Embed Size (px)
Citation preview

БЮЛЛЕТЕНЬМОСКОВСКОГО ОБЩЕСТВА
ИСПЫТАТЕЛЕЙ ПРИРОДЫ
НОВАЯ СЕРИЯ
143-й год издания
ОТДЕЛ БИОЛОГИЧЕСКИЙТом LXXVII, вып. 1
ЯНВАРЬ — ФЕВРАЛЬ
В ы ходит 6 р а з в год
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА 1972

БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXXVII (I), 1972
УДК 576.2.
БОЛЬШОЙ Ж ИЗНЕННЫ Й ЦИКЛ GALEOBDOLON LUTEUM HUDS.
О. В. Смирнова, Н. А. Торопова
Зеленчук желтый (Galeobdolon luteum Huds.) относится к весьма своеобразной группе наземно-ползучих растений, еще недостаточно изученных в онтогенетическом отношении. При исследовании большого жизненного цикла этого вида была сделана попытка сопоставить возрастные состояния и фазы морфогенеза. По нашему мнению, при таком сравнении более отчетливо выявятся следующие моменты: 1) .закономерное становление и деградация жизненной формы в ходе онтогенеза; 2) структурное своеобразие особи в период ее максимального влияния на среду; 3) место вегетативного размножения в онтогенезе и роль его в самоподдержании видовых популяций.
Материал по жизненному циклу зеленчука желтого был собран в дубравах и липняках Московской обл., в равнинных дубовых и предгорных дубово-буковых лесах Закарпатья. В жизненном цикле этого вп да выделены возрастные периоды и состояния согласно схеме, предложенной Т. А. Работновым (1950).
I. Латентный период
Описан только на основании литературных данных. Семена зеленчука созревают к июню. Семенная продуктивность, по данным Г. П. Ры синой (1968), в зависимости от типа сообщества может быть различной, в среднем около 60 семян на генеративный побег. Период покоя семян не установлен. М. М. Шик (1953) считает, что прорастание семян возможно сразу после их созревания. Есть указания о прорастании семян, несколько лет пролежавших в почве (Рысин, Рысина, 1965). В ненарушенном травяном покрове семенное возобновление у зеленчука подавлено (Кожевников, 1929). Более энергичному появлению всходов способствует удаление травяного покрова и подстилки с опытных площадок (Рысин, Рысина, 1965).
II. Виргинильный период
В начале мая в дубравах начинается массовое появление пророст ков зеленчука. Прорастание семян надземное, семядоли ассимилирующие, черешковые, пластинка их округлой формы с выемчатой верхуш-
76
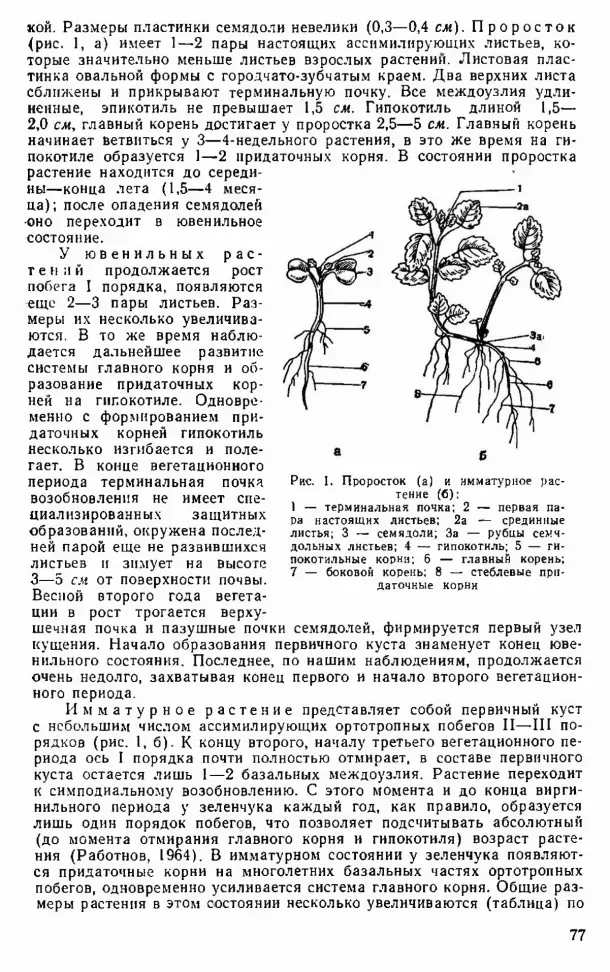
кой. Размеры пластинки семядоли невелики (0,3—0,4 см). П р о р о с т о к (рис. 1, а) имеет 1—2 пары настоящих ассимилирующих листьев, которые значительно меньше листьев взрослых растении. Листовая пластинка овальной формы с городчато-зубчатым краем. Два верхних листа сближены и прикрывают терминальную почку. Все междоузлия удлиненные, эпикотиль не превышает 1,5 см. Гипокотиль длиной 1,5— 2,0 см, главный корень достигает у проростка 2,5—5 см. Главный корень начинает ветвиться у 3—4-недельного растения, в это же время на ги- покотиле образуется 1—2 придаточных корня. В состоянии проростка растение находится до середины—конца лета (1,5—4 месяца); после опадения семядолей •оно переходит в ювенильное состояние.
У ю в е н и л ь н ы х р а с т е н и й продолжается рост побега I порядка, появляются еще 2—3 пары листьев. Размеры их несколько увеличиваются. В то же время наблюдается дальнейшее развитие системы главного корня и образование придаточных корней на гипокотиле. Одновременно с формированием придаточных корней гипокотиль несколько изгибается и полегает. В конце вегетационного периода терминальная почка возобновления не имеет специализированных защитных образований, окружена последней парой еще не развившихся листьев и зимует на высоте 3—5 см от поверхности почвы.Весной второго года вегетации в рост трогается верхушечная почка и пазушные почки семядолей, фирмируется первый узел кущения. Начало образования первичного куста знаменует конец ювенильного состояния. Последнее, по нашим наблюдениям, продолжается очень недолго, захватывая конец первого и начало второго вегетационного периода.
И м м а т у р н о е р а с т е н и е представляет собой первичный куст с небольшим числом ассимилирующих ортотропных побегов II—III порядков (рис. 1, б). К концу второго, началу третьего вегетационного периода ось I порядка почти полностью отмирает, в составе первичного куста остается лишь 1—2 базальных междоузлия. Растение переходит к симподиальному возобновлению. С этого момента и до конца вирги- нильного периода у зеленчука каждый год, как правило, образуется лишь один порядок побегов, что позволяет подсчитывать абсолютный (до момента отмирания главного корня и гипокотиля) возраст растения (Работнов, 1964). В имматурном состоянии у зеленчука появляются придаточные корни на многолетних базальных частях ортотропных побегов, одновременно усиливается система главного корня. Общие размеры растения в этом состоянии несколько увеличиваются (таблица) по
Рис. 1. Проросток (а) и имматурное растение (б):
1 — терминальная почка; 2 — первая пара настоящих листьев; 2а — срединные листья; 3 —- семядоли; За — рубцы семядольных листьев; 4 — гипокотиль; 5 — ги- покотильные корни; 6 — главный корень; 7 — боковой корень; 8 — стеблевые при
даточные корни
77

Нек
отор
ые
возр
астн
ые
пока
зате
ли з
елен
чука
— см см —о о о оо’ о" о о-н -н-н -нСО — ■Ч- см<3> оО ■'Г Оо* о* о" —*
о-н
фш О
8 Iо.ьб "
о о о+1 -н-нш ю —— СГ> ’»— о" о
о-н
о■нсмо>СО*
S 3ч- 00 соо о оо" о* о*+i +1+1со СО
Ч « СО* -см
оо*-н00
см — о* о*+1 -нсо h- со —
(N — (М (Nо ОО О Оо* О* О* о* О*+1 +Ж М +1— ЮЮ Ю СО- со-» — о см . . . .
------- см —
о+1
58S s
о о о+ 1 + 1 +1-> с~. со
оо*+СО
см смО О О +1 +1 +1S с£ осм* ь-* о*
см о о
юсо*
Ю СО о юсо" см* V —*
о о со* ю*— Ч"
юю ю со*
*58?О Юсо юсо со см ч-
ост>* оч«
— см —О 0 ^ 0
о* о о*•н -Н-Н ю.ш со со со• -о
о+1
о+ 1сосо
о о —«-н-нЧ* Ч"смо>
— — о
смо+1о
о о о оо’ о' о' о*+1 +1 +1 +1Ю Ю СО оо ' о ” . —*о
S2 2 се “з; -22 Ж ™ ° -f- *>3 3
*5 5о
роCl и(у QJ
# X 3s : х Е ф С р ю о ? о й о 5 .2 у S .u
2 ° £ « > . о . Я m ® со
<С О О О?*> qju
0J о
У Ш Ш UJЛ О О о о Л к ^XpL-XuXH R3 s u ^ и i S ™t-J-p - с-Ч
I- ^ 3яч 5с вГ О я О гSfc 8я; 'о >5
d <3 о « 5
сравнению с растениями в ювенильном состоянии. По мере появления в первичном кусте побегов III и более высоких порядков некоторые из них приобретают специфические особенности: сначала растут как ортотроп- ные, а затем по мере увеличения числа и длины междоузлий полегают на землю и укореняются. Такие побеги-плети несут черешковые листья, пластинки которых располагаются в горизонтальной плоскости. Это достигается скручиванием черешков листьев и междоузлий побега (Шик, 1953). Образование ползучих побегов мы считаем началом нового возрастного состояния — молодого вегетативного-
У м о л о д о г о в е г е т а т и в н о г о р а с т е н и я (рис. 2) из почек, развивающихся в пазухах нижних листьев полегающих плетей, образуются побеги, входящие в состав первичного куста. Верхушка плети с 2—3 парами листьев приподнята над землей и продолжает расти. Узлы средней части побега укореняются, из пазушных почек укоренившихся узлов развиваются боковые побеги. Такие побеги и кусты, возникающие на некотором расстоянии от первичного куста из почек специализированного ползучего побега, можно назвать парциальными (Серебряков и Чернышева, 1955).
Таким образом, молодое вегетативное растение зеленчука представляет собой куртину: первичныйкуст, соединенный ползучими побегами с одним или несколькими парциальными побегами или кустами. У молодого вегетативного растения продолжается рост и раз-
78

витие первичного куста. Увеличивается число и размеры боковых и придаточных корней, хорошо сохраняется главный корень. Возрастает число и размеры ортотропных побегов (таблица) и число ежегодно трогающихся в 'рост пазушных почек.
Уже в молодом вегетативном состоянии начинается вегетативное размножение зеленчука. Дочерние парциальные кусты разрастаются.
Рис. 2. Молодое вегетативное растение:1а — верхушечная почка побега IV порядка; римские цифры — порядок побе
гов. Остальные обозначения те же, что н на рис. 1
развивают свою собственную корневую систему и при разрушении пла- гиотропной плети (связующего звена между материнским и дочерними кустами) переходят полностью к самостоятельному существованию, формируя клон. Первичный куст при этом не отмирает. Продолжая энергично нарастать, он в дальнейшем переходит к образованию генеративных побегов.
Таким образом, в виргинальном периоде заканчивается формирование основных структурных черт зеленчука желтого. Общая продолжительность этого периода составляет в среднем 4—5 лет (таблица).
111. Генеративный период
Ц и к л р а з в и т и я п о б е г а (малый жизненный цикл). По данным М М. Шик (1953), растения зеленчука в генеративном периоде имеют два типа побегов: генеративные и вегетативные. Полный цикл развития наземно-ползучего (Высоцкий, 1906) вегетативного побега взрослого растения состоит из ортотропной и плагиотропной фаз.
Среди вегетативных побегов зеленчука М. М. Шик (1953) выделила побеги весенней и летней генераций. Побеги, начавшие расти орто- тропно рано весной, к середине лета превращаются в плагиотропные плети. Из пазушных почек этих плетей к концу вегетационного периода развиваются ортотропные побеги, почки возобновления которых зиму
79

ют на высоте 10 см и более над уровнем почвы. Часть из них формирует соцветие, остальные весной продолжают расти, удлиняются и превращаются в плагиотропные плети.
Как показали наши наблюдения, рост побегов у зеленчука отличается большим своеобразием и не укладывается в схему, предложенную М. М. Шик (1953). Ниже рассматриваются возможные варианты роста
J
%% ? % f
1
чу /-
Й Б, Б, Б 3
* \\
✓ „ v|L1 ' JL / / / 0-0П
А В, В , В 3
а ~> б
Рис. 3. Образование дициклического, полициклического, дикарпического побегов:I — побеги летней генерации; II — побеги весенней генерации; а — многолетние части побега; б — однолетние; пунктир — отмершие листья и соцветия; сплошная линия — живые; штриховка — уровень почвы. Оостальчые обозначения в тек
сте. Пазушные почки н придаточные корни не обозначены
и расположения побеговг зеленчука по отношению к поверхности почвы. Рост всех побегов начинается ортотропно. У побегов летней генерации ортотропный рост продолжается до конца вегетационного периода. У части этих побегов в конце лета закладывается соцветие. Осенью генеративные побеги хорошо отличаются от вегетативных довольно крупной рыхлой почкой на верхушке. Почка содержит почти сформированное соцветие. Никаких специальных защитных образований, кроме сильно опушенных верхних листьев, почка не имеет и в таком состоянии перезимовывает. Спорогенез происходит в ранневесенний период или непосредственно перед распусканием бутона (Шик, 1953). Весной полностью развившийся генеративный побег имеет 8—9 междоузлий, 3—4 верхних узла несут мутовки по 6 цветков каждая. Нижние листья черешковые, прицветники сидячие. Судьба этого побега может быть различной. 1. После созревания семян отмирает вся часть генеративного побега выше первого — второго базального узла с почками возобновления. Этот случай описан М. М. Шик (1953) (рис. 3, Bt). 2. После созревания семян терминальная почка генеративного побега трогается в рост.
80

Образуется вегетативная часть побега, состоящая из 7 и более междоузлий. Весь побег полегает и укореняется, давая начало новым парциальным побегам и кустам. Рост возникшего таким образом плагиотроп- ного побега-плети продолжается в течение 1-го, редко 2-го вегетационного периода. После этого терминальная почка или отмирает, или часть побега с терминальной почкой изгибается, принимает ортотропное положение и снова переходит к цветению (рис. 3, Б2-з). Дальнейшую судьбу такого побега нам не удалось проследить, так как на второй-третий год жизни плети происходит отделение ее от материнского побега в области первых пяти—семи базальных междоузлий. Через 3—4 года после образования плети от нее остаются лишь небольшие участки в непосредственной близости от узлов, давших начало новым парциальным образованиям.
Побеги весенней генерации к середине лета полегают, превращаясь в плечи. В следующий вегетационный период то же самое происходит с побегами летней генерации, которые не переходят к цветению. В дальнейшем часть этих побегов некоторое время нарастает верхушкой, сохраняя плагиотропное положение. Рост продолжается 1—2 вегетационных периода, после этого терминальная почка отмирает (рис. 3, Bi). Этот вариант побегов у зеленчука описан М. М. Шик (1953). Другие плагиотропные побеги изменяют направление роста. Верхушка побега изгибается, принимает ортотропное положение и через некоторое время побег переходит к цветению (рис. 3, В2).
В дальнейшем либо верхушечный рост полностью прекращается, либо после созревания семян терминальная почка снова трогается в рост, и побег переходит к плагиотропной фазе роста (рис. 3, В3). Вскоре после этого базальные междоузлия плети разрушаются и, таким образом, становится невозможным проследить дальнейшую судьбу побегов.
Изучение циклов развития побегов различных генераций показало, что у зеленчука кроме дициклических монокарпических побегов можно выделить полициклические ди-, а возможно, и поликарпические побеги.
В пределах парциального куста побеги возобновления развиваются из пазушных почек первого-второго базальных узлов побегов предыдущих порядков. Базальные части побегов, остающиеся в пределах парциального куста, формируют своеобразное симподиально нарастающее корневище, подобное корневищу парциальных кустов Carex pilosa Scop., Orobtis vcrnus L. (Серебряков, Серебрякова, 1965).
На плагиотропных плетях боковые побеги могут развиваться из пазушных почек на протяжении всей плети за исключением ее генеративной части. Возобновление плагиотропных побегов в течение 2—3 лет происходит моноподиально. Так как побеги возобновления у зеленчука появляются в ннжней и средней частях материнского побега (как орто- тропного, так и плагиотропного), а также в верхней части материнского плагпотропного побега, у этого вида возможно акро-, мезо- и базисим- подиальное возобновление (Гатцук, 1967).
Рост пазушных почек может происходить при разной степени сфор- мироЕзнности материнского побега. При этом возможны следующие варианты: 1) почки одного узла или разных узлов одного побега трогаются в рост одновременно; 2) одна из почек узла трогается в рост, другая в течение нескольких лет остается в покоящемся состоянии, а уже затем формирует побег; 3) почки разных узлов одного и того же побега трогаются в рост разновременно. Довольно часто у зеленчука наблюдается образование силлептических побегов. Развитие почек наблюдается в течение большей части вегетационного периода (Шик,
6 Бюллетень биологии, вып. 1 81

1953). поэтому выделение побегов весенней и летней генерации в некоторой степени условно.
Образование более одного порядка побегов в год в пределах одного парциального куста чрезвычайно затрудняет определение возраста взрослых растений. Если допустить, что на побеге весенней генерации побеги следующего порядка появляются только в конце лета — осенью, а ветвление этих побегов начинается следующей весной, то в год сме-
Рис. 4. Схема онтогенеза парциального куста:1 — парциальный побег; I I — парциальный куст 1—2-го года жизни (А — образовавшийся из побега летней генерации, Б — из побега зесенней генерации); III — парциальный куст 3—4-го года жизни; IV — парциальный
куст 4 и более лег
няется два порядка побегов. Условный возраст при этом будет равен половине числа порядков побегов. В случае более энергичного ветвления и образования силлептических побегов в год может появиться 3— 4 порядка побегов. По нашим наблюдениям, наиболее часто у зеленчука сменяется два порядка побегов в год, поэтому при подсчете возраста число порядков побегов делилось на два.
Растение становится способным к плодоношению на 5—б-й год жизни. В группу генеративных особей могут входить куртина, семенной куст, возникший при ее распаде, но основную массу составляют парциальные образования. Быстрое разрушение связи между материнским и дочерними парциальными образованиями приводит к большому разнообразию в структуре генеративных особей и усложняет классификацию их по возрастным состояниям. Формирование особей разной структуры прослеживается при изучении о н т о г е н е з а п а р ц и а л ь ног о куста .
Начало парциальному кусту дает ортотропный побег, развивающийся из пазушной почки плети (рис. 4, I). Если это побег весенней генерации, то к концу лета он превратится в плеть. Одновременно из его базальных пазушных почек образуются новые ортотропные побеги, которые входят в состав куста (рис. 4, II, Б). Побег летней генерации, дающий начало парциальному кусту зацветает весной, а из пазушных почек базальных узлов появляются ортотропные побеги (рис. 4. II, А).
82

Парциальные кусты двух-трех лет имеют в своем составе несколько ортотропных вегетативных и генеративных побегов и плагиотропных побегов-плетей. В это время наблюдается образование максимального числа новых парциальных побегов и кустов. У всех парциальных кустов этого возраста имеется хорошо развитая корневая система и большой запас почек возобновления. Парциальные кусты четырех-пятилетнего возраста обычно имеют только один генеративный побег. Плагиотроп- ные побеги уже не образуются, число почек возобновления резко уменьшается (рис. 4, IV). Следующим этапом обычно является отмирание парциального куста.
По мощности развития (числу и размерам ассимилирующих побегов, числу и размерам листьев, наличию почек возобновления и растущих побегов, по числу и степени развития генеративных побегов) в генеративном периоде выделено три возрастных состояния: молодое, средневозрастное и старое.
К м о л о д ы м г е н е р а т и в н ы м относятся в первую очередь растения семенного происхождения, с хорошо сохранившейся системой главного корня. Они представлены либо куртиной, либо частью куртины с первичным кустом, от которой отделились парциальные побеги и кусты. Первичный куст состоит из небольшого числа ассимилирующих побегов. в нижней части которых развиваются побеги возобновления. Генеративный побег (V—VII порядка), как правило, один. К молодым генеративным относятся и растения, возникшие вегетативным путем: это могут быть парциальные кусты на первых этапах их онтогенеза и системы парциальных кустов и побегов 1—3 лет. По количеству и размерам генеративных и вегетативных побегов такие растения подобны особям семенного происхождения.
С р е д н е в о з р а с т н ы е г е н е р а т и в н ы е р а с т е н и я представлены мощно развитыми одиночными парциальными кустами и системами парциальных кустов и побегов 3—4-летнего возраста. Иногда до средневозрастного генеративного состояния доживает первичный куст. Средневозрастные особи имеют наибольшее число ассимилирующих побегов; большая часть ортотропных является генеративными. В течение вегетационного периода возможно образование двух и более генераций плагиотропных побегов. Большое число и быстрый рост их способствует энергичному распространению вида в ценозе. Длина побегов, число пар и размеры листьев в этом возрастном состоянии достигают максимума (таблица). Среди средневозрастных растений явно выделяются 2 группы: 1) растения с трогающимися в рост почками и молодыми только начавшими рост побегами; 2) без таковых.
В первом случае к следующему вегетационному периоду не произойдет заметного изменения жизненности и возрастного состояния особи: на смену отмирающим побегам придут выросшие побеги возобновления. Во втором случае будущее особи может быть различно: а) отсутствие побегов возобновления может быть временным явлением, связанным с неблагоприятными условиями существования, тогда на следующий год из спящих почек могут развиться новые побеги и растение удержится на прежнем уровне; б) отсутствие побегов возобновления может быть признаком перехода к следующему возрастному состоянию и говорить о старении особи. Кроме того, возможно отмирание особи без перехода в старое генеративное состояние.
С т а р ы е г е н е р а т и в н ы е р а с т е н и я представлены парциальными кустами большого собственного возраста. Эти кусты заканчивают цикл развития, они лишены плагиотропных побегов — растение теряет способность захватывать новые территории. У старых генеративных рас-
6* 83

тений заметно сокращается число вегетативных и генеративных побегов в кусте, уменьшаются их размеры. У генеративных побегов сокращается число цветочных мутовок в соцветии и цветков в мутовках, у вегетативных— размеры и количество листьев (таблица). В этом возрастном состоянии почти полностью отсутствуют тронувшиеся в рост почки и молодые побеги.
В популяции зеленчука часто встречаются растения, по числу и размерам вегетативных побегов соответствующие средневозрастным генеративным. Такие особи рассматривались нами как временно нецветущие средневозрастные особи генеративного периода. По наличию или отсутствию молодых побегов эти растения были разделены на группы, аналотчные выделенным среди цветущих средневозрастных особен. Вр емснно нецветущих, соответствующих молодому генеративному состоянию. не обнаружено. Все нецветущие старые особи отнесены к старым вегетативным растениям (сенильный период).
IV. Сенильный период
Включает два возрастных состояния: старое вегетативное и сенильное.
С т а р ы е в е г е т а т и в н ы е р а с т е н и я — небольшие парциальные кусты с ортотропными ассимилирующими побегами (рис. 5, а). По
размерам и числу листьев эти побеги можно сравнить с таковыми у имматурных растений. Число придаточных корней невелико. Пазушные почки почти полностью отсутствуют. У старых вегетативных и сенильных растений каждый год образуется один порядок побегов, поэтому условный возраст равен числу порядков побегов.
С е н и л ь н ы е р а с т е ния имеют один, реже два ортотропных побега, которые по размерам и количеству листьев почти не отличаются от побегов ювенильных растений (рис. 5, б). Условный возраст таких растений очень мал (2—3 года), корневая система состоит из нескольких слабо ветвящихся коротких придаточных корней. Корневище находится на поверхности почвы. Как уже отмечалось выше, при переходе от генеративного периода к сенильному растение полностью те
ряет способность к образованию плагиотропных плетей и перестает размножаться вегетативным путем. Наблюдать деление парциального
Рис. 5. Старое вегетативное (а) и сенильное (б) растения:
9 — базальные основания побегов предыдущих лет. Остальные обозначения те же,
что и на рис. 1 и 2
84

куста на части, способные к самостоятельному существованию, нам ни разу не приходилось.
В работе использована периодизация морфогенеза, предложенная рядом авторов (Серебряков и Чернышева, 1955; Серебряков, 1965; Гат- цук, 1967; Шафранова, 1967; Иванова, 1968 и др.). В морфогенезе G. luteuin мы выделили следующие фазы: I — моноподиально нарастающий одноосный побег; II — первичный куст; III — куртина; IV — клон.
I. Ф а з а о д н о о с н о г о м о н о п о д и а л ь н о н а р а с т а ю щ е г о п о б е г а включает следующие возрастные состояния: проросток и юве-
Рис. 6. Схема фаз морфогенеза зеленчука. Обозначения в тексте
пильное (рис. 6). Проросток может быть выделен как отдельная фаза морфогенеза, так как у некоторых видов он является совершенно особым образованием. В разбираемом случае проросток представляет собой одноосный побег так же, как и ювенильное растение, отличаясь от последнего только наличием семядолей и числом пар срединных листьев. Продолжительность пребывания растения в фазе одноосного побега невелика, в среднем один год.
II. Ф а з а п е р в и ч н о г о к у с т а более продолжительна. Начинается формирование первичного куста со второго года жизни зеленчука, когда из пазушных почек семядолей развиваются побеги II порядка. Эта фаза соответствует нмматурному возрастному состоянию и продолжается обычно 2 года.
III. Фа з а к у р т и н ы начинается с момента укоренения первых плагиотропных побегов-плетей и возникновения из их пазушных почек парциальных побегов. Она соответствует молодому вегетативному и, по-видимому, части молодого генеративного состояния. Продолжительность этой фазы 2—3 года.
IV. Ф а з а к л о н а начинается в молодом вегетативном — молодом генеративном состояниях и охватывает весь генеративный и сенильный периоды. Она господствует в жизненном цикле зеленчука желтого (рис. 6). На протяжении этой фазы происходит окончательное формирование всех структурных особенностей разбираемого вида, а затем постепенное упрощение структуры в процессе деградации жизненной формы.
Минимальная продолжительность фазы клона 7—10 лет. Реальная продолжительность значительно больше и, по нашим представлениям, составляет несколько десятков лет. Увеличение продолжительности фазы клена обусловлено большей протяженностью генеративного периода
85

в жизненном цикле зеленчука. В серии сменяющих друг друга особей, которые возникают на протяжении генеративного периода, длительное время не происходит изменения возрастного состояния (старения). Последнее и приводит к господству в популяциях генеративных особей.
Учитывая относительную длительность фазы клона и многообразные структурные изменения, которые происходят с особью в это время, мы попытались подразделить эту фазу на три этапа, которым соответствуют особи различного строения: 1) система парциальных кустов или побегов; 2) взрослый парциальный куст; 3) парциальный куст с побегом ювенильного типа. Так как связь между соседними парциальными кустами у зеленчука разрушается очень быстро, системы парциальных побегов или кустов можно обнаружить лишь в том случае, когда интенсивность образования ползучих побегов очень велика. Это наблюдается в средневозрастном генеративном состоянии, когда растение достигает наибольшей вегетативной мощи. Одиночные парциальные кусты преобладают, с одной стороны, когда растение еще не достигло максимальной сложности структуры (молодое генеративное состояние), а с другой — в процессе постепенного упрощения формы (старое генеративное — старое вегетативное состояние).
В возобновлении популяций зеленчука желтого большую роль играет вегетативное размножение. Оно происходит на протяжении большей части фазы клона, наиболее интенсивно в средневозрастном генеративном состоянии. При отделении парциальных образований возможны следующие случаи: 1) дочерние кусты и побеги принадлежат к той же возрастной группе, что и материнский куст; 2) дочерние кусты и побеги не равноценны по возрастному состоянию материнскому кусту: они Moiут быть более молодыми или более старыми. Частным случаем этого варианта является образование сенильных парциальных побегов на плагиотропных побегах генеративных растений.
Образование двух и более генераций плагиотропных побегов в год приводит к чрезвычайно быстрому захвату территории: скорость распространения особи составляет в среднем 1 м за вегетационный период. Это значительно больше, чем скорость распространения таких вегетативно подвижных видов синузии дубравного широкотравья, как Сагех pilosa Scop, и Aegopodium podagraria L. (Шик, 1953; Любарский, 1967). При чрезвычайно быстром захвате территории продолжительность существования парциального куста невелика — в среднем 3 года, в то время как парциальные кусты осоки волосистой и сныти произрастают на захваченной территории не менее 6—8 лет (Смирнова, 1967).
Малая продолжительность жизни парциальных образований п быстрое разрушение связей между ними приводит к тому, что в ненарушенных ценозах зеленчук почти никогда не образует сплошных зарослей, вытесняясь другими видами.
Проведенное исследование позволяет заключить, что по особенностям структурных изменений особи в ходе большого жизненного цикла, по продолжительности фаз морфогенеза, по быстроте захвата поверхности и относительной устойчивости особи на занятой территории зеленчук желтый может быть отнесен к особому биологическому и фито- ценотическому типу. Этот вид можно охарактеризовать как неустойчивый доминант, имеющий черты эксплерента.
Л И Т Е Р А Т У Р А
В ы с о ц к и й Г. Н. 1906. Почвенио-ботаиическне исследования в южных тульскихзасеках. «Тр. опыты, лесничих», вып. 4. СПб.
86

Г а т ц у к Л. Е. 1967. Морфогенез копеечника кустарникового прн постоянном уровне песчаного субстрата. В сб.: «Онтогенез и возрасти, состав популяций цветк. растений». М„ «Наука».
И в а н о в а И. В. 1968. Морфогенез жизненной формы в роде Rubus L. Автореф. канд. дне. М.
К о ж е в н и к о в А. В. 1929. Материалы к фитосоциологической характеристике 38 кв. Погонно-Лосинного острова. «Тр. по леей, опыти. делу ЦЛОС», вып. 6. М.
Л ю б а р с к и й Е. Л. 1967. Экология вегетативного размножения высших растений. Казань.
Р а б о т н о в Т. А. 1950. Жизненный цикл многолетних травянистых растений в л у говых ценозах. «Тр. БИНа», сер. III. геобот., вып. 6. М.— Л., Изд-во АН СССР.
Р а б о т н о в Т. А. 1964. Определение возрастного состава популяций видов в сообществе. В сб.: «Полевая геоботаника», т. 3. М.— Л., «Наука».
Р ы с н н Л. П., Р ы с п н а Г. П. 1965. Почвенный запас семян травянистых растений в лесу п факторы, влияющие на их прорастание. В сб.: «Леса Подмосковья». М., «Наука».
Р ы с и н а Г. П. 1968. К характеристике семенной продуктивности некоторых лесных травянистых растений. В сб.: «Сложи, боры хвойно-широколиств. лесов и пути ведения лесн. хоз-ва в лесопарк, условиях Подмосковья». М., «Наука».
С е р е б р я к о в И. Г. 1965. Экологическая морфология растений. М., «Высшая школа».
С е р е б р я к о в Н. Г., С е р е б р я к о в а Т. И. 1965. О двух типах формирования корневищ у травянистых многолетников. «Бюл. МОИП», отд. биол., т. LXX, вып. 1.
С е р е б р я к о в И. Г., Ч е р н ы ш е в а М. Б. 1955. О морфогенезе жизненной формы кустарничка у черники, брусники и некоторых болотных Ericaceae. «Бюл. МОИП», отд. биол., т. LX, вып. 2.
С м и р н о в а О. В. 1967. Онтогенез и возрастные группы осоки волосистой (Сагех pilosa Scop.) н сныти обыкновенной (Aegopodium podagraria L.). В сб.: «Онтогенез н возрасти, состав популяций цветк. растений». М., «Наука».
Ш а ф р а н о в а Л. М. 1967. Морфогенез и жизненная форма лапчатки мелколистной (Potentilla parvifolia Fisch.) в связи с переходом от кустарников к травам у лапчаток (Potentilla L. s. I.). В сб.: «Онтогенез и возрасти, состав популяций цветк. растении». М., «Наука».
Ш н к М. М. 1953. Сезонное развитие травяного покрова дубрав. «Уч. зап. МГПИ им. Ленина», т. 73, вып. 2, каф. ботаники.
THE GREAT LIFE CYCLE OF GALEOBDOLON LUTEUM HUDS.
О. V. Smirnova, N. A. Toropova
Summary
The investigations were carried out in the foothill oak-beech forests of the Transcarpathians and the oak forests of the Moscow region. The ontogenetic stages recognised according to the scheme of T. A. Rabotnov (1950) were compared with the morphogenetic phases. Features of exle- rentness is appeared in G. luteum as it is evident from its structural and rhvthmological singularities, rapid invasion and instability of individuals on the invaded territory.




















