Embed Size (px)
Citation preview

[CANCER RESEARCH 53, 106-111. January 1. 1993]
Autoreactive and Heat Shock Protein 60-recognizing CD4+ T-Cells Show AntitumorActivity against Syngeneic Fibrosarcoma1
Manioru Harada, Goro Matsuzaki,2 Yasunobu Yoshikai, Noritada Kobayashi, Shin Kurosawa, Hiroaki Takimoto, and
Kikuo Nomoto
Department of Immunology, Medical Institute of Bioregulation. Kyushu University, Fukuoka 812 ¡M.H., G. M., N. K., S. K., H. T., K. N.¡,and Laboratory of Cermfree Life,Research Institute for Disease Mechanism and Control, Nagoya University School of Medicine, Nagova 466 IY. Y.j, Japan
ABSTRACT
A <'1)4' heat shock protein (hsp) 60-recognizing autoreactive T-cell line
(BASL1) and clone (BASL1.1) were examined for their antitumor activityagainst major histocompatibility complex class li syngeneic Meth A fibrosarcoma (Meth A), which was immunofluorescently stained with monoclonal antibody specific for hsp 60. In in vitro proliferativi' assay, BASL1.1was suggested to recognize Meth A-derived hsp 60 presented by syngeneicantigen-presenting cells in a major histocompatibility complex class II-
restricted manner. This cell line and clone showed antitumor activity intumor-neutralizing (Winn) assay. BASL1 and BASL1.1 cells producedy-interferon, tumor necrosis factor, and interleukin 2 but not interleukin
4 by the stimulation with syngeneic spleen cells. In cytolytic assay, thesecell lines and clones showed neither direct nor indirect (bystander) cytol-
ysis against Meth A. In cytostatic assay, these cells inhibited the proliferation of Meth A in the presence of syngeneic macrophages, and thisactivity was abrogated by the addition of anti-y-interferon monoclonalantibody. Recombinant y-interferon could induce cytostatic activity only
in the presence of macrophages, and tumor necrosis factor synergized thisactivity. Antitumor activity induced by BASL1 was abrogated by theadministration of anti-CD8 monoclonal antibody in vivo, suggesting thatCDS* cytotoxic T-lymphocytes are essential and final effector cells forBASL1 -mediated Meth A rejection. These findings indicate that CD4+
autoreactive and hsp 60-recognizing T-cells show two types of antitumoractivity: cytostasis and induction of tumor-specific cytotoxic T-lymphocytes. Furthermore, these results imply that tumor-specific immunitycould be elicited by CD4* helper T-cells which recognize hsp.
INTRODUCTION
It has been reported that tumor cells express various degrees ofTAA3 (1, 2). However, only a few tumors were identified by their
unique TAAs (3-5), resulting in difficulty in the application of im-
munotherapy to cancer. Furthermore, the notable finding that a majortumor rejection antigen of P815 mastocytoma is identical to the geneproduct expressed in normal syngeneic mice (6) impresses on us thedifficulties of immunotherapy using TAAs.
Recently, several tumor cells have been reported to express highlevels of hsp (2, 7-9). hsp is known to be a highly conserved protein
from prokaryote to eukaryote and to be induced in cells by varioustypes of stress, including transformation (10-12), suggesting a possibility that hsp can be a so-called common tumor antigen. In fact, some
Received 6/1/92; accepted 10/15/92.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by a grant to G. M. from the Ministry of Education, Science and
Culture, and grants to Y. Y from the Ministry of Education. Science and Culture, theMinistry of Health and Welfare, Sapporo Research Foundation, Fukuoka Cancer Society,and Special Coordination Funds of the Science and Technology Agency of the JapaneseGovernment. This work also received financial support from the Comprehensive 10-year
Strategy for Cancer Control and the UNDP/World BankAVHO Special Program forResearch and Training in Tropical Disease.
2 To whom requests for reprints should be addressed.' The abbreviations used are: hsp. heat shock protein; SC, spleen cells; PPD, purified
protein derivative; LN, lymph node; TAA(s), tumor-associated antigen(s); MHC, majorhistocompatibility complex; F1TC, fluorescein isothiocyanate; CTL(s), cytotoxic T-lym-
phocyte(s); mAb, monoclonal antibody; IL, interleukin; IFN, interferon; TNF, tumornecrosis factor; Meth A, Meth A fibrosarcoma; HBSS, Hanks' balanced salt solution;
dThd, deoxythymidine.
TAAs were revealed to be hsp (2, 7). According to these observations,we supposed that hsp can be used as a "common" antigen to achieve
an efficient immunotherapy against various tumors even when TAAsare not identified.
In the present study, we examined the antitumor activity of a CD4*
hsp 60-recognizing autoreactive T-cell line and clone against MHCclass II~ syngeneic Meth A fibrosarcoma, which was immunofluores
cently stained with mAb specific for hsp 60, and confirmed that theseT-cells show two types of antitumor activities: cytostatic activity andthe induction of tumor-specific CTLs.
MATERIALS AND METHODS
Mice. Eight- to 10-week-old female BALB/c (H-2d, Lyt2.2*, Thy 1.2+),C57BL/6 (H-2b), and C3H/He (H-2k) mice were purchased from Japan SLC
(Shizuoka, Japan). All mice were bred in specific pathogen-free conditions.
Cell Preparation. To prepare SC, spleens were removed aseptically andteased into suspension in complete culture medium. Peritoneal exúdate cellswere obtained by lavage of the peritoneal cavity with 10 ml of HBSS. The cellswere washed and then adjusted to proper concentrations in complete culturemedium. RPMI 1640 (Gibco, Grand Island, NY) supplemented with 10%heat-inactivated fetal calf serum (HyClone, Logan, UT), 5 x 10~5 M 2-mer-
captoethanol, 20 ITIM4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid, 30
ug/ml gentamicin (Schering Corporation, Kenilworth, NJ), and 0.2% sodiumbicarbonate was used as the complete culture medium.
Autoreactive T-Cell Line and Clones. We established an autoreactiveT-cell line, BASLI, as described previously (13), and two clones were isolated
through the limiting dilution technique by plating at a concentration of 0.3cells/well. Because these two clones showed similar cell surface phenotype andproliferative response in vitro, we used one of them, called BASL 1.1, in thisstudy.
Tumors. Meth A is a 3-methylcholanthrene-induced fibrosarcoma of BALB/c
origin. It is maintained in an ascites form in syngeneic BALB/c mice by weeklypassages. Before experiments, B-cells and macrophages were removed fromMeth A suspension by the incubation in plastic dishes (Falcon 3003; Becton-Dickinson, Lincoln Park, NJ) coated with goat anti-mouse Ig (G -t-M) mAb(Tago, Inc., Burlingame, ÇA) for 45 min at 37°C.Removal of B-cells and
macrophages was confirmed by flow cytometric analysis.Proliferation Assay of Autoreactive T-Cells. Proliferation of cells was
assayed with standard [3H]dThd incorporation and scintillation counting.
Briefly, autoreactive T-cells were collected 14 days after the last stimulation.
The viable cells were collected by density gradient centrifugation by usingLympholyte M (Cedarlane, Ontario, Canada). Two hundred thousand viableBASL1 or BASL1.1 were seeded in each well of flat-bottomed 96-well culture
plates (Corning Glass Works, Corning, NY) in a total volume of 0.2 ml with2 x IO5 30 Gy-irradiated SC for 72 h. During the last 8 h of the culture, 37 kBqof ['H]dThd was added to each well. Cells were then harvested and incorporation of [3H]dThd was counted with a Beta Plate system (Pharmacia LKB
Biotechnology, Uppsala, Sweden). To block the proliferation of these cells,anti-IAd mAbs (mouse IgG2a; Meiji Institute of Health Science, Tokyo, Japan)or anti-IAb-k mAbs (mouse IgG2a; Meiji Institute of Health Science) were
added to the culture at 1:400 dilution. To analyze the proliferation of BASL1and BASL1.1 to hsp 60 derived from Mycobacterium bovis (kindly providedfrom Dr. R. van der Zee, National Institute of Public Health and EnvironmentalProtection, Bilthoven, the Netherlands) and PPD of Mycobacterium tuberculosis Aoyama B (BCG, Inc., Tokyo, Japan), hsp 60 and PPD were added toeach well at 5 ug/ml.
106
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTITUMOR ACTIVITY OF hsp «(-RECOGNIZING T-CELLS
To analyze the proliferative response of BASL1.I Io Meth A tumor cells. 2x 10s BASLI.l and 1 x IO4 mitomicine C (Kyowa Hakko Kogyo, Co., Ltd.,
Tokyo, Japan)-treated Meth A tumor cells were seeded in each well of flat-
bottomed 96-well culture plates (Corning Glass Works) in a total volume of 0.2ml with or without 2 x IO5 30 Gy-irradiated BALB/c SC for 48 h. During thelast 8 h of the culture. 37 kBq of [*H]dThd were added to each well, cells wereharvested, and the incorporation of |'H]dThd was counted with a Beta Plate
system (Pharmacia LKB Biotechnology). To block the proliferation of thesecells, anti-H-2Dd mAbs (mouse IgG2a; Meiji Institute of Health Science) oranti-IAd mAbs (mouse IgG2a; Meiji Institute of Health Science) were added to
the culture at 1:400 dilution. Mitomycin C (Kyowa Hakko Kogyol-treated
Meth A tumor cells were prepared by coculture with 100 ug/ml mitomycin C
for 60 min.Tumor-neutralizing (Winn) Assay. The antitumor activities of BASLI
and BASLI.l were assayed by modified Winn's technique (14). Briefly. Meth
A tumor cells were injected s.c. into the right abdomen with or without effectorcells in a volume of 0.2 ml. After tumor inoculation, tumor growth wasinspected every 3 or 4 days, and tumor acceptance was determined on day 30.
Assay of Cytostasis. Cytostatic activity was determined by the inhibitionof DNA synthesis of Meth A. To prepare adherent macrophages, peritonealexúdate cells (2 x lOVwell) were incubated in 96-well flat-bottomed plates(Corning 25860) for 90 min at 37°Cfollowed by washing with completemedium three times. Cell suspensions containing 1 x 10J Meth A tumor cells
were cultured with or without 2 x IO5 BASL1 or BASLI.l and/or adherent
cells prepared from peritoneal exúdate cells in 0.2 ml in flat-bottomed micro-tiler plates (Corning 25860) in a humidified atmosphere at 5% CO2. 37°Cfor20 h. Cultures were pulsed with 37 kBq of |'H]dThd during the last 2 h of theincubation. The cultures were harvested, and incorporation of |'H]dThd was
counted with a Beta Plate system (Pharmacia LKB Biotechnology). To evaluate the contribution of IFN-y in cytostasis, hamster anti-murine IFN--y mAb
(hamster IgG; Genzyme. Boston. MA) was added to each well at 10ug/ml. Tosearch the cytostatic mechanism, Meth A tumor cells were incubated with orwithout recombinant mouse IFN--y (Genzyme) and/or recombinant mouse
TNF-a (Genzyme) with or without adherent cells.
Cytokine Assay. TNF activity of culture supernatants was assayed by theirability to inhibit the proliferation of murine L929 fibroblasts (15). The TNFunit was determined by probit analysis, in which 1 unit of TNF activity resultsin 50<7r inhibition of growth of L929 cells. IFN-y concentration was deter
mined by two-site sandwich enzyme-linked immunosorbent assay (16) withmonoclonal and polyclonal anti-IFN-y antibodies (Genzyme). IL-2 and IL-4activities were determined by using HT-2 cells, an IL-2- and IL-4-dependentcell line. HT-2 cells ( 1 x IO4 cells/well) were cultured for 20 h in the presence
of 2-fold diluted supernatants in 96-well flat-bottomed plates (Corning 25860).Cultures were pulsed with 37 kBq of ['H]dThd during the last 4 h of theincubation. IL-2 and IL-4 activities were determined by the ['HjdThd uptake
in the presence of anti-IL-4 mAb (Collaborative Research. Inc.. Bedford. MA)or anti-IL-2 mAb (Collaborative Research. Inc.) at the concentration of 3
ug/ml, respectively.Flow Cytometric Analysis. Expression of MHC class I. MHC class II. and
hsp 60 on Meth A tumor cells was analyzed by FACScan (Becton Dickinson.Mountain View. CA) as follows. Meth A tumor cells were incubated for 15 minin complete medium supplemented with 5% rabbit serum to block Fc receptor.FITC-conjugated anti-H-2Kd MAb (mouse IgM; Meiji Institute of Health
Science) was used for the analysis of MHC class I expression. For the analysisof MHC class II and hsp 60 expression on the cell surface. Meth A tumor cellswere treated with anti-I-Ad mAb (mouse IgG2a: Meiji Institute of Health
Science) or anti-hsp 60 mAb (ML30; kindly provided by Dr. J. Ivanyi. Royal
Postgraduate Medical School, London), which recognize both mycobacterialand murine hsp 60 (17). for 30 min at 4°C.These cells were stained withsecondary antibody FITC-labeled affinity-purified F(ab')2 fragments of anti-
mouse IgG (Tago, Inc.). Labeled cells were analyzed by FACScan (BectonDickinson) and fluorescence histograms were accumulated on a logarithmicscale. Staining background was determined by cells treated only with secondary antibody. In some cases, Meth A tumor cells were fixed with 10%paraformaldehyde before blocking Fc receptor.
Depletion of CDS* T-Cell Subset in BALB/c Mice. To deplete the CDS*T-cell subset in BALB/c (Lyt 2.2*. Thy 1.2*) mice, anti-CD8 (anti-Lyt2.2;
mouse IgM. Meiji Institute of Health Science) mAb (50 ug in 0.5 ml of HBSS)was injected i.v. into BALB/c mice on days -3. -2. and -1. As control mAb.
anti-Thyl.l (mouse IgM; Meiji Institute of Health Science) mAb (50 ug in0.5 ml of HBSS) was injected i.v. into BALB/c mice on days -3. -2. and -1.Depletion of CD8* T-cells by the administration of anti-Lyt2.2 mAb was
confirmed by flow cytometric analysis.Statistics. The statistical significance of the data was determined using the
Student's t test. A P value of less than 0.05 was considered to be statistically
significant.
RESULTS
Characterization of an Autoreactive T-Cell Line and Clone.
BASLI cells proliferate in response to the syngeneic BALB/c SC butnot to the allogeneic C57BL/6 or C3H SC (Fig. 1; Ref. 13). Theproliferation to the syngeneic SC was blocked in the presence ofanti-lAd mAb but not in the presence of anti-IAK k mAb, showing that
the autoreactivity of BASLI is restricted to MHC class II. Interestingly, the proliferation of BASLI to the syngeneic BALB/c SC wassignificantly enhanced by the presence of mycobacterial hsp 60, amajor component of PPD. or PPD. The presence of unrelated antigensuch as ovalbumin could not enhance the autoreactivity of BASLI(data was not shown), hsp and PPD induced no proliferative responseof BASLI in the presence of allogeneic stimulator cells. These resultssuggest that this cell line recognizes endogenous A/r 65,000 hsp in thecontext of IAd, as previously reported ( 13). To examine this specificity
more strictly, we established two clones from the BASLI cell line andanalyzed their antigen recognition. Because both of them are CD4*Vß6*T-cells and showed the same specificity (data not shown), one
of them, BASLI.l, was used in this study. As shown in Fig. 1,BASLI.l showed the same proliferative response as BASLI. indicating that the BASLI.l clone is a representative of the BASLI cell line.
Flow Cytometric Analysis of Meth A Tumor Cells. We examinedMHC expression of Meth A tumor cells. Meth A tumor cells expressMHC class I molecules (Fig. 2A ) but not MHC class II molecules(Fig. 2B). The incubation with recombinant mouse IFN-y (250 units/
ml) for 24 h could not induce Meth A tumor cells to express MHCclass II molecules (data not shown).
Stimulation
(-)
BALBc
C57BL6
C3H
BALB/cBALB'C
BALB'C
C57BL6
BALB'C
C57BL/6
Ag or Ab
Stimulation Ag or AbBASL1.1
BALBc
C57BU6
BALBc
BALB/c
BALBc
C57BL/6
BALB/c
C57BL76
X103 (cpm)
Fig. 1. Proliferative response of an autoreactive T-cell line and clone. BASLI andBASLI.l cells (2 x IO5) were incubated with 2 x IO5 irradiated stimulator SC with orwithout mAb or antigen for 72 h. ( 'HJdThd was added to the cultures 8 h before harvest.
The data are expressed as the mean ±SD of four samples, hsp 60 and PPD were derivedfrom Mycohacterium hmis and Mycobacierium luben-ulnsis. respectively.
107
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTITUMOR ACTIVITY OF hsp 60-RECOCNEING T-CELLS
0).0
3
CDO
0>
(50>DC
IB1 10* 10
anti-H-2Kdla" la1 na2 la
anti-IA
anti-hsp 60
Fig. 2. Flow cytometric analysis of Meth A tumor cells with mAb specific for MHCmolecules and hsp 60. A, Meth A tumor cells directly stained with FITC-conjugatedanti-H-2Ku mAb. contro! background. B, Meth A tumor cells indirectly stained withanti-I-Ad mAb, followed by FITC-conjugated Fiab'h fragment of goat anti-mouse IgG.
Nonfixed Meth A (C) and fixed Meth A (D) with 10% paraformaldehyde/phosphate-buffered saline were indirectly stained with anti-hsp 60 mAb followed by FITC-conjugated F(ab' )Tfragments of goat anti-mouse IgG. In the case of indirect staining, the doited
lìneaindicate control staining with secondary mAb alone. Data were displayed by histograms on a logarithmic scale.
Recently, several tumor cells have been reported to express highlevels of hsp (2, 7-9). Furthermore, a TAA of Meth A was suggested
to be a member of the hsp 90 family (7). Then, we tried to determinewhether or not Meth A cells express a member of the hsp 60 family.As shown in Fig. 2C, flow cytometric analysis revealed that a part ofMeth A tumor cells were stained with mAb specific for hsp 60,suggesting that Meth A tumor cells express hsp 60 on their cellsurface. Furthermore, paraformaldehyde-fixed Meth A tumor cells
were highly stained with this mAb (Fig. 2D), showing that Meth Atumor cells constitutively express hsp 60 intracellularly.
Proliferative Response of BASL1.1 against Meth A. We attempted to determine whether the autoreactive hsp 60-recognizingCD4* T-cell clone (BASL1.1) respond against hsp-expressing Meth A
tumor cells in vitro (Fig. 3). BASL1.1 could not respond to MHC classI!" surface hsp 60* Meth A tumor cells in the absence of irradiated
syngeneic SC (group 5), suggesting that BASL1.1 fails to recognizehsp 60 expressed on cell surface directly. In contrast, the addition ofmitomycin C-treated Meth A remarkably augmented the proliferative
response of BASL1.1 against syngeneic SC (P < 0.01; group 7), andthis proliferation was blocked not by the presence of anti-H-2Dd mAbbut of anti-IAd mAb. These results indicate that BASL1.1 recognizes
Meth A-derived hsp 60 presented by syngeneic antigen-presentingcells in a MHC class II-restricted manner.
Tumor-neutralizing Activity of BASL1 and BASL1.1 Cells. Weexamined antitumor activity of autoreactive and hsp 60-recognizingT-cells against syngeneic Meth A fibrosarcoma by Winn assay. As
shown in Table 1, more than 70% of BALB/c mice rejected Meth Awhen they were coinoculated with 1 x IO5 Meth A and 1 x IO5 or 1x IO6 of BASL1, while almost all BALB/c mice accepted 1 x IO5Meth A coinoculated with naive unseparated or CD4+ LN cells. Fur
thermore, BASL1.1 showed Meth A-neutralizing activity similar to
that of BASL1.Cytokine Production of BASL1 and BASL1.1 Cells. To inves
tigate the mechanism of the antitumor effect of the autoreactive
hsp 60-recognizing T-cells, we examined the cytokine production
of BASL1 and BASL1.1 cells. The supernatants of BASL1 andBASL1.1 stimulated with SC or mitogen were examined. As shown inTable 2, BASL1 and BASL1.1 secreted IFN-y and TNF when they
were stimulated with the syngeneic BALB/c SC or concanavalin A.Secretion of IFN-y and TNF by BASL1.1 cells stimulated withBALB/c SC was inhibited by anti-IAd mAb.
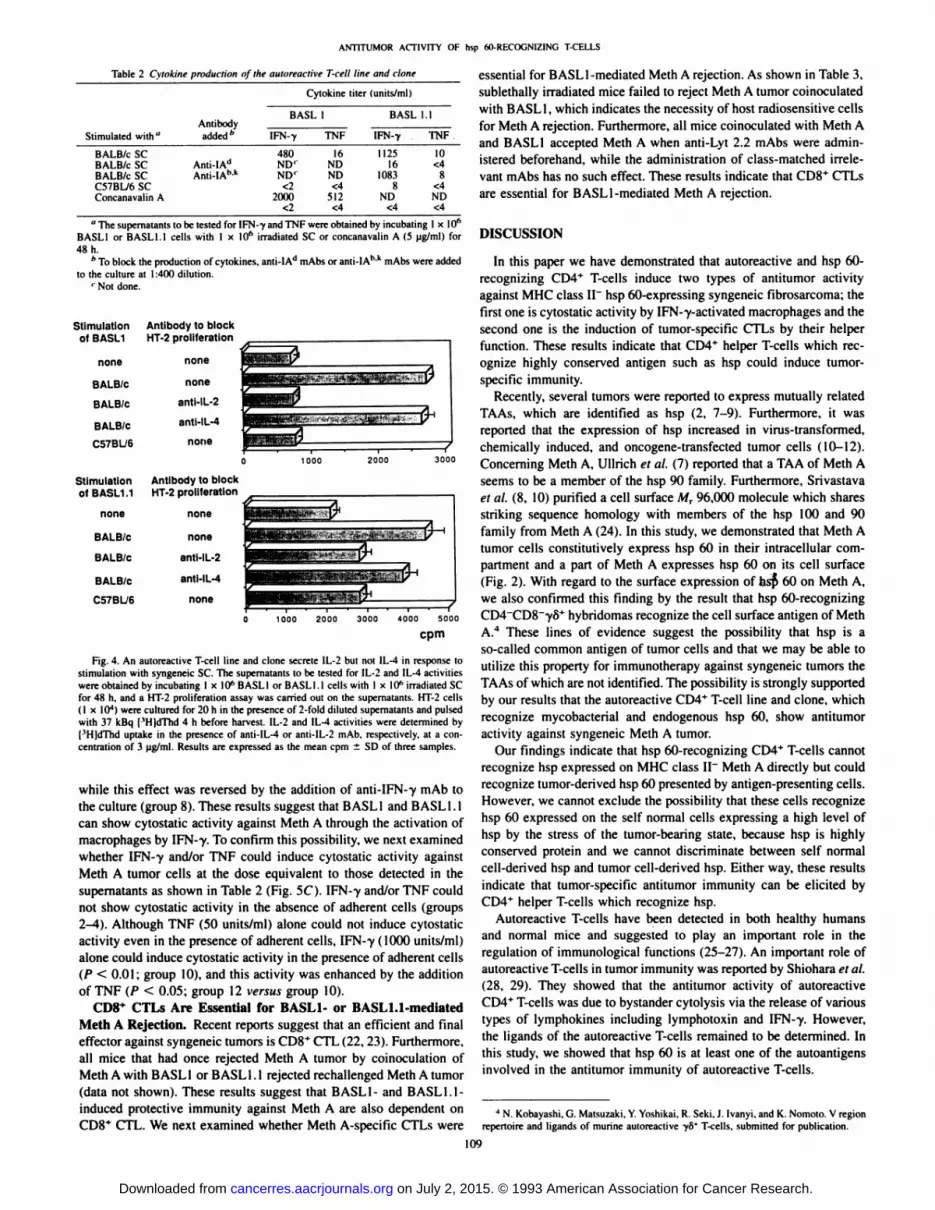
As shown in Fig. 4, BASL1 and BASL1.1 cells secreted IL-2 by thestimulation with syngeneic SC, while they did not secrete IL-4. Recentstudies have subdivided CD4* helper T-cells into Th 1 or Th2 subsets
by their secretion pattern of cytokines (18). BASL1 and BASL1.1 canbe assigned to the Th 1 subset by their secretion pattern of cytokines.
Cytostatic Activity of BASL1 and BASL1.1 Cells. To furtherexamine the antitumor mechanism of these cells, cytolytic and cyto-
static activities were examined. BASL1 and BASL1.1 could not lyseMHC class II " Meth A tumor cells but showed a low level of cytolytic
activity (15%) against syngeneic MHC class II* targets, lipopolysac-charide-stimulated B-cell blasts, in a 8-h 5lCr release assay (effector:
target ratio = 30:1). BASL1 and BASL1.1 showed no indirect(bystander) lytic activity against Meth A in the presence of syngeneiclipopolysaccharide blasts (data not shown).
We have reported that a cytostatic, but not cytolytic, mechanismplayed a principal role in antitumor resistance in the syngeneic BALE/c-Meth A system (19-21). We next investigated the cytostatic activity
of BASL1 and BASL1.1 against Meth A tumor (Fig. 5, A and B).BASL1 (or BASL1.1) alone showed no significant cytostatic activityagainst Meth A (group 4), probably because of the inability to recognize MHC class II" Meth A directly. Macrophages alone showed a low
level of cytostatic activity against Meth A (P < 0.05; group 5). In thepresence of both BASL1 (or BASL1.1) and macrophages, however,proliferation of Meth A was extremely decreased (P < 0.01 ; group 7),
Group BASL1.1 BALB/cSC
Meth A mAb
Fig. 3. Proliferative response of BASLI.I against Meth A tumor cells. BASL1.1 cells(2 x IO5) were incubated with I x IO4 mitomycin C-treated Meth A tumor cells with orwithout irradiated syngeneic SC or mAb for 48 h. [ 'HJdThd was added to the cultures
8 h before harvest. Results were expressed as the mean cpm ±SD of three samples.
Table I Meth A tumor-neutralising activity of BASL1 and BASLI.I cells
Tumor acceptance on day30"Experiment12Meth
A1x 10*
1 x IO51 x IO51 x IO51 xIO52
x IO52 x IO52 x IO5Coinoculaled
cellsBASL1
(1 x IO5)BASLI (1 x IO6)Whole LN cells (1 x IO5)CD4-LN cells *(1 xIO5)BASLI.I
<2x IO5)BASLI. 1 <2x IO6)Tumor-accepted
mice/tumor-inoculatedmice
(%)6/7
(86)2/7 (29)IH (29)6/6(100)5/6(83)5/5(100)
2/5 (40)1/5 (20)
" Meth A tumor cells were injected s.c. into the right abdomen with or without
indicated effector cells. After inoculation, tumor growth was inspected every 3 or 4 days,and tumor acceptance was determined on day 30.
''To prepare CD4* LN cells, nylon-wool nonadherent LN cells were treated with
anti-Lyt2.2 mAb plus C.
108
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
ANTITUMOR ACTIVITY OK hsp MI-RECOGNIZING T-CELLS
Table 2 Cylnkine production of Ihr aulorf active T-cell Une ami ciane
Cytokine liter(units/ml)Stimulated
with"BALB/c
SCBALB/c SCBALB/c SCC57BL/6 SCConcanavalin AAntibody
added*Anti-IAd
Anti-IAhABASLIFN-Y480ND'ND'
<22000
<21TNF16
NDND<4
512<4BASL
1IFN-y1125
161083
8ND
<4.1TNF10
<48
<4ND<4
" The supematants to be tested for IFN-y and TNF were obtained by incubating 1 x IO6BASL1 or BASL1.I cells with I x 10" irradiated SC or concanavalin A (5 ug/ml) for
48 h.''To block the production of cytokines, anti-IAd mAbs or anti-IAh'k mAbs were added
to the culture at 1:40(1 dilution.' Not done.
Stimulation Antibody to blockof BASL1 HT-2 proliferation
none
BALB/c
BALB/c
BALB/c
C57BL/6
none
none
anti-IL-2
anti-IL-4
none
1000 2000
Stimulation Antibody to blockofBASL1.1 HT-2 proliferation
none
BALB/c
BALB/c
BALB/c
C57BÜ6
none
none
anti-IL-2
anti-IL-4
none
Fig. 4. An autoreactive T-cell line and clone secrete 1L-2 but
stimulation with syngeneic SC. The supematants to be tested forwere obtained by incubating 1 x 10" BASLI or BASLI.l cells wifor 48 h, and a HT-2 proliferation assay was carried out on the(I x IO4) were cultured for 20 h in the presence of 2-fold dilutedwith 37 kBq I'HldThd 4 h before harvest. IL-2 and IL-4 activit['H]dThd uptake in the presence of anti-IL-4 or anti-IL-2 mAb,
centratimi of 3 ug/ml. Results are expressed as the mean cpm ±
cpm
not IL-4 in response toIL-2 and IL-4 activitiesth I x 10" irradiated SCupernalants. HT-2 cellssupematants and pulsed
were determined by. respectively, at a con-
SD of three samples.
while this effect was reversed by the addition of anti-IFN-y mAb to
the culture (group 8). These results suggest that BASLI and BASLI.lcan show cytostatic activity against Meth A through the activation ofmacrophages by IFN-y. To confirm this possibility, we next examinedwhether IFN-y and/or TNF could induce cytostatic activity against
Meth A tumor cells at the dose equivalent to those detected in thesupematants as shown in Table 2 (Fig. 5C). IFN-y and/or TNF could
not show cytostatic activity in the absence of adherent cells (groups2-4). Although TNF (50 units/ml) alone could not induce cytostaticactivity even in the presence of adherent cells. IFN-y ( 1000 units/ml)
alone could induce cytostatic activity in the presence of adherent cells(P < 0.01; group 10), and this activity was enhanced by the additionof TNF (P < 0.05; group 12 versus group 10).
CD8+ CTLs Are Essential for BASLI- or BASLI.1-mediated
Meth A Rejection. Recent reports suggest that an efficient and finaleffector against syngeneic tumors is CD8+ CTL (22, 23). Furthermore,
all mice that had once rejected Meth A tumor by coinoculation ofMeth A with BASLI or BASL 1.1 rejected rechallenged Meth A tumor(data not shown). These results suggest that BASLI- and BASLI.1-
induced protective immunity against Meth A are also dependent onCDS* CTL. We next examined whether Meth A-specific CTLs were
essential for BASL 1-mediated Meth A rejection. As shown in Table 3,
sublethally irradiated mice failed to reject Meth A tumor coinoculatedwith BASLI, which indicates the necessity of host radiosensitive cellsfor Meth A rejection. Furthermore, all mice coinoculated with Meth Aand BASLI accepted Meth A when anti-Lyt 2.2 mAbs were administered beforehand, while the administration of class-matched irrelevant mAbs has no such effect. These results indicate that CD8+ CTLs
are essential for BASL I-mediated Meth A rejection.
DISCUSSION
In this paper we have demonstrated that autoreactive and hsp 60-recognizing CD4* T-cells induce two types of antitumor activityagainst MHC class II~ hsp 60-expressing syngeneic fibrosarcoma: the
first one is cytostatic activity by IFN-y-activated macrophages and thesecond one is the induction of tumor-specific CTLs by their helperfunction. These results indicate that CD4+ helper T-cells which rec
ognize highly conserved antigen such as hsp could induce tumor-
specific immunity.Recently, several tumors were reported to express mutually related
TAAs. which are identified as hsp (2, 7-9). Furthermore, it wasreported that the expression of hsp increased in virus-transformed,chemically induced, and oncogene-transfected tumor cells (10-12).
Concerning Meth A, Ullrich et al. (7) reported that a TAA of Meth Aseems to be a member of the hsp 90 family. Furthermore, Srivastavaet al. (8. 10) purified a cell surface Mr 96,000 molecule which sharesstriking sequence homology with members of the hsp 100 and 90family from Meth A (24). In this study, we demonstrated that Meth Atumor cells constitutively express hsp 60 in their intracellular compartment and a part of Meth A expresses hsp 60 on its cell surface(Fig. 2). With regard to the surface expression of hsp 60 on Meth A,we also confirmed this finding by the result that hsp 60-recognizingCD4~CD8~yo+ hybridomas recognize the cell surface antigen of Meth
A.4 These lines of evidence suggest the possibility that hsp is a
so-called common antigen of tumor cells and that we may be able to
utilize this property for immunotherapy against syngeneic tumors theTAAs of which are not identified. The possibility is strongly supportedby our results that the autoreactive CD4+ T-cell line and clone, which
recognize mycobacterial and endogenous hsp 60, show antitumoractivity against syngeneic Meth A tumor.
Our findings indicate that hsp 60-recognizing CD4* T-cells cannotrecognize hsp expressed on MHC class II~ Meth A directly but could
recognize tumor-derived hsp 60 presented by antigen-presenting cells.
However, we cannot exclude the possibility that these cells recognizehsp 60 expressed on the self normal cells expressing a high level ofhsp by the stress of the tumor-bearing state, because hsp is highly
conserved protein and we cannot discriminate between self normalcell-derived hsp and tumor cell-derived hsp. Either way. these resultsindicate that tumor-specific antitumor immunity can be elicited byCD4* helper T-cells which recognize hsp.
Autoreactive T-cells have been detected in both healthy humans
and normal mice and suggested to play an important role in theregulation of immunological functions (25-27). An important role ofautoreactive T-cells in tumor immunity was reported by Shiohara et al.
(28, 29). They showed that the antitumor activity of autoreactiveCD4+ T-cells was due to bystander cytolysis via the release of various
types of lymphokines including lymphotoxin and IFN-y. However,the ligands of the autoreactive T-cells remained to be determined. In
this study, we showed that hsp 60 is at least one of the autoantigensinvolved in the antitumor immunity of autoreactive T-cells.
J N. Kobayashi. G. Matsu/aki. Y. Yoshikai. R. Seki. J. Ivanyi. and K. Nomoto. V regionrepertoire and ligands of murine autoreactive ye* T-cells. submitted for publication.
\m
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Group
1
2
3
4
S
6
7
8
9
ANTITUMOR ACTIVITY OF hsp 60-RECOONIZING T-CELLS
Meth A BASL1 M O mAb
anti-IFN-y
Ham IgG
Fig. 5. Cytostatic activity of BASLI andBASLI.l cells against syngeneic Meth A tumorcells. Meth Acells ( I x IO4) were incubated with orwithout BASLI or BASLI.l (2 x IO"), and adher
ent cells were prepared from peritoneal exúdatecells (2 x 10-Vwelll for 20 h. Preparation of adherent cells was performed as described in "Materialsand Methods." [ 'HjdThd was added to the cultures
2 h before harvest. To block the activity of IFN-y,hamster anti-mouse IFN-y mAb was added to eachwell al IO Mg/ml. To search the cytostatic mechanism. Meth A tumor cells were incubated with orwithout IFN-y 11000 units/ml) and/or TNF (50
units/mil in the absence or presence of adherentcells. Results are expressed as the mean cpm ±SDof three samples.
Group
1
2
3
4
5
6
7
Meth A BASL1.1 MJ mAb
Group
1
2
3
4
5
6
7
8
9
10
1112
Meth A M IFN-y
1000
anti-IFN-v
Ham IgG
TNF
50
50
50
SO
50
50
10X 10 3(cpm)
Table 3 inhibition of BASÃŒJ-mediatedMeth A rejection by sublethal irradiationor anti-CDH mAb administration
Tumor acceptance on day 30"
CoinoculatedcellsBASLI
BASLIBASLI
BASLITreatment350
rad*Anti-Lyt2.2'
Mouse IgM 'Tumor-accepted
mice/tumor-inoculatedmice(%)6/6
(IOO)2/12 (17)6/6 (100)6/6 (100)1/5 (20)
" Meth A tumor cells (2 x IO5) were injected s.c. into the right abdomen with orwithout BASLI cells (2 x IO6) in a volume of 0.2 ml. After inoculation, tumor growth was
inspected on every 3 or 4 days and tumor acceptance was determined on day 30.* Mice were sublethally irradiated 4 h before inoculation.' Mice received i.v. injections of mAb (50 ug in 0.5 ml of HBSSl three times on days
-3, -2, and -I.
Recently, the importance of the local secretion of cytokines inuntitumor immunity has been reevaluated by the experiments ofgene-transfected tumors secreting IFN--y or IL-2 (22, 23, 30). Watanabe et
al. (30) reported that local IFN-y secretion reduces tumorigenicity and
suppresses tumor growth. Furthermore, Fearon et al. (22) suggestedan important principle that the absence of effective in vivo immuneresponse to tumors may be largely due to the failure of helper T-cellsto effectively provide IL-2. rather than the absence of CTL precur
sors capable of recognizing TAAs. This principle is compatible withthe hypothesis that IL-2 secreted from autoreactive and hsp-recog-nizing T-cells in the vicinity of tumor cells is useful for the induction
of antitumor CTLs. However, we have no direct evidence of thishypothesis.
Although cell-mediated cytotoxicity can be detected as two differ
ent types of phenomenon, namely, cytolysis and cytostasis (31 ), manyreports seem to focus on the former activity in tumor immunity. Inprevious studies, we found that cytostasis played a principal role in theantitumor immunity in the syngeneic BALB/c mouse-Meth A tumorsystem (19-21). As shown in this report, autoreactive and hsp-recog-nizing T-cells showed cytostatic activity against Meth A fibrosarcoma
in the presence of adherent macrophages. Although autoreactive andhsp-recognizing T-cells secrete a moderate level of IFN--y and a low
level of TNF by the stimulation with syngeneic SC (Table 2), IFN-7
no
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTITUMOR ACTIVITY OF hsp 60-RECOGNIZING T-CELLS
and/or TNF at a dose equivalent to those detected in the supernatantsshowed no direct cytostatic activity against Meth A tumor cells in theabsence of macrophages (Fig. 5C). In contrast, IFN-y could show
cytostatic activity in the presence of macrophages, and its activity wasaugmented by the addition of TNF, suggesting that the main mechanism of cytostasis is due to IFN-y-activated macrophages, and TNF
could show a synergic effect. However, we cannot neglect the possibility that other cytokines are induced by IFN-y and participate in the
cytostatic mechanism.Recent studies have subdivided the CD4* T-cells into two mutually
exclusive subsets: the Thl subset that produces IL-2 and IFN-y andthe Th2 subset that produces IL-4 and IL-5 (18). The relative role of
these two subsets in antitumor immunity is not clear. The autoreactivehsp 60-recognizing CD4+ T-cell line was assigned to the Thl group.
Interestingly, Nagarkatti el al. (32) reported that the tumor-infiltratingCD4+ T-cells have the characteristics of Thl cells. Taken together, the
Thl cells may participate in tumor rejection.In conclusion, we demonstrated that autoreactive and hsp 60-
recognizing CD4* T-cells show antitumor activity against syngeneic
fibrosarcoma. These results may indicate a possibility of novel anti-tumor immunotherapy by CD4* helper T-cells which recognize a"common" antigen such as hsp. Therefore, we are further studying
whether or not these T-cells show antitumor activity against syngeneic
tumors other than Meth A.
ACKNOWLEDGMENTS
We specially thank Dr. R. van der Zee for providing mycobacterial hsp 60and Dr. J. Ivanyi for providing anti-hsp 60 mAb (ML30).
REFERENCES
1. LeGrue. S. J., Pellis. N. R., Riley. L. B., and Kahan. B. D. Biochemical characterization of l-butanol-extracted murine tumor-specific transplantation antigens. CancerRes.. 45:3164-3172, 1985.
2. Srivastava, P. K.. DeLeo. A. B., and Old. L. J. Tumor rejection antigens of chemicallyinduced sarcomas of inbred mice. Proc. Nati. Acad. Sci. USA, 83:3407-3411. 1986.
3. Old. L. J. Cancer immunology: the search for specificity. Cancer Res., 4/.-361-375,
1981.4. Hearing. V. J.. Gresten. D. M.. Montague, P. M. Vieira, W. D.. Gaietto. G., and Law.
L. W. Murine melanoma-specific tumor rejection activity elicited by a purified,melanoma-associated antigen. J. Immunol., /J7:379-384, 1986.
5. Sato. N., Yagishita, A.. Okubo, M.. Torigoe, T.. Takahashi, S., Sato. T., and Kikuchi.K. Characterization of tumor rejection antigen molecules of chemically inducedmurine colon tumor C-26. Cancer Res., 47:3147-3151, 1987.
6. Eynde, B. V., Lethe, B., Pel, A. V., Plaen. E. D.. and Boon. Y. The gene coding for amajor tumor rejection antigen of tumor P8I5 is identical to the normal gene ofsyngeneic DBA/2 mice. J. Exp. Med., 137:1373-1384. 1991.
7. llrich, S. J.. Robinson, E. A.. Law. L. W.. Willingham. M.. and Appella. E. A mousetumor-specific transplantation antigen is a heat shock-related protein. Proc. Nail.Acad. Sci. USA, 8^:3121-3125, 1986.
8. Maki, R. G., Old. L. J.. and Srivastava, P. K. Human homologue of murine tumorrejection antigen gp96: 5.-regulately and coding regions and relationship to stress-induced proteins. Proc. Nati. Acad. Sci. USA. «7:5658-5662, 1990.
9. Fisch, P., Malkovsky. M., Kovats, S.. Sturm. E.. Braakman. E.. Klein, B. S., Voss. S.D.. Morrissey, L. M., DeMars. R.. Welch, W. J.. Bolhuis. R. L. H.. and Sondel, P. M.Recognition by human V-y9/VS2 T cells of a groEL homolog on Daudi Burkitt's
lymphoma cells. Science (Washington DC). 259:1269-1273, 1990.10. Srivastava. P. K., and Maki. R. G. Stress-induced proteins in immune response to
cancer. Curr. Top. Microbiol. Immunol.. /67.-109-I23. 1991.
11. Young. R. A., and Elliott. T. J. Stress proteins, infection and immune surveillance.Cell, 59:5-8, 1989.
12. Konno. A.. Sato. N., Yagishita. A.. Torigoe, T., Cho, J.. Torimoto. K., Hará.!.. Wada.Y.. Okubo. M.. Takahashi. N.. and Kikuchi. K. Heat- or stress-inducible transformation-associated cell surface antigen on the activated H-ra.s oncogene-transfected ratfibroblast. Cancer Res., 49:6578-6582. 1989.
13. Matsuzaki, G., Yoshikai, Y., Harada, M., and Nomoto, K. Autoreactive T cells fromnormal mice recognize mycobacterial 65 kd heat-shock protein from Mvcobacteriumbons. Int. Immunol., 3:215-220. 1991.
14. Winn. H. J. Immune mechanisms in Homotransplantation. II. Quantitative assay of theimmunologie activity of lymphoid cells stimulated by tumor homograft. J. Immunol..86:228-295. 1961.
15. Aggarwal, B., Kohr, W. J., Hass, P. E., Mofft, B., Spencer, S. A.. Henzel. W. J..Bringman, T. S., Nedwin, G. E., Goeddel. D. V., and Harkins, R. N. Human tumornecrosis factor: production, purification and characterization. J. Biol. Chem..260:2345-2354, 1985.
16. Cherwinski, H. M., Schumacher, J. H., Brown, K. D., and Mosmann, T. R. Two typesof mouse helper T cells clone. III. Further differences in lymphokine synthesisbetween Thl and Th2 clones revealed by RNA hybridization, functionally monospe-cific bioassays and monoclonal antibodies. J. Exp. Med., /66:1229-1244. 1987.
17. Ivanyi, J., Sinha. S., Aston, R., Cussell, D.. Keen, M., and Sengupta, U. Definition ofspecies specific and cross-reactive antigenic determinants of Mycahacteriutn lepraeusing monoclonal antibodies. Clin. Exp. Immunol.. 52:528-536, 1983.
18. Mosmann. T. R.. Cherwinski. H.. Bond, M. W., Giedlin. M. A., and Coffmann, R. L.Two types of murine helper T cell clone. I. Definition according to profiles oflymphokine activities and secreted proteins. J. Immunol., 756:2348-2357, 1986.
19. Milani. M.. Mori. K.. Himeno. K.. Matsumoto. Y.. Taniguchi. K., and Nomoto. K. Therole of cytostasis in tumor immunity: comparison between syngeneic and allogene»systems. Cell. Immunol.. 92:22-30, 1985.
20. Matsumoto, T., Himeno, K., Milani, M.. Mori, K.. Miake, S., and Nomoto. K.Differing contribution of various effector cells in the elimination of syngeneic orallogene» cells. J. Clin. Lab. Immunol., /9:83-89, 1986.
21. Milani, M.. Mori. K., Himeno, K., and Nomoto, K. Anti-tumor cytostatic mechanismand delayed-type hypersensitivity against a syngeneic murine tumor: comparison
between neonalally thymectomized mice and congenially athymic nude mice. J.Immunol.. /42.-2148-2I54, 1989.
22. Fearon, E. R., Pardoll. D. M.. haya. T.. Golumbek. P.. Levitsky. H. I., Simons. J. !..Karasuyama. H.. Vogelstein, B., and Forest, P. Interleukin-2 production by tumor cellsbypasses T helper function in the generation of an antitumor response. Cell, 60:397-
403. 1990.23. Gansbacher, B„Zier. K.. Daniels, B„Cronin, K.. Bannerji. R., and Gilboa, E.
Interleukin 2 gene transfer into tumor cells abrogates tumorigenicity and inducesprotective immunity. J. Exp. Med.. / 72:1217-1224, 1990.
Lotze, M. T., and Finn, O. J. Recent advances in cellular immunology: implicationsfor immunity to cancer. Immunol. Today. //:I9(>-193, 1990.
Nagarkatti, P., Snow. E. C.. and Kaplan, A. M. Characterization of autoreactive Tlymphocyte clones isolated from normal, unprimed mice. Cell. Immunol., 94:32^48.1985.
26. Kotani. H., Mitsuya. H., Jarrett, R. F., Yenokida, G. G., James, S. P., and Strober. W.An autoreactive T cell clone that can be activated to provide both helper and suppressor function. J. Immunol.. ¡3d:1951-1959, 1986.
27. Tilkin, A. F.. Michon. J., Juy, D.. Kayibanda. M.. Henin. Y., Sterkers, G.. Beluel. H.,and Levy, J. P. Autoreaclive T cell clones of MHC class 11specificities are producedduring responses against foreign antigens in man. J. Immunol.. /.î8:674-679. 1987.
28. Shiohara. T.. Ruddle. N. M., Horowitz. M.. Moellmann, G. E.. and Lemer, A. B.Anti-tumor activity of class II MHC antigen-restricted cloned autoreactive T cells. I.Destruction of B16 melanoma cells mediated bystander cytolysis m vivo. J. Immunol.,138:1971-1978, 1987.
29. Shiohara. T., Moellmann, G. E.. Jacobson. K., Kuklinska. E., Ruddle, N. H., andLcmer, A. B. Anti-tumor activity of class II MHC antigen-restricted cloned autoreactive T cells. II. Novel immunotherapy of B16 melanoma by local and systemicadoptive transfer. J. Immunol.. 1JX: 1979-1986. 1987.
30. Watanabe. Y., Kuribayashi, K., Miyatake, S., Nishihara. K., Nakayama, E.. Taniyama.T.. and Sakata, T. Exogenous expression of mouse interferon cDNA in mouse neuroblastoma C1300 celts results in reduced tumorigenicity by augmented anti-tumorimmunity. Proc. Nati. Acad. Sci. USA, 86:9456-9460, 1989.
31. Farram, E., and Festenstein, H. Mechanism of "cytostasis" of tumors in vivo by
syngeneic lymphoid cells of tumor bearers. Cell. Immunol., 44:352-366. 1979.32. Nagarkatti. M.. Clay, S. R., and Nagarkatti, P. S. Characterization of tumor-infiltrating
CD4* T cells as Thl cells based on lymphokine secretion and functional properties.J. Immunol.. /44.-4898-4905. 1990.
Ill
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1993;53:106-111. Cancer Res Mamoru Harada, Goro Matsuzaki, Yasunobu Yoshikai, et al. FibrosarcomaT-Cells Show Antitumor Activity against Syngeneic
+Autoreactive and Heat Shock Protein 60-recognizing CD4
Updated version
http://cancerres.aacrjournals.org/content/53/1/106
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on July 2, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























