Embed Size (px)
Citation preview

Ovulation Involves the Luteinizing Hormone-Dependent Activation of Gq/11 in Granulosa Cells
Shawn M. Breen, Nebojsa Andric, Tai Ping, Fang Xie, Stefan Offermans,Jan A. Gossen, and Mario Ascoli
Department of Pharmacology (S.M.B., N.A., T.P., M.A.), Carver College of Medicine, University of Iowa,Iowa City, Iowa 52242; Center for Reproductive Sciences (F.X.), University of California, San Francisco,San Francisco, California 94143; Department of Pharmacology (S.O.), Max Planck Institute for Heart andLung Research, 61231 Bad Nauheim, Germany; Women’s Health Department (J.A.G.), Merck Sharp andDohme Research Laboratories, 5340 BH Oss, The Netherlands
The LH receptor (LHR) activates several families of heterotrimeric G proteins, but only the activa-tion of Gs and subsequent generation of cAMP are universally accepted as important mediators ofLH actions. To examine the involvement of the Gq/11 family on the actions of LH, we crossedCyp19Cre and G�q
f/f;G�11�/� mice to generate mice with a granulosa cell-specific deletion of G�q
in the context of a global deletion of G�11. Granulosa cells from G�qf/f;G�11
�/�;Cre� mice havebarely detectable levels of G�q/11, have a normal complement of LHR, and respond to LHR acti-vation with a transient increase in cAMP accumulation, but they fail to respond with increasedinositol phosphate accumulation, an index of the activation of G�q/11. The LHR-provoked resump-tion of meiosis, cumulus expansion, and luteinization are normal. However, the G�q
f/f;G�11�/�;
Cre� mice display severe subfertility because many of the oocytes destined for ovulation becomeentrapped in preovulatory follicles or corpora lutea. Because follicular rupture is known to bedependent on the expression of the progesterone receptor (Pgr), we examined the LHR-inducedexpression of Pgr and 4 of its target genes (Adamts-1, Ctsl1, Edn2, and Prkg2). These actions of theLHR were impaired in the ovaries of the G�q
f/f;G�11�/�;Cre� mice. We conclude that the defect in
follicular rupture is secondary to the failure of the LHR to fully induce the expression of the Pgr. Thisis the first conclusive evidence for the physiological importance of the activation of Gq/11 by the LHRand for the involvement of G�q/11 in ovulation. (Molecular Endocrinology 27: 1483–1491, 2013)
Ovulation is a complex process that can be subdividedinto at least 3 sequential steps: the resumption of
meiosis of the oocyte, the expansion of cumulus granu-losa cells, and finally the rupture of the follicular wall thatallows for the release of cumulus/oocyte complexes.These events are triggered by the binding of LH to the LHreceptor (LHR) in mural granulosa cells, a signal that ispropagated to LHR-negative cumulus cells in a paracrinefashion through the epidermal growth factor (EGF) net-work and by gap junction-mediated diffusion of intracel-lular cGMP to LHR-negative oocytes.
Oocytes are maintained in meiotic arrest by the highlevel of intraoocyte cAMP generated by endogenous Gs-
coupled receptors (1, 2), but a cGMP-mediated inhibitionof phosphodiesterase 3A is essential for the maintenanceof high intraoocyte cAMP and thus meiotic arrest. Therequired cGMP is derived from intracellular cGMP gen-erated in the somatic cells that diffuses into the oocytethrough gap junctions. The generation of cGMP in thesomatic compartment is in turn controlled by the para-crine actions of atrial natriuretic peptides generated inmural granulosa cells that engage the atrial natriureticpeptide receptors present in mural and cumulus cells(3–5).
The binding of LH to the LHR in mural granulosa cellsactivates Gs and increases cAMP accumulation (6, 7). An
ISSN Print 0888-8809 ISSN Online 1944-9917Printed in U.S.A.Copyright © 2013 by The Endocrine SocietyReceived May 3, 2013. Accepted July 3, 2013.First Published Online July 8, 2013
Abbreviations: ADAMTS-1, A disintegrin and metalloproteinase with thrombospondin-like repeats 1; EGF, epidermal growth factor; EGFR, EGF receptor; GAPDH, glyceralde-hyde-3-phosphate dehydrogenase; hCG, human chorionic gonadotropin; LHR, LH recep-tor; P, phosphorylated; PGR, progesterone receptor; PMSG, pregnant mare serumgonadotropin; qPCR, quantitative PCR; StAR, steroidogenic acute regulatory protein.
O R I G I N A L R E S E A R C H
doi: 10.1210/me.2013-1130 Mol Endocrinol, September 2013, 27(9):1483–1491 mend.endojournals.org 1483
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022

important consequence of the activation of this signalingpathway is the increased expression of 3 EGF-like growthfactors (amphiregulin, epiregulin, and betacellulin) thatare released in a soluble form and transactivate the EGFreceptor (EGFR) in mural and cumulus cells (6). Thisresults in the activation of the ERK1/2 cascade, a processthat is essential for the expression of several genes, includ-ing Has2, Ptgs2, and Tnfaip6, which are expressed inmural and cumulus cells and are necessary for cumulusexpansion (8, 9). Acting through the EGF network (butperhaps not exclusively), the LHR also decreases cGMPlevels in mural and cumulus granulosa cells, and anERK1/2-dependent closure of gap junctions between mu-ral and cumulus cells ultimately lowers cGMP levels in thesomatic compartment and in the oocyte. The decrease inintraoocyte cGMP activates phosphodiesterase 3A andreduces intraoocyte cAMP levels, thus allowing meiosis toresume (3, 4, 10–12).
The rupture of the follicular wall, the last step in theprocess of ovulation, is also an LH-dependent event, andit requires increased expression and activation of the pro-gesterone receptor (PGR) in mural granulosa cells (13–15). The activated PGR in turn enhances the expression ofa complex gene network including other nuclear recep-tors, transcription factors, and extracellular proteasesthat degrade the follicular wall (14, 15). Pgr null mice areanovulatory because of a defect in follicular rupture (16).Some of the ovulatory actions of the PGR are mediated byA disintegrin and metalloproteinase with thrombospon-din-like repeats 1 (ADAMTS-1), an extracellular proteasesynthesized and secreted by mural granulosa cells in re-sponse to PGR target activation (17), because Adamts-1null mice have a partial defect in follicular rupture (18).
Despite the importance of follicular rupture to the ovu-latory response, the second-messenger pathways that me-diate the LHR-dependent increase in the expression of thePGR have not been defined. The most relevant informa-tion on this pathway comes from studies done in primarycultures of granulosa cell showing that optimal inductionof Pgr expression can be achieved only by simultaneouspharmacological activation of the protein kinase A andprotein kinase C pathways (19). This is an interestingfinding because, in addition to activating adenylyl cy-clase, engagement of the LHR in granulosa cells (20–23)or in heterologous cells expressing the recombinant LHR(24–26) also results in the activation of phospholipase Cand the generation of inositol phosphates and diacylglyc-erol, which activate protein kinase C. These findings sug-gest that induction of the Pgr and ovulation involve theLHR-dependent activation of G protein families distinctfrom Gs.
To investigate the role of the Gq/11 family of G pro-teins on ovulation and female fertility, we used a Cre/LoxP approach to generate mice with a granulosa cell-specific deletion of G�q in the context of a global deletionof G�11. The reproductive phenotype of G�q
f/f;G�11�/�;
Cre� mice resembles that of the Pgr�/� (16) or Ad-amts1�/� mice (18) in which many of the oocytes destinedfor ovulation become entrapped in preovulatory folliclesor corpora lutea. Moreover, the LHR-induced expressionof Pgr and 4 of its target genes (Adamts-1, Ctsl1, Edn2,and Prkg2) is reduced in the ovaries of the G�q
f/f;G�11�/�;
Cre� mice.
Materials and Methods
MiceG�q
f/f;G�q�/� (27–29) and Cyp19Cre transgenic mice (9,
30) are in a C57BL/6 background. A colony of G�qf/f;G�q
�/�;Cre� mice was generated and subsequently maintained by cross-ing G�q
f/f;G�q�/�;Cre� females with G�q
f/f;G�q�/�;Cre�
males. The resulting G�qf/f;G�q
�/�;Cre� and G�qf/f;G�q
�/�;Cre� females were used as experimental and control animals,respectively. Genotyping was done using tail genomic DNA andPCR amplification as described (9, 27–30). The segregation ofthe Cre allele in the female pups closely approximated Mende-lian inheritance (47% Cre� and 53% Cre�).
Most experiments were done using immature female mice(21–24 days old) that had been injected ip with 5 IU pregnantmare serum gonadotropin (PMSG; National Hormone and Pep-tide Program) or injected with 5 IU PMSG followed by an ipinjection of an ovulatory dose (5 IU) of human chorionic gonad-otropin (hCG; Sigma Chemicals). Ovaries were collected at dif-ferent times after the injection of hCG as indicated in the figurelegends.
For the experiments using primary cultures of differentiatedgranulosa cells, the mice were injected sc with 4.5 �g of dieth-ylstilbestrol (Sigma Chemicals) and 15 hours later with 5 IU ofPMSG. Differentiated granulosa cells were isolated 44–48hours after the injection of PMSG. All animal procedures wereapproved by the Institutional Animal Care and Use Committeefor the University of Iowa.
Granulosa cell culturesDifferentiated granulosa cells were isolated (31) and plated
in 24-well plates precoated with bovine fibronectin in medium(DMEM/F12 with 10 mM HEPES and 50 �g/mL gentamicin)containing 1 mg/mL BSA (23). We usually plated 3 wells (2–5 �105 cells/well) from each mouse in a total volume of 500 �L. Thecells were allowed to attach for 4–5 hours and collected imme-diately for the Western blots of G�q/11 or the medium wasreplaced. The cells were then incubated with buffer only or withhCG (1 �g/mL) prior to the collection for cAMP determinationor for Western blots of phosphorylated (P)-ERK1/2 orP-CCAAT/enhancer-binding protein/�.
For the hCG binding and inositol phosphate assays, the cellswere plated in culture medium that was also supplemented withinsulin (1 �g/mL), transferrin (1 �g/mL), and selenium (1 ng/mL)
1484 Breen et al Activation of Gq/11 Is Involved in Ovulation Mol Endocrinol, September 2013, 27(9):1483–1491
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022

as well as 15 ng/mL of T (Sigma Chemicals) and 100 ng/mL orrecombinant human FSH (EMD Serono) (23). After 24 hoursthe medium was replaced and the cells were cultured for anadditional 24 hours prior to measuring 125I-hCG binding asdescribed earlier (23). For the inositol phosphate assays, themedium was replaced with the same medium supplemented with4 �Ci/mL of 3H-myoinositol, and the cells were cultured for anadditional 24 hours. Finally, the labeling medium was aspiratedand the cells were washed and used to measure the accumulationof 3H-inositol phosphates during 1 hour incubation in mediumcontaining 20 mM LiCl without or with 1 �g/mL recombinanthCG as described earlier (23).
HistologyOvaries were fixed in Bouin’s fixative, embedded in paraffin,
sectioned, and stained with hematoxylin and eosin using stan-dard conditions. The sections were used to count the number ofentrapped oocytes or assess germinal vesicle breakdown.
Entrapped oocytes were counted in every 10th serial section(5 �m) of an entire ovary 24 hours after an injection of anovulatory dose of hCG to PMSG-primed mice. Depending onthe genotype of the mice and the structure being scored (antralfollicles or corpora lutea), this resulted in the counting of 0–30entrapped oocytes. The counting was performed by 2 differentindividuals who were blinded to the genotypes of the mice. Theaverages were used.
Germinal vesicle breakdown, a measurement of the resump-tion of meiosis, was assessed 4 hours after an injection of anovulatory dose of hCG to PMSG-primed mice using serial sec-tions of the entire ovary. It resulted in the scoring of 15–35oocytes/ovary, and it was done by a single individual who wasblinded to the genotypes of the mice.
Fertilization and embryo transferThe percent fertilization of ovulated oocytes was measured
using immature female mice that had been injected with PMSGand hCG as described above and mated with G�q
f/f;G�q�/�;
Cre� male mice of known fertility at the time of the hCG injec-tion (in the afternoon). Females with a vaginal plug the follow-ing morning were euthanized 2 days later and the uterine hornswere flushed with isotonic saline. Embryos and unfertilizedoocytes were counted and the fertilization rate was calculated(32).
For the embryo transfer experiments, mature females (80–120 days old) were mated with vasectomized males in the after-noon. Females with a vaginal plug on the next day were used 3days later as recipients of wt blastocysts. Blastocysts were ob-tained 3–4 days after the detection of a vaginal plug from wtimmature females (21–26 days old) that were superovulated(injected with 5 IU PMSG followed by 5 IU hCG 44 hours later)and mated with males of known fertility in the afternoon.Twelve to 14 healthy blastocysts were implanted in each uterinehorn and the implanted embryos were counted 3 days later (32).
Real-time PCROvarian RNA was prepared from 2 ovaries homogenized
in 2 mL of Trizol using a Polytron homogenizer (23). RNAfrom granulosa cell cultures was prepared from individualwells (see above) using the QIAGEN RNeasy minikit. Equalamounts of purified RNA were reversed transcribed using
dN6 random primers (Applied Biosystems) and Moloney mu-rine leukemia virus reverse transcriptase (Promega) as describedelsewhere (33). Real-time PCR reactions were performed in a 25�L volume using 300 nM of each primer and 1� iQ SYBRGreen Super Mix (Bio-Rad Laboratories), and fluorescence wasdetected on the CFX96 real-time PCR detection system (Bio-Rad Laboratories). The conditions for the quantitative PCR(qPCR) were optimized for each gene, and the target gene ex-pression was normalized to an internal control, glyceraldehyde-3-phosphate dehydrogenase (Gapdh). The primers used forEreg, Areg, Btc, and Gapdh were previously described (34, 35).The primers for Adamts-1, Edn2, and Prkg2 were those de-scribed by Kim et al (36), and those for Cyp19a1 and theLhcgr have been described by us (23). Those used for Ptgs2are: forward, AAAGGTTCTTCTACGGAGAGAGTTCA, re-verse, TGGGCAAAGAATGCAAACATC; Tnfaip6: forward,GCAGCTAGAGGCAGCCAGAA, reverse, ACTCTACCCT-TGGCCATCCA; and Has2: forward, GGGAACTCAGAC-GACGACCTT, and reverse, GATGTACGTGGCCGATTT-GTC. All data were analyzed using the method of Pfaffl (37).
Other methodsFor Western blots, cells or ovaries were lysed in a buffer (150
mM NaCl; 50 mM Tris; 1 mM EDTA; 1% Nonidet P-40; 0.5%sodium deoxycholate; 0.1% sodium dodecyl sulfate; 1 mMNa3VO4; and 1 mM NaF, pH 7.4) supplemented with a com-mercial mixture of protease inhibitors (Roche). Homogenateswere kept on ice for 30 minutes, with occasional mechanicaldisruption using a pipette, followed by a centrifugation at13 000 � g for 10 minutes. Supernatants were collected andassayed for protein content using a bicinchoninic assay proteinassay kit from Bio-Rad Laboratories. Cellular lysates were usedimmediately or stored at �80°C until used. Western blots weredone as described earlier (34, 38, 39) using 2–20 �g of lysateprotein. The solutions used to block and wash the membranesand perform the primary and secondary antibody incubationscontained either 5% milk or 1%–5% BSA, depending on theantigen being detected. The length (1–18 hours) and tempera-ture (4°C or ambient) of the incubation and the dilutions of theprimary antibodies also varied, depending on the antigen beingdetected. The secondary antibody dilution (1:3000) and lengthand temperature of incubation (1 hour at room temperature)were constant, however. The secondary antimouse or antirabbitantibodies coupled to horseradish peroxidase were from Bio-Rad Laboratories (catalog number 170–6515). The immunecomplexes in the Western blots were eventually visualized usingenhanced chemiluminescence (Pierce Chemical, Rockford, Illi-nois) and exposed to film. The source of antibodies was asfollows: G�q/G�11 was from Santa Cruz Biotechnology (itemnumber sc392). P-EGFR (item number 3377), P-ERK1/2 (itemnumber 9122), P-CCAAT/enhancer-binding protein/�(item number 3084), AKT (item number 9272), and GAPDH (itemnumber 2118) were from Cell Signaling, and Cyp11a1 (item num-ber AB1244) was from Chemicon. The antibodies to the precursorand mature forms of ADAMTS-1 were generously provided by DrJoAnne Richards (the Baylor College of Medicine) (17). The anti-body to steroidogenic acute regulatory protein (StAR) was kindlyprovided by Dr Doug Stocco (Texas Tech University).
Progesterone assays were done using a commercially avail-able kit from Cayman Chemicals.
doi: 10.1210/me.2013-1130 mend.endojournals.org 1485
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022
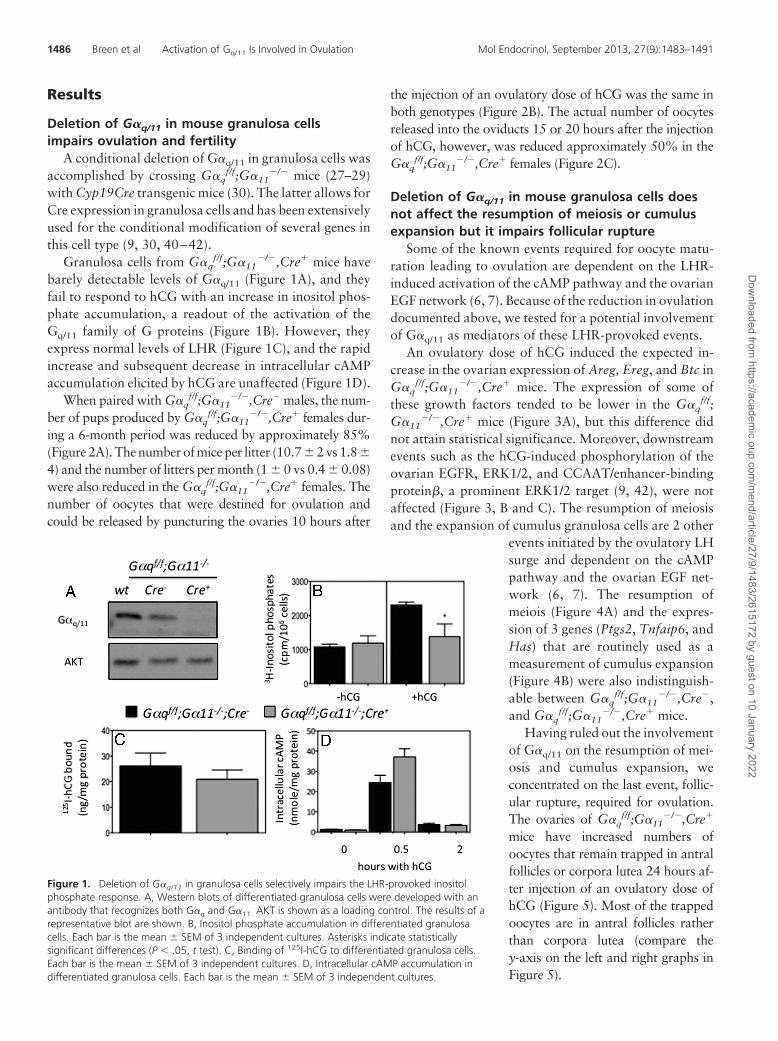
Results
Deletion of G�q/11 in mouse granulosa cellsimpairs ovulation and fertility
A conditional deletion of G�q/11 in granulosa cells wasaccomplished by crossing G�q
f/f;G�11�/� mice (27–29)
with Cyp19Cre transgenic mice (30). The latter allows forCre expression in granulosa cells and has been extensivelyused for the conditional modification of several genes inthis cell type (9, 30, 40–42).
Granulosa cells from G�qf/f;G�11
�/�,Cre� mice havebarely detectable levels of G�q/11 (Figure 1A), and theyfail to respond to hCG with an increase in inositol phos-phate accumulation, a readout of the activation of theGq/11 family of G proteins (Figure 1B). However, theyexpress normal levels of LHR (Figure 1C), and the rapidincrease and subsequent decrease in intracellular cAMPaccumulation elicited by hCG are unaffected (Figure 1D).
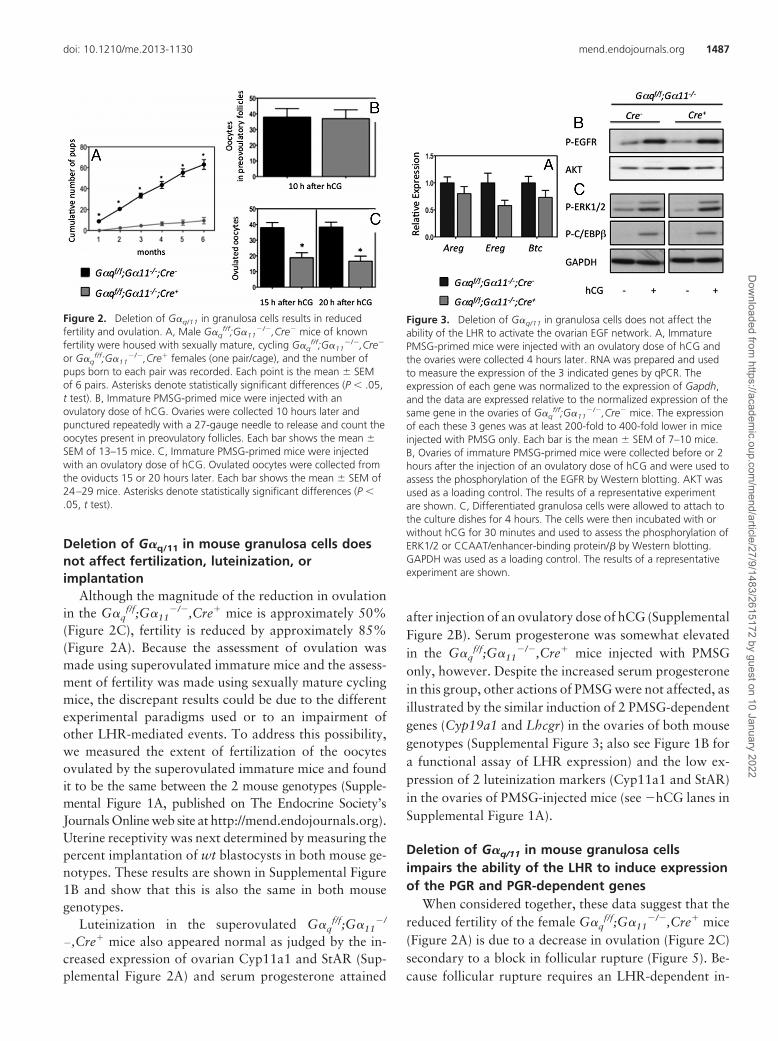
When paired with G�qf/f;G�11
�/�,Cre� males, the num-ber of pups produced by G�q
f/f;G�11�/�,Cre� females dur-
ing a 6-month period was reduced by approximately 85%(Figure 2A). The number of mice per litter (10.7 � 2 vs 1.8 �4) and the number of litters per month (1 � 0 vs 0.4 � 0.08)were also reduced in the G�q
f/f;G�11�/�,Cre� females. The
number of oocytes that were destined for ovulation andcould be released by puncturing the ovaries 10 hours after
the injection of an ovulatory dose of hCG was the same inboth genotypes (Figure 2B). The actual number of oocytesreleased into the oviducts 15 or 20 hours after the injectionof hCG, however, was reduced approximately 50% in theG�q
f/f;G�11�/�,Cre� females (Figure 2C).
Deletion of G�q/11 in mouse granulosa cells doesnot affect the resumption of meiosis or cumulusexpansion but it impairs follicular rupture
Some of the known events required for oocyte matu-ration leading to ovulation are dependent on the LHR-induced activation of the cAMP pathway and the ovarianEGF network (6, 7). Because of the reduction in ovulationdocumented above, we tested for a potential involvementof G�q/11 as mediators of these LHR-provoked events.
An ovulatory dose of hCG induced the expected in-crease in the ovarian expression of Areg, Ereg, and Btc inG�q
f/f;G�11�/�,Cre� mice. The expression of some of
these growth factors tended to be lower in the G�qf/f;
G�11�/�,Cre� mice (Figure 3A), but this difference did
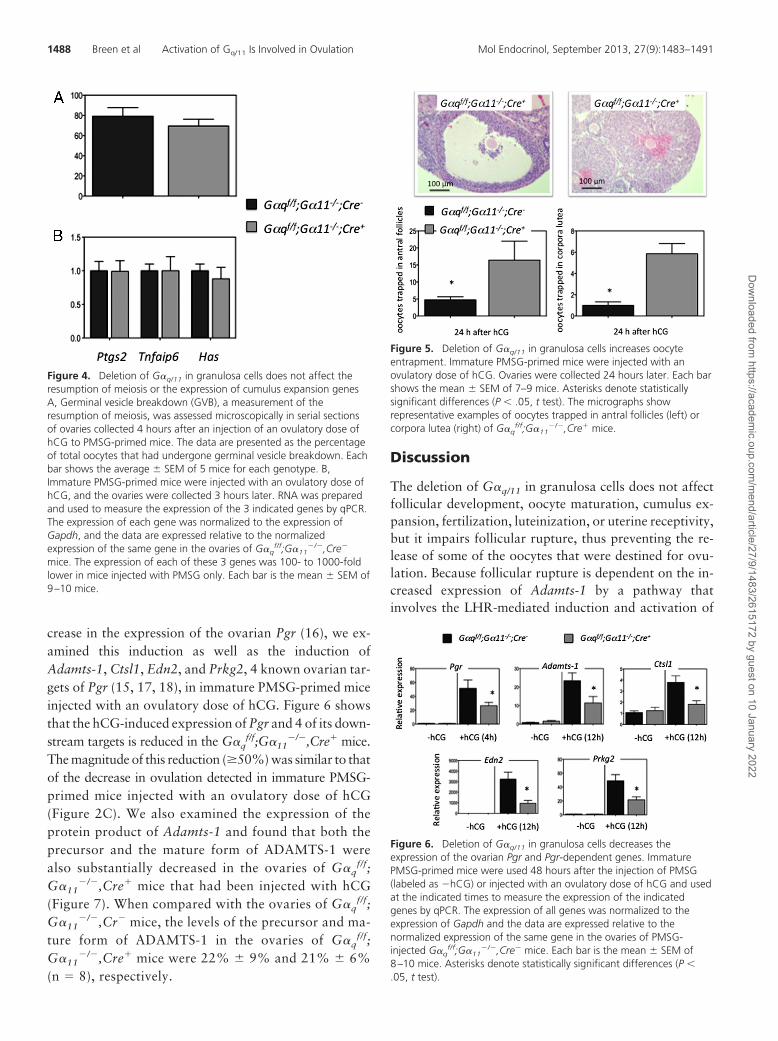
not attain statistical significance. Moreover, downstreamevents such as the hCG-induced phosphorylation of theovarian EGFR, ERK1/2, and CCAAT/enhancer-bindingprotein�, a prominent ERK1/2 target (9, 42), were notaffected (Figure 3, B and C). The resumption of meiosisand the expansion of cumulus granulosa cells are 2 other
events initiated by the ovulatory LHsurge and dependent on the cAMPpathway and the ovarian EGF net-work (6, 7). The resumption ofmeiois (Figure 4A) and the expres-sion of 3 genes (Ptgs2, Tnfaip6, andHas) that are routinely used as ameasurement of cumulus expansion(Figure 4B) were also indistinguish-able between G�q
f/f;G�11�/�,Cre�,
and G�qf/f;G�11
�/�,Cre� mice.Having ruled out the involvement
of G�q/11 on the resumption of mei-osis and cumulus expansion, weconcentrated on the last event, follic-ular rupture, required for ovulation.The ovaries of G�q
f/f;G�11�/�,Cre�
mice have increased numbers ofoocytes that remain trapped in antralfollicles or corpora lutea 24 hours af-ter injection of an ovulatory dose ofhCG (Figure 5). Most of the trappedoocytes are in antral follicles ratherthan corpora lutea (compare they-axis on the left and right graphs inFigure 5).
Figure 1. Deletion of G�q/11 in granulosa cells selectively impairs the LHR-provoked inositolphosphate response. A, Western blots of differentiated granulosa cells were developed with anantibody that recognizes both G�q and G�11. AKT is shown as a loading control. The results of arepresentative blot are shown. B, Inositol phosphate accumulation in differentiated granulosacells. Each bar is the mean � SEM of 3 independent cultures. Asterisks indicate statisticallysignificant differences (P � .05, t test). C, Binding of 125I-hCG to differentiated granulosa cells.Each bar is the mean � SEM of 3 independent cultures. D, Intracellular cAMP accumulation indifferentiated granulosa cells. Each bar is the mean � SEM of 3 independent cultures.
1486 Breen et al Activation of Gq/11 Is Involved in Ovulation Mol Endocrinol, September 2013, 27(9):1483–1491
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022
Deletion of G�q/11 in mouse granulosa cells doesnot affect fertilization, luteinization, orimplantation
Although the magnitude of the reduction in ovulationin the G�q
f/f;G�11�/�,Cre� mice is approximately 50%
(Figure 2C), fertility is reduced by approximately 85%(Figure 2A). Because the assessment of ovulation wasmade using superovulated immature mice and the assess-ment of fertility was made using sexually mature cyclingmice, the discrepant results could be due to the differentexperimental paradigms used or to an impairment ofother LHR-mediated events. To address this possibility,we measured the extent of fertilization of the oocytesovulated by the superovulated immature mice and foundit to be the same between the 2 mouse genotypes (Supple-mental Figure 1A, published on The Endocrine Society’sJournals Online web site at http://mend.endojournals.org).Uterine receptivity was next determined by measuring thepercent implantation of wt blastocysts in both mouse ge-notypes. These results are shown in Supplemental Figure1B and show that this is also the same in both mousegenotypes.
Luteinization in the superovulated G�qf/f;G�11
�/
�,Cre� mice also appeared normal as judged by the in-creased expression of ovarian Cyp11a1 and StAR (Sup-plemental Figure 2A) and serum progesterone attained
after injection of an ovulatory dose of hCG (SupplementalFigure 2B). Serum progesterone was somewhat elevatedin the G�q
f/f;G�11�/�,Cre� mice injected with PMSG
only, however. Despite the increased serum progesteronein this group, other actions of PMSG were not affected, asillustrated by the similar induction of 2 PMSG-dependentgenes (Cyp19a1 and Lhcgr) in the ovaries of both mousegenotypes (Supplemental Figure 3; also see Figure 1B fora functional assay of LHR expression) and the low ex-pression of 2 luteinization markers (Cyp11a1 and StAR)in the ovaries of PMSG-injected mice (see �hCG lanes inSupplemental Figure 1A).
Deletion of G�q/11 in mouse granulosa cellsimpairs the ability of the LHR to induce expressionof the PGR and PGR-dependent genes
When considered together, these data suggest that thereduced fertility of the female G�q
f/f;G�11�/�,Cre� mice
(Figure 2A) is due to a decrease in ovulation (Figure 2C)secondary to a block in follicular rupture (Figure 5). Be-cause follicular rupture requires an LHR-dependent in-
Figure 2. Deletion of G�q/11 in granulosa cells results in reducedfertility and ovulation. A, Male G�q
f/f;G�11�/�,Cre� mice of known
fertility were housed with sexually mature, cycling G�qf/f;G�11
�/�,Cre�
or G�qf/f;G�11
�/�,Cre� females (one pair/cage), and the number ofpups born to each pair was recorded. Each point is the mean � SEMof 6 pairs. Asterisks denote statistically significant differences (P � .05,t test). B, Immature PMSG-primed mice were injected with anovulatory dose of hCG. Ovaries were collected 10 hours later andpunctured repeatedly with a 27-gauge needle to release and count theoocytes present in preovulatory follicles. Each bar shows the mean �SEM of 13–15 mice. C, Immature PMSG-primed mice were injectedwith an ovulatory dose of hCG. Ovulated oocytes were collected fromthe oviducts 15 or 20 hours later. Each bar shows the mean � SEM of24–29 mice. Asterisks denote statistically significant differences (P �.05, t test).
Figure 3. Deletion of G�q/11 in granulosa cells does not affect theability of the LHR to activate the ovarian EGF network. A, ImmaturePMSG-primed mice were injected with an ovulatory dose of hCG andthe ovaries were collected 4 hours later. RNA was prepared and usedto measure the expression of the 3 indicated genes by qPCR. Theexpression of each gene was normalized to the expression of Gapdh,and the data are expressed relative to the normalized expression of thesame gene in the ovaries of G�q
f/f;G�11�/�,Cre� mice. The expression
of each these 3 genes was at least 200-fold to 400-fold lower in miceinjected with PMSG only. Each bar is the mean � SEM of 7–10 mice.B, Ovaries of immature PMSG-primed mice were collected before or 2hours after the injection of an ovulatory dose of hCG and were used toassess the phosphorylation of the EGFR by Western blotting. AKT wasused as a loading control. The results of a representative experimentare shown. C, Differentiated granulosa cells were allowed to attach tothe culture dishes for 4 hours. The cells were then incubated with orwithout hCG for 30 minutes and used to assess the phosphorylation ofERK1/2 or CCAAT/enhancer-binding protein/� by Western blotting.GAPDH was used as a loading control. The results of a representativeexperiment are shown.
doi: 10.1210/me.2013-1130 mend.endojournals.org 1487
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022
crease in the expression of the ovarian Pgr (16), we ex-amined this induction as well as the induction ofAdamts-1, Ctsl1, Edn2, and Prkg2, 4 known ovarian tar-gets of Pgr (15, 17, 18), in immature PMSG-primed miceinjected with an ovulatory dose of hCG. Figure 6 showsthat the hCG-induced expression of Pgr and 4 of its down-stream targets is reduced in the G�q
f/f;G�11�/�,Cre� mice.
The magnitude of this reduction (�50%) was similar to thatof the decrease in ovulation detected in immature PMSG-primed mice injected with an ovulatory dose of hCG(Figure 2C). We also examined the expression of theprotein product of Adamts-1 and found that both theprecursor and the mature form of ADAMTS-1 werealso substantially decreased in the ovaries of G�q
f/f;G�11
�/�,Cre� mice that had been injected with hCG(Figure 7). When compared with the ovaries of G�q
f/f;G�11
�/�,Cr� mice, the levels of the precursor and ma-ture form of ADAMTS-1 in the ovaries of G�q
f/f;G�11
�/�,Cre� mice were 22% � 9% and 21% � 6%(n � 8), respectively.
Discussion
The deletion of G�q/11 in granulosa cells does not affectfollicular development, oocyte maturation, cumulus ex-pansion, fertilization, luteinization, or uterine receptivity,but it impairs follicular rupture, thus preventing the re-lease of some of the oocytes that were destined for ovu-lation. Because follicular rupture is dependent on the in-creased expression of Adamts-1 by a pathway thatinvolves the LHR-mediated induction and activation of
Figure 4. Deletion of G�q/11 in granulosa cells does not affect theresumption of meiosis or the expression of cumulus expansion genesA, Germinal vesicle breakdown (GVB), a measurement of theresumption of meiosis, was assessed microscopically in serial sectionsof ovaries collected 4 hours after an injection of an ovulatory dose ofhCG to PMSG-primed mice. The data are presented as the percentageof total oocytes that had undergone germinal vesicle breakdown. Eachbar shows the average � SEM of 5 mice for each genotype. B,Immature PMSG-primed mice were injected with an ovulatory dose ofhCG, and the ovaries were collected 3 hours later. RNA was preparedand used to measure the expression of the 3 indicated genes by qPCR.The expression of each gene was normalized to the expression ofGapdh, and the data are expressed relative to the normalizedexpression of the same gene in the ovaries of G�q
f/f;G�11�/�,Cre�
mice. The expression of each of these 3 genes was 100- to 1000-foldlower in mice injected with PMSG only. Each bar is the mean � SEM of9–10 mice.
Figure 5. Deletion of G�q/11 in granulosa cells increases oocyteentrapment. Immature PMSG-primed mice were injected with anovulatory dose of hCG. Ovaries were collected 24 hours later. Each barshows the mean � SEM of 7–9 mice. Asterisks denote statisticallysignificant differences (P � .05, t test). The micrographs showrepresentative examples of oocytes trapped in antral follicles (left) orcorpora lutea (right) of G�q
f/f;G�11�/�,Cre� mice.
Figure 6. Deletion of G�q/11 in granulosa cells decreases theexpression of the ovarian Pgr and Pgr-dependent genes. ImmaturePMSG-primed mice were used 48 hours after the injection of PMSG(labeled as �hCG) or injected with an ovulatory dose of hCG and usedat the indicated times to measure the expression of the indicatedgenes by qPCR. The expression of all genes was normalized to theexpression of Gapdh and the data are expressed relative to thenormalized expression of the same gene in the ovaries of PMSG-injected G�q
f/f;G�11�/�,Cre� mice. Each bar is the mean � SEM of
8–10 mice. Asterisks denote statistically significant differences (P �.05, t test).
1488 Breen et al Activation of Gq/11 Is Involved in Ovulation Mol Endocrinol, September 2013, 27(9):1483–1491
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022

the ovarian PGR (13–18, 43), we examined the ability ofthe LHR to activate this pathway. The increased expres-sion of the Pgr and several of its downstream targets(Adamts-1, Ctsl1, Edn2, and Prkg2) induced by an ovu-latory dose of hCG was impaired in the ovaries of G�q
f/
f;G�11�/�,Cre� mice. Progesterone levels after an injec-
tion of hCG were normal, however. Based on these data
and the similarity of the reproductive phenotype of femaleG�q
f/f;G�11�/�,Cre� (this paper), Pgr�/� (16), and Ad-
amts-1�/� (18) mice, we conclude that the LHR-mediatedactivation of G�q/11 in granulosa cells is involved in fol-licular rupture because this G protein family is a mediatorof the LHR-provoked induction of the granulosa cell Pgr.
We also note that the reduction in the precursorand mature forms of ADAMTS-1 is more pronouncedthan the reduction in Pgr or Adamts-1 expression, thusraising the possibility that Gq/11 may also affect the sta-bility of ADAMTS-1. ADAMTS-1, however, is not theonly Pgr target that mediates follicular rupture becauseAdamts-1�/� mice display only a partial defect in ovula-tion (18). Cathepsin L, another extracellular protease, isalso a Pgr target (16), but its involvement in follicularrupture has not been studied in detail. Like Adamts-1,however, ovarian Ctsl1 expression is also reduced in theG�q
f/f;G�11�/�,Cre� mice. Another Pgr target, Edn2
(and one of its downstream targets, Prkg2) are also de-creased in the ovaries of G�q
f/f;G�11�/�;Cre� mice, but
the involvement of Edn2 on follicular rupture and ovula-tion is unresolved (43–45).
When compared with the anovulatory phenotype ofthe Pgr�/� mice (16), the partial decrease in ovulation andinduction of the Pgr in the G�q
f/f;G�11�/�,Cre� mice sug-
gests that G�q/11 is not the only family of G proteins thatmediates the effects of the LHR on Pgr induction. Thelatter conclusion is supported by previous studies on theinduction of the Pgr or a Pgr reporter gene in granulosacell cultures by second-messenger analogs (19). Additionof phorbol-12-myristate-13-acetate has little or no effecton Pgr expression, whereas the addition of forskolin hasa substantial stimulatory effect. The addition of forskolinand phorbol-12-myristate-13-acetate, however, actedsynergistically to induce Pgr expression (19). Thus, a par-tial reduction in the induction of the ovarian Pgr andovulation in the G�q
f/f;G�11�/�,Cre� mice injected with
an ovulatory dose of hCG would be expected because theGs/adenylyl cyclase/protein kinase A pathway is intact inthese mice. Full induction of the Pgr and normal ovula-tion would not be possible, however, because of the ge-netic ablation of G�q/11 (Figure 8). G�q/11-dependentpathways could also impact ovulation in a Pgr-indepen-dent fashion by directly modulating gene transcription,protein stability, or posttranslational modifications ofother proteins that participate in follicular rupture (Fig-ure 8). Such an effect has already been demonstrated forthe expression of Adamts-1 that is coordinately regulatedby the LHR in Pgr-dependent and -independent fashions(46). Our data support the possibility that G�q/11 alsoimpacts the expression of ADAMTS-1 at a posttranscrip-tional level because the reduction in the expression Ad-
Figure 7. Deletion of G�q/11 in granulosa cells decreases theexpression of ovarian ADAMTS-1. Immature PMSG-primed mice wereused 48hours after the injection of PMSG (labeled �hCG) or injectedwith an ovulatory dose of hCG and used 12 hours later. Western blotswere developed with antibodies that recognize the precursor (Pro-ADAMTS-1) or the mature (MP-ADAMTS-1) forms of ADAMTS-1. Theresults of a representative experiment showing 2 mice of eachgenotype for each hormonal treatment are shown.
Figure 8. Ovulation involves the LH-dependent activation of 2families of heterotrimeric G proteins. When engaged by an agonist,the LHR present in mural granulosa cells activates Gs and Gq/11. G�s-dependent and -independent pathways that result in the activation ofthe EGF network and the ERK1/2 cascade are involved in 2 of the 3steps required for ovulation, oocyte maturation, and cumulusexpansion. These are not impaired by the deletion of G�q/11ingranulosa cells. Follicular rupture, the final step required for ovulation,is dependent on G�s and G�q/11 and is thus partially disrupted by thedeletion of G�q/11 in granulosa cells. G�q/11-dependent pathways areinvolved in the induction and transactivation of the progesteronereceptor, an event that is required for the induction of Adatms-1 andother genes involved in follicular rupture. G�q/11-dependent pathwaysmay also act downstream of the PGR and participate in follicularrupture by directly modulating gene transcription, protein stability, orby posttranslational protein modifications as shown by the dashedarrow marked with a question mark.
doi: 10.1210/me.2013-1130 mend.endojournals.org 1489
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022

amts-1 is less pronounced than the reduction in the ex-pression of its protein products (Figures 6 and 7). Lastly,because the expression of Cyp19Cre is induced by PMSG(30), we must also consider the possibility that variationin the level of Cre expression among different folliclescould result in mosaicism in the recombination of thefloxed G�q allele. We cannot exclude this possibility, butit appears unlikely because the loss of expression ofG�q/11 in granulosa cells is extensive (Figure 1A) and thepenetrance of the phenotype caused by the Cyp19Cre-mediated recombination of other floxed alleles in granu-losa cells is complete (9, 30, 40–42).
The impairment in ovulation and follicular rupture inthe G�q
f/f;G�11�/�,Cre� mice is very similar to that re-
ported for the Adamts-1�/� mice (18). In contrast to theAdamts-1�/� mice, which also have impaired fertilization(18), the G�q
f/f;G�11�/�,Cre� mice do not (Supplemental
Figure 1). This discrepancy could be explained by thefinding that Adamts-1 is expressed in both mural andcumulus granulosa cells (17), whereas Cyp19Cre and Pgrare expressed only in mural granulosa cells (9, 14, 16, 30,40–42, 47). Thus, the decrease in Adamts-1 expression inthe G�q
f/f;G�11�/�,Cre� mice is restricted to mural gran-
ulosa cells, whereas the decrease in Adamts-1 expressionin the Adamts-1�/� is global and includes cumulus andmural granulosa cells.
There is also a discrepancy in the magnitude of thereduction in ovulation (�50%) and fertility (�85%) inthe G�q
f/f;G�11�/�,Cre� mice (Figure 2). This could be
due to the different experimental paradigms used (cyclingvs superovulated animals), the pharmacokinetics of thehormones that induce ovulation (LH in the cycling fe-males and hCG in the superovulation protocol), or a dis-ruption of other physiological events required for fertilityin cycling mice. We know that the granulosa cell-specificdeletion of G�q/11 does not impair fertilization of theovulated oocytes or luteinization in superovulated mice,and we also know that it does not affect uterine receptiv-ity in pseudopregnant females. On the other hand, wehave not yet examined the effects of the granulosa cell-specific deletion of G�q/11 on the estrous cycle or lutein-ization in cycling or pregnant mice, and thus, it is possiblethat changes in these physiological events also contributeto the larger magnitude of the decrease in fertility com-pared with the decrease in ovulation.
In summary, we have generated a novel mouse modelwith a granulosa cell-specific deletion of G�q/11, and wehave shown that the females experience severe subfertil-ity, a defect in follicular rupture, and a failure of the LHRto fully induce the expression of the Pgr. This is the firstconclusive evidence demonstrating that the in vivo acti-vation of Gq/11 by the LHR is involved in ovulation.
Acknowledgments
We thank JoAnne Richards (Baylor College of Medicine, Hous-ton, Texas) and Doug Stocco (Texas Tech University, Lubbock,Texas) for supplying us with the ADAMTS-1 and StARantibodies.
Address all correspondence and requests for reprints to: Ma-rio Ascoli, PhD, Department of Pharmacology, 2–530 BSB, 51Newton Road, The University of Iowa, Iowa City, Iowa 52242.E-mail: [email protected].
S.M.B. work was supported by a postdoctoral fellowshipfrom the National Institutes of Health (Grant 5F32HD06510).
Disclosure Summary: The authors have nothing to disclose.
References
1. Mehlmann LM. Stops and starts in mammalian oocytes: recentadvances in understanding the regulation of meiotic arrest andoocyte maturation. Reproduction. 2005;130:791–799.
2. Hinckley M, Vaccari S, Horner K, Chen R, Conti M. The G-protein-coupled receptors GPR3 and GPR12 are involved in cAMP signal-ing and maintenance of meiotic arrest in rodent oocytes. Dev. Biol.2005;287:249–261.
3. Norris RP, Ratzan WJ, Freudzon M, et al. Cyclic GMP from thesurrounding somatic cells regulates cyclic AMP and meiosis in themouse oocyte. Development. 2009;136:1869–1878.
4. Vaccari S, Weeks Ii JL, Hsieh M, Menniti FS, Conti M. Cyclic GMPsignaling is involved in the luteinizing hormone-dependent meioticmaturation of mouse oocytes. Biol. Reprod. 2009;81:595–604.
5. Zhang M, Su YQ, Sugiura K, Xia G, Eppig JJ. Granulosa cell ligandNPPC and its receptor NPR2 maintain meiotic arrest in mouseoocytes. Science. 2010;330:366–369.
6. Conti M, Hsieh M, Musa Zamah A, OH JS. Novel signaling mech-anisms in the ovary during oocyte maturation and ovulation. MolCell Endocrinol. 2012;356:65–73.
7. Richards JS, Pangas SA. The ovary: basic biology and clinical im-plications. J Clin Invest. 2010;120:963–972.
8. Fan HY, Liu Z, Mullany LK, Richards JS. Consequences of RASand MAPK activation in the ovary: the good, the bad and the ugly.Mol Cell Endocrinol. 2012;356:74–79.
9. Fan H-Y, Liu Z, Shimada M, et al. MAPK3/1 (ERK1/2) in ovariangranulosa cells are essential for female fertility. Science. 2009;324:938–941.
10. Norris RP, Freudzon M, Mehlmann LM, et al. Luteinizing hormonecauses MAP kinase-dependent phosphorylation and closure of con-nexin 43 gap junctions in mouse ovarian follicles: one of two pathsto meiotic resumption. Development. 2008;135:3229–3238.
11. Norris RP, Freudzon M, Nikolaev VO, Jaffe LA. Epidermal growthfactor receptor kinase activity is required for gap junction closureand for part of the decrease in ovarian follicle cGMP in response toLH. Reproduction. 2010;140:655–662.
12. Hsieh M, Thao K, Conti M. Genetic dissection of epidermal growthfactor receptor signaling during luteinizing hormone-inducedoocyte maturation. PLoS ONE. 2011;6:e21574.
13. Park OK, Mayo KE. Transient expression of progesterone receptormessenger RNA in ovarian granulosa cells after the preovulatoryluteinizing hormone surge. Mol Endocrinol. 1991;5:967–978.
14. Robker RL, Akison LK, Russell DL. Control of oocyte release byprogesterone receptor-regulated gene expression. Nucl Recept Sig-nal. 2009;7:e012.
15. Kim J, Bagchi IC, Bagchi MK. Control of ovulation in mice byprogesterone receptor-regulated gene networks. Mol Hum Reprod.2009;15:821–828.
1490 Breen et al Activation of Gq/11 Is Involved in Ovulation Mol Endocrinol, September 2013, 27(9):1483–1491
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022

16. Robker RL, Russell DL, Espey LL, Lydon JP, O’Malley BW, Rich-ards JS. Progesterone-regulated genes in the ovulation process: AD-AMTS-1 and cathepsin L proteases. Proc Natl Acad Sci USA. 2000;97:4689–4694.
17. Russell DL, Doyle KM, Ochsner SA, Sandy JD, Richards JS. Pro-cessing and localization of ADAMTS-1 and proteolytic cleavage ofversican during cumulus matrix expansion and ovulation. J BiolChem. 2003;278:42330–42339.
18. Brown HM, Dunning KR, Robker RL, et al. ADAMTS1 cleavage ofversican mediates essential structural remodeling of the ovarianfollicle and cumulus-oocyte matrix during ovulation in mice. Biol.Reprod. 2010;83:549–557.
19. Sriraman V, Sharma SC, Richards JS. Transactivation of the pro-gesterone receptor gene in granulosa cells: evidence that Sp1/Sp3binding sites in the proximal promoter play a key role in luteinizinghormone inducibility. Mol Endocrinol. 2003;17:436–449.
20. Davis JS, Weakland LL, West LA, Farese RV. Luteinizing hormonestimulates the formation of inositol triphosphate and cyclic AMP inrat granulosa cells. Biochem J. 1986;238:597–604.
21. Dimino MJ, Snitzer J, Brown KM. Inositol phosphates accumula-tion in ovarian granulosa cells after stimulation by luteinizing hor-mone. Biol Reprod. 1987;37:1129–1134.
22. Donadeu FX, Esteves CL, Doyle LK, Walker CA, Schauer SN, DiazCA. Phospholipase C�3 mediates LH-Induced granulosa cell differ-entiation. Endocrinology. 2011;152:2857–2869.
23. Donadeu FX, Ascoli M. The differential effects of the gonadotropinreceptors on aromatase expression in primary cultures of immaturerat granulosa cells are highly dependent on the density of receptorsexpressed and the activation of the inositol phosphate cascade.Endocrinology. 2005;146:3907–3916.
24. Gudermann T, Birnbaumer M, Birnbaumer L. Evidence for dualcoupling of the murine luteinizing hormone receptor to adenylylcyclase and phosphoinositide breakdown and Ca�2 mobilization.J Biol Chem. 1992;267:4479–4488.
25. Gudermann T, Nichols C, Levy FO, Birnbaumer M, Birnbaumer L.Ca�2 mobilization by the LH receptor expressed in Xenopusoocytes independent of 3�,5�-cyclic adenosine monophosphate for-mation: evidence for parallel activation of two signaling pathways.Mol Endocrinol. 1992;6:272–278.
26. Zhu X, Gilbert S, Birnbaumer M, Birnbaumer L. Dual signalingpotential is common among Gs-coupled receptors and dependenton receptor density. Mol Pharmacol. 1994;46:460–469.
27. Offermanns S, Zhao L-P, Gohla A, Sarosi I, Simon MI, Wilkie TM.Embryonic cardiomyocyte hypoplasia and craniofacial defects inG�q/G�11-mutant mice. EMBO J. 1998;17:4304–4312.
28. Wettschureck N, Lee E, Libutti SK, Offermanns S, Robey PG, Spie-gel AM. Parathyroid-specific double knockout of Gq and G11�-subunits leads to a phenotype resembling germline knockout of theextracellular Ca2�-sensing receptor. Mol Endocrinol. 2007;21:274–280.
29. Kero J, Ahmed K, Wettschureck N, et al. Thyrocyte-specific Gq/G11 deficiency impairs thyroid function and prevents goiter devel-opment. J Clin Invest. 2007;117:2399–2407.
30. Fan H-Y, Shimada M, Liu Z, et al. Selective expression ofKrasG12D in granulosa cells of the mouse ovary causes defects infollicle development and ovulation. Development. 2008;135:2127–2137.
31. Campbell KL. Ovarian granulosa cells isolated with EGTA and
hypertonic sucrose: cellular integrity and function. Biol Reprod.1979;21:773–786.
32. Kaushik D, Reese J, Paria BC. Methodologies to study implantationin mice. In: Soares MJ, Hunt JS, ed. Methods in Molecular Medi-cine: Placenta and Trophoblast: Methods and Protocols. Vol 1.Totowa, NJ: Humana Press; 2006:9–34.
33. Frungieri MB, Weidinger S, Meineke V, Kohn FM, Mayerhofer A.Proliferative action of mast-cell tryptase is mediated by PAR2,COX2, prostaglandins, and PPAR�: possible relevance to humanfibrotic disorders. Proc Natl Acad Sci USA. 2002;99:15072–15077.
34. Andric N, Ascoli M. The lutropin receptor activated ERK1/2 cas-cade stimulates epiregulin release from granulosa cells. Endocrinol-ogy. 2008;149:5549–5556.
35. Andric N, Ascoli M. A delayed, gonadotropin-dependent andgrowth-factor mediated activation of the ERK1/2 cascade nega-tively regulates aromatase expression in granulosa cells. Mol Endo-crinol. 2006;20:3308–3320.
36. Kim J, Sato M, Li Q, et al. Peroxisome proliferator-activated recep-tor � is a target of progesterone regulation in the preovulatoryfollicles and controls ovulation in mice. Mol Cell Biol. 2008;28:1770–1782.
37. Pfaffl MW. A new mathematical model for relative quantificationin real-time RT-PCR. Nucleic Acids Res. 2001;29:e45.
38. Shiraishi K, Ascoli M. Lutropin/choriogonadotropin (LH/CG)stimulate the proliferation of primary cultures of rat Leydig cellsthrough a pathway that involves activation of the ERK1/2 cascade.Endocrinology. 2007;148:3214–3225.
39. Andric N, Thomas M, Ascoli M. Transactivation of the epidermalgrowth factor receptor is involved in the lutropin receptor mediateddown regulation of ovarian aromatase expression in vivo. Mol.Endocrinol. 2010;24:552–560.
40. Fan H-Y, Liu Z, Cahill N, Richards JS. Targeted disruption of ptenin ovarian granulosa cells enhances ovulation and extends the lifespan of luteal cells. Mol Endocrinol. 2008;22:2128–2140.
41. Fan H-Y, Liu Z, Paquet M, et al. Cell type-specific targeted muta-tions of Kras and Pten document proliferation arrest in granulosacells versus oncogenic insult to ovarian surface epithelial cells. Can-cer Res. 2009;69:6463–6472.
42. Fan HY, Liu Z, Johnson PF, Richards JS. CCAAT/enhancer-bind-ing proteins (C/EBP)-� and -� are essential for ovulation, luteiniza-tion, and the expression of key target genes. Mol Endocrinol. 2011;25:253–268.
43. Palanisamy GS, Cheon Y-P, Kim J, et al. A novel pathway involvingprogesterone receptor, endothelin-2, and endothelin receptor Bcontrols ovulation in mice. Mol Endocrinol. 2006;20:2784–2795.
44. Bridges PJ, Jo M, Al Alem L, et al. Production and binding ofendothelin-2 (EDN2) in the rat ovary: endothelin receptor subtypeA (EDNRA)-mediated contraction. Reprod Fertil Dev. 2010;22:780–787.
45. Ko C, Gieske MC, Al-Alem L, et al. Endothelin-2 in ovarian folliclerupture. Endocrinology. 2006;147:1770–1779.
46. Doyle KM, Russell DL, Sriraman V, Richards JS. Coordinate tran-scription of the ADAMTS-1 gene by luteinizing hormone and pro-gesterone receptor. Mol Endocrinol. 2004;18:2463–2478.
47. Conneely OM, Mulac-Jericevic B, DeMayo F, Lydon JP, O’MalleyBW. Reproductive functions of progesterone receptors. RecentProg Horm Res. 2002;57:339–355.
doi: 10.1210/me.2013-1130 mend.endojournals.org 1491
Dow
nloaded from https://academ
ic.oup.com/m
end/article/27/9/1483/2615172 by guest on 10 January 2022





























