Embed Size (px)
Citation preview

CIinical and Experimental Pharmacology & PhysioIogy (1987) 14, 437-447
AUTORADIOGRAPHIC LOCALIZATION OF RECEPTORS IN THE MAMMALIAN CARDIOVASCULAR SYSTEM
R. J. Summers, P. Molenaar, 1. A. Stephenson and C. R. Jones* Department of Pharmacology, University of Melbourne, Parkville, Victoria, Australia
(Received 26 February 1987)
SUMMARY
1. Autoradiographic techniques have been used to examine the location of p- adrenoceptors in the heart and P-adrenoceptors, substance P receptors and muscarinic cholinoceptors in blood vessels.
2. Both &-adrenoceptors and &adrenoceptors were present in guinea-pig and human heart, on the myocardium and associated with the cardiac nerves and blood vessels.
3. Nerves on the vasculature and vascular smooth muscle contained P-adrenoceptors and muscarinic cholinoceptors.
4. Receptors for substance P and 0-adrenoceptors, but not muscarinic cholinoceptors were present on endothelial cells.
Key words: autoradiography, P-adrenoceptors, guinea-pig heart, human heart, muscarinic cholinoceptors, substance P.
INTRODUCTION
The technique of receptor autoradiography has developed from radioligand binding and has a number of important advantages. The retention of the anatomical organization of the tissue allows the identification of areas and often cell types which possess particular receptors (Kuhar 1985), information which is often useful in identifying their roles. Receptor binding techniques are often used to determine changes in density or characteristics of receptors in response to drug treatment, disease or pathological changes. Attempts to link changes in the receptor population with a response in heart or blood vessels may be complicated if the receptor under study is not only linked to this response but also several others in different cells. The presence of receptors on different cell types will be revealed clearly by autoradiography. The other main advantage of autoradiography is sensitivity since areas of receptor concentration as small as 10 pm in diameter can be identified and receptors in small amounts of tissue can be studied. This advantage is of particular importance for human tissues which are often available only in small amounts.
The presence of P-adrenoceptors in the heart has been known for many years (Lands et al.
Correspondence: Dr R. J. Summers, Department of Pharmacology, University of Melbourne, Parkville,
* Present address: Dr C. Richard Jones, Department Innere Medizin, Abteilung fur Kardiologie, Kan- Vic. 3052, Australia.
tonsspital, Basel, Switzerland, CH-4031.

438 R. J. Summers et al.
1967). More recent functional studies using selective agonists (Johansson & Persson 1983; Molenaar & Summers 1987) or radioligand binding studies with selective antagonists (Minnemann et al. 1981; Nahorski 1981; Stiles et al. 1984) have demonstrated P,-adrenoceptors as well as PI- adrenoceptors in the heart of most mammalian species. In some species such as man (Robberecht et al. 1983), cat (Carlsson et al. 1972, 1977; Ablad et al. 1976) and guinea-pig (Hedberg et al. 1980; Molenaar & Summers 1987) stimulation of either P-adrenoceptor subtype leads to a posi- tive inotropic or chronotropic effect, whereas in others such as the rat (Juberg et al. 1985) these responses are only elicited by P,-adrenoceptor stimulation. There are therefore indications at least for the P,-adrenoceptor population that they perform functions in addition to mediation of the positive inotropic and chronotropic response. Determination of the location of the P-adrenoceptor subtypes could therefore provide useful indications of the functions that these receptors perform. tors perform.
In blood vessels 6-adrenoceptors may be associated with muscle where they mediate increases in CAMP accumulation, Ca2+ sequestration, and relaxation (Taira et al. 1977), and with nerves where they facilitate transmitter release (Rand et al. 1980, Majewski 1983). Receptors of several types may also be located on the vascular endothelium where they mediate the release of endothelium-derived relaxing factor (EDRF) which is thought to be important in relaxation of blood vessels to substances such as acetylcholine (Ach), substance P and catecholamines (Furch- gott & Zawadzki 1980; Furchgott et al. 1981; Rubanyi & Vanhoutte 1985). In these vascular prepa- rations the amounts of tissue involved are so small that the sensitivity of the autoradiographic technique makes it the method of choice for the study of endothelial receptors.
METHODS
The methods used to produce and label microscope slide mounted tissue sections are essentially the same as those described previously (Molenaar et al. 1987; Stephenson & Summers 1987). Briefly, guinea-pig atria, human atrial appendage, papillary muscle and mammary artery, dog carotid arteries and rabbit pulmonary arteries were frozen in isopentane cooled in liquid nitrogen and mounted onto cold cryostat chucks using OCT embedding medium. To test the endothelium de- pendence of binding in blood vessels, endothelium was removed in some segments of blood ves- sels by gently threading a piece of roughened polyethylene tubing through the lumen. Serial 10 l m sections were cut and mounted onto gelatinized microscope slides. Labelling was carried out at 25°C and the labelled sections placed in apposition to emulsion-coated coverslips (Kodak NTB3) and exposed at 4°C for 2-10 days ([lZ5I] labelled ligands) or 1-3 months (['HI-ligands). The pho- tographic medium was developed and fixed and sections of blood vessels stained with methylene blue (0.5% in borax) and cardiac tissues stained with pyronin Y (1Vo). For cardiac tissues every fifth slide mounted section was set aside and stained with haeniatoxylin and eosin for histological examination. In cardiac sections P-adrenoceptors were labelled with [1251]-CYP (Molenaar et al. 1987) and in blood vessel sections P-adrenoceptors, muscarinic cholinoceptors and substance P receptors were labelled using [lzsI]-CYP, [3H]-QNB and [1251]-BHSP as described previously (Stephenson & Summers 1987). In biochemical studies of [IzsI]-CYP binding, sections were labelled in the same way as for autoradiography except that the sections were wiped from the slides using Whatman GF/B filters and counted in a Packard gamma counter (model 5301 at an efficiency of 79%). Results of these experiments were analysed using the computer programs EBDA (McPher- son 1983) which performed preliminary Scatchard, Hill and Hofstee analysis and produced a file for subsequent analysis using the iterative curve fitting program LIGAND (Munson & Rodbard 1980).

Localization of receptors in cardiovascular system 439
Drugs used were: (-) and ( +)-propranolol, ICI 118,551 (erythro-DL-l(7-methylindan-4-xylox~)- 3-isopropylamino butan-2-01; Imperial Chemical Industries). CGP 207 12A (2-hydroxy-5-(2-( (2- hydroxy-3-(4-( ( 1-methyl-4-trifluoro methyl) 1 H-imidazole-2-yl)-phenoxy)propyl)amino)ethoxy)- benzamide monomethane sulphonate; Ciba-Geigy); R0363 ( (-)-[ (3/,4’-dihydroxyphenoxy)-2- hydroxy-3-(3~’,4~’-dimethoxyphenethylamino)-propane]oxalate) was a gift from Dr E. Malta of the Victorian College of Pharmacy Ltd; procaterol hydrochloride (Warner Lambert); Na [L251] (Amersham International); (-) cyanopindolol (Sandoz, Basle). Substance P (Boehringer Mann- heim); atropine sulphate, methylene blue (BDH); [1251]-BHSP was a gift from Dr E. Burcher, Deakin University. [1251]-CYP was prepared from (-)CUP and [1251] Na (Lew & Summers 1985).
RESULTS
@-Adrenoceptors in guinea-pig heart
Binding of [1251]-CYP to slide mounted sections of guinea-pig atria was saturable, of high affinity (KD 32 pmol/l left atria; 25 pmol/l right atria) to a population of non-interacting sites (nH values 1.01 and 0.98) with a mean density of 41 and 40 fmol/mg protein (Molenaar et al. 1987). Delineation of the PI- and P,-adrenoceptors was carried out in competition experiments using the highly selective P,-adrenoceptor antagonist CGP 20712A and the &adrenoceptor selective antagonist ICI 118,551. Competition curves obtained with these two antagonists are biphasic (Fig. 1) and can be analysed using LIGAND to give affinities of the competitors for the high and low affinity site and the proportions of each site. The pKD values for CGP 20712A were 8.85 and 4.29 and for ICI 118,551 9.00 and 6.85 at the high and low affinity binding sites. Similar values were obtained for the left and right atria and the sites revealed by CGP 20712A correspond to PI- and &adrenoceptors while the reverse is true for ICI 118,551. The proportions of sites with the properties of P1- and &adrenoceptors were 85 and 15% in both left and right atria.
-12 - 8 - L - 12 - 8 -L Log concentrotion ( rno\ / l )
Fig. 1. Mean competition binding curves for [1251]-CYP and the &-adrenoceptor antagonist CGP 2071244 (0) and P,-adrenoceptor antagonist ICI 118,551 (A) at [1251]-CYP binding sites. The binding curves were biphasic and were characterized by low pseudo Hill coefficients. Further analysis with the non-linear curve fitting program LIGAND revealed two binding sites in the proportions of 85:15% in both preparations with the properties of PI- and &-adrenoceptors,
respectively. From Molenaar et al. (1987).

440 R . J. Summers et al.
Autoradiography was used to map the distribution of P-adrenoceptors in sections of guinea- pig heart (Molenaar et al. 1987). Concentrations of ICI 118,551 (70 nmol/l) were chosen to block selectively &adrenoceptors and CGP 207 12A (100 nmol/l) to selectively block &-adrenoceptors based on affinities determined in the competition experiments. (-)-Propranolol (I pmol/l) was used to define non-specific binding. The micrograph of the haematoxylin and eosin stained sec- tion containing the atrioventricular bundle shows a number of structures (Fig. 2a). High levels of binding of (-)-[1251]-CYP are found over the right atrium (RA), right ventricle (RV), and AV bundle as well as the nerve associated with the AV bundle (Fig. 2b). However, the binding of the ligand to the P-adrenoceptor subtypes is not evenly distributed over these structures. Recep- tors over the myocardium of the RA and RV are predominantly P1-adrenoceptors as shown by the darkfield micrographs in Fig. 2c and d. However, receptors over the AV node and the ner- vous tissue contained much higher proportions of &adrenoceptors. Incubation of sections labelled with [1251]-CYP in the presence of (-)-propranolol (1 pmol/l) showed no localization of grains (Fig. 2e). These studies indicate that the AV bundle and the nerves associated with it contain a higher proportion of 0,-adrenoceptors than the surrounding myocardium. The superior and inferior vena cava shown in Fig. 3 contained predominantly &adrenoceptors. Receptors are local- ized over the adventitia and there is a relatively low density associated with the medial smooth muscle. In the vena cava there is also a concentration of &adrenoceptors associated with the vascular endothelium.
P-Adrenoceptors in the human heart
Similar types of experiments have been carried out using human cardiac tissues. The results of the biochemical experiments again illustrate the presence of P1- and P,-adrenoceptors but the proportion of &adrenoceptors is much higher than in the guinea-pig. The pKD values for CGP 20712A in ventricular papillary muscle were 9.10 and 5.41 at P1- and P,-adrenoceptors, respec- tively. The values obtained for ICI 118,551 were 6.39 and 8.90 (Jones et al. unpubl. data). The affinities obtained were therefore comparable to those seen in the guinea-pig atria. However, the proportion of 0,-adrenoceptors in the papillary muscle ranged from 34 to 37%. Autoradiographs of the distribution of P1- and P,-adrenoceptors in sections of papillary muscle indicated both recep- tor types located over the cardiac muscle cells and in the small intramyocardial blood vessels a concentration of P,-adrenoceptors was observed over the intimal surface (Fig. 4). However, in the human cardiac tissue the major concentrations of both PI- and @,-adrenoceptors are found over the myocardium. Both the PI- and the P,-adrenoceptors are clearly linked to a functional response and maximal positive inotropic and chronotropic responses can be produced by isoprena- line, the &-adrenoceptor selective agonist R0363 and the 6,-adrenoceptor selective agonist procaterol (Fig. 5).
Receptors in blood vessels
Autoradiographic localization of receptors in sections of blood vessels has shown that there are receptors associated with the endothelium, with the smooth muscle cells and with the nerves and other structures in the adventitia (Lipe & Summers 1986). Of particular importance was the ability of the autoradiographic technique to examine directly the locations of receptors which are known to be involved in endothelium-dependent relaxation. Localization of 0-adrenoceptors was examined in sections of human mammary artery. As shown in Fig. 6A and B P-adrenoceptors were associated predominantly with the endothelium. Substance P receptors were labelled in sec- tions of dog carotid artery using [lZSI]-BHSP and again showed a high degree of localization to
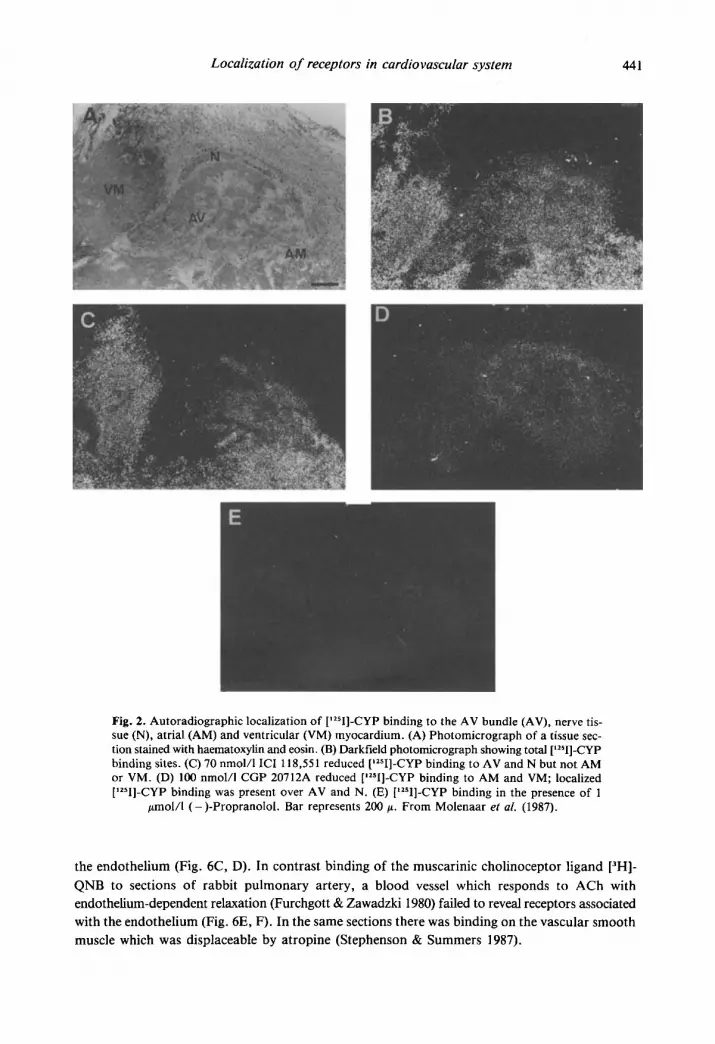
Localization of receptors in cardiovascu far system 441
Fig. 2. Autoradiographic localization of ['2sI]-CYP binding to the AV bundle (AV), nerve tis- sue (N), atrial (AM) and ventricular (VM) myocardium. (A) Photomicrograph of a tissue sec- tion stained with haematoxylin and eosin. (B) Darkfield photomicrograph showing total ['2sI]-CYP binding sites. (C) 70 nmol/l ICI 118,551 reduced ['zsI]-CYP binding to AV and N but not AM or VM. (D) 100 nmol/l CGP 20712A reduced ['*sI]-CYP binding to AM and VM; localized [1251]-CYP binding was present over AV and N. (E) ['2sI]-CYP binding in the presence of 1
fimol/l (-)-Propranolol. Bar represents 200 f i . From Molenaar ef ai. (1987).
the endothelium (Fig. 6C, D). In contrast binding of the muscarinic cholinoceptor ligand [3H]- QNB to sections of rabbit pulmonary artery, a blood vessel which responds to ACh with endothelium-dependent relaxation (Furchgott & Zawadzki 1980) failed to reveal receptors associated with the endothelium (Fig. 6E, F). In the same sections there was binding on the vascular smooth muscle which was displaceable by atropine (Stephenson & Summers 1987).

442 R. J. Summers et al.
Fig. 3. Autoradiographic localization of [12sI]-CYP binding to endothelial (EN), media (ME) and adventitial (AD) cells of the superior vena cava and atrial myocardium (AM). (A) Pho- tomicrograph of a section stained with haematoxylin and eosin. The darkfield photomicrograph (B) shows the total distribution of [12sI]-CYP binding over EN, ME, A D and AM. In compari- son with ME, AM has a much higher density of binding sites. [lzsI]-CYP binding in the presence of 70 nmol/l ICI 118,551 was reduced over EN, ME and AD; however, a high and even distri- bution of binding sites remained over AM (C). In the presence of 100 nmol/l CGP 20712A (D) [1z51]-CYP binding sites remained over EN, ME and AD. A low and even distribution of binding sites was present over ME. [1251]-CYP binding in the presence of 1 fimol/l (-)-propranolol
is shown in (E). Bar represents 100 pm. From Molenaar et al. (1987).
DISCUSSION
Autoradiography has a number of important advantages over radioligand binding techniques for the study of receptors. The increased sensitivity allows the study of receptors in small areas of

Localization of receptors in cardiovascuiar system 443
Fig. 4. Light microscopic autoradiographic localization of (-)['zsI]-CYP binding to sections of left ventricular papillary muscle. Panel (A) shows a photomicrograph of a haematoxylin and eosin stained section showing a coronary artery and the surrounding myocardium. Panels (B-E) are darkfield micrographs of sections incubated with (-) [Iz51]-CYP in the absence (B) or presence of 70 nmol/l ICI 118,551 (C) or 100 nmol/l CGP 20712A (D) or 1 pmol/l (-)-propranolol (E) and apposed to emulsion coated coverslips. Note the high density of binding sites over the my- ocardium (MY) and the absence of binding over the media (ME) of the coronary artery. There are receptors of the &-subtype (B, D) over the endothelial surface (EN). Scale bar represents
tissue and the localization of receptors to particular regions or cells within a tissue provides use- ful information in determining the processes these receptors control. In the guinea-pig heart PI- adrenoceptors are associated with cells of myocardial origin whereas &-adrenoceptors were also found on cardiac nerves, the endothelium of the great veins and the surface of the aortic valve as well as the adventitial surface of the cardiac blood vessels (Molenaar et al. 1987). The p2-
100 pm.

444
100 - s I
3 E E
E
.- 2 5 0 -
01 C
0 C
Lz e s
.- -
R . J. Summers et al.
-
1
adrenoceptors on myocardial cells are functional in the guinea-pig since a positive inotropic and chronotropic response is obtained to the &adrenoceptor agonist procaterol (Molenaar & Sum- mers 1987) although the response is weak compared with that obtained by stimulation of PI- adrenoceptors. The role of P,-adrenoceptors in the cardiac vasculature is not clear although & adrenoceptors have been shown to mediate vasodilatation in guinea-pig pulmonary artery (O’Don- nell & Wanstall 1985). Receptors on the nerves may either be involved in presynaptic regulation of transmitter release or may be receptors undergoing axonal transport along the nerve (Zarbin et al. 1983). The role of the &adrenoceptors found in other areas is unclear at present.
In the human tissue studied, the major population of both PI- and 0,-adrenoceptors is found on the myocardial cells. This correlates well with the observation that the response obtained to the stimulation of P,-adrenoceptors in human atrial appendage is as great as that to stimulation of &adrenoceptors. The phenomenon of isoprenaline-induced tachycardia, resistant to PI- adrenoceptor blockade is well documented (Arnold et al. 1985; Brown et al. 1983) and it is likely that this resistant response is mediated through 6,-adrenoceptors.
The results of the experiments which localized receptors on blood vessels raise a number of important points. The presence of high levels of BHSP binding localized to the endothelium of the dog carotid artery is in agreement with previous reports which show that this preparation is particularly sensitive to substance P (D’Orleans-Juste et al. 1985; Satoh et al. 1986). Both the binding (Stephenson & Summers 1987) and the response (D’Orleans-Juste et al. 1985) are endothelium-dependent. The correlation between endothelium-dependent relaxation and p- adrenoceptors is not as well established. It has been reported that dog coronary arteries relax in response to isoprenaline and that a component of this response is endothelium-dependent (Rubanyi & Vanhoutte 1985). The results reported here demonstrate &-adrenoceptors at the in- timal surface of human coronary arteries and the mammary artery, a blood vessel which is in- creasingly used in cardiac bypass surgery. In the case of the mammary artery the binding is endothelium-dependent (Summers et al. unpubl. data) and isoprenaline produces a relaxation of this preparation in vitro (Malta, pers. comm.) Autoradiographic studies of muscarinic cholinocep- tors associated with endothelium produced an interesting result. In a total of 14 different arteries from six species, all of which are known to respond to ACh with endothelium-dependent relaxa- tion, no evidence was obtained for receptors associated with the endothelium. In a number of

Localization 0 f receptors in cardiovascular system
A
445
Fig. 6. Autoradiographic localization of (top to bottom): 0-adrenoceptors in human mammary artery, substance P receptors in dog carotid artery, and muscarinic cholinoceptors in rabbit pul- monary artery. Panels A, C and E are bright field micrographs of sections of each blood vessel and panels B, D and F show the corresponding autoradiographic localization of receptors. Bar
represents 100 pm (A, C) and SO Fm (E). E, F from Stephenson and Summers (1987).
blood vessels, including the rabbit pulmonary artery, receptors were clearly present on the medial smooth muscle concentrated towards the adventitial surface (Fig. 6). These autoradiographic find- ings are in agreement with studies which have been unable to demonstrate binding of [‘HI-QNB or release of EDRF from cultured endothelial cells treated with muscarinic agonists (Peach et 02. 1985; Loeb et al. 1985). Similar findings have been reported for endothelial cells grown on sephadex beads which do not respond to ACh but release EDRF in response to the calcium iono- phore A23187 and bradykinin (Cocks et al. 1985). This suggests that ACh may produce endothelium-dependent relaxation of blood vessels by acting on the smooth muscle cells to release

446 R. J. Summers et al.
a substance which in turn releases EDRF from the endothelial cells. This view is supported by the recent observation that the muscarinic cholinoceptor agonist methacholine failed to elevate cGMP levels in cultures of vascular smooth muscle or endothelial cells but is effective in mixed cultures containing both types of cell (Loeb et al. 1985).
In conclusion, the autoradiographic technique provides some useful insights into the roles of receptors in the cardiovascular system. In the heart both PI- and &adrenoceptors are present and in most species activation of either receptor subtype produces positive inotropic and chronotropic responses. However, the autoradiographic studies clearly show that a proportion of the &adrenoceptors fulfil other functions associated with the vasculature and innervation. In blood vessels receptors are found associated with nerves, smooth muscle and with the endothelium.
ACKNOWLEDGMENTS
This study was supported by a grant from the National Heart Foundation of Australia. PM is a NH & MRC Biomedical Research Scholar.
REFERENCES
Ablad, B., Cadsson, E., Dahlof, C. & Ek, L. (1976) Some aspects of pharmacology of 8-adrenoceptor blockers. Drugs, 11 (suppl. l), 100-111.
Arnold, J.M.O., O’Connor, P.C., Riddell, J.G., Harron, D.W.G., Shanks, R.G. & McDevitt, D.G. (1985) Effects of the PI-adrenoceptor antagonist ICI 118,551 on exercise tachycardia and isoprenaline induced /3-adrenoceptor responses in man. British Journal of Clinical Pharmacology, 19, 619-630.
Brown, J.E., McLeod, A.A. & Shand, D.G. (1983) Evidence for cardiac PI-adrenoceptors in man. Clinical Pharmacology and Therapeutics, 33, 424-428.
Carlsson, E., Ablad, B., Brandstrom, A. & Carlsson, B. (1972) Differentiated blockade of the chronotropic effects of various adrenergic stimuli in the cat heart. Life Sciences, 11, 953-958.
Carlsson, E., Dahlof, C.-G., Hedberg, A., Persson, H., & Tangstrand, B. (1977) Differentiation of cardiac chronotropic and inotropic effects of P-adrenoceptor agonists. Naunyn-Schmiedeberg’s Archives of Phar- macology, 300, 101-105.
Cocks, T.M., Angus, J.A., Campbell, J.A. & Campbell, G.R. (1985) Release and properties of endothelium derived relaxing factor (EDRF) from endothelial cells in culture. Journalof CelIPhysiology, 123,310-320.
D’Orleans-Juste, P., Dion, S., Mizrahi, J., & Regoli, D. (1985) The effect of peptides and non-peptides in iso- lated arterial smooth muscles: role of endothelium. European Journal of Pharmacology, 114, 9-21.
Furchgott, R.F. & Zawadzki, J.V. (1980) The obligatory role of endothelial cells in the relaxation of vascular smooth muscle by acetylcholine. Nature, 288, 373-376.
Furchgott, R.F., Zawadzki, J.V. & Cherry, P.D. (1981) Role of endothelium in the vasodilator response to acetylcholine. In: Vasodilatation, (Ed. P.M. Vanhoutte and I. Leusen). Raven Press, New York.
Hedburg, A., Minneman, K.P. & Molinoff, P.B. (1980) Differential distribution of beta-1 and beta-2 adrenergic receptors in cat and guinea-pig heart. Journal of Pharmacology and Experimental Therapeutics, 212,
Johansson, L.-H. & Persson, H. (1983) PI-adrenoceptors in guinea-pig atria. Journal of Pharmacy and Phar- macology, 35, 804-807.
Juberg, E.N., Minneman, K.P. & Abel, P.W. (1985) PI- and PI-adrenoceptor binding and functional response in right and left atria of rat heart. Naunyn-Schmiedeberg’s Archives of Pharmacology, 330, 193-202.
Kuhar, M.J. (1985) Receptor localization with the microscope. In: Neurotransmitter Receptor Binding, 2nd edn, (Ed. H.I. Yamamura, S.J. Enna & M.J. Kuhar), p. 153. Raven Press, New York.
Lipe, S. & Summers, RJ. (1986) Autoradiographic analysis of the distribution of 6-adrenoceptors in the dog
Lands, A.M., Arnold, A., McAuliff, J.P., Luduena, F.P. & Brown, T.G. (1967) Differentiation of receptor
503-508.
splenic vasculature. British Journal of Pharmacology, 87, 603-609.
systems activated by sympathomimetic amines. Nature, 214, 597-598.

Localization of receptors in cardiovascular system 447
Lew, R. & Summers, R.J. (1985) Autoradiographic localization of 6-adrenoceptor subtypes in guinea-pig
Loeb, A.L., Owens, G.K. &Peach, M.J. (1985) Evidence for endothelium derived relaxing factor in cultured
Majewski, H. (1983) Modulation of noradrenaline release through activation of presynaptic 0-adrenoceptors.
McPherson, G.A. (1983) A practical computer based approach to the analysis of radioligand binding experi-
Minnemann, K.P., Pittman, R.N. & Molinoff, P.B. (1981) 0-adrenergic receptor subtypes: properties, distri- bution and regulation. Annual Review of Neuroscience, 4, 419-461.
Molenaar, P., Canale, E. & Summers, R.J. (1987) Autoradiographic localization of PI- and &adrenoceptors in guinea-pig atrium and regions of the conducting system. Journal of Pharmacology and Experimen- tal Therapeutics, (in press).
Molenaar, P. & Summers, R.J. (1987) Characterisation of PI- and &adrenoceptors in guinea-pig atrium: functional and receptor binding studies. Journal of Pharmacology and Experimental Therapeutics, (in press).
Munson, P.J. & Rodbard, D. (1980) LIGAND: a versatile computerized approach for the characterization of ligand binding systems. Analytical Biochemistry, 107, 220-239.
Nahorski, S.R. (1981) Identification and significance of beta-adrenoceptor subtypes. Trends in Pharmaco- logical Sciences, 2, 95 -98.
O’Donnell, S.R. & Wanstall, J.C. (1985) Responses to the &-selective agonist procaterol of vascular and atrial preparations with different functional P-adrenoceptor populations. British Journal of Pharmacology,
Peach, M.J., Singer, H.A. & Loeb, A.L. (1985) Mechanisms of endothelium-dependent smooth muscle relax- ation. Biochemical Pharmacology, 34, 1867-1874.
Rand, M.J., Majewski, H., Medgett, I.C., McCulloch, M.W. & Story, D.F. (1980) Prejunctional receptors modulating autonomic neuroeffector transmission. Circulation Research, 46, 170-176.
Robberecht, P., Delhaye, M., Talon, C., et al. (1983) The human heart 0-adrenergic receptors 1: heterogeneity of the binding sites: presence of 50% beta, and 50% beta,-adrenergic receptors. Molecular Pharma-
Rubanyi, G. & Vanhoutte, P.M. (1985) Endothelium removal decreases relaxations of canine coronary arter- ies caused by 6-adrenergic agonists and adenosine. Journal of Cardiovascular Pharmacology, 7 , 139-144.
Satoh, K., Angus, J.A. & Cocks, T.M. (1986) Alpha,-adrenoceptors and endothelium-dependent relaxation in canine large arteries. Journal of Molecular and Cellular Cardiology, 18 (suppl. 51), 293.
Stephenson, J.A. & Summers, R.J. (1987) Autoradiographic localization of receptors on vascular endotheli- um. European Journal of Pharmacology, 134, 35-43.
Stiles, G.L., Caron, M.G. & hfkowitz, R.J. (1984) 0-adrenergic receptors: biochemical mechanisms of phys- iological regulation. Physiological Reviews, 64, 661-743.
Taira, N., Yabuuchi, Y. & Yamashita, S. (1977) Profile of P-adrenoceptors in femoral, superior mesenteric and renal vascular beds of dogs. Britkh Journal of Pharmacology, 59, 577-583.
Zarbin, M.A., Palacios, J.M., Wamsley, J.K. & Kubar, M.J. (1983) Axonal transport of @-adrenergic recep- tors. Antero- and retrogradely-transported receptors differ in agonist affinity and nucleotide sensitivi- ty. Molecular Pharmacology, 24, 341-348.
kidney. British Journal of Pharmacology, 85, 341 -348.
cells. Hypertension, 7, 804-807.
Journal of Autonomic Pharmacology, 3,47-60.
ments. Computer Programs in Biomedicine, 17, 107-1 14.
84, 221-235.
cology, 24, 169-173.





























