Embed Size (px)
Citation preview

Mol Genet Genomics (2010) 283:289–303
DOI 10.1007/s00438-010-0513-zORIGINAL PAPER
AtfA bZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus nidulans
Anita Balázs · Imre Pócsi · Zsuzsanna Hamari · Éva Leiter · Tamás Emri · Márton Miskei · Judit Oláh · Viktória Tóth · Nikoletta Heged:s · Rolf A. Prade · Mónika Molnár · István Pócsi
Received: 20 March 2009 / Accepted: 18 January 2010 / Published online: 4 February 2010© Springer-Verlag 2010
Abstract The aim of the study was to demonstrate thatthe bZIP-type transcription factor AtfA regulates diVerenttypes of stress responses in Aspergillus nidulans similarlyto Atf1, the orthologous ‘all-purpose’ transcription factor ofSchizosaccharomyces pombe. Heterologous expression ofatfA in a S. pombe �atf1 mutant restored the osmotic stresstolerance of Wssion yeast in surface cultures to the samelevel as recorded in complementation studies with the atf1gene, and a partial complementation of the osmotic andoxidative-stress-sensitive phenotypes was also achieved insubmerged cultures. AtfA is therefore a true functionalortholog of Wssion yeast’s Atf1. As demonstrated by RT-PCR experiments, elements of both oxidative (e.g. catalase B)
and osmotic (e.g. glycerol-3-phosphate dehydrogenase B)stress defense systems were transcriptionally regulated byAtfA in a stress-type-speciWc manner. Deletion of atfAresulted in oxidative-stress-sensitive phenotypes while thehigh-osmolarity stress sensitivity of the fungus was notaVected signiWcantly. In A. nidulans, the glutathione/gluta-thione disulWde redox status of the cells as well as apoptoticcell death and autolysis seemed to be controlled by regula-tory elements other than AtfA. In conclusion, the orchestra-tions of stress responses in the aspergilli and in Wssion yeastshare several common features, but further studies areneeded to answer the important question of whether aWssion yeast-like core environmental stress response alsooperates in the euascomycete genus Aspergillus.
Keywords Stress signaling · Oxidative stress · Osmotic stress · Apoptosis · Autolysis · Auxotrophy · CESR
Introduction
In the aspergilli, SskB [mitogen activated protein kinasekinase kinase (MAPKKK)]—PbsB [mitogen activated pro-tein kinase kinase (MAPKK)]—HogA/SakA [mitogen acti-vated protein kinase (MAPK)] stress signaling plays apivotal role in the regulation of diVerent types of stressresponses, e.g. responses to osmotic (Han and Prade 2002;Kawasaki et al. 2002; Furukawa et al. 2005; Hagiwara et al.2007; Vargas-Pérez et al. 2007), oxidative (Kawasaki et al.2002; Furukawa et al. 2005; Hagiwara et al. 2007) and star-vation (Xue et al. 2004) stress. Although some upstreamelements of the SskB–HogA/SakA signaling pathwayincluding the phosphotransfer protein YpdA and theresponse regulator SskA have been identiWed and func-tionally characterized (Furukawa et al. 2005; Hagiwara
A. Balázs and Imre Pócsi contributed equally to this paper.
Communicated by J. Perez-Martin.
A. Balázs · Imre Pócsi · É. Leiter · T. Emri · V. Tóth · N. Heged:s · M. Molnár · István Pócsi (&)Department of Microbial Biotechnology and Cell Biology, Faculty of Science and Technology, University of Debrecen, P.O. Box 63, Debrecen 4010, Hungarye-mail: [email protected]; [email protected]
Z. Hamari (&)Department of Microbiology, Faculty of Science and Informatics, University of Szeged, Közép fasor 52, Szeged 6726, Hungarye-mail: [email protected]
M. Miskei · J. OláhDepartment of Horticulture and Plant Biotechnology, Faculty of Agricultural Science, University of Debrecen, Egyetem tér 1, Debrecen 4010, Hungary
R. A. PradeDepartment of Microbiology and Molecular Genetics, Oklahoma State University, 307 Life Sciences East, Stillwater, OK 74078, USA
123

290 Mol Genet Genomics (2010) 283:289–303
et al. 2007; Vargas-Pérez et al. 2007), no stress–responsetranscription factor downstream to HogA/SakA has beendescribed until now. Therefore, our main goal was to iden-tify and functionally characterize a transcriptional regulatorthat may transmit YpdA–SskA–SskB–PbsB–HogA/SakAstress signals to a wide array of stress response genes. Thecardinal importance of MAPK signaling in the coordinationof genome-wide fungal stress responses has been empha-sized by several authors (Gasch et al. 2000; Chen et al.2003, 2008; Gasch 2007; Miskei et al. 2009).
To Wnd a promising candidate for acting downstream ofHogA/SakA MAPK, we relied on a recent in silico recon-struction of yeast-like stress–response systems in the asper-gilli (Miskei et al. 2009). The success of the application ofyeast-based models to identify and annotate stress–response proteins in the genus Aspergillus led us to theconclusions that (1) the aspergilli are closer to the Wssionyeast Schizosaccharomyces pombe than to the buddingyeast Saccharomyces cerevisiae when the elements and theorganization of their stress–response systems are consid-ered and compared and, as a consequence, (2) A. nidulansSskB–PbsB–HogA/SakA signaling resembles the Wssionyeast’s Wak1/Win1 (MAPKKK)–Wis1 (MAPKK)–Sty1/Spc1 (MAPK) system, which is an ‘all-purpose’ signalingpathway transmitting various environmental stress signalsto core environmental stress response (CESR) genes (Chenet al. 2003, 2008; Gasch 2007; Miskei et al. 2009). It isimportant to note that, downstream to Sty1/Spc1 MAPK,environmental stress signals proceed via Atf1, a bZIP-typetranscriptional factor which primarily controls CESR genesin S. pombe (Shiozaki and Russell 1996; Wilkinson et al.1996; Chen et al. 2003), and that a putative ortholog ofS. pombe Atf1, called AtfA, has been annotated in A. nidulans(locus ID in Broad Institute Aspergillus nidulans databaseat http://www.broad.mit.edu/annotation/genome/: AN2911.3;expectation value for homology between A. nidulans AtfAand S. pombe Atf1 is E = 1 £ 10¡36, coming from BLASTPsearch performed in the NCBI protein database at http://www.blast.ncbi.nlm.nih.gov/Blast.cgi; Aguirre et al. 2005;Hagiwara et al. 2008; Miskei et al. 2009).
The Wrst functional analysis of AtfA was performed byHagiwara et al. (2008), who generated and phenotypicallycharacterized a �atfA deletion mutant in A. nidulans. Theseauthors found that AtfA—similar to SskA response regula-tor and HogA/SakA MAPK (Kawasaki et al. 2002; Hagiwaraet al. 2007; Vargas-Pérez et al. 2007)—was required for theformation of conidia with appropriate tolerance of oxidativeand heat stress. On the other hand, they did not observe anystress-sensitive phenotypes when the fungus was subjectedto diVerent types of high-osmolarity and oxidative stress insurface cultures (Hagiwara et al. 2008).
In this study, the atfA gene of A. nidulans was deleted,the oxidative stress (caused by tert-butylhydroperoxide,
H2O2 or menadione sodium bisulWte) sensitive phenotypesof a group of �atfA mutants were described, and a set ofoxidative and osmotic-stress-responsive genes regulated byAtfA was identiWed. Moreover, the osmotic and oxidativestress sensitivities of a �atf1 mutant Wssion yeast strain(Takeda et al. 1995) were successfully complemented withthe heterologous expression of AtfA. The possible roles ofAtfA in the regulation of important and complex physiolog-ical processes such as environmental stress responses,apoptotic cell death and autolysis are also discussed here.
Materials and methods
Strains and culture media
The genotypes and origin of the A. nidulans and S. pombestrains are summarized in Table 1. For cultivation of theA. nidulans strains, standard complete medium and mini-mal nitrate medium (MNM) were used with appropriatenutritional supplements (Barratt et al. 1965; Cove 1966;Scazzocchio et al. 1982; http://www.gla.ac.uk/ibls/molgen/aspergillus/supplement.html). S. pombe strains were grownon minimal medium agar (MMA; 1% glucose, 0.67% YeastNitrogen Base Difco™, 2% agar) and Edinburgh minimalmedium 2 (EMM2, US Biological) plates or liquid cultures.When required, EMM2 was supplemented with 100 mg l¡1
leucine. Unless otherwise indicated, S. pombe and A. nidu-lans cultures were incubated at 30 and 37°C and wereshaken at 2.5 and 3.3 Hz frequencies, respectively.
Complementation of the osmotic and oxidative-stress-sensitive phenotypes of �atfA mutant S. pombe cells with atf1 and atfA
To isolate RNA from A. nidulans, the FGSC 26 strain wascultured overnight in liquid complete medium (Cove 1966).Mycelia were washed and transferred into MNM (Barrattet al. 1965), and supplemented with 0.8 mmol l¡1 menadi-one sodium bisulWte to induce the transcription of oxida-tive-stress-responsive genes (Pócsi et al. 2005). Afterharvesting mycelia at 5 h incubation time, RNA wasextracted with TRIZOL® Reagent (Invitrogen), atfA mRNAwas reverse-transcribed with the SUPERSCRIPT™ II Kit(Invitrogen), and atfA cDNA was PCR-ampliWed with theprimers shown in Table 2. The cDNA was cloned intopGEM® T Easy vector (Promega), and its sequence wasveriWed by sequencing both strands. After digestions withNdeI restriction enzyme (Fermentas Ltd., Vilnius, Lithua-nia), the atfA cDNA was re-cloned into the S. pombeexpression vectors pREP1, pREP41, and pREP81 (Maund-rell 1993; Basi et al. 1993). The NT146 �atf1 strain wastransformed using the lithium-acetate method of Ito et al.
123

Mol Genet Genomics (2010) 283:289–303 291
(1983) with the following plasmids: pREP41-atf1 (positivecontrol carrying the Wssion yeast atf1 gene; a kind gift fromDr. T. Toda, Cancer Research, UK), pREP41 (negativecontrol), and the plasmids carrying the A. nidulans atfAgene (pREP1-atfA, pREP41-atfA, and pREP81-atfA).Transformed cells were plated on MMA.
To examine the osmotic stress sensitivity on plates,serial dilutions of the transformed and control cultures pre-grown in liquid EMM2 overnight were pipetted ontoEMM2 agar plates supplemented with 1.1 mol l¡1 KCl or0.4 mol l¡1 NaCl, and were grown for 5 days. When theosmotic stress sensitivity was tested in liquid culture,EMM2 containing either 1.0 mol l¡1 KCl or 0.5 mol l¡1
NaCl was inoculated with S. pombe cells at a cell density ofOD630 t 0.10. Propagation of the cells was monitoredspectrophotometrically at � = 630 nm with an EL 340Microplate Bio Kinetics Reader (Bio-Tek™ Instruments) at2 h intervals. EMM2-cultured S. pombe cells (OD630 = 0.10–0.15) were also exposed to oxidative stress by supplement-ing the liquid culture media with 30% (v/v) H2O2 at a Wnalconcentration of 2.0 mmol l¡1. After 1 h incubation, Wssionyeast cells were washed with water, and were re-suspendedin H2O2-free EMM2. When cells recovered from the oxidative
stress (t13 h), their propagation was monitored by measuringOD630 at 2 h intervals.
Construction and stress sensitivity of A. nidulans atfA deletion mutants
The atfA gene was replaced with the riboB marker gene inthe HZS 120 atfA+ strain. The substitution cassette wasconstructed by double-joint PCR (Yu et al. 2004; primersfor the deletion construct are summarized in Table 2; thetargeting sequences for homologous recombination were1,325 and 2,451-bp-long upstream and downstream to theatfA gene, respectively), and was transformed directly inthe HZS 120 recipient strain (Tilburn et al. 1983). Selectedtransformants were pre-screened for the absence of the atfAgene product by PCR (Table 2), and transformants withoutamplicon were also checked by genomic DNA Southern blotanalysis. Three single-copy transformants (genotype pabaA1riboB2 �atfA::riboB) characterized with light green conidiawere crossed with the CS 2902 strain, and a set of �atfAprogenies with diVerent auxotrophy markers was collected.All riboB+ progenies of the independent crosses proved tobe atfA deleted mutants by Southern analyses.
Table 1 Genotypic characterization and origin of the S. pombe and A. nidulans strains used in this study
a All A. nidulans strains used in this study carried the veA1 mutation. leu1, ura4, pyrG89, pabaA1, biA1, pyroA1, and riboB2 are leucine, uracil,uracil, p-amino-benzoic acid (PABA), biotin, pyridoxine, and riboXavin auxotroph mutations, respectively. Chromosome localization of the abovelisted genes can be found at websites http://www.broadinstitute.org/annotation/genome/aspergillus_group/Home.html or http://www.gla.ac.uk/ibls/molgen/aspergillus/index.html for A. nidulans and http://old.genedb.org/genedb/pombe/ for S. pombeb veA¡ genotype was checked by incubating the strains on MNM agar for 3 days at 37°C in complete darkness similarly to the procedure of Moo-ney and Yager (1990), and veA1 mutation in the initiation codon of the VeA ORF was also veriWed according to Kim et al. (2002)
Strain Genotypea Origin/reference
S. pombe strains
NT146 strain h¡ leu1-32 ura4-D18 atf1::ura4 T. Toda (Cancer Research UK); Takeda et al. (1995)
HM123 h¡ leu1-32 Takeda et al. (1995)
A. nidulans strains
CS 2902 pyrG89 biA1 pyroA4 riboB2 Yu et al. (2004)
FGSC 26 biA1 S. Rosén (University of Lund)
G1071b J. Clutterbuck (University of Glasgow)
HZS 120 pabaA1 riboB2 Strain Collection of the Department of Microbiology, University of Szeged
HZS 145b Strain Collection of the Department of Microbiology, University of Szeged
HZS 187 pabaA1 biA1 riboB2 �atfA::riboB This study
HZS 188 biA1 riboB2 �atfA::riboB This study
HZS 189 pabaA1 riboB2 �atfA::riboB This study
HZS 190, LE 1–24 riboB2 �atfA::riboB This study
HZS 191 pyrG89 biA1 riboB2 �atfA::riboB This study
HZS 192 pyrG89 biA1 pyroA4 riboB2 �atfA::riboB This study
HZS 193 pyrG89 pabaA1 pyroA4 riboB2 �atfA::riboB This study
HZS 194 pabaA1 pyroA4 riboB2 �atfA::riboB This study
123

292 Mol Genet Genomics (2010) 283:289–303
In stress sensitivity studies, the agar plate assays of Hag-iwara et al. (2007, 2008) were used with minor modiWca-tions. Freshly grown (7 days) conidia (105 suspended in 5 �laliquots of PBS–0.1% Tween 20) were spotted on MNMplates (Barratt et al. 1965) containing the following stress-generating agents: oxidative stress: 0.6–0.8 mmol l¡1 tert-butylhydroperoxide (tBOOH; stimulates lipid peroxidation),6.0 mmol l¡1 H2O2 (increases the peroxide content ofthe cells), 0.12 mmol l¡1 menadione sodium bisulphite(MSB; elevates intracellular superoxide concentrations),2.0 mmol l¡1 diamide [disturbs the glutathione status of thecells via instantaneous and stoichiometrical oxidation of glu-tathione (GSH) to glutathione disulphide (GSSG)], osmoticstress: 1.5 mol l¡1 KCl, 1.5 mol l¡1 NaCl, 2.0 mol l¡1 sorbi-tol. Stress plates were incubated for 5 days, and the osmoticstress tolerances of selected prototroph strains were tested atboth 30 and 37°C temperatures (Han and Prade 2002).
To test the tBOOH-sensitivity of A. nidulans mycelia onagar plates, freshly-harvested conidia were pipetted ontocircle-shaped, 2-cm-diameter cellophane sheets laid onthe surfaces of MNM agar plates and were incubatedfor 24 h. Mycelial mats were transferred with the sheetsonto freshly prepared MNM agar also supplemented with0–3.0 mmol l¡1 tBOOH.
The oxidative stress sensitivity of �atfA mutants wasalso studied in submerged cultures. Late exponentialgrowth phase vegetative tissues of selected mutant and con-trol strains were transferred into fresh MNM and were cul-tivated further for 30 h according to the protocol of Pócsiet al. (2005). The starting dry cell mass (DCM) was set toapproximately 4 mg ml¡1 for all strains, and culture mediawere either supplemented with 75–300 mmol l¡1 H2O2,
0.38–2.0 mmol l¡1 tBOOH, 0.4–0.8 mmol l¡1 MSB or3.0 mmol l¡1 diamide (stress-exposed cultures) or leftuntreated (controls).
The oxidative, heat, and cold stress sensitivities of A.nidulans conidia were estimated according to Hagiwaraet al. (2007, 2008). Freshly grown conidia (105 ml¡1) weretreated with 100 and 200 mmol l¡1 H2O2 at 37°C for20 min, were subjected to heat stress at 50°C for 30 and60 min or were kept at 4°C for 7 days in glucose-freeMNM. Stress-exposed conidia were spread on nutrient agarplates, and the numbers of colonies were counted after2 days incubation at 37°C. The conidiospore forming capa-bilities of A. nidulans strains were determined as publishedpreviously by Vargas-Pérez et al. (2007).
Cell death and autolysis of the HZS 190 �atfA mutant
The small molecular mass antifungal protein produced byPenicillium chrysogenum (PAF) causes apoptosis-like celldeath in sensitive Wlamentous fungi like A. nidulans (Leiteret al. 2005; Marx et al. 2008). The PAF sensitivity of theA. nidulans strains was examined by the microplate assayof Kaiserer et al. (2003) and the agar diVusion test ofBinder et al. (2010).
To monitor progressing apoptosis in carbon-depletedsubmerged cultures, phosphatidylserine externalization onprotoplasts was determined by Annexin V assay at 48 and72 h incubation times as described elsewhere (Leiter et al.2005; Pócsi et al. 2009). Concomitant autolysis was charac-terized with DCM and cell vitality declinations as well aswith extracellular chitinase and proteinase productions(Emri et al. 2004, 2005 and Pócsi et al. 2009).
Table 2 PCR primers used in the genetic manipulations
Primers Sequences
Reverse transcription of atfA mRNA
atfA RT 5�-cgccagttagtcaagtgatattattcc-3�
AmpliWcation of atfA cDNAa
atfA amp frw 5�-cgcatatgtctgccgccgtggcttc-3�
atfA amp rev 5�-ggcatatgtcaagtgtatggaggatt-3�
Double-joint PCR
atfA upstream frw 5�-gatagaaacccgcagaacgaagagg-3�
atfA upstream nested frw 5�-gagtattggctaaactgcagtgcggcaag-3�
atfA upstream rev 5�-ccggtcgataaatcagctgaataatagag-3�
atfA riboB chimera frw 5�-ctctattattcagctgatttatcgaccggacgtagtgtagattcaggcacattgaagcg-3�
atfA riboB chimera rev 5�-acagaaaccgttgcgccagttagtcaagtggccatgactactaggtggtgctatc-3�
atfA downstream frw 5�-cacttgactaactggcgcaacggtttctgt-3�
atfA downstream nested rev 5�-cctatgtagaatctcagaaactgtcttc-3�
atfA downstream rev 5�-agctggacatagtgtttgacctgcacg-3�
Screening the absence of atfA in transformants
frw 5�-gtactgacttgatgatcttccagaggc-3�
rev 5�-gtctgttagtatactgataaattagagg-3�a The primers included NdeI restriction sites (italics)
123

Mol Genet Genomics (2010) 283:289–303 293
IdentiWcation of stress-responsive AtfA-target genes using real-time polymerase chain reaction (RT-PCR) assays
Late exponential growth phase mutant and control strainmycelia (Pócsi et al. 2005) were exposed to stress-generat-ing agents for 0.5 h according to Asano et al. (2007) andHagiwara et al. (2008). The selected agents and concentra-tions were 75 mmol l¡1 H2O2, 1.0 mmol l¡1 tBOOH and0.6 mol l¡1 NaCl. In a separate set of experiments, oxida-tive stress was also generated by treating mycelia with5.0 mmol l¡1 H2O2, 0.8 mmol l¡1 tBOOH or 60 �mol l¡1
MSB (Asano et al. 2007). Total RNA was extracted asdescribed elsewhere (Pócsi et al. 2005), and RT-PCRassays were optimized and performed according toPusztahelyi et al. (2006) using QuantiTect™SYBR®GreenRT-PCR Kit (Qiagen, Germany). The putative stress-responsive genes tested in these assays are summarized inTable 3 together with the primer pairs used to amplify speciWcgene transcripts. In each RNA sample, 18S rRNA transcriptswere also quantiWed as reference gene transcripts using the5�-ttctgccctatcaact-3� (frw) and 5�-ggctgaaacttaaaggaattg-3�
(rev) primers (Kato et al. 2003). Normalized relative tran-script levels were estimated by the ‘delta–delta method’and quantiWed with ¡��CP = ¡[�CPsample(target ¡reference) ¡ �CPcontrol(target ¡ reference)] = �CPtarget(control ¡ sample) ¡ �CPreference(control ¡ sample) RT-PCRcycle number of crossing point diVerences, where CP val-ues stand for the cycle numbers of crossing points recordedfor the tested target gene (CPtarget) and for the reference 18SrRNA (CPreference), respectively, either in stress-treatedsamples (CPsample) or in non-stressed controls (CPcontrol)(PfaZ 2001). This model presumes that RT ampliWcationeYciencies (E values) are optimal and identical(Etarget = Ereference = 2) for both the target and the reference(18S rRNA) genes and the gene expression ratio of the tar-get gene in the sample and the control is 2¡��CP (PfaZ2001).
Data statistics
The variations between experiments were estimated bystandard deviations (SDs) and the statistical signiWcance ofchanges in the parameters studied was estimated by the Stu-dent’s t-test. Only changes with probability levels ofP · 5% were regarded as statistically signiWcant (Pócsiet al. 2005; Pusztahelyi et al. 2006). In RT-PCR measure-ments, the SD values of CPs determined for each gene andculture conditions tested were always within the range of0.6–1.5 cycles, which were comparable to those found pre-viously for other A. nidulans genes (Emri et al. 2006; Mol-nár et al. 2006; Pusztahelyi et al. 2006). SDs for ¡��CPvalues were calculated following the rules for the linearcombination of measurements (Barford 1967). In statistical
signiWcance calculations, ¡��CP (mean § SD) valueswere compared to ¡��CP values calculated for unstressedcontrol cultures alone with the simpliWcations �CPtarget =0 § SD and �CPreference = 0 § SD (Pusztahelyi et al. 2006).
Results
The bZIP-type transcription factor AtfA of A. nidulans isconsidered as a putative ortholog of Wssion yeast’s ‘all-pur-pose’ transcription factor Atf1 and, hence, may regulate abroad spectrum of stress responses (Miskei et al. 2009). Totest this hypothesis, numerous genetic, physiological, andgene expression experiments were carried out and someresults heavily supported the hypothesized stress responseregulatory function of AtfA.
Aspergillus nidulans’s AtfA is a true functional ortholog of S. pombe’s Atf1
Functional characterization of AtfA was Wrst performed inWssion yeast by heterologously expressing AtfA in theS. pombe NT146 �atf1 deletion strain. A. nidulans atfAcDNA was isolated with primers designed to the predictedcDNA sequence of XM_655423 (http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nuccore&id=67524906; Galaganet al. 2005). The sequence of the cDNA fully matched theXM_655423 sequence and was submitted to the NCBI(accession number: EU877709; http://www.ncbi.nlm.nih.gov/). The isolated atfA cDNA was cloned subsequentlyinto pREP expression vectors, which provide diVerentexpression levels in Wssion yeast (Maundrell 1993; Basiet al. 1993). Overexpression of Atf1 under the wild-typenmt1 promoter (pREP1 plasmid) in S. pombe is lethal(Takeda et al. 1995). Similarly, expression of the pREP1-atfA construct in Wssion yeast proved to be deleterious (datanot shown). Therefore, moderate expression of AtfA wasachieved by transforming the �atf1 S. pombe strain withthe pREP41-atfA construct. Moderate expression of thenative protein (pREP41-atf1 construct) was applied as apositive control.
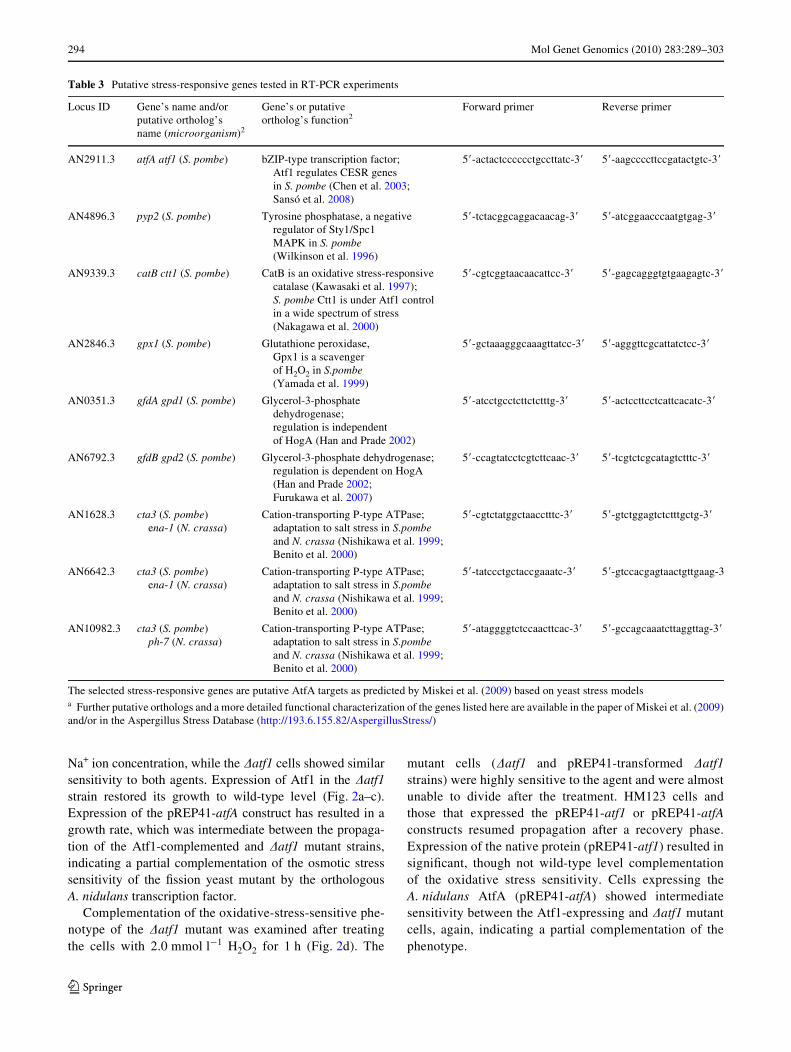
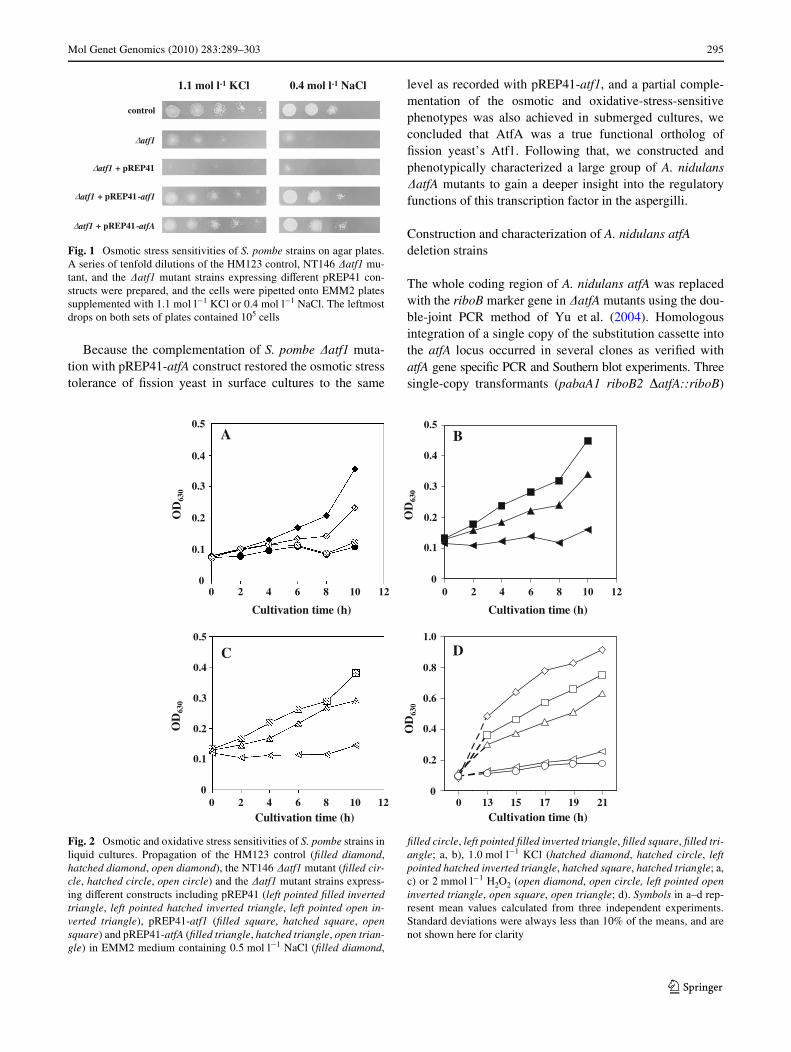
Schizosaccharomyces pombe �atf1 mutant cells werehighly sensitive to the osmotic stress provoked by high con-centrations of K+ or Na+ ions. Complementation of thisstress sensitivity by AtfA was tested Wrst on agar plates.Moderate expression of AtfA complemented the osmoticstress sensitivity of the �atf1 mutant, approximately to thesame level as the expression of the native transcription fac-tor Atf1 (Fig. 1). Following that the osmotic stress sensitiv-ity of the strains was also tested by monitoring theirpropagation in liquid cultures. As shown in Fig. 2a, HM123control cultures were more sensitive to the selected1.0 mol l¡1 concentration of K+ ions than to 0.5 mol l¡1
123
294 Mol Genet Genomics (2010) 283:289–303
Na+ ion concentration, while the �atf1 cells showed similarsensitivity to both agents. Expression of Atf1 in the �atf1strain restored its growth to wild-type level (Fig. 2a–c).Expression of the pREP41-atfA construct has resulted in agrowth rate, which was intermediate between the propaga-tion of the Atf1-complemented and �atf1 mutant strains,indicating a partial complementation of the osmotic stresssensitivity of the Wssion yeast mutant by the orthologousA. nidulans transcription factor.
Complementation of the oxidative-stress-sensitive phe-notype of the �atf1 mutant was examined after treatingthe cells with 2.0 mmol l¡1 H2O2 for 1 h (Fig. 2d). The
mutant cells (�atf1 and pREP41-transformed �atf1strains) were highly sensitive to the agent and were almostunable to divide after the treatment. HM123 cells andthose that expressed the pREP41-atf1 or pREP41-atfAconstructs resumed propagation after a recovery phase.Expression of the native protein (pREP41-atf1) resulted insigniWcant, though not wild-type level complementationof the oxidative stress sensitivity. Cells expressing theA. nidulans AtfA (pREP41-atfA) showed intermediatesensitivity between the Atf1-expressing and �atf1 mutantcells, again, indicating a partial complementation of thephenotype.
Table 3 Putative stress-responsive genes tested in RT-PCR experiments
The selected stress-responsive genes are putative AtfA targets as predicted by Miskei et al. (2009) based on yeast stress modelsa Further putative orthologs and a more detailed functional characterization of the genes listed here are available in the paper of Miskei et al. (2009)and/or in the Aspergillus Stress Database (http://193.6.155.82/AspergillusStress/)
Locus ID Gene’s name and/or putative ortholog’s name (microorganism)2
Gene’s or putative ortholog’s function2
Forward primer Reverse primer
AN2911.3 atfA atf1 (S. pombe) bZIP-type transcription factor; Atf1 regulates CESR genes in S. pombe (Chen et al. 2003; Sansó et al. 2008)
5�-actactcccccctgccttatc-3� 5�-aagccccttccgatactgtc-3�
AN4896.3 pyp2 (S. pombe) Tyrosine phosphatase, a negative regulator of Sty1/Spc1 MAPK in S. pombe (Wilkinson et al. 1996)
5�-tctacggcaggacaacag-3� 5�-atcggaacccaatgtgag-3�
AN9339.3 catB ctt1 (S. pombe) CatB is an oxidative stress-responsive catalase (Kawasaki et al. 1997); S. pombe Ctt1 is under Atf1 control in a wide spectrum of stress (Nakagawa et al. 2000)
5�-cgtcggtaacaacattcc-3� 5�-gagcagggtgtgaagagtc-3�
AN2846.3 gpx1 (S. pombe) Glutathione peroxidase, Gpx1 is a scavenger of H2O2 in S.pombe (Yamada et al. 1999)
5�-gctaaagggcaaagttatcc-3� 5�-agggttcgcattatctcc-3�
AN0351.3 gfdA gpd1 (S. pombe) Glycerol-3-phosphate dehydrogenase; regulation is independent of HogA (Han and Prade 2002)
5�-atcctgcctcttctctttg-3� 5�-actccttcctcattcacatc-3�
AN6792.3 gfdB gpd2 (S. pombe) Glycerol-3-phosphate dehydrogenase; regulation is dependent on HogA (Han and Prade 2002; Furukawa et al. 2007)
5�-ccagtatcctcgtcttcaac-3� 5�-tcgtctcgcatagtctttc-3�
AN1628.3 cta3 (S. pombe) ena-1 (N. crassa)
Cation-transporting P-type ATPase; adaptation to salt stress in S.pombe and N. crassa (Nishikawa et al. 1999; Benito et al. 2000)
5�-cgtctatggctaacctttc-3� 5�-gtctggagtctctttgctg-3�
AN6642.3 cta3 (S. pombe) ena-1 (N. crassa)
Cation-transporting P-type ATPase; adaptation to salt stress in S.pombe and N. crassa (Nishikawa et al. 1999; Benito et al. 2000)
5�-tatccctgctaccgaaatc-3� 5�-gtccacgagtaactgttgaag-3
AN10982.3 cta3 (S. pombe) ph-7 (N. crassa)
Cation-transporting P-type ATPase; adaptation to salt stress in S.pombe and N. crassa (Nishikawa et al. 1999; Benito et al. 2000)
5�-ataggggtctccaacttcac-3� 5�-gccagcaaatcttaggttag-3�
123
Mol Genet Genomics (2010) 283:289–303 295
Because the complementation of S. pombe �atf1 muta-tion with pREP41-atfA construct restored the osmotic stresstolerance of Wssion yeast in surface cultures to the same
level as recorded with pREP41-atf1, and a partial comple-mentation of the osmotic and oxidative-stress-sensitivephenotypes was also achieved in submerged cultures, weconcluded that AtfA was a true functional ortholog ofWssion yeast’s Atf1. Following that, we constructed andphenotypically characterized a large group of A. nidulans�atfA mutants to gain a deeper insight into the regulatoryfunctions of this transcription factor in the aspergilli.
Construction and characterization of A. nidulans atfA deletion strains
The whole coding region of A. nidulans atfA was replacedwith the riboB marker gene in �atfA mutants using the dou-ble-joint PCR method of Yu et al. (2004). Homologousintegration of a single copy of the substitution cassette intothe atfA locus occurred in several clones as veriWed withatfA gene speciWc PCR and Southern blot experiments. Threesingle-copy transformants (pabaA1 riboB2 �atfA::riboB)
Fig. 1 Osmotic stress sensitivities of S. pombe strains on agar plates.A series of tenfold dilutions of the HM123 control, NT146 �atf1 mu-tant, and the �atf1 mutant strains expressing diVerent pREP41 con-structs were prepared, and the cells were pipetted onto EMM2 platessupplemented with 1.1 mol l¡1 KCl or 0.4 mol l¡1 NaCl. The leftmostdrops on both sets of plates contained 105 cells
control
∆atf1
∆atf1 + pREP41
∆atf1 + pREP41-atf1
∆atf1 + pREP41-atfA
1.1 mol l-1 KCl 0.4 mol l-1 NaCl
Fig. 2 Osmotic and oxidative stress sensitivities of S. pombe strains inliquid cultures. Propagation of the HM123 control (Wlled diamond,hatched diamond, open diamond), the NT146 �atf1 mutant (Wlled cir-cle, hatched circle, open circle) and the �atf1 mutant strains express-ing diVerent constructs including pREP41 (left pointed Wlled invertedtriangle, left pointed hatched inverted triangle, left pointed open in-verted triangle), pREP41-atf1 (Wlled square, hatched square, opensquare) and pREP41-atfA (Wlled triangle, hatched triangle, open trian-gle) in EMM2 medium containing 0.5 mol l¡1 NaCl (Wlled diamond,
Wlled circle, left pointed Wlled inverted triangle, Wlled square, Wlled tri-angle; a, b), 1.0 mol l¡1 KCl (hatched diamond, hatched circle, leftpointed hatched inverted triangle, hatched square, hatched triangle; a,c) or 2 mmol l¡1 H2O2 (open diamond, open circle, left pointed openinverted triangle, open square, open triangle; d). Symbols in a–d rep-resent mean values calculated from three independent experiments.Standard deviations were always less than 10% of the means, and arenot shown here for clarity
0 2 4 6 8 10 12Cultivation time (h)
0.5
0.4
0.3
0.2
0.1
0
C
OD
630
Cultivation time (h)
0 2 4 6 8 10 12
0.5
0.4
0.3
0.2
0.1
0
A
OD
630
0 2 4 6 8 10 12
Cultivation time (h)
0.5
0.4
0.3
0.2
0.1
0
B
OD
630
Cultivation time (h)0 13 15 17 19 21
1.0
0.8
0.6
0.4
0.2
0
D
OD
630
123

296 Mol Genet Genomics (2010) 283:289–303
were crossed to the CS 2902 strain, and proper deletionof atfA was conWrmed again in several progenies withor without auxotrophy markers. All mutants grew well onMNM agar plates under unstressed conditions with com-parable growth rates to those of the control (G1071, HZS145, FGSC 26, HZS 120, and CS 2902) strains (data notshown).
Similar to the Wndings of Hagiwara et al. (2008), therewas no signiWcant diVerence in the KCl (1.5 mol l¡1) sensi-tivities of the tested mutants [7 prototroph (HZS 190, LE 1–6) and 7 auxotroph (HZS 187–189 and 191–194) �atfAprogenies were tested, and 55–69% growth inhibitions wererecorded] and the control strains (strains G 1071, HZS 145,FGSC 26, HZS 120 and CS 2902 with 48–65% growthreductions). Furthermore, NaCl (1.5 mol l¡1) and sorbitol(2 mol l¡1) also inhibited the growths of selected mutants(HZS 188, HZS 190; 62–67% reductions in growth withNaCl and 35–38% with sorbitol) and the control strains(56–65% growth inhibitions with NaCl and 26–41% withsorbitol) at similar levels. Importantly, the diVerences in thegrowth rates of the HZS 145 and HZS 190 strains observedat 30°C incubation temperature in the presence of KCl,NaCl or sorbitol were also comparable with those recordedat 37°C.
Again, in good accordance with the observations of Hag-iwara et al. (2008), HZS 190 mutant conidia possessed astress-sensitive phenotype. For example, they were highlysensitive to 100–200 mmol l¡1 H2O2 treatments, producingvirtually no survivals at the concentration of 200 mmol l¡1
H2O2 (data not shown). Furthermore, mutant conidia lostviability much more signiWcantly than spores from the HZS145 control strain after 1 week storage at 4°C (84 and 10%losses in the numbers of surviving conidia, respectively).Interestingly, heat stress (50°C) aVected mutant conidiaonly after prolonged, 60 min, incubation but not at 30 minincubation time (data not shown). It is worth noting thatboth the HZS 190 �atfA mutant and the HZS 145 controlstrains produced conidiospores in similar numbers withinthe range of 47–58 £ 107 spores cm¡2 colony area.
The deletion of atfA did not inXuence either the PAF-triggered (Leiter et al. 2005; Marx et al. 2008; Binder et al.2010) or the carbon starvation-triggered (Pócsi et al. 2009)apoptosis of the A. nidulans HZS 190 mutant and HZS 145control strains (data not shown). As far as the autolysismarkers (Emri et al. 2004, 2005; Pócsi et al. 2009) are con-cerned, there was no signiWcant diVerence between theautolytic loss of biomass and cell vitality and the kinetics ofthe age-dependent chitinase and proteinase productions ofthe HZS 190 and the HZS 145 strains up to 168 h of incu-bation at 37°C (data not shown).
Up to this point, the regulatory function of A. nidulansAtfA seemed to be limited to the coordination of the stressdefense of conidiospores (Hagiwara et al. 2008). However,
this view changed considerably after performing a large-scale oxidative stress sensitivity study on the available�atfA mutant and control strains.
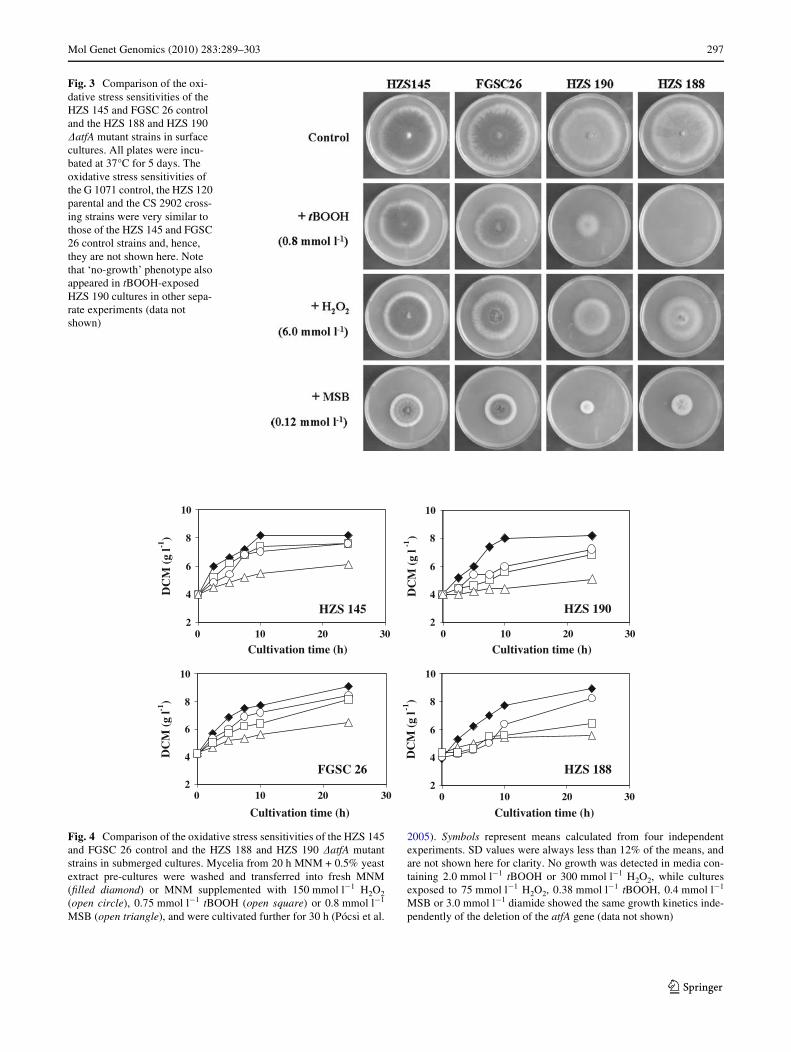
All single-copy transformants (three strains with geno-type pabaA1 riboB2 �atfA::riboB) and their progenies withdiVerent auxotrophy markers or prototrophs [25 prototroph(HZS 190, LE 1–24) and 7 auxotroph (HZS 187–189 and191–194) strains] were more sensitive to 0.8 mmol l¡1
tBOOH (65–100% growth inhibitions) than the controlG1071, HZS 145, HZS 120, CS 2902 and FGSC 26 strains(15–30% growth reductions) (Fig. 3). The appearance of‘no-growth’ phenotype in tBOOH-exposed mutants wasnot strain-speciWc because both high growth inhibition and‘no-growth’ phenotypes were often observed in separate setsof experiments with the same �atfA strains (data not shown).
The oxidative stress sensitivities of the HZS 188 biotinauxotroph and the HZS 190 prototroph strains were alsohigher than those of the control strains in the presence of6.0 mmol l¡1 H2O2 (30–40 and 10–24% growth reductions,respectively) and 0.12 mmol l¡1 MSB (37–58 and 66–78%growth inhibitions, respectively) (Fig. 3). Unexpectedly,the diVerence in the diamide sensitivities of the control andmutant strains recorded at 2.0 mmol l¡1 concentration wassurprisingly low with 57–62 and 65–68% growth reduc-tions, respectively.
In submerged cultures, the �atfA mutant and controlstrains reached quite similar, 4 ! 8–9 g l¡1 increases inDCM after 10–25 h incubation under unstressed condi-tions (Fig. 4). The HZS 188 and HZS 190 �atfA mutantswere more sensitive to H2O2 (150 mmol l¡1), tBOOH(0.75 mmol l¡1) and MSB (0.8 mmol l¡1) initiated oxida-tive stress at 10 h incubation time than the HZS 145 andFGSC 26 control strains (Fig. 4). Nevertheless, the H2O2-treated HZS 188 and HZS 190 and the tBOOH-exposedHZS 190 cultures reached approximately the same DCMproduction as their unstressed counterparts at 25 h cultiva-tion clearly indicating AtfA-independent cell recovery afterperoxide stress (Fig. 4). Importantly, no such recovery wasobserved in MSB-exposed mutant and control cultures,which can be explained by the redox-cycling character ofthis chemical, which causes accumulating oxidative stressin sensitive fungi (Emri et al. 1999; Pócsi et al. 2005).Although none of the control strains grew well in the pres-ence of 0.8 mmol l¡1 MSB the increases in DCMs observedwith the HZS 145 and FGSC 26 strains at 25 h cultivationtime (4 ! 6.1–6.5 g l¡1) exceeded signiWcantly thoserecorded with the mutants (4 ! 5.1–5.6 g l¡1; Fig. 4).Diamide, when employed in a concentration of 3.0 mmol l¡1,did not inXuence the growth of either the control or themutant strains (Fig. 4).
It is remarkable that the �atfA mutant generated byHagiwara et al. (2008) did not show any oxidative-stress-sensitive phenotypes. To try to solve this contradiction, we
123
Mol Genet Genomics (2010) 283:289–303 297
Fig. 3 Comparison of the oxi-dative stress sensitivities of the HZS 145 and FGSC 26 control and the HZS 188 and HZS 190 �atfA mutant strains in surface cultures. All plates were incu-bated at 37°C for 5 days. The oxidative stress sensitivities of the G 1071 control, the HZS 120 parental and the CS 2902 cross-ing strains were very similar to those of the HZS 145 and FGSC 26 control strains and, hence, they are not shown here. Note that ‘no-growth’ phenotype also appeared in tBOOH-exposed HZS 190 cultures in other sepa-rate experiments (data not shown)
Fig. 4 Comparison of the oxidative stress sensitivities of the HZS 145and FGSC 26 control and the HZS 188 and HZS 190 �atfA mutantstrains in submerged cultures. Mycelia from 20 h MNM + 0.5% yeastextract pre-cultures were washed and transferred into fresh MNM(Wlled diamond) or MNM supplemented with 150 mmol l¡1 H2O2(open circle), 0.75 mmol l¡1 tBOOH (open square) or 0.8 mmol l¡1
MSB (open triangle), and were cultivated further for 30 h (Pócsi et al.
2005). Symbols represent means calculated from four independentexperiments. SD values were always less than 12% of the means, andare not shown here for clarity. No growth was detected in media con-taining 2.0 mmol l¡1 tBOOH or 300 mmol l¡1 H2O2, while culturesexposed to 75 mmol l¡1 H2O2, 0.38 mmol l¡1 tBOOH, 0.4 mmol l¡1
MSB or 3.0 mmol l¡1 diamide showed the same growth kinetics inde-pendently of the deletion of the atfA gene (data not shown)
2
4
6
8
10
0 10 20 30
Cultivation time (h)
DC
M (
g l-1
)
HZS 145
2
4
6
8
10
0 10 20 30
Cultivation time (h)
DC
M (
g l-1
)
FGSC 262
4
6
8
10
0 10 20 30
Cultivation time (h)
DC
M (
g l-1
)
HZS 188
2
4
6
8
10
0 10 20 30
Cultivation time (h)
DC
M (
g l-1
)
HZS 190
123

298 Mol Genet Genomics (2010) 283:289–303
tested the possibilities that auxotrophy supplements mayinterfere with oxidative stress plate assays and/or that oxi-dative-stress-sensitive phenotypes may originate in theincreased stress sensitivity of �atfA mutant conidia.
As presented in Table 4, a mixture of commonly usedauxotrophy supplements in A. nidulans genetics mitigatedthe oxidative stress sensitivity of the HZS 190 mutantexposed to 0.6 mmol l¡1 tBOOH. When added separately,
riboXavin, PABA and pyridoxine moderated signiWcantlythe tBOOH sensitive phenotype. On the other hand, supple-mentation of MNM agar with the combination of thesecompounds did not aVect the ‘no-growth’ phenotype ofHZS 190 when this phenotype appeared in the presence of0.8 mmol l¡1 tBOOH (data not shown).
Importantly, A. nidulans HZS 190 mycelia pre-grown onMNM agar under unstressed conditions grew more slowlythan their HZS 145 counterparts or even did not grow at allwhen they were exposed to high concentrations (2.0–3.0 mmol l¡1) of tBOOH in MNM surface cultures (Fig. 5).This means that, in addition to mutant conidia, vegetativetissues of �atfA mutants also had an oxidative-stress-sensi-tive phenotype.
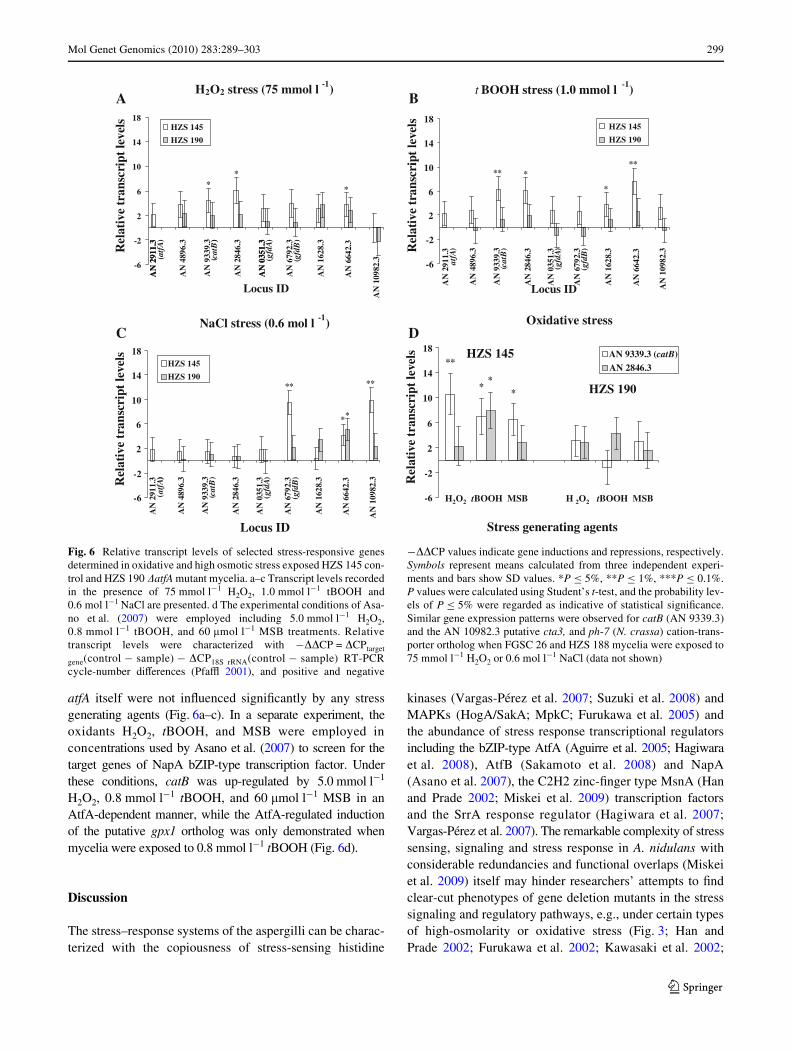
AtfA-regulated stress-responsive genes
As shown in Fig. 6a–c, AtfA regulated the transcription ofseveral stress-responsive genes under versatile stress condi-tions including 75 mmol l¡1 H2O2 [catB catalase, AN2846.3putative gpx1 (S. pombe) glutathione peroxidase ortholog,AN6642.3 putative cta3 (S. pombe) and ena-1 (N. crassa) cat-ion-transporting P-type ATPase ortholog], 1.0 mmol l¡1
tBOOH (catB, AN 2846.3 putative gpx1 ortholog, AN1628.3and AN6642.3 putative cta3 and ena-1 cation-transporterorthologs) and 0.6 mol l¡1 NaCl [gfdB glycerol-3-phosphatedehydrogenase, AN10982.3 putative cta3 and ph-7 (N. crassa)cation-transporter ortholog] treatments. The AN6642.3 puta-tive cta3 and ena-1 cation-transporting P-type ATPase ortho-log was equally induced in control and �atfA mutant strains,which indicated a salt-stress-responsive but AtfA-independentregulation for this gene. Among the tested genes, the transcrip-tions of AN4896.3 putative pyp2 (S. pombe) tyrosine phospha-tase ortholog, gfdA glycerol-3-phosphate dehydrogenase and
Table 4 Mitigation of the tBOOH-sensitivity of the HZS 190 mutantwith auxotrophy supplements
a The concentrations of the supplements were 50 �g ml¡1 uracil/5 mmol l¡1 uridine, 5 �mol l¡1 PABA, 0.02 �g ml¡1 biotin, 0.05 �g ml¡1
pyridoxine and/or 0.1 �g ml¡1 riboXavin in accordance with theAspergillus Media Supplements Recommendations of the FungalGenetics Stock Center (http://www.gla.ac.uk/ibls/molgen/aspergillus/supplement.html)b Mean § SD values calculated from nine independent experimentsare shown. *P · 5%, **P · 1%, ***P · 0.1%, where P values werecalculated using the Student’s t-testc Growth inhibitions with 0.6 mmol l¡1 tBOOH are presentedd HZS 190 grown on MNM agar without any tBOOH supplementationwas regarded as controle The mixture of all supplements listed earlier
Supplementsa Growthb colony diameter (cm)
Growth inhibition by tBOOHc (%)
Controld 7.03 § 0.05 0
– 4.6 § 0.3 34
Biotin 4.9 § 0.3 31
PABA 5.4 § 0.1*** 23
Pyridoxine 5.1 § 0.3** 27
RiboXavin 5.6 § 0.3*** 20
Uracil/uridine 4.8 § 0.4 32
Mixturee 6.0 § 0.3*** 15
Fig. 5 DiVerences in the oxidative stress sensitivities of A. nidulansHZS 145 and HZS 190 mycelia exposed to increasing concentrationsof tBOOH. Conidia (105) were incubated on cellophane sheets placedon the surfaces of MNM agar plates at 37°C for 24 h. Following that
colonies were lifted and laid on MNM stress plates supplemented withtBOOH and were incubated for 5 days. The tBOOH sensitivity of theG 1071 control strain was identical to that of the HZS 145 strain (datanot shown)
123
Mol Genet Genomics (2010) 283:289–303 299
atfA itself were not inXuenced signiWcantly by any stressgenerating agents (Fig. 6a–c). In a separate experiment, theoxidants H2O2, tBOOH, and MSB were employed inconcentrations used by Asano et al. (2007) to screen for thetarget genes of NapA bZIP-type transcription factor. Underthese conditions, catB was up-regulated by 5.0 mmol l¡1
H2O2, 0.8 mmol l¡1 tBOOH, and 60 �mol l¡1 MSB in anAtfA-dependent manner, while the AtfA-regulated inductionof the putative gpx1 ortholog was only demonstrated whenmycelia were exposed to 0.8 mmol l¡1 tBOOH (Fig. 6d).
Discussion
The stress–response systems of the aspergilli can be charac-terized with the copiousness of stress-sensing histidine
kinases (Vargas-Pérez et al. 2007; Suzuki et al. 2008) andMAPKs (HogA/SakA; MpkC; Furukawa et al. 2005) andthe abundance of stress response transcriptional regulatorsincluding the bZIP-type AtfA (Aguirre et al. 2005; Hagiwaraet al. 2008), AtfB (Sakamoto et al. 2008) and NapA(Asano et al. 2007), the C2H2 zinc-Wnger type MsnA (Hanand Prade 2002; Miskei et al. 2009) transcription factorsand the SrrA response regulator (Hagiwara et al. 2007;Vargas-Pérez et al. 2007). The remarkable complexity of stresssensing, signaling and stress response in A. nidulans withconsiderable redundancies and functional overlaps (Miskeiet al. 2009) itself may hinder researchers’ attempts to Wndclear-cut phenotypes of gene deletion mutants in the stresssignaling and regulatory pathways, e.g., under certain typesof high-osmolarity or oxidative stress (Fig. 3; Han andPrade 2002; Furukawa et al. 2002; Kawasaki et al. 2002;
Fig. 6 Relative transcript levels of selected stress-responsive genesdetermined in oxidative and high osmotic stress exposed HZS 145 con-trol and HZS 190 �atfA mutant mycelia. a–c Transcript levels recordedin the presence of 75 mmol l¡1 H2O2, 1.0 mmol l¡1 tBOOH and0.6 mol l¡1 NaCl are presented. d The experimental conditions of Asa-no et al. (2007) were employed including 5.0 mmol l¡1 H2O2,0.8 mmol l¡1 tBOOH, and 60 �mol l¡1 MSB treatments. Relativetranscript levels were characterized with ¡��CP = �CPtarget
gene(control ¡ sample) ¡ �CP18S rRNA(control ¡ sample) RT-PCRcycle-number diVerences (PfaZ 2001), and positive and negative
¡��CP values indicate gene inductions and repressions, respectively.Symbols represent means calculated from three independent experi-ments and bars show SD values. *P · 5%, **P · 1%, ***P · 0.1%.P values were calculated using Student’s t-test, and the probability lev-els of P · 5% were regarded as indicative of statistical signiWcance.Similar gene expression patterns were observed for catB (AN 9339.3)and the AN 10982.3 putative cta3, and ph-7 (N. crassa) cation-trans-porter ortholog when FGSC 26 and HZS 188 mycelia were exposed to75 mmol l¡1 H2O2 or 0.6 mol l¡1 NaCl (data not shown)
H2O2 stress (75 mmol l -1)
-6
-2
2
6
10
14
18
Locus ID
Rel
ativ
etr
ansc
ript
leve
ls HZS 145
HZS 190
A
(A
N 2
911.
3at
fA)
AN
291
1.3
AN
489
6.3
AN
933
9.3
( cat
B))
AN
284
6.3
AN
035
1.3
(gfd
A )A
N 0
351.
3
AN
679
2.3
(gfd
B)
AN
162
8.3
AN
664
2.3
AN
109
82.3
**
*
NaCl stress (0.6 mol l -1)
-6
-2
2
6
10
14
18
Locus ID
Rel
ativ
etr
ansc
ript
leve
ls
C
**
**
**
)A
N 2
911.
3at
fA)
(
AN
489
6.3
AN
933
9.3
(cat
B)
AN
284
6.3
AN
035
1.3
(gfd
A)
AN
679
2.3
( gfd
B)
AN
162
8.3
AN
664
2.3
AN
109
82.3
Oxidative stress
-6
-2
2
6
10
14
18
Stress generating agents
Rel
ativ
etr
ansc
ript
leve
ls AN 9339.3 (catB)AN 2846.3
H2O2 tBOOH MSB H 2O2 tBOOH MSB
D
**
**
* HZS 190
HZS 145
t BOOH stress (1.0 mmol l -1)
-6
-2
2
6
10
14
18
Locus ID
Rel
ativ
etr
ansc
ript
leve
ls
B
** ***
*
AN
291
1.3
atfA
)
AN
489
6.3
AN
933
9.3
(cat
B)
AN
284
6.3
AN
035
1.3
( gfd
A)
AN
679
2.3
(gfd
B)
AN
162
8.3
AN
664
2.3
AN
109
82.3
HZS 145
HZS 190
HZS 145
HZS 190
123

300 Mol Genet Genomics (2010) 283:289–303
Vargas-Pérez et al. 2007). When faced with the complexityof stress signaling and regulation of stress response in theaspergilli (Miskei et al. 2009), complementation studies inyeast mutants, where orthologs of the corresponding Asper-gillus genes have been deleted, are of especially great valuein the functional analyses of the gene products (Kawasakiet al. 2002; Furukawa et al. 2002, 2005). In this study, thesuccessful complementation of the osmotic and oxidative-stress-sensitive phenotypes of the S. pombe �atf1 strainwith atfA (Figs. 1, 2) demonstrated that AtfA is a true func-tional ortholog of Wssion yeast’s Atf1. This observationsuggests that AtfA functions downstream of the SskB–PbsB–HogA/SakA stress signaling pathway and transmitsdiVerent types of stress signals to an array of stress-respon-sive genes (Pócsi et al. 2005; Miskei et al. 2009).
The pivotal role of AtfA in the regulation of the osmoticand oxidative stress responses of A. nidulans was demon-strated by the observation that the A. nidulans orthologs ofwell-known target genes of Wssion yeast Atf1 did notrespond to stress in �atfA mutants grown in liquid MNMcultures (Fig. 6). Among the putative S. pombe orthologs ofthe tested A. nidulans genes, atf1, pyp2, ctt1, gpx1, andgpd1 were induced in Wssion yeast by diVerent types ofstress as part of the CESR (Chen et al. 2003, 2008). In A.nidulans, the induction of the majority of the selected geneswas stress-type-speciWc rather than stress-responsive ingeneral, and the transcription of the putative pyp2 ortholog,and atfA itself did not respond signiWcantly to stress(Fig. 6). These Wndings do not support the idea that aWssion yeast-like CESR may exist in A. nidulans, butwhole-genome expression proWling studies have not beenperformed yet in this fungus to answer the important ques-tion of whether a group of genes signiWcantly induced orrepressed by all or most types of stress also operates in thegenome of A. nidulans (Miskei et al. 2009). Interestingly,the expression of the putative gpx1 ortholog (locus ID AN2846.3) was induced by tBOOH and high (75 mmol l¡1)concentration H2O2 treatments in A. nidulans but was notaVected signiWcantly by MSB and 5.0 mmol l¡1 H2O2
(Fig. 6). These Wndings indicate that, similar to S. pombe(Chen et al. 2008), diverse gene expression programs mayexist in A. nidulans to protect the cells from oxidative dam-age in a stress-type and dose-dependent manner.
In Wssion yeast, mutants defective in the Sty1/Spc1-Atf1regulatory pathway are hypersensitive to diVerent types ofenvironmental stress (Nguyen et al. 2000; Quinn et al.2002; Chen et al. 2008). In good accordance with this butcontrary to the previous Wndings of Hagiwara et al. (2008),the deletion of atfA resulted in oxidative-stress-sensitivephenotypes in A. nidulans (Figs. 3, 4, 5). On the other hand,the �atfA mutations did not aVect signiWcantly the osmoticand diamide stress tolerance of the fungus, which is inagreement with the observations of Hagiwara et al. (2008).
Contradictory phenotypes have also been presented for theA. nidulans �sskA mutants by Hagiwara et al. (2007) andVargas-Pérez et al. (2007), and these discrepancies remainyet to be explained.
In addition to likely genotypic variations and the lack ofinterlaboratory standardization of stress-sensitivity assays,commonly used auxotrophy supplements and their combi-nations (Table 4) may mitigate or even mask stress-sensitive phenotypes, which subsequently may lead toambiguous phenotype descriptions. Among the additivestested, riboXavin, which may protect cells from oxidativeinjuries (Sugiyama 1991; Perumal et al. 2005), consider-ably decreased the tBOOH-sensitivity of the HZS 190mutant (Table 4). Considering the antioxidant eVect of pyr-idoxine, pyridoxine derivatives have been demonstrated toprevent the death of MSB-exposed Wssion yeast cells(Chumnantana et al. 2005). The oxidative stress protectiveeVect of PABA may originate in its precursor function infolic acid biosynthesis. Folic acid plays a pivotal role in theprevention of mitochondria-associated oxidative injuries(Huang et al. 2004; Chang et al. 2007), which are intensi-Wed in tBOOH-exposed fungal cultures (Fekete et al. 2007).Relying on the outcomes of these studies, we argue here forthe use of prototrophic strains to promote interlaboratorystandardization of future stress assays.
In good accordance with the Wndings of Hagiwara et al.(2008), conidia of HZS 190 were highly sensitive to H2O2-treatments, heat stress, and storage in glucose-free MNM at4°C for 1 week. Importantly, the oxidative-stress-sensitivephenotypes observed with �atfA strains were not a mere con-sequence of the increased stress sensitivity of mutant conidiaas demonstrated by transferring pre-grown A. nidulans myce-lial mats onto tBOOH-supplemented stress plates (Fig. 5).Nevertheless, colonies growing out from the mats (eithercontrol or mutant) tolerated tBOOH-treatments at approxi-mately twice higher concentrations than their conidia-inocu-lated counterparts (Figs. 3, 5). This phenomenon may beexplained by the bioWlm-like texture of the mycelial matspre-grown on cellophane sheets. More recently, baker’s yeastcells growing as bioWlms were characterized with anincreased oxidative stress tolerance (Gales et al. 2008).Because the generation of stress signal transduction pathwaymutants frequently leads to the formation of stress-sensitiveconidia (Hagiwara et al. 2007, 2008; Vargas-Pérez et al.2007) and the contribution of the increased stress-sensitivityof conidiospores to the apparent stress-sensitive phenotypesof the mutants cannot be quantiWed properly (especially noton stress agar plates when ‘no-growth’ phenotypes mayappear unpredictably; Fig. 3), we also recommend the use ofmycelium-inoculated liquid cultures to compare the stress-sensitivities of mutant and control strains (Fig. 4).
There are numerous papers reporting on the apoptosis-eliciting eVect of the phosphorylated form of ATF-2
123

Mol Genet Genomics (2010) 283:289–303 301
(Cho et al. 2001; Makino et al. 2006; Breitwieser et al. 2007),the human ortholog of S. pombe Atf1 (Degols and Russell1997; Lawrence et al. 2007). On the other hand, the overex-pression of another bZIP-type oxidative stress responsetranscription factor, Yap1p, strongly delayed chronologicalaging-induced apoptosis in S. cerevisiae (Herker et al.2004). We addressed therefore the question of whether thedeletion of atfA would have any impact on the apoptoticcell death processes in A. nidulans. Contrary to our expec-tations, the deletion of atfA did not aVect either PAF-elic-ited or carbon-starvation-triggered apoptosis in A. nidulans.The unaVected apoptosis rates indicate that cell-death sig-nals may be transmitted by signal transduction pathwaysother than the SskA–SskB–PbsB–HogA/SakA–AtfA sys-tem, e.g. via FadA heterotrimeric G-protein signaling(Leiter et al. 2005; Marx et al. 2008). It is important to notethat neither SskA (Hagiwara et al. 2007) nor AtfA (Figs. 3,5) are necessary to respond to GSH/GSSG redox imbalancein A. nidulans, which is a well-known elicitor of pro-grammed cell death in fungi (Madeo et al. 1999; Pócsi et al.2004).
The deletion of atfA did not aVect the autolysis of thefungus in submerged cultures either and, therefore, its par-ticipation in the regulation of autolysis also seems to beunlikely. This is understandable because the autolytic phaseof growth is preceded by a severe GSH/GSSG redox imbal-ance in carbon-depleted submerged cultures of A. nidulans(i.e. by a physiological change that the �atfA mutant is notlargely sensitive to), but not by the accumulation of reac-tive oxygen species (i.e. not by changes that the �atfAmutant is largely sensitive to; Fig. 3; Emri et al. 2004). InA. nidulans, autolysis is initiated by the FluG-BrlA conidia-tion initiation pathway (Emri et al. 2005; Pócsi et al. 2009),while the age-dependent degradation of GSH and, hence,the pre-autolytic changes in the GSH/GSSG redox balanceis under the control of FadA/FlbA and GanB/RgsA hetero-trimer G-protein signaling pathways (Molnár et al. 2004,2006; Emri et al. 2008).
In conclusion, the AtfA bZIP-type transcription factor,which is a true ortholog of the Wssion yeast’s ‘generalstress’ transcription factor Atf1, plays an important role inthe regulation of the elements of the oxidative and osmoticstress responses in the vegetative tissues of A. nidulans.AtfA is also involved in stress-tolerant conidia formation(Hagiwara et al. 2008) but does not seem to play a majorrole in the regulation of the GSH/GSSG redox balance ofthe cells or any role in the initiation and regulation of pro-grammed cell death and autolysis. The �atfA mutants andcontrol strains, which were physiologically characterized inthis study, will be used in genome-wide gene expressionexperiments with oligomer-based DNA microarrays to mapthe elements of general and stress-type-speciWc stressresponses in A. nidulans.
During the revision of this paper, Hagiwara et al. (2009)published DNA microarray data on Xudioxonil (this anti-mycotic exerts its action through Hog MAPK signalingpathway) and sorbitol-responsive A. nidulans genes using aseries of mutants impaired in SskA—HogA/SakA—AtfAsignaling. The authors found considerable overlapsbetween sskA-, hogA/sakA-, and atfA-dependent genes,which were up-regulated by Xudioxonil and sorbitol treat-ments. Moreover, the Xudioxonil-triggered induction ofatfA itself was both sskA- and hogA/sakA-dependent. Basedon these observations, Hagiwara et al. (2009) reached theconclusions that AtfA functions downstream of HogA/SakA, and SskA—HogA/SakA—AtfA signaling is impli-cated in the transcriptional response to Xudioxonil andosmotic stress. In another recent study, Sakamoto et al.(2009) demonstrated that some oxidative stress responsegenes encoding catalases, thioredoxin, �-glutamylcysteinesynthase, 6-phosphogluconate dehydrogenase, etc. weredownregulated in the colony surface cells (aerial hyphaeand conidia) of an Aspergillus oryzae �atfA mutant. TheA. oryzae �atfA mutant conidia were also sensitive tooxidative stress and possessed a low germination ratio evenwithout any stress (Sakamoto et al. 2009).
Acknowledgments The Authors are indebted to Mrs. LászlónéGábor Tóth for her valuable technical assistance and Mr. MatthewBritschgi for editing the English of the paper. One of us (ZH) wasawarded a János Bolyai Research Scholarship by the Hungarian Academyof Sciences. This work was supported Wnancially by the HungarianNational OYce for Research and Technology (grant reference numberOMFB 01501/2006) and by the GENOMNANOTECH-DEBRET(RET-06/2004).
References
Aguirre J, Ríos-Momberg M, Hewitt D, Hansberg W (2005) Reactiveoxygen species and development in microbial eukaryotes. TrendsMicrobiol 13:111–118
Asano Y, Hagiwara D, Yamashino T, Mizuno T (2007) Characteriza-tion of the bZIP-type transcription factor NapA with reference tooxidative stress response in Aspergillus nidulans. Biosci Biotech-nol Biochem 71:1800–1803
Barford NC (1967) Experimental measurements: precision, error andtruth. Addison-Wesley, London, pp 29–32
Barratt RW, Johnson GB, Ogata WN (1965) Wild-type and mutantstocks of Aspergillus nidulans. Genetics 52:233–246
Basi G, Schmid E, Maundrell K (1993) TATA box mutations in theSchizosaccharomyces pombe nmt1 promoter aVect transcriptioneYciency but not the transcription start point or thiamine repress-ibility. Gene 123:131–136
Benito B, Garciadeblás B, Rodríguez-Navarro A (2000) Molecularcloning of the calcium and sodium ATPases in Neurospora cras-sa. Mol Microbiol 35:1079–1088
Binder U, Oberparleiter C, Meyer V, Marx F (2010) The antifungalprotein PAF interferes with PKC/MPK and cAMP/PKA signal-ling of Aspergillus nidulans. Mol Microbiol 75:294–307
Breitwieser W, Lyons S, Flenniken AM, Ashton G, Bruder G, WillingtonM, Lacaud G, KouskoV V, Jones N (2007) Feedback regulation of
123

302 Mol Genet Genomics (2010) 283:289–303
p38 activity via ATF2 is essential for survival of embryonic livercells. Genes Dev 21:2069–2082
Chang CM, Yu CC, Lu HT, Chou YF, Huang RF (2007) Folatedeprivation promotes mitochondrial oxidative decay: DNA largedeletions, cytochrome c oxidase dysfunction, membrane depolar-ization and superoxide overproduction in rat liver. Br J Nutr97:855–863
Chen D, Toone WM, Mata J, Lyne R, Burns G, Kivinen K, Brazma A,Jones N, Bähler J (2003) Global transcriptional responses of Ws-sion yeast to environmental stress. Mol Biol Cell 14:214–229
Chen D, Wilkinson CRM, Watt S, Penkett CJ, Toone WM, Jones N,Bähler J (2008) Multiple pathways diVerentially regulate globaloxidative stress responses in Wssion yeast. Mol Biol Cell 19:308–317
Cho SG, Bhoumik A, Broday L, Ivanov V, Rosenstein B, Ronai Z(2001) TIP49b, a regulator of activating transcription factor 2 re-sponse to stress and DNA damage. Mol Cell Biol 21:8398–8413
Chumnantana R, Yokochi N, Yagi T (2005) Vitamin B6 compoundsprevent the death of yeast cells due to menadione, a reactive oxy-gen generator. Biochim Biophys Acta 1722:84–91
Cove DJ (1966) The induction and repression of nitrate reductase in thefungus Aspergillus nidulans. Biochim Biophys Acta 113:51–56
Degols G, Russell P (1997) Discrete roles of the Spc1 kinase and theAtf1 transcription factor in the UV response of Schizosaccharo-myces pombe. Mol Cell Biol 17:3356–3363
Emri T, Pócsi I, Szentirmai A (1999) Analysis of the oxidative stressresponse of Penicillium chrysogenum to menadione. Free RadicRes 30:125–132
Emri T, Molnár Z, Pusztahelyi T, Pócsi I (2004) Physiological andmorphological changes in autolyzing Aspergillus nidulans cul-tures. Folia Microbiol 49:277–284
Emri T, Molnár Z, Pusztahelyi T, Varecza Z, Pócsi I (2005) The FluG-BrlA pathway contributes to the initialisation of autolysis in sub-merged Aspergillus nidulans cultures. Mycol Res 109:757–763
Emri T, Molnár Z, Veres T, Pusztahelyi T, Dudás G, Pócsi I (2006)Glucose-mediated repression of autolysis and conidiogenesis inEmericella nidulans. Mycol Res 110:1172–1178
Emri T, Zs Molnár, Szilágyi M, Pócsi I (2008) Regulation of autolysisin Aspergillus nidulans. Appl Biochem Biotechnol 151:211–220
Fekete A, Emri T, Gyetvai A, Gazdag Z, Pesti M, Varga Z, Balla J,Cserháti C, Emödy L, Gergely L, Pócsi I (2007) Development ofoxidative stress tolerance resulted in reduced ability to undergomorphologic transitions and decreased pathogenicity in a t-butyl-hydroperoxide-tolerant mutant of Candida albicans. FEMS YeastRes 7:834–847
Furukawa K, Katsumo Y, Urao T, Yabe T, Yamada-Okabe T,Yamada-Okabe H, Yamagata Y, Abe K, Nakajima T (2002) Iso-lation and functional analysis of a gene, tcsB, encoding a trans-membrane hybrid-type histidine kinase from Aspergillusnidulans. Appl Environ Microbiol 68:5304–5310
Furukawa K, Hoshi Y, Maeda T, Nakajima T, Abe K (2005) Aspergil-lus nidulans HOG pathway is activated only by two-componentsignalling pathway in response to osmotic stress. Mol Microbiol56:1246–1261
Furukawa K, Yoshimi A, Furukawa T, Hoshi Y, Hagiwara D, Sato N,Fujioka T, Mizutani O, Mizuno T, Kobayashi T, Abe K (2007)Novel reporter gene expression systems for monitoring activationof the Aspergillus nidulans HOG pathway. Biosci BiotechnolBiochem 71:1724–1730
Galagan JE, Calvo SE, Cuomo C, Ma LJ, Wortman JR, Batzoglou S,Lee SI, Baotürkmen M, Spevak CC, Clutterbuck J, Kapitonov V,Jurka J, Scazzocchio C, Farman M, Butler J, Purcell S, Harris S,Braus GH, Draht O, Busch S, D’Enfert C, Bouchier C, GoldmanGH, Bell-Pedersen D, GriYths-Jones S, Doonan JH, Yu J,Vienken K, Pain A, Freitag M, Selker EU, Archer DB, PeñalvaMA, Oakley BR, Momany M, Tanaka T, Kumagai T, Asai K,
Machida M, Nierman WC, Denning DW, Caddick M, Hynes M,Paoletti M, Fischer R, Miller B, Dyer P, Sachs MS, Osmani SA,Birren BW (2005) Sequencing of Aspergillus nidulans andcomparative analysis with A. fumigatus and A. oryzae. Nature438:1105–1115
Gales G, Penninckx M, Block JC, Leroy P (2008) Role of glutathionemetabolism status in the deWnition of some cellular parametersand oxidative stress tolerance of Saccharomyces cerevisiae cellsgrowing as bioWlms. FEMS Yeast Res 8:667–675
Gasch AP (2007) Comparative genomics of the environmental stressresponse in ascomycete fungi. Yeast 24:961–976
Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB, StorzG, Botstein D, Brown PO (2000) Genomic expression programsin the response of yeast cells to environmental changes. Mol BiolCell 11:4241–4257
Hagiwara D, Asano Y, Marui J, Furukawa K, Kanamaru K, Kato M,Abe K, Kobayashi T, Yamashino T, Mizuno T (2007) The SskAand SrrA response regulators are implicated in oxidative stress re-sponses of hyphae and asexual spores in the phosphorelay signal-ing network of Aspergillus nidulans. Biosci Biotechnol Biochem71:1003–1014
Hagiwara D, Asano Y, Yamashino T, Mizuno T (2008) Characteriza-tion of bZip-type transcription factor AtfA with reference to stressresponses of conidia of Aspergillus nidulans. Biosci BiotechnolBiochem 72:2756–2760
Hagiwara D, Asano Y, Marui J, Yoshimi A, Mizuno T, Abe K (2009)Transcriptional proWling of Aspergillus nidulans HogA MAPKsignaling pathway in response to Xudioxonil and osmotic stress.Fungal Genet Biol 46:868–878
Han KH, Prade RA (2002) Osmotic stress-coupled maintenance ofpolar growth in Aspergillus nidulans. Mol Microbiol 43:1065–1078
Herker E, Jungwirth H, Lehmann KA, Maldener C, Fröhlich KU,Wissing S, Büttner S, Fehr M, Sigrist S, Madeo F (2004) Chrono-logical aging leads to apoptosis in yeast. J Cell Biol 164:501–507
Huang RF, Yaong HC, Chen SC, Lu YF (2004) In vitro folate supple-mentation alleviates oxidative stress, mitochondria-associateddeath signaling and apoptosis induced by 7-ketocholesterol. BrJ Nutr 92:887–894
Ito H, Fukuda Y, Murata K, Kimura A (1983) Transformation of intactyeast cells treated with alkali cations. J Bacteriol 153:163–168
Kaiserer L, Oberparleiter C, Weiler-Görz R, Burgstaller W, Leiter E,Marx F (2003) Characterization of the Penicillium chrysogenumantifungal protein PAF. Arch Microbiol 180:204–210
Kato N, Brooks W, Calvo AN (2003) The expression of sterigmatocy-stin and penicillin genes in Aspergillus nidulans is controlled byveA, a gene required for sexual development. Eukaryot Cell2:1178–1186
Kawasaki L, Wysong D, Diamond R, Aguirre J (1997) Two divergentcatalase genes are diVerentially regulated during Aspergillus nidu-lans development and oxidative stress. J Bacteriol 179:3284–3292
Kawasaki L, Sánchez O, Shiozaki K, Aguirre J (2002) SakA MAP ki-nase is involved in stress signal transduction, sexual developmentand spore viability in Aspergillus nidulans. Mol Microbiol45:1153–1163
Kim H, Han K, Kim K, Han D, Jahng K, Chae K (2002) The veA geneactivates sexual development in Aspergillus nidulans. FungalGenet Biol 37:72–80
Lawrence CL, Maekawa H, Worthington JL, Reiter W, Wilkinson CR,Jones N (2007) Regulation of Schizosaccharomyces pombe Atf1protein levels by Sty1-mediated phosphorylation and heterodi-merization with Pcr1. J Biol Chem 282:5160–5170
Leiter É, Szappanos H, Oberparleiter C, Kaiserer L, Csernoch L,Pusztahelyi T, Emri T, Pócsi I, Salvenmoser W, Marx F (2005)Antifungal protein PAF severely aVects the integrity of the plas-ma membrane of Aspergillus nidulans and induces an apoptosis-like phenotype. Antimicrob Agents Chemother 49:2445–2453
123

Mol Genet Genomics (2010) 283:289–303 303
Madeo F, Fröhlich E, Ligr M, Grey M, Sigrist SJ, Wolf DH, FröhlichKU (1999) Oxygen stress. A regulator of apoptosis in yeast. J CellBiol 145:757–767
Makino C, Sano Y, Shinagawa T, Millar JB, Ishii S (2006) Sin1 bindsto both ATF-2 and p38 and enhances ATF-2-dependent transcrip-tion in an SAPK signaling pathway. Genes Cells 11:1239–1251
Marx F, Binder U, Leiter É, Pócsi I (2008) The Penicillium chrysoge-num antifungal protein PAF, a promising tool for the developmentof new antifungal therapies and fungal cell biology studies. CellMol Life Sci 65:445–454
Maundrell K (1993) Thiamine-repressible expression vectors pREPand pRIP for Wssion yeast. Gene 123:127–130
Miskei M, Karányi Z, Pócsi I (2009) Annotation of stress-responseproteins in the aspergilli. Fungal Genet Biol 46:S105–S120
Molnár Z, Mészáros É, Szilágyi Z, Rosén S, Emri T, Pócsi I (2004)InXuence of fadAG203R and �XbA mutations on morphology andphysiology of submerged Aspergillus nidulans cultures. ApplBiochem Biotechnol 118:349–360
Molnár Z, Emri T, Zavaczki E, Pusztehelyi T, Pócsi I (2006) EVects ofmutations in the GanB/RgsA G protein mediated signaling on theautolysis of Aspergillus nidulans. J Basic Microbiol 46:495–603
Mooney JL, Yager LN (1990) Light is required for conidiation inAspergillus nidulans. Genes Dev 4:1473–1482
Nakagawa CW, Yamada K, Mutoh N (2000) Role of Atf1 and Pap1 inthe induction of the catalase gene of Wssion yeast Schizosaccha-romyces pombe. J Biochem 127:233–238
Nguyen AN, Lee A, Place W, Shiozaki K (2000) Multistep phospho-relay proteins transmit oxidative stress signals to the Wssion yeaststress-activated protein kinase. Mol Biol Cell 11:1169–1181
Nishikawa T, Aiba H, Mizuno T (1999) The cta3+ gene that encodes acation-transporting P-type ATPase is induced by salt stress undercontrol of the Wis-Sty1 MAPKK-MAPK cascade in Wssion yeast.FEBS Lett 455:183–187
Perumal SS, Shanthi P, Sachdanandam P (2005) Augmented eYcacyof tamoxifen in rat breast tumorigenesis when gavaged along withriboXavin, niacin, and CoQ10: eVect on lipid peroxidation andantioxidants in mitochondria. Chem Biol Interact 152:49–58
PfaZ MW (2001) A new mathematical model for relative quantiWca-tion in real-time RT-PCR. Nucleic Acids Res 29: article no. e45
Pócsi I, Prade RA, Penninckx MJ (2004) Glutathione, altruistic metab-olite in fungi. Adv Microb Physiol 49:1–76
Pócsi I, Miskei M, Karányi Z, Emri T, Ayoubi P, Pusztahelyi T, BallaG, Prade RA (2005) Comparison of gene expression signatures ofdiamide, H2O2 and menadione exposed Aspergillus nidulans cul-tures—linking genome-wide transcriptional changes to cellularphysiology. BMC Genomics 6: article no. 182
Pócsi I, Leiter É, Kwon NJ, Shin KS, Kwon GS, Pusztahelyi T, EmriT, Abuknesha RA, Price RG, Yu JH (2009) Asexual sporulationsignaling regulates autolysis of Aspergillus nidulans via modulat-ing the chitinase ChiB production. J Appl Microbiol 107:514–523
Pusztahelyi T, Molnár Z, Emri T, Klement É, Miskei M, Kerékgyártó J,Balla J, Pócsi I (2006) Comparative studies of diVerential expres-sion of chitinolytic enzymes encoded by chiA, chiB, chic and nagAgenes in Aspergillus nidulans. Folia Microbiol 51:547–554
Quinn J, Findlay VJ, Dawson K, Millar JB, Jones N, Morgan BA,Toone WM (2002) Distinct regulatory proteins control the graded
transcriptional response to increasing H2O2 levels in Wssion yeastSchizosaccharomyces pombe. Mol Biol Cell 13:805–816
Sakamoto K, Arima TH, Iwashita K, Yamada O, Gomi K, Akita O(2008) Aspergillus oryzae atfB encodes a transcription factor re-quired for stress tolerance in conidia. Fungal Genet Biol45:922–932
Sakamoto K, Iwashita K, Yamada O, Kobayashi K, Mizuno A, AkitaO, Mikami S, Shimoi H, Gomi K (2009) Aspergillus oryzae atfAcontrols conidial germination and stress tolerance. Fungal GenetBiol 46:887–897
Sansó M, Gogol M, Ayté J, Seidel C, Hidalgo E (2008) Transcriptionfactors Pcr1 and Atf1 have distinct roles in stress- and Sty1-dependent gene regulation. Eukaryot Cell 7:826–835
Scazzocchio C, Sdrin N, Ong G (1982) Positive regulation in aneukaryote, a study of the uaY gene of Aspergillus nidulans. I.Characterization of alleles, dominance and complementationstudies, and a Wne structure map of the uaY–oxpA cluster. Genet-ics 100:185–208
Shiozaki K, Russell P (1996) Conjugation, meiosis, and the osmoticstress response are regulated by Spc1 kinase through Atf1 tran-scription factor in Wssion yeast. Genes Dev 10:2276–2288
Sugiyama M (1991) EVects of vitamins on chromium(VI)-induceddamage. Environ Health Perspect 92:63–70
Suzuki A, Kanamaru K, Azuma N, Kato M, Kobayashi T (2008) GFP-tagged expression analysis revealed that some histidine kinases ofAspergillus nidulans show temporally and spatially diVerentexpression during the life cycle. Biosci Biotechnol Biochem72:428–434
Takeda T, Toda T, Kominami K, Kohnosu A, Yanagida M, Jones N(1995) Schizosaccharomyces pombe atf1+ encodes a transcriptionfactor required for sexual development and entry into stationaryphase. EMBO J 14:6193–6208
Tilburn J, Scazzocchio C, Taylor GG, Zabicky-Zissman JH, Locking-ton RA, Davies RW (1983) Transformation by integration inAspergillus nidulans. Gene 26:205–221
Vargas-Pérez I, Sánchez O, Kawasaki L, Georgellis D, Aguirre J(2007) Response regulators SrrA and SskA are central compo-nents of a phosphorelay system involved in stress signal transduc-tion and asexual sporulation in Aspergillus nidulans. EukaryotCell 6:1570–1583
Wilkinson MG, Samuels M, Takeda T, Toone WM, Shieh JC, Toda T,Millar JB, Jones N (1996) The Atf1 transcription factor is a targetfor the Sty1 stress-activated MAP kinase pathway in Wssion yeast.Genes Dev 10:2289–2301
Xue T, Nguyen K, Romans A, May GS (2004) A mitogen-activatedprotein kinase that senses nitrogen regulates conidial germinationand growth in Aspergillus fumigatus. Eukaryot Cell 3:557–560
Yamada K, Nakagawa CW, Mutoh N (1999) Schizosaccharomycespombe homologue of glutathione peroxidase, which does not con-tain selenocysteine, is induced by several stresses and works as anantioxidant. Yeast 15:1125–1132
Yu JH, Hamari Z, Han KH, Seo JA, Reyes-Domínguez Y, ScazzocchioC (2004) Double-joint PCR: a PCR-based molecular tool forgene manipulations in Wlamentous fungi. Fungal Genet Biol41:973–981
123





























