Embed Size (px)
Citation preview

CALL FOR PAPERS Cellular Mechanisms of Tissue Fibrosis
Biomechanical strain vehicles for fibroblast-directed skeletal myoblastdifferentiation and myotube functionality in a novel coculture
Michael R. Hicks,1,2 Thanh V. Cao,1 and Paul R. Standley1
1The University of Arizona College of Medicine-Phoenix, Phoenix, Arizona; and 2School of Life Sciences, Arizona StateUniversity, Tempe, Arizona
Submitted 31 October 2013; accepted in final form 13 July 2014
Hicks MR, Cao TV, Standley PR. Biomechanical strain vehiclesfor fibroblast-directed skeletal myoblast differentiation and myotube func-tionality in a novel coculture. Am J Physiol Cell Physiol 307: C671–C683,2014. First published August 13, 2014; doi:10.1152/ajpcell.00335.2013.—Skeletal muscle functionality is governed by multiple stimuli, includ-ing cytokines and biomechanical strain. Fibroblasts embedded withinmuscle connective tissue respond to biomechanical strain by secretingcytokines that induce myoblast differentiation and, we hypothesize,regulate myotube function. A coculture was established to allow crosstalk between fibroblasts in Bioflex wells and myoblasts on nonde-formable coverslips situated above Bioflex wells. Cyclic short-dura-tion strain (CSDS) modeling repetitive stress/injury, acyclic long-duration strain (ALDS) modeling manipulative therapy, and combinedstrain paradigms (CSDS � ALDS) were applied to fibroblasts. Non-strained myoblasts in uniculture and coculture served as controls.After fibroblasts had induced myoblast differentiation, myotubecontraction was assessed by perfusion of ACh (10�11–10�3 M).CSDS-treated fibroblasts increased myotube contractile sensitivityvs. uniculture (P � 0.05). As contraction is dependent on AChbinding, expression and clustering of nicotinic ACh receptors(nAChRs) were measured. CSDS-treated fibroblasts increasednAChR expression (P � 0.05), which correlated with myotubecontraction. ALDS-treated fibroblasts did not significantly affectcontraction or nAChR expression. Agrin-treated myotubes werethen used to design a computer algorithm to identify �-bungaro-toxin-stained nAChR clusters. ALDS-treated fibroblasts increasednAChR clustering (P � 0.05), while CSDS-treated fibroblastsdisrupted cluster formation. CSDS-treated fibroblasts producednAChRs preferentially located in nonclustered regions (P � 0.05).Strain-activated fibroblasts mediate myotube differentiation withmultiple functional phenotypes. Similar to muscle injury, CSDS-treated fibroblasts disrupted nAChR clusters and hypersensitizedmyotube contraction, while ALDS-treated fibroblasts aggregatednAChRs in large clusters, which may have important clinicalimplications. Cellular strategies aimed at improving muscle func-tionality, such as through biomechanical strain vehicles that acti-vate fibroblasts to stabilize postsynaptic nAChRs on nearby skel-etal muscle, may serve as novel targets in neuromuscular disorders.
fibroblasts; skeletal muscle; contraction; nicotinic acetylcholine re-ceptors; biomechanical strain; cytokines
THE LONG-TERM GOAL for muscle regeneration strategies is re-covery of the functional properties of injured muscle. Giventhat 6,000,000 Americans are diagnosed with musculoskeletaldiseases each year, the potential to improve muscle regenera-
tion and drive regeneration to functional maturity is significant(5). Essential to muscle regeneration are satellite cells residingbeneath the basal lamina of each myofiber. After injury, qui-escent satellite cells are activated to form a proliferative pool ofmyoblasts that differentiate and fuse to provide myonuclei forrepair or replacement of damaged myofibers (12).
Once myotubes reach functional maturity, they express andmaintain nicotinic ACh receptors (nAChRs), which mediatemuscle contraction through ligand-binding voltage-gated Na�
channels. Plasticity of nAChRs clustered at the motor endplate, in the context of size, shape, and density, is crucial formuscle homeostasis and function (16, 48). For example, syn-aptic transmission progressively weakens with advanced aging,and its dysfunction is a major pathological feature of neurode-generative diseases such as amyotrophic lateral sclerosis (47).However, physical activity can increase the functional plastic-ity of neuromuscular junctions and may improve motor func-tion for athletes and patients undergoing physical rehabilitation(17, 26). Myoblast differentiation and synthesis of proteinsregulating functional outcomes are dependent on multipleinputs from cytokines and growth factors, including IGF-I,hepatocyte growth factor, IL-6, TNF�, myostatin, and othersderived from the cellular milieu (9). Thus strategies aimed atpromoting the functional regeneration of muscle, such asthrough endogenous cells within the body that supply thecellular milieu with these mediators, are timely for manydemographics (46).
Fibroblasts comprise �50% of the extramuscular cell pop-ulations within skeletal muscle (22) and are a major cellularcomponent of regenerating muscle, ensuring effective andefficient repair (37). Fibroblasts also synthesize a unifyingconnective tissue designed to meet the biophysical demands ofa dynamically functioning muscle and can respond to mechan-ical stimuli within minutes (49). Thus fibroblasts serve assensitive mechanotransducers by modifying expression of ex-tracellular matrix proteins and secreting cytokines that havedocumented effects on nearby muscle (46). We and others havealso shown that different versions of biomechanical strain canuniquely influence behavior of fibroblasts and their resultingsecretions (7, 36); therefore, strain may serve as a vehicle todirect fibroblast-mediated muscle repair. Supporting evidenceshows that regimental stretching can reorganize fibroblast-richconnective tissue and induce muscle hypertrophy (14), whichhas lasting synergistic effects on strength gains and muscleactivation (32). However, fibroblasts also synthesize fibroticand inflammatory molecules, which are pathogenic to skeletalmuscle (18, 44), and repetitive tasks that result in excessive use
Address for reprint requests and other correspondence: M. R. Hicks, TheUniv. of Arizona College of Medicine-Phoenix, 425 N. 5th St., Rm. 331H-ABC1, Phoenix, AZ 85004 (e-mail: [email protected]).
Am J Physiol Cell Physiol 307: C671–C683, 2014.First published August 13, 2014; doi:10.1152/ajpcell.00335.2013.
0363-6143/14 Copyright © 2014 the American Physiological Societyhttp://www.ajpcell.org C671

of skeletal muscle and its surrounding muscle fascia can resultin fibroblast-mediated inflammation, strain-induced injury, anddystonia (29). Understanding the mechanisms by which bio-mechanical strain influences the bipotent roles of fibroblasts isa major step in shifting fibroblasts toward a phenotype prefer-ential to the functional regeneration of skeletal muscle.
Our laboratory has developed methods to mechanicallystrained fibroblasts seeded on flexible membranes (35). With aclinically relevant stretch treatment as a reference, we devel-oped an acyclic long-duration strain (ALDS) regimen aftermyofascial release, a form of manipulative treatment thatapplies sustained force to fibroblast-rich tissues to releaserestriction points (41). By analyzing video recordings of clini-cians manually performing myofascial release, we determinedthe strain magnitude and duration required to develop an invitro model. We have shown that a single ALDS treatment hassignificant effects on fibroblast secretions, including angioge-nin, granulocyte colony-stimulating factor, thymus and activa-tion-regulated chemokine, IL-1�, IL-3, and IL-8 (7). We alsodeveloped a cyclic short-duration strain (CSDS) regimen toinduce a stress injury response in the fibroblasts and a com-bined strain regimen (CSDS � ALDS) to model a repetitivestrain injury followed by a manipulative treatment, as mighttypically be experienced clinically. We have shown that 1 minof ALDS can reverse CSDS-induced disorganization of fibro-blast actin architecture and proximally decreases lamellipodiaas to change fibroblast morphology from an elongated to astellate shape (35).
Using these models, we built a fibroblast-skeletal musclecoculture and demonstrated that strain-activated fibroblastssignificantly enhance myoblast differentiation through IL-6(27). The current study extends our previous work byinvestigating the functional phenotype of myotubes differ-entiated in the presence of strain-activated fibroblasts. Wehypothesized that fibroblasts subjected to specific variationsof strain magnitude, duration, and frequency could modifythe differentiation or development of nearby myotubes anduse contractility and nAChR expression and organization asdifferentiation markers. We also hypothesized that CSDS-treated fibroblasts would induce an injury-like myotubephenotype relative to nonstrained fibroblasts and that ALDSapplied after CSDS would recover CSDS-mediated effects.We developed a novel method to investigate in vitro myo-tube contraction in response to incremental doses of ACh.While others used electric pulse stimulation to quantifyskeletal myotube contraction in vitro (33), our approachallowed for a more direct correlation to nAChR expressionand clustering in myotube populations. We also used quan-titative fluorescence imaging to measure nAChR clustersbased on previous studies (1, 25) and show that strained andnonstrained fibroblasts are capable of regulating the pheno-typic outcomes of myotubes. Thus fibroblasts may serve asa novel target for strategies aimed at facilitating the func-tional regeneration of skeletal muscle.
METHODS
Coculture setup and application of biomechanical strain tofibroblasts. A coculture was established to allow paracrine interac-tions between fibroblasts (normal human dermal fibroblasts; CambrexLaboratories) and myoblasts [C2C12 cells; American Type CultureCollection (ATCC)]. Fibroblasts were seeded (6.5 � 104 cells/cm2,
2% FBS) in 35-mm Bioflex wells, which contain an elastomericsurface membrane, allowing adherent cells to deform under vacuum(model BF3001-C, Flexcell International, Hillsborough, NC). Myo-blasts were seeded (4.1 � 104 cells/cm2, 10% FBS) on 34 � 13-mmnondeformable glass coverslips custom-made to be affixed 2 mmabove the Bioflex wells in an arrangement that allowed fibroblasts, butnot myoblasts, to be mechanically strained (Fig. 1A). Cell cultureswere kept at 5% CO2 and 37°C in all experiments. Differentiationmedium (DM, 2% horse serum; ATCC) was used as a positive controlto verify that lot-specific myoblasts were capable of differentiation. Inno instance were myotubes from DM and FBS media statisticallycompared, as differences in sera, seeding densities, and culture timemay have confounded the analyses. DMEM (catalog no. 11885076,Life Technologies) served as a basal medium for FBS and horseserum.
After 24 h, coverslips containing myoblasts were transferred toBioflex wells containing fibroblasts and oriented such that the myo-blasts faced the fibroblasts. The coverslip and Bioflex membranedimensions allowed for 5.2-cm2 of gas exchange per 9.6-cm2 surfacearea. Immediately after the coculture was established, fresh growthmedium (2% FBS in DMEM) was added, and strain regimens wereinitiated. The FX-4000 Tension Plus System (Flexcell International) isa computer-controlled device designed to deliver negative-pressurevacuum through user programming of strain magnitude, duration, andfrequency. Bioflex membranes were strained equiradially using fourprotocols, as described previously (27, 35). 1) For the nonstrainprotocol, fibroblasts were grown on Bioflex membranes but were notsubjected to strain. 2) For the ALDS protocol, fibroblasts grown onBioflex membranes were subjected to a single acyclic strain. Mem-branes were elongated 3%/s until they were 6% beyond resting length.Strain was held for 60 s and returned to baseline at 1.5%/s. 3) For theCSDS protocol, fibroblasts grown on Bioflex membranes were cycli-cally strained at 0.6 Hz for 8 h. Membranes were elongated 33%/suntil they were 10% beyond resting length and then immediatelyreturned to baseline at 33%/s. There was a 1-s rest phase between theend of one cycle and the beginning of the next cycle. 4) For the CSDS �ALDS protocol, fibroblasts on Bioflex membranes were subjected toCSDS, allowed to rest for 3 h, and then subjected to ALDS.
After strain regimens, cells remained in cocultures without mediumchange for 96 h to allow secreted mediators from strain-activatedfibroblasts to induce myotube differentiation, as we previously re-ported (27). Coverslips containing differentiated myotubes were thenassessed for myotube differentiation parameters and resulting func-tional phenotypes (see below).
Measurements of strain-induced fluid forces on myoblast viabi-lity and myotube differentiation in the presence and absence offibroblasts. To test whether myotubes exposed to the strain environ-ment were affected by fluid forces and media agitation, C2C12 cellsin non-fibroblast-stretch experiments were analyzed for proliferation,differentiation, and viability at 3, 48, and 96 h poststrain (Fig. 1).Myoblast proliferation was measured by counting DAPI-labeled nu-clei per millimeter squared, myotube differentiation was measured bycounting cells expressing myosin with at least three nuclei, and fusionindex was calculated as the percentage of nuclei in myotubes relativeto the entire C2C12 cell population. C2C12 cells are highly metabol-ically active, and because 96 h without medium change could poten-tially impact cell viability, extracellular DNA produced as a result ofcell death was assayed from conditioned medium (catalog no. G8742,Promega). Manufacturer-recommended culture conditions for C2C12cells that include a medium change every 24 h and a starting celldensity of 13.3 � 104 cells/ml in 10% FBS were used for comparison.Data are presented in raw fluorescence units per culture well. Trypanblue exclusion was also performed for all time points and treatments.While we previously analyzed myotube differentiation from all straintreatments in 21 separate cell cultures (27), in this set of experimentswe exposed C2C12 cells in uniculture and coculture to only the CSDSand nonstrain paradigms to extend our previous work on the effects of
C672 FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
non-fibroblast-stretch experiments. We included a 3-h post-CSDStime point, when ALDS would typically be applied in the CSDS �ALDS paradigm, and a 48-h time point, half-way through the exper-iments, to measure earlier effects on myoblast proliferation and cellviability. To establish proof of concept for early myogenic activationby fibroblasts and strain, we also measured myogenin and MyoD(catalog nos. QT00112378 and QT00101983, Qiagen) by poolingmRNA from two culture wells per treatment (1 g) and analyzing byreal-time PCR, with expression of each gene normalized to that of
GAPDH using the threshold cycle (CT) method (catalog no.QT01658692, Qiagen). Six independent samples were analyzed (n �6, see Statistics).
Myotube contraction studies. Next, we were interested in determin-ing whether the myotubes differentiated in the presence of strainedand nonstrained fibroblasts were functionally different from oneanother. Thus a perfusion system was devised to induce in vitromyotube contraction and test the functional capacity of coculturedmyotubes. Coverslips containing myotubes were removed from co-
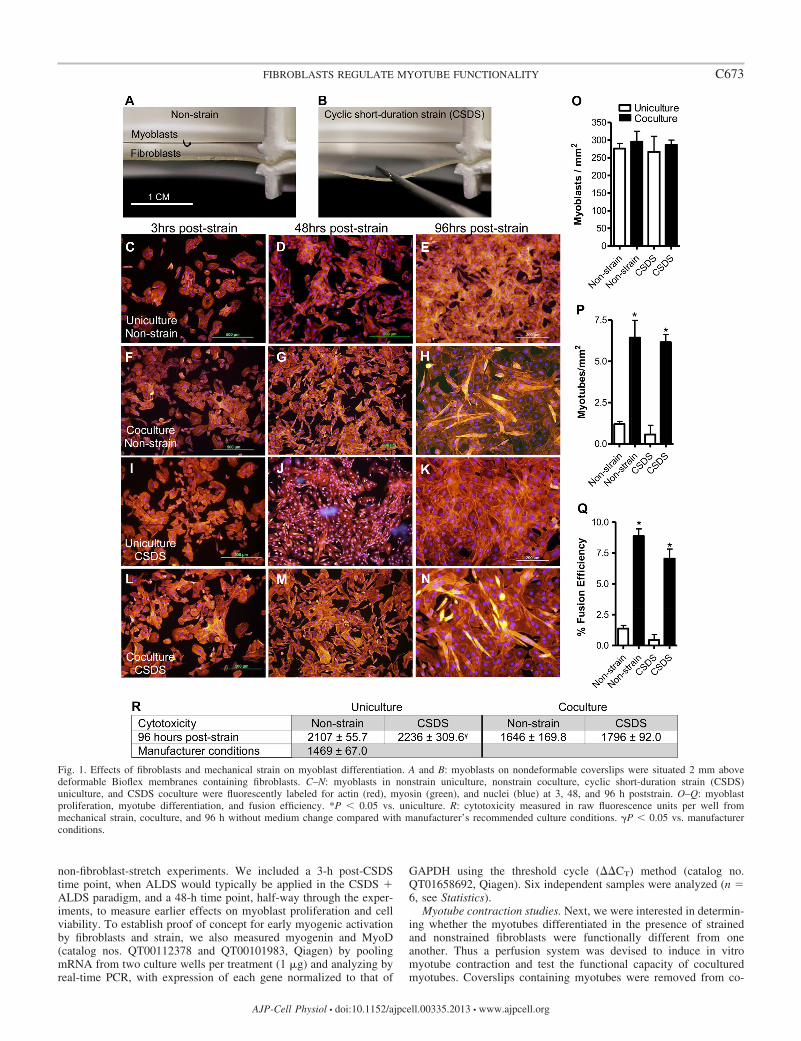
Fig. 1. Effects of fibroblasts and mechanical strain on myoblast differentiation. A and B: myoblasts on nondeformable coverslips were situated 2 mm abovedeformable Bioflex membranes containing fibroblasts. C–N: myoblasts in nonstrain uniculture, nonstrain coculture, cyclic short-duration strain (CSDS)uniculture, and CSDS coculture were fluorescently labeled for actin (red), myosin (green), and nuclei (blue) at 3, 48, and 96 h poststrain. O–Q: myoblastproliferation, myotube differentiation, and fusion efficiency. *P � 0.05 vs. uniculture. R: cytotoxicity measured in raw fluorescence units per well frommechanical strain, coculture, and 96 h without medium change compared with manufacturer’s recommended culture conditions. �P � 0.05 vs. manufacturerconditions.
C673FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org

cultures and outfitted in 35-mm perfusion plates with attached inflowand outflow tubing holders. The apparatus allowed for continuoussuperfusion of myotubes with various contractile agonists and washbuffers. The velocity of perfusate was controlled by two variable-flowperistaltic pumps (model 138761, Fisher Scientific), and the perfusatewas delivered through 1/16-in. tubing at 4.2 cm/s. The outflow tubingwas situated 3 mm above the inflow tubing to ensure constantsubmersion of myotubes with perfusate. Perfusion plates were posi-tioned on the stage of an IX71 Olympus inverted microscope thatmonitored perfusions by video microscopy, capturing 29 frames persecond. High-powered fields (HPFs) with at least three myotubes wererandomly selected for perfusion to account for fields of view contain-ing no myotubes, as typically found in uniculture (Fig. 1E). Myotubeswere defined as syncytial cells with at least three nuclei (27).
Myotubes were sequentially perfused for 1 min with log10 incre-ments of ACh (10�11–10�4 M). Myotubes were washed with 1� PBSfor 1 min between perfusions to remove previously perfused ACh. Allperfusates were isosmotic and kept at 37°C, pH 7.4, to negate theeffects of osmolality, temperature, and pH on myotube contraction.The perfusion plate was placed on a transparent 37°C thermoplatestage-top incubator (model MATS-U55R30, Tokai Hit) during videocapture for additional temperature control.
To separate medium- and perfusion-related changes from ACh-specific effects relating to progressive decreases in myotube area, weperformed additional experiments in which myotubes differentiated inthe presence of nonstrained fibroblasts were perfused with one of fourperfusates. Perfusate groups included PBS only, PBS containing ACh(10�11–10�3 M), PBS containing ACh (10�11–10�3 M) and 1.8 mMCa2�, and PBS containing ACh (10�11–10�3 M) perfused in myo-tubes treated with unconjugated �-bungarotoxin (�-BGT; catalog no.ab120542, Abcam). Myotubes treated with �-BGT were immediatelytransferred to our perfusion apparatus, and ACh was administered (seeFig. 3B). A final dose of KCl (10�2 M) was added to all treatments todepolarize the myotube sarcolemma independently of nAChR activa-tion.
Sequential videos of ACh- and KCl-induced myotube contractionswere analyzed with Adobe Premiere Pro 5.0. The last still frame fromeach perfusion video was extracted, and images were input intoImageJ for analyses. For length-shortening experiments, 10 randomcoordinate points were identified on each myotube and measuredusing ImageJ. Alternatively, sarcolemma perimeters (Fig. 2B, shownin phase-contrast mode) were carefully outlined and computationallyfilled to produce binary images representative of myotube areas on ablank canvas. To avoid potential bias, all fully visible myotubes perfield, except those that overlapped each other in a manner that couldpotentially influence contraction, were outlined. Binary images ofoutlined myotubes were then exported to ImageJ, and myotube areaswere measured. All myotubes per HPF video were analyzed as percentcontraction relative to their own precontractile area. ACh-inducedmyotube contraction is reported as the percentage of fractional short-ening relative to a precontractile value of 100%. Maximum fractionalshortening in response to ACh, half-maximum (50%) effective dose(ED50), and Hill slope, a measure describing the fraction of nAChRsbound by ACh (40), were calculated (see Statistics). The R2 valuequantifies Hill slope goodness of fit. The minimal fractional shorten-ing required to cause statistically significant contraction from aprecontractile value of 100% was also determined. Myotube contrac-tion values from each HPF were averaged from one video per HPF (n �1). A total of 12 perfusion videos (corresponding to 105–140 myo-tubes) were independently captured for each treatment (n � 12).
Quantification of AChRs by Western blotting. To measure nAChRexpression, myotubes were removed from coverslips using a cellscraper and lysis buffer (catalog no. P8340, Sigma). Three coverslipsper treatment were pooled to acquire sufficient protein (50 g/lane).Proteins were separated on 4–20% Tris·HCl gels in a Mini-Protean IIelectrophoresis cell (Bio-Rad). Samples were transferred to nitrocel-lulose membranes and probed for AChR (57 kDa; catalog no. sc-
14000, Santa Cruz Biotechnology). Fluorescent secondary antibodies(catalog no. 926-32211, LI-COR) were used to label AChR at awavelength of 800 nm and the molecular weight ladder (catalog no.928-40000, LI-COR) at 700 nm. Membranes were stripped using 0.2M NaOH for 1 min. They were washed five times with PBS andreprobed for GAPDH (33 kDa; catalog no. sc-25778, Santa CruzBiotechnology). Protein expression from Western blotting was calcu-lated using relative quantification from fluorescence densitometry datagenerated by LI-COR Odyssey software. Myoblasts in uniculture,seeded on coverslips and transferred to Bioflex wells devoid offibroblasts (2% FBS), served as a negative control. Treatment resultsare expressed as fold change from uniculture and normalized toendogenous control GAPDH. Seven independent samples (21 pooledwells) were analyzed per treatment (n � 7).
Identification of AChR clusters for quantitative analyses. AChRspresent on differentiated myotubes were fluorescently labeled withtetramethylrhodamine-�-BGT (catalog no. T1175, Molecular Probes),and quantitative fluorescence imaging was performed using an open-source software package in CellProfiler (Broad Institute). The auto-mated image cytometry system identifies and measures objects’ size,shape, pixel intensity, and topology, and resulting acquired data arecontrasted against a background-subtraction algorithm (8). Settings toidentify nAChR clusters in CellProfiler were established using agrin-treated myotubes (16 h, 1 g/ml recombinant rat agrin; catalog no.550-AG100, R & D Systems) grown in DM. Agrin, normally secretedby motor neurons, stabilizes and aggregates nAChRs into distinguish-able cluster regions (�65% intensity), whereas outside these regions,nAChRs largely exist in dispersed microclusters (�65% intensity)denoted by lower-intensity pixels, as described by others (1, 19).Myotubes from coculture experiments were incubated with 1 g/ml�-BGT for 30 min; then coverslips containing myotubes were re-moved from coculture, fixed with 2% formaldehyde, and mounted onprelabeled microscope slides in DAPI-VECTASHIELD (catalog no.H1200, Sigma). Treatment identification labels were concealed andcoded, and slides were imaged by a blinded observer. Microscopesettings for intensity, exposure time, and staining techniques werestringently controlled to maintain fluorescence consistency over themonths �-BGT experiments were conducted. As a gain control, a HPFdevoid of myotubes was selected, and gain settings were decreaseduntil the HPF was completely black. Then 6–16 images were ran-domly obtained at �20 magnification (0.58 mm2/HPF) for eachindependent cell culture. To further account for variations in fluores-cence intensity, data were analyzed using two-factor ANOVA, withthe date of staining and treatment group as independent variables.Twelve randomly and blindly selected myotubes from each indepen-dent cell culture were analyzed by CellProfiler. In this manner,nAChR area measurements were obtained for individual myotubes(108 myotubes/treatment, n � 9). To further investigate if strain-activated fibroblasts remodeled �-BGT-stained regions, a histogramthat quantified the size distribution of AChR clusters, 4–1,024 m2,was created.
Statistical analysis. Data were analyzed using one-factor ANOVAwith post hoc Tukey’s tests and P � 0.05 to test significance, unlessspecifically stated. For cytotoxicity studies, post hoc Dunnett’s testwas used to compare treatment groups with the manufacturer-recom-mended culture conditions (P � 0.05). For contraction studies, AChconcentrations were converted to log10, and a four-parameter logisticssigmoid-shaped dose-response curve with variable slope was used todetermine log(ED50) and Hill slope for each treatment. A precontrac-tile value of 100% was assigned to all groups, and a one-sample t-testwith null hypothesis that the mean is equal to 100% was used todetermine the threshold concentration of ACh that caused a differencefrom precontraction. For nAChR cluster distribution studies, welogarithmically transformed our histogram to reflect a distributionwhere the y-axis vs. the logarithm of the x-axis was normal (orGaussian) in nature as surface area of the receptors (m2) increasedlogarithmically. The geometric mean was used to calculate the aver-
C674 FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
age AChR cluster size of the lognormal distribution. Data betweenexperiments were then correlated by plotting the combined means SE derived from each experimental procedure and each treatmentgroup. Linear regression analysis was used to calculate R2. Student’st-tests were used to compare differences between agrin-treated anduntreated myotubes grown in DM (P � 0.05). All figures were madeusing GraphPad PRISM 5.0.
RESULTS
Fibroblasts enhance myoblast differentiation in coculture.We have developed a novel fibroblast-skeletal muscle cocul-ture to allow myoblast differentiation in the immediate vicinityof strained fibroblasts that does not require replacement offibroblast-conditioned medium. At no point did coculture resultin increased cytotoxicity. Although cell number in coculturewas more than double that in uniculture, cytotoxicity wasreduced, suggesting that fibroblasts and their secretions in-crease the vitality of nearby myoblasts. Strain-dependenttrends exerted a 6.0–9.1% increase in cytotoxicity vs. non-
strain, which resulted in significance between ATCC-recom-mended culturing conditions and CSDS applied to emptyBioflex wells (P � 0.05; Fig. 1R). Myoblast differentiation wasnot measured until 96 h poststrain, perhaps due to the lowseeding density and reduced FBS concentration in our system.Importantly, however, at 96 h fibroblasts significantly in-creased myoblast differentiation and fusion index vs. unicul-ture (P � 0.05; Fig. 1, P and Q). Application of CSDS did notsignificantly affect myoblast differentiation vs. nonstrain, butstrain-dependent trends show decreased myoblast fusion. Myo-blast proliferation was also not found to be different acrosstreatment groups (Fig. 1O). However, at 3 and 48 h poststrain,the density of myoblasts per square millimeter varied within asingle coverslip, resulting in large 95% confidence intervals(data not shown). In a separate study, we pooled two indepen-dent cultures from each treatment and measured myogenin andMyoD. Myogenin expression increased in nonstrain cocultureby 8.8-fold and in CSDS coculture by 13.3-fold vs. uniculture
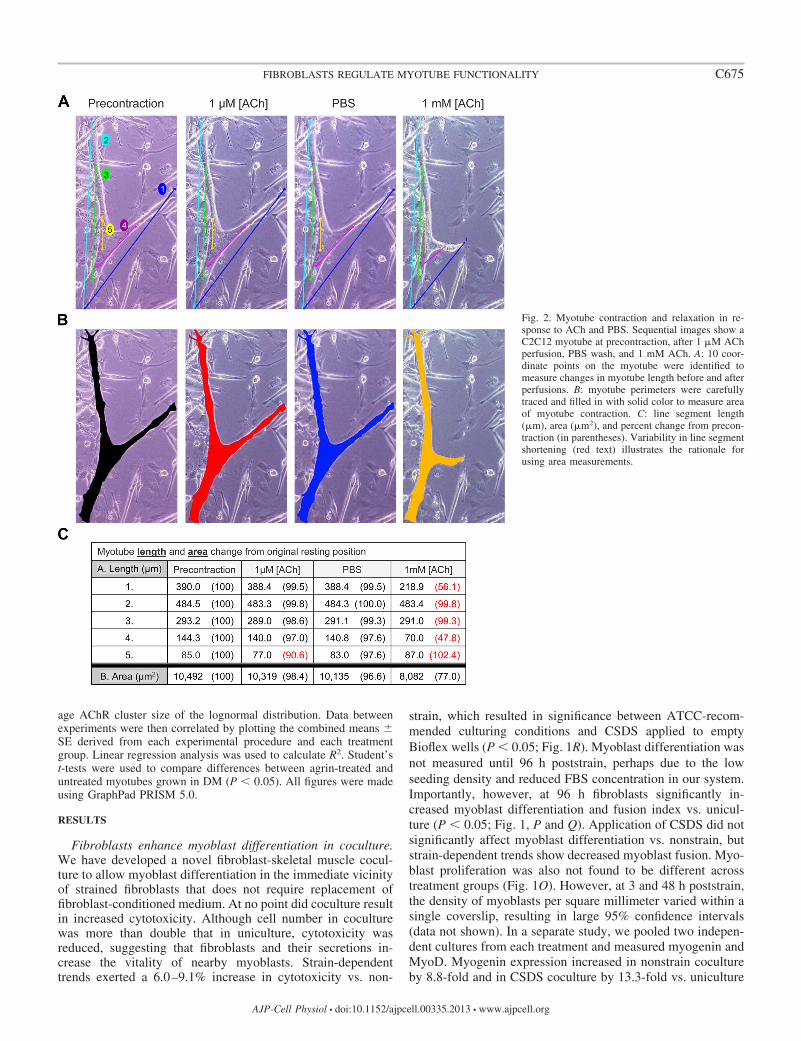
Fig. 2. Myotube contraction and relaxation in re-sponse to ACh and PBS. Sequential images show aC2C12 myotube at precontraction, after 1 M AChperfusion, PBS wash, and 1 mM ACh. A: 10 coor-dinate points on the myotube were identified tomeasure changes in myotube length before and afterperfusions. B: myotube perimeters were carefullytraced and filled in with solid color to measure areaof myotube contraction. C: line segment length(m), area (m2), and percent change from precon-traction (in parentheses). Variability in line segmentshortening (red text) illustrates the rationale forusing area measurements.
C675FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
at 24 h poststrain (P � 0.05; data not shown). However, we didnot measure a change in MyoD, perhaps because it is alsoexpressed by proliferating myoblasts. In combination with ourlaboratory’s previous findings (27), these data served as a basisfor investigating the functional phenotype of myotubes differ-entiated in the presence of strained fibroblasts.
Length measurements of intracellular myotube organellesshow a contraction-relaxation-like response to ACh. Myotubelength measurements identified shortening of intracellularstructures in response to low doses of ACh that returned tooriginal resting length during PBS wash, suggesting that con-traction-relaxation occurred in myotubes (Fig. 2A). However,other coordinates on the same the myotube did not changeposition. Thus measurements of myotube length producedvariable results, depending on the placement of the line seg-ment, making it impossible to reproducibly measure changesbetween myotubes.
Because of the variability in ACh-induced length shortening,the total area of the myotube was measured (Fig. 2B). How-ever, the contraction-relaxation of intracellular structures iden-tified by line segment shortening could not be ascertained whenchanges in myotube area were measured. Therefore, ACh-induced loss of cell adhesion was used as a surrogate formyotube contraction. Importantly, we show that increaseddoses of ACh resulted in progressive decreases in myotubearea consistently measurable across all treatment groups.
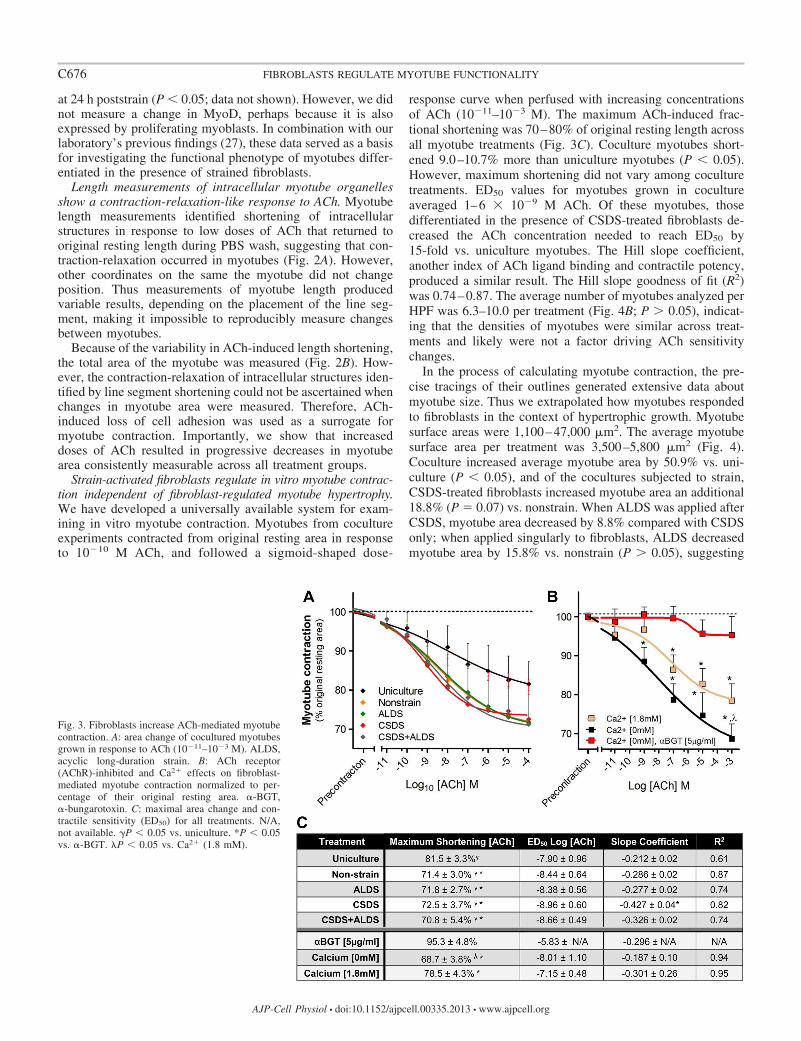
Strain-activated fibroblasts regulate in vitro myotube contrac-tion independent of fibroblast-regulated myotube hypertrophy.We have developed a universally available system for exam-ining in vitro myotube contraction. Myotubes from cocultureexperiments contracted from original resting area in responseto 10�10 M ACh, and followed a sigmoid-shaped dose-
response curve when perfused with increasing concentrationsof ACh (10�11–10�3 M). The maximum ACh-induced frac-tional shortening was 70–80% of original resting length acrossall myotube treatments (Fig. 3C). Coculture myotubes short-ened 9.0–10.7% more than uniculture myotubes (P � 0.05).However, maximum shortening did not vary among coculturetreatments. ED50 values for myotubes grown in cocultureaveraged 1–6 � 10�9 M ACh. Of these myotubes, thosedifferentiated in the presence of CSDS-treated fibroblasts de-creased the ACh concentration needed to reach ED50 by15-fold vs. uniculture myotubes. The Hill slope coefficient,another index of ACh ligand binding and contractile potency,produced a similar result. The Hill slope goodness of fit (R2)was 0.74–0.87. The average number of myotubes analyzed perHPF was 6.3–10.0 per treatment (Fig. 4B; P � 0.05), indicat-ing that the densities of myotubes were similar across treat-ments and likely were not a factor driving ACh sensitivitychanges.
In the process of calculating myotube contraction, the pre-cise tracings of their outlines generated extensive data aboutmyotube size. Thus we extrapolated how myotubes respondedto fibroblasts in the context of hypertrophic growth. Myotubesurface areas were 1,100–47,000 m2. The average myotubesurface area per treatment was 3,500–5,800 m2 (Fig. 4).Coculture increased average myotube area by 50.9% vs. uni-culture (P � 0.05), and of the cocultures subjected to strain,CSDS-treated fibroblasts increased myotube area an additional18.8% (P � 0.07) vs. nonstrain. When ALDS was applied afterCSDS, myotube area decreased by 8.8% compared with CSDSonly; when applied singularly to fibroblasts, ALDS decreasedmyotube area by 15.8% vs. nonstrain (P � 0.05), suggesting
Fig. 3. Fibroblasts increase ACh-mediated myotubecontraction. A: area change of cocultured myotubesgrown in response to ACh (10�11–10�3 M). ALDS,acyclic long-duration strain. B: ACh receptor(AChR)-inhibited and Ca2� effects on fibroblast-mediated myotube contraction normalized to per-centage of their original resting area. �-BGT,�-bungarotoxin. C: maximal area change and con-tractile sensitivity (ED50) for all treatments. N/A,not available. �P � 0.05 vs. uniculture. *P � 0.05vs. �-BGT. �P � 0.05 vs. Ca2� (1.8 mM).
C676 FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
that specific strain paradigms applied to fibroblasts regulatemyotube hypertrophy.
To determine whether myotube contraction was a functionof surface area, myotubes from nonstrain coculture were brack-eted into �2,500-, 2,500- to 5,000-, 5,000- to 7,500-, 7,500- to10,000-, and �10,000-m2 areas, and sigmoid-shaped dose-response curves were analyzed. We found that Hill slopecoefficients were greatest for small-area (�5,000-m2) myo-tubes. However, there were no significant differences in ED50
values among myotube areas (2.1–6 nM, data not shown).Thus we concluded that fibroblast-induced myotube hypertro-phy did not account for the contractile responses induced byACh among coculture treatments.
AChR binding is required for in vitro myotube contraction.To show that ACh-AChR binding is a necessary component forthe progressive decreases in myotube area measured by our invitro apparatus, unconjugated �-BGT was added to myotubesto block nAChRs. We found that unconjugated �-BGT (5g/ml) incubated in coculture for 1 h was sufficient to blockthe binding of rhodamine-conjugated �-BGT without detect-able cytotoxic effects (data not shown). �-BGT-treated myo-tubes were then immediately perfused with ACh (10�11–10�3
M). No concentration of ACh could decrease fractional short-ening of myotubes �95% of their original resting length (Fig.3B). To show that �-BGT-treated myotubes were contractileindependent of nAChRs, we administered a final dose of KCl(10�2 M), which induced area shortening greater than precon-traction (P � 0.05).
Our PBS-based perfusate (catalog no. P3813, Sigma) did notcontain Ca2�, and many adhesion molecules are Ca2�-depen-dent. To test whether Ca2�-dependent adhesion affected frac-
tional shortening, 1.8 mM Ca2� was added to the perfusate,and ACh (10�11–10�3 M) was administered to myotubesdifferentiated in the presence of nonstrained fibroblasts. Ca2�-containing perfusate had no effect on the log(ACh) ED50 vs.Ca2�-free perfusate (�7.42 0.5 and �7.45 1.1, respec-tively). However, Ca2� could not induce the maximal frac-tional shortening of Ca2�-free perfusate by 78.5 4.3% and68.7 3.8, respectively (P � 0.05; Fig. 3B). PBS only wasalso used to verify that the superficial velocity of the perfusatewas not shearing myotubes from the coverslips. PBS only wasnot capable of inducing loss of cell adhesion (data not shown).
Together, these data show that our in vitro contraction modeldecreases the original resting length of myotubes within therange of sarcomere-dependent shortening (42). Of the param-eters tested, Ca2� in the ACh medium blocked 31% of cellcontraction (between 32% shortening in Ca2�-free mediumand 22% shortening in 1.8 mM Ca2�), suggesting that adhesionmolecules in myotube cultures may attest to the isometric-likecontractions identified in length-shortening experiments (Fig.2A). However, �-BGT inhibition of nAChRs prevented 84% ofcell contraction and suggests that the major source of fractionalshortening in our system is derived from an ACh-nAChR-mediated process.
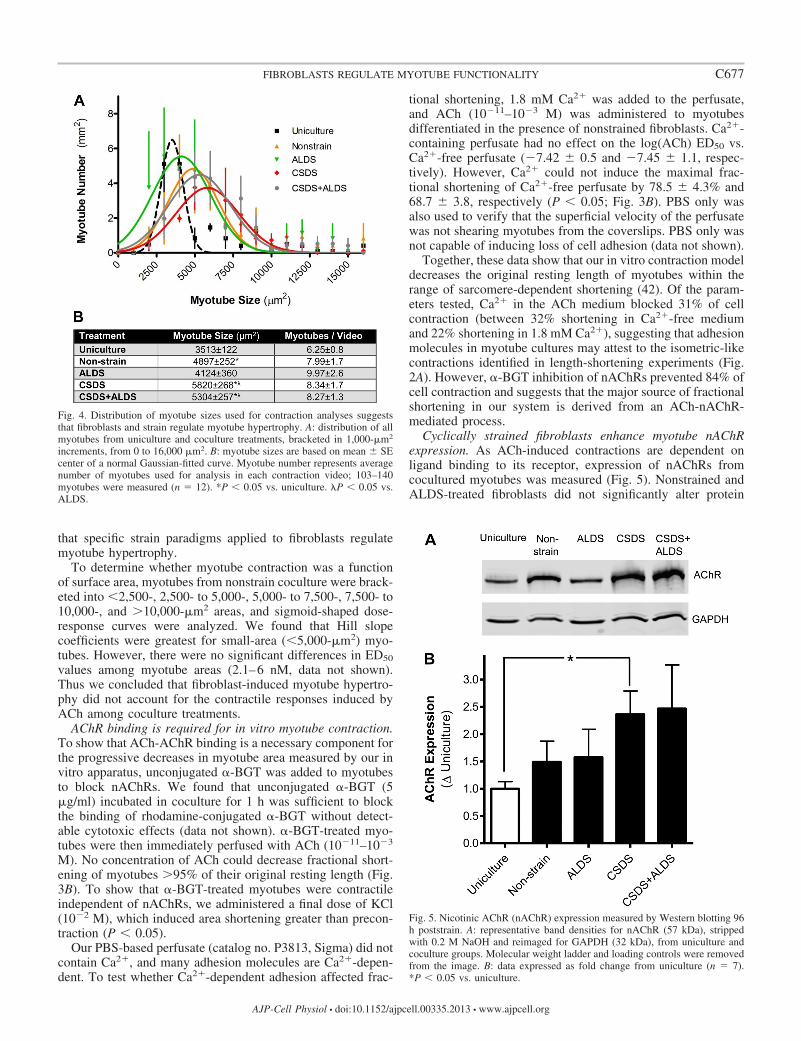
Cyclically strained fibroblasts enhance myotube nAChRexpression. As ACh-induced contractions are dependent onligand binding to its receptor, expression of nAChRs fromcocultured myotubes was measured (Fig. 5). Nonstrained andALDS-treated fibroblasts did not significantly alter protein
Fig. 4. Distribution of myotube sizes used for contraction analyses suggeststhat fibroblasts and strain regulate myotube hypertrophy. A: distribution of allmyotubes from uniculture and coculture treatments, bracketed in 1,000-m2
increments, from 0 to 16,000 m2. B: myotube sizes are based on mean SEcenter of a normal Gaussian-fitted curve. Myotube number represents averagenumber of myotubes used for analysis in each contraction video; 103–140myotubes were measured (n � 12). *P � 0.05 vs. uniculture. �P � 0.05 vs.ALDS.
Fig. 5. Nicotinic AChR (nAChR) expression measured by Western blotting 96h poststrain. A: representative band densities for nAChR (57 kDa), strippedwith 0.2 M NaOH and reimaged for GAPDH (32 kDa), from uniculture andcoculture groups. Molecular weight ladder and loading controls were removedfrom the image. B: data expressed as fold change from uniculture (n � 7).*P � 0.05 vs. uniculture.
C677FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
expression vs. uniculture (1.50 0.37 and 1.57 0.51 fold,respectively). However, CSDS-treated fibroblasts increasedAChR expression vs. uniculture (2.36 0.43 fold, P � 0.05),and CSDS � ALDS-treated fibroblasts followed a similar trend(2.46 0.79 fold, P � 0.10). Expression among coculturegroups was not significantly different. We also measurednAChR expression from agrin-treated and untreated myotubesgrown in DM. Both groups expressed nAChR but were notsignificantly different from one another (data not shown).
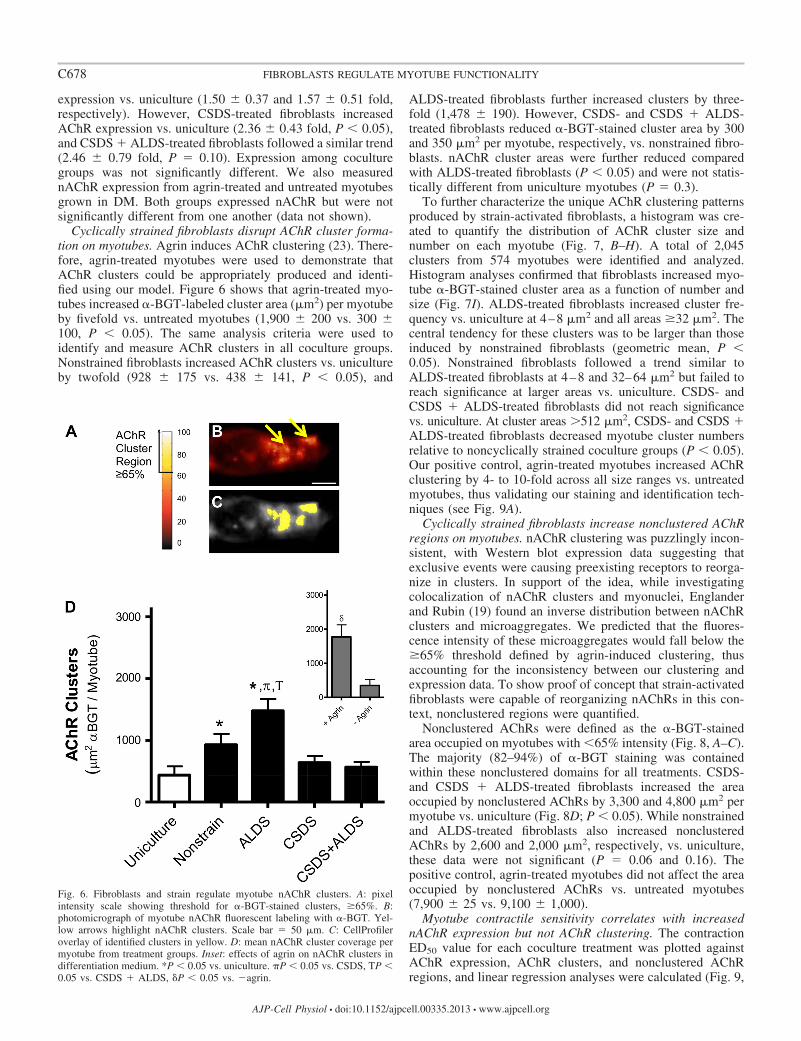
Cyclically strained fibroblasts disrupt AChR cluster forma-tion on myotubes. Agrin induces AChR clustering (23). There-fore, agrin-treated myotubes were used to demonstrate thatAChR clusters could be appropriately produced and identi-fied using our model. Figure 6 shows that agrin-treated myo-tubes increased �-BGT-labeled cluster area (m2) per myotubeby fivefold vs. untreated myotubes (1,900 200 vs. 300 100, P � 0.05). The same analysis criteria were used toidentify and measure AChR clusters in all coculture groups.Nonstrained fibroblasts increased AChR clusters vs. unicultureby twofold (928 175 vs. 438 141, P � 0.05), and
ALDS-treated fibroblasts further increased clusters by three-fold (1,478 190). However, CSDS- and CSDS � ALDS-treated fibroblasts reduced �-BGT-stained cluster area by 300and 350 m2 per myotube, respectively, vs. nonstrained fibro-blasts. nAChR cluster areas were further reduced comparedwith ALDS-treated fibroblasts (P � 0.05) and were not statis-tically different from uniculture myotubes (P � 0.3).
To further characterize the unique AChR clustering patternsproduced by strain-activated fibroblasts, a histogram was cre-ated to quantify the distribution of AChR cluster size andnumber on each myotube (Fig. 7, B–H). A total of 2,045clusters from 574 myotubes were identified and analyzed.Histogram analyses confirmed that fibroblasts increased myo-tube �-BGT-stained cluster area as a function of number andsize (Fig. 7I). ALDS-treated fibroblasts increased cluster fre-quency vs. uniculture at 4–8 m2 and all areas �32 m2. Thecentral tendency for these clusters was to be larger than thoseinduced by nonstrained fibroblasts (geometric mean, P �0.05). Nonstrained fibroblasts followed a trend similar toALDS-treated fibroblasts at 4–8 and 32–64 m2 but failed toreach significance at larger areas vs. uniculture. CSDS- andCSDS � ALDS-treated fibroblasts did not reach significancevs. uniculture. At cluster areas �512 m2, CSDS- and CSDS �ALDS-treated fibroblasts decreased myotube cluster numbersrelative to noncyclically strained coculture groups (P � 0.05).Our positive control, agrin-treated myotubes increased AChRclustering by 4- to 10-fold across all size ranges vs. untreatedmyotubes, thus validating our staining and identification tech-niques (see Fig. 9A).
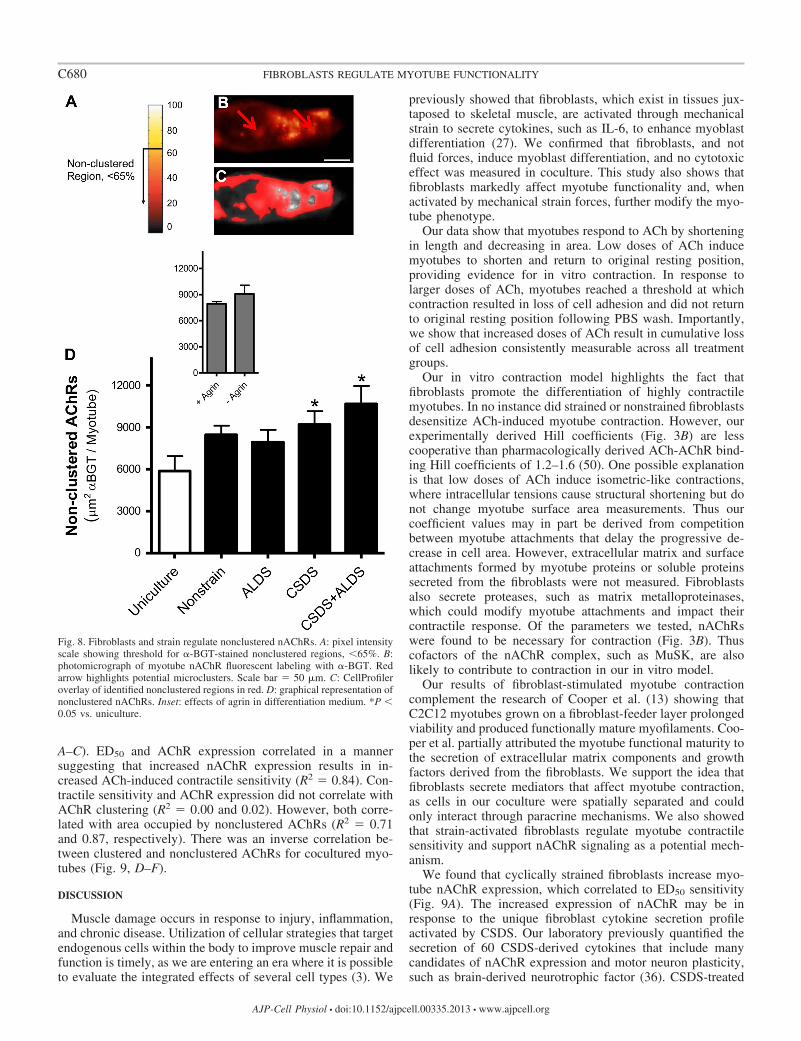
Cyclically strained fibroblasts increase nonclustered AChRregions on myotubes. nAChR clustering was puzzlingly incon-sistent, with Western blot expression data suggesting thatexclusive events were causing preexisting receptors to reorga-nize in clusters. In support of the idea, while investigatingcolocalization of nAChR clusters and myonuclei, Englanderand Rubin (19) found an inverse distribution between nAChRclusters and microaggregates. We predicted that the fluores-cence intensity of these microaggregates would fall below the�65% threshold defined by agrin-induced clustering, thusaccounting for the inconsistency between our clustering andexpression data. To show proof of concept that strain-activatedfibroblasts were capable of reorganizing nAChRs in this con-text, nonclustered regions were quantified.
Nonclustered AChRs were defined as the �-BGT-stainedarea occupied on myotubes with �65% intensity (Fig. 8, A–C).The majority (82–94%) of �-BGT staining was containedwithin these nonclustered domains for all treatments. CSDS-and CSDS � ALDS-treated fibroblasts increased the areaoccupied by nonclustered AChRs by 3,300 and 4,800 m2 permyotube vs. uniculture (Fig. 8D; P � 0.05). While nonstrainedand ALDS-treated fibroblasts also increased nonclusteredAChRs by 2,600 and 2,000 m2, respectively, vs. uniculture,these data were not significant (P � 0.06 and 0.16). Thepositive control, agrin-treated myotubes did not affect the areaoccupied by nonclustered AChRs vs. untreated myotubes(7,900 25 vs. 9,100 1,000).
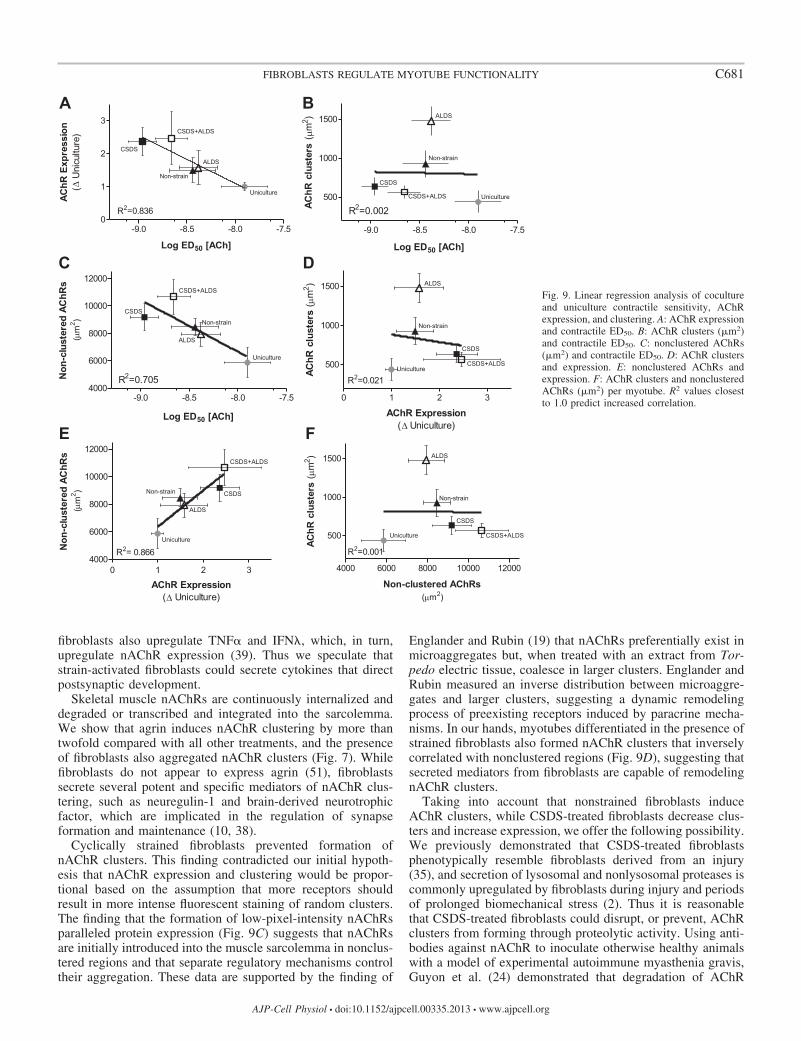
Myotube contractile sensitivity correlates with increasednAChR expression but not AChR clustering. The contractionED50 value for each coculture treatment was plotted againstAChR expression, AChR clusters, and nonclustered AChRregions, and linear regression analyses were calculated (Fig. 9,
Fig. 6. Fibroblasts and strain regulate myotube nAChR clusters. A: pixelintensity scale showing threshold for �-BGT-stained clusters, �65%. B:photomicrograph of myotube nAChR fluorescent labeling with �-BGT. Yel-low arrows highlight nAChR clusters. Scale bar � 50 m. C: CellProfileroverlay of identified clusters in yellow. D: mean nAChR cluster coverage permyotube from treatment groups. Inset: effects of agrin on nAChR clusters indifferentiation medium. *P � 0.05 vs. uniculture. �P � 0.05 vs. CSDS, TP �0.05 vs. CSDS � ALDS, �P � 0.05 vs. �agrin.
C678 FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
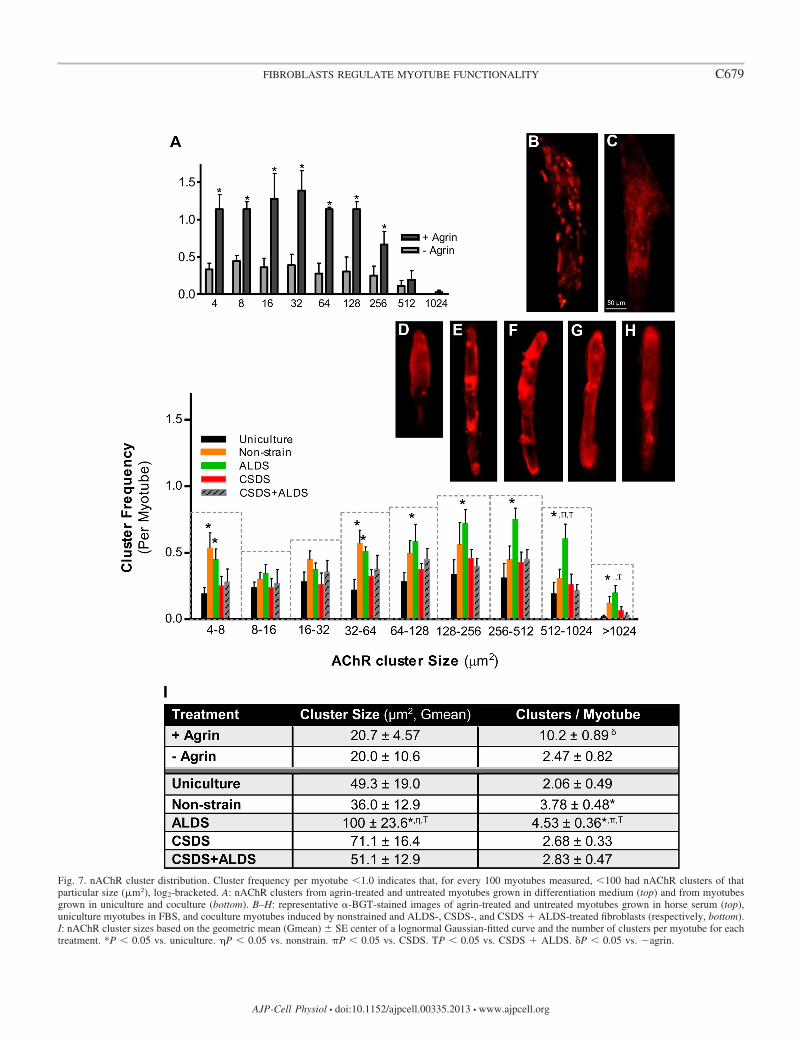
Fig. 7. nAChR cluster distribution. Cluster frequency per myotube �1.0 indicates that, for every 100 myotubes measured, �100 had nAChR clusters of thatparticular size (m2), log2-bracketed. A: nAChR clusters from agrin-treated and untreated myotubes grown in differentiation medium (top) and from myotubesgrown in uniculture and coculture (bottom). B–H: representative �-BGT-stained images of agrin-treated and untreated myotubes grown in horse serum (top),uniculture myotubes in FBS, and coculture myotubes induced by nonstrained and ALDS-, CSDS-, and CSDS � ALDS-treated fibroblasts (respectively, bottom).I: nAChR cluster sizes based on the geometric mean (Gmean) SE center of a lognormal Gaussian-fitted curve and the number of clusters per myotube for eachtreatment. *P � 0.05 vs. uniculture. �P � 0.05 vs. nonstrain. �P � 0.05 vs. CSDS. TP � 0.05 vs. CSDS � ALDS. �P � 0.05 vs. �agrin.
C679FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
A–C). ED50 and AChR expression correlated in a mannersuggesting that increased nAChR expression results in in-creased ACh-induced contractile sensitivity (R2 � 0.84). Con-tractile sensitivity and AChR expression did not correlate withAChR clustering (R2 � 0.00 and 0.02). However, both corre-lated with area occupied by nonclustered AChRs (R2 � 0.71and 0.87, respectively). There was an inverse correlation be-tween clustered and nonclustered AChRs for cocultured myo-tubes (Fig. 9, D–F).
DISCUSSION
Muscle damage occurs in response to injury, inflammation,and chronic disease. Utilization of cellular strategies that targetendogenous cells within the body to improve muscle repair andfunction is timely, as we are entering an era where it is possibleto evaluate the integrated effects of several cell types (3). We
previously showed that fibroblasts, which exist in tissues jux-taposed to skeletal muscle, are activated through mechanicalstrain to secrete cytokines, such as IL-6, to enhance myoblastdifferentiation (27). We confirmed that fibroblasts, and notfluid forces, induce myoblast differentiation, and no cytotoxiceffect was measured in coculture. This study also shows thatfibroblasts markedly affect myotube functionality and, whenactivated by mechanical strain forces, further modify the myo-tube phenotype.
Our data show that myotubes respond to ACh by shorteningin length and decreasing in area. Low doses of ACh inducemyotubes to shorten and return to original resting position,providing evidence for in vitro contraction. In response tolarger doses of ACh, myotubes reached a threshold at whichcontraction resulted in loss of cell adhesion and did not returnto original resting position following PBS wash. Importantly,we show that increased doses of ACh result in cumulative lossof cell adhesion consistently measurable across all treatmentgroups.
Our in vitro contraction model highlights the fact thatfibroblasts promote the differentiation of highly contractilemyotubes. In no instance did strained or nonstrained fibroblastsdesensitize ACh-induced myotube contraction. However, ourexperimentally derived Hill coefficients (Fig. 3B) are lesscooperative than pharmacologically derived ACh-AChR bind-ing Hill coefficients of 1.2–1.6 (50). One possible explanationis that low doses of ACh induce isometric-like contractions,where intracellular tensions cause structural shortening but donot change myotube surface area measurements. Thus ourcoefficient values may in part be derived from competitionbetween myotube attachments that delay the progressive de-crease in cell area. However, extracellular matrix and surfaceattachments formed by myotube proteins or soluble proteinssecreted from the fibroblasts were not measured. Fibroblastsalso secrete proteases, such as matrix metalloproteinases,which could modify myotube attachments and impact theircontractile response. Of the parameters we tested, nAChRswere found to be necessary for contraction (Fig. 3B). Thuscofactors of the nAChR complex, such as MuSK, are alsolikely to contribute to contraction in our in vitro model.
Our results of fibroblast-stimulated myotube contractioncomplement the research of Cooper et al. (13) showing thatC2C12 myotubes grown on a fibroblast-feeder layer prolongedviability and produced functionally mature myofilaments. Coo-per et al. partially attributed the myotube functional maturity tothe secretion of extracellular matrix components and growthfactors derived from the fibroblasts. We support the idea thatfibroblasts secrete mediators that affect myotube contraction,as cells in our coculture were spatially separated and couldonly interact through paracrine mechanisms. We also showedthat strain-activated fibroblasts regulate myotube contractilesensitivity and support nAChR signaling as a potential mech-anism.
We found that cyclically strained fibroblasts increase myo-tube nAChR expression, which correlated to ED50 sensitivity(Fig. 9A). The increased expression of nAChR may be inresponse to the unique fibroblast cytokine secretion profileactivated by CSDS. Our laboratory previously quantified thesecretion of 60 CSDS-derived cytokines that include manycandidates of nAChR expression and motor neuron plasticity,such as brain-derived neurotrophic factor (36). CSDS-treated
Fig. 8. Fibroblasts and strain regulate nonclustered nAChRs. A: pixel intensityscale showing threshold for �-BGT-stained nonclustered regions, �65%. B:photomicrograph of myotube nAChR fluorescent labeling with �-BGT. Redarrow highlights potential microclusters. Scale bar � 50 m. C: CellProfileroverlay of identified nonclustered regions in red. D: graphical representation ofnonclustered nAChRs. Inset: effects of agrin in differentiation medium. *P �0.05 vs. uniculture.
C680 FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org
fibroblasts also upregulate TNF� and IFN�, which, in turn,upregulate nAChR expression (39). Thus we speculate thatstrain-activated fibroblasts could secrete cytokines that directpostsynaptic development.
Skeletal muscle nAChRs are continuously internalized anddegraded or transcribed and integrated into the sarcolemma.We show that agrin induces nAChR clustering by more thantwofold compared with all other treatments, and the presenceof fibroblasts also aggregated nAChR clusters (Fig. 7). Whilefibroblasts do not appear to express agrin (51), fibroblastssecrete several potent and specific mediators of nAChR clus-tering, such as neuregulin-1 and brain-derived neurotrophicfactor, which are implicated in the regulation of synapseformation and maintenance (10, 38).
Cyclically strained fibroblasts prevented formation ofnAChR clusters. This finding contradicted our initial hypoth-esis that nAChR expression and clustering would be propor-tional based on the assumption that more receptors shouldresult in more intense fluorescent staining of random clusters.The finding that the formation of low-pixel-intensity nAChRsparalleled protein expression (Fig. 9C) suggests that nAChRsare initially introduced into the muscle sarcolemma in nonclus-tered regions and that separate regulatory mechanisms controltheir aggregation. These data are supported by the finding of
Englander and Rubin (19) that nAChRs preferentially exist inmicroaggregates but, when treated with an extract from Tor-pedo electric tissue, coalesce in larger clusters. Englander andRubin measured an inverse distribution between microaggre-gates and larger clusters, suggesting a dynamic remodelingprocess of preexisting receptors induced by paracrine mecha-nisms. In our hands, myotubes differentiated in the presence ofstrained fibroblasts also formed nAChR clusters that inverselycorrelated with nonclustered regions (Fig. 9D), suggesting thatsecreted mediators from fibroblasts are capable of remodelingnAChR clusters.
Taking into account that nonstrained fibroblasts induceAChR clusters, while CSDS-treated fibroblasts decrease clus-ters and increase expression, we offer the following possibility.We previously demonstrated that CSDS-treated fibroblastsphenotypically resemble fibroblasts derived from an injury(35), and secretion of lysosomal and nonlysosomal proteases iscommonly upregulated by fibroblasts during injury and periodsof prolonged biomechanical stress (2). Thus it is reasonablethat CSDS-treated fibroblasts could disrupt, or prevent, AChRclusters from forming through proteolytic activity. Using anti-bodies against nAChR to inoculate otherwise healthy animalswith a model of experimental autoimmune myasthenia gravis,Guyon et al. (24) demonstrated that degradation of AChR
-9.0 -8.5 -8.0 -7.50
1
2
3
R2=0.836
Log ED50 [ACh]
noisserpxERh
CA(
)erutlucinU
Δμ
μ
μμ
μ
μ
Uniculture
Non-strain
ALDS
CSDS
CSDS+ALDS
-9.0 -8.5 -8.0 -7.54000
6000
8000
10000
12000
R2=0.705
Log ED50 [ACh]
sRh
CAderetsulc-no
N(
m2 )
Uniculture
Non-strain
ALDS
CSDS
CSDS+ALDS
-9.0 -8.5 -8.0 -7.5
500
1000
1500
R2=0.002
Log ED50 [ACh]
AChR
clus
ters
(m
2 )
Uniculture
Non-strain
ALDS
CSDS
CSDS+ALDS
0 1 2 34000
6000
8000
10000
12000
R2= 0.866
AChR Expression( Uniculture)
sRh
CAderetsulc-no
N(
m2 )
Uniculture
Non-strain
ALDS
CSDS
CSDS+ALDS
4000 6000 8000 10000 12000
500
1000
1500
R2=0.001
Non-clustered AChRs( m2)
AChR
clus
ters
(m
2 )
Uniculture
Non-strain
ALDS
CSDS
CSDS+ALDS
0 1 2 3
500
1000
1500
R2=0.021
AChR Expression( Uniculture)
AChR
clus
ters
(m
2 )
Uniculture
Non-strain
ALDS
CSDS
CSDS+ALDS
A
C
E
B
D
F Δ
Δ
Fig. 9. Linear regression analysis of cocultureand uniculture contractile sensitivity, AChRexpression, and clustering. A: AChR expressionand contractile ED50. B: AChR clusters (m2)and contractile ED50. C: nonclustered AChRs(m2) and contractile ED50. D: AChR clustersand expression. E: nonclustered AChRs andexpression. F: AChR clusters and nonclusteredAChRs (m2) per myotube. R2 values closestto 1.0 predict increased correlation.
C681FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org

clusters is followed by a compensatory increase in nAChRexpression. Guyon et al. found that experimental autoimmunemyasthenia gravis is associated with increased TNF� andIFN�, which we have shown are secreted by CSDS-treatedfibroblasts. Thus we speculate that CSDS-mediated disruptionof nAChR cluster formation is followed by a CSDS-mediatedcompensatory upregulation of nAChR expression.
The hypothesis that nAChR cluster aggregation would cor-relate with increased contractile sensitivity, perhaps throughsynchronous depolarization, was found to be incorrect. ED50
values correlated with the expression of nonclustered AChRs.One potential source for consistency in the literature is in thefield of muscle denervation, where following the loss of inner-vation, nAChR clusters disperse across the sarcolemma andprotein expression increases (11). Bernareggi et al. (4) foundthat microtransplantation of sarcolemma into Xenopus oocytesderived from denervated muscle generated ACh currents 50times stronger than those elicited from innervated muscle.Similarly, our CSDS regimen dispersed clusters, increasednAChR expression, and hypersensitized myotube contraction(Fig. 3). We suggest that fascia fibroblasts may be one exog-enous source for noninvasive treatment options, such asthrough the targeted stretching of fibroblast-rich tissues, tosensitize nearby muscle contraction. Of clinical relevance,focal dystonias are common risk factors of repetitive straininjuries that are treated with botulinum toxin (BOTOX)-inhib-ited ACh release to stop hypersensitive muscle contraction(43). While our coculture does not contain motor neurons, it istempting to speculate about the roles of strain and fibroblasts insuch clinical settings, and we advocate the future use of animaland human models to validate our findings, which could lead tonovel therapeutics for motor dysfunction through attenuationof the fibroblast stress response.
Manual therapy techniques that apply long-duration stretchto fibroblast-rich tissues are clinically prescribed to correctsomatic dysfunction associated with musculoskeletal injuries(34) to restore muscle function (15, 31). Therefore, we werekeenly interested in applying a modeled manipulative treat-ment to fibroblasts and measuring functional outcomes inmyotubes. We show that ALDS-treated fibroblasts increasednAChR cluster number and size on myotubes (Fig. 7B). Main-tenance of the postsynaptic receptors is critical for preventingaxonopathies associated with motor neuron diseases such asamyotrophic lateral sclerosis and spinal muscular atrophy(21), as they send retrograde signals that are necessary forpresynaptic differentiation (52). We do not want to overreachon any interpretation for the significance of ALDS-treatedfibroblasts in our in vitro setting; however, it is conceivablethat just as Schwann cells and motor neurons regulate synapseformation through paracrine processes (20), fibroblasts, whichare highly adaptable to biomechanical strain and juxtaposedto nAChRs during strain-induced muscle contraction, mightalso contribute to this process. Cellular strategies to improvenAChR clustering and expression, e.g., through engraftment ofbioengineered skeletal muscle seeded with strain-activatedfibroblasts or through physical therapy regimens targetingfibroblast-rich tissue, may help neuromuscular innervation (30)and improve clinical outcomes associated with neuromusculardisorders.
While many experimental approaches are currently beingdeveloped to address the effects of multiple cell types on
muscle regeneration (6, 28, 45), our simple coculture systemmodeled after a myofascial junction shows proof of conceptthat fibroblasts are a novel cell type directing all stages ofmyoblast differentiation, including myotube contractility andnAChR organization. Our coculture has the potential to beexpanded to investigate interactions between multiple celltypes, such as fibroblast-regulated innervation of skeletal mus-cles by motor neurons, and strain-activated class switching offibroblast subpopulations, such as the platelet-derived growthfactor receptor-� and ADAM12 lineages (18). We speculatethat biomechanical strain serves as one stimulus regulating thephenotypic shift in fibroblasts that leads to pathogenesis orregeneration-favoring milieus for skeletal muscle (22) andprovide evidence that cyclically and acyclically strained fibro-blasts have profoundly different effects on nearby muscle cells.However, we examined only three strain paradigms, and werecently showed that variations to our ALDS model in strainmagnitude and duration elicit many different fibroblast re-sponses (7). We support the future use and optimization ofstrain paradigms and our coculture model to further investigatefibroblast responses that could lead to novel therapeutics forimproved skeletal muscle regeneration.
ACKNOWLEDGMENTS
The authors thank Dr. Wade Grow for helping with �-BGT labeling andDavid Campbell for helping with development of the initial CellProfileralgorithms, which formed the basis of the AChR cluster studies.
GRANTS
This work was supported by grants from the American Osteopathic Asso-ciation, the Arizona Biomedical Research Collaborative, and A. T. StillUniversity.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
M.R.H., T.V.C., and P.R.S. are responsible for conception and design of theresearch; M.R.H. and T.V.C. performed the experiments; M.R.H. and T.V.C.analyzed the data; M.R.H. and P.R.S. interpreted the results of the experi-ments; M.R.H. prepared the figures; M.R.H., T.V.C., and P.R.S. drafted themanuscript; M.R.H., T.V.C., and P.R.S. edited and revised the manuscript;M.R.H., T.V.C., and P.R.S. approved the final version of the manuscript.
REFERENCES
1. Akaaboune M, Culican SM, Turney SG, Lichtman JW. Rapid andreversible effects of activity on acetylcholine receptor density at theneuromuscular junction in vivo. Science 286: 503–507, 1999.
2. Belcastro AN, Shewchuk LD, Raj DA. Exercise-induced muscle injury:a calpain hypothesis. Mol Cell Biochem 179: 135–145, 1998.
3. Bentzinger CF, Wang YX, von Maltzahn J, Rudnicki MA. The emerg-ing biology of muscle stem cells: implications for cell-based therapies.Bioessays 35: 231–241, 2013.
4. Bernareggi A, Reyes-Ruiz JM, Lorenzon P, Ruzzier F, Miledi R.Microtransplantation of acetylcholine receptors from normal or denervatedrat skeletal muscles to frog oocytes. J Physiol 589: 1133–1142, 2011.
5. Borselli C, Storrie H, Benesch-Lee F, Shvartsman D, Cezar C, Lich-tman JW, Vandenburgh HH, Mooney DJ. Functional muscle regener-ation with combined delivery of angiogenesis and myogenesis factors.Proc Natl Acad Sci USA 107: 3287–3292, 2010.
6. Briggs D, Morgan JE. Recent progress in satellite cell/myoblast engraft-ment—relevance for therapy. FEBS J 280: 4281–4293, 2013.
7. Cao TV, Hicks MR, Campbell D, Standley PR. Dosed myofascialrelease in three-dimensional bioengineered tendons: effects on humanfibroblast hyperplasia, hypertrophy, and cytokine secretion. J Manipula-tive Physiol Ther 36: 513–521, 2013.
C682 FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org

8. Carpenter AE, Jones TR, Lamprecht MR, Clarke C, Kang IH,Friman O, Guertin DA, Chang JH, Lindquist RA, Moffat J, GollandP, Sabatini DM. CellProfiler: image analysis software for identifying andquantifying cell phenotypes. Genome Biol 7: R100, 2006.
9. Charge SB, Rudnicki MA. Cellular and molecular regulation of muscleregeneration. Physiol Rev 84: 209–238, 2004.
10. Choi W, Wolber R, Gerwat W, Mann T, Batzer J, Smuda C, Liu H,Kolbe L, Hearing VJ. The fibroblast-derived paracrine factor neuregu-lin-1 has a novel role in regulating the constitutive color and melanocytefunction in human skin. J Cell Sci 123: 3102–3111, 2010.
11. Cohen TJ, Waddell DS, Barrientos T, Lu Z, Feng G, Cox GA, BodineSC, Yao TP. The histone deacetylase HDAC4 connects neural activity tomuscle transcriptional reprogramming. J Biol Chem 282: 33752–33759,2007.
12. Collins CA, Olsen I, Zammit PS, Heslop L, Petrie A, Partridge TA,Morgan JE. Stem cell function, self-renewal, and behavioral heterogene-ity of cells from the adult muscle satellite cell niche. Cell 122: 289–301,2005.
13. Cooper ST, Maxwell AL, Kizana E, Ghoddusi M, Hardeman EC,Alexander IE, Allen DG, North KN. C2C12 co-culture on a fibroblastsubstratum enables sustained survival of contractile, highly differentiatedmyotubes with peripheral nuclei and adult fast myosin expression. CellMotil Cytoskeleton 58: 200–211, 2004.
14. Coutinho EL, DeLuca C, Salvini TF, Vidal BC. Bouts of passivestretching after immobilization of the rat soleus muscle increase collagenmacromolecular organization and muscle fiber area. Connect Tissue Res47: 278–286, 2006.
15. Dalbello-Haas V, Florence JM, Krivickas LS. Therapeutic exercise forpeople with amyotrophic lateral sclerosis or motor neuron disease. Co-chrane Database Syst Rev CD005229, 2008.
16. Deschenes MR. Motor unit and neuromuscular junction remodeling withaging. Curr Aging Sci 4: 209–220, 2011.
17. Deschenes MR, Judelson DA, Kraemer WJ, Meskaitis VJ, Volek JS,Nindl BC, Harman FS, Deaver DR. Effects of resistance training onneuromuscular junction morphology. Muscle Nerve 23: 1576–1581, 2000.
18. Dulauroy S, Di Carlo SE, Langa F, Eberl G, Peduto L. Lineage tracingand genetic ablation of ADAM12� perivascular cells identify a majorsource of profibrotic cells during acute tissue injury. Nat Med 18: 1262–1270, 2012.
19. Englander LL, Rubin LL. Acetylcholine receptor clustering and nuclearmovement in muscle fibers in culture. J Cell Biol 104: 87–95, 1987.
20. Feng Z, Ko CP. Schwann cells promote synaptogenesis at the neuromus-cular junction via transforming growth factor-�1. J Neurosci 28: 9599–9609, 2008.
21. Ferri A, Sanes JR, Coleman MP, Cunningham JM, Kato AC. Inhib-iting axon degeneration and synapse loss attenuates apoptosis and diseaseprogression in a mouse model of motoneuron disease. Curr Biol 13:669–673, 2003.
22. Flavell SJ, Hou TZ, Lax S, Filer AD, Salmon M, Buckley CD.Fibroblasts as novel therapeutic targets in chronic inflammation. Br JPharmacol 153 Suppl 1: S241–S246, 2008.
23. Grow WA, Gordon H. Acetylcholine receptors are required for postsyn-aptic aggregation driven by the agrin signalling pathway. Eur J Neurosci12: 467–472, 2000.
24. Guyon T, Wakkach A, Poea S, Mouly V, Klingel-Schmitt I, LevasseurP, Beeson D, Asher O, Tzartos S, Berrih-Aknin S. Regulation ofacetylcholine receptor gene expression in human myasthenia gravis mus-cles. Evidences for a compensatory mechanism triggered by receptor loss.J Clin Invest 102: 249–263, 1998.
25. Henriquez JP, Webb A, Bence M, Bildsoe H, Sahores M, Hughes SM,Salinas PC. Wnt signaling promotes AChR aggregation at the neuromus-cular synapse in collaboration with agrin. Proc Natl Acad Sci USA 105:18812–18817, 2008.
26. Hewett TE, Zazulak BT, Myer GD, Ford KR. A review of electromyo-graphic activation levels, timing differences, and increased anterior cru-ciate ligament injury incidence in female athletes. Br J Sports Med 39:347–350, 2005.
27. Hicks MR, Cao TV, Campbell DH, Standley PR. Mechanical strainapplied to human fibroblasts differentially regulates skeletal myoblastdifferentiation. J Appl Physiol 113: 465–472, 2012.
28. Joe AW, Yi L, Natarajan A, Le Grand F, So L, Wang J, Rudnicki MA,Rossi FM. Muscle injury activates resident fibro/adipogenic progenitorsthat facilitate myogenesis. Nat Cell Biol 12: 153–163, 2010.
29. Kietrys DM, Barr AE, Barbe MF. Exposure to repetitive tasks inducesmotor changes related to skill acquisition and inflammation in rats. JMotor Behav 43: 465–476, 2011.
30. Ko IK, Lee BK, Lee SJ, Andersson KE, Atala A, Yoo JJ. The effect ofin vitro formation of acetylcholine receptor (AChR) clusters in engineeredmuscle fibers on subsequent innervation of constructs in vivo. Biomate-rials 34: 3246–3255, 2013.
31. Kokkonen J, Nelson AG, Eldredge C, Winchester JB. Chronic staticstretching improves exercise performance. Med Sci Sports Exerc 39:1825–1831, 2007.
32. Kokkonen J, Nelson AG, Tarawhiti T, Buckingham P, Winchester JB.Early-phase resistance training strength gains in novice lifters are en-hanced by doing static stretching. J Strength Conditioning Res 24: 502–506, 2010.
33. Langhammer CG, Zahn JD, Firestein BL. Identification and quantifi-cation of skeletal myotube contraction and association in vitro by videomicroscopy. Cytoskeleton 67: 413–424, 2010.
34. Licciardone JC, Minotti DE, Gatchel RJ, Kearns CM, Singh KP.Osteopathic manual treatment and ultrasound therapy for chronic low backpain: a randomized controlled trial. Ann Fam Med 11: 122–129, 2013.
35. Meltzer KR, Cao TV, Schad JF, King H, Stoll ST, Standley PR. Invitro modeling of repetitive motion injury and myofascial release. J BodywMov Ther 14: 162–171, 2010.
36. Meltzer KR, Standley PR. Modeled repetitive motion strain and indirectosteopathic manipulative techniques in regulation of human fibroblastproliferation and interleukin secretion. J Am Osteopath Assoc 107: 527–536, 2007.
37. Murphy MM, Lawson JA, Mathew SJ, Hutcheson DA, Kardon G.Satellite cells, connective tissue fibroblasts and their interactions arecrucial for muscle regeneration. Development 138: 3625–3637, 2011.
38. Ngo ST, Cole RN, Sunn N, Phillips WD, Noakes PG. Neuregulin-1potentiates agrin-induced acetylcholine receptor clustering through mus-cle-specific kinase phosphorylation. J Cell Sci 125: 1531–1543, 2012.
39. Poea-Guyon S, Christadoss P, Le Panse R, Guyon T, De Baets M,Wakkach A, Bidault J, Tzartos S, Berrih-Aknin S. Effects of cytokineson acetylcholine receptor expression: implications for myasthenia gravis.J Immunol 174: 5941–5949, 2005.
40. Prinz H. Hill coefficients, dose-response curves and allosteric mecha-nisms. J Chem Biol 3: 37–44, 2010.
41. Ramsey SM. Holistic manual therapy techniques. Prim Care 24: 759–786, 1997.
42. Rassier DE, MacIntosh BR, Herzog W. Length dependence of activeforce production in skeletal muscle. J Appl Physiol 86: 1445–1457, 1999.
43. Rogozhin AA, Pang KK, Bukharaeva E, Young C, Slater CR. Recov-ery of mouse neuromuscular junctions from single and repeated injectionsof botulinum neurotoxin A. J Physiol 586: 3163–3182, 2008.
44. Serrano AL, Munoz-Canoves P. Regulation and dysregulation of fibrosisin skeletal muscle. Exp Cell Res 316: 3050–3058, 2010.
45. Tidball JG, Villalta SA. Regulatory interactions between muscle and theimmune system during muscle regeneration. Am J Physiol Regul IntegrComp Physiol 298: R1173–R1187, 2010.
46. Uezumi A, Ikemoto-Uezumi M, Tsuchida K. Roles of nonmyogenicmesenchymal progenitors in pathogenesis and regeneration of skeletalmuscle. Front Physiol 5: 68, 2014.
47. Valdez G, Tapia JC, Lichtman JW, Fox MA, Sanes JR. Sharedresistance to aging and ALS in neuromuscular junctions of specificmuscles. PLos One 7: e34640, 2012.
48. Vincent A. Unravelling the pathogenesis of myasthenia gravis. Nat RevImmunol 2: 797–804, 2002.
49. Vogel V, Sheetz M. Local force and geometry sensing regulate cellfunctions. Nat Rev Mol Cell Biol 7: 265–275, 2006.
50. Watty A, Methfessel C, Hucho F. Fixation of allosteric states of thenicotinic acetylcholine receptor by chemical cross-linking. Proc Natl AcadSci USA 94: 8202–8207, 1997.
51. Yang JF, Cao G, Koirala S, Reddy LV, Ko CP. Schwann cells expressactive agrin and enhance aggregation of acetylcholine receptors on musclefibers. J Neurosci 21: 9572–9584, 2001.
52. Yumoto N, Kim N, Burden SJ. Lrp4 is a retrograde signal for presyn-aptic differentiation at neuromuscular synapses. Nature 489: 438–442,2012.
C683FIBROBLASTS REGULATE MYOTUBE FUNCTIONALITY
AJP-Cell Physiol • doi:10.1152/ajpcell.00335.2013 • www.ajpcell.org









![[3H]acetylcholine synthesis in cultured ciliary ganglion neurons: Effects of myotube membranes](https://img.pdfslide.net/doc/110x75/635c6c4fa3fa66b45c0de58c/3hacetylcholine-synthesis-in-cultured-ciliary-ganglion-neurons-effects-of-myotube.jpg)



















