediaor plates) only represent a subset of species enrichedbecause of specific laboratory treatment (e.g., carbonsource, available electron acceptors). Nevertheless, thegeneral trends in As speciation and changes in microbialpopulation are similar to other studies on mine tailings(e.g., Harrington et al., 1998; Macur et al., 2001).
Activity of iron, sulfate, and arsenate-reducingbacteria are known to mobilize As from mine tailingsand aquifer sediments (e.g., Harrington et al., 1998;Cummings et al., 1999; Macur et al., 2001; Malasarnet al., 2004). These microbes act in consortia or some-times alone, and affect redox-based reactions. Hence, itremains to be tested if: 1) similar types of bacteria occurin the Adak sediments, and 2) they act alone, or inconsortium with arsenate-reducing microorganisms(i.e., respirers or detoxifiers) in remobilizing As.
Previous studies indicate that As(V) reductioninvolving microbial respiration only occurs under strictanaerobic conditions (Newman et al., 1997), and only afew microorganisms are capable of As(V) respiration(Oremland and Stolz, 2003). In contrast, As(V)reduction coupled to detoxification reactions is func-tional under both aerobic and anaerobic conditions, andmany diverse microorganisms are capable of metabo-lizing the reaction (Oremland and Stolz, 2003; Macur etal., 2004). Therefore, we believe that As reduction inthese sediments predominantly occurs via a processother than As(V) respiration. During detoxification pro-cess, after As(V) enters the cells via the phosphatetransport system, As(V) is reduced to As(III) by the Asresistance system (ArsC). Subsequently, a secondarycarrier protein (ArsB) or an anion-translocating ATPase(ArsAB) helps in extruding As(III) from the cytoplasm(Diorio et al., 1995; Rosen, 1999). Ultimately, which-ever metabolic pathway is adapted by these heterotro-phic microorganisms As(V) reduction would continueas long as: 1) organic matter is available, 2) microbesneed to detoxify themselves for survival purposes, or 3)energy is derived from As(V) reduction. The inescap-able consequence of these metabolic processes is thebuildup of toxic levels of As(III) in the system.
The unusual distribution of As species in the Adaksurface and ground water highlights the role of in situmicroorganisms in As speciation and cycling. In fact,there is similarity with few other studies where As(III)concentrations are high compared to As(V) in oxicsurface and groundwater environments (e.g., Kuhn and
Sigg, 1993; Aurillo et al., 1994; Sohrin et al., 1997).These microbial processes make prediction of As va-lence and behavior based solely on redox status a prob-lematic issue. Moreover, these processes could also havean impact on water treatment and remediation methodsif they are widespread in sub-surface environments.
3.3. Arsenic remobilization and environmentalimplications
It is important to understand the implications of thesebiogeochemical processes in natural environments eventhough the experiments are primarily laboratory based.Appropriate scaling of these experiments could providea likelihood of these processes, and explain the unusualtrends observed in the Adak surface and ground watersamples and microcosm experiments. In order to modelAs transformation and remobilization in these time-series experiments we assumed: 1) dissolution of As andinorganic transformation of As(V) to As(III) are slowreactions compared to microbial processes (e.g., Kuhnand Sigg, 1993; Newman et al., 1997, 1998), and 2)increase in dissolved As concentration in the mediumrepresents microbially mediated transport of As acrossthe sediment–water interface. These assumptions arebased on the following observations in the microcosmexperiments: 1) decrease in Astot concentration in sed-iments is coupled to an equivalent increase in Astotconcentration in the aqueous phase, and 2) change in As(V) reduction coincides with the increase in heterotro-phic plate counts. We calculated As remobilization ratein the time-series experiments as:
Arsenic remobilization ðAM cm−2 yr−1Þ¼ DðAs concÞ
SA*Timeð2Þ
where As concentration represents the change in dis-solved Astot concentration in 500 ml of medium over a4-week period; SA is the surface area of the bottle (5 cmradius).
Arsenic remobilization rates in Lake Ruttjejaurerange between 6.64×10−3 and 9.13×10−3 μM cm−2
yr−1 in the anaerobic samples, and between 13.3×10−3
and 18.3×10−3 μM cm−2 yr−1 in the aerobic samples.Estimated values are slightly higher compared to thoseobserved in Lake Balmar in northwestern Ontario(Martin and Pedersen, 2002). Lake Balmar affected bymine tailings indicates As remobilization rates between0.03×10−3 and 2.42×10−3 μM cm−2 yr−1. The authorssuggested that the change in redox status is indirectlyassociated with a postulated increase in the lake's

52 J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
primary productivity, and an increase in sediment-derived As. Likewise, the study in the Aberjonawatershed by Ahmann et al. (1997) indicate microbialprocesses actively involved in solubilization of As fromsediments, and comparable As remobilization rates. Thepresent study and others (e.g., Ahmann et al., 1997;Martin and Pedersen, 2002; Macur et al., 2004; Routh etal., 2005) imply that biogeochemical processes play animportant role in controlling As remobilization acrossthe sediment–water interface. Nevertheless, it is impor-tant to bear in mind that these estimates are based onlaboratory studies, which are substantially differentfrom natural conditions. Moreover, As(III) released intothe medium is strongly influenced by: 1) sorption of Asto ferrihydrite, alumina, sulfide, and dissolved organicmatter, and 2) microbial oxidation of As(III) (e.g.,Belzille and Tessier, 1990; Harrington et al., 1998;Zobrist et al., 2000; Santini et al., 2000; Redman et al.,2002). Hence, these results should not be taken literally.Instead, the data should only be used to understand theprocesses involved in microbially mediated As cycling.
In As affected areas, groundwater treatment mostcommonly involves employing different innovativemethods to keep the conditions aerobic (Brannon andPatrick, 1987; Masscheleyn et al., 1991; Murcott, 2001).The rationale is that aerobic conditions will inhibit thereduction of As(V) to As(III). The present study showsthat microbial reduction of As(V) could remobilize As(V) from sediments after converting it to As(III). It islikely that similar processes could occur in other Asaffected sites too. This could be one of the reasons for thelow efficiency (and sometimes even failure) of commonin situ remediation methods involving the use of chem-ical oxidants and aeration in aquifers across India andBangladesh (Murcott, 2001; Hossain et al., 2005). Thereasons for the poor outcome are often unknown, or theyhave not been thoroughly investigated. Hence, it is rec-ommended to explore these microbial processes as partof regular on-site investigations before undertakingcostly As remediation methods. In the long run,elucidating the important pathways responsible for Asreduction could be beneficial for designing appropriateremediation methods that minimize surface and ground-water contamination.
4. Future directions
It is important to continue investigating the specificconditions, or processes favoring reactions that directlyreduce As(V), and release the bound arsenical fractionfrom sediments. Ongoing studies at Adak focus onexperiments involved with: 1) isolation and genetic
characterization of microorganisms from enrichmentscultures using different electron donors and acceptors,2) microbial growth and As(V) reduction rates, and 3)specific conditions that favor the growth of thesemicroorganisms.
Genetic characterization (16S RNA technique)indicates that microorganisms isolated from the enrich-ment cultures are predominantly aerobic or facultativeanaerobic species (Routh and Saraswathy, 2005; Routhet al., in press; Routh, unpublished data). Positive matchwith GeneBank helped in identifying some of thesemicrobial species namely Arthrobacter globiformis,Bacillus pumilus, Pseudomonas mandelii, and Rhodo-coccus erythropolis. In addition, tentative identificationhas been suggested for four other microbial species(Enevold Falsen, CCUG, personal communication), butfurther work is needed to establish them as new strains.While most of the microorganisms isolated so far areassociated with As(V) reduction, none of them aredirectly involved in AS(III) oxidation (J. Santini,personal communication). For example, we recentlyidentified a novel species of actinomycete Arsenicicoc-cus bolidensis in these sediments (Collins et al., 2004),and described its metabolic characteristics (Routh et al.,in press). A. bolidensis is capable of reducing As(V) andnitrate, and As(V) reduction is related to growth (Routhet al., in press). However, it remains critical togenetically identify the arsenate reductase genes (in A.bolidensis and in other microorganisms isolated fromthis site) to ascertain whether As(V) reduction resultsfrom respiration or detoxification process or a combi-nation of both (e.g., Malasarn et al., 2004). It isinteresting to note that in their review on the ecology ofAs in sedimentary environments Oremland and Stolz(2003) concluded that ‘…presence of an enzymaticdetoxification pathway does not preclude the As(V)respiration capability in bacteria…’.
5. Conclusions
Weathering of mine tailings result in acid minedrainage and trace element contamination in Adak.Water, sediment, and soil samples indicate high concen-trations of several toxic elements (e.g., As, Cu, and Zn).The unusual distribution of As species characterized byhigh As(III) levels in the oxic waters suggests activeinvolvement of microbial processes in As cycling.
We investigated the possible biological mechanismscontributing towards transformation and remobilizationof As. The microcosm experiments indicate that mi-croorganisms are capable of: 1) surviving in As-richsediments, and 2) reducing As(V) to As(III), and 3)

53J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
enhancing remobilization of As from sediments. Con-sequently, As(III) concentrations increase in the aqueousmedia. In contrast, As(V) concentrations increasesteadily in the controls implying that active metabolismis important for As reduction. The decrease in Astotconcentration in the controls however, suggests thatinorganic processes (desorption or dissolution type re-actions) are also involved in solubilization of As.Microbial As reduction and subsequent remobilizationaffects the distribution of As at this site, and poses acritical challenge. Therefore, it is recommended to in-vestigate the efficiency of these microbial processes infield settings before undertaking expensive As removaltechniques for water treatment.
Acknowledgements
The study was funded through grants from theSwedish Geological Survey (1321401). Tomas Hjorthhelped us with the analyses on the graphite furnace andICP-OES. Alan Martin made suggestions with the fluxcalculation. Joanne Santini helped us with the arseniteoxidation experiments. We thank Chad Saltikov whoreviewed an earlier version of the manuscript. Com-ments from two anonymous reviewers were helpful.
References
Ahmann, D., Krumholz, L.R., Hemond, H.F., Lovley, D.R., Morel,F.M.M., 1997. Microbial mobilization of arsenic from the sedimentsof the Aberjona watershed. Environ. Sci. Technol. 31, 2923–2930.
Aurillo, A.C., Mason, R.P., Hemond, H.F., 1994. Speciation and fateof arsenic in three lakes of the Aberjona watershed. Environ. Sci.Technol. 28, 577–585.
Axelsson, C.L., Karlqvist, L., Lintu, Y., Olsson, T., 1986. Gruvindus-trins restproduktupplag- fältundersökningar med vattenbalansstu-die i Kristineberg. Uppsala Geosystem AB, Sweden (in Swedish).
Belzille, N., Tessier, A., 1990. Interactions between arsenic and ironoxyhydroxides in lacustrine sediments. Geochim. Cosmochim.Acta 54, 103–109.
Bhattacharya, A., Routh, J., Jacks, G., Bhattacharya, P.,Mörth,M., 2006.Environmental assessment of abandoned mine tailings in Adak,Västerbotten district (northern Sweden). Appl. Geochem. 21,1760–1780.
Brannon, M.J., Patrick, W.H., 1987. Fixation, transformation andmobilization of arsenic in sediments. Environ. Sci. Technol. 21,450–459.
Cai, J., Salmon, K., DuBow, M.S., 1998. A chromosomal ars operonhomologue of Pseudomonoas aeruginosa confers increasedresistance to arsenic and antimony in Escherichia coli. Microbi-ology 144, 2705–2713.
Carlsson, E., Öhlander, B., Holmström, H., 2003. Geochemistry of theinfiltrating water in the vadose zone of a remediated tailingsimpoundment, Kristineberg mine, northern Sweden. Appl. Geo-chem. 18, 659–674.
Chappell, J., Chiswell, B., Olszowy, H., 1995. Speciation of arsenic incontaminated soil by solvent extraction. Talanta 42, 323–329.
Collins, M.D., Routh, J., Saraswathy, A., Lawson, P.A., Schumann, P.,Welinder-Olsson, C., Falsen, E., 2004. Arsenicicoccus bolidensisgen. nov., sp. nov., a novel actinomycete isolated from contam-inated lake sediments. Int. J. Syst. Evol. Microbiol. 54, 605–608.
Cullen, W.R., Reimer, K.J., 1989. Arsenic speciation in theenvironment. Chem. Rev. 89, 713–764.
Cummings, D.E., Caccavo, F.J., Fendorf, S., Rosenzweig, R.F., 1999.Arsenic mobilization by dissimilatory Fe (III)-reducing bacteriumShewanella alga BrY. Environ. Sci. Technol. 33, 723–729.
Diorio, C., Cai, J., Marmor, J., Shinder, R., DuBow, M.S., 1995. AnEscherichia coli chromosomal ars operon homolog is functionalin arsenic detoxification and gram-negative bacteria. J. Bacteriol.177, 2050–2056.
Frietsch, R., Billström, K., Perdahl, J.A., 1995. Sulphur isotopes inLower Proterozoic iron and sulphide ores in northern Sweden.Miner. Depos. 30, 275–284.
Gao, S., Fujii, R., Chalmers, A.T., Tanji, K.K., 2004. Evaluation ofadsorbed arsenic and potential contribution to shallow groundwa-ter in Tulare Lake bed area, Tulare Basin, California. Soil Sci. Soc.Am. J. 68, 89–95.
Harrington, J.M., Fendorf, S.E., Rozenzweig, R.F., 1998. Bioticgeneration of Arsenic (III) in metal(loid)-contaminated freshwaterlake sediments. Environ. Sci. Technol. 32, 2425–2430.
Hossain, M.A., Sengupta, M.K., Ahamed, S., Rahman, M.M., Mondal,D., Lodh, D., Das, B., Nayak, B., Roy, B.K., Mukherjee, A.,Chakraborti, D., 2005. Ineffectiveness and poor reliability ofarsenic removal plants in West Bengal, India. Environ. Sci.Technol. 39, 4300–4306.
Jacks, G., Bhattacharya, P., Routh, J., Martin, M.T., 2003. Arsenic cyclingin a covered mine tailings deposit, Northern Sweden. In: Schulz, H.D.,Hadeler, A. (Eds.), Geo Proc 2002. Wiley Publ., pp. 303–309.
Johnson, D.L., Pilson, M.E.Q., 1972. Spectrophotometric determina-tion of arsenite, arsenate and phosphate in natural waters. Anal.Chem. Acta 58, 289–299.
Jones, C.A., Lagner, H.W., Anderson, K., McDermott, T.R., Inskeep,W.P., 2000. Rates of microbially mediated arsenate reduction andsolubilization. Soil Sci. Soc. Am. J. 64, 600–608.
Korte, N.E., Fernando, Q., 1991. A review of As(III) in groundwater.Critical Reviews inEnvironmental Control. CRCPress, Inc., pp. 1–39.
Kuhn, A., Sigg, L., 1993. Arsenic cycling in eutrophic Lake Greifen,Switzerland: influence of seasonal redox processes. Limnol.Oceanogr. 38, 1052–1059.
Laverman, A.M., Blum, J.K., Schaefer, J.K., Phillips, E.J.P., Lovley, D.R., Oremland, R.S., 1995. Growth of strain SES-3with arsenate andother diverse electron acceptors. Appl. Environ. Microbiol. 61,3556–3561.
Macur, R.E., Wheeler, J.T., McDermott, T.R., Inskeep, W.P., 2001.Microbial populations associated with the reduction and enhancedmobilization of arsenic in mine tailings. Environ. Sci. Technol. 35,3676–3682.
Macur, R.E., Jackson, C.R., Botero, L.M., McDermott, T.R., Inskeep,W.P., 2004. Bacterial populations associated with the oxidation andreduction of arsenic in an unsaturated soil. Environ. Sci. Technol.38, 104–111.
Malasarn, D., Saltikov, C.W., Campbell, K.M., Santini, J.M., Hering,J.G., Newman, D.K., 2004. arrA is a reliable marker for As(V)respiration. Science 306, 455.
Manning, B.A., Goldberg, S., 1997. Adsorption and stability of arsenic(III). Environ. Sci. Technol. 31, 2005–2011.

54 J. Routh et al. / Journal of Geochemical Exploration 92 (2007) 43–54
Martin, A.J., Pedersen, T.F., 2002. Seasonal and interannual mobilityof arsenic in a lake impacted by metal mining. Environ. Sci.Technol. 36, 1516–1523.
Masscheleyn, P.H., Delaune, R.D., Patrick, W.H., 1991. Effect ofredox potential and pH on arsenic speciation and solubility incontaminated soil. Environ. Sci. Technol. 25, 1414–1419.
Meng, X., Wang, W., 1998. Speciation of As by disposable cartridges.3rd International Conference on As Exposure and Health Effects,San Diego, CA.
Mukhopadhyay, R., Rosen, B.P., Phung, L.T., Silver, S., 2002.Microbial arsenic: from geocycles to genes and enzymes. FEMSMicrobiol. Rev. 26, 311–325.
Murcott, S., 2001. A comprehensive review of low cost, tubewell watertreatment technologies for arsenic removal. In: Chappell, W.R.,Abernathy, C.O., Calderon, R.L. (Eds.), Arsenic Exposures andHealth Effects, vol. IV. Elsevier Sci. Ltd., pp. 419–429.
Newman, D.K., Kennedy, E.K., Coates, J.D., Ahmann, D., Ellis, D.J.,Lovley, D.R., Morel, F.M.M., 1997. Dissimilatory arsenate andsulfate reduction in Desulfotomaculum auripigmentum sp. nov.Arch. Microbiol. 168, 380–388.
Newman, D.K., Ahmann, D., Morel, F.M.M., 1998. A brief review ofmicrobial arsenate respiration. Geomicrobiol. J. 15, 255–268.
Oremland, R.S., Stolz, J.F., 2003. The ecology of arsenic. Science 300,939–944.
Pantsar-Kallio, M.,Manninen, P.K.G., 1997. Speciation of arsenic in soilsamples as a function of soil pH. Sci. Total Environ. 204, 193–200.
Redman, A.D., Macalady, D.L., Ahmann, D., 2002. Natural organicmatter affects arsenic speciation and sorption onto hematite.Environ. Sci. Technol. 36, 2896–2899.
Rochette, E.A., Bostick, B.C., Li, G., Fendorf, S., 2000. Kinetics ofarsenate reduction by dissolved sulfide. Environ. Sci. Technol. 34,4714–4720.
Rosen, B.P., 1999. Families of arsenic transporters. Trends Microbiol.7, 207–212.
Routh, J., Saraswathy, A., 2005. Microbial processes and arsenicmobilization in mine tailings and shallow aquifers. In: Bundschuh,J., Bhattacharya, P., Chandrasekharam, D. (Eds.), Natural Arsenicin Groundwater: Occurrence, Remediation, and Management.Taylor and Francis, London, pp. 145–153.
Routh, J., Saraswathy, A., Sinha-Ray, S.P., Nag, S.K., Jacks, G., 2005.Arsenic reduction by indigenous microorganisms in shallow aquifers
from Ambikanagar, West Bengal (India). In: O'Day, P.A., Vlassopoulos,D., Meng, X., Benning, L.G. (Eds.), Advances in Arsenic Research:Integration of Experimental and Observational Studies and Implicationsfor Mitigation. ACS Symposium Series, vol. 915. American ChemicalSociety, Washington, D.C., pp. 132–147.
Routh, J., Saraswathy, A., Collins, M.D., in press. Microbial arsenicreduction in mine tailings and impact on water chemistry at Adak,Västerbotten district (northern Sweden). Sci. of Tot. Environ.
Santini, J.M., Sly, L.I., Schnagl, R.D., Macy, J.M., 2000. A new chemo-lithoautotrophic arsenite-oxidizing bacterium isolated from a goldmine: phylogenetic, physiological, and preliminary biochemicalstudies. Appl. Environ. Microbiol. 66, 92–97.
Santini, J.M., Stolz, J.F., Macy, J.M., 2002. Isolation of a new arsenaterespiring bacterium — physiological and phylogenetic studies.Geomicrobiol. J. 19, 41–52.
Sohrin, Y., Matsui, M., Kawashima, M., Hojo, M., Hasegawa, H.,1997. Arsenic biogeochemistry affected by eutrophication in LakeBiwa, Japan. Environ. Sci. Technol. 31, 2712–2720.
Swedish Standards Institute, 1993. Measuring trace metal concentra-tion in sediment: Method SIS028150.
Turpeinen, R., Pantsar-Kallio, M., Häggblom, M., Kairesalo, T., 1999.Influence of microbes on the mobilization, toxicity and biomethy-lation of arsenic in the soil. Sci. Total Environ. 236, 173–180.
Vivallo, W., 1987. Early Proterozoic bimodal volcanism, hydrothermalactivity, and massive sulfide deposition in the Boliden–Långdalarea, Skellefteå district, Sweden. Econ. Geol. 82, 440–456.
Wagner, T., Boyce, A.J., Jonsson, E., Fallick, A.E., 2004. Lasermicroprobe sulphur isotope analysis of arsenopyrite: experimentalcalibration and application to the Boliden Au–Cu–As massivesulphide deposit. Ore Geol. Rev. 25, 311–325.
Willden, M., 1986. Geology of western part of Skellefteå field andKristineberg and Hornträsk sulphide deposits. In: Rikard, D.T.(Ed.), The Skellefte field, 7th IAGOD Symposium ExcursionGuide No 4, Sveriges Geologiska Undersökning, pp. 46–52.
Zobrist, J., Dowdle, P.A., Davis, J.A., Oremland, R.S., 2000. Mobi-lization of arsenite by dissimilatory reduction of adsorbed arsenate.Environ. Sci. Technol. 34, 4747–4753.





























