Embed Size (px)
Citation preview

Mycological Society of America
Balansia pilulaeformis, an Epiphytic SpeciesAuthor(s): Keith Clay and Irene C. FrentzSource: Mycologia, Vol. 85, No. 4 (Jul. - Aug., 1993), pp. 527-534Published by: Mycological Society of AmericaStable URL: http://www.jstor.org/stable/3760498 .
Accessed: 16/09/2013 14:36
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Mycological Society of America is collaborating with JSTOR to digitize, preserve and extend access toMycologia.
http://www.jstor.org
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

,, , 85(4)
MyCOlOgia July-August 1993
Official Publication of the Mycological Society of America
BALANSIA PILULAEFORMIS, AN EPIPHYTIC SPECIES
Keith Clay and Irene C. Frentz
Department of Biology, Indiana University, Bloomington, Indiana 47405
ABSTRACT
Living specimens of the grass Chasmanthium laxum infected with the fungus Balansia pilulaeformis were examined for growth habit of the fungus. The fungus was found to grow epiphytically on meristems and in the axils of young leaves in a manner essentially identical to Atkinsonella species and B. cyperi. Living specimens of Cenchrus echinatus infected by B. obtecta and dried specimens of rice (Oryza sativa) infected by B. pallida were also examined and the latter species exhibited an epiphytic growth form as well. As now understood, the majority of species and genera in the tribe Balansieae (Clavicip- itaceae) critically examined are epiphytes, belying the notion that most species are endophytic. These additional findings of epiphytism provide further motivation for examining other taxa and have im? plications for generic classification in the tribe.
Key Words: Balansia, Chasmanthium, endophyte, epiphyte
The tribe Balansieae, Clavicipitaceae, consists of several genera of fungi that systematically in? fect grasses and, to a lesser extent, sedges. Epi? chloe typhina (Fr.) Tul. and a large number of related anamorphs exist as endophytes within the leaves and stems of many cool-season grasses and have been associated with livestock toxicity, insect resistance, and enhanced host vigor (Clay, 1988). Evidence of their endophytic habit comes from direct microscopic examination of hyphae within host tissues, including seeds, and from culture tests where the fungus grows out from surface sterilized host tissues. Although conidio? mata and ascomata or fruiting bodies develop externally with the egress of fungal mycelium, these fungi grow vegetatively within their hosts as endophytes (White et al., 1991a). It has gen- erally been assumed that all members of this tribe (including species ofAtkinsonella, Balansia, and Balansiopsis) were endophytic (Clay, 1986a; Sie? gel et al., 1987) and, indeed, the whole tribe is often referred to by the generic term grass en?
dophyte. Diehl's (1950) monograph is definitive on the
subject of endophytism. For example, regarding Balansia he states (p. 3): "This evidence of sys- temic infection is supported by the presence of characteristic hyphae in nodes and to some ex-
tent internodes and even in leaves. . . . These hy? phae are most readily detected in meristematic tissues and in intercellular spaces." Later, dis-
cussing Atkinsonella hypoxylon (Peck) Diehl states (incorrectly) (p. 53), "The mycelium of this species is intercellular and perennial in the tis? sues of the host."
The species Myriogenospora atramentosa (Berk. & Curt.) Diehl, added to the tribe by Lut? trell and Bacon (1977) based on the development of perithecia and the anamorph, differs from oth? er members of the tribe in being entirely epi- phytic on the outer epidermis of its hosts (Smith et al., 1985). More recently, Leuchtmann and Clay (1988) demonstrated that A. hypoxylon and B. cyperi Edg. are also epiphytic. The establish- ment of the new species A. texensis (Diehl) Leuchtmann & Clay, which is epiphytic on Stipa leucotricha Trin. & Rupr., raised to four the number of epiphytic species, encompassing three genera in the Balansieae (Leuchtmann and Clay, 1988, 1989). These findings indicated that the epiphytic habit is not uncommon in the tribe.
In this paper we describe the epiphytic growth habit of another species of Balansia, B. pilulae- formis (Berk. & Curt.) Diehl. This taxon was placed in the genus Balansiopsis by Diehl (1950) based on the absence of conidia, but it was later
Mycologia, 85(4), 1993, pp. 527-534. ? 1993, by The New York Botanical Garden, Bronx, NY 10458-5126 Issued 3 September 1993
527
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

528 Mycologia
shown by Rykard et al. (1984) to produce co? nidia. Two additional Balansia species were ex? amined in less detail and one was found to be
epiphytic as well. This raises further questions about the growth habit of other members ofthe tribe and the uncritical assumption of endophy- tism. We suggest that growth habit is a heretofore
underappreciated taxonomic character; the sig- nificant differences between epiphytic and en?
dophytic infections of hosts may necessitate ge- neric reclassification.
MATERIALS AND METHODS
Plant material. ?The grass Chasmanthium laxum (L.) Yates infected by Balansia pilulaeformis was collected in the summer of 1991 and 1992 from the Hobcaw Barony, owned by the Belle W. Baruch Foundation in Georgetown Co. near Georgetown, South Carolina. In? fected plants were evident as every inflorescence was aborted and bore the stroma of B. pilulaeformis. In? fected plants and uninfected controls were dug up and returned to the greenhouses at Indiana University where several specimens are currently growing. In addition, living plants of Cenchrus echinatus L. infected by B. obtecta Diehl were collected in Baseball City, Lakeland Co., Florida along the perimeter of an orange grove and dried specimens of Oryza sativa L. infected by B. pallida (Wint.) Diehl from Sierra Leone were kindly provided by Dr. S. N. Fomba at the West Africa Rice Development Association's Regional Mangrove Swamp Rice Research Station in Freetown, Sierra Leone (Fom? ba, 1984). Infected specimens of all three host species have been deposited in the U.S. National Fungal Col? lections, Beltsville, Maryland. Culture studies.?The growth habit of B. pilulaeformis infecting C laxum was investigated by two different approaches. First, attempts were made to culture the fungus from several tissue types including nodes and internodes of the aborted inflorescences, leaves, culm bases, stromata, and ascospores. All tissues except for the culm bases and ascospores were surface sterilized by immersing them for 1 min in 70% ethanol, then 3 min in 50% bleach (2.63% sodium hypochlorite), and finally 0.5 min in 70% ethanol. The cut surfaces ex? posed to the sterilants were then trimmed with a scalpel and the remaining tissue was cut into small pieces and placed on plates containing corn meal-malt (CMM) agar with oxytetracycline (17 g cornmeal agar, 20 g malt extract, 2 g yeast extract, 50 mg oxytetracycline hydrochloride per L distilled water). The plates were stored at room temperature and checked periodically for outgrowth of B. pilulaeformis. If the fungus grew endophytically, then surface sterilization should not kill it, and it should grow out of all surface-sterilized plant samples. However, if the fungus grew epiphyti- cally, only mycelium protected from the sterilant (e.g., inner leaf whorls, interior ofthe stroma) should survive the sterilization process.
Ascospores were isolated by placing an ascostroma in 250 /ul distilled, sterile water for about 1 h. A few lA of this water were then transferred to a glass slide
and examined under a microscope for the long, needle- shaped ascospores. A further 50 y\ of the ascospore- containing water was spread on a CMM plate contain? ing a higher concentration of agar (6% total). Using a dissecting microscope at 40 x, single ascospores were transferred to separate CMM plates and observed for growth. Ascospores were plated in order to ensure that our techniques were suitable for the culture of B. pi- lulaeformis.
Histological studies.?Tissues of infected C. laxum ex? amined for the presence of the fungus included leaf sheaths, pith, and tiller bases. For leaf sheaths, the epidermis from the inner side of the leaf was carefully peeled off, placed on a slide torn side up, and stained with lactophenol cotton blue. For pith, a node and surrounding internode of a stroma-bearing culm were eut in half longitudinally and the pith was scraped from around the node, and stained as above. Initially, tiller bases were sectioned longitudinally by hand but the toughness of the material made it difficult to obtain thin sections. As a result, parafnn sections were made. Tissues were fixed in FAA (formaldehyde-acetic acid- alcohol), dehydrated in a tertiary butyl alcohol series, and embedded in Paraplast (melting point 52-54 C). Eight-Aim thick sections were stained with tannic acid- ferric chloride, safranin and fast green, according to the method of Foster as modified by Giffbrd (Schnei- der, 1981), while 6-^m thick sections were stained with thionin-orange G stain (Stoughton, 1930). Five-jum sections were stained with a modification of the pre- vious stain as described by Margolena (1932).
For C. echinatus infected by B. obtecta, leaf sheaths and stem pith form freshly collected plants were ex? amined as described above. Dried specimens of rice infected with B. pallida were soaked for several hours in warm soapy water to soften the tissues. Leaf sheaths, stem pith, and nodes and internodes subtending the aborted panicle were examined as described above. The specimens did not include the base of tillers, so they could not be examined.
RESULTS
Culture studies.?The results of culture studies are summarized in Table I. The growth of most
ascospores clearly demonstrates that the fungus is culturable under our experimental conditions.
Therefore, growth could be expected from plant tissues as well. Most ascospores germinated with? in 2 wk. On CMM agar, colonies from ascospores were greenish-yellow with a dark brown reverse. The mycelium was velvety to densely cottony in
texture, and the center was raised. Conidia were not observed in either 2-wk-old or 4-month-old colonies. All of the attempted isolations from stromata were contaminated, probably reflecting the age of the stromata and the fact that infected
plants were collected in the field. The results from the culture studies of the internodes suggested that the fungus was not present in these tissues.
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

Clay and Frentz: Epiphytic Balansia Species 529
Although all ofthe pieces from the nodes became contaminated, the majority of pieces from in- ternodes remained sterile. The tightly appressed leaf sheath surrounding the nodes prevents com? plete sterilization of the protected interior. In contrast, the fungus grew from the culm bases and inner leaves near the base, indicating that the fungus was present in or on the meristematic region and in or on leaf bases near the culm base, where the fungus is protected from the sterilant. Fungal cultures growing out from plant pieces matched in all respects cultures derived from single ascospores. The lack of fungal growth from internodes suggests that B. pilulaeformis is not an endophyte, but fungal growth from the culm bases does not in and of itself prove that the fungus in an epiphyte.
Histological studies.?The stromata of B. pilu? laeformis, which are produced at the apex of culms, are initially light grey and then turn darker as perithecia develop (Fig. 1). They bear a strong resemblance to the stromata of Atkinsonella hy- poxylon infecting Danthonia and similarly sur- round plant tissue. Erumpent ascostromata are distinctly papillate (Fig. 2). Microscopic exam? ination of tissues from infected plants of C. lax? um revealed no fungal hyphae in pith or leaf sheaths. However, longitudinal sections of tiller bases revealed that there were mats of fungi in leaf axils and around leaf primordia and inter- calary meristems (Fig. 3), more or less identical to the appearance and location of Atkinsonella hypoxylon infecting Danthonia spicata (Leucht- mann and Clay, 1988, Figs. 1, 3). Hyphae of B. pilulaeformis are hyaline and approximately 2 Mm in diameter. They are arranged in mats of parallel, densely packed hyphae (Figs. 4, 5). The hyphae run radially in several layers in the leaf axils (Fig. 4) but longitudinally on the surfaces above (Fig. 5). Single hyphae, which may have been sheared offa mat during sectioning, can be seen as well (Fig. 6).
Although dried specimens of rice infected with B. pallida did not provide high quality material for detailed histological studies, they provided clear evidence of epiphytism in this species. The inflorescences of rice were aborted by the fungus. Unlike B. pilulaeformis and B. obtecta, which completely mummify the entire inflorescence, individual grains of rice are replaced by individ? ual stromata enclosed within single lemmas and paleas (Fig. 7). The tips of stromata could be
Table I Results of cultural studies of Chasmanthium laxum infected with balansia pilulaeformis
a Not surface sterilized.
observed just barely emerging from the floret. No hyphae were found in leaf sheaths or pith. In contrast, mats of hyphae similar to those of B. pilulaeformis were observed on the bases of leaves and inside leaf sheaths subtending the aborted inflorescences (Fig. 9). Mats consisted of thin layers of densely packed, parallel hyphae.
The fruiting bodies of B. obtecta are elongate black cylinders 3-10 cm in length and 3-8 mm wide (Fig. 8). They are borne at the apex of culms where inflorescences are normally produced. On microscopic examination, hyphae of B. obtecta were found in the pith as well as the leaf sheath ofCenchrus echinatus (Figs. 10, 11). The hyphae in the leaf sheath are intercellular and very straight (Fig. 11), resembling the hyphae of B. epichloe (Weese) Diehl infecting smutgrass (Sporobolus poiretii) (Roem. & Schult.) Hitchc. (see Fig. 15, Leuchtmann and Clay, 1988). In the pith, the hyphae are more convoluted (Fig. 10) as in B. henningsiana (Moell.) Diehl (Fig. 16, Leucht? mann and Clay, 1988).
The number of epiphytic species in the tribe Balansieae now numbers six (Table II). To our knowledge, only four members of the tribe have been definitively shown to be endophytes, in? cluding B. obtecta here. This does not include the large number of Acremonium derivatives of E. typhina. Most members of the tribe still re- main unexamined as to their endophytic or epi? phytic growth habit. The hosts of epiphytic Bal? ansieae are taxonomically diverse and include members from at least 5 of the 6 subfamilies (Pooideae, Panicoideae, Oryzoideae, Bambuso- ideae, and Arundinoideae) of grasses, and the sedge family Cyperaceae. The endophytic genera of the Balansieae are also widespread in their host relations but are not known to occur regu- larly outside of the grass family.
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions
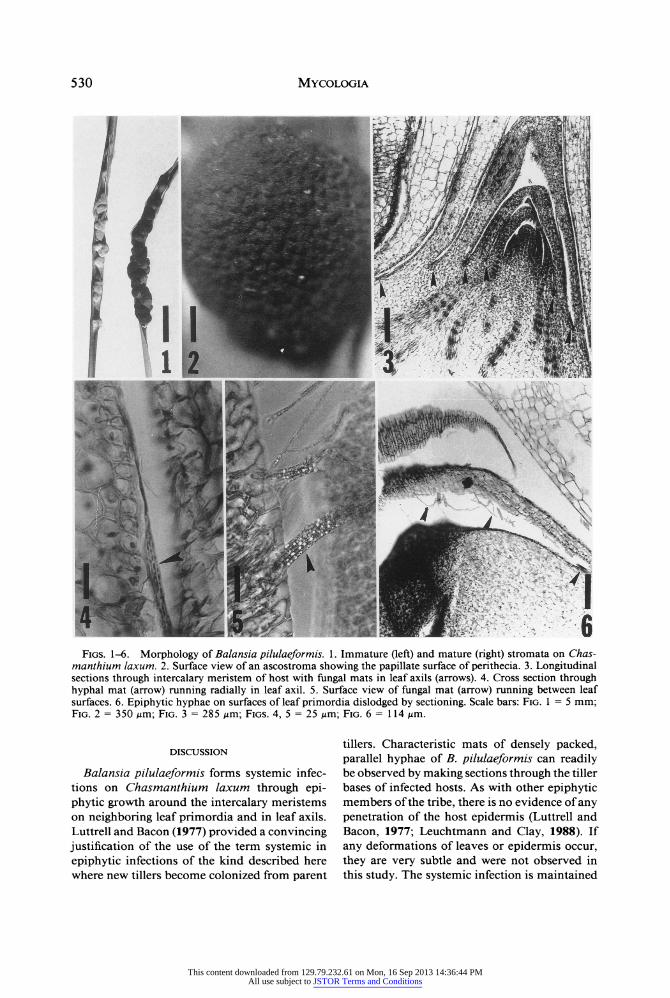
530 Mycologia
Figs. 1-6. Morphology of Balansia pilulaeformis. 1. Immature (left) and mature (right) stromata on Chas? manthium laxum. 2. Surface view of an ascostroma showing the papillate surface of perithecia. 3. Longitudinal sections through intercalary meristem of host with fungal mats in leaf axils (arrows). 4. Cross section through hyphal mat (arrow) running radially in leaf axil. 5. Surface view of fungal mat (arrow) running between leaf surfaces. 6. Epiphytic hyphae on surfaces of leaf primordia dislodged by sectioning. Scale bars: Fig. 1 = 5 mm; Fig. 2 = 350 ^m; Fig. 3 = 285 /im; Figs. 4, 5 = 25 nm; Fig. 6=114 /xm.
DISCUSSION
Balansia pilulaeformis forms systemic infec- tions on Chasmanthium laxum through epi? phytic growth around the intercalary meristems on neighboring leaf primordia and in leaf axils. Luttrell and Bacon (1977) provided a convincing justification of the use of the term systemic in epiphytic infections of the kind described here where new tillers become colonized from parent
tillers. Characteristic mats of densely packed, parallel hyphae of B. pilulaeformis can readily be observed by making sections through the tiller bases of infected hosts. As with other epiphytic members ofthe tribe, there is no evidence of any penetration of the host epidermis (Luttrell and Bacon, 1977; Leuchtmann and Clay, 1988). If any deformations of leaves or epidermis occur, they are very subtle and were not observed in this study. The systemic infection is maintained
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

Clay and Frentz: Epiphytic Balansia Species 531
Figs. 7-11. Morphology of Balansia pallida and B. obtecta. 7. Inflorescence of rice infected with B. pallida illustrating tips of stromata emerging through the lemmas (arrows). 8. Stromata of B. obtecta (arrows) formed around aborted inflorescences of host plant. 9. Hyphal mats of B. pallida on leaf surface (arrow). 10. Intercellular hypha (arrow) of B. obtecta in pith of host. 11. Linear hyphae (arrows) of B. obtecta in leaf sheath of host. Scale bars: Fig. 7=10 mm; Fig. 8 = 24 mm; Fig. 9 = 55 nm; Figs. 10, 11 = 25 Mm.
Table II List of epiphytic and endophytic species in the tribe Balansieae and their taxonomic distribution
among tribes in the grass family
Species Host range Reference
Epiphytes: Atkinsonella hypoxylon Atkinsonella texensis Balansia cyperi Balansia pallida Balansia pilulaeformis
Myriogenospora atramentosa
Endophytes: Balansia epichloe
Balansia henningsiana
Balansia obtecta Epichloe typhina
Danthonieae Stipeae Cyperus, Cyperaceae Paniceae, Oryzeae Bambuseae, Centosteceae,
Paniceae Andropogoneae, Paniceae
Andropogoneae, Aveneae, Chlorideae, Eragrosteae, Paniceae
Andropogoneae, Chlorideae, Paniceae
Chlorideae, Paniceae Aveneae, Brachyelytreae, Meli-
ceae, Poeae, Triticeae
Leuchtmann and Clay (1988) Leuchtmann and Clay (1988,1989) Leuchtmann and Clay (1988) (This paper) (This paper)
Luttrell and Bacon (1977)
Rykard et al. (1985)
Leuchtmann and Clay (1988)
(This paper) Sampson (1933)
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

532 Mycologia
as new meristems and tiller primordia are col? onized from prexisting infected tillers. The fun?
gus sterilizes its host by the development of stro? mata around immature inflorescences. All plants observed bore either only completely normal
flowering culms or only aborted culms with B.
pilulaeformis stromata. The mechanism of con-
tagious spread ofthe fungus is not known.
Presumably the fungus infects other hosts epi- phytically although C. laxum was the only host examined. Hosts of B. pilulaeformis reported by Diehl (1950) include Uniola laxa (= C. laxum), Paspalum ciliatifolium Michx., P. pubescens MuhL, and an unidentified bamboo. Limited ob- servations of preserved material of B. pallida infecting rice indicate a very similar growth hab? it. Diehl (1950), Ou (1972), and Fomba (1984) report that the species infects several oryzoid grasses in addition to the panicoid grass Impera- ta cylindrica (L.) Beauv. Ou (1972) cites Deigh- ton's (1956) (original not seen) reports of B. pal? lida infecting 13 wild grass species in Sierra Leone.
This research has revealed additional epi? phytic members of a group generally considered to be endophytic. Based on current knowledge ofthe tribe, the majority of species are epiphytic and three of four genera (or five depending on the disposition ofthe genus Balansiopsis) consist
entirely or partly of epiphytic species. Thus, ex-
cepting E. typhina and its anamorphic deriva- tives, endophytism in the tribe should not be assumed without microscopic examination, and
may prove to be the exception rather than the rule. Our results point to the need for critical examination of other species. The term endo?
phyte should be used more rigorously and not as a blanket term.
Epiphytism as represented by B. pilulaeformis, and Atkinsonella and other Balansia species, has
implications for the physiology of host/parasite interactions. Epiphytic species presumably ob? tain their nutrition from substances found on the surfaces of young leaves and leaf primordia. In contrast, endophytic species occur intercellularly in leaves and flowering culms where they have more direct access to the products of photosyn- thesis. The enzymatic capabilities of these fungi may differ. Stovall (1988) compared B. cyperi, an epiphyte, and B. obtecta, an endophyte, and found that, while both species generally exhib- ited similar enzymatic capabilities, the endo? phytic species had pectate lyase activity, whereas the epiphyte did not. This difference is consistent
with the growth of B. obtecta in intercellular
spaces where it could utilize pectin in the middle lamella as a nutritional source. White et al.
(1991b) recently demonstrated that epiphytic A.
hypoxylon had the greater ability to utilize oil and pararfm compared to endophytic Balansia and Acremonium taxa. An epiphyte might be
expected to be able to exploit the cuticle as an
energy source whereas an endophyte would never
directly contact the cuticle except when fruiting. The location of epiphytic fungi around the mer- istems also provides an ideal location for influ-
encing growth and developmental processes of their hosts (Leuchtmann and Clay, 1988). The
endophyte B. epichloe has been shown to pro? duce plant growth regulators in culture (Porter et al., 1985), and several species infected with the epiphytes B. cyperi and M. atramentosa are known to produce developmental abnormalities (viviparous plantlets) from aborted inflores? cences (Clay, 1986b).
As pointed out by Rykard et al. (1984), stro? mata of Balansia pilulaeformis bear a striking re- semblance to those of A. hypoxylon infecting spe? cies of Danthonia. Here we have shown the
similarity extends to their epiphytic growth habit as well. Further, recent systematic treatments have removed several species formerly placed in the genus Uniola in the tribe Unioleae, subfamily Chloridoideae and placed them in the genus Chasmanthium in the tribe Centosteceae, sub?
family Arundinoideae, to which Danthonia and the tribe Danthoneae also belong (Gould and
Shaw, 1983). Thus, the common hosts of A hy? poxylon and B. pilulaeformis occur in the same
subfamily. However, other reported hosts of B.
pilulaeformis occur in the more distantly related subfamilies Panicoideae and Bambusoideae, and A. texensis infects only Stipa leucotrica in the tribe Stipeae (subfamily Pooideae). It is apparent that species of Atkinsonella and Balansia encom-
pass a broader range of hosts than do Epichloe typhina and its Acremonium anamorphs (Clay, 1988).
Rykard et al. (1984) suggested that, based on their similarities, there was little utility in sep- arating A. hypoxylon and B. pilulaeformis into different genera, and that the genus Atkinsonella was hard to justify. The results presented here, along with earlier work (Leuchtmann and Clay, 1988, 1989), lead us to agree that the two taxa do not belong in separate genera but to disagree that both taxa necessarily belong in the genus
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

Clay and Frentz: Epiphytic Balansia Species 533
Balansia. We suggest that epiphytic species of Atkinsonella and Balansia should be placed in a distinct genus from endophytic Balansia species. Balansia claviceps Speg. is of special interest with respect to growth habit in that it is the type spe? cies of the genus (Spegazzini, 1885). If it is epi? phytic, then Atkinsonella species could be re- classified in the genus Balansia, while endophytic species of Balansia, such as B. obtecta, could be classified into a distinct genus. In contrast, were B. claviceps endophytic, then epiphytic species of Balansia should be transferred to the genus Atkinsonella (or another genus), including B. cy- peri, B. pallida, and B. pilulaeformis along with all other species of Balansia shown to be epi? phytic in the future.
This presumes that epiphytism is a monophy- letic character. The distinction between the epi? phytic and endophytic growth habits represents an important taxonomic character that has con- siderable functional significance. This trait has
largely been ignored, even though it can be easily determined by microscopic examination of in? fected host material, living or dead. Recently published DNA sequence data (Schardl et al., 1991) indicate that A. hypoxylon and B. obtecta are distinct from Epichloe and Acremonium but similar data are required from additional epi? phytic and endophytic species to test the hy- pothesis that epiphytic species of Atkinsonella and Balansia form a monophyletic group.
There is a precedent for recognizing growth habit on host plants as an important taxonomic character. Subfamilies in the powdery mildews
(Erysiphaceae, Ascomycotina) are differentiated
largely on the basis of epiphytic versus endo?
phytic mycelium (Braun, 1987). This represents a higher level of taxonomic classification than genera.
One possible objection to our revised generic concept concerns the primacy of conidial devel? opment and morphology as a taxonomic char? acter. This represented the basis of DiehFs clas? sification of the tribe, with the genera being distinguished on the basis ofthe presence or ab? sence of ephelidial and typhodial (Acremonium- like) conidia. However, the significance of the two forms of conidia in Atkinsonella has been questioned (Rykard et al., 1984) and the micro- conidal state in A. hypoxylon and A. texensis has been shown to differ significantly in morphology and development (Leuchtmann and Clay, 1989). Further, the variable production of the Ephelis
state in B. pilulaeformis as reported by Diehl
(1950) and Rykard et al. (1984) suggests that en- vironmental effects may be important. Diehl has also reported that some Balansia species may produce phialidic conidia under conditions of extreme moisture. The reports of several poorly studied Epichloe species with distinct and un- usual host ranges (Morgan-Jones et al., 1992) further muddy the systematic relationships in the tribe. While reproductive characters form the ba? sis of most taxonomies, significant differences in
vegetative morphology should not be ignored, especially when they are correlated with differ? ences in fungal ecology, physiology, and repro- duction.
ACKNOWLEDGMENTS
We thank Dr. Val Monteiro for help with the sec- tioning and staining, Amos Yan for help with the pho- tography, and John Schmidt for bringing infected C. laxum to our attention, and to Clint Turbeville, who suffered numerous chigger bites while collecting plants. Permission to collect material from the Hobcaw Bar- ony was kindly provided by Dr. George Askew. Drs. M. R. Tansey and D. Moore, and one anonymous re- viewer provided helpful comments on an earlier ver- sion of this manuscript. This research was supported by NSF grant BSR-9006858 to K. Clay.
LITERATURE CITED
Braun, U. 1987. A monograph ofthe Erysiphales (powdery mildews). Nova Hedwigia 89: 1-700.
Clay, K. 1986a. Grass endophytes. Pp. 188-204. In: Microbiology ofthe phyllosphere. Eds., N. J. Fok- kema and J. van den Heuvel. Cambridge Univer? sity Press, Cambridge, United Kingdom.
-. 1986b. Induced vivipary in the sedge Cyperus virens and the transmission ofthe fungus Balansia cyperi (Clavicipitaceae). Canad. J. Bot. 64: 2984- 2988.
1988. Fungal endophytes of grasses: a defen- sive mutualism between plants and fungi. Ecology 69: 10-16.
Deighton, F. C. 1956. Diseases of cultivated and eco? nomic plants in Sierra Leone. Eyre and Spottis- wood Ltd., Margate, United Kingdom.
Diehl, W. W. 1950. Balansia and the Balansiae in America. USDA, Washington, D.C.
Fomba, S. N. 1984. Rice disease situation in man? grove and associated swamps in Sierra Leone. Trop. Pest Managem. 30: 73-81.
Gould, F. W., and R. B. Shaw. 1983. Grass system? atics. Texas A&M University Press, College Sta? tion.
Leuchtmann, A., & K. Clay. 1988. Atkinsonella hy? poxylon and Balansia cyperi, epiphytic members ofthe Balansiae. Mycologia 80: 192-199.
-, and-. 1989. Morphological, cultural
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions

534 Mycologia
and mating studies on Atkinsonella, including A. texensis. Mycologia 81: 692-701.
Luttrell, E. S., and C. W. Bacon. 1977. Classification of Myriogenospora in the Clavicipitaceae. Canad. J. Bot. 55: 2090-2097.
Margolena, L. A. 1932. Erythrosin for Stoughton's thionin orange G. Stain Technol. 7: 25-27.
Morgan-Jones, G., R. A. Phelps, and J. F. White, Jr. 1992. Systematic and biological studies in the Balansieae and related anamorphs. I. Prologue. Mycotaxon 43: 401-415.
Ou, S. 1972. Rice diseases. Eastern Press Ltd., Lon? don, United Kingdom.
Porter, J. K., C. W. Bacon, H. G. Cutler, R. F. Arren- dale, and J. D. Robbins. 1985. In vitro auxin production by Balansia epichloe. Phytochemistry 24: 1429-1431.
Rykard, D. M., C. W. Bacon, and E. S. Luttrell. 1985. Host relations of Myriogenospora atramentosa and Balansia epichloe (Clavicipitaceae). Phytopathol? ogy 75: 950-956.
-,E.S. Luttrell, and C.W. Bacon. 1984. Conid- iogenesis and conidiomata in the Clavicipitoideae. Mycologia 76: 1095-1103.
Sampson, K. 1933. The systematic infection of grass? es by Epichloe typhina (Pers.) Tul. Trans. Brit. Mycol. Soc. 18: 30-47.
Schardl, C. L., J. Liu, J. F. White, Jr., R. A. Finkel, Z. An, and M. R. Siegel. 1991. Molecular phy? logenetic relationships of nonpathogenic grass my- cosymbionts and clavicipitaceous plant patho? gens. Pl. Syst. Evol. 178: 27-41.
Schneider, H. 1981. Plant anatomy and general bot? any. Pp. 315-333. In: Stainingprocedures used by
the Biological Stain Commission. 4th ed. Ed., G. Clark. The Williams and Wilkins Co., Baltimore, Maryland.
Siegel, M. R., G. C. M. Latch, and M. C. Johnson. 1987. Fungal endophytes of grasses. Annual Rev. Phytopathol. 25: 293-315.
Smith, K. T., C. W. Bacon, and E. S. Luttrell. 1985. Reciprocal translocation of carbohydrates be? tween host and fungus in bahiagrass infected with Myriogenospora atramentosa. Phytopathologyl5: 407-411.
Speggazini, C. 1885. Fungi Guaranitici. Pugillis I. Soc. Cient. ArgentinaAn. 19: 34-48, 91-96, 241- 265.
Stoughton, R. H. 1930. Thionin and orange G for the differential staining of bacteria and fungi in plant tissues. Ann. Appl. Biol. 17: 162-164.
Stovall, M. E. 1988. An investigation of the fungus Balansia cyperi Edg. and its effects on purple nut- sedge Cyperus rotundus L. Ph.D. Dissertation, Louisiana State Univ., Baton Rouge. 165 pp.
White, J. F., Jr., J. P. Breen, and G. Morgan-Jones. 1991b. Substrate utilization in selected Acremon? ium, Atkinsonella and Balansia species. Mycologia 83: 601-610.
-, A. C. Morrow, G. Morgan-Jones, and D. A. Chambless. 1991a. Endophyte-host associations in forage grasses. XIV. Primary stromata forma? tion and seed transmission in Epichloe typhina: developmental and regulatory aspects. Mycologia 83: 72-81.
Accepted for publication March 8, 1993
This content downloaded from 129.79.232.61 on Mon, 16 Sep 2013 14:36:44 PMAll use subject to JSTOR Terms and Conditions





























