Embed Size (px)
Citation preview

Scientia Horticulturae 101 (2004) 417–426
Genetic relationships among some cultivars of seabuckthorn from China, Russia and Mongolia based
on RAPD analysis
Chengjiang Ruana,b,∗, Pei Qina, Jianwei Zhenga, Zhenxiang Heaa Halophyte Research Laboratory of Nanjing University, 22 Hankou Road, Nanjing 210093, China
b Department of Marine, Yancheng Institute of Technology, Yancheng 224003, Jiangsu, China
Accepted 21 November 2003
Abstract
Sea buckthorn (Hippophae rhamnoides) is important environmentally and as a new commercialberry crop. Fourteen cultivars of sea buckthorn from China, Russia and Mongolia were evaluatedfor genetic relationships using RAPD data. Nine random primers were used to generate 114 RAPDmarkers by PCR. Among them, 103 (90.35%) were polymorphic. The genetic similarity ranged from0.45 to 0.80 with a mean of 0.67. Genetic distance ranged from 0.23 to 0.80 with a mean of 0.40.Cluster analysis identified Zhongguoshaji from China (H. r. subsp.sienses) as the most geneticallydistinct cultivar. Hongguo (H. r. subsp.sienses), Zhongguoyou (H. r. subsp.sienses), the hybridLiaohuerhao from China, Zeliang and Huoguang from Russia (H. r. subsp.mongolica) were clusteredmore distantly. The seven cultivars from Russia and Mongolia (H. r. subsp.mongolica) and thehybrid Liaohuyihao from China were grouped into two main clusters. Cluster I contained Chengsi,Liaohuyihao, Xiangyang, Chuyi and Wulangemu. Cluster II consists of Nuyou, Hongyun and Aleiyi.Data provide a scientific basis for selection and management of germplasm collections and crossingstrategies for sea buckthorn.© 2004 Elsevier B.V. All rights reserved.
Keywords: Hippophae rhamnoides; Similarity index; Genetic distance; Germplasm; Breeding
1. Introduction
Sea buckthorn (Hippophae rhamnoidesL.) is a deciduously perennial shrub or tree be-longing to the Elaeagnaceae family. There are 6 species and 12 subspecies in the world(Ruan and Xie, 2001). It is naturally distributed from Asia to Europe and was introduced to
∗ Corresponding author. Tel.:+86-25-3592684; fax:+86-25-3592684.E-mail address:[email protected] (C. Ruan).
0304-4238/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.scienta.2003.11.013
__________________________________________________________www.paper.edu.cn

418 C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426
South and North America (Ruan and Li, 1999). The distribution is from longitude 2–123◦Eand latitude 27–69◦N and from 0 to 3300 m a.s.l. Sea buckthorn has proven highly adaptableto extreme conditions, including temperatures ranging from−43 to 40◦C, drought, highaltitudes, salinity, alkalinity, and inundation (Ruan and Li, 2002). This broad geographicaldistribution and diverse growing conditions contribute to an extensive genetic diversity ofsea buckthorn.
Commercially sea buckthorn is a hardy, multi-purpose plant with orange, red or yellowberries. It has a strong ability to fix atmospheric nitrogen, and it rapidly develops an extensiveroot system and canopy, thus, quickly covering large areas of soil. These properties makeit an ideal candidate for soil and water conservation in extreme or marginal areas. Itsberries and leaves contain more than 100 different kinds of phytonutrients and bioactivesubstances including vitamins, fatty acids, free amino acids and minerals, and a range ofhealth products is being manufactured from sea buckthorn. Applications include preventionof cardiovascular diseases and cancer, as well as, treatment of skin problems, burns anddigestive tract disorders, and for anti-senilism, anti-inflammation and anti-radiation, andfor improvement of the capacity of the immune system. (Ruan and Li, 2000).
Randomly amplified polymorphic DNA (RAPD) markers have proven to be a reliablemethod for determining genetic relationships among germplasm collections. RAPD markershave the advantages of simplicity and the ability to detect relatively small amounts ofgenetic variation. The technique has already been successfully applied to estimate geneticrelationships such asAlstroemeriaL. (Dubouzet et al., 1998), cocoyam (Schnell et al., 1999),Cucumis meloL. (Silberstein et al., 1999; Garcia-Mas et al., 2000), Eucalyptus(Rossettoet al., 1999), Ficus caricaL. (Cabrita et al., 2001), Clivia (Ran et al., 2001) andCasuarina(Ho et al., 2002). Limited studies on the genetic relationships and diversity in sea buckthornhave been published.Yao and Tigerstedt (1993)using isoenzymes found a within populationgenetic diversity of 0.168. They also noted that the populations ofH. r. ssp.rhamnoideswere more diverse (as revealed by Nei’s genetic distance of 0.037) thanH. r. ssp.sinensis(0.007).Bartish et al. (1999)studied 10 populations ofH. r. ssp.rhamnoidesfrom northernEurope using randomly amplified polymorphic DNA (RAPD). In one population ofH. r.ssp.mongolicathey estimate that 85% of the genetic variation was within the populationswith 15% due to differences among the populations, and this result was supported by thefurther analysis using RAPD markers ofBartish et al. (2000). Within population variabilityof H. r. ssp.rhamnoideswas also found to be low (0.192 or 0.159). However, the geneticrelationships of the many cultivars from China, Russia and Mongolia remain unknown.
To characterize genetic relationships of 14 cultivars of sea buckthorn from China, Russiaand Mongolia, we screened the selected accessions using a RAPD protocol. Sea buckthornfrom China is adaptable and fast growing, but more prone to withered disease. Withereddisease is a major disease of sea buckthorn at present cause is unknown. Leaves becomeyellow and abscure. Lesions occur in the trunk or branches and are 5–10 cm in length. Plantsslow low vigor, and finally wither and die. Germplasm from Russia and Mongolia showsmany promising agronomic traits, including withered disease resistance. Crosses betweenthese germplasms may lead to improved varieties. In this study, we report on the usefulnessof RAPD markers for estimating genetic relationships in 14 cultivars of sea buckthorn fromChina, Russia and Mongolia. This data provides a scientific basis for future selection andmanagement of germplasm.
中国科技论文在线__________________________________________________________www.paper.edu.cn

C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426 419
2. Materials and methods
2.1. Plant materials
The 14 cultivars of sea buckthorn were obtained from China, Russia and Mongolia(Table 1). Annual seedlings of LHYH, LHEH and HG were introduced from Institute ofFuxing Sea Buckthorn, Liaoling Province in 1999. Seeds of ZGSJ and ZGY were collectedin the experimental field at Ansai County of Shaanxi Province in 1997 and sowed in 1998.WLGM seeds were donated by Professor Ochirtat Laagang of Mongolia Academy Scienceand sowed in 1999. Annual seedlings of eight cultivars from Russia were introduced fromthe Institute of Cytology and Geneties Siberan Branch, Novossibirsk, Russia in 1999. Plantswere maintained at the YCIT (Institute of Yancheng Technology) test garden, Yancheng,Jiangsu Province, China. Leaf samples of all cultivars were sent to the Nanjing Universityfor molecular markers analysis.
2.2. DNA extraction and quantification
DNA was isolated from each leaf sample. Samples were placed in a mortar with liquidnitrogen and ground to a fine power, and then transferred to a 1.5 ml microcentrifuge tube.To the tube, 800�l extraction buffer (100 mM Tris–HCl (pH= 8.0), 1% PVP–40,000,2% (w/v) CTAB, 25 mM EDTA (pH= 8.0), and 10 mM diethyldithio-carbamic acid) wasadded. The tube was then incubated at 65◦C for a total of 30 min, with one inversion ofthe tube at 15 min. The tube was centrifuged at 10,000× g for 10 min at 4◦C. To removeenzymes and any other contaminating protein, the pellet was rinsed twice with hydroxyben-zene/chloroform/isopentanol (25:24:41, v/v/v). The resulting DNA pellet was washed with75% ethanol, and then the isolated DNA was resuspended in TE buffer. Typically, 1�l ofleaf DNA was used in a PCR amplification reaction. The purity and concentration of DNAsamples were assessed by 1% agarose gel electrophoresis compared to known quantities ofcalf thymus DNA and by UV-spectrophotometry.
2.3. RAPD analysis
Primers used to generate RAPD profiles were as follows: OPB11, 17; OPC01, 02, 04,11; OPD03, 05, 13. PCR amplification reactions were carried out in 25�l final volume ofreaction mixture containing 50 mM KCl, 1.5 mM MgCl2, 10 mM Tris–HCl pH 9.0, 1 unitof TaqDNA polymerase (Pharmacia/Amersham), 0.24 mM of each dNTP (Life Technolo-gies), 0.4�M of primer (Operon Technologies) and 5 ng of genomic DNA. Amplificationswere performed to a Perkin-Elmer thermocycler (Gene Amp PCR System 2700) usingthe following program: an initial cycle of 180 s at 94◦C, followed by 45 cycles of 45 sat 94◦C, 45 s at 36◦C and 90 s at 72◦C. A final extension step was carried out at 72◦Cfor 7 min. The RAPDs produced were resolved in 2% agarose gels and visualized underUV-transillumination after ethidium bromide staining. Gels were recorded using a Kodakelectrophoresis documentation and analysis system 120 (EDAS).
中国科技论文在线__________________________________________________________www.paper.edu.cn
420 C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426
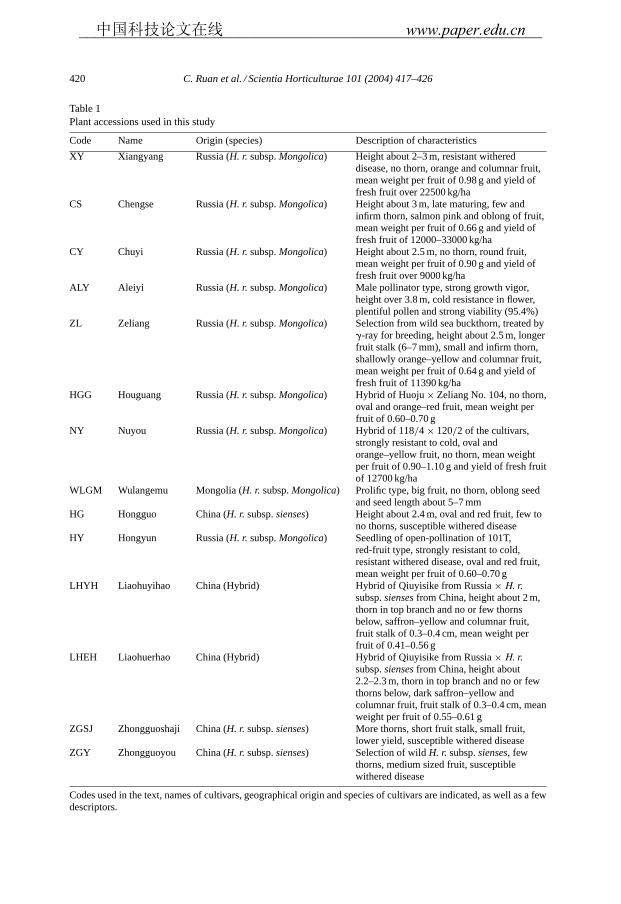
Table 1Plant accessions used in this study
Code Name Origin (species) Description of characteristics
XY Xiangyang Russia (H. r. subsp.Mongolica) Height about 2–3 m, resistant withereddisease, no thorn, orange and columnar fruit,mean weight per fruit of 0.98 g and yield offresh fruit over 22500 kg/ha
CS Chengse Russia (H. r. subsp.Mongolica) Height about 3 m, late maturing, few andinfirm thorn, salmon pink and oblong of fruit,mean weight per fruit of 0.66 g and yield offresh fruit of 12000–33000 kg/ha
CY Chuyi Russia (H. r. subsp.Mongolica) Height about 2.5 m, no thorn, round fruit,mean weight per fruit of 0.90 g and yield offresh fruit over 9000 kg/ha
ALY Aleiyi Russia (H. r. subsp.Mongolica) Male pollinator type, strong growth vigor,height over 3.8 m, cold resistance in flower,plentiful pollen and strong viability (95.4%)
ZL Zeliang Russia (H. r. subsp.Mongolica) Selection from wild sea buckthorn, treated by�-ray for breeding, height about 2.5 m, longerfruit stalk (6–7 mm), small and infirm thorn,shallowly orange–yellow and columnar fruit,mean weight per fruit of 0.64 g and yield offresh fruit of 11390 kg/ha
HGG Houguang Russia (H. r. subsp.Mongolica) Hybrid of Huoju× Zeliang No. 104, no thorn,oval and orange–red fruit, mean weight perfruit of 0.60–0.70 g
NY Nuyou Russia (H. r. subsp.Mongolica) Hybrid of 118/4 × 120/2 of the cultivars,strongly resistant to cold, oval andorange–yellow fruit, no thorn, mean weightper fruit of 0.90–1.10 g and yield of fresh fruitof 12700 kg/ha
WLGM Wulangemu Mongolia (H. r. subsp.Mongolica) Prolific type, big fruit, no thorn, oblong seedand seed length about 5–7 mm
HG Hongguo China (H. r. subsp.sienses) Height about 2.4 m, oval and red fruit, few tono thorns, susceptible withered disease
HY Hongyun Russia (H. r. subsp.Mongolica) Seedling of open-pollination of 101T,red-fruit type, strongly resistant to cold,resistant withered disease, oval and red fruit,mean weight per fruit of 0.60–0.70 g
LHYH Liaohuyihao China (Hybrid) Hybrid of Qiuyisike from Russia× H. r.subsp.siensesfrom China, height about 2 m,thorn in top branch and no or few thornsbelow, saffron–yellow and columnar fruit,fruit stalk of 0.3–0.4 cm, mean weight perfruit of 0.41–0.56 g
LHEH Liaohuerhao China (Hybrid) Hybrid of Qiuyisike from Russia× H. r.subsp.siensesfrom China, height about2.2–2.3 m, thorn in top branch and no or fewthorns below, dark saffron–yellow andcolumnar fruit, fruit stalk of 0.3–0.4 cm, meanweight per fruit of 0.55–0.61 g
ZGSJ Zhongguoshaji China (H. r. subsp.sienses) More thorns, short fruit stalk, small fruit,lower yield, susceptible withered disease
ZGY Zhongguoyou China (H. r. subsp.sienses) Selection of wildH. r. subsp.sienses, fewthorns, medium sized fruit, susceptiblewithered disease
Codes used in the text, names of cultivars, geographical origin and species of cultivars are indicated, as well as a fewdescriptors.
中国科技论文在线__________________________________________________________www.paper.edu.cn
C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426 421
2.4. Data analysis
The RAPD fragments were measured using GelReader V. 2.05 from the National Cen-ter for Supercomputing Applications at the University of Illinois at Urbana-Champaign(IL, USA). The RAPD fragments were scored as present (1) or absent (0) for each ofthe markers. Genetic similarity was calculated asS = 2mxy/(mx + my), whereS is thegenetic similarity,mxy the number of polymorphic bands shared betweenmx and my,mx the number of bands present inx and absent iny and my is the number of bandspresent iny and absent inx. The genetic distance was calculated according toNei andLi (1979). The genetic distance matrix was used in UPGMA cluster analysis using TF-PGA software. GelReader and TFPGA are available by anonymous FTP on theInternet.
3. Results and analysis
3.1. RAPD analysis
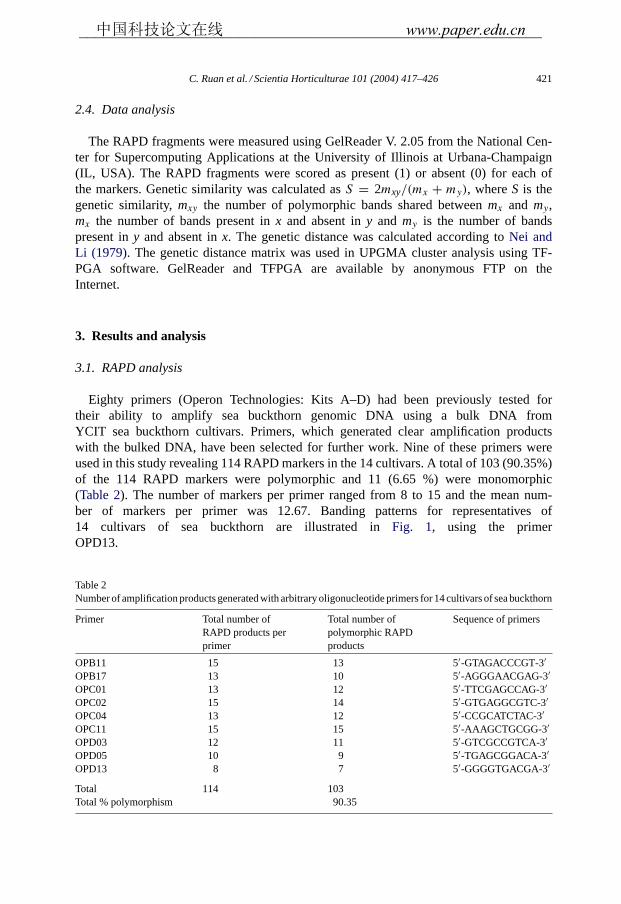
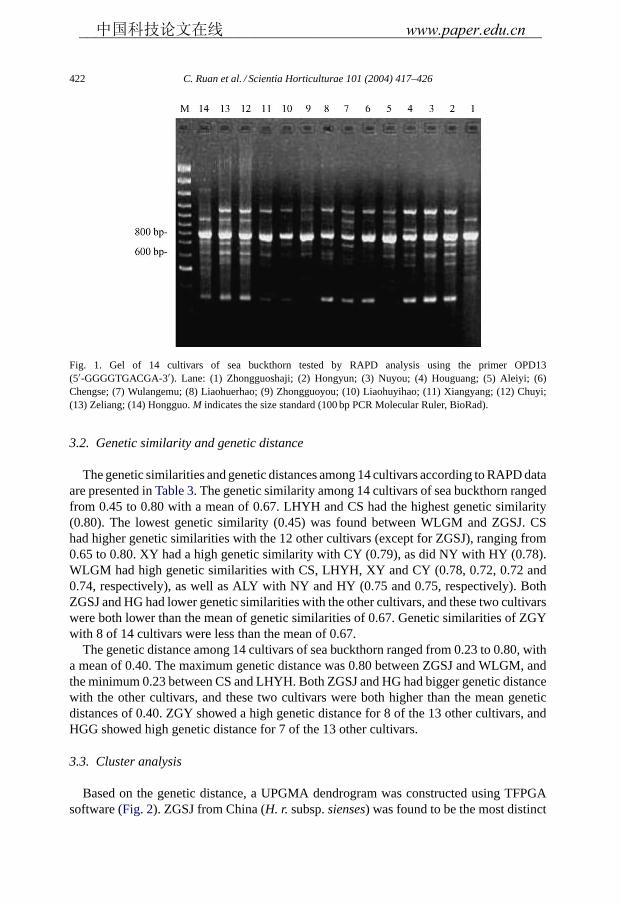
Eighty primers (Operon Technologies: Kits A–D) had been previously tested fortheir ability to amplify sea buckthorn genomic DNA using a bulk DNA fromYCIT sea buckthorn cultivars. Primers, which generated clear amplification productswith the bulked DNA, have been selected for further work. Nine of these primers wereused in this study revealing 114 RAPD markers in the 14 cultivars. A total of 103 (90.35%)of the 114 RAPD markers were polymorphic and 11 (6.65 %) were monomorphic(Table 2). The number of markers per primer ranged from 8 to 15 and the mean num-ber of markers per primer was 12.67. Banding patterns for representatives of14 cultivars of sea buckthorn are illustrated inFig. 1, using the primerOPD13.
Table 2Number of amplification products generated with arbitrary oligonucleotide primers for 14 cultivars of sea buckthorn
Primer Total number ofRAPD products perprimer
Total number ofpolymorphic RAPDproducts
Sequence of primers
OPB11 15 13 5′-GTAGACCCGT-3′OPB17 13 10 5′-AGGGAACGAG-3′OPC01 13 12 5′-TTCGAGCCAG-3′OPC02 15 14 5′-GTGAGGCGTC-3′OPC04 13 12 5′-CCGCATCTAC-3′OPC11 15 15 5′-AAAGCTGCGG-3′OPD03 12 11 5′-GTCGCCGTCA-3′OPD05 10 9 5′-TGAGCGGACA-3′OPD13 8 7 5′-GGGGTGACGA-3′
Total 114 103Total % polymorphism 90.35
中国科技论文在线__________________________________________________________www.paper.edu.cn
422 C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426
Fig. 1. Gel of 14 cultivars of sea buckthorn tested by RAPD analysis using the primer OPD13(5′-GGGGTGACGA-3′). Lane: (1) Zhongguoshaji; (2) Hongyun; (3) Nuyou; (4) Houguang; (5) Aleiyi; (6)Chengse; (7) Wulangemu; (8) Liaohuerhao; (9) Zhongguoyou; (10) Liaohuyihao; (11) Xiangyang; (12) Chuyi;(13) Zeliang; (14) Hongguo.M indicates the size standard (100 bp PCR Molecular Ruler, BioRad).
3.2. Genetic similarity and genetic distance
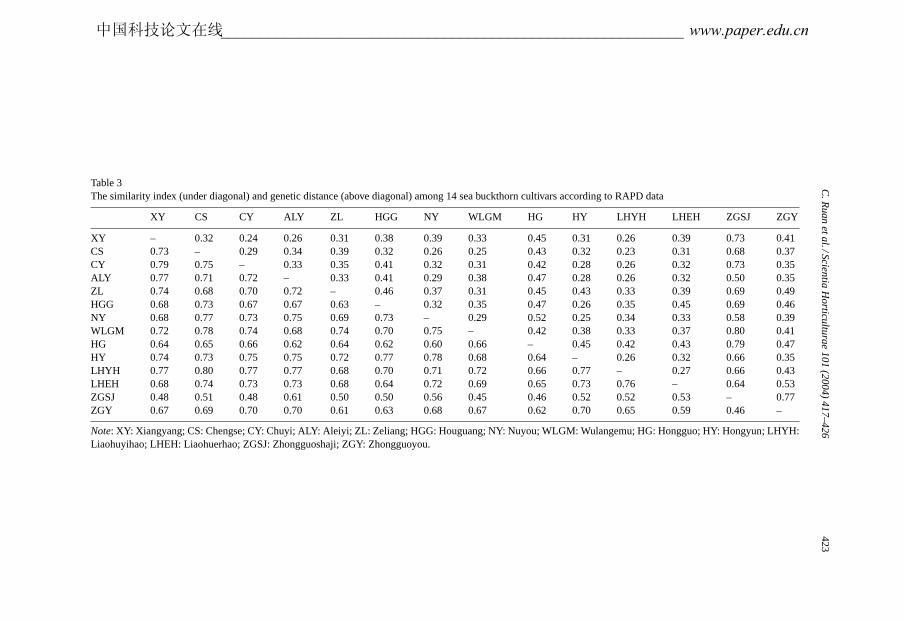
The genetic similarities and genetic distances among 14 cultivars according to RAPD dataare presented inTable 3. The genetic similarity among 14 cultivars of sea buckthorn rangedfrom 0.45 to 0.80 with a mean of 0.67. LHYH and CS had the highest genetic similarity(0.80). The lowest genetic similarity (0.45) was found between WLGM and ZGSJ. CShad higher genetic similarities with the 12 other cultivars (except for ZGSJ), ranging from0.65 to 0.80. XY had a high genetic similarity with CY (0.79), as did NY with HY (0.78).WLGM had high genetic similarities with CS, LHYH, XY and CY (0.78, 0.72, 0.72 and0.74, respectively), as well as ALY with NY and HY (0.75 and 0.75, respectively). BothZGSJ and HG had lower genetic similarities with the other cultivars, and these two cultivarswere both lower than the mean of genetic similarities of 0.67. Genetic similarities of ZGYwith 8 of 14 cultivars were less than the mean of 0.67.
The genetic distance among 14 cultivars of sea buckthorn ranged from 0.23 to 0.80, witha mean of 0.40. The maximum genetic distance was 0.80 between ZGSJ and WLGM, andthe minimum 0.23 between CS and LHYH. Both ZGSJ and HG had bigger genetic distancewith the other cultivars, and these two cultivars were both higher than the mean geneticdistances of 0.40. ZGY showed a high genetic distance for 8 of the 13 other cultivars, andHGG showed high genetic distance for 7 of the 13 other cultivars.
3.3. Cluster analysis
Based on the genetic distance, a UPGMA dendrogram was constructed using TFPGAsoftware (Fig. 2). ZGSJ from China (H. r. subsp.sienses) was found to be the most distinct
中国科技论文在线__________________________________________________________www.paper.edu.cn
C.R
ua
ne
tal./S
cien
tiaH
orticu
ltura
e1
01
(20
04
)4
17
–4
26
423
Table 3The similarity index (under diagonal) and genetic distance (above diagonal) among 14 sea buckthorn cultivars according to RAPD data
XY CS CY ALY ZL HGG NY WLGM HG HY LHYH LHEH ZGSJ ZGY
XY – 0.32 0.24 0.26 0.31 0.38 0.39 0.33 0.45 0.31 0.26 0.39 0.73 0.41CS 0.73 – 0.29 0.34 0.39 0.32 0.26 0.25 0.43 0.32 0.23 0.31 0.68 0.37CY 0.79 0.75 – 0.33 0.35 0.41 0.32 0.31 0.42 0.28 0.26 0.32 0.73 0.35ALY 0.77 0.71 0.72 – 0.33 0.41 0.29 0.38 0.47 0.28 0.26 0.32 0.50 0.35ZL 0.74 0.68 0.70 0.72 – 0.46 0.37 0.31 0.45 0.43 0.33 0.39 0.69 0.49HGG 0.68 0.73 0.67 0.67 0.63 – 0.32 0.35 0.47 0.26 0.35 0.45 0.69 0.46NY 0.68 0.77 0.73 0.75 0.69 0.73 – 0.29 0.52 0.25 0.34 0.33 0.58 0.39WLGM 0.72 0.78 0.74 0.68 0.74 0.70 0.75 – 0.42 0.38 0.33 0.37 0.80 0.41HG 0.64 0.65 0.66 0.62 0.64 0.62 0.60 0.66 – 0.45 0.42 0.43 0.79 0.47HY 0.74 0.73 0.75 0.75 0.72 0.77 0.78 0.68 0.64 – 0.26 0.32 0.66 0.35LHYH 0.77 0.80 0.77 0.77 0.68 0.70 0.71 0.72 0.66 0.77 – 0.27 0.66 0.43LHEH 0.68 0.74 0.73 0.73 0.68 0.64 0.72 0.69 0.65 0.73 0.76 – 0.64 0.53ZGSJ 0.48 0.51 0.48 0.61 0.50 0.50 0.56 0.45 0.46 0.52 0.52 0.53 – 0.77ZGY 0.67 0.69 0.70 0.70 0.61 0.63 0.68 0.67 0.62 0.70 0.65 0.59 0.46 –
Note: XY: Xiangyang; CS: Chengse; CY: Chuyi; ALY: Aleiyi; ZL: Zeliang; HGG: Houguang; NY: Nuyou; WLGM: Wulangemu; HG: Hongguo; HY: Hongyun; LHYH:Liaohuyihao; LHEH: Liaohuerhao; ZGSJ: Zhongguoshaji; ZGY: Zhongguoyou.
中国科技论文在线__________________________________________________________ www.paper.edu.cn

424 C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426
Fig. 2. UPGMA dendrogram estimating the genetic distance among 14 cultivars of sea buckthorn (H. rhamnoides)from China, Russia and Mongolia.
cultivar, with HG and ZGY from China (H. r. subsp.sienses) also clustered more distantly.The seven cultivars from Russia and Mongolia (H. r. subsp.mongolica) and LHYH of hy-brid from China were grouped into two main clusters, Cluster I contained CS, LHYH, XY,CY and WLGM, Cluster II consisted of NY, HY and ALY. Within Cluster I, two sub-groupswere resolved, one consisting of CS, LHYH, XY, CY and another only containing WLGM.Within the first sub-group, CS was closely related to LHYH, and the two grouped to-gether; XY was closely related to CY, and the two grouped together; within Cluster II,NY was closely related to HY, and the two formed a sub-group with ALY. Hybrid LHEHfrom China and ZL and HGG from Russia (H. r. subsp.mongolica) were clustered moredistantly.
4. Discussion
We observed 114 bands for the 9 primers used and estimated that 90.35% of the totalobserved bands were polymorphic. Genetic similarity ranged from 0.45 to 0.80 with a meanof 0.67, which indicated extensive genetic diversity. The genetic similarity values obtainedwith the RAPD markers were analogous with the results of the UPGMA dendrogram.
The genetic distance between ZGSJ and WLGM was the biggest (0.80), as was expectedsince they represent different subspecies. ZGY and HG were also selected from wildH. r.subsp.sienses, however, the RAPD results showed the two to be distinct from cultivatedZGSJ. This suggests that ZGY and HG may represent new species. Several cultivars fromRussia derived fromH. r. subsp.mongolicahad similar agronomic traits with WLGM andcommon origin. This may explain WLGM from Mongolia was grouped with CS, XY andCY.
中国科技论文在线__________________________________________________________www.paper.edu.cn

C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426 425
It is difficult to distinguish HG from HY by phenotypic characteristics, however, theRAPD results clearly show the two to be distinct. Though HGG was hybrid of Huoju×Zeliang No. 104 (selected from ZL), genetic distance between HGG and ZL was relativelylarge (0.4595), and the two were not grouped together.
While LHYH and LHEH have common origins (Qiuyesike× H. r. subsp.sienses) andmorphological characters, RAPD analysis grouped LHYH into Cluster I with CS ofH. r.subsp.mongolica, while LHEH was grouped in Cluster II. The two, however, originatedfrom different maternal plant. Female parents of LHYH were Nos. 15 and 18 of the cultivatedQiuyesike from Russia, while female parents of LHEH were Nos. 42 and 51.
The hybrid LHYH grouped into Cluster I with CS may be from matroclinal inheritance,or in the long term of artificial selection matroclinous variations were selected, For example,matroclinous characteristics were important selective traits, including big fruit, few or nothorns and low susceptability to withered disease.
RAPD analysis also has certain limitations. The low reproducibility of RAPDs introducesa problem for their use in cultivar identification. Moreover, it is worth noting that putativelysimilar bands originating from RAPDs in different individuals are not necessarily homolo-gous although they may share the same size in base pairs. This situation may lead to errorwhen calculating genetic relationships. However, the ease of RAPD analysis, which onlyrequires PCR technology, offers advantages for high throughput breeding programs.
In conclusion, results demonstrate the utility of RAPDs for detection of genetic rela-tionships between and within sea buckthorn cultivars from China, Russia and Mongolia.Resulting information can be used as guidelines for improved germplasm collection andbreeding. Resultant molecular markers can also be used for pre-selection in seedling pop-ulations to discard unfavorable genotypes at an earlier stage. Results reported here alsosupport relationships predicted by using traditional phenotypic evaluation.
Acknowledgements
This study was funded by the China–Russia Project No. BZ2001028 from Jiangsu Sci-entific and Technological Committee of China, partly supported by the Special Project ofSea Buckthorn No. 20010416 from the Chinese Ministry of Water Resources and partlysupported by the Young Natural Science Foundation of Institute of Yancheng TechnologyProject No. 2002XYK04. We thank Dr. ChangFang Zhou for technical help. We are verygrateful to Professor Daiqiong Li, for his constructive suggestions and important revisionof the manuscript.
References
Bartish, I.V., Jeppsson, N., Nybom, H., 1999. Population genetic structure in the dioecious pioneer plant speciesHippophae rhamnoidesinvestigated by random amplified polymorphic DNA (RAPD) markers. Mol. Ecol. 8,791–802.
Bartish, I.V., Jeppsson, N., Bartish, G.I., Lu, R., Nybom, H., 2000. Inter- and intra-specific genetic variation inHippophae (Elaeagnaceae) investigated by RAPD markers. Plant Syst. Evol. 225, 85–101.
中国科技论文在线__________________________________________________________www.paper.edu.cn

426 C. Ruan et al. / Scientia Horticulturae 101 (2004) 417–426
Cabrita, L.F., Alsoy, U., Hepaksoy, S., Leitao, J.M., 2001. Suitability of isozyme, RAPD and AFLP markers toassess genetic differences and relatedness among fig (Ficus caricaL.) clones. Sci. Hort. 87, 261–273.
Dubouzet, J.G., Murata, N., Shinoda, K., 1998. Relationships among some cultivated species and varieties ofAlstroemeriaL. based on RAPD analysis. Sci. Hort. 73, 37–44.
Garcia-Mas, J., Oliver, M., Gomez-Paniagua, H., de Vicente, M.C., 2000. Comparing AFLP, RAPD and RFLPmarkers for measuring genetic diversity in melon. Theor. Appl. Genet. 101, 860–864.
Ho, K.Y., Yang, J.C., Hsiao, J.Y., 2002. An assessment of genetic diversity and documentation of hybridization ofcasuarinas grown in Taiwan using RAPD markers. Int. J. Plant Sci. 163, 831–835.
Nei, N., Li, W., 1979. Mathematical model for studying genetic variation in term of restriction endonucleases.Proc. Natl. Acad. Sci. U.S.A. 76, 5269–5273.
Ran, Y., Murray, B.G., Hammett, K.R.W., 2001. Evaluating genetic relationships between and withinclivia speciesusing RAPDs. Sci. Hort. 90, 167–179.
Rossetto, M., Jezierski, G., Hopper, S.D., Dixon, K.W., 1999. Conservation genetics and clonality in two criticallyendangered eucalyptus from the highly endemic south-western Australian flora. Biol. Conserv. 88, 321–331.
Ruan, C.J., Li, D.Q., 1999. Study on indices of water physiology and drought resistance of sea buckthorn(Hippophae rhamnoides) in the Semiarid Loess Hilly Region of China. In: Proceedings of the InternationalSymposium on Sea Buckthorn. Forestry Press, Beijing, China, pp. 94–101.
Ruan, C.J., Li, D.Q., 2000. Function and benefits of sea buckthorn improving eco-environment of loess plateau.Environ. Prot. 5, 30–31 (in Chinese).
Ruan, C.J., Xie, Q.L., 2001. Preliminary report on sea buckthorn variety introduction at beach in Yancheng city.Hippophae 14, 15–19 (in Chinese).
Ruan, C.J., Li, D.Q., 2002. Analysis on the community characteristics ofHippophae rhamnoidesL. plantationand water and nutrition of woodland in Loess Hilly Region. J. Appl. Ecol. 13, 1061–1064 (in Chinese).
Schnell, R.J., Goenaga, R., Olano, C.T., 1999. Genetic similarities among cocoyam cultivars based on randomlyamplified polymorphic DNA (RAPD) analysis. Sci. Hort. 80, 267–276.
Silberstein, L., Kovalski, I., Huang, R.G., Anagnostou, K., Jahn, M., Perl-Treves, R., 1999. Molecular variationin melon (Cucumis meloL.) as revealed by RFLP and RAPD markers. Sci. Hort. 79, 101–111.
Yao, Y.M., Tigerstedt, P.M.A., 1993. Isozyme studies of genetic diversity and evolution in Hippophae. Gen. Resour.Crop Evol. 40, 153–164.
中国科技论文在线__________________________________________________________www.paper.edu.cn





























