Embed Size (px)
Citation preview

Vol. 49, No. 2APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Feb. 1985, p. 279-2880099-2240/85/020279-10$02.00/0Copyright C 1985, American Society for Microbiology
Bacterial 0-Methylation of Chloroguaiacols: Effect of SubstrateConcentration, Cell Density, and Growth Conditions
ANN-SOFIE ALLARD, MIKAEL REMBERGER, AND ALASDAIR H. NEILSON*Swedish Environmental Research Institute, Box 21060, S-100 31 Stockholm, Sweden
Received 13 September 1984/Accepted 2 November 1984
0-methylation of chloroguaiacols has been examined in a number of gram-positive and gram-negativebacteria to elucidate the effects of substrate concentration, growth conditions, and cell density. Substrateconcentrations between 0.1 and 20.0 mg liter-1 were used, and it was found that (i) yields of the 0-methylatedproducts were significantly higher at the lowest concentrations and (ii) rates of 0-methylation were not linearfunctions of concentration. With 3,4,5-trichloroguaiacol, the nature of the metabolites also changed withconcentration. During growth with a range of substrates, 0-methylation of chloroguaiacols also took place.With vanillate, however, de-O-methylation occurred: the chlorocatechol formed from 4,5,6-trichloroguaiacolwas successively 0-methylated to 3,4,5-trichloroguaiacol and 3,4,5-trichloroveratrole, whereas that producedfrom 4,5-dichloroguaiacol was degraded without 0-methylation. Effective 0-methylation in nonproliferatingsuspensions occurred at cell densities as low as 105 cells ml-', although both the yields and the rates were lowerthan in more dense cultures. By using disk assays, it was shown that, compared with their precursors, all ofthe 0-methylated metabolites were virtually nontoxic to the strains examined. It is therefore proposed that0-methylation functions as a detoxification mechanism for cells exposed to chloroguaiacols and chlorophenols.In detail, significant differences were observed in the response of gram-positive and gram-negative cell strainsto chloroguaiacols. It is concluded that bacterial 0-methylation is to be expected in the natural environmentsubjected to discharge of chloroguaiacols.
Tests for biodegradability occupy a central position inprocedures for assessing the impact of chemicals dischargedinto the environment. Chlorinated organic compounds suchas polychlorinated biphenyls (14) and chlorophenols (1) haveattracted particular interest on account of their apparentpersistence. A variety of polychlorinated guaiacols andcatechols (references in reference 23) are formed during theproduction of fully bleached chemical pulp; they have beenfound in the discharge from aerated lagoons handling kraftmill effluent (22) and in water and sediment samples from theGulf of Bothnia (T.-M. Xie, Ph.D. thesis, Goteborg Univer-sity, Goteborg, Sweden, 1984). It may therefore be con-cluded that these compounds are not readily biodegradable.Although it has been shown that, during degradation of
halogenated aromatic compounds, metabolites may be pro-duced which are toxic to the cells producing them (refer-ences in reference 4), less attention has been paid to thepossibility that the products of microbial metabolism mightbe toxic to other components of the ecosystem such as fish.We have previously shown, in laboratory experiments withbacteria, that various chlorinated phenols and guaiacolswere 0-methylated to the corresponding anisoles and vera-troles (29). In a further study (30), we observed that thesemetabolites possessed a number of potentially hazardousbiological properties (e.g., bioconcentration potential, tox-icity to embryos and larvae of zebra fish [Brachydaniorerio]) which had not been observed in their precursors.
It was therefore important to assess the environmentalsignificance of bacterial 0-methylation by taking into con-sideration a number of factors which were not resolved inthe previous study. Among the most important are thefollowing: (i) the effect of the substrate concentration, whichwill be much lower in receiving waters than those used in the
* Corresponding author.
earlier laboratory experiments; (ii) the possibility that 0-methylation could occur during growth of the cells, which isimportant in view of the wide range of potential substratesavailable under natural conditions, and the possibility thatonly some of these might support 0-methylation; and (iii) theeffect of cell density (the earlier experiments used onlydense cell suspensions which will not be encountered natu-rally).The results of experiments designed to resolve these
issues are given here. The rates of 0-methylation were notlinear functions of the substrate concentrations, and it wasshown that, for the transforming cells, the phenolic com-pounds were appreciably more toxic than the metabolitesproduced, the corresponding 0-methyl ethers. It is thereforeproposed that, for cells carrying out 0-methylation, thisreaction serves as a detoxification mechanism. Experimentswith cells growing at the expense of a range of substratesshowed that 0-methylation generally occurred: for gram-pos-itive organisms growing with vanillate, however, a numberof other transformations took place with the less toxicsubstrates. Attention is drawn to significant differences inthe behavior of gram-positive and gram-negative organismstowards chlorinated guaiacols, and an attempt is made toassess the environmental significance of these observations.
MATERIALS AND METHODS
Isolation and characterization of strains. Pure strains ofbacteria were used throughout these studies. The isolationand characterization of strains from three localities havealready been described (29). Strains were isolated from threeadditional samples of Baltic Sea and freshwater lake sedi-ments by similar procedures, using methane (strains 1637,1678), methylamine (strains 1623, 1624, 1631, 1632), and3,4-dimethoxybenzoate (strains 1539, 1571) as substrates for
279

280 ALLARD, REMBERGER, AND NEILSON
TABLE 1. Retention times of chloroguaiacol and chlorocatechol0-acetates and 0-trimethylsilyl ethers relative to that of
tetrachloroguaiaol 0-acetateRelative retention time
Substance O-Acetate 0-Trimethylsilylether
3,4,5-Trichloroguaiacol 0.73 0.684,5,6-Trichloroguaiacol 0.79 0.82Tetrachloroguaiacol 1.00 0.984,5-Dichlorocatechol 0.68 0.633,4,5-Trichlorocatechol 0.98 0.94Tetrachlorocatechol 1.29 1.303,4,5-Trichloro-2,6- 1.04 0.89dimethoxyphenol
Pentachlorophenol 0.94 1.01
enrichment. Biochemical characterization of all of the strainswas carried out as previously described (27, 28).
Substrates and metabolites. The synthesis of substratesand metabolites has already been described (29). Theirpurity was assessed before use and was >99% by gaschromatographic (GC) analysis.
Analytical procedures. (i) Phenols, guaiacols, and catechols.The methods used previously had to be modified (a) toovercome problems due to adsorption to bacterial cells whenlow substrate concentrations were used, (b) to analyzecatechols with minimum interference from aerial oxidationduring extraction and analysis, and (c) to remove sufficientamounts of aromatic acids when used as growth substratessince these interfered with analysis of the catechols. Thefollowing method was used for preparation of O-acetates andincorporated the procedure used by Perdue and Wolfe (33)for releasing lipophilic substances from humic acids. Thesample (1.0 ml) was mixed with acetonitrile (0.5 ml) and 0.25ml of a 20% (wt/vol) solution of ascorbic acid and 0.25 ml ofH2SO4 (98%) were added. The mixture was heated at 70°Cfor 30 min and then held for 15 min in an ultrasonic cleaningbath. A mixture of n-hexane-di-isopropyl ether (3:1; 1.5 ml)containing 100 ng each of pentachlorobenzene and 2,4,6-tribromophenol as internal standards was added, and thetubes were shaken for 2 min. The mixture was centrifuged ina clinical centrifuge, the organic phase was removed, and theaqueous phase was extracted once again with 0.5 ml ofn-hexane-di-isopropyl ether lacking internal standards. Thecombined extracts were acetylated by adding ca. 100 mg ofanhydrous sodium acetate followed by acetic anhydride (100,ul). The tubes were heated in a block at 70°C for 15 min andshaken occasionally. After cooling, 0.8 M K2CO3 solution (4ml) was added and the tubes were shaken gently for 5 min.The organic phase was transferred to fresh tubes, acetoni-trile (1.5 ml) was added, and the tubes were shaken for 1min. A solution of 0.05 M K2CO3 was added and the tubeswere shaken vigorously for exactly 1 min. The organic phasewas removed and used for GC analysis as described previ-ously, except that an oven temperature of 195°C was used.Analysis and identification of guaiacols and catechols, pro-duced as metabolites, also used conversion into the 0-hept-afluorobutyrates and 0-trimethylsilyl ethers. For the latter,the sample was extracted with toluene, dried (Na2SO4), andheated with N,O-bis(trimethylsilyl)acetamide (250 pAl) for 15min at 70°C. The reaction mixture was cooled and shakenwith n-hexane-water (1:3) for 30 s, and the organic phasewas removed and used for GC analysis. Relative retentiontimes for 0-acetates and 0-trimethylsilyl ethers of chloro-
guaiacols, chlorocatechols, and pentachlorophenol are givenin Table 1.
(ii) Anisoles, veratroles, and 1,2,3-trichloro-4,5,6-trimethoxybenzene. The sample (1.0 or 2.0 ml) was treatedwith 1 M NaOH and held in an ultrasonic cleaning bath for15 min. A mixture of n-hexane-di-isopropyl ether (3:1; 1.5ml) containing 50 ,ug of lindane liter-' as internal standardwas added, the tubes were shaken for 2 min, the phases wereseparated by centrifugation, and the organic phase was usedfor GC analysis as described previously (29).
Identification of metabolites by GC-mass spectrometrywas carried out as described before (29).
Toxicity of the substrates. The MICROTOX system, usingthe standard protocol (8), was used for the relatively solublechlorocatechols, chloroguaiacols, and chlorophenols. Con-centrated solutions were prepared in water, and the pH wasadjusted to ca. 7.2 before use. Due to lack of a solvent withsuitably low toxicity, the MICROTOX system could not beused for the much less toxic and poorly soluble anisoles andveratroles. Values are given as 50% effective concentration(EC50) after 15 min (15°C).A growth inhibition assay (45), based on standard proce-
dures for determining resistance to antibiotics (3), was usedfor all of the compounds except the readily oxidizablecatechols. Cells were grown in 3-ml volumes of a standardmineral medium (VV 2) containing 4-hydroxybenzoate (1.0 gliter-') as sole carbon source. This culture was used directlyto inoculate plates of tryptone soya broth (Oxoid) solidifiedwith agar (15.0 g liter-'). Concentrated solutions of thesubstances were prepared in n-hexane and applied to sterile5.5-mm paper disks in the following amounts: 5, 10, 25, 75,and 150 ,ug for 3,4,5-trichloroguaiacol and pentachloro-phenol; 25, 75, 150, 225, and 300 p.g for 4,5-dichloro-,4,5,6-trichloro-, and tetrachloroguaiacols and 3,4,5-trichloro-2,6-dimethoxyphenol (3,4,5-trichlorosyringol). Plates wereincubated at 30°C and diameters of zones of inhibition weremeasured after 3 and 7 days. The same method was used fordetermining sensitivity to erythromycin, clindamycin, andlincomycin (15 ,ug disk-'). These antibiotics were chosensince all three function by blocking protein synthesis onribosomes (13) and since resistance to erythromycin ingram-positive bacteria has been shown to involve N,N-dimethylation of adenine in 23S rRNA (39).
Metabolic experiments. All metabolic experiments werecarried out in VV 2 medium and with the precautionsdescribed previously (29).
(i) Experiments with nonproliferating cells. Experimentswith dense cell suspensions were carried out as previouslydescribed (29) and were used (a) to screen a range oforganisms for 0-methylating capacity, using 4,5,6-trichloro-guaiacol at concentrations of 20 mg liter-' and 100 ,ugliter-'; (b) to examine in detail the effect of substrateconcentration, using 3,4,5- and 4,5,6-trichloroguaiacol atconcentrations of ca. 10, 1.0, and 0.1 mg liter-'; and (c) toexamine rates of 0-methylation with a range of chlorinatedguaiacols and with pentachlorophenol at concentrations of100 jxg liter-'. Rates of transformation and 0-methylationwere calculated from linear portions of the appropriatecurves (i.e., pseudozero-order rates in concentration) andconverted into specific rates by dividing by the cell density.We have given rates of synthesis, although these generallyagreed well with rates of transformation of the substrates.Where several metabolites were formed, rates for formationof the principal one have been given. It should be empha-sized that these rates are subject to a number of inherentuncertainties: analytical errors in determining substrate con-
APPL. ENVIRON. MICROBIOL.

BACTERIAL 0-METHYLATION OF CHLOROGUAIACOLS 281
TABLE 2. Biochemical reactions of strains capable of carryingout 0-methylation of chloroguaiacolsa
No. of positive strains in groupReaction (no. of strains):
A (5) B (2) C (1) D (1) E (2)
Oxidase 5 2 + + 0Catalase 5 2 + - 0P-Galactosidase 2 2 - - 0Lipase (Tween 80) 0 2 + - 0Phosphatase 1 2 - NDb NDHydrolysis of esculin 0 2 - ND NDNitrate to nitrite 0 2 + - 2Acid from (ASS)c:Glucose 5 2 + + 0Sucrose 5 2 + ND NDMaltose 5 2 + + 0Arabinose 5 2 + + 0Xylose ND ND ND + 0Mannitol ND ND ND + 0
Growth with:4-Hydroxybenzoate 5 2 - + 23-Hydroxybenzoate 5 0 + + 2Salicylate 2 0 + - 2Guaiacol 4 0 - - 0Vanillate 5 0 - - 2Syringate 0 0 - - 14-Hydroxyphenylacetate 5 0 - - 2Succinate 5 2 - + 1Decanoate 4 0 - + 0Pelargonate 5 0 - + 1Mandelate 2 0 - - 03,4-Dimethoxybenzoate 3 0 - - 0Methanol 1 0 - - 0Glucose ND ND ND + 0Gluconate ND ND ND + 0
a Individual strains were as follows: group A, strains 1366, 1395, 1487, 1539,and 1571; group B, strains 1623 and 1624; group C, strain 1632; group D, strain1631; group E, strains 1637 and 1678.
b ND, Not determined.c ASS, Ammonia salts sugar (28).
centrations; errors in cell densities obtained by plating densecultures; variability between experiments.The influence of cell density on 0-methylation was exam-
ined with 4,5,6-trichloroguaiacol (100 ,ug liter-'): 1,000-folddilutions of the dense cell suspensions were made to yieldcell densities of ca. 108, 105, and 102 cells ml-'. The numberof viable cells during the experiment was determined bydiluting a sample of the cultures, spreading 0.1-ml portionsonto plates of nutrient agar (Oxoid), and counting thenumber of colonies after incubation for 3 days at 30°C.
(ii) Experiments with growing cells. All of the experimentswith growing cells used substrate concentrations of 100 pLgliter-' since this approximates more closely to those whichmight be encountered in receiving waters and was pragmaticsince rates of 0-methylation could then be expected to becompatible with growth rates. Mineral medium VV 2 wasused, supplemented with 1.0 g of the appropriate growthsubstrates per liter: 4-hydroxy-, 3,4-dihydroxy-, and 3,4-dimethoxybenzoates, vanillate (4-hydroxy-3-methoxybenzo-ate), betaine, succinate, or gluconate. The last was added asa concentrated filter-sterilized solution to the mineral basesterilized by autoclaving; other carbon sources were auto-claved with the mineral base. Inocula were grown with thecorresponding substrate before use in metabolic experi-ments. These were carried out in 250-ml side-arm flaskscontaining 100 ml of medium. The relevant non-growthsubstrate (prepared as a concentrated solution in water) was
added when the medium was inoculated. Flasks were shakenat 23°C on an orbital shaker (ca. 125 rpm). Cell density wasestimated from the light scattering at 630 nm and from thenumber of colonies obtained after plating onto nutrient agaras described above. Samples were periodically removed foranalysis of the concentrations of non-growth substrates andtheir metabolites.
RESULTSDistribution and characterization of strains. Strains capa-
ble of 0-methylating 4,5,6-trichloroguaiacol were isolatedfrom six independent localities.The biochemical reactions of all of the gram-positive
strains (groups A, B, and C) and of the three new gram-neg-ative strains (groups D and E) have been assembled in Table2. All strains were negative for decarboxylation of arginine,ornithine, and lysine and for hydrolysis of gelatin and DNA.Some tests were carried out only for gram-positive orgram-negative strains: all gram-positive strains were positivefor urease and negative for hydrolysis of starch, and allgram-negative strains were negative for production of gasfrom nitrate, formation of fluorescent pigment, and toleranceof cetrimide (0.03%). The strains isolated from methaneenrichments (1395, 1487, 1637, 1678) were nutritionallyversatile. Facultatively methanotrophic bacteria have beendescribed, and, indeed, in the most recent description of thegenus Methylobacterium (17), the obligate requirement forthe ability to utilize methane has been removed.On the basis of the available data, groups A, B, and C
were assigned to the to the genus Rhodococcus (16, 19),instead of Arthrobacter and was done previously; group D,to the genus Pseudomonas; and group E, to the genusAcinetobacter. In no case did the reactions of the strainscorrespond closely to those of well-circumscribed taxa.
Toxicity of the substrates. The toxicities estimated from thediameters of zones of inhibition (disk assay) and from theMICROTOX EC50 (15 min, 15°C) values are given in Table3. For the disk assay, the most convenient amount forcomparison was 150 jLg disk-'. For only one gram-negativestrain (1559) was there any marked reduction in sensitivitytowards the compounds after 7 days of incubation. Forgram-positive strains, on the other hand, there were gener-ally significant changes: there were marked reductions inzone diameters for the more toxic substances (3,4,5,-trichloro-, tetrachloroguaiacol, and pentachlorophenol) andsmaller reductions for the more weakly toxic ones (4,5-di-
TABLE 3. Growth inhibition of groups of seven gram-positiveand seven gram-negative bacteria by chloroguaiacols and
pentachlorophenolDiam (mm) of zones of
Chloroguaiacol/pentachlo- inhibition (range)' MICROTOXrophenol (150 ,ug Gram- Gram- EC5,, (mg5.5-mm disk-1) positive negative liter-)b
organisms organisms
4,5-Dichloroguaiacol 6-11 (55) 6-12 (70) 153,4,5-Trichloro-2,6- 8-15 (0) 6-9 (85) 26dimethoxyphenol
4,5,6-Trichloroguaiacol 8-16 (0) 6-9 (70) 10Tetrachloroguaiacol 17-32 (0) 6-13 (70) 53,4,5-Trichloroguaiacol 36-45 (0) 6-17 (30) 2.5Pentachlorophenol 35-45 (0) 7-27 (0) 1.0
a The range of diameters of zones of inhibition after 3 days of incubation isgiven, and the percentage of resistant strains is given in parentheses.
' Values are for 15-min assays at 15°C.
VOL. 49, 1985

282 ALLARD, REMBERGER, AND NEILSON
TABLE 4. Final yields of 3,4,5-trichloroveratrole formed fromhigh (20 mg liter-') and low (100 ,ug liter-') concentrations of
4,5,6-trichloroguaiacol by groups of bacteriaYield (%)
Bacteria StrainHigh Low
Gram positive 1395 45 1001539 87 1001571 100 1001623 0.5 181624 1.0 181632 76 100
Gram negative 1556 0.3 91557 0.7 301558 0.4 211559 1.8 561631 1.0 471637 1.3 711678 6.0 100
chloro-, 4,5,6-trichloroguaiacol, and 3,4,5-trichloro-2,6-dime-thoxyphenol). By comparison, 4,5-dichloro-, 3,4,5-trichloro-and tetrachloroveratrole, 1,2,3-trichloro-4,5,6-trimethoxy-benzene, and pentachloroanisole were virtually nontoxiceven to the most sensitive test strains.The gram-positive strains were uniformly sensitive to
erythromycin, lincomycin, and clindamycin and the gram-negative strains were resistant.
0-methylation of chloroguaiacols. To make data from thedifferent types of experiments as cohesive as possible, thegram-positive strain 1395 was examined comprehensively:the similar, though independent, strains 1539 and 1571 wereused when necessary. For comparison with gram-negativeorganisms, strain 1678 was used. A wider range of strainswas used to examine the effect of concentration of yields of0-methylated products and the toxicity of the various sub-strates: these results provided a broader foundation for theconclusions drawn from this study.
Effect of substrate concentration, using dense cell suspen-sions. The yields of 3,4,5-trichloroveratrole produced fromhigh (20 mg liter-' and low (100 ,ug liter-') concentrations of4,5,6-trichloroguaiacol by a range ofgram-positive and gram-negative bacteria are given in Table 4.For strain 1395, 3,4,5- and 4,5,6-trichloroguaiacol, tetra-
chloroguaiacol, 3,4,5-trichloro-2,6-dimethoxyphenol, penta-chlorophenol, and 3,4,5-trichloro- and tetrachlorocatecholwere examined at a concentration of 100 ,ug liter-'. Theyields were virtually quantitative (80 to 95%), and the ratesof transformation of the guaiacols mirrored those of forma-tion of the 0-methylated products. The rates of 0-methyla-tion, however, varied: 17 (4,5,6-trichloroguaiacol), 11(3,4,5-trichloro- and tetrachlorocatechol), 10 (pentachlorophenol),5.5 (tetrachloroguaiacol), and 1.6 (3,4,5-trichloroguaiacoland 3,4,5-trichloro-2,6-dimethoxyphenol) ,ug h-1.The products from the chloroguaiacols and pentachloro-
phenol were the 0-methyl ethers; the chlorocatecholsformed, first, chloroguaiacols and then chloroveratroles.The initial product from 3,4,5-trichlorocatechol was shownto be 3,4,5-trichloroguaiacol by GC comparison of the 0-ac-etate, 0-heptafluorobutyrate, and 0-trimethylsilyl ether withauthentic compounds. In the earlier study (29), 0-methyla-tion of 3,4,5-trichloro-2,6-dimethoxyphenol, formed from3,4,5-trichloroguaiacol at higher substrate concentrations,was not observed. The structure of the 1,2,3-trichloro-4,5,6-trimethoxybenzene observed in this study was therefore
confirmed by mass spectrometric comparison with the au-thentic compound (Fig. 1).
For the chlorocatechols, it was particularly convenient tocarry out these experiments at a substrate concentration of100 p.g liter-', since interference due to chemical oxidationwas negligible during the short duration of the experiments.The effect of substrate concentration on the rates of
0-methylation of 3,4,5- and 4,5,6-trichloroguaiacols bystrains 1395 and 1678 is shown in Fig. 2A and B. It should benoted that, for strain 1395, the rates at 10.0 mg liter-' referto the synthesis of 3,4,5-trichloro-2,6-dimethoxyphenol,which is the principal metabolite. In all other cases, 3,4,5-trichloroveratrole was the dominant metabolite. Figure 3A,B, C, and D shows how the rates of transformation of3,4,5-trichloroguaiacol and the nature of the metabolitesformed by strain 1395 varied with the substrate concentra-tion. Although strain 1678 also formed 3,4,5-trichloro-2-dimethoxyphenol from 3,4,5-trichloroguaiacol at a concen-tration of 10.0 mg liter-', this was a minor metabolite.accounting for only ca. 10% of the chloroveratrole.
Transformation by growing cells. When the chloroguai-acols were added to the growth medium at a concentration of100 p.g liter-' at the time of inoculation, there was no delayin the onset of growth, no effect on the growth rate, and nodiminution in the final yield.
(i) Gram-positive strains. The kinetics of synthesis of3,4,5-trichloroveratrole from 4,5,6-trichloroguaiacol bystrain 1395 during growth with betaine is shown in Fig. 4.Similar results were obtained during growth of this strainwith succinate, gluconate, and 4-hydroxybenzoate and dur-ing growth of strains 1539 and 1571 with 4-hydroxybenzoate,3,4-dihydroxybenzoate, and 3,4-dimethoxybenzoate (strain1395 brought about no change during growth with 3,4-dihydroxybenzoate and was unable to use 3,4-dimethoxy-benzoate as a growth substrate). Only the veratrole wasformed from 3,4,5-trichloroguaiacol during growth of strain1395 with succinate, gluconate, betaine, and vanillate andfrom tetrachloroguaiacol during growth of strains 1395,1539, and 1571 with vanillate. On the other hand, duringgrowth with vanillate all three strains brought about acomplex sequence of transformations of 4,5,6-trichloroguai-acol (Fig. 5). The totally unexpected formation of 3,4,5-tri-chloroguaiacol from the 4,5,6-isomer required an unam-biguous confirmation of its structure. The mass spectra of3,4,5- and 4,5,6-trichloroguaiacol derivatives are virtuallyindistinguishable; identification was therefore made by com-paring the GC retention times of three different derivatives,the 0-acetate, 0-heptafluorobutyrate, and 0-trimethylsilylether, with those of authentic compounds. They were iden-tical. A similar procedure was used for identifying 3,4,5-tri-chlorocatechol.
De-O-methylation of 4,5-dichloroguaiacol also took placeduring growth of strain 1395 with vanillate: only 4,5-dichlo-rocatechol was formed and was identified by comparison ofthe 0-acetate, 0-trimethylsilyl ether, and O-heptafluorobu-tyrate with authentic samples. The dichlorocatechol wasdegraded without 0-methylation.
It should be emphasized that de-O-methylation (followedin some cases by 0-methylation) was encountered onlyduring growth with vanillate and only with the less toxicnon-growth substrates.
(ii) Gram-negative strains. Results with the gram-nezativestrains (1637, 1678) were significantly different. 4,5,6-Trichlo-roguaiacol was completely unchanged during growth of thecells with vanillate, and with 4-hydroxy- and 3,4-dihydroxybenzoate, 0-methylation occurred only after cells
APPL. FNVIRON. MICROBIOL.

BACTERIAL 0-METHYLATION OF CHLOROGUAIACOLS 283
1.1 ,2,3-TRICHLORO-4,5,6-TRIMETHOXYBENZENE STANDARD
50 100 150 200 250 300 m/e
50.0
METABOLITE FROM 3,4,5-TRICHLOROGUAIACOLSTRAIN 1395
100.0-FIG. 1. Mass spectra of metabolite from 3,4,5-trichloro-2,6-dimethoxyphenol and of authentic 1,2,3-trichloro-4,5,6-trimethoxybenzene.
had reached the stationary phase and dense cultures hadbeen established.
Effect of cell density. The final yields of 3,4,5-trichloro-veratrole produced by strains 1395 and 1678 from 4,5,6-trichloroguaiacol, the time required to reach a maximalconcentration of the metabolite, and the specific rates (ex-cluding those for the lowest cell densities) are given in Table5. Whereas the cell densities of the two most dense cultureswere virtually constant throughout the experiment, therewas a considerable increase for the least dense cultures.
50
z0
2
UO.
Cl,
z
4
0
IL'
ti4x
10
1.0
0.1 10.01.0
Values given in Table 5 for these cultures therefore overes-timate the true values.
DISCUSSION
Bacteria with the capability of 0-methylating chloroguai-acols were widely distributed in both brackish water andfreshwater sediments and included a diversity of taxonomictypes. The present study was based upon experiments withthe representative members of this collection of bacteria,
50
z
0
za4
LI.
0
LiJ
4r
10
1.0
0.1 1.0 10.0
SUBSTRATE CONCENTRATION (mg.lW) SUBSTRATE CONCENTRATION (mg.I-')
FIG. 2. Dependence of rates of 0-methylation of 4,5,6-trichloroguaiacol (0) and 3,4,5-trichloroguaiacol (0) on substrate concentration.(A) Strain 1395 (3 x 10i cells ml-1); (B) strain 1678 (6 x 109 cells ml-1).
A
VOL. 49, 1985
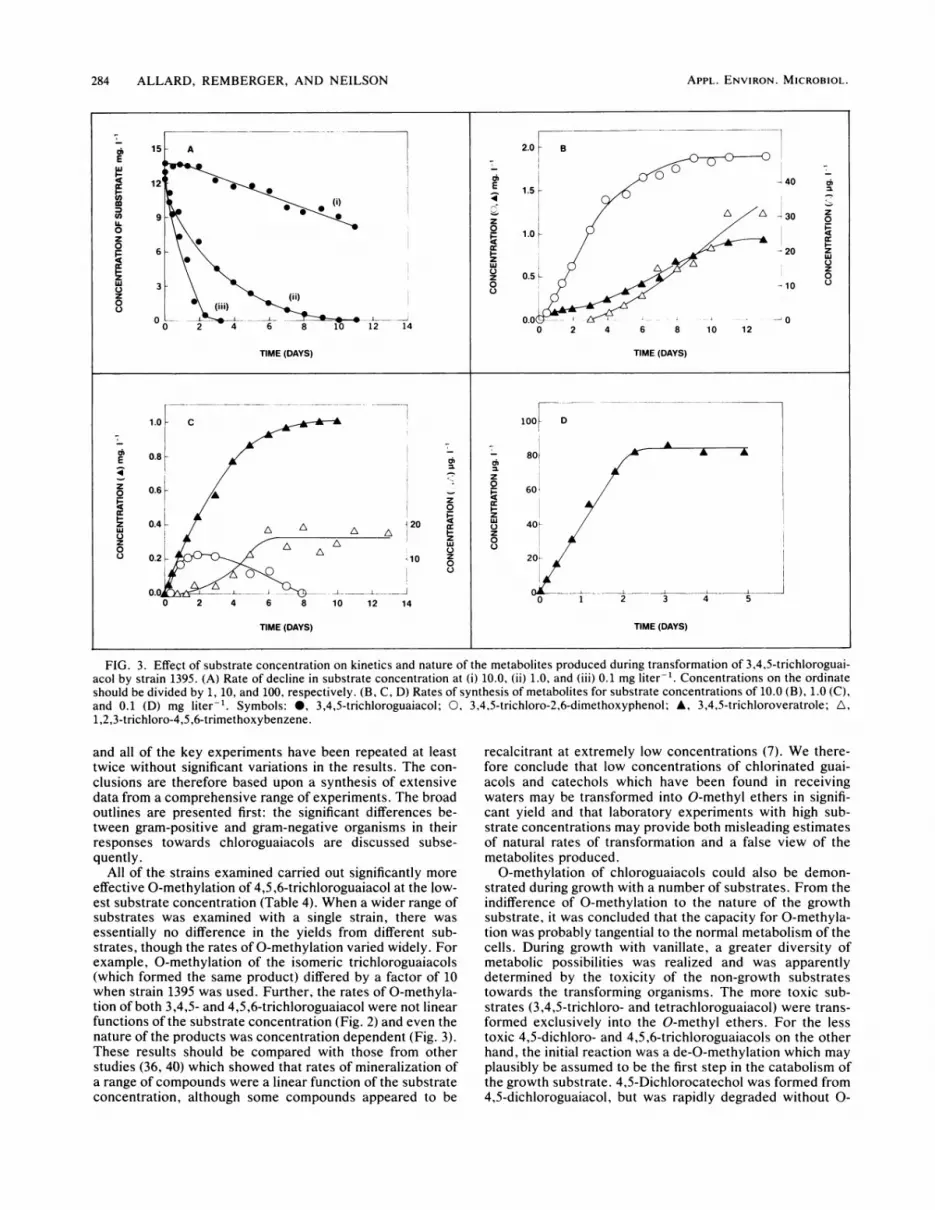
284 ALLARD, REMBERGER, AND NEILSON
FIG. 3. Effect of substrate concentration on kinetics and nature of the metabolites produced during transformation of 3,4,5-trichloroguai-acol by strain 1395. (A) Rate of decline in substrate concentration at (i) 10.0, (ii) 1.0, and (iii) 0.1 mg liter-'. Concentrations on the ordinate
should be divided by 1, 10, and 100, respectively. (B, C, D) Rates of synthesis of metabolites for substrate concentrations of 10.0 (B), 1.0 (C),and 0.1 (D) mg liter-'. Symbols: 0, 3,4,5-trichloroguaiacol; 0, 3,4,5-trichloro-2,6-dimethoxyphenol; A, 3,4,5-trichloroveratrole; A,1,2,3-trichloro-4,5 ,6-trimethoxybenzene.
and all of the key experiments have been repeated at leasttwice without significant variations in the results. The con-clusions are therefore based upon a synthesis of extensivedata from a comprehensive range of experiments. The broadoutlines are presented first: the significant differences be-tween gram-positive and gram-negative organisms in theirresponses towards chloroguaiacols are discussed subse-quently.
All of the strains examined carried out significantly moreeffective 0-methylation of 4,5,6-trichloroguaiacol at the low-est substrate concentration (Table 4). When a wider range ofsubstrates was examined with a single strain, there was
essentially no difference in the yields from different sub-strates, though the rates of 0-methylation varied widely. Forexample, 0-methylation of the isomeric trichloroguaiacols(which formed the same product) differed by a factor of 10when strain 1395 was used. Further, the rates of 0-methyla-tion of both 3,4,5- and 4,5,6-trichloroguaiacol were not linearfunctions of the substrate concentration (Fig. 2) and even thenature of the products was concentration dependent (Fig. 3).These results should be compared with those from otherstudies (36, 40) which showed that rates of mineralization ofa range of compounds were a linear function of the substrateconcentration, although some compounds appeared to be
recalcitrant at extremely low concentrations (7). We there-fore conclude that low concentrations of chlorinated guai-acols and catechols which have been found in receivingwaters may be transformed into 0-methyl ethers in signifi-cant yield and that laboratory experiments with high sub-strate concentrations may provide both misleading estimatesof natural rates of transformation and a false view of themetabolites produced.
0-methylation of chloroguaiacols could also be demon-strated during growth with a number of substrates. From theindifference of 0-methylation to the nature of the growthsubstrate, it was concluded that the capacity for 0-methyla-tion was probably tangential to the normal metabolism of thecells. During growth with vanillate, a greater diversity ofmetabolic possibilities was realized and was apparentlydetermined by the toxicity of the non-growth substratestowards the transforming organisms. The more toxic sub-strates (3,4,5-trichloro- and tetrachloroguaiacol) were trans-formed exclusively into the 0-methyl ethers. For the lesstoxic 4,5-dichloro- and 4,5,6-trichloroguaiacols on the otherhand, the initial reaction was a de-O-methylation which mayplausibly be assumed to be the first step in the catabolism ofthe growth substrate. 4,5-Dichlorocatechol was formed from4,5-dichloroguaiacol, but was rapidly degraded without 0-
U. F~~~~~~~~~~~~~~~~~~~~~~~~~.
0 0 1.0 4~~~~~~~~~~~~~~~~~~~~~2z cr~~~~~~~~~~0~~~~~~~~~~~~~~~~~~~~~~~~~6.)-0 20z ~~~~~~~~~~~~~~~~~~~~~~~~UU
U 80z~~~~~~~~~~~~~~
z 0o 0.6- 0.A
1.0c 100D~~0~
A ~ ~~~~~~~~~~~A A A
z~~~~~~~~~~~~~~~O 0.2- A600~~~~~~~~~~~~~~~1
cc
0. ____ __ - L___A --____
0 2 4 6 8 10 12 14 1 2 35
TIME (DAYS)TIME (DAYS)
APPL. ENVIRON. MICROBIOL.

BACTERIAL 0-METHYLATION OF CHLOROGUAIACOLS 285
:. 80 /z2
60-
O 40 i40
20
1 2 3 4 5
TIME (DAYS)
FIG. 4. Kinetics of synthesis of 3,4,5-trichloroveratrole from4,5,6-trichloroguaiacol by strain 1395 during growth with betaine. Atthe points indicated by arrows, the cell densities were ca. 102, 105,and 108 cells ml-'.
methylation. With 4,5,6-trichloroguaiacol, a more complexsequence of reactions took place: an initial de-O-methylationto 3,4,5-trichlorocatechol followed by selective mono-O-methylation to 3,4,5-trichloroguaiacol and a slow further 0-methylation to yield the final product (3,4,5-trichlorovera-trole). These de-0-methylations are strictly analogous tothose of 5-chlorovanillate and 2-chloroisovanillate duringgrowth of Nocardia corallina with vanillate and isovanillate,respectively (10).
It may seem bizarre that some of the strains were able tocarry out both de-O-methylation and 0-methylation (thoughnot necessarily simultaneously): such a metabolic possibilityhas, however, been demonstrated, e.g., in fungi whichmetabolize 1,4-dichloro-2,5-dimethoxybenzene (44) and inStreptomyces griseus which de-O-ethylates 7-ethoxycou-marin and mono-O-methylates the product to a mixture of6-methoxy-7-hydroxy- and 6-hydroxy-7-methoxycoumarins(37).
In contrast to N. corallina (10), a comparable de-O-meth-ylation did not occur during growth of our strains with
TABLE 5. Effect of cell density on 0-methylation of 4,5,6-trichloroguaiacol (100 ,ug liter') by strains 1395 and 1678"
SpecificTime to transfor-
Final maximal Transforma- mation rateCell density yield concn of tion rate (,g
(%S) metabo- (jig h-') [h (cellslite (h) ml-i)]- l
X io-9
Strain 1395High 100 10 18 99Medium 38 232 0.36 600Low 20 232 b
Strain 1678High 100 25 5.5 1.8Medium 6 148 0.037 2.3Low 4 334
a High and medium cell densities were essentially constant during theexperiment and were (log1o) 8.3 and 5.8 for strain 1395 and 9.5 and 7.3 forstrain 1678. At low cell densities, cryptic growth of both strains occurred sothat cell densities increased from 2.8 to 5.3 (strain 1395) and from 4.3 to 7.7(strain 1678).b-, Not given due to cryptic growth.
3,4-dimethoxybenzoate (only 0-methylation took place). Itmust then be concluded that the pathways for the metabo-lism (or their regulation) of vanillate and 3,4-dimethoxyben-zoate differ in some essential way. The simplest hypothesiswould be that growth with vanillate induces a relativelynonspecific de-O-methylase (6) and that the product ofvanillate catabolism (3,4-dihydroxybenzoate) induces thecapacity for 0-methylation. Consistent with this is the factthat only 0-methylation occurred during growth of strains1539 and 1571 with 3,4-dihydroxybenzoate (strain 1395 wasunique in that 4,5,6-trichloroguaiacol was completely un-changed during growth with this substrate). A plausiblemetabolic pathway could proceed by an initial ring hydroxyla-tion instead of a de-O-methylation: subsequent ring cleavagewould result in formation of a methyl ester and final elimi-nation of the methyl group as methanol (Fig. 6a). Ananalogous pathway has indeed been demonstrated for thecatabolism of 3-0-methylgallate formed from 3,4,5-trime-thoxybenzoate by Pseudomonas putida (Fig. 6b) (12).
Although on the basis of these experiments it is temptingto speculate that transformations of chloroguaiacols willoccur during growth with structurally related substrates
100l
80k-0z
0
FIG. 5. Kinetics of transformation of 4,5,6-trichloroguaiacol (0) and synthesis of 3,4,5-trichlorocatechol (0), 3,4,5-trichloroguaiacol (-),and 3,4,5-trichloroveratrole (A) by strain 1571 during growth with vanillate. At the points indicated by arrows, the cell densities were ca. 105,106, and i07 cells ml-l.
VOL. 49, 1985

286 ALLARD, REMBERGER, AND NEILSON
OMe OMe
,OH(a)
OMe
,- CO2Me
H02C CO2H
(b)
OMe
MeO ON OMe HO
CO2H CO2H
HO2C CO2Me
CO2H
0
FIG. 6. Hypothetical catabolism of vanillate by gram-positivebacteria without de-O-methylation of the aromatic ring (a) andestablished pathway for catabolism of 3,4,5-trimethoxybenzoate byP. putida (12) (b).
(vanillate), evidence both in support of (46) and against (18)this general hypothesis has been presented. All of thetransformations studied here might be termed cometabolic(11, 20). In view of the conflicting views on the value andsignificance of this term (21), however, we prefer to desig-nate these transformations simply as concurrent metabo-lism, using this as a purely descriptive term.These experiments with growing cells showed that 0-
methylation could be detected even in cultures of low celldensity. This was confirmed in experiments with nonproli-ferating cultures: the extent and rate of 0-methylation were
strongly dependent on the cell density, although the specificrates were remarkably similar for a given strain (Table 5).Values for the lowest cell densities were unreliable due tocryptic growth during the experiment. This is probablyunavoidable in the light of increasing evidence (references inreference 25) that many bacteria are able to utilize forgrowth extremely low substrate concentrations. The twolowest cell densities used in this study approximate to valuesfor the total number of heterotrophic bacteria in oligotrophicand eutrophic environments (5, 24). Although these valuesdo not apply to a single type of organism, we have shownthat a variety of both gram-positive and gram-negativebacteria were capable of carrying the 0-methylation of
chloroguaiacols and that they were widely distributed andmust occur in substantial numbers in sediments. Unpub-lished experiments in which sediment samples were incu-bated with 100 ,ug of 4,5,6-trichloroguaiacol liter-' showedthat 3,4,5-trichloroveratrole was produced in yields of 15 to75%. We therefore conclude that the reactions describedhere will occur in the natural environment and are notlaboratory curiosities.
Increasing interest has been shown in the quantitativeaspects of biodegradation (2, 38), and we give below twoexamples of such an application of our data.
Rates of transformation were clearly concentration de-pendent (Fig. 2). If, however, we arbitrarily select a givenconcentration, e.g., 100 ,ug liter-', and divide the rates bythe cell density, we obtain a function which may be used ina number of ways: (i) to compare the effectiveness ofdifferent strains, (ii) to assess the relative recalcitrance of arange of substrates, or (iii) to provide plausible estimates ofrates which might be applied to natural situations and tocompare these with measured values.
It is also instructive to compare our rates of 0-methylationwith those which have been found for a number of othersingle-step transformations. To make the comparison withsecond-order rates (i.e., first order in substrate concentra-tion and cell biomass), we have divided our specific trans-formation rates (i.e., rates divided by the cell density) by thesubstrate concentration. At a concentration of 100 ,ug liter-',our rates ranged from 6 x 10-13 (strain 1395, 4,5,6-trich-loroguaiacol) to 7.5 x 10-15 (strain 1678, 3,4,5-trichloroguai-acol) [h (cells liter-1')]. These values may be comparedwith those for microbial hydrolysis of a number of esters (5x 10-12 to 2 x 10-14) (31) or hydroxylation of substitutedphenols to catechols (7 x 10-12 to 1.5 x 10-14) (32). Inanother study (2) in which rates were zero order in substrateconcentration, specific rates for a range of chlorinatedphenols varied from 10-8 to 2 x 10-13 ,ug [h (cells liter-')]-1.Our values are therefore of the same order as all of these.
In the course of this investigation, striking differences inthe behavior of gram-positive and gram-negative organismstowards chloroguaiacols were revealed. Gram-positive or-ganisms carried out effective 0-methylation of 4,5,6-trich-loroguaiacol at high substrate concentrations (up to 20 mgliter-1) and, concomitant with growth, were able to bringabout 0-methylation (most substrates) or de-O-methylationfollowed by 0-methylation (vanillate). By contrast, gram-negative organisms carried out effective 0-methylation onlyat low substrate concentrations (100 ,ug liter-') and, duringgrowth, brought about 0-methylation only after cells hadreached the stationary phase. This may plausibly be attrib-uted to the high cell densities of such cultures.The toxicity data further underlined these differences. The
disk tests clearly showed that, compared with the chloro-guaiacols, the chloroveratroles were virtually nontoxic. Thedata also permitted a toxicity ranking of the chloroguaiacolsand of pentachlorophenol which agreed substantially withthat obtained from the MICROTOX EC50 values. It istherefore suggested that 0-methylation is a detoxificationmechanism for cells exposed to chloroguaiacols. It wouldtherefore be expected that organisms sensitive to a givensubstrate would most effectively bring about its 0-methyla-tion and vice versa. This was broadly the case. For example,gram-positive organisms were sensitive to the more highlychlorinated substrates and, at high concentrations, effec-tively carried out 0-methylation of 4,5,6-trichloroguaiacol(Table 4). Conversely, the gram-negative organisms wereconspicuously less sensitive to these substances and at high
APPL. ENVIRON. MICROBIOL.

BACTERIAL 0-METHYLATION OF CHLOROGUAIACOLS 287
concentrations yielded only low yields of the 0-methylethers. Indeed, many of the gram-negative organisms weretotally insensitive to these compounds, whereas this wasnever encountered with the gram-positive organisms. Wetherefore propose that 0-methylation of chloroguaiacols andchlorophenols is essentially a mechanism for detoxifyingthese substrates and converting them into virtually nontoxic0-methyl esters. It should, however, be pointed out thatother detoxification reactions for phenols have been demon-strated: acetylation in bacteria (35), and, in fungi, bothformation of sulfate esters (9, 15) and synthesis of glucuronicacid conjugates (9).There are salient analogies between the behavior of the
strains towards chlorinated guaiacols and erythromycin.Gram-positive strains were sensitive to both of these, andgram-negative strains were virtually resistant: resistance ofgram-positive strains involves 0-methylation of chloroguai-acols and N-dimethylation of 23S ribosomal adenine (39).Gram-negative organisms are inherently resistant to eryth-romycin through impermeability to the antibiotic, and ini-tially sensitive gram-positive strains may acquire resistancealso through development of impermeability (43). Theseobservations suggest an additional possible role of permea-bility in determining resistance of bacteria to chloroguai-acols.
This investigation has addressed the issues outstandingfrom the earlier study: extrapolation of the results of thelaboratory experiments to the natural environment necessi-tated the detailed examination of a number of factors. Threeof the more significant have now been examined. We there-fore feel justified in postulating that 0-methylation of chlo-roguaiacols will occur under natural conditions: (i) therelevant organisms were widely distributed, (ii) all of themwere able to transform low substrate concentrations in highyields, (iii) they were capable of carrying out 0-methylationduring growth with a wide variety of substrates which mayoccur in natural environments, and (iv) even in the absenceof growth substrates, they brought about 0-methylation atlow cell densities.
Apart from our own observations, the wider significanceof 0-methylation of halogenated phenols is supported bysuch diverse observations as the detection of pentachloroan-isole in oysters (26) and the muscle tissue of fish (34) and thatof di-O-methyl-tetrabromobisphenol A in sediment (41, 42)and mussel (42) samples.
It is suggested that, in general, it may be prudent inenvironmental hazard assessments to examine the possibil-ity that a given substance may be transformed in theenvironment into compounds at least as toxic as theirprecursors.
ACKNOWLEDGMENTSThis investigation has received financial support from the Re-
search Committee of the National Swedish Environment ProtectionBoard and from the Swedish Paper and Pulp Association.
LITERATURE CITED1. Ahlborg, U. G., and T. M. Thunberg. 1980. Chlorinated ph-
enols: occurrence, toxicity, metabolism and environmental im-pact. Crit. Rev. Toxicol. 7:1-35.
2. Banerjee, S., P. H. Howard, A. M. Rosenberg, A. E. Dom-browski, H. Sikka, and D. L. Tullis. 1984. Development of ageneral kinetic model for biodegradation and its application tochlorophenols and related compounds. Environ. Sci. Technol.18:416-422.
3. Barry, A. L., and C. Thornsberry. 1980. Susceptibility testing:diffusion test procedures, p. 463-474. In E. H. Lennette, A.
Balows, W. J. Hausler, and J. P. Truant (ed.), Manual of clinicalmicrobiology, 3rd ed. American Society for Microbiology,Washington, D.C.
4. Bartels, I., H.-J. Knackmuss, and W. Reinecke. 1984. Suicideinactivation of catechol 2,3-dioxygenase from Pseudomonasputida mt-2 by 3-halocatechols. Appl. Environ. Microbiol.47:500-505.
5. Baughman, G. L., and R. R. Lassiter. 1978. Prediction ofenvironmental pollutant concentration, p. 35-54. In J. Cairns,K. L. Dickson, and A. W. Maki (ed.), Estimating the hazard ofchemical substances to aquatic life. ASTM STP 657. AmericanSociety for Testing and Materials, Philadelphia.
6. Bernhardt, F. H., N. Erdin, H. Staudinger, and V. Ullrich. 1973.Interactions of substrates with a purified 4-methoxybenzoatemonooxygenase system (O-demethylating) from Pseudomonasputida. Eur. J. Biochem. 35:126-134.
7. Boethling, R. S., and M. Alexander. 1979. Effect of concentra-tion of organic chemicals on their biodegradation by naturalmicrobial communities. Appl. Environ. Microbiol. 37:1211-1216.
8. Bulich, A. A., M. W. Greene, and D. L. Isenberg. 1981.Reliability of the bacterial luminescence assay for determinationof the toxicity of pure compounds and complex effluents, p.338-347. In D. R. Branson and K. L. Dickson (ed.), Aquatictoxicology and hazard assessment: fourth conference. ASTMSTP 737. American Society for Testing and Materials, Philadel-phia.
9. Cerniglia, C. E., J. P. Freeman, and R. K. Mitchum. 1982.Glucuronide and sulfate conjugation in the fungal metabolismof aromatic hydrocarbons. Appl. Environ. Microbiol. 43:1070-1075.
10. Crawford, R. L., E. McCoy, J. M. Harkin, T. K. Kirk, and J. R.Obst. 1973. Degradation of methoxylated benzoic acid by aNocardia from a lignin-rich environment: significance to lignindegradation and effect of chloro substituents. Appl. Microbiol.26:176-184.
11. Dalton, H., and D. I. Stirling. 1982. Co-metabolism. Philos.Trans. R. Soc. London Ser. B 297:481-496.
12. Donnelly, M. I., and S. Dagley. 1980. Production of methanolfrom aromatic acids by Pseudomonas putida. J. Bacteriol.142:916-924.
13. Franklin, T. J., and G. A. Snow. 1981. Biochemistry of antimi-crobial action, 3rd ed. Chapman and Hall, London.
14. Furukawa, K. 1982. Microbial degradation of polychlorinatedbiphenyls (PCBs), p. 33-57. In A. M. Chakrabarty (ed.), Bio-degradation and detoxification of environmental pollutants.CRC Press, Boca Raton, Fla.
15. Golbeck, J. H., S. A. Albaugh, and R. Radmer. 1983. Metabo-lism of biphenyl by Aspergillus toxicarius: induction ofhydroxylating activity and accumulation of water-soluble me-tabolites. J. Bacteriol. 156:49-57.
16. Goodfellow, M., and G. Alderson. 1977. The actinomycete-ge-nus Rhodococcus: a home for the 'rhodochrous' complex. J.Gen. Microbiol. 100:99-122.
17. Green, P. N., and I. J. Bousefield. 1983. Emendation of Meth-ylobacterium Patt, Cole, and Hanson 1976; Methylobacteriumrhodinum (Heumann 1692) comb. nov. corrig.; Methylobacter-ium radiotolerans (Ito and lizuka 1971) comb. nov. corrig.; andMethylobacterium mesophilicum (Austin and Goodfellow 1979)comb. nov. Int. J. Syst. Bacteriol. 33:875-877.
18. Haller, H. D., and R. K. Finn. 1978. Kinetics of biodegradationof p-nitrobenzoate and inhibition of benzoate in a pseu-domonad. Appl. Environ. Microbiol. 35:890-896.
19. Helmke, E., and H. Weyland. 1984. Rhodococcus marinonas-cens sp. nov., an actinomycete from the sea. Int. J. Syst.Bacteriol. 34:127-138.
20. Horvath, R. S. 1972. Microbial co-metabolism and the degrada-tion of organic compounds in nature. Bacteriol. Rev. 36:146-155.
21. Hulbert, M. H., and S. Krawiec. 1977. Cometabolism: a critique.J. Theor. Biol. 69:287-291.
22. Kovacs, T. G., R. H. Voss, and A. Wong. 1984. Chlorinatedphenolics of bleached kraft mill origin. An olefactory evalua-tion. Water Res. 18:911-916.
VOL. 49, 1985

288 ALLARD, REMBERGER, AND NEILSON
23. Kringstad, K., and K. Lindstrom. 1984. Spent liquors from pulpbleaching. Environ. Sci. Technol. 18:236A-248A.
24. Mallory, L. M., B. Austin, and R. R. Colwell. 1977. Numericaltaxonomy and ecology of oligotrophic bacteria isolated from theestuarine environment. Can. J. Microbiol. 23:733-750.
25. Martin, P., and R. A. MacLeod. 1984. Observations on thedistinction between oligotrophic and eutrophic marine bacteria.Appl. Environ. Microbiol. 47:1017-1022.
26. Miyazaki, T., S. Kaneko, S. Horii, and T. Yamagishi. 1981.Identification of polyhalogenated anisoles and phenols in oys-ters collected from Tokyo Bay. Bull. Environ. Contam. Toxi-col. 26:577-584.
27. Neilson, A. H. 1980. Isolation and characterization of bacteriafrom the Baltic Sea. J. Appl. Bacteriol. 49:199-213.
28. Neilson, A. H. 1980. Isolation and characterization of bacteriafrom the Swedish west coast. J. Appl. Bacteriol. 49:215-223.
29. Neilson, A. H., A.-S. Allard, P.-A. Hynning, M. Remberger, andL. Landner. 1983. Bacterial methylation of chlorinated phenolsand guaiacols: formation of veratroles from guaiacols andhigh-molecular-weight chlorinated lignin. Appl. Environ. Mi-crobiol. 45:774-783.
30. Neilson, A. H., A.-S. Allard, S. Reiland, M. Remberger, A.Tarnholm, T. Viktor, and L. Landner. 1984. Tri- and tetrachlo-roveratrole, metabolites produced by bacterial O-methylation oftri- and tetrachloroguaiacol: an assessment of their bioconcen-tration potential and their effects on fish reproduction. Can. J.Fish. Aquat. Sci. 41:1502-1512.
31. Paris, D. F., W. C. Steen, G. L. Baughmann, and J. T. Barnett.1981. Second-order model to predict microbial degradation oforganic compounds in natural waters. Appl. Environ. Mi-crobiol. 41:603-609.
32. Paris, D. F., N. F. Wolfe, and W. C. Steen. 1982. Structure-ac-tivity relationships in microbial transformation of phenols.Appl. Environ. Microbiol. 44:153-158.
33. Perdue, E. M., and N. L. Wolfe. 1982. Modification of pollutanthydrolysis kinetics in the presence of humic substances. Envi-ron. Sci. Technol. 16:847-852.
34. Renberg, L., E. Marell, G. Sundstrom, and M. Adolfsson-Erici.1983. Levels of chlorophenols in natural waters and fish after anaccidental discharge of a wood-impregnating solution. Ambio
12:121-123.35. Rott, B., S. Nitz, and F. Korte. 1979. Microbial decomposition of
pentachlorophenolate. J. Agric. Food Chem. 27:306-310.36. Rubin, H. E., R. V. Subba-Rao, and M. Alexander. 1982. Rate of
mineralization of trace concentrations of aromatic compoundsin lake water and sewage samples. Appi. Environ. Microbiol.43:1133-1138.
37. Sariaslani, F. S., and J. P. Rosazza. 1983. Novel biotransforma-tions of 7-ethyoxycoumarin by Streptomyces griseus. Appl.Environ. Microbiol. 46:468-474.
38. Simkins, S., and M. Alexander. 1984. Models for mineralizationkinetics with the variables of substrate concentration and pop-ulation density. Appl. Environ. Microbiol. 47:1299-1306.
39. Skinner, R. H., and E. Cundcliffe. 1982. Dimethylation ofadenine and the resistance of Streptomyces erythraeus to eryth-romycin. J. Gen. Microbiol. 128:2411-2416.
40. Subba-Rao, R. V., H. E. Rubin, and M. Alexander. 1982.Kinetics and extent of mineralization of organic chemicals attrace levels in freshwater and sewage. Appl. Environ. Mi-crobiol. 43:1139-1150.
41. Watanabe, I., T. Kashimoto, and R. Tatsukawa. 1983. The flameretardant tetrabromobisphenol-A and its metabolites found inriver and marine sediments in Japan. Chemosphere12:1533-1539.
42. Watanabe, I., T. Kashimoto, and R. Tatsukawa. 1983. Identifi-cation of the flame retardant tetrabromobisphenol-A in the riversediment and the mussel collected in Osaka. Bull. Environ.Contam. Toxicol. 31:48-52.
43. Weisblum, B., C. Siddhikol, C. J. Lai, and V. Demohn. 1971.Erythromycin-inducible resistance in Staphylococcus aureus:requirements for induction. J. Bacteriol. 106:835-847.
44. Wiese, M. V., and J. M. Vargas. 1973. Degradation and synthe-sis of chloroneb and 2,5-dichloro-4-methoxyphenol by soil mi-croorganisms. Pestic. Biochem. Physiol. 3:214-222.
45. Wright, S. J. L. 1975. A simple agar plate method, usingmicroalgae for herbicide bio-assay or detection. Bull. Environ.Contam. Toxicol. 14:65-70.
46. You, I.-S., and R. Bartha. 1982. Stimulation of 3,4-dichloroan-iline mineralization by aniline. Appl. Environ. Microbiol.44:678-681.
APPL. ENVIRON. MICROBIOL.





























