Embed Size (px)
Citation preview

279
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
© Agricultural and Food Science in FinlandManuscript received September 2000
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
Assessment of genetic diversity using RAPD analysisin a germplasm collection of sea buckthorn
Galyna I. BartishBalsgård – Department of Horticultural Plant Breeding, Swedish University of Agricultural Sciences,
Fjälkestadsvägen 123-1, S-29194 Kristianstad, Sweden. Current address: Natural Science Section,Södertörn University College, PO Box 4101, S-14104 Huddinge, Sweden
Niklas JeppssonBalsgård – Department of Horticultural Plant Breeding, Swedish University of Agricultural Sciences,
Fjälkestadsvägen 123-1, S-29194 Kristianstad, Sweden
Igor V. BartishBalsgård – Department of Horticultural Plant Breeding, Swedish University of Agricultural Sciences,
Fjälkestadsvägen 123-1, S-29194 Kristianstad, Sweden. Current address: Institute of Botany, Academy ofSciences, CZ-25243 Pruhonice 1, Czech Republic
Hilde NybomBalsgård – Department of Horticultural Plant Breeding, Swedish University of Agricultural Sciences,
Fjälkestadsvägen 123-1, S-29194 Kristianstad, Sweden, e-mail: [email protected]
Random amplified polymorphic DNA (RAPD) markers were used to characterize a part of a seabuckthorn gene bank collected for plant breeding purposes. Molecular markers were generated in 55cultivars and accessions, representing five subspecies of Hippophae rhamnoides L. and intraspecifichybrids between different subspecies. Sixty-three markers were used to generate a Dice’s similaritycoefficient matrix of pairwise comparisons between individual RAPD profiles. Cluster (UPGMA)and principal co-ordinate analyses, based on this matrix, revealed clustering of plants into groupswhich generally correspond to their taxonomic classification or geographic origin. The analysis ofmolecular variance (AMOVA) was found useful for estimating components of genetic variation be-tween and within taxonomic and geographic groups of accessions and cultivars. Whereas both alter-natives for grouping the material (taxonomic or geographic origin) resulted in significant between-group variation, the major part of molecular variance (approximately 75%) was still attributed tovariation within groups. We conclude that the RAPD analysis is useful for clarification of taxonomicand geographic origin of accessions and cultivars of sea buckthorn.
Key words: sea buckthorn, Hippophae rhamnoides, genetic diversity, RAPD, core collection, hybrid-ization

280
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Bartish, G.I. et al. RAPD diversity in sea buckthorn
Introduction
A number of plant species, that have only rarelybeen cultivated in the past, are now being inves-tigated for possible domestication with the goalto diversify farming and to introduce crops thatare amenable to organic production methods.One such crop, that has recently received in-creased attention, is the sea buckthorn, Hip-pophae rhamnoides L. This is an outcrossingdioecious pioneer plant, that occurs mainly alongrivers in mountainous regions but also on seashores. It has a wide but discontinuous distribu-tion throughout Europe and Asia, from the northof Europe to central China. The species is fur-ther divided into nine subspecies (Rousi 1971).Its berries can be used for juice and jam produc-tion. However, these berries also contain aromat-ic and medicinal compounds that can be extract-ed and used e.g. in functional food products, inmedicine and in cosmetics. The plant itself iswidely used for its ornamental value and for theprevention of soil erosion (Trajkovski and Jepps-son 1999).
Domestication of sea buckthorn started inSiberia in the 1930s (Kalinina and Panteleyeva1987) with local germplasm (ssp. mongolica).The interest increased and it was soon introducedto other regions of Russia and to neighbouringcountries (Trajkovski and Jeppsson 1999). Theneed for plant material, that was adapted to thelocal climate, prompted the onset of breedingprogrammes, which have generated numerouscultivars. At present, more than 60 cultivars havebeen described (Trajkovski and Jeppsson 1999).In addition to ssp. mongolica and ssp. turkesta-nica of central Asian origin, the European sub-species rhamnoides and carpatica and ssp. cau-casica from Minor Asia have also been utilizedfor breeding purposes. Hybridization betweenthese different subspecies has been used exten-sively in efforts to combine traits from superiorgenotypes.
Possibly one of the world’s most diverse Hip-pophae gene banks is now available at Balsgård– Department of Horticultural Plant Breeding,
S. Sweden. This gene bank is composed of allthe wild taxa in the genus from different parts ofthe world, as well as cultivars and selectionsfrom different breeding programmes. Knowledgeof genetic relatedness among accessions of agermplasm collection is necessary for develop-ment of conservation strategies, since the aim isto preserve as much variability as possible witha minimum repetitiveness. Most crop germplasmcollections contain a considerable percentage ofmisclassified accessions, which may seriouslyaffect the use of germplasm for horticulturalimprovement. The efficient organization of genebank resources is therefore an important task,and a core collection should be developed(Hokanson et al. 1998). Such a subset of the en-tire germplasm collection should, ideally, rep-resent the genetic diversity found within thewhole collection. In addition, information aboutlevels of relatedness can, for example, enable thebreeder to take advantage of the heterosis effectproduced by hybridization between geneticallydistant genotypes. Unfortunately, informationabout taxonomic and geographic origin is notalways available for gene bank accessions, noteven when these consist of recently marketedcultivars.
The present study was performed to studygenetic relatedness and diversity in a small sub-set of the Hippophae gene bank at Balsgård. Thissubset was composed of accessions from nativeH. rhamnoides populations, cultivars and selec-tions from different breeding programmes. Weused random amplified polymorphic DNA(RAPD), which has been shown to be one of themost cost-effective methods for obtaining poly-morphic markers in many plant genera (Tivanget al. 1996, Bartish et al. 1999b, Virk et al. 2000).In sea buckthorn, RAPD analysis has recentlybeen applied to studying the partitioning of ge-netic variation in plants from native stands (Bar-tish et al. 1999a) and to develop a marker forsex determination (Persson and Nybom 1999).In addition, a representative sample of all pres-ently recognized taxa in the genus Hippophaehas been subjected to a phenetic analysis usingRAPD markers (Bartish et al. 2000). This infor-

281
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
mation will be used in the process of develop-ing a core collection of sea buckthorn.
Material and methods
Plant material and DNA extractionWe selected 55 plants of Hippophae rhamnoidesL. including subspecies carpatica, caucasica,fluviatilis, mongolica, and rhamnoides. Informa-tion about geographic origin of these accessionswas available in most cases (Table 1) and wasused to divide the collection into groups. Twoapproaches for this division were applied: geo-graphic origin (10 groups) and classification (6groups; i.e. 5 subspecies and the inter-subspe-cific hybrids). Dormant shoots were collected inJanuary and forced, in a greenhouse, to producefresh leaves. DNA extraction from leaf tissue andgeneration of RAPD markers was performedaccording to Bartish et al. (2000).
RAPD analysisInitial screening of decamer primers has beenperformed previously on Hippophae sp. materi-al (Persson and Nybom 1999, Bartish et al.1999a). We selected eight primers from OperonTechnologies, Inc., based on the presence ofpolymorphism, the ability to discriminate be-tween populations, and the reproducibility ofamplifications (Table 2). PCR amplificationswere performed according to Bartish et al.(2000). Amplification products were separatedby electrophoresis in 1.8% agarose gels in 1 xTPE buffer (Weising et al. 1994), stained withethidium bromide, and documented under UVlight with polaroid photography.
Data analysisEach polymorphic band in the RAPD profile was
treated as an independent locus with two alle-les, based on the presence or absence of the band.Amplification products were scored manually,1 for presence and 0 for absence. A binary ma-trix was generated based on 63 polymorphicRAPD bands. Only reliably scored bands with asize of 200–1600 bp were included in the analy-sis.
Dice’s coefficient of similarity was calculat-ed for all pairwise comparisons between indi-vidual samples from the binary RAPD data ma-trix according to the formula
%SAB
=[2NAB
/(NA+N
B)]100,
where %SAB
is the coefficient of similaritybetween individuals A and B, N
A and N
B is the
number of bands for sample A and B respective-ly, and N
AB is the number of shared bands for A
and B. This matrix was analysed by cluster anal-ysis (UPGMA method, SAHN program, NTSYS-pc, Rohlf 1997). Additional information aboutgenetic relationships between individual plantswas derived by principal co-ordinate analysis(PCO, DECENTER and EIGENVECTOR pro-grams, NTSYS-pc) by which a two-dimension-al plot was obtained.
A second matrix, this time using Euclideandistances among all individual plants, was com-puted from the binary RAPD data matrix. Weused this new matrix as an input distance matrixfor an Analysis of Molecular Variance (AMO-VA), as in Excoffier et al. (1992) and Huff et al.(1993), for partitioning the genetic variance intocomponents within and among plant groups andfor testing the significance of the estimates ob-tained.
Results and discussion
RAPD analysis of 8 decamer primers revealed63 polymorphic (86.3%) and 10 monomorphicmarkers (Table 2), indicating a relatively highdiversity in our plant sample, which thus pro-vides a promising foundation for further breed-
282
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Bartish, G.I. et al. RAPD diversity in sea buckthorn
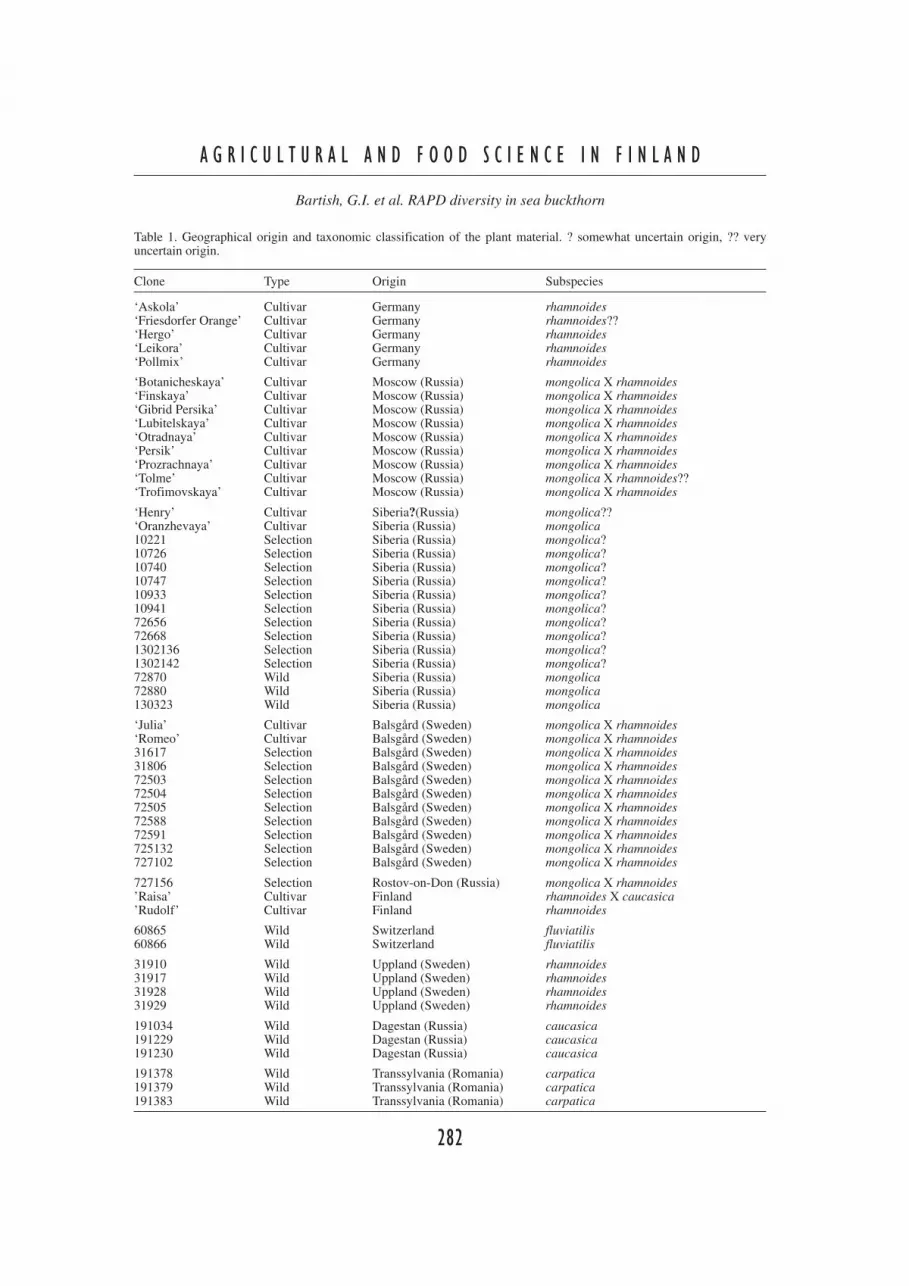
Table 1. Geographical origin and taxonomic classification of the plant material. ? somewhat uncertain origin, ?? veryuncertain origin.
Clone Type Origin Subspecies
‘Askola’ Cultivar Germany rhamnoides‘Friesdorfer Orange’ Cultivar Germany rhamnoides??‘Hergo’ Cultivar Germany rhamnoides‘Leikora’ Cultivar Germany rhamnoides‘Pollmix’ Cultivar Germany rhamnoides
‘Botanicheskaya’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Finskaya’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Gibrid Persika’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Lubitelskaya’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Otradnaya’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Persik’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Prozrachnaya’ Cultivar Moscow (Russia) mongolica X rhamnoides‘Tolme’ Cultivar Moscow (Russia) mongolica X rhamnoides??‘Trofimovskaya’ Cultivar Moscow (Russia) mongolica X rhamnoides
‘Henry’ Cultivar Siberia?(Russia) mongolica??‘Oranzhevaya’ Cultivar Siberia (Russia) mongolica10221 Selection Siberia (Russia) mongolica?10726 Selection Siberia (Russia) mongolica?10740 Selection Siberia (Russia) mongolica?10747 Selection Siberia (Russia) mongolica?10933 Selection Siberia (Russia) mongolica?10941 Selection Siberia (Russia) mongolica?72656 Selection Siberia (Russia) mongolica?72668 Selection Siberia (Russia) mongolica?1302136 Selection Siberia (Russia) mongolica?1302142 Selection Siberia (Russia) mongolica?72870 Wild Siberia (Russia) mongolica72880 Wild Siberia (Russia) mongolica130323 Wild Siberia (Russia) mongolica
‘Julia’ Cultivar Balsgård (Sweden) mongolica X rhamnoides‘Romeo’ Cultivar Balsgård (Sweden) mongolica X rhamnoides31617 Selection Balsgård (Sweden) mongolica X rhamnoides31806 Selection Balsgård (Sweden) mongolica X rhamnoides72503 Selection Balsgård (Sweden) mongolica X rhamnoides72504 Selection Balsgård (Sweden) mongolica X rhamnoides72505 Selection Balsgård (Sweden) mongolica X rhamnoides72588 Selection Balsgård (Sweden) mongolica X rhamnoides72591 Selection Balsgård (Sweden) mongolica X rhamnoides725132 Selection Balsgård (Sweden) mongolica X rhamnoides727102 Selection Balsgård (Sweden) mongolica X rhamnoides
727156 Selection Rostov-on-Don (Russia) mongolica X rhamnoides’Raisa’ Cultivar Finland rhamnoides X caucasica’Rudolf’ Cultivar Finland rhamnoides
60865 Wild Switzerland fluviatilis60866 Wild Switzerland fluviatilis
31910 Wild Uppland (Sweden) rhamnoides31917 Wild Uppland (Sweden) rhamnoides31928 Wild Uppland (Sweden) rhamnoides31929 Wild Uppland (Sweden) rhamnoides
191034 Wild Dagestan (Russia) caucasica191229 Wild Dagestan (Russia) caucasica191230 Wild Dagestan (Russia) caucasica
191378 Wild Transsylvania (Romania) carpatica191379 Wild Transsylvania (Romania) carpatica191383 Wild Transsylvania (Romania) carpatica

283
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
ing activities. For comparison, the following lev-els of within-species RAPD polymorphism havebeen reported in other woody plant species: 58%in the endangered subtropical shrub Halora-godendron lucasii (Sydes and Peakall 1998),61% in the tropical tree Gliricidia sepium(Chalmers et al. 1992), 80% in the tropical treeTheobroma cacao (Russell et al. 1993), 92% inthe tropical tree Cedrela odorata (Gillies et al.1997), 97% in the endangered subtropical treeCaesalpinia echinata (Cardoso et al. 1998), and98% in the endangered subtropical shrub Gre-villea scapigera (Rossetto et al. 1995).
Partitioning molecular variation by AMOVArevealed that most variation resides within geo-graphic or taxonomic groups (75.3% and 76.0%,respectively; P<0.001; Table 3) in spite of thediverse geographic and taxonomic origins of theaccessions studied. Similarly, within-subspeciesvariation was found to be considerably higherthan between-subspecies variation in a previousRAPD study based on natural populations ofseveral Hippophae species (Bartish et al. 2000).
The UPGMA dendrogram divides the stud-ied individuals into three main groups and a fewrather isolated single accessions. The first ofthese main groups consists mostly of cultivars,selections and wild accessions of ssp. mongoli-ca from Siberia (Fig. 1). One noteworthy pointis that within this group we find the Finnish cul-tivar ‘Henry’. According to records at the nurs-ery Jalaja Oy (Mr. Kettonen, personal commu-nication), this clone was introduced from Nizh-
ny Novgorod (Russia). Since breeding pro-grammes on H. rhamnoides in Russia rely most-ly on ssp. mongolica (Kalinina and Panteleyeva1987), it is reasonable to believe that ‘Henry’belongs to this subspecies. In somewhat periph-eral positions in this group we find accession31617, which is a genotype selected from a hy-brid progeny (mongolica x rhamnoides x cau-casica) and accessions 1302136 and 1302142,which are claimed to have been raised from openpollination of ‘Zyryanka’, a cultivar reported tobe a gamma-ray induced mutation of Baltic Seaorigin (ssp. rhamnoides, Burmistrov 1995).
The second main group (Fig. 1) consistsmostly of plants from different breeding pro-grammes, and all of them, with only one excep-
Table 2. Primers used, number of polymorphic and monomorphic bands, and percentages of polymorphicbands.
Primers Sequence Number of Number of Polymorphism(5’ to 3’) polymorphic monomorphic %
bands bands
OPA-11 CAATCGCCGT 8 0 100OPA-15 TTCCGAACCC 6 2 75OPB-4 GGACTGGAGT 10 0 100OPB-18 CCACAGCAGT 7 3 70OPD-12 CACCGTATCC 8 2 80OPD-18 GAGAGCCAAC 9 1 90OPE-6 AAGACCCCTC 9 0 100
Table 3. Partitioning of molecular variance into componentsby AMOVA at different hierarchical levels. Groups weredefined according to Table 1. All values are significant(P<0.001).
Hierarchial level of variance Percentage of variation
Variation between taxonomic groups 19.5Variation between geographic groups
within taxonomic groups 7.0Variation within geographic groups
within taxonomic groups 73.5
Variation between geographic groups 24.7Variation within geographic groups 75.3
Variation between taxonomic groups 24.0Variation within taxonomic groups 76.0

284
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Bartish, G.I. et al. RAPD diversity in sea buckthorn
Fig. 1. UPGMA dendrogram based on RAPD data (Dice’s pairwise coefficients of similarity) for 55 accessions of Hip-pophae rhamnoides. The symbols correspond to the division by geographic groups presented in Table 1. For labelling ofsymbols, see Fig. 2.

285
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
tion (accession 31929 from Uppland), belong tossp. mongolica or are hybrids between ssp. mon-golica and ssp. rhamnoides. ‘Friesdorfer Or-ange’, a German cultivar of unknown taxonom-ic origin, also clusters within this group.
The third main group (Fig. 1) consists of tax-onomically diverse samples: wild accessions ofsubspecies carpatica, caucasica, fluviatilis andrhamnoides and cultivars obtained from Germa-ny and Finland. All of the German cultivars inthis cluster are derived from ssp. rhamnoides.By contrast, the Finnish cultivars represent aheterogeneous group: ‘Tolme’ was originallyobtained from Moscow and its taxonomic originis unknown, ‘Raisa’ is a hybrid between ssp.rhamnoides and ssp. caucasica, and ‘Rudolf’ isderived from ssp. rhamnoides (Dr. Saila Karhu,personal communication). The pronounced dif-ferentiation between ssp. mongolica on the onehand, and the other four subspecies on the otherhand, is in good accordance with a previousRAPD-based study on natural populations (Bar-tish et al. 2000), which found ssp. mongolica tobe rather well separated from the other four pres-ently investigated subspecies, and instead veryclose to ssp. turkestanica.
In the present study, some sibling plants clus-tered closely as expected, e.g. the three halfsibpairs 1302142/1302136 (group I), 10933/10941and 72656/72668 (group II). However, in othercases siblings were placed into different groups,e.g. the halfsib pair 10726 (group II) /10740(group I). Another example is given by the half-sib pair 60865/60866 with 60866 grouped to-gether with the other native European samples(group III), whereas 60865 has diverged strong-ly from all other accessions. Yet other siblingprogenies show intermediate clustering; e.g. thehalfsibs ‘Romeo’, ‘Julia’ and 31806 groupedclosely (group II) whereas the fourth sibling,31617, was placed in group I. The fullsib group72503/72504/72505 was rather dispersed but stillall were placed within group II.
Results of the principal component analysis(Fig. 2) were generally congruent with the UPG-MA dendrogram and the same main groups couldbe distinguished. Two plants did not cluster with
any of these main groups; one is a wild acces-sion of ssp. fluviatilis (60865) and another(31617) is a hybrid, obtained at Balsgård fromopen pollination with participation of three sub-species (ssp. caucasica, ssp. mongolica and ssp.rhamnoides).
One of the main groups (ellipse A) includesonly plants from ssp. mongolica and the secondgroup (ellipse B) includes mainly hybrids be-tween ssp. rhamnoides and ssp. mongolica. Sev-eral plants within this group were obtained froma Siberian breeding program (Lisavenko insti-tute, Barnaul, Siberia). In addition to plants be-longing to ssp. mongolica, this breeding pro-gramme used wild accessions of ssp. rhamnoidesfrom Baltic populations as a source of addition-al genetic diversity (Kalinina and Panteleyeva1987). A hybrid origin was actually verified foraccessions 10933 and 10941 by the original let-ter that was included with the seed introduction.The cultivars ‘Trofimovskaya’ and ‘Botan-icheskaya’ were obtained at the Botanical gar-den of Moscow University, by open pollinationof plants from ssp. rhamnoides which grew to-gether with plants from ssp. mongolica(Mikheyev and Demenko 1990). Accessions10740 and 10747 are derived from the cultivar‘Zolotoy Pochatok’, which was selected fromwild populations of ssp. mongolica from Sibe-ria (Kalinina and Panteleyeva 1987). Since ac-cession 10747 was placed among hybrids ofmongolica x rhamnoides (Fig. 2, ellipse B), wecan hypothesise that this plant also contains ge-netic material from ssp. rhamnoides. The culti-vars ‘Persik’ and ‘Otradnaya’ were developed atthe Botanical Garden of Moscow University, byopen pollination of wild populations from theSaint-Petersburg area (Mikheyev and Demenko1990). Although they have a similar origin, thePCO plot reveals considerable differentiationbetween them and suggests that introgressionfrom ssp. mongolica was more pronounced inthe cultivar ‘Otradnaya’.
Ellipse C on the PCO plot contains represent-atives of all subspecies, apart from ssp. mongol-ica. The cultivars from Germany and Finland aremore heterogeneous genetically than the wild

286
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Bartish, G.I. et al. RAPD diversity in sea buckthorn
Fig. 2. Two-dimensional representation of the results of a principal co-ordinate analysis based on pairwise coefficients ofsimilarity among 55 accessions of Hippophae rhamnoides. Percentages of variability explained by the two components:PCO1 18.1%, PCO2 9.2%.
accessions of the same subspecies, which indi-cates that some of those cultivars may have beenderived through hybridization between differenttaxa. The Finnish cultivar ‘Raisa’ is actually ahybrid between ssp. rhamnoides and ssp. cau-casica. The German cultivar ‘Friesdorfer Or-ange’ may also be of hybrid origin according toits placement far from the wild accessions. Thisis further supported by the UPGMA analysiswhere it was placed in group II, which consistsmainly of hybrids.
Random mating of selected superior pheno-types, followed by mass selection in progenies,is a commonly practiced breeding strategy infruit tree crops (Alston and Spiegel-Roy 1985).However, regardless of whether inbreeding
should be avoided or promoted, informationabout the genetic relatedness of the breedingmaterial is useful when trying to locate materialwith traits of interest and when designing abreeding programme. In the present study,RAPDs proved to be relatively efficient in group-ing sea buckthorn genotypes according to theirtaxonomic classification and to their geograph-ic origin. RAPDs have previously been used forsuccessful geographic classification of cultivarsof unknown origin, e.g. pistachio (Pistacia vera)(Hormaza et al. 1994).
It is not always straightforward to estimategenetic relatedness among accessions whenbased exclusively on pedigree information, es-pecially not when only the maternal parent is

287
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
known as was the case for all halfsibs in ourstudy. However, Garcia et al. (1998) reported ahigh correlation between RAPD-based similari-ty estimates and pedigree information in melonbreeding lines. Moreover, in that material, RAPDmarkers appeared to be more suitable than agro-nomic traits in predicting genetic distance amongdifferent breeding lines. A good concordancebetween RAPD data and pedigree informationwas found in a set of strawberry cultivars (Han-cock et al. 1994). Also in strawberries, a lack ofagreement between RAPD-based similarity es-timates and known pedigree data was, however,reported in another study (Graham et al. 1996),as well as a relatively weak agreement in a thirdstudy (Degani et al. 1998). In these latter stud-ies, incorrect naming of parental clones and alow number of applied RAPD markers were sug-gested as the causes. Although no associationwas found between DNA markers and pedigreerelationships in plantain and banana, differentploidy levels were represented and that may haveseriously biased the expected contribution ofparental genomes to the analyzed offspring(Tenkouano et al. 1999).
To set up a core collection, Brown (1989)suggests a stratified sampling approach: thewhole collection is first divided into non-over-lapping groups from which samples are taken byrandom. The hierarchy of grouping starts withtaxonomy. The correct classification of the cul-tivated material of sea buckthorn is, in many
cases, unknown. In addition, hybridization be-tween subspecies is a common practise in manybreeding programmes. Classification using mor-phological characters is not always reliable inthese cases since neither leaves nor flowers of-fer many diagnostic features.
In the present study, the investigated plantcollection was divided into three distinct groupsby use of RAPD markers; 1) ssp. mongolica,2) the European subspecies and 3) hybrids be-tween ssp. mongolica and the European subspe-cies. To obtain a representation of wild materialfrom different taxa, further collection in nativestands can be recommended, since the exact or-igin will then be known. For the more heteroge-neous cultivated material, however, RAPDs mayserve as a useful classification tool. Agronomi-cal as well as morphological traits should alsobe scored in addition to molecular markers sincethe core collection must provide a good repre-sentation of all traits found in the whole germ-plasm collection. However, in some cases a traitmay be of polyphyletic origin (Gepts 1995). Thismay actually be quite common in cultivated plantmaterial, since the aims of different breeding pro-grammes are often similar, even if material fromdifferent origins has been used. In such cases,molecular markers can be used to ensure theconservation of different evolutionary lineages.
Acknowledgements. This study was funded by a grant fromthe Swedish Research Council for Forestry and Agricul-ture to HN.
ReferencesAlston, F.H. & Spiegel-Roy, P. 1985. Fruit tree breeding:
strategies, achievements and constraints. In: Cannel,M.G.R. & Jackson, J.E. (eds.). Attributes of trees ascrop plants. Institute of Terrestrial Ecology, Hunting-don, UK. p. 49–67.
Bartish, I.V., Jeppsson, N., Bartish, G.I., Lu, R. & Ny-bom, H. 2000. Inter- and intraspecific genetic varia-tion in Hippophae (Elaeagnaceae) investigated byRAPD markers. Plant Systematics and Evolution225 (in press).
– , Jeppsson, N. & Nybom, H. 1999a. Population genet-
ic structure in the dioecious pioneer plant speciesHippophae rhamnoides investigated by random am-plified polymorphic DNA (RAPD) markers. MolecularEcology 8: 791–802.
– , Rumpunen, K. & Nybom, H. 1999b. Genetic diversi-ty in Chaenomeles (Rosaceae) revealed by RAPDanalysis. Plant Systematics and Evolution 214: 131–145.
Brown, A.H.D. 1989. Core collections: a practical ap-proach to genetic resources management. Genome31: 818–824.

288
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Bartish, G.I. et al. RAPD diversity in sea buckthorn
Burmistrov, L.A. 1995. Genetic resources and breedingpotential of sea buckthorn (Hippophae rhamnoides)in Russia. WANATCA Yearbook 19: 9–13.
Cardoso, M.A., Provan, J., Powell, W., Ferreiras, P.C.G.& De Oliveira, D.E. 1998. High genetic differentia-tion among remnant populations of the endangeredCaesalpinia echinata Lam. (Leguminosae-Caesalpi-noideae). Molecular Ecology 7: 601–608.
Chalmers, K.J., Waugh, R., Sprent, J.I., Simons, A.J. &Powell, W. 1992. Detection of genetic variation be-tween and within populations of Gliricidia sepium andG. maculata using RAPD markers. Heredity 69: 465–472.
Degani, C., Rowland, L.J., Levi, A., Hortynski, J.A. &Galletta, G.J. 1998. DNA fingerprinting of strawberry(Fragaria x ananassa) cultivars using randomly am-plified polymorphic DNA (RAPD) markers. Euphyti-ca 102: 247–253.
Excoffier, L., Smouse, P.E. & Quattro, J.M. 1992. Analy-sis of molecular variance inferred from metric dis-tances among DNA haplotypes: application to humanmitochondrial DNA restriction data. Genetics 131:479–491.
Garcia, E., Jamilena, M., Alvarez, J.I., Arnedo, T., Oliver,J.L. & Lozano, R. 1998. Genetic relationships amongmelon breeding lines revealed by RAPD markers andagronomic traits. Theoretical and Applied Genetics96: 878–885.
Gepts, P. 1995. Genetic markers and core collections.In: Hodgkin, T. et al. (eds.). Core collections of plantgenetic resources. John Wiley & Sons, Chichester,UK. p. 127–146.
Gillies, A.C.M., Cornelius, J.P., Newton, A.C., Navarro,C., Hernández, M. & Wilson, J. 1997. Genetic varia-tion in Costa Rican populations of the tropical timberspecies Cedrela odorata L., assessed using RAPDs.Molecular Ecology 6: 1133–1145.
Graham, J., McNicol, R.J. & McNicol, J.W. 1996. A com-parison of methods for estimation of genetic diversi-ty in strawberry cultivars. Theoretical and AppliedGenetics 93: 402–406.
Hancock, J.F., Callow, P.A. & Shaw, D.V. 1994. Random-ly amplified polymorphic DNAs in the cultivated straw-berry (Fragaria x ananassa). Journal of the Ameri-can Society of Horticultural Science 118: 862–864.
Hokanson, S.C., Szewc-McFadden, A.K., Lamboy, W.F.& McFerson, J.R. 1998. Microsatellite (SSR) mark-ers reveal genetic identities, genetic diversity andrelationships in a Malus x domestica Borkh. coresubset collection. Theoretical and Applied Genetics97: 671–683.
Hormaza, J.I., Dollo, L. & Polito, V.S. 1994. Determina-tion of relatedness and geographical movements ofPistacia vera (Pistachio; Anacardiaceae) germplasm
by RAPD analysis. Economic Botany 48: 349–358.Huff, D.R., Peakall, R. & Smouse, P.E. 1993. RAPD var-
iation within and among natural population of out-crossing buffalograss (Buchloe dactyloides (Nutt.)Engelm.). Theoretical and Applied Genetics 86: 927–934.
Kalinina, I.P. & Panteleyeva, Y.I. 1987. Breeding of seabucktorn in the Altaj. In: Hryukov, A.B. (ed.). Advancesin agricultural science. Moscow. p. 76–87.
Mikheyev, A.M. & Demenko, V.I. 1990. Oblepiha. Rosa-gropromizdat. Moscow.
Persson, H.A. & Nybom, H. 1999. Genetic sex determi-nation and RAPD marker segregation in the dioeciousspecies sea buckthorn (Hippophae rhamnoides L.).Hereditas 129: 45–51.
Rohlf, F.J. 1997. NTSYS-pc. Numerical taxonomy andmultivariate analysis system, version 1.80. ExeterSoftware, Setauket, NY.
Rossetto, M., Weaver, P.K. & Dixon, K.W. 1995. Use ofthe RAPD analysis in devising conservation strate-gies for the rare and endangered Grevillea scapig-era (Proteaceae). Molecular Ecology 4: 321–329.
Rousi, A. 1971. The genus Hippophae L. A taxonomicstudy. Annales Botanici Fennici 8: 177–227.
Russell, J.R., Hosein, F., Johnson, E., Waugh, R. & Pow-ell, W. 1993. Genetic differentiation of cocoa (Theo-broma cacao L.) populations revealed by RAPD anal-ysis. Molecular Ecology 2: 89–97.
Sydes, M.A. & Peakall, R. 1998. Extensive clonality inthe endangered shrub Haloragodendron lucasii (Hal-oragaceae) revealed by allozymes and RAPDs. Mo-lecular Ecology 7: 87–93.
Tenkouano, A., Crouch, J.H., Crouch, H.K., Vuylsteke,D. & Ortiz, R. 1999. Comparison of DNA marker andpedigree-based methods of genetic analysis of plan-tain and banana (Musa spp.) clones. I. Estimation ofgenetic relationships. Theoretical and Applied Ge-netics 98: 62–68.
Tivang, J., Skroch, P. & Nienhuis, J. 1996. Randomlyamplified polymorphic DNA (RAPD) variation amongand within artichoke (Cynara scolymus L.) cultivarsand breeding populations. Journal of the AmericanSociety of Horticultural Science 121: 783–788.
Trajkovski, V. & Jeppsson, N. 1999. Domestication of seabuckthorn. Botanica Lithuanica, suppl. 2: 37–46.
Virk, P.S., Zhu, J., Newbury, H.J., Bryan, G.J., Jackson,M.T. & Ford-Lloyd, B.V. 2000. Effectiveness of differ-ent classes of molecular marker for classifying andrevealing variation in rice (Oryza sativa) germplasm.Euphytica 112: 275–284.
Weising, K., Nybom, H., Wolff, K. & Meyer, W. 1994. DNAfingerprinting in plants and fungi. CRC Press, BocaRaton, FL.

289
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Vol. 9 (2000): 279–289.
SELOSTUSTyrnin geneettisen monimuotoisuuden arviointi RAPD analyysillä
Galyna I. Bartish, Niklas Jeppsson, Igor V. Bartish ja Hilde NybomSwedish University of Agricultural Sciences, Ruotsi
RAPD (Random amplified polymorphic DNA)-mark-kerien avulla kuvailtiin osaa tyrnin geenipankista,joka oli koottu jalostustarkoituksiin. Molekyylimark-kereita monistettiin 55 näytteestä, jotka edustivat viit-tä tyrnin (Hippophae rhamnoides L.) alalajia ja erialalajien välisiä hybridejä. Kuudenkymmenenkolmenmarkkerin avulla laskettiin Dicen samankaltaisuus-kerroinmatriisi (Dice’s similarity coefficient matrix)yksittäisten RAPD profiilien parittaisista vertailuis-ta. Matriisiin perustuvien analyysien (UPGMA- andprincipal co-ordinate analyses) avulla kasvit jakau-tuivat ryhmiin, jotka vastaavat niiden taksonomistaluokkaa ja maantieteellistä alkuperää. AMOVA-ana-
lyysi (analysis of molecular variance) todettiin käyt-tökelpoiseksi menetelmäksi arvioitaessa näytteidentaksonomisten ja maantieteellisten ryhmien välisiä jasisäisiä geneettisen muuntelun osatekijöitä. Vaikkamolemmista aineiston ryhmittelyvaihtoehdoista (tak-sonominen ja maantieteellinen alkuperä) paljastuiryhmien välistä muuntelua, pääosa molekyylivarians-sista (arviolta 75 %) luettiin yhä ryhmien sisäisenmuuntelun aiheuttamaksi. Yhteenvetona toteamme,että RAPD analyysi on käyttökelpoinen menetelmätyrninäytteiden taksonomisen ja maantieteellisenalkuperän selvittämiseen.

290
A G R I C U L T U R A L A N D F O O D S C I E N C E I N F I N L A N D
Bartish, G.I. et al. RAPD diversity in sea buckthorn





























