Embed Size (px)
Citation preview

J. Mol. Biol. (1977) 112, 317-329
Amanitin Binding to R N A Polymerase II in 0~-Amanitin-resistant Rat Myoblast Mutants
MICHAEL M. C~ERAR, STEVE J. ANDREWS, EMn~A S. DAVID DONALD G. SOMERST, JEAN-LOUIS MANDEL$ AND MARK L. PEARSON
Department of Medical Genetic, s, University of Toronto Toronto, Canada, MSS 1A8
(Received 8 October 1976)
A series of independent ~-amanit in-resistant (Area R) mutan t s have been isolated from diploid and te t raplo id strains of L6 ra t myoblas t cells. Wi th one exception, these mutan t s contain both a-amanit in-sensit ive wild-type and ~-amanit in-resistant m u t a n t R N A polymerase I I activities. The relative resist- ance of the m u t a n t enzymes in extracts of the different Ama a isolates, deter- mined by measuring the inhibit ion constant , Kl , for a-amanit in, parallels their relat ive resistance to growth inhibi t ion b y the toxin. The binding affinities of the different m u t a n t forms of R N A polymeraso I I for y-amanit in, determined by measuring the equil ibrium dissociation constaut , Kd, for y-[aH]amanitin§ bound to the enzyme, are also reduced relat ive to the wild-type enzyme, and the Kd values are proport ional to the K l values. Thus the spectrum of Ama R phenotypes in these m u t a n t s trains l ikely reflects the presence of muta t ions a t different sites in the s t ructura l gone coding for the ~-amanit in binding subuni t of R N A polymerase I I . No difference has been found between m u t a n t and wild- type forms of the enzyme with respect to their turnover numbers, Kra values for UTP, or thermal stabilit ies.
Diploid mutan t s contain equal amounts of wild-type and mu tan t forms of R N A polymerase I I . This indicates tha t t hey possess one wild- type and one m u t a n t allele for the ~-amanit in binding subuni t of the enz)rme, and t h a t both alleles are expressed in a codominant fashion. Similarly, the te t raplo id mutan t s possess three copies of the wild- type and one of the m u t a n t allele.
One te t raploid m u t a n t has been isolated which is only sl ightly resistant to growth inhibit ion by ~-amanitin and does not contain a de tec tably al tered form of R N A polymerase I I . This s t ra in m a y be defective in ~-amanitin t ransport .
1. I n t r o d u c t i o n
~-Aman i t i n is a m u s h r o o m t o x i n which acts as a specific inh ib i to r of R N A po lymerase I I in vitro (Ked inger etal., 1970; L inde l l etal., 1970). Soma t i c cell m u t a n t s r e s i s t an t to t he cy to tox i c ac t i on o f ~ -aman i t i n have been i so la ted in severa l cell s t r a ins inc luding Chinese h a m s t e r o v a r y (CHO) cells (Chan et al., I972), r a t m y o b t a s t (L6) cells (Soraers etal., 1975a), b a b y h a m s t e r k i d n e y ( B H K ) cells ( A m a t i e t a l . , 1975) a n d h u m a n
Present address : Department of Botany and Genetics, University of Guelph, Guelph, Canada N1G 2Wl.
:1: Present address: Insti tut de Chimie Biologlque, Faeulte de Medeein, 11 Rue Humann, 67085 Strasbourg, France.
§ See footnote on p. 318.
317

318 M. M. CRERAR ET AJS.
diploid fibroblast cells (Buchwald & Ingles, 1976). All these AmaRt mutants possess altered drug-resistant RNA polymerase II activities when extracts of these cells are assayed in vitro, indicating that the mutations lie in one of the structural genes for RNA polymerase II. This view is reinforced by recent studies of several CHO Area R mutants. CHO Amal cells have a temperature-sensitive RNA polymerase II (Lobban et al., 1976), and different mutants exhibiting varying degrees of resistance show corresponding decreases in the binding affinity of their enzyme for r-[3H]amanitin (Ingles et al., 1976b). Therefore, it appears that the Area ~ phenotype results from mutations in the structural gene coding for the ~¢-amauitin-binding subunit of RNA polymerase II.
Both the CHO and BHK Area R mutants contain only the mutant form of RNA polymerase II and so appear to be functionally hemizygous for the c¢-amauitin- binding subunit structural gene (Ingles et al., 1976a). In contrast, the L6 myoblast and human fibroblast Area R mutants contain both wild-type and mutant alleles of this gene. The resistant phenotype of both these latter cells indicates that the Area R mutation behaves codominantly, as shown subsequently using cell hybrids formed between Ama s cells and either CHO Areal (Lobban & Siminovitch, 1975) or BHK Ama R mutants (Amati et al., 1975).
Quite fortuitously, the presence of both wild-type and mutant forms of RNA polymerase II in L6 Areal02 (Somers et al., 1975a) has made it possible to examine the regulation of RNA polymerase II activity in vivo. Growth of this Ama R mutant in the presence of c¢-amanitin results in the inactivation of the sensitive form of RNA polymerase II, and this is followed by a compensatory increase in the resistant activity. Consequently, the net RNA polymerase II activity is maintained at a constant level (Somerset al., 1975b).
Besides ¢¢-amanitin resistance, another property distinguishes the L6 myoblast mutant Amal02 from its wild-type parent. When grown to confluence, Amal02 cells do not form multinucleate myotubes, nor do they contain the elevated levels of creatine kinase activity characteristic of wild-type myoblasts (Mandel & Pearson, 1974). I t has been suggested that this myogenesis negative phenotype may be a pleiotropic effect of the RNA polymerase II mutation (Somerset al., 1975a). However, both the regulation of RNA polymerase II and the myogenesis defect attributed to the Ama R mutation are based, on studies of a single mutant, Areal02. Therefore, in order to examine further the nature of the Ama ~ phenotype in L6 myoblasts, we have isolated a series of independent Ama R mutants with a wide spectrum of Area ~ phenotypes.
We report here the characteristics of these mutants with respect to their sensitivity to growth inhibition and RNA polymerase II inactivation by ~-amanitin, and the binding affinity of their RNA polymerase II enzymes for ~-[3H]amanitin. In addition, utilizing both diploid and tetraploid strains as parents, we have examined the effect of gene dosage on the Area R phenotype. The properties of these mutants have facili- tated studies on the mechanism of the regulation of RNA polymerase II when these mutants are grown in ~-amanitin, the results of which are reported in the accompany- ing paper (Crerar & Pearson, 1977). A preliminary account of part of this work has been presented (Ingles et al., 1976a).
tAbbreviations used: Area R, ~-amanitin-resistant; Ama s, ~,-amanitin-sensitive; Thg r, thio- guanine-resistant; y-[3H]amanitin, O-[SH]methyl.demethyl_y_amanitin.

~ - A M A N I T I N - R E S I S T A N T R A T M Y O B L A S T S 319
2. Materials and Methods
(a) Gell tinez and culture conditions
Two subelones of the ra t myoblas t continuous cell line L6 isolated by Yaffe (1968) were used as paren ta l cell lines for selection of a-amanit in-resis tant mutants . L6(H9) is a pseudotetraploid cell line, isolated as a subclone of the L6 cell line k indly provided by S. Heinemann (Salk Inst i tute) . This line is called L6 wild- type in our previous work (Somerse t al., 1975a). L6(Do) is a pseudodiploid subclone of L6 isolated b y D. Luzzat i and kindly provided by J . P. Wahrmann (Paris) (Loomis et al., 1973). (For simplici ty, we have referred to these lines as being te t raplo id and diploid, respectively, since their chromosome numbers approach 4n and 2n. I n fact, these lines and their mutan t s are aneuploid, as indicated b y their modal chromosome numbers l is ted in Table 1.) These cell lines were cultured in DME ~ 10% FCS (90~/o (v/v) Dulbeeco's modified Eagle ' s medium plus 10% (v/v) fetal calf serum (Gibco)) as described previously (Somerse t al., 1975a).
(b) Selection of a-amanitin-resistant mutants
A culture of a freshly cloned isolate of the te t raploid line L6(H9) was divided and port ions (106 cells/100+mm dish) were mutagenized separa te ly by exposure to 400 ~g e thylmethanesulfonate (Eastman)/ml in DME T 10~o FCS for a period of 24 h. This t rea tment with e thylmethanesulfonate reduces the relat ive pla t ing efficiency to 0-5. After mutagenesis, the cultures were grown for 96 h in fresh medium to allow phenotypic fixation and expression of mutat ions. The cultures were then challenged with various concentrat ions of a-amanit in (Mallinckrodt, Canada) determined b y the ahsorhance a t 305 n m (e---- 15,400 l/reel per era, -~W ~ 918, Coehet-Meilhac & Chambon, 1974). Throughout the selection, the cultures were incubated a t ei ther 37°C or 34°C, as indicated for the mutan t s l isted in Table 1. After 28 days exposure of the cells to 1 to 3 ~M+~-amanitin, during which the medium was changed every 4 to 5 days, 2 or 3 colonies from each muta- genized culture were picked, recloned and re tes ted for their ab i l i ty to grow in the con- centrat ion of a-amanit in in which they were selected. The muta t ion frequency for ~- amani t in resistance in this line has been repor ted by Seiners et al. (1975a).
A similar selection procedure was used to obta in an a-amanit in-resis tant mutan t , Area731, of the diploid line, L6(Do). I n addit ion, a thioguanine-resistant , a-amanit in- res is tant double mutan t , Thg l Area733 was selected in 2 steps from L6(Do). To select first for Thg l , cultures of L6(Do) were mutagenized as above, grown for 4 days in the absence of e thylmethanesulfonate , and then exposed to 12 ~M-6-thioguanine (2-amino- 6-mereaptopurine; Sigma) for a period of approx. 2 weeks a t 37°C. Resis tant colonies were picked and retested for their ab i l i ty to grow in 12 ~M-thioguanine. The apparen t muta t ion frequency for thioguanine resistance was 3 × 10 -e. Thg l is resistant to 18 ~M-thioguanine bu t does not survive in H A T medium (10 -4 y~-hypoxanthine, 4 × 10 -~ M-aminopterin, 1.6 × 10 -8 )[- thymidine; Littlefield, 1964), whereas L6(Do) wi ld- type cells do not survive in 3 ~ - t h i o g u a n i n e bu t grow well in H A T medium. One a-amanit in-resis tant clone, derived from Thg l and designated T h g l Area733, was chosen for fur ther characterizat ion. The apparen t muta t ion frequency for ~-amanit in resistance in these selections was 2 × 10 -6.
(c) R1VA polymerase activity assays
Cell lysates were assayed for R N A polymerase ac t iv i ty in either a high (0.4 M) or low (0"1 M) concentrat ion of ammonium sulfate to assay either R N A polymerase in flagrante transcripto or to ta l R N A polymerase act ivi ty , respectively, as described previously (Somers et al., 1975a,b).
High (0.4 ~) ammonium sulfate assays for determinat ions of R N A polymcrase ac t iv i ty in frozen and thawed cell lysates were performed a t 30°C in 120-~1 react ion mixtures using 2 ~Ci [aH]UTP (1.5 Ci/mmol; New England Nuclear). Under these conditions, R N A polyrnerase molecules are unable to reini t ia te t ranscr ipt ion (Gissinger et cd., 1974; Hossenlopp et al., 1974), so only R N A po lymerase - I )NA ~emplate-nascent R N A com- plexes formed in rive are able to direct U T P incorporation. I n addit ion, R N A polymerase

320 M. M. C R E R A R E T A L .
I and I I I ac t iv i ty is low in high salt, contr ibut ing approximate ly 20% to the to ta l R N A polymerase act ivi ty. In contrast , using the low (0" 1 M) salt assay to measure both chroma- t in-bound and free R N A polymerase in sonicated extracts , act ivi ty is dependent ( :> 90~/o) on an exogenous D N A template , and R N A polymerase I and I I I ac t iv i ty contributes 40 to 50~o to the to ta l R N A polymerase act ivi ty. I n both the low and high ammonium sulfate assays, 1 et /min corresponds to 1-2 × 10 - i s reel of [3H]UMP incorporated.
All assays for DEAE-Sephadex-pur i f ied R N A polymerase I I activit ies (Somers e$ al., 1975a,b) util ized the low ammonium sulfate assay, except tha t 6 ~Ci [3H]UTP (4.1 Ci/ nnnol) and 875 ~g bovine serum albumin/ml (Pentex, c rys ta l l ine)were added to each 120-~1 reaction mixture. DEAE-Sephadex-pur i f ied R N A polymerase I ac t iv i ty was assayed in the same way as DEAE-Sephadex-pur i f ied R N A polymerase I I act ivi ty , except assays were performed in 0.06 m-ammonium sulfate. In this assay, 1 ct /min corresponds to 0.44 × 10-15 reel of [3H]UMP incorporated.
(d) r-[3H]amanitin binding to R N A polymerass I I
Equilibri t tm binding of r-[3H]amanit in (5.4 Ci/mmol; lY[. Cochet-Meilhac, personal communication) to R N A polymerase I I in extracts of myoblas t mutan t s and wild-type cells was measured by precipi tat ion with ammonium sulfate a t 0°C, essentially as des- cribed by Cochet-Meilhac etal. (1974). Cell lysates were prepared b y 4 cycles of freezing and thawing of a cell suspension (~-~1-5 × 10 v eells/ml) in buffer C (50 mM-Tris.HC1 (pH 7-9), 0"1 mM-EDTA, 30% glycerol, plus 1 mM-dithiothreitol). Ammonium sulfate was added to the ext rac t to a final concentration of 0-3 ~. After sonicating for 10 s, 150-~1 portions of this cell lysato were added to 15-ml centrifuge tubes (Corex 8441) containing 300 ~1 buffer D (0.1 ~-ammonium sulfate, 200 ~g bovine serum albumin/ml, 400 ~g bovine ~-globulin (Pentex, fraction I I ) /ml , 3% (v/v) dimethyl sulfoxido, all in buffer C) and 50 ~l of various di luted stock solutions of r-[3H]amanitin. An equal volume of cold, sa tura ted and neutral ized ammonium sulfate was added to the reaction mixture, which was then incubated at 0°C for 23 h to allow r-[3H]amanit in binding to prec ip i ta ted enzyme to reach equilibrium. Blank values were determined b y incubat ing cell extracts plus buffer D with 10 ~1 12 ~M-unlabeled ~-amanitin (at least 100-fold excess) for 1 h at 0°C prior to the addi t ion of r-[3H]amanit in and ammonium sulfate. After 23 h incuba- tion, bound r-[ZH]amanitin was determined by centrifuging and dissolving the ammonium sulfate precipi ta te with buffer D, reprecipi tat ing with sa tura ted ammonium sulfate as above and repeat ing this washing procedure twice with blfffer D minus bovine r-globulin. The washed ammonium sulfate pellet was resuspended in 150 ~1 water, dissolved in 1.0 ml NCS (Amersham-Searle) and counted in 15 ml toluene/Onmifluor solution (15.2 g Omnifluor, New England Nuclear,/3-8 1 toluene). The to ta l (bound plus free) concentra- tions of r-[3H]amanit in added to the cell lysates were determined by adding 50 ~l of the diluted stock solutions of r-[3H]amanit in to 100 ~1 water then adding 1 ml of NCS and counting in toluene/Omnifluor. (The same p ipe t was used for addi t ion of the r-[3H]- amani t in stock dilutions to both the cell lysate and water in order to minimize addi t ion errors.) The concentrations of r-[3H]amanit in in the di luted stock solutions ranged from 10- 9 to 3.3 × 10- 8 ~ (3 × 102 to 104 cts/min per 50 ~1, based on a counting efficiency of 50~o measured using [3H]toluene (New England Nuclear) as an internal s tandard).
For determinat ion of the R N A polymerase ac t iv i ty in the sonicated cell lysates used for the binding experiments above, these lysates were made to 5 mM-MgC12 and 0-1 mM- phenylmethylsulfonylfluoride, di luted with 2 vol. buffer C made to 5mM-MgC12 and 10 -4 M-phenylmethylsulfonylfluoride, centrifuged and assayed for R N A polymerase ac t iv i ty as described above for the low ammonium sulfate assay.
3. Resul t s
(a) Selection of mutants
A series of i n d e p e n d e n t Area g m u t a n t s have been i so la ted f rom e t h y l m e t h a n e - su l fonate m u t a g e n i z e d cul tures of d ip lo id a n d t e t r a p l o i d L6 r a t m y o b l a s t cells. (Al though the lines used a re a c t u a l l y aneuplo id , t h e y a p p e a r to be d ip lo id and
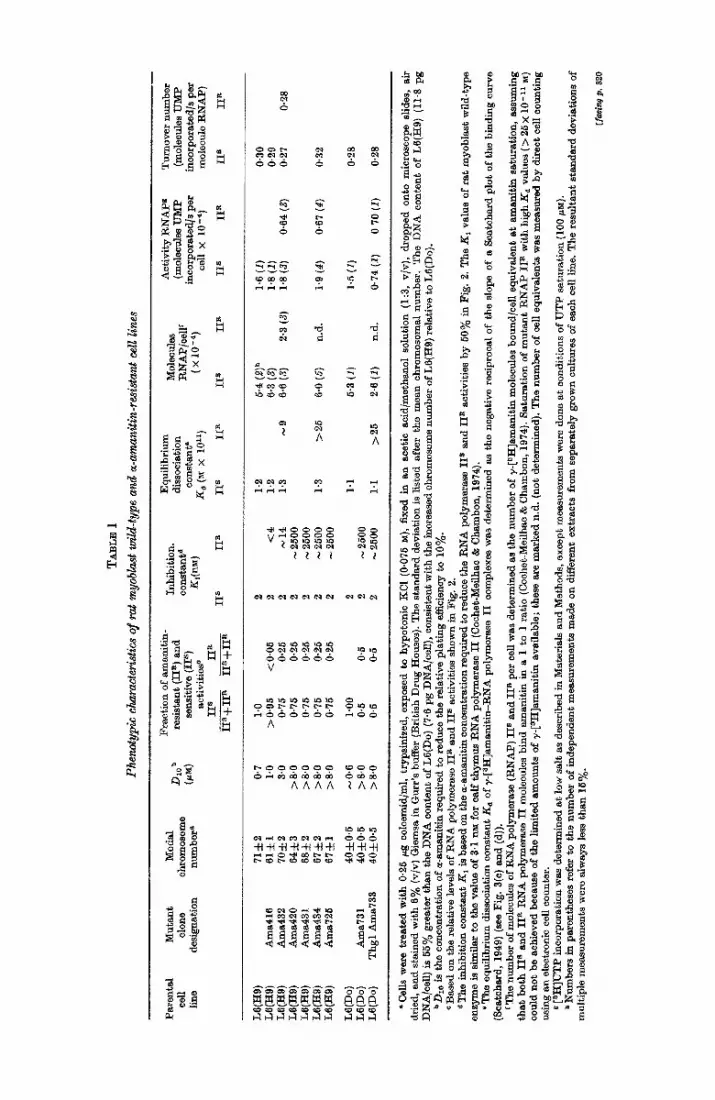
TA
BL
E 1
Phe
noty
pic
char
acte
rist
ics o
f rat
myo
blas
t wild
-typ
e an
d c~
-am
aniti
n-re
sist
ant c
ell l
ines
Par
enta
l M
uta
nt
Mod
al
coil
cl
one
chro
mos
ome
line
de
sign
atio
n n
mn
ber
~
Fra
ctio
n o
f am
anlt
in.
Eq
uil
ibri
um
M
olec
ules
D
~o b
re
sist
ant
(II"
) an
d
Inhi
biti
on.
diss
ocia
tion
R
NA
P/s
ellr
(p
~)
sens
itiv
e (I
I s)
c°n
stan
td
con
stan
t e
acti
viti
es"
Ki(
r~)
Ka
(I~
X 1
011)
( X
10
-a)
IIS
II ~
II s
II ~
IIS
II ~
II s
II R
II s
+ II r~
II s
+ I
I r~
Act
ivit
y R
NA
P K
T
urn
ov
er n
um
ber
(m
olec
ules
UM
P
(mol
ecul
es U
MP
in
oorp
orat
od/s
per
in
corp
orat
od]s
per
ce
ll ×
10
-4)
mol
ecul
e R
NA
P)
II s
H a
II
S
II ~
L0(
Hg)
7
1±
2
0'7
1.0
2 1.
2 L
6(H
9)
Are
a416
61
=1=1
1.
0 >
0.9
5
<0
-05
2
<4
1.
2 L
6(H
g)
Are
a432
70
.-I= 2
3.
0 0"
75
0"25
2
~ 14
1'
3 ~
9 L
6(H
g)
Are
a420
64
:h 3
>
8.0
0.75
0.
25
2 ~
2500
L
6(H
9)
Are
a431
6
8±
2
> 8.
0 0.
75
0-25
2
~ 25
00
L6(
Hg)
A
rea4
34
67 4
- 2
> 8.
0 0.
75
0.25
2
~ 25
00
1.3
> 25
L
6(H
g)
Are
a725
07
:~ 1
>
8-0
0-75
0-
25
2 ~
2500
L6(
Do)
40
4-0.
5 ~
0.6
1.00
2
1.1
L6(
Do)
A
rea7
31
40=}
=0.5
>
8.0
0.
5 0.
5 2
~2
50
0
L6(
Do)
T
hg
l A
rea7
33
404-
0"5
>8
.0
0.5
0-5
2 ~
25
00
1"
1 >
25
5.4
(2)h
1.
6 (1
) 0.
30
6.3
(z)
1.8
(1)
0.29
6.
0 (3
) 2.
8 (3
) 1.
8 (3
) 0.
64 (
3)
0.27
6.0
(5)
n.d.
1'
9 (4
) 0.
67 (
4)
0"32
5-3
(0
1.5
(0
o.28
2'(~
(1)
n.d.
0.
74 (
I)
0 70
(1)
0'
28
0.28
~Cel
ls w
are
trea
ted
wit
h 0
.25
~g
colc
emid
/ml,
try
psin
ised
, ex
pose
d to
hy
po
ton
ic K
CI
(0.0
75 M
), fi
xed
in
an a
ceti
c ac
id/m
eth
ano
l so
luti
on
(1:3
, v/
v),
dro
pp
ed o
nto
mic
rosc
ope
slid
es,
air
drie
d, a
nd
sta
ined
wit
h 6
% (
v]v)
Gie
msa
in
Gu
rr's
buf
fer
(Bri
tish
Dru
g H
ouse
s).
The
s~
anda
rd d
evia
tion
is
list
ed a
fter
the
mea
n c
hro
mo
som
al n
um
ber
. T
he D
NA
co
nte
nt
of L
6(H
9)
(11.
8 p
g
DN
A/c
oll)
is
55
% g
reat
er t
han
the
DN
A c
on
ten
t of
L6(
Do)
(7.
6 p
g D
NA
/col
l),
cons
iste
nt w
ith
the
inc
reas
ed c
hrom
osom
e n
um
ber
of
L6(
H9)
rel
ativ
e to
L6(
Do)
. b D
ie i
s th
e co
ncen
f~ra
tion
of
~-a
man
ltln
req
uire
d to
red
uce
the
rela
tive
pla
tin
g e
ffic
ienc
y to
10%
. B
ased
on
the
rela
tive
lev
els
of R
NA
pol
ymer
aso
II ~
an
d I
I s a
ctiv
itie
s sh
own
in F
ig.
2.
The
inh
ibit
ion
con
stan
t K
I is
bas
ed o
n th
e ~
-arn
anit
in o
on
esn
trat
ion
req
uire
d to
red
uce
the
RN
A p
oly
mer
ase
II s
and
II R
act
ivit
ies
by
60
% i
n F
ig.
2, T
he K
t va
lue
of r
at m
yo
bla
st w
ild-
type
en
zym
e is
sim
ilar
to
the
valu
e of
3.1
n~
for
cal
f th
ym
us
RN
A p
olym
eras
e II
(C
oche
t-M
eilh
ac &
Ch
amb
on
, 19
74).
e T
he e
quil
ibri
um d
isso
oiat
lon
con
sen
t K
a o
f 7-
[3H
]am
anit
in-l
q, N
A p
olym
oras
e II
co
mpl
exes
was
det
erm
ined
as
the
nega
tive
rec
ipro
cal
of t
he s
lope
of
a S
cag¢
hard
plo
t of
the
bin
ding
cur
ve
(Sca
tcha
rd,
1949
) (s
ee F
ig.
3(e)
an
d (
d)).
T
he n
um
ber
of
mol
ecul
es o
f R
NA
pol
ymer
a3e
(RN
AP
) II
s an
d Z
I s p
er c
ell w
as d
eter
min
ed a
s th
e n
um
ber
0£
y-[S
H]a
man
ltin
mol
ecul
es b
ound
[coi
l eq
uiva
lent
at
aman
ltln
sat
ura
tio
n,
assu
min
g
that
bo
th I
I s
and
II ~
RN
A p
olym
eras
o II
mol
ecul
es b
ind
am
anig
in i
n a
1 to
I r
atio
(C
oohe
t-M
oilh
ac &
Ch
amb
on
, 19
74).
Sat
ura
tio
n o
f m
uta
nt
RN
AP
II R
wit
h h
igh
Kd
valu
es (
>2
5X
10
"1~
M)
CO
Uld
no
t be
ach
ieve
d be
caus
e of
the
lim
ited
am
ou
nts
of
7-[S
H]a
man
itin
ava
ilab
le;
thes
e ar
e m
ark
ed n
.d,
(not
det
erm
ined
). T
he n
um
ber
of
cell
equ
ival
ents
was
mea
sure
d b
y d
irec
t ce
ll c
ou
nti
ng
us
ing
an e
lect
roni
c ce
ll c
ount
er.
[sR
]UT
P i
ncor
pora
tion
was
det
erm
ined
at
love
sal
t as
des
crib
ed i
n M
ater
ials
an
d l
~ot
hods
, o
xes
pt
mea
sure
men
ts w
er~
don
e at
oen
ditl
ons
of U
TP
sat
ura
tio
n (
100
tzM
). h
l~Y
umbe
rs i
n pa
rent
hese
s re
fer
to t
he
nu
mb
er o
f in
dep
end
ent
mea
sure
men
ts m
ade
on
dif
fere
nt e
xtr
acts
fro
m s
epar
atel
y g
row
n c
ultu
res
of e
ach
cel
l li
ne.
The
res
ult
ant
stan
dar
d d
evia
tion
s of
m
ulti
ple
mea
sure
men
ts w
ere
alw
ays
less
th
an 1
5%.
[fa~ ~
. 82
0

0t-AMANITIN-RESISTANT RAT i~IYOBLASTS 321
tetraploid for the gene coding for the ~-amanitin binding subunit of RNA polymerase II, as documented below. Therefore the parental lines and mutants derived from them are referred to here as being diploid or tetraploid, for convenience.) Mutants were selected for their ability to grow in the presence of various concentrations of ~-amanitin at either 34 or 37°C in order to generate a wide range of resistant pheno- types, some of which might be temperature dependent. Out of 13 independent tetraploid L6(Hg) isolates, six &ma R mutants, which represented the spectrum of phenotypes obtained, were selected for further study. In order to examine the effects of ploidy on the Area ~ phenotype, two additional mutants were selected from the diploid L6(Do) strain and its thioguanine-resistant derivative, Thgl. The designation and many of the properties of these cells and their RNA polymerase II activities, as derived from the experiments outlined below, are summarized in Table 1.
Area R mutants are designated here by three digits, the first representing the tem- perature (34°C or 37°C) at which the selection was carried out, the second the con- centration (1, 2 or 3/xM) of m-amanitin used in the selection, and the third the isolate number.
The doubling times of all the mutants are 14 to 16 hours, the same as the parental lines, and are unaffected by the presence of at least 1/x~I-m-amanitin, with one excep- tion, Area416. This latter mutant grows exponentially with a doubling time of 22 hours in 1/zM-m-amanitin, unlike either of the wild-type parents, whose growth ceases after two or three doublings under these conditions.
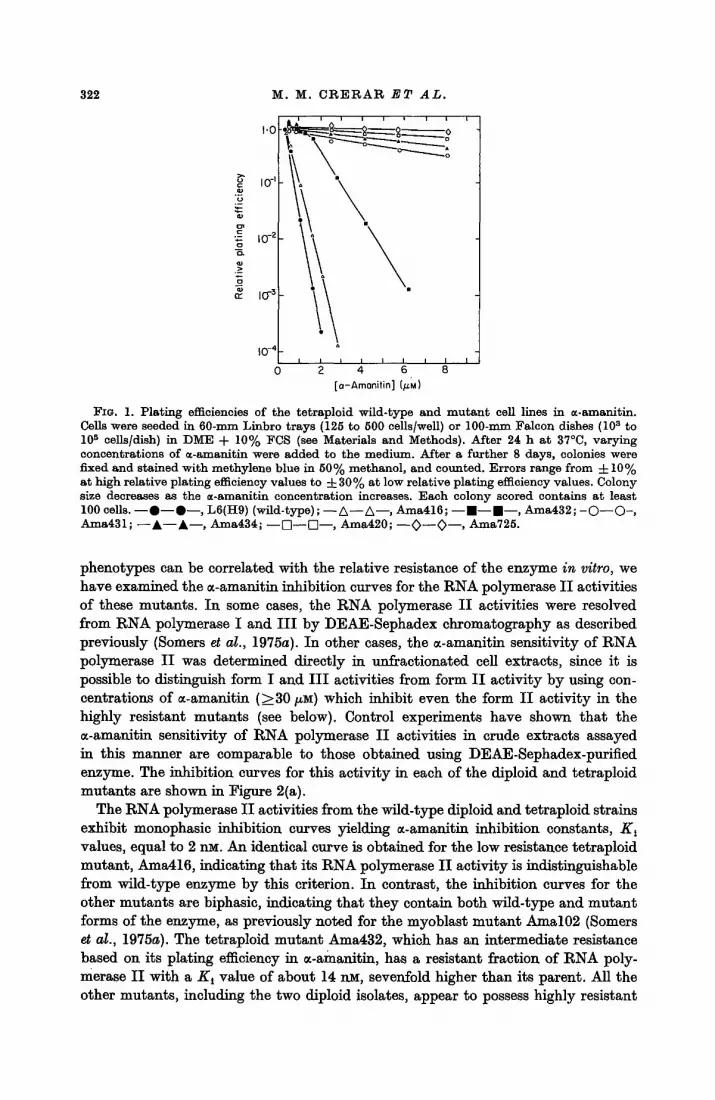
The relative plating efficiences of the different tetraploid mutants in the presence of varying concentrations of ~-amanitin are shown in Figure 1, and the D10 values estimated from these curves are listed in Table 1. The relative plating efficiences of Area416 and Ama432 are drastically reduced at concentrations of m-amanitin only slightly greater than the concentrations of m-amanitin in which they were selected (1 and 3/xM, respectively). In contrast, Area420, Ama431, Area434 and Ama725 are highly resistant to m-amanitin concentrations of 8/~M or greater, as previously noted for the original mutant Amal02 (Seiners et a/., 1975a). Based on comparisons of the ~-amanitin concentrations necessary to reduce the relative plating efficiencies of wild-type cells and these highly resistant mutants to 50%, these mutants are at least tenfold more resistant to growth inhibition by ~-amanitin than wild-type cells. The mutants, Area731 and Thgl Area733, selected from the diploid parent L6(Do), are also at least tenfold more resistant than their parent (data not shown). None of the mutants selected for their Ama R phenotype at 34°C was Area s when retested at 39°C.
It is worth noting here that mutants selected at low concentrations of ~-amanitin (1 /~M) usually exhibit the low resistance phenotype of Area416, (five out of five mutants tested), whereas mutants selected at 3 /xM-a-amanitin exhibit the high resistance phenotype of Area431 (seven out of eight mutants tested). In all eases, the Area R phenotype has bred true through at least six months continuous culture in the absence of a-amanitin.
(b) Amanitin resistant.RigA polymeraze I1
The &ma R phenotype in CHO, BHK, human fibroblast and L6 myoblast cells has been shown to be associated with alterations in the drug sensitivity of their RNA polymerase II activities. To ascertain that these new myoblast mutants also contain altered RNA polymerase II enzymes, and to determine if the speotrum of resistant
322 M. M. CRERAR E T AL.
1.0
I0-' "5
o
o
i0 -3
10-4 0
i \ \
[a-Amonitin] {/.~M I
FIG, 1. Plating efficiencies of the tetraploid wild-type and mutant cell lines in ~-amanltin. Cells were seeded in 60-ram Linbro trays (125 to 500 cells/well) or 100-ram Falcon dishes (108 to l0 s cells/dish} in DME + 10% FCS (see Materials and Methods). After 24 h at 37°C, varying concentrations of ~-amanitin were added to the medium. After a further 8 days, colonies were fixed and stained with methylene blue in 50% methanol, and counted. Errors range from 4-10% at high relative plating efficiency values to 4- 30% at low relative plating efficiency values. Colony size decreases as the ~-amanitin concentration increases. Each colony scored contains at least 100 cells. - - 0 - - 0 - - , L6(Hg) (wild-type); - - A - - A - - , Ama416; - - , - - . - - , Area432; - O - - O - , Ama431; - - A - - A - - , Ama434; - - [~--[Z-- , Ama420; - - ~ - - 0 - - , Ama725.
phenotypes can be correlated with the relative resistance of the enzyme in vitro, we have examined the ~-amanitin inhibition curves for the RNA polymerase I I activities of these mutants . In some cases, the RNA polymerase I I activities were resolved from RNA polymerase I and I I I by DEAE-Sephadex chromatography as described previously (Somerset al., 1975a). In other cases, the ~-amanitin sensitivity of RNA polymerase I I was determined directly in unfractionated cell extracts, since it is possible to distinguish form I and I I I activities from form I I act ivi ty by using con- centrations of ~-amanitin ( > 3 0 ~M) which inhibit even the form I I act ivi ty in the highly resistant mutan ts (see below). Control experiments have shown tha t the ~-amanitin sensitivity of RNA polymerase I I activities in crude extracts assayed in this manner are comparable to those obtained using DEAE-Sephadex-purified enzyme. The inhibition curves for this act ivi ty in each of the diploid and tetraploid mutan ts are shown in Figure 2(a).
The RNA polymerase I I activities from the wild-type diploid and tetraploid strains exhibit monophasic inhibition curves yielding ~-amanitin inhibition constants, Kl values, equal to 2 nM. An identical curve is obtained for the low resistance tetraploid mutant , Ama416, indicating tha t its RNA polymerase I I act ivi ty is indistinguishable from wild-type enzyme by this criterion. In contrast, the inhibition curves for the other mutants are biphasic, indicating tha t they contain both wild-type and mu tan t forms of the enzyme, as previously noted for the myoblas t mu tan t Amal02 (Somers et al., 1975a). The tetraploid mutan t Ama432, which has an intermediate resistance based on its plating efficiency in ~-amanitin, has a resistant fraction of RNA poly- merase I I with a K t value of about 14 nM, sevenfold higher than its parent. All the other mutants , including the two diploid isolates, appear to possess highly resistant

~ - A M A N I T I N - R E S I S T A N T R A T M Y O B L A S T S 323
"r-
0 ~1110 4 I~) 3 I0 ~ l~l'~O-m-~° " ~ ~1~
(e)
t I I I v 4 ~ ¢1~111 I [0 -4 I 0 -3 I 0 -2 I0 "1 I lO
[c~-Amanit in] (~M) (b)
100
FIG. 2. Inhibition by ~-amanitin of RNA polymerase I I activities from wild-type and ~-amanltin- resistant mutants grown minus and plus ~-amanitin. The tetraploid mutants Ama420, Ama431, Area434 and Ama725 were grown either minus or plus 3 p~-~-amanitin for 24 h. DEAE-Sephadex- purified RNA polymerase I I activities from these mutants were then assayed as described in Materials and Methods. The RNA polymerase activities assayed in the absence of ~-amanitin (100% levels) ranged from 400 to N 3500 cts/min [3H]LrMP incorporated above a background of about 50 cts/min. L6(H9) wild-type, Area416 and Area432 mutant cell lines were grown either minus or plus 1 ~-~-amanit in for 40 h. RNA polymerase I I activities from these 3 cell lines were measured in cell lysates at 0-4 M-ammonium sulfate as described in Materials and Methods. RNA polymerase I and I I I activities (resistant to 30/~M-~-amanitin) in these cases made up roughly 20% of the total activity assayed in the absence of ~-amanitin and these values were subtracted from the total activity values. The RNA polymerase activity assayed in the absence of ~-amanitin (100% levels) ranged from 1000 to 6000 cts/min [3H]UMP incorporated over a background of about 100 ets]min.
RNA polymerase I I activities for the diploid L6(Do) parent and the mutants Area731 and Thgl Area733 were measured in cell lysates at 0.1 M-ammonium sulfate as described in Materials and Methods. RNA polymerase I and I I I activities contributed approximately 40% of the total activity assayed in the absence of ~-amanitin and were subtracted from the total RNA poly- merase activity values. The RNA polymerase activities assayed in the absence of ~-amanitin (100% levels) ranged from 4000 to 8000 cts/min above a background of about 100 cts/min.
(a) RNA polymerase I I activity from cells grown without ~-amanitin; (b) RNA polymerase I I activity from cells grown plus --amanitin. - -C)- -C)- - , L6(H9); - - 0 - - 0 - - , Ama416; - - / k - - / k - - . Ama432; - - E l - - D - - , A m a 4 2 0 ; - - A - - A - - , Ama431; - - I - - I - - , Ama434; - - ~ ) - - 0 - - , Area725; - - ® - - ® - - , L6(Do) ; - - ~ - - ~ - - , Ama731 ; - - @ ~ @ - - , Thgl Area733.
forms of t he enzyme in add i t ion to wi ld- type enzyme. T h e K 1 values for these l a t t e r
m u t a n t s are similar, abou t 2-5 ~M, rough ly 1000-fold h igher t h a n the i r pa r en t a l K l
value.
F r o m the inhib i t ion curves shown in F igure 2(a), we h a v e e s t ima ted t h e r e l a t ive
p ropor t ion o f sensi t ive a n d res is tan t R N A polymerase I I ac t iv i ty . Such calculat ions
indica te t h a t all t he t e t r ap lo id m u t a n t s conta in rough ly 2 5 % res is tan t R N A poly-
merase I I ac t iv i ty , while t h e diploids con ta in abou t 50%, as shown in Tab le 1.
Note , however , t h a t all t h e res is tant forms of R N A polymerase I I in t h e h ighly
res is tan t m u t a n t s are even tua l l y inh ib i ted a t v e r y high ~-amani t in concent ra t ions

324 M . M . C R E R A R E T A L .
(__~10 pM), which are nevertheless still below the values required to inhibit RNA polymerase I I I (Weinmann & Roeder, 1974).
To determine if there were small differences in the sensitivity of the resistant forms of the mutant enzymes, we studied the ~-amanitin senstivities of the enzymes obtained from cells grown in the presence of ~-amanitin. Under these conditions the sensitive form should be inactivated in vivo, leaving mainly the resistant form to be titrated in vitro, so the inhibition curves should tend to become monophasic (Somers et al., 1975b). The inhibition curves for the RNA polymerase II activities of Area416, Ama432 and several highly resistant tetraploid mutants grown in this fashion are shown in Figure 2(b). There is still no discernible difference between Ama416 and L6(H9) in the ~-amanitin sensitivities of their residual RNA polymerase II activities. As expected, the inhibition curves for the intermediate resistance mutant, Ama432, and the highly resistant mutants, Ama420, Ama431, Ama434 and Ama725, are all close to monophasic and have the same K l values as the resistant form found in the same mutants grown in the absence of ~-amanitin (see Table 1). Combining extracts of mutant and wild-type cells prior to assaying their RNA polymerase activity results in strictly additive inhibitions of the wild-type plus mutant enzymes, as expected if resistance is an intrinsic property of the mutant RNA polymerase II.
The mutant enzyme activities described above have been s h o ~ by criteria other than their DEAE-Sephadex elution profiles and ~-amanitin sensitivity to be genuine form II RNA polymerases. As reported previously for Amal02 (Somerset al., 1975a), the Area s RNA polymerase activities purified on DEAE-Sephadex from Ama420, Ama431, Ama434 and Ama725 cells are inhibited to the same extent as wild-type RI~A polymerase II by 10 m~t-potassium phosphate (45%±3%), and by anti-calf thymus RNA polymerase II antiserum (50%±5%) (Ingles, 1973). This is in contrast to the findings with RNA polymerase I from both wild-type and mutant cells, where inhibition by phosphate is 90%-4-4% and inhibition by the antiserum is only 2%.
To determine if the alteration to ~-amanitin resistance of the mutant RNA poly- merase II leads to any other changes in the properties of the enzyme, their thermal stabilities and their Km values for UTP were examined. The total (mutant plus wild-type) RNA polymerase II activities isolated from the mutants, Ama420, Ama431, Ama434 and Ama725 and wild-type cells have K m values for [3H]UTP of 19±3 ~M, similar to the Km values (18±2 ~M) determined for the resistant RNA polymerase II activities isolated from these mutants after 24 hours exposure to 3 ~-~-amanitin, and also to the Km values determined for calf thymus RNA polymerase II (Gissinger et al., 1974). In addition, the thermal stabilities of the wild-type and mutant myoblast enzymes do not differ significantly, unlike the difference noted between that of the CHO Amal mutant and its parent (Lobban et al., 1976). After a preineubation for 15 minutes at 50°C, 4 3 % ± 3 % of the total and 46% ~=3% of the resistant fraction of the DEAE-Sephadex-purified RNA polymerase II activities from Ama420, Area434 and Ama725 are inactivated. Hence, the sensitive and resistant fractions of the RNA polymerase II activities from these mutants are not detectably altered in either their thermal stabilities or their K m values for UTP.
(c) Amani t in binding
In order to determine if the increased resistance to ~-amanitin inhibition of the altered RNA polymerase II activities in these mutants is the direct result of a decrease in their binding affinities for the drug (as has been observed for mutant RNA

a-AMANITIN-RESISTANT RAT MYOBLASTS 325
polymerase II enzymes isolated from a variety of CHO Ama R mutants (Ingles etal. , 1976b)), equilibrium binding of 7-[SH]amanitin to the mutant and wild-type forms of RNA polymerase II was measured in crude cell lysates. Measuring binding in crude cell lysates is a valid approach for determining the binding affinities of these enzymes for amanitin, since Cochet-•eilhac et al. (1974) have shown in a variety of eukaryotic cells that RNA polymerase II is the only component capable of binding 7-[3H]amanitin with high amnity in cell lysates. Such measurements allow estimates to be made of the number of enzyme molecules per cell, assuming a stoichiometry of one bound y-amanitin per enzyme molecule (Cochet-lVfeilhac & Chambon, 1974), and thus it becomes possible to assess the effect of ploidy on the number of sensitive and resistant enzyme molecules in these mutants. Also, when the binding measure- ments are coupled with RNA polymerase II activity measurements, it is possible to estimate the turnover number and to compare that of the wild-type and mutant forms of the enzyme.
Binding isotherms for 7-[3H]amanltin and RNA polymerase II in representative tetraploid and diploid Area R mutants are shown in Figure 3(a) and (b). The corres- ponding Scatchard plots (Scatchard, 1949) of this data are shown in Figure 3(e) and (d). For comparative purposes, y-[3H]amanitin binding to RNA polymerase II in both mutant and wild-type cell extracts has been normalized in these Figures to the total (sensitive plus resistant) RNA polymerase II activity in these extracts. (Absolute values for the binding and polymerizing activities are listed in Table 1.)
Ama416 cell tysates bind the same amount of y-3[H]amanitin at saturation per unit total RNA polymerase II activity as wild-type cell lysates and the slope of the Seatchard plot yields Ka values (~-~1.2 × 10 -11 M, Table I) that are similar to each other and to the K~ values determined for the enzymes from calf thymus, rat testis, rat liver and chick oviduct (Coehet-lVIeilhac et al., 1974) (Fig. 3(a) and (c)). Hence these data provide further evidence that there is only wild-type RNA polymerase II in Area416 cells. In contrast, Area434 cell lysates bind only 75~/o as much y-[ZH]- amanitin as wild-type cell lysates at the highest amanitin concentrations tested, and this binding component has a Kd value of 1.3 × 10 -11 M, similar to the Kd value of the wild-type cell lysate. Consequently, the sensitive form of RNA polymerase II in Ama434 appears to be the only component capable of binding y-[3H]amanitin with a high affinity, implying that the resistant form must therefore have a much lower binding affinity (higher dissociation constant) for amanitin than the wild-type enzyme. On the other hand, two binding components are discernible in the binding isotherm for Ama432, the mutant with an intermediate resistance phenotype. The resultant biphasic Seatchard plot shows one component that binds at amanitin concentrations below 0.15 nM and which has a Kd value identical to that of the wild- type enzyme. The other binds ~-[3H]amanitin at higher concentrations and has a K d value estimated to be approximately 9×10 -11 M, sevenfold higher than the wild-type Ka value. The intercepts on the abscissa of the Scatehard plots in Figure 3(e) indicate that Ama432 contains about 75~/o wild-type and 25o//o mutant enzyme, as suggested earlier by the activity measurements shown in Figure 2.
A similar analysis of the binding isotherm and corresponding Scatehard plot in Figure 3(b) and (d) for RNA polymerase II in the diploid high resistance mutant, Thgl Area733, indicates that it contains only a single detectable binding component, binding 50~/o as much y-[3H]amanitin per unit total RNA polymerase II activity as the wild-type cell lysate, with a K d value equal to its wild-type counterpart. As in

M. M. C R E R A R E T A L .
I I I
• c,
o , , 0.,o o . 2 0 0 . 2 4
326
i f I I I I I I
1.0 ~I °o-O.o.o. ~ o o t - o - • ~ • . ~ iO ~j • o
:~ o.8 2 . . . - . _ . . - . - - - - - - - - i ' - " , - - -a
O" 6 J ~ , ' -
ko.4- ~"
~ o.z
0 0.2 0.4 0.6 "8"-~ II. 0 0.2
Tota l X- [ '3H]amoni l in (nld)
(o) I I I I I - - - - I - - . . . . . I I
8
6 -
~ 4 -
m I
2 - - o
I i o 0.04 0.08
n n n U n ua u U n m n n l • •
I 0"04
7"- [3H] °moni|in bound (riM)
i i ?
/ ° "~'°'~ "°~°'~'a- °' °'°'~-g'° °--~
oo" ~o,
i I I I
0.4 0.6 0-8
0.08
(b) i I I I_.~
/
)\. -
\ , -
, \
_ I .I ~ , _ _ L 0 - 1 2 0 " i 6 0 . 2 0 0 - 2 4
{cl (d)
FIG. 3. Binding of y-[3H]amanitin to RNA polymerase I I in extracts of wild-type and mutant lines.
(a) Binding isotherms for the tetraploid wild-type and mutant lines at 0°C showing ~,-[3H]- amanitin bound at various concentrations of added amanitin relative to the maximum amount bound to cell extracts of the L6(Hg) parental line at saturating levels of amanitin (~1200 cts/min bound per 2 X 106 cell equivalents). All values of y-[aH]amanitin bound were normalized to the same total RNA polymerase I I activity (that sensitive to 30 F~[-c¢-amanitin).
(b) Binding isotherms for the diploid L6(Do) parental line and its mutant Thgl Ama733 as in (a). (e) Seatehard plot of the data in (a) for the tetraploid lines. (d) Scatehard plot of the data in (b) for the diploid lines. - - A - - A - - , L6(H9); - - O - - O - - , Ama416; - - 0 - - 0 - - , Ama432; - - A - - A - - , Ama434;
- - I 1 - - 1 1 - - , L6(Do); - - [3-- [Z--, Thgl Area733.
t he case of Area434, th is aga in impl ies t h a t t he r e s i s t an t form of R N A po]ymerase I I in T h g l Area733 has a much lower b ind ing aff ini ty for a m a n i t i n t h a n the w i l d - t y p e enzyme. These da t a , t a k e n in con junc t ion wi th t he R N A po lymerase I I a c t i v i t y d a t a s t rong ly suggest t h a t t he t e t r a p l o i d m u t a n t s of i n t e r m e d i a t e a n d high res i s tance
con ta in 75% wi ld - type a n d 25% m u t a n t enzyme, and t h e d ip lo id m u t a n t s con ta in 5 0 % wi ld - type a n d 50% m u t a n t enzyme.
E s t i m a t e s of t he n u m b e r of R N A po lymerase I I molecules pe r cell and the corres- pond ing t u r n o v e r number s for t he m u t a n t and w i ld - type enzymes based on th is b ind ing d a t a a re l i s ted in Tab le 1. The t u r n o v e r numbe r s of these enzymes a re s imi lar to those r e p o r t e d p rev ious ly for the enzyme from o the r sources (Weaver et a/., 1971; K e d i n g e r & Chambon, 1972; Sugden & Kel ler , 1973). T o t a l R N A po lyme ra se

¢ c - A M A N I T I N - R E S I S T A N T R A T M Y O B L A S T S 327
I I enzyme levels, ranging from 5× 104 to 9 × 104 molecules/cell, also agree with similar values based on binding and activity measurements in a variety of rat tissues (Weaver et al., 1971; Cochet-Meilhae et al., 1974), and immunological determinations in CHO cells (Guialis et al., 1977).
4. Discussion
We have characterized a series of L6 rat myoblast mutants exhibiting a spectrum of Area B phenotypes. These mutants have been selected in a single step following mutagenesis with ethylmethanesulfonate. By varying the concentration of ~-amanitin used in the selections, mutants with varying degrees of resistance can be isolated. Selections carried out at low ~-amanitin concentrations (1 /z~) preferentially yield mutants like Ama416 with a low resistance phenotype, while mutants selected at higher concentrations (3 /~M) tend to be resistant to much higher levels of drug (> 8/zM). Attempts to isolate mutants with a temperature-sensitive Ama B phenotype have so far been unsuccessful. This phenotype may be difficult to isolate since RNA polymerase II is a reasonably thermostable enzyme, requiring temperatures above 50°C for inactivation in vitro (Somerset al., 1975a). Therefore, single amino acid substitutions in the a-amauitin binding site may not suffice to destabilize the mutant enzyme at 40°C, the maximum temperature compatible with myoblast growth.
The present experiments establish the molecular basis of the Ama B mutation in these myoblast mutants as an alteration in the 7-amanitia binding site of I~NA polymerase II. A similar conclusion has recently been reached in studies of several CH0 Area B mutants (Ingles et al., 1976b). 1VIyoblast mutants derived from both diploid and tetraploid strains possess an ~-amanitin-resistant BNA polymerase II activity whose inhibition constant, Ki, parallels the relative resistance to growth inhibition among the different mutants. Furthermore, measurements of the equilib- rium dissociation constant, Kd, for y-amauitin bound to resistant RNA polymerase II in extracts of mutants with differing degrees of resistance show that the mutants are defective in binding, and that the K d value increases directly with the increase in Kl. The discrepancy in absolute value between Ka and K i is due to the different salt concentrations and temperatures used in the determination of these two para- meters (Cochet-Yleflhac et al., 1974). Examination of the turnover numbers, Km values for UTP, and thermolabilities of the mutant and wild-type enzymes have shown no significant difference in the catalytic properties of the mutant and wild- type enzymes. Since all the mutant enzymes are eventually inactivated at sufficiently high ~-amanitin concentrations, they all retain a partially functional ~-amanitin binding site. Perhaps mutations which completely destroy this site are lethal (in which case they would have to be dominant lethals), or perhaps we have not yet examined a sufficient number of mutants to find an RNA polymerase II unable to bind the toxin.
Ama416, the low level resistance mutant, appears to possess an RNA polymerase II activity indistinguishable from wild-type with respect to :¢-amanitin sensitivity and r-amanitin binding. This suggests that the Ama B phenotype of this mutant is due to some other defect. The simplest possibility is that Ama416 is less permeable to ~-amanitin than wild-type, so that for any given concentration of a-amanitin in the medium, the intraeellular concentration is lower ia Area416 than in wild-type cells. This would fit the observation that the concentration of a-amanitin required

328 M . M . C R E R A R E T A L .
for the inhibition of wild-type cell growth is roughly two to three orders of magnitude greater than that required to inhibit RNA polymerase I I activity in vitro, indicating tha t there is a permeability barrier blocking the free diffusion of ~-amanitin from the medium into the nucleus.
All the mutants containing resistant RNA polymerase I I also contain wild-type sensitive enzyme, based on the biphasic character of the inhibition curves and the wild-type amanitin binding component of the binding isotherms. Since the mutant phenotype is resistant, the Area R mutation is codominant in myoblasts as it is in CHO (Lobban & Siminovitch, 1975), B H K (Amati et al., 1975) and human fibroblasts (Buchwald & Ingles, 1976). In addition, the close correlation between the ploidy of these mutants and the ratios of their mutant-to-wild-type enzyme levels strongly suggests tha t these ratios simply reflect the dosage of the gene coding for the a-amanitin binding subunit. Hence, the tetraploid mutants appear to contain one mutant and three wild-type alleles, and the diploid mutants contain one mutant and one wild-type allele for this gene.
Measurements of ~-amanitin binding in crude extracts allow estimates to be made of the number of RNA polymerase I I molecules per cell. We find tha t both the wild-type diploid and tetraploid strains contain approximately the same absolute number of molecules per cell. This is surprising, since characterization of the mutants derived from these cells indicates tha t tetraploid cells contain twice the number of genes coding for the a-amanitin binding subunit than that in the diploid cells. Another surprising aspect of the binding data is the apparent increase in the total RNA polymerase IX levels (up to 65~/o in Area432) in the tetraploid mutants relative to their L6(H9) parent. An independent check on the validity of these binding deter- minations is provided by measuring the total RNA polymerase IX activity per cell, which correlates closely with the 7-amanitin binding determinations. Furthermore, the differences in the tetraploid cell lines do not appear to he due to the presence of the Ama R mutat ion per se, since the diploid mutant Thgl Ama733 seems to have the same number of enzyme molecules per cell as its Area s parent. Therefore these results may suggest tha t cellular levels of RNA polymerase I I are regulated by factors other than gene dosage (such as nuclear volume or perhaps RNA polymerase I I itself), although the trivial possibility tha t the observed variation in enzyme levels is due to subclonal variation has not been excluded.
Because of the presence of both wild-type and mutant RNA polymerase I I in Amal02, the first Ama x isolate of L6 (Somers et al., 1975a), it was possible to demon- strate tha t RNA polymerase I I activity was under regulatory control such tha t the ~-amanitin-resistant RNA polymerase I I activity level increased during growth of the mutant in the drug, compensating for the inactivation of the wild-type form. A more detailed study of this aspect of the regulation of RNA polymerase I I and the mechanism underlying this phenomenon in these new mutants is presented in the following paper (Crerar & Pearson, 1977).
We are grateful for discussions with our colleagues C. J. Ingles and L. Sirninoviteh. We thank P. Chambon, Th. Wiela~d and H. Faulstieh for a gift of ~-[3I-I]amanitin. We also thar~k C. J. Ingles for a gift of anti-calf thymus RNA polymerase II antiserum. One author (M. M. C) was supported by a postdoctoral fellowship from the Muscular Dystrophy Association of Canada. This work was supported by grants from the National Cancer Institute of Canada, the Medical Research Cotmefl of Canada and a contract from the National Cancer Institute, the United States National Institutes of Health.

~ - A M A N I T I N . R E S I S T A N T RAT MYOBLASTS 329
REFERENCES
Alnati, P., Blasi, F., Di Porzio, U., Riceio, A. & Traboni, C. (1975). Proc. Nat. Acad. Sci., U.S.A. 72, 753-757.
Buehwald, M. & Ingles, C. J. (1976). Somatic Cell Genet. 2, 225-233. Chan, V. L., Whitmore, G. F. & Siminoviteh, L. (1972). Proc. Nat. Acad. Sci., U.S.A. 69,
3119-3123. Coehet-Meilhae, M. & Chambon, P. (1974). Biochim. Biophys. Acts , 353, 160-184. Coehet-Meilhae, M., Nuret, P., 0ourvalin, J. C. & Chambon, P. (1974). Biochim. Biophy~.
Acts , 353, 185-192. Crerar, M. M. & Pearson, M. L. (1977). J . Mol. Biol. 113, 327-338. Gissinger, F., Kedinger, C. & Chambon, P. (1974). Biochimie, 56, 319-333 Guialis, A., Beatty, B. G., Ingles, C. J. & Crerar, M. M. (1977). Cell, 10, 53-60. Hossenlopp, P., Oudet, P. & Chambon, P. (1974). Eur. J . Bioohem. 41, 397-41I. Ingles, C. J. (1973). Biochem. Biophys. Res. Commun. 55, 364-371. Ingles, C. J. , Beatty, B. G., Guialis, A., Pearson, M. L., Crerar, M. M., Lobban, P. E.,
Siminoviteh, L., Somers, D. G. & Buchwald, M. (1976a). In R N A Polymerase (Losick, R. & Chamberlin, M., eds), lap. 835-853, Cold Spring Harbor Laboratory Press, New York.
Ingles, C. J., Guialis, A., Lam, J. & Siminovitch, L. (1976b). J. Biol. Chem. 251, 2729-2734. Kedinger, C. & Chambon, P. (1972). Eur. J . Biochem. 28, 283-290. Kedinger, A. C., Gniazdowski, M., Mandel, ft.-L, Gissinger, F. & Chambon, P. (1970).
Biochem. Biophy8. Res. Commun. 38, 165-171. Lindell, T. J., Weinberg, F., Morris, P. W., l~oeder, R. G. & l~utter, W. J. {1970). Science,
170, 447-449. Littlefield, J. W. (1964). Science, 145, 709-710. Lobban, P. E. & Siminovitch, L. (1975). Cell, 4, 167-172. Lobban, P. E., Siminovitch, L. & Ingles, C. J. (1976). Cell, 8, 65-70. Loom_is, W. F. Jr, Wahrmarm, ft. P. & Luzzati, D. (1973), Proc. Nat. Acad. Sci., U.S.A.
70, 425-429. Mandel, J.-L. & Pearson, M. L. (1974). Nature (London), 251,618-620. Scatchard, G. (1949). Ann. N . Y . Acad. Sci. 51, 660-672. Somers, D. G., Pearson, M. L. & Ingles, C. J. (1975a). J . Biol. Chem. 250, 4825-4831. Somers, D. G., Pearson, M. L. & Ingles, C. J. (1975b). Nature (London), 253, 372-374. Sugden, B. & Keller, W. (1973). J . Biol. Chem.. 248, 3777-3788. Weaver, l~. F., Blatti, S. P. & l~utter, W. J. (1971). Proc. Nat. Acad. Svi., U.S.A. 68,
2994-2999. Weinmann, 1~. & l~oeder, 1~. G. (1974). Proc. Nat. Acad. Sci., U.S.A. 71, 1790-1794. Yaffe, D. (1968). Proc. Nat. Acad. Sci., U.S.A. 61, 477-483.





























