Embed Size (px)
Citation preview

BLS
Ba
o1b
R
ArcmgvcdrHliecodtLwtBpgigpt
dnLagI
Ki
Gwt
*EAtNp
Neuroscience 127 (2004) 695–707
0d
I-DIRECTIONAL ASSOCIATIONS BETWEEN GALANIN ANDUTEINIZING HORMONE-RELEASING HORMONE NEURONAL
YSTEMS IN THE HUMAN DIENCEPHALONwicrhga1
witl(dpgttigmndvgihn
arecw(gcloonopc1na
. DUDASa* AND I. MERCHENTHALERb
Laboratory of Neuroendocrine Organization (NEO), Lake Erie Collegef Osteopathic Medicine, 1858 West Grandview Boulevard, Erie, PA6509-1025, USA
Women Health Research Institute, Wyeth Research, 500 Arcolaoad, Collegeville, PA 19426, USA
bstract—Evidence suggests that galanin plays an importantole in the regulation of reproduction in the rat. Galanin isolocalized with luteinizing hormone (LH)–releasing hor-one (LHRH) in a subset of LHRH neurons in female rats andalanin-immunoreactive (galanin-IR) nerve terminals inner-ate LHRH neurons. Recent studies indicate that galanin mayontrol gonadal functions in rats at two different levels: (i) viairect modulation of pituitary LH secretion and/or (ii) indi-ectly via the regulation of the hypothalamic LHRH release.owever, the morphological substrate of any similar modu-
ation is not known in human. In the present series of exper-ments we first mapped the galanin-IR and LHRH-IR neurallements in human brain, utilizing single label immunohisto-hemistry. Then, following the superimposition of the mapsf these systems, the overlapping sites were identified withouble labeling immunocytochemistry and examined in ordero verify the putative juxtapositions between galanin-IR andHRH-IR structures. LHRH and galanin immunoreactivityere detected mainly in the medial basal hypothalamus, in
he medial preoptic area and along the diagonal band ofroca. Careful examination of the IR elements in the overlap-ing areas revealed close, bi-directional contacts betweenalanin-IR and LHRH-IR structures, which have been verified
n semithin plastic sections. These galanin-LHRH and LHRH-alanin juxtapositions were most numerous in the medialreoptic area and in the infundibulum/median eminence ofhe human diencephalon.
In conclusion, the present study is the first to reveal bi-irectional juxtapositions between galanin- and LHRH-IReural elements in the human diencephalon. These galanin–HRH and LHRH–galanin contacts may be functional syn-pses, and they may be the morphological substrate of thealanin-controlled gonadal functions in humans. © 2004
BRO. Published by Elsevier Ltd. All rights reserved.
ey words: GnRH, hypothalamus, immunocytochemistry,nnervation, interaction, synapse.
alanin, a 29 amino acid peptide (Tatemoto et al., 1983) isidely distributed in the CNS and peripheral nervous sys-
em. In rodents, galanin is an important cellular messenger
Corresponding author. Tel: �1-814-866-8142; fax: �1-814-866-8411.-mail address: [email protected] (B. Dudas).bbreviations: DAB, diaminobenzidine; IR, immunoreactive; LH, lu-
einizing hormone; LHRH, luteinizing hormone–releasing hormone;HS, normal horse serum; PAP, peroxidase–antiperoxidase; PBS,
Mhosphate buffer containing 0.9% sodium chloride.
306-4522/04$30.00�0.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.05.018
695
ithin the CNS, modulating diverse physiological functionsncluding nociception (Liu et al., 2001; Wynick et al., 2001),ognition (Steiner et al., 2001), and many aspects of neu-oendocrine activity that are associated with feeding be-avior and reproduction. As a neuroendocrine regulator,alanin induces the release of prolactin, growth hormone,nd luteinizing hormone (LH; see Merchenthaler et al.,993; Crawley, 1995; Hokfelt et al., 1998 for review).
Galanin may regulate LH secretion in two possibleays: (i) directly at the level of the pituitary and/or (ii)
ndirectly, at the level of the diencephalon via the regula-ion of hypothalamic LH-releasing hormone (LHRH) re-ease. Galanin may directly control pituitary LH secretionKaplan et al., 1988) and/or galanin synthesized in theiencephalon may also reach gonadotroph cells in theituitary via the hypophyseal portal circulation. In fact,alanin levels are higher in the hypophyseal portal bloodhan in the peripheral circulation and galanin release intohe hypophyseal portal circulation follows a pulsatile fash-on (Lopez et al., 1991). These observations suggest thatalanin functions as a hypothalamic, hypophysiotropic hor-one. Retrograde labeling studies from the median emi-ence confirmed these functional observations in rats byemonstrating labeled neurons in the arcuate and para-entricular nuclei following peripheral injection of fluoro-old (Merchenthaler, 1991a,b). A pituitary action of galanin
s also supported by the observations that galanin en-ances LHRH binding to LHRH receptors of pituitary go-adotrophs (Sahu et al., 1994).
Galanin (Skofitsch and Jacobowitz, 1985; Melander etl., 1986b; Gai et al., 1990; Merchenthaler et al., 1993) and itseceptors (Melander et al., 1986a; Servin et al., 1987; Mitchellt al., 1999; Branchek et al., 2000) are present in largeoncentration in the preoptic area of the hypothalamus,here the majority of LHRH synthesizing neurons reside
Merchenthaler et al., 1984). In addition to being in a strate-ically important location of the hypothalamus, galanin iso-expressed with LHRH in neurons of the rat (Merchentha-
er et al., 1990), mouse (Rajendren and Gibson, 1999) andvine (Dufourny et al., 2003) medial preoptic area and diag-nal band of Broca. The colocalization of these peptides hasot been reported in primates, including humans. The degreef co-expression is estrogen-dependent and sexually dimor-hic (Merchenthaler et al., 1990, 1991) (for reviews see Mer-henthaler et al., 1993; Herbison, 1998; Merchenthaler,998). Interestingly, the expression of galanin within LHRHeurons is stimulated by estrogen in rats (Merchenthaler etl., 1991) but inhibited in mice (Rajendren and Gibson, 1999).
oreover, galanin-immunoreactive (IR) fiber varicosities abutved.

LcbL
t(toiZlrtoh
aee2dt1pecg
gTpgqmtmuL
T
Fynwao
T
Fw02v3((ic
isLd
I
SawGfiemtt1ssTgCosbgchs21vTddpnfM
wginwd
tseD
r(n
C
TsUTTs6aa(
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707696
HRH-IR perikarya in the preoptic area of the rat (Mer-henthaler et al., 1991) and mouse (Rajendren and Li, 2001)rain, suggesting a direct synaptic control on hypothalamicHRH secretion.
Although numerous data suggest that galanin is one ofhe major modulators of reproductive functions in ratLopez et al., 1991; Sahu et al., 1994), little is known abouthe role galanin plays in the regulation of gonadal functionsf human. It has been suggested that galanin participates
n the generation of the LHRH surge (Pau and Spies, 1997;drojewicz and Sowinska, 2000); however, the morpho-
ogical substrate of the putative galanin–LHRH interactionemains elusive. In contrast to known actions in the hypo-halamus, galanin has not been shown to stimulate basalr LHRH-stimulated LH concentrations in the pituitary ofumans (Giustina et al., 1994; Arvat et al., 1995).
The distribution and morphology of LHRH system haslready been described in humans (King et al., 1985; Stopat al., 1991; Rance et al., 1994; Silverman et al., 1994; Dudast al., 2000; Dudas and Merchenthaler, 2001, 2002a,b,003), however, no detailed information is available on theistribution of galanin immunoreactivity in the human hypo-halamus (Skofitsch and Jacobowitz, 1985; Melander et al.,986b; Gai et al., 1990; Merchenthaler et al., 1993). Since theutative juxtapositions between galanin-IR and LHRH-IRlements may be the morphological substrate of galanin-ontrolled LH release, it is crucial to reveal the sites wherealanin and LHRH systems overlap.
In the present study, we first mapped the LHRH-IR andalanin-IR neuronal elements of the human hypothalamus.hen, the maps of LHRH and galanin systems were su-erimposed, and the putative synapses between thealanin-IR and LHRH-IR elements were identified anduantified in the overlapping areas using double-label im-unohistochemistry. Since the long post mortem time of
he human samples made the application of the electronicroscopy virtually impossible, semithin sections weresed to study the fine structure of the putative galanin-HRH synapses.
EXPERIMENTAL PROCEDURES
issue samples
our human brains (two adult women and two adult men, 36–84ears of age) from individuals with no history of neurological andeuroendocrinological diseases were used. The brain samplesere obtained from autopsies at 24–48 h post mortem period, inccordance with the regulation and permission of the Ethics Boardf Szent-Gyorgyi Albert Medical University, Szeged, Hungary.
issue preparation
ollowing the removal of the brains from the skull, the hypothalamiere dissected and the tissue blocks were fixed by immersion in.1 M phosphate-buffered (pH 7.4) 4% formaldehyde at 4 °C for–24 weeks. Each block contained half of the hypothalamus di-ided in the midsagittal line. The samples were cryoprotected with0% sucrose in phosphate buffer containing 0.9% sodium chloridePBS) supplemented with 0.1% sodium-azide. Coronal sections30 �m) were cut on a freezing microtome (Reichert) and collectedn four series of wells of plastic 24-compartment plates with PBS
ontaining 0.2% sodium-azide, and stored at 4 °C until process- ang. The adjacent sections were processed as follows: (i) Nissltaining with Cresyl Violet, (ii) immunohistochemical detection ofHRH, (iii) immunohistochemical detection of galanin and (iv)ouble-label immunohistochemistry of galanin and LHRH.
mmunohistochemistry
ingle-labeling was carried out using either peroxidase–ntiperoxidase (PAP) or streptavidin–biotin methods combinedith silver intensification introduced by Gallyas et al. (1982) andallyas and Merchenthaler (1988). The samples were pretreatedrst with 10% thioglycolic acid for 30 min to suppress the endog-nous tissue argentophilia, then with 0.2% Triton X-100 for 20in, and 10% normal horse serum (NHS) in PBS for 1 h at room
emperature to block non-specific staining. Thereafter, the sec-ions were incubated in primary antisera solution composed of0% NHS, PBS and 0.1% sodium azide for 24 h. The LHRH-IRtructures were identified using a monoclonal mouse LHRH anti-erum (Chemicon, Temecula, CA, USA) at a dilution of 1:20,000.he galanin-IR structures were visualized with a rabbit anti-alanin serum (RAS 7153N; Peninsula Laboratories, Belmont,A, USA; 1:3000). The sections were then incubated in a sec-ndary antiserum containing unlabeled goat anti-rabbit IgG (Jack-on, West Grove, PA, USA; at a dilution of 1:50) for galanin, oriotin-labeled horse anti-mouse IgG (Vector Laboratories, Burlin-ame, CA, USA; 1:500) for 2 h to detect LHRH. In each case, theross-reactions of the secondary antibodies with the endogenousuman IgGs were eliminated by the addition of 2% normal humanerum to these reagents. Finally, the sections were incubated forh in rabbit PAP (Jackson Immunocytochemical; at a dilution of
:2000) for the detection of galanin, or peroxidase-labeled strepta-idin (Jackson; at a dilution of 1:500) for the detection of LHRH.he chromogen solution was composed of 0.05% diaminobenzi-ine (DAB), 0.125% nickel–ammonium–sulfate and 0.005% hy-rogen peroxide in 0.1 M Tris–HCl (pH 7.6). The resulting DABolymer was silver-intensified in a solution containing 0.1% silveritrate, 0.1% ammonium-nitrate, 1% silicotungstic acid and 0.2%ormaldehyde for 3–4 min (Gallyas et al., 1982; Gallyas anderchenthaler, 1988).
Simultaneous detection of galanin-IR and LHRH-IR structuresas performed using double-label immunohistochemistry. First,alanin immunohistochemistry was carried out using silver-
ntensified Ni-DAB as chromogen, and then the LHRH-containingeuronal structures were immunolabeled using DAB chromogenithout nickel ions. The anti-LHRH serum was used at a 1:8000ilution.
Some of the double-labeled sections were embedded in plas-ic in order to obtain thin (1 �m) sections. The large, frozenections were cut in smaller pieces, processed through a gradedthanol series and propylene oxide, and then flat embedded inurcupan. Semithin sections were cut with an ultramicrotome.
In control sections, the primary antibodies were omitted oreplaced with non-immune rabbit sera at the dilution of 1:3000galanin) or 1:20,000 (LHRH), respectively. Immunoreaction wasot observed.
omputer-assisted mapping
he Nissl-stained hypothalamic sections were systematicallycanned using a plain-scanner (Hewlett-Packard, Palo Alto, CA,SA), and the outlines of the sections were traced with the Corel-race software 4.0 (Corel Corporation, Ottawa, Ontario, Canada).he computer-generated superimposition of eight, consecutiveections was summarized in each of the slides of Figs. 1, 2, 5 anddenoted by letters from A–P. Each of these slides demonstrates960 �m thick section of the human diencephalon. The neurons
nd fibers have been marked on these figures using a ZeissThornwood, NY, USA) Axiophot microscope with camera lucida,
nd Adobe (San Jose, CA, USA) Photoshop software, version 3.0.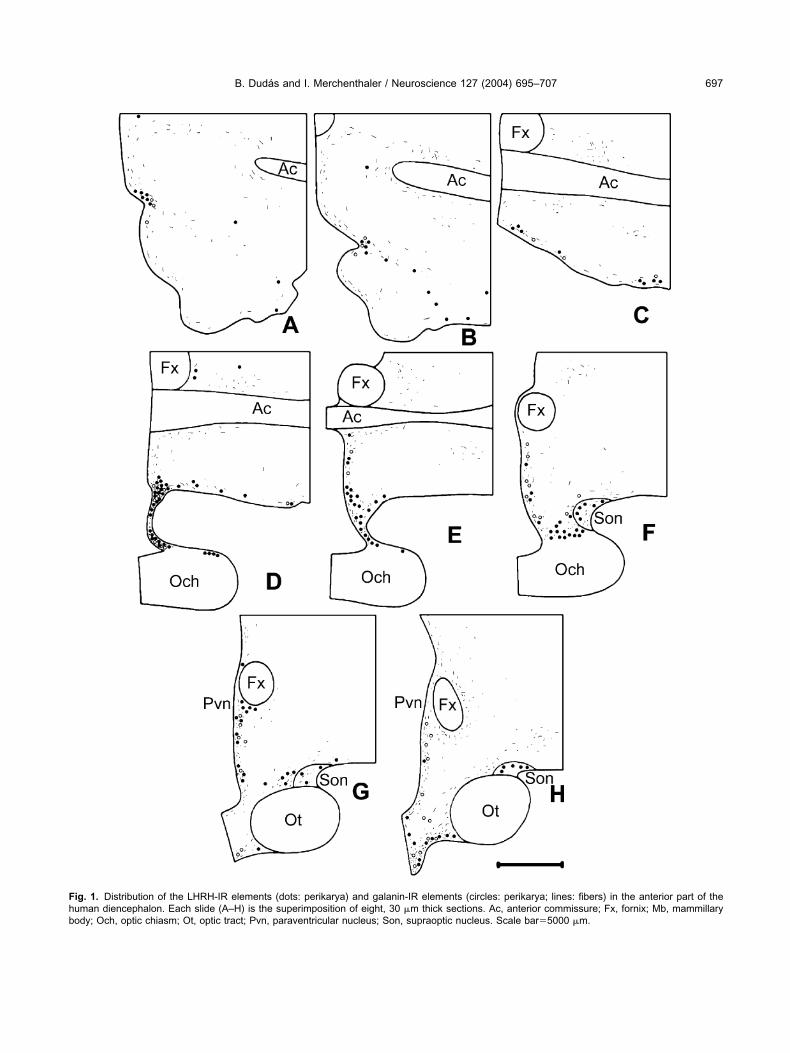
Fhb
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707 697
ig. 1. Distribution of the LHRH-IR elements (dots: perikarya) and galanin-IR elements (circles: perikarya; lines: fibers) in the anterior part of theuman diencephalon. Each slide (A–H) is the superimposition of eight, 30 �m thick sections. Ac, anterior commissure; Fx, fornix; Mb, mammillaryody; Och, optic chiasm; Ot, optic tract; Pvn, paraventricular nucleus; Son, supraoptic nucleus. Scale bar�5000 �m.
Fhp
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707698
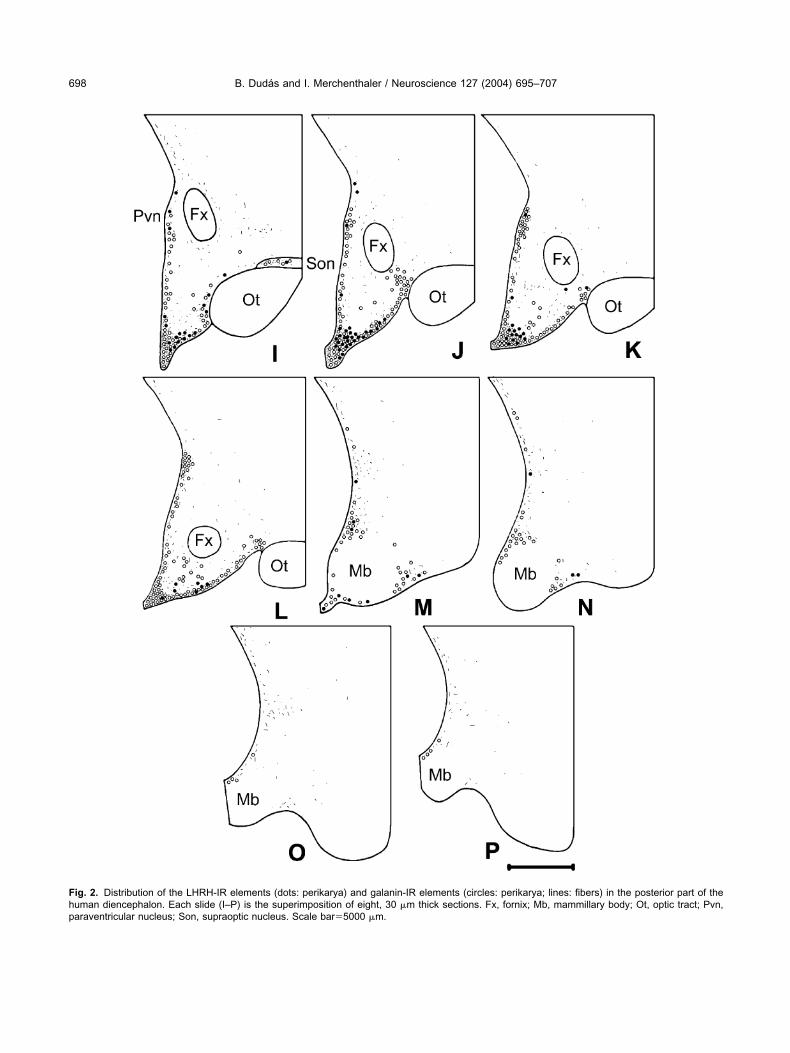
ig. 2. Distribution of the LHRH-IR elements (dots: perikarya) and galanin-IR elements (circles: perikarya; lines: fibers) in the posterior part of theuman diencephalon. Each slide (I–P) is the superimposition of eight, 30 �m thick sections. Fx, fornix; Mb, mammillary body; Ot, optic tract; Pvn,araventricular nucleus; Son, supraoptic nucleus. Scale bar�5000 �m.

T
TB(
L
Lp(flzph12focDiivttsf
G
TcdlaI
suotra(rtnp(gbl
spzrswp3
iLtFbstbpcl
Fta n eminen
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707 699
erminology
he terminology of the diencephalic structures was adapted fromraak and Braak (1987), Saper (1990), and Silverman et al.
1994).
RESULTS
HRH system in the human hypothalamus
HRH-IR perikarya were located mainly in the medialreoptic area and in the infundibulum/median eminenceFigs. 1H, 2I–L). Numerous LHRH-IR cell bodies wereound along the diagonal band of Broca (Fig. 1B), in theamina terminalis (Fig. 1D), and in the periventricularone of the tuberal region (Figs. 1G–H, 2I–K). Fewerikarya were detected periventricularly in the posteriorypothalamus (Fig. 2M, N), and in the supraoptic (Figs.F–H, 2I) and the paraventricular nuclei (Figs. 1G, H,I). The vast majority of the LHRH-IR neurons were
usiform in shape with processes emanating from thepposite poles of the cell body (Fig. 4). Few multipolarells were also detected in the medial preoptic region.ense LHRH-IR fiber networks were observed in the
nfundibulum/median eminence, where the fiber varicos-ties were often in intimate relationship with the portalessels. The periventricular area of the chiasmatic anduberal regions also contained scattered LHRH-IR axonshat were running, characteristically, parallel with theurface of the 3rd ventricle. Few LHRH-IR fibers wereound along the diagonal band of Broca.
alanin system in the human hypothalamus
he majority of the galanin-IR perikarya (96%) wereoncentrated posteriorly to the lamina terminalis in fourifferent regions. (i) The infundibulum contained dense-
y-packed galanin-IR cell bodies that were often locatedround the portal vessels (Figs. 2I, J; 3C). (ii) Numerous
ig. 3. Galanin-IR elements in the human diencephalon. (A) Fusiformuberal region. (B) Galanin-IR fibers abut on a multipolar galanin-IR nexon varicosities in intimate proximity of the portal vessels in the media
R perikarya were detected in a narrow zone at the basal l
urface of the tuberal region (Fig. 2J–L). This cell pop-lation extended from the infundibular nucleus to theptic tract. (iii) Several galanin-IR perikarya were de-
ected periventricularly, in the chiasmatic and tuberalegions (Figs. 1E–H, 2I–L). The paraventricular nucleuslso contained galanin-IR cell bodies (Figs. 1G, H, 2I).iv) In the posterior hypothalamus, the galanin-IR neu-ons were concentrated in the posterior subdivision ofhe periventricular nucleus and in the tuberomammillaryucleus (Fig. 2M, N). Few perikarya were detected in theeriventricular region of the posterior hypothalamusFig. 2M–P). In front of the lamina terminalis, scatteredalanin-IR cell bodies were observed along the diagonaland of Broca (Fig. 1B, C). Few cells were found in the
amina terminalis itself (Fig. 1D).The galanin-IR neurons were mainly fusiform in
hape with processes emanating from the oppositeoles of the perikarya (Fig. 3A, C). In the periventricularone, the galanin-IR cells were characteristically ar-anged with their longitudinal axis running parallel to theurface of the 3rd ventricle. Multipolar galanin-IR cellsere detected mainly in the infundibulum and in theosterior subdivision of the periventricular nucleus (Fig.B).
Galanin-IR fiber varicosities formed a dense networkn the infundibulum/median eminence (Figs. 1G, H, 2I–). Numerous axons were detected periventricularly, in
he chiasmatic and tuberal regions (Figs. 1E–H, 2I–L).ew galanin-IR fibers were observed along the diagonaland of Broca (Fig. 1B, C), around the anterior commis-ure and fornix (Figs. 1B–H, 2I–L), and in the laminaerminalis (Fig. 1D). In the posterior hypothalamus, fi-ers were detected in the posterior subdivision of theeriventricular nucleus, in the tuberomammillary nu-leus and in the periventricular region (Fig. 2M–P). Ga-
anin-IR axon varicosities were scarcely observed in the
R neuron receiving a galanin-IR fiber in the periventricular area of thehe infundibular nucleus. (C) Galanin-IR cell body receives galanin-IRce. The lumen of the vessel is marked by asterisk. Scale bar�20 �m.
galanin-Iuron in t
ateral hypothalamus.
Fn1cipapa
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707700
ig. 4. Juxtapositions between the LHRH (brown) and galanin (black) IR elements in the human diencephalon. The position of the demonstrated LHRH-IReurons is shown by asterisks in the upper corners of the micrographs. The schematic drawings correspond to the slices denoted by the same letters in Fig.. Most of the galanin-LHRH juxtapositions are located in the medial part of the infundibulum/median eminence, where 77% of the LHRH-IR perikarya wereontacted with more than three galaninergic fiber varicosities (A–C, G–I). Interestingly, several LHRH-IR perikarya received both galanin and LHRH-IR axons
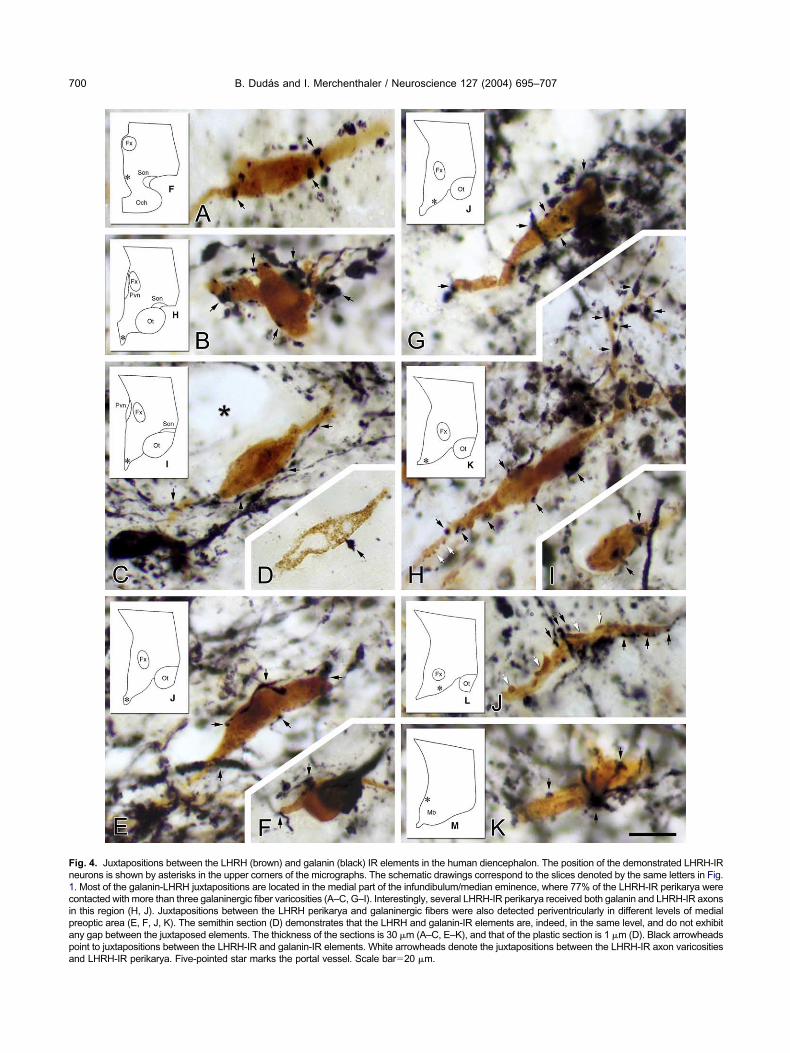
n this region (H, J). Juxtapositions between the LHRH perikarya and galaninergic fibers were also detected periventricularly in different levels of medialreoptic area (E, F, J, K). The semithin section (D) demonstrates that the LHRH and galanin-IR elements are, indeed, in the same level, and do not exhibitny gap between the juxtaposed elements. The thickness of the sections is 30 �m (A–C, E–K), and that of the plastic section is 1 �m (D). Black arrowheadsoint to juxtapositions between the LHRH-IR and galanin-IR elements. White arrowheads denote the juxtapositions between the LHRH-IR axon varicositiesnd LHRH-IR perikarya. Five-pointed star marks the portal vessel. Scale bar�20 �m.
FpiLLb
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707 701
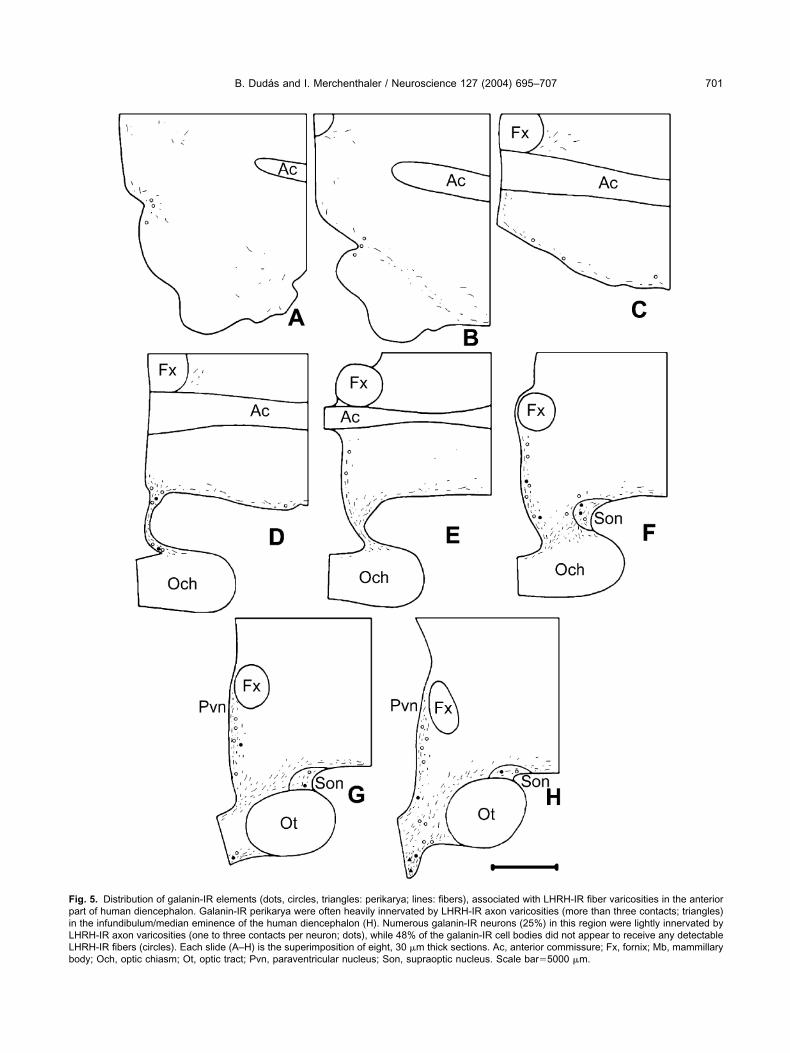
ig. 5. Distribution of galanin-IR elements (dots, circles, triangles: perikarya; lines: fibers), associated with LHRH-IR fiber varicosities in the anteriorart of human diencephalon. Galanin-IR perikarya were often heavily innervated by LHRH-IR axon varicosities (more than three contacts; triangles)
n the infundibulum/median eminence of the human diencephalon (H). Numerous galanin-IR neurons (25%) in this region were lightly innervated byHRH-IR axon varicosities (one to three contacts per neuron; dots), while 48% of the galanin-IR cell bodies did not appear to receive any detectableHRH-IR fibers (circles). Each slide (A–H) is the superimposition of eight, 30 �m thick sections. Ac, anterior commissure; Fx, fornix; Mb, mammillaryody; Och, optic chiasm; Ot, optic tract; Pvn, paraventricular nucleus; Son, supraoptic nucleus. Scale bar�5000 �m.
FpibLp
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707702
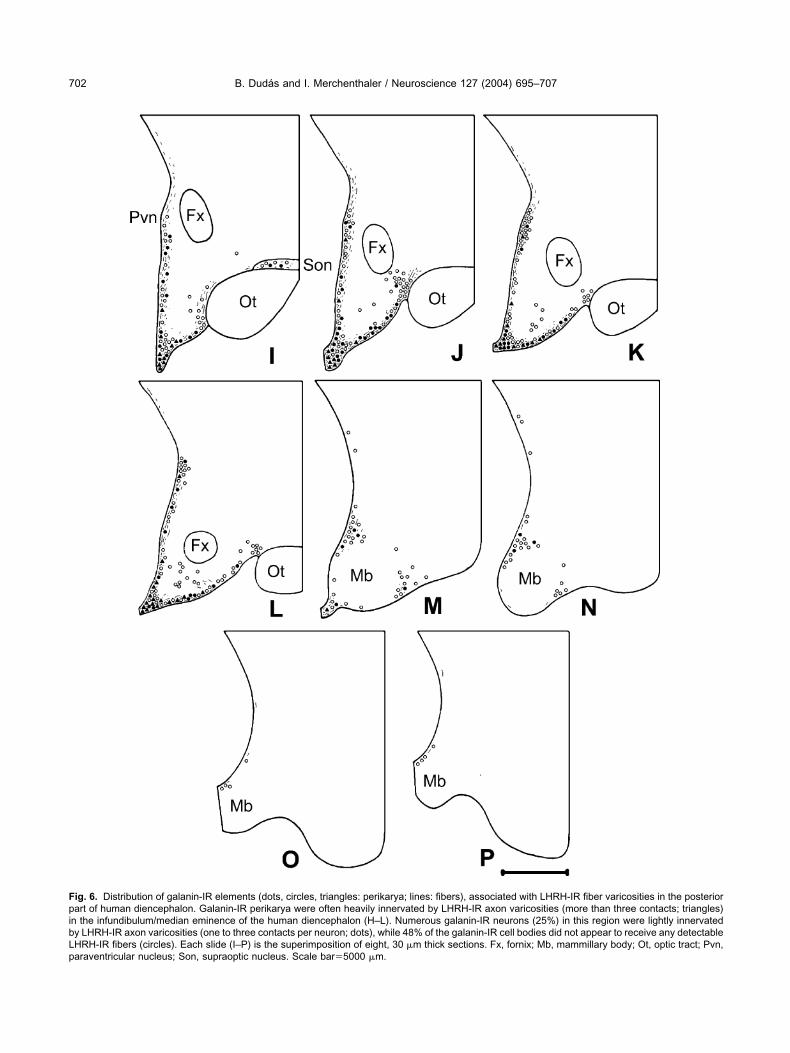
ig. 6. Distribution of galanin-IR elements (dots, circles, triangles: perikarya; lines: fibers), associated with LHRH-IR fiber varicosities in the posteriorart of human diencephalon. Galanin-IR perikarya were often heavily innervated by LHRH-IR axon varicosities (more than three contacts; triangles)
n the infundibulum/median eminence of the human diencephalon (H–L). Numerous galanin-IR neurons (25%) in this region were lightly innervatedy LHRH-IR axon varicosities (one to three contacts per neuron; dots), while 48% of the galanin-IR cell bodies did not appear to receive any detectableHRH-IR fibers (circles). Each slide (I–P) is the superimposition of eight, 30 �m thick sections. Fx, fornix; Mb, mammillary body; Ot, optic tract; Pvn,araventricular nucleus; Son, supraoptic nucleus. Scale bar�5000 �m.

G
IittlbttpLbpattptpatrg
L
IaimgsctctdwvattLl
J
Ia(wpr3et
J
Nfi
(aatt
TeocmiaprpLlpr
ohptcthwgcriAtvtl(stt
L(rcsBJettttt
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707 703
alanin–LHRH juxtapositions
ntimate contacts between the galanin-IR axon varicos-ties and LHRH-IR perikarya were observed mainly inhe infundibulum/median eminence of the human hypo-halamus (Fig. 4B–D). Here 77% of the brown, DAB-abeled LHRH-IR neurons were heavily contacted bylack, silver-intensified galanin fibers receiving morehan three contacts. The rest of the LHRH neurons inhis region received only a few (less then three contactser neuron). Numerous, LHRH-IR cells received bothHRH and galanin-IR axon varicosities in the medialasal hypothalamus (Fig. 4H, J). Galanin-LHRH juxta-ositions have also been found in the medial preopticrea (Fig. 4A) and in the periventricular zone of theuberal region where 24% of the LHRH-IR were con-acted by galanin-IR nerve processes. Few LHRH-IRerikarya were juxtaposed by galanin-IR fibers (one to
hree contacts) in the posterior subdivision of theeriventricular nucleus (Fig. 4K), in the lamina terminalisnd along the diagonal band of Broca. Close examina-ion of semithin sections of these intimate associationsevealed no gap between the juxtaposing LHRH-IR andalanin-IR neural elements (Fig. 4D).
HRH-galanin juxtapositions
ntimate associations between LHRH-IR nerve terminalsnd galanin-IR cell bodies were observed mainly in the
nfundibulum/median eminence of the human hypothala-us (Figs. 6I–M; 7). Here 27% of the brown, DAB-labeledalanin-IR perikarya were heavily contacted by black,ilver-intensified LHRH-IR fibers receiving more than threeontacts (Fig. 7). Numerous galanin-IR neurons (25%) inhis region were lightly innervated by LHRH-IR axon vari-osities (one to three contacts per neuron), while 48% ofhe galanin-IR cell bodies did not appear to receive anyetectable LHRH-IR fibers. Fewer galanin-IR perikaryaere contacted by LHRH-IR nerve terminals in the para-entricular nucleus (Fig. 6I–K), anterior periventricularrea (Fig. 5D–G) and in the periventricular area of theuberal region, between the infundibulum and the paraven-ricular nucleus (Figs. 5H, 6I–K). No colocalization ofHRH and galanin was observed in any of the hypotha-
amic regions studied (not shown).
uxtapositions between galanin-IR elements
ntimate associations between galanin-IR cell bodies andxon varicosities were often observed in the infundibulumFig. 3B, C). Here, the majority of the galanin-IR perikaryaere heavily contacted by galanin-IR fibers. The medialreoptic area and the periventricular zone of the tuberalegion also contained some galanin-galanin contacts (Fig.A). A few juxtapositions between the galanin-IR neurallements were also detected in the posterior subdivision of
he periventricular nucleus.
uxtapositions between LHRH-IR elements
umerous LHRH-IR perikarya juxtaposed with LHRH-IR
ber varicosities in the infundibulum/medial preoptic area mFig. 7C, F, J). Here, 8% of the LHRH-IR cell bodiesssociated by galanin-IR fibers also received LHRH-IRxon varicosities (Fig. 7C, F, J). These intimate associa-ions were en passant type juxtapositions without any de-ectable spaces between the contacting elements.
DISCUSSION
he morphology and distribution of the LHRH-IR neurallements reported in this paper are in good agreement withur previous studies (Dudas et al., 2000; Dudas and Mer-henthaler, 2001, 2002a,b, 2003). LHRH-IR perikarya areainly located in the infundibulum/median eminence and
n the medial preoptic area; few cell bodies are observedlong the diagonal band of Broca, in the lamina terminalis,eriventricularly in the tuberal and posterior hypothalamicegions, and in the tuberomammillary nucleus. The su-raoptic and paraventricular nucleus also contained fewHRH-IR neurons. LHRH-IR axon varicosities are mainly
ocalized in the infundibulum/median eminence and in theeriventricular zone of the suprachiasmatic and tuberalegions.
While previous studies provided a general descriptionf the pattern and morphology of galanin-IR neurons in theuman hypothamus (Gai et al., 1990), the present studyrovides a high-resolution map of the galanin-IR system inhe hypothalamus where LHRH-IR elements are also lo-ated. Moreover, this is the first study that precisely mapshe pattern of galanin-IR axon varicosities not only in theypothalamus but also in front of the lamina terminalis,here a subpopulation of LHRH perikarya is located. Sincealanin-IR fibers may innervate several neurotransmitter-ontaining structures in the human diencephalon and thus,egulate numerous hypothalamic functions, it is extremelymportant to describe their distribution in the diencephalon.lthough the pattern of distribution of galanin-IR neurons in
he human hypothalamus is in good agreement with pre-ious studies (Gai et al., 1990), an additional subpopula-ion of galanin-IR cell bodies was observed inside theamina terminalis and along the diagonal band of Brocapresent paper). These galanin-IR perikarya may be theource of some of the delicate galanin-IR fibers detected inhe basal part of the hypothalamus in front of the laminaerminalis.
Several animal studies indicate that galanin modulatesH release acting either directly at the level of the pituitaryKaplan et al., 1988; Sahu et al., 1994) or indirectly, via theegulation of LHRH release into the hypophysial portalirculation (Lopez et al., 1991). Since galanin is synthe-ized in both the anterior pituitary (Kaplan et al., 1988;ennet et al., 1991) and the hypothalamus (Skofitsch andacobowitz, 1985; Melander et al., 1986b; Merchenthalert al., 1993), it may regulate reproductive functions in bothhe periphery and the brain. Galanin, synthesized locally inhe anterior pituitary, has been shown to induce LH secre-ion in rats (Kaplan et al., 1988) and galanin, synthesized inhe hypothalamus, has been shown to stimulate LH secre-ion in rats (Lopez et al., 1991; Sahu et al., 1994) and
onkeys (Finn et al., 2000). Therefore, galanin, similarly to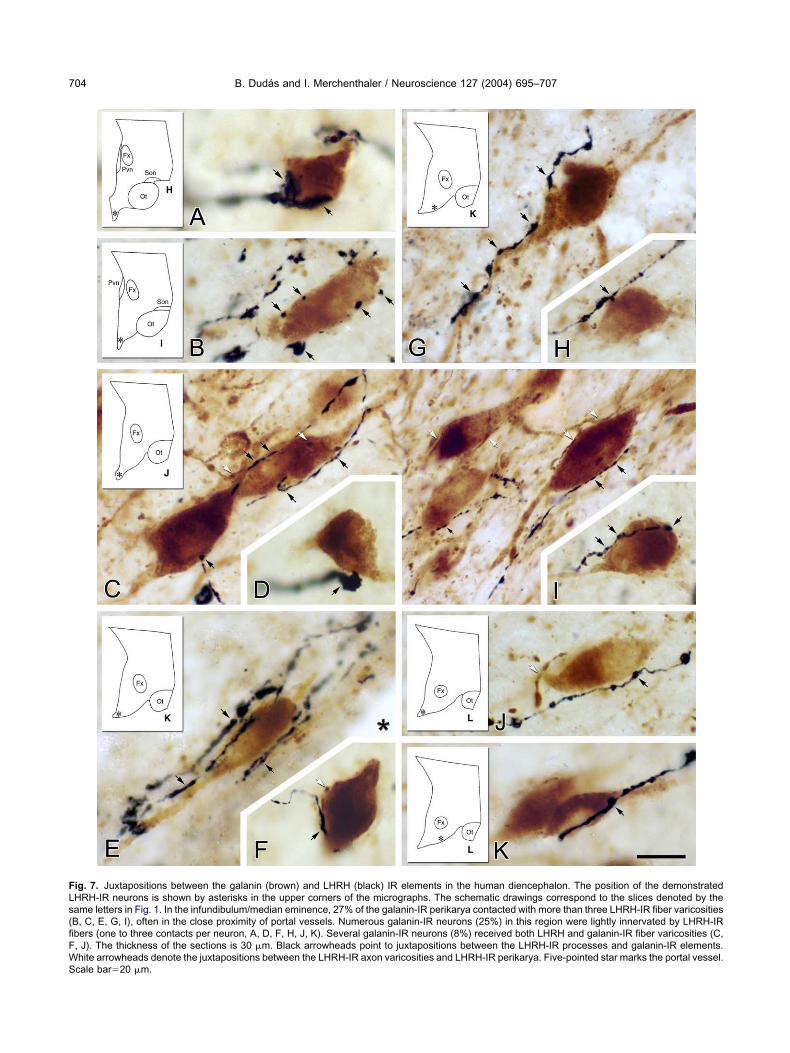
FLs(fiFWS
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707704
ig. 7. Juxtapositions between the galanin (brown) and LHRH (black) IR elements in the human diencephalon. The position of the demonstratedHRH-IR neurons is shown by asterisks in the upper corners of the micrographs. The schematic drawings correspond to the slices denoted by theame letters in Fig. 1. In the infundibulum/median eminence, 27% of the galanin-IR perikarya contacted with more than three LHRH-IR fiber varicositiesB, C, E, G, I), often in the close proximity of portal vessels. Numerous galanin-IR neurons (25%) in this region were lightly innervated by LHRH-IRbers (one to three contacts per neuron, A, D, F, H, J, K). Several galanin-IR neurons (8%) received both LHRH and galanin-IR fiber varicosities (C,, J). The thickness of the sections is 30 �m. Black arrowheads point to juxtapositions between the LHRH-IR processes and galanin-IR elements.hite arrowheads denote the juxtapositions between the LHRH-IR axon varicosities and LHRH-IR perikarya. Five-pointed star marks the portal vessel.cale bar�20 �m.

Lsgvip
LtensthtplbtsimasLtdm
oa(ntowsorcatlgagocs
tdm1semai
i1faairlsio1ocpcstc
LtmppsiL
gtmambbLmaam
AGLCab
A
B
B
B
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707 705
HRH, is also considered to be a hypothalamic hypophy-iotropic hormone. The intimate associations betweenalanin-IR fiber varicosities and the hypophyseal portalessels in the infundibulum described in the present stud-es provide morphological support for galanin as a hy-ophysiotropic peptide in the human hypothalamus.
Although several data suggest that the modulation ofH secretion by galanin occurs via the regulation of hypo-
halamic LHRH release in rats (Lopez et al., 1991; Merch-nthaler et al., 1993), the morphological basis of this phe-omenon is completely unknown in human. The presenttudy is the first to detect well-defined juxtapositions be-ween galanin-IR and LHRH-IR neural elements in theuman diencephalon. One however, has to keep in mindhat because of the shrinkage, alteration in size and mor-hology due to exposure the brain tissue to formalin for a
ong period of time, some of these close associations coulde artifacts. However, the majority of them may be func-ional synapses and may serve as a morphological sub-trate of the galanin-controlled LHRH release. Interest-ngly, numerous LHRH-IR perikarya in the infundibulum/
edian eminence were found to receive both LHRH-IRnd galanin-IR axon varicosities. These multiple juxtapo-itions were not observed on the subpopulation ofHRH-IR neurons in the medial preoptic area and periven-ricular region, indicating that LHRH release in the infun-ibulum/median eminence may be uniquely modulated byultiple galanin/LHRH inputs.
In addition to the presence of galaninergic juxtapositionn LHRH neurons in mice (Rajendren and Gibson, 2001)nd humans (present paper), galanin receptor subtype 1GAL-R1) is also expressed in a subpopulation of LHRHeurons in the female rat (Mitchell et al., 1999). Moreover,he expression of GAL-R1 within LHRH neurons is presentnly in proestrous rats or in ovariectomized rats treatedith estrogen, and progesterone blocks GAL-R1 expres-ion (Mitchell et al., 1999). Although not shown in humans,ther types of galanin receptors are also present in theodent preoptic area (Floren et al., 2000) and therefore,ould be involved in galanin-regulated LHRH neuronalctivity. Endogenous or intracerebroventricularly adminis-ered galanin probably stimulates LHRH-mediated LH re-ease via these receptors. The receptor-mediated action ofalanin is further supported by the findings that galantide,specific partial antagonist of galanin receptors, blocks
alanin-induced LH release (Sahu et al., 1994). Thesebservations provide further support for galanin as a criti-al factor in the initiation of the LHRH and subsequent LHurges prior to ovulation in the rat and possibly in human.
Another level of galanin regulated LHRH neuronal ac-ivity may involve galanin, colocalized with LHRH. A highegree of colocalization of galanin-IR and LHRH-IR ele-ents has been reported in the rat (Merchenthaler et al.,990, 1991), mouse (Rajendren and Gibson, 2001) andheep (Dufourny et al., 2003) medial preoptic area; how-ver, no such phenomenon has been reported in the hu-an hypothalamus. The lack of colocalization of LHRHnd galanin in human was confirmed by the present stud-
es. The level of galanin expression within LHRH neurons
s sexually dimorphic in rodents (Merchenthaler et al.,991; Rajendren and Gibson, 1999) but not in ovine (Du-
ourny et al., 2003) with more co-expressing neurons in thedult female than male animals. Interestingly, however, themount of galanin in LHRH neurons is negatively regulated
n mice (Rajendren and Gibson, 1999) and positively inats (Merchenthaler et al., 1991) by estrogen. Nonethe-ess, several lines of evidence suggest that galanin expres-ion increases within LHRH neurons during their activation
n proestrus or following administration of estrogen tovariectomized rats (for review see Merchenthaler et al.,993; Herbison, 1998; Gundlah et al., 2002). The functionf galanin, co-localized and co-released with LHRH (Mer-henthaler et al., 1990), probably is to further enhance theulsatile release of LHRH into the hypophyseal portal cir-ulation. The findings in rats, mice, and sheep suggestpecies differences not only in the presence of colocaliza-ion of galanin with LHRH but in the mechanism of galanin-ontrolled LHRH release.
Our studies also indicate, for the first time, thatHRH-IR nerve terminals juxtapose galanin-IR perikarya inhe human hypothalamus. Via these connections, LHRHay modulate galanin neuronal activity. Although no mor-hological or functional/pharmacological data have beenublished addressing such connections, the present ob-ervations in the human hypothalamus should provide the
mpetus to confirm the existence and functionality ofHRH-galanin connections in rodent models.
In conclusion, the bi-directional juxtapositions betweenalanin-IR and LHRH-IR neural elements demonstrated inhe present study may be functioning synapses forming theorphological basis of the galanin-controlled LH releasend open up the possibility for the existence of LHRH-odulated galanin neuronal activity. The juxtapositionsetween galanin-IR elements, similarly to those existingetween LHRH-IR structures, indicate that galanin andHRH may regulate their own release by a feedbackechanism. Moreover, the dense network of galanin-IRxon varicosities in the infundibulum/median eminencend medial preoptic area raises the possibility that galaninay control other diencephalic functions as well.
cknowledgements—The authors thank Professor Thackery S.ray (Department of Cell Biology, Neurobiology and Anatomy,oyola University Chicago) for access to the microscope, Dr.saba Bohata for the computer work, and Ms. Andrea Lakatosnd Ms. Gabriella Papp for their expert technical assistance (Al-ert Szent-Gyorgyi University, Szeged, Hungary).
REFERENCES
rvat E, Gianotti L, Ramunni J, Grottoli S, Brossa PC, Bertagna A,Camanni F, Ghigo E (1995) Effect of galanin on basal and stimu-lated secretion of prolactin, gonadotropins, thyrotropin, adrenocor-ticotropin and cortisol in humans. Eur J Endocrinol 133:300–304.
ennet WM, Hill SF, Ghatei MA, Bloom SR (1991) Galanin in thenormal human pituitary and brain and in pituitary adenomas. JEndocrinol 130:463–467.
raak H, Braak E (1987) The hypothalamus of the human adult:chiasmatic region. Anat Embryol 175:315–330.
ranchek TA, Smith KE, Gerald C, Walker MW (2000) Galanin recep-
tor subtypes. Trends Pharmacol Sci 21:109–117.
CD
D
D
D
D
D
F
F
G
G
G
G
G
H
H
K
K
L
L
M
M
M
M
M
M
M
M
M
M
P
R
R
R
R
S
S
S
S
S
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707706
rawley JN (1995) Biological actions of galanin. Regul Pept 59:1–16.udas B, Merchenthaler I (2001) Catecholaminergic axons innervate
luteinizing hormone-releasing hormone immunoreactive neurons ofthe human diencephalon. J Clin Endocrinol Metab 86:5620–5626.
udas B, Merchenthaler I (2002a) Close juxtapositions betweenLHRH immunoreactive neurons and substance P immunoreactiveaxons in the human diencephalon. J Clin Endocrinol Metab 87:2946–2953.
udas B, Merchenthaler I (2002b) Close juxtapositions between lu-teinizing hormone-releasing hormone-immunoreactive neuronsand corticotropin-releasing factor-immunoreactive axons in the hu-man diencephalon. J Clin Endocrinol Metab 87:5778–5784.
udas B, Merchenthaler I (2003) Topography and associations ofleu-enkephalin and luteinizing hormone-releasing hormone neuro-nal systems in the human diencephalon. J Clin Endocrinol Metab88:1842–1848.
udas B, Mihaly A, Merchenthaler I (2000) Topography and associa-tions of luteinizing hormone-releasing hormone and neuropeptideY-immunoreactive neuronal systems in the human diencephalon.J Comp Neurol 427:593–603.
ufourny L, Schofield N, Skinner DC (2003) Immunoreactive galaninexpression in ovine gonadotropin-releasing hormone neurones: noeffects of gender or reproductive status. J Neuroendocrinol 15:1062–1069.
inn PD, Pau KY, Spies HG, Cunningham MJ, Clifton DK, Steiner RA(2000) Galanin’s functional significance in the regulation of theneuroendocrine reproductive axis of the monkey. Neuroendocrinol-ogy 71:16–26.
loren A, Land T, Langel U (2000) Galanin receptor subtypes andligand binding. Neuropeptides 34:331–337.
ai WP, Geffen LB, Blessing WW (1990) Galanin immunoreactive neu-rons in the human hypothalamus: colocalization with vasopressin-containing neurons. J Comp Neurol 298:265–280.
allyas F, Gorcs T, Merchenthaler I (1982) High-grade intensificationof the end-product of the diaminobenzidine reaction for peroxidasehistochemistry. J Histochem Cytochem 30:183–184.
allyas F, Merchenthaler I (1988) Copper-H2O2 oxidation strikinglyimproves silver intensification of the nickel-diaminobenzidine (Ni-DAB) end-product of the peroxidase reaction. J Histochem Cyto-chem 36:807–810.
iustina A, Licini M, Schettino M, Doga M, Pizzocolo G, Negro-Vilar A(1994) Physiological role of galanin in the regulation of anteriorpituitary function in humans. Am J Physiol 266:E57–E61.
undlah C, Lu NZ, Bethea CL (2002) Ovarian steroid regulation ofmonoamine oxidase-A and -B mRNAs in the macaque dorsal rapheand hypothalamic nuclei. Psychopharmacology (Berl) 160:271–282.
erbison AE (1998) Multimodal influence of estrogen upon gonadotropin-releasing hormone neurons. Endocr Rev 19:302–330.
okfelt T, Xu ZQ, Shi TJ, Holmberg K, Zhang X (1998) Galanin inascending systems: focus on coexistence with 5-hydroxytryptamineand noradrenaline. Ann NY Acad Sci 863:252–263.
aplan LM, Gabriel SM, Koenig JI, Sunday ME, Spindel ER, Martin JB,Chin WW (1988) Galanin is an estrogen-inducible, secretory prod-uct of the rat anterior pituitary. Proc Natl Acad Sci USA 85:7408–7412.
ing JC, Anthony EL, Fitzgerald DM, Stopa EG (1985) Luteinizinghormone-releasing hormone neurons in human preoptic/hypothalamus: differential intraneuronal localization of immunore-active forms. J Clin Endocrinol Metab 60:88–97.
iu HX, Brumovsky P, Schmidt R, Brown W, Payza K, Hodzic L, PouC, Godbout C, Hokfelt T (2001) Receptor subtype-specific prono-ciceptive and analgesic actions of galanin in the spinal cord: selec-tive actions via GalR1 and GalR2 receptors. Proc Natl Acad SciUSA 98:9960–9964.
opez FJ, Merchenthaler I, Ching M, Wisniewski MG, Negro-Vilar A
(1991) Galanin: a hypothalamic-hypophysiotropic hormone modu-lating reproductive functions. Proc Natl Acad Sci USA 88:4508–4512.
elander T, Hokfelt T, Nilsson S, Brodin E (1986a) Visualization ofgalanin binding sites in the rat central nervous system. Eur J Phar-macol 124:381–382.
elander T, Hokfelt T, Rokaeus A (1986b) Distribution of galaninlikeimmunoreactivity in the rat central nervous system. J Comp Neurol248:475–517.
erchenthaler I (1991a) The hypophysiotropic galanin system of therat brain. Neuroscience 44:643–654.
erchenthaler I (1991b) Neurons with access to the general circula-tion in the central nervous system of the rat: a retrograde tracingstudy with fluoro-gold. Neuroscience 44:655–662.
erchenthaler I (1998) LHRH and sexual dimorphism. Ann NY AcadSci 863:175–187.
erchenthaler I, Gorcs T, Setalo G, Petrusz P, Flerko B (1984)Gonadotropin-releasing hormone (GnRH) neurons and path-ways in the rat brain. Cell Tissue Res 237:15–29.
erchenthaler I, Lopez FJ, Lennard DE, Negro-Vilar A (1991) Sexualdifferences in the distribution of neurons coexpressing galanin andluteinizing hormone-releasing hormone in the rat brain. Endocrinol-ogy 129:1977–1986.
erchenthaler I, Lopez FJ, Negro-Vilar A (1990) Colocalization ofgalanin and luteinizing hormone-releasing hormone in a subset ofpreoptic hypothalamic neurons: anatomical and functional corre-lates. Proc Natl Acad Sci USA 87:6326–6330.
erchenthaler I, Lopez FJ, Negro-Vilar A (1993) Anatomy and phys-iology of central galanin-containing pathways. Prog Neurobiol 40:711–769.
itchell V, Bouret S, Prevot V, Jennes L, Beauvillain JC (1999) Evi-dence for expression of galanin receptor Gal-R1 mRNA in certaingonadotropin releasing hormone neurones of the rostral preopticarea. J Neuroendocrinol 11:805–812.
au KY, Spies HG (1997) Neuroendocrine signals in the regulation ofgonadotropin-releasing hormone secretion. Chin J Physiol 40:181–196.
ajendren G, Gibson MJ (1999) Expression of galanin immunoreac-tivity in gonadotropin-releasing hormone neurons in mice: a confo-cal microscopic study. Brain Res 821:270–276.
ajendren G, Gibson MJ (2001) Increased synaptic input to gonado-tropin releasing hormone cells in preoptic area grafts that supportreproductive development in female hypogonadal mice. J Neuroen-docrinol 13:270–274.
ajendren G, Li X (2001) Galanin synaptic input to gonadotropin-releasing hormone perikarya in juvenile and adult female mice:implications for sexual maturity. Brain Res Dev Brain Res 131:161–165.
ance NE, Young WS 3rd, McMullen NT (1994) Topography of neu-rons expressing luteinizing hormone-releasing hormone gene tran-scripts in the human hypothalamus and basal forebrain. J CompNeurol 339:573–586.
ahu A, Xu B, Kalra SP (1994) Role of galanin in stimulation ofpituitary luteinizing hormone secretion as revealed by a specificreceptor antagonist, galantide. Endocrinology 134:529–536.
aper C (1990) Hypothalamus. In: The human nervous system (Paxi-nos G, ed), pp 389–413. San Diego: Academic Press.
ervin AL, Amiranoff B, Rouyer-Fessard C, Tatemoto K, LaburtheM (1987) Identification and molecular characterization of galaninreceptor sites in rat brain. Biomed Biophys Res Com 144:298 –306.
ilverman AJ, Livne I, Witkin JW (1994) The gonadotropin-releas-ing hormone (GnRH) neuronal systems: immunocytochemistryand in situ hybridization. In: The physiology of reproduction(Knobil ENJ, ed), pp 1683–1709. New York: Raven Press.
kofitsch G, Jacobowitz DM (1985) Immunohistochemical mapping ofgalanin-like neurons in the rat central nervous system. Peptides
6:509–546.
S
S
T
W
Z
B. Dudas and I. Merchenthaler / Neuroscience 127 (2004) 695–707 707
teiner RA, Hohmann JG, Holmes A, Wrenn CC, Cadd G, Jureus A,Clifton DK, Luo M, Gutshall M, Ma SY, Mufson EJ, Crawley JN(2001) Galanin transgenic mice display cognitive and neurochem-ical deficits characteristic of Alzheimer’s disease. Proc Natl AcadSci USA 98:4184–4189.
topa EG, Koh ET, Svendsen CN, Rogers WT, Schwaber JS, King JC(1991) Computer-assisted mapping of immunoreactive mammaliangonadotropin-releasing hormone in adult human basal forebrainand amygdala. Endocrinology 128:3199–3207.
atemoto K, Rokaeus A, Jornvall H, McDonald TJ, Mutt V (1983)Galanin: a novel biologically active peptide from porcine intestine.FEBS Lett 164:124–128.
ynick D, Thompson SW, McMahon SB (2001) The role of galanin asa multi-functional neuropeptide in the nervous system. Curr OpinPharmacol 1:73–77.
drojewicz Z, Sowinska E (2000) The significance of galanin in phys-iologic and pathologic processes in humans. Postepy Hig MedDosw 54:819–833.
(Accepted 5 May 2004)(Available online 17 July 2004)





























