Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Bryozoans, corals, and microfacies of Lower Eifelian(Middle Devonian) limestones at Kierspe, Germany
Andrej Ernst • Andreas May • Stephan Marks
Received: 29 June 2011 / Accepted: 13 December 2011 / Published online: 20 January 2012
� Springer-Verlag 2012
Abstract The Lower Eifelian Meinerzhagener Korallen-
kalk (= upper Cultrijugatus Beds) at Kierspe, Sauerland,
contains a rich reefal fauna. Eight bryozoan species are
described, two of them are new: the cystoporate Fistuliporella
kierspensis n. sp. and the trepostome Leptotrypella sophiae n.
sp. The bryozoans from the Meinerzhagener Korallenkalk
shows distinct similarities to the Lower–Middle Devonian of
Spain (Santa Lucıa Formation, Emsian–Eifelian), and to the
Middle Devonian (Eifelian) of Transcaucasia. The coral fauna
comprises five tabulate corals and one rugose coral that
document a paleobiogeographic relationship between Central
and Eastern Europe and Central Asia. The associated fauna is
represented by brachiopods, ostracods, and echinoderms. The
studied limestones also commonly contain calcimicrobes
represented by three species. The faunal and microfacial
characteristics indicate a shallow marine depositional envi-
ronment just above the storm wave base, with a supposed
depth of 20 m, within the photic zone. The nutrient regime was
at least a mesotrophic. The upper boundary of the Cultrijug-
atus Beds coincides with the Chotec-Event that strongly
affected brachiopods, whereas corals and bryozoans were
insensitive to this event.
Keywords Middle Devonian � Germany � Microfacies �Bryozoa � Corals � Brachiopods � Calcimicrobes �Taxonomy � Paleoecology � Paleobiogeography
Introduction
The regional geology, stratigraphy, and paleontology of the
Middle Devonian sequences of the northern Rhenish
Massif (Eifel, Bergisches Land, and Sauerland) have been
investigated for many decades. As a result of this extended
research, there are many paleontological, stratigraphical
and regional geological publications (e.g., Richter 1971;
Grabert 1980; Langenstrassen 1983; Meyer 1986; Koch
et al. 1990; Meyer and Stets 1996; Ribbert 1998; Weddige
and Ziegler 2000; May 2003). Paleontological publications
include numerous studies of invertebrates such as bra-
chiopods (e.g., Solle 1953, 1971; Struve 1964, 1982a,
1992; May 1989, 1991; Gad 1995, 1997; Thormann and
Weddige 2001), stromatoporoids (May 1993d; Krebedun-
kel 1995; Salerno 2008), rugose and tabulate corals (e.g.,
Birenheide 1979, 1985; Byra 1983; Coen-Aubert and Lutte
1990; Lutte 1987, 1990, 1993; May 1993 a, b; Avlar and
May 1997; Schroder 1998; Bruhl 1999; Schroder and
Salerno 2001; Stadelmaier et al. 2005), trilobites (e.g.,
Struve 1996; Basse and Lemke 1996; Basse 1998, 2002,
2003, 2006, 2010) and gastropods (e.g., Heidelberger and
Koch 2005; Heidelberger 2003, 2008).
The current knowledge of bryozoan diversity in the
Devonian of the Eifel area was originally restricted to a few
papers in the last century (e.g., Solle 1937, 1952, 1968;
Toots 1951; Krausel 1953, 1957, 1981), but it has recently
increased (Ernst and Schroder 2007; Ernst 2008a, b; Ernst
and Bohaty 2009; Ernst et al. 2011b), Paleozoic bry-
ozoans belong mainly to the Class Stenolaemata and are
A. Ernst (&)
Institut fur Geowissenschaften der
Christian-Albrechts-Universitat zu Kiel,
Ludewig-Meyn-Str. 10, 24118 Kiel, Germany
e-mail: [email protected]
A. May
C/Canada 69, 5 B, 28850 Torrejon de Ardoz, Madrid, Spain
e-mail: [email protected]
S. Marks
Dauvemuhle 49, 48159 Munster, Germany
e-mail: [email protected]
123
Facies (2012) 58:727–758
DOI 10.1007/s10347-011-0289-6

often diverse and abundant in different marine sediments
from Ordovician to Permian time. Due to their stable cal-
cite skeletons, bryozoans usually show better preservation
than many other animals, which makes them an important
fossil group. They are significant in regional stratigraphy,
paleobiogeography, and paleoecology. Bryozoans can be
especially useful in the interpretation of past environments,
because they are sessile organisms strongly dependent on
environmental conditions. Bryozoans are abundant and
diverse in the Devonian worldwide (Cuffey and McKinney
1979). This period was a time of important changes in the
structure and global composition of bryozoan faunas
(Bigey 1985).
This paper is a study of representative material from the
Meinerzhagener Korallenkalk from the limestone quarry at
Kierspe in the western Sauerland, Germany (Fig. 1). The
Meinerzhagener Korallenkalk is a facies of the upper part of
the Cultrijugatus Beds and belongs to the Lower Eifelian.
The Cultrijugatus Beds are equivalent to the Hohenhof
Formation of northern Sauerland. The UTM coordinates
of the studied quarry are r 3402658.71, h 5666076.91 to
r 3402660.03, h 5666087.76. All investigated material was
collected in the quarry during excavating a new access to
the Hulloch, a locally famous cave (Marks 2010).
The study includes an overview of the macrofauna and
microflora and provides microfacies analysis as well as
systematic descriptions of the bryozoans and corals. The
results obtained from the bryozoan faunal analysis can thus
be compared with those from the coral fauna. A recon-
struction of the paleoenvironment of this horizon is given,
stratigraphic correlation and paleobiogeographic connec-
tions are also discussed.
Geological setting
The Rhenish Massif is generally assigned to the Avalonia
terrain, which become separated from Gondwana in the
Early Ordovician and drifted northwards (e.g., Oncken et al.
2000; Tait et al. 2000; Romer and Hahne 2010). Avalonia
collided with Baltica in the Late Ordovician—Early Silurian,
which led to the closure of the Tornquist Sea. Due to the
collision of Baltica and Avalonia with Laurentia, the Iapetus
Ocean was closed and as a result of this Laurussia was
formed (e.g., Linnemann et al. 2008; Kroner et al. 2007;
Nance et al. 2010). According to Kroner et al. (2007),
Sanchez Martınez et al. (2007), and Nance et al. (2010), the
closure of the Rheic Ocean began in the Late Silurian—Early
Devonian and continued until the Early Carboniferous by
successive closing from west to east.
Marine Devonian sediments in Europe can be generally
assigned to two facies settings, the ‘‘Rhenish’’ facies and
the ‘‘Hercynian’’ facies (Erben 1962). The Rhenish facies
is characterized by thick siliciclastic successions, which are
regarded to have been deposited in deltaic shallow marine
environments (e.g., Stets and Schafer 2002; Wehrmann
et al. 2005). Typical sediments of the ‘‘Hercynian’’ facies
are pure limestones and argillaceous shales; sandstones are
rare. Beginning in the Early Devonian, the Rhenohercynian
Basin developed as a narrow (about 250–350 km wide) but
rather elongate (more than 2,000 km) trough south of
the Old Red Sandstone Continent. Towards the south, the
trough was confined by the Mid-German High during the
Early Devonian (Lochkovian and Pragian). Detrital sedi-
ment was delivered from northern (Wierich 1999) as well as
southern source areas (Hahn 1990; Hahn and Zankl 1991).
The Lower Devonian sequences in the Rhenish Massif are
dominated by sandstones and siltstones, which were rapidly
deposited in sedimentary troughs and on swells (e.g., Meyer
and Stets 1980, 1996; Mittmeyer 1982) that formed during a
rift stage. In the southern and western Rhenish Massif,
Middle Devonian successions are predominantly composed
of shales, platy limestones and reef limestones. In contrast,
within Bergisches Land and Sauerland, siliciclastic sedi-
ments were delivered from the north, i.e., from the Old Red
Sandstone Continent (Langenstrassen 1982, 1983; May
2003) during the Lower Devonian and early Middle Devo-
nian (Eifelian and Lower Givetian). This siliciclastic input
diminished during Givetian time when shallow subtropical
carbonates were established over much of the region. The
occurrence of biostromal and biohermal limestones in the
Bergisches Land and Sauerland can be directly correlated
with sea level changes (see May 2003: 60–61, fig. 3): Sea-
level rises pushed back the siliciclastic sedimentation and
permitted the growth of reef builders.
The study area lies within the southwestern part of the
Ebbe saddle and comprises sediments of Emsian to Eifelian
age (Fig. 2). The outcrop (a limestone quarry in Kierspe)
exposes deposits of the so-called Meinerzhagener Koral-
lenkalk that belongs in the upper Cultrijugatus Beds (Avlar
and May 1997).
The Cultrijugatus Beds are slightly more marine time
equivalents of the Hohenhof Formation of northwestern
Sauerland. The Cultrijugatus Beds comprise a ca. 200-m-
thick succession of greenish, yellowish or yellowish-grey
marly or sandy argillites, Upper Emsian and Lower Eif-
elian age, and can be divided into three parts (Fig. 2).
Carbonate lenses occasionally are developed in the middle
and upper part, whereas the lower part contains rare
sandstones and gray-blue argillites. Biostromal or bioher-
mal limestones and fossiliferous marlstones within the
upper Cultrijugatus Beds of the geological map sheet
Meinerzhagen have been named ‘‘Meinerzhagener Koral-
lenkalk’’ by Fuchs (1923).
The Emsian/Eifelian boundary occurs within the middle
part of the Cultrijugatus Beds, and is constrained because
the lower part of the middle Cultrijugatus Beds contains a
728 Facies (2012) 58:727–758
123

spiriferid brachiopod fauna that suggests an Upper Emsian
age (Avlar and May 1997: 106); whereas the upper part of
the middle Cultrijugatus Beds contains the Eifelian index
fossil Arduspirifer intermedius (Schlotheim, 1820) (Solle
1953; Struve 1982b; Avlar and May 1997). Furthermore, a
fauna from the upper part of the middle Cultrijugatus Beds
documented by May (1989: 16) also suggests a Middle
Devonian age. The Cultrijugatus Beds overlie the siliciclastic
Remscheid Formation (Emsian) and are covered by the
Hobracke Formation (higher Lower Eifelian), which repre-
sents a succession of mixed carbonate and siliciclastic
deposits. The transition between the Cultrijugatus Beds and
4˚
4˚
6˚
6˚
8˚
8˚
10˚
10˚
12˚
12˚
50˚ 50˚
52˚ 52˚
54˚ 54˚
s’-Gravenhage
Brussels
Antwerpen
Rotterdam
Amsterdam
Kierspe
KölnBonn
Koblenz
Frankfurt
Bremen
Kassel
Hannover
Hamburg
Kiel
Maastricht
0 50 100
km
Düsseldorf
0 50 100
m
Quarry
Fig. 1 Geographic position of
the quarry near Kierspe,
Germany (map sketch created at
http://www.aquarius.geomar.de/
omc/make_map.html; local map
based on an illustration pro-
vided by � Markischer Kreis,
Katasterbehorde 11-04615 with
approval of Markischer Kreis on
8th June 2011, Az FD
67-Kundenzentrum)
Facies (2012) 58:727–758 729
123

the Hobracke Formation is defined by detailed lithological
differences (Cultrijugatus Beds with more greenish-yellowish
shales and Hobracke Formation with more siltstones and fine-
grained sandstones) and the disappearance of Paraspirifer
cultrijugatus (Roemer, 1844), Paraspirifer sandbergeri Solle,
1971 and Alatiformia alatiformis (Drevermann, 1907) (Avlar
and May 1997).
Materials and methods
Bryozoans, corals, and sediment were investigated using
thin-sections studied with a transmitted light binocular
microscope. Fifty-five thin-sections of different sizes
(28 mm 9 48 mm and 50 mm 9 50 mm) were made from
five representative rock samples and two coral colonies
collected from the outcrop. The thin-sections were used for
bryozoan and coral taxonomy and microfacies study. The
material is housed at the Senckenberg Museum (Frankfurt
am Main, Germany) under collection numbers SMF
21.385–SMF 21.499, SMF 21.683–SMF 21.684.
The terminology of Hill (1981) is used for the corals.
The corallite diameter is measured from the dark median
suture of the outer wall of one side to the other side. The
common wall thickness represents the complete thickness
of the both outer walls of neighboring corallites. For the
corallite diameter and wall thickness, 15–20 measurements
were taken in each case. Given are the observed minimum,
maximum and mean values.
For bryozoans, the morphologic character terminology
is adopted from Anstey and Perry (1970) for trepostomes
and from Hageman (1993) for cryptostomes. The spacing
of structures is measured as the distance between their
centers. Statistics were summarized using arithmetic mean,
sample standard deviation, coefficient of variation, and
minimum and maximum values.
Macrofauna (corals and brachiopods)
The Meinerzhagener Korallenkalk within this studied sec-
tion contains abundant tabulate corals represented by the
dominant species Pachyfavosites vilvaensis (Sokolov,
1952) (Fig. 3a, b, i, 4d, e) and Coenites vermicularis
(M‘Coy, 1850) (Fig. 3h, i, Fig. 4f, g), and the less common
species Thamnopora angusta Lecompte, 1939 (Fig. 3c–f).
Fig. 2 Stratigraphy and lithofacies of the Cultrijugatus Beds in the southwestern Ebbe saddle (modified after Avlar and May 1997)
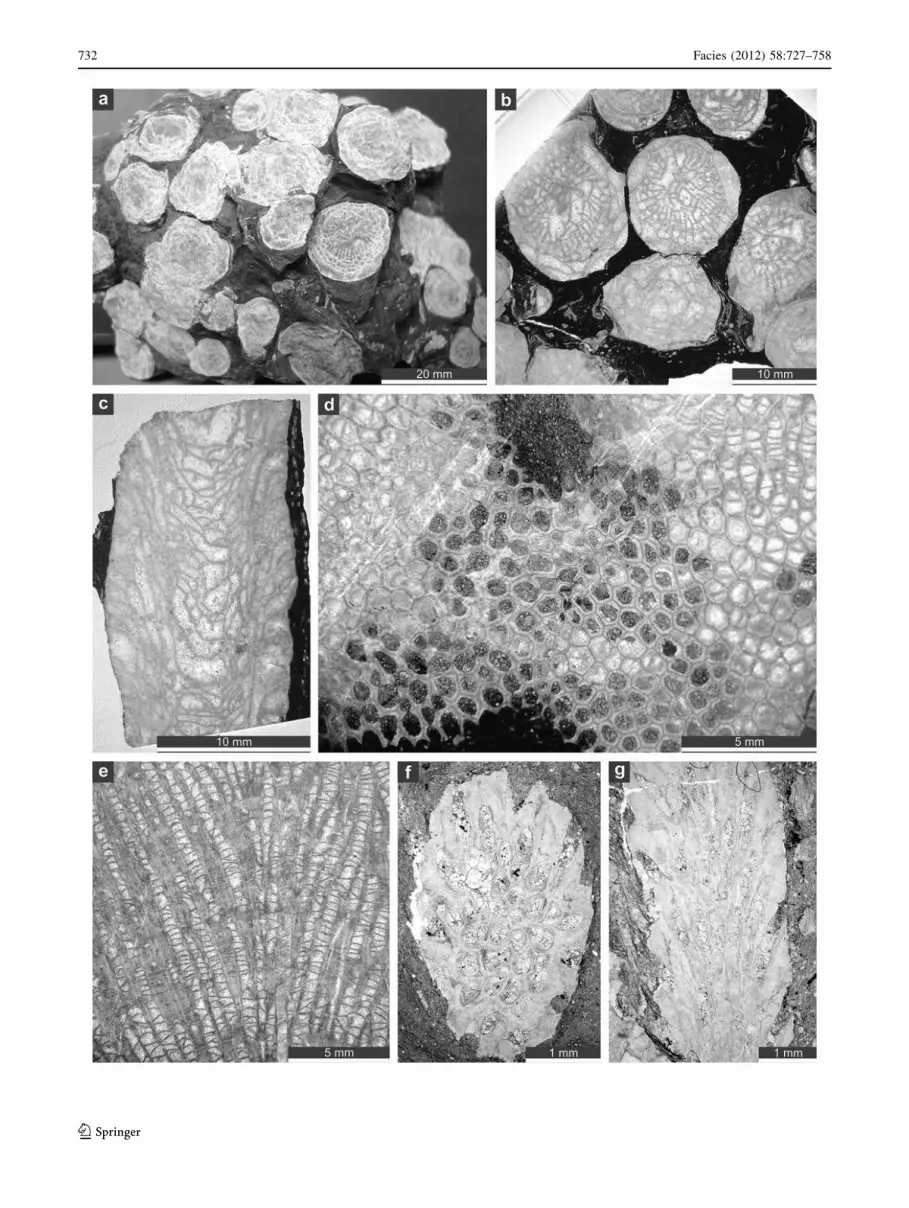
Fig. 3 Macrofauna (corals and brachiopods). Pachyfavosites vilva-ensis (Sokolov, 1952). a External view. b Exposed broken side of the
colony with ‘‘ragged’’ margins within rock sample. c–f Thamnoporaangusta Lecompte, 1939. c Thin-section of the encrusting basal part of
a colony, SMF 21.488. d Broken branch of colony within rock sample.
e Several branches of a colony embedded in rock, transverse view.
f Thin-section of a branch SMF 21.489. g Platyaxum (Roseoporella)
gradatum (Lecompte, 1939), SMF 21.683. h Colony of Roemerolitestenuis (Schluter, 1885), stabilizing the sediment, a bafflestone to
floatstone composed of branches of Coenites vermicularis (McCoy,
1850), SMF 21.684. i Bafflestone to floatstone containing isolated
corallites of Sociophyllum semiseptatum rolfwerneri Birenheide, 1979,
abundant branches of Coenites vermicularis (McCoy, 1850), and an
irregularly laminated colony of Pachyfavosites vilvaensis (Sokolov,
1952). j Alatiformia alatiformis (Drevermann, 1907)
c
730 Facies (2012) 58:727–758
123

Facies (2012) 58:727–758 731
123
732 Facies (2012) 58:727–758
123

Platyaxum (Roseoporella) gradatum (Lecompte, 1939)
(Fig. 3g) and Roemerolites tenuis (Schluter, 1885)
(Fig. 3h) are rather rare species here. Rugose corals are
represented by the relatively rare Sociophyllum semisept-
atum rolfwerneri Birenheide, 1979 (Figs. 3i, 4a–c) and
rare, small, horn-like solitary corals.
The common brachiopod Alatiformia alatiformis
(Drevermann, 1907) (Fig. 3j) was collected from marly
argillites at the top of the Meinerzhagener Korallenkalk at
this locality, along with rare specimens of the brachiopod
Xystostrophia sp.
Microfacial characteristics of the limestones at Kierspe
Limestones from Kierspe display different types of mi-
crofacies. The most common microfacies is represented by
crinoid-coral-bryozoan floatstones (Fig. 5a). Furthermore,
autochthonous limestones are represented by coral-bryo-
zoan bafflestones (Fig. 5b), algal-bryozoan bindstones
(Fig. 6d–g), and locally by pure bryozoan bindstones
(Figs. 7d, e, 8a, b). Bryozoan bindstones are produced by
encrusting cystoporates (Fistuliphragma gracilis, Fistu-
lporella kierspensis) and trepostomes (Leioclema passita-
bulatum and two unidentified species).
Among the corals, Coenites vermicularis (M‘Coy, 1850),
Thamnopora angusta Lecompte, 1939 and Sociophyllum
semiseptatum rolfwerneri Birenheide, 1979 were bafflers,
whereas Pachyfavosites vilvaensis (Sokolov, 1952), Platy-
axum (Roseoporella) gradatum (Lecompte, 1939) and
Roemerolites tenuis (Schluter, 1885) (Fig. 3h) were binders.
Besides abundant bryozoans, corals and crinoid fragments,
the following components were identified in thin-sections:
brachiopod shells, thick-walled ostracods (Fig. 5d), echi-
noid spines (Fig. 5c), and rare trilobites (Fig. 5e).
The most common calcareous alga is Girvanella weth-
eredii Chapman 1908 that occurs in about 30% of thin-
sections. This alga produces locally significant masses and
sheets 0.12–0.35 mm in thickness (Fig. 5f–j), preferen-
tially occurring on the surface of the tabulate corals Coe-
nites vermicularis (M‘Coy, 1850). Girvanella wetheredii
develops masses consisting of small, densely spaced tubes
(tube diameter: 0.005–0.0075 mm; 10–20 tubes per
0.1 mm distance). Another species, Girvanella problem-
atica Nicholson and Etheridge, 1878, is rather rare,
developing loose tubes 0.010–0.015 mm in diameter.
Rothpletzella munthei (Rothpletz, 1913) Wood, 1948 is
the rarest species, observed here in only three thin sections.
This species was found encrusting an undescribed trepos-
tome bryozoan and Acanthoclema parvula (Fig. 6d–g).
Rothpletzella munthei produces unilaminar encrusting
sheets consisting of tubes 0.025–0.050 mm in diameter,
spaced 22–27 per 1 mm distance.
The microfacies of the Meinerzhagener Korallenkalk is
similar to the microfacies of the limestones of the
Grunewiese Member of the Ihmert Formation in north-
western Sauerland (May 1992a, 1993d), which are earliest
Givetian in age (Schone et al. 1998). Furthermore, the coral
fauna of the Grunewiese Member shows several similarities
(May 1993a, b), and Girvanella wetheredii Chapman 1908,
Girvanella problematica Nicholson and Etheridge, 1878 and
Rothpletzella munthei (Rothpletz, 1913) Wood, 1948 have
been described by May (1992b) from the limestones of the
Grunewiese Member. At first glance it seems that the
Meinerzhagener Korallenkalk contains more bryozoans and
fewer stromatoporoids than the Grunewiese Member.
Remarkably, in this case the same trend is evident, which
May (2005) observed comparing Pragian and Middle
Devonian limestones of Bohemia. May (2005: 234)
suggested that an increase in the water temperature from the
Lower Devonian to the Middle Devonian could possibly
explain these variations. Further investigations must be
undertaken in order to prove whether this could be also an
explanation for the observed differences between the
Meinerzhagener Korallenkalk and the Grunewiese Member.
Systematic paleontology
Phylum Cnidaria Hatschek, 1888
Class Anthozoa Ehrenberg, 1834
Subclass Tabulata Milne-Edwards and Haime, 1850
Order Favositida Wedekind, 1937
Family Favositidae Dana, 1846
Genus Pachyfavosites Sokolov, 1952
Pachyfavosites: Sokolov, 1952: 43; Hill 1981: 548;
Birenheide 1985: 67; May 1993a: 123; Lutte 1993: 61;
Bruhl 1999: 39; Stadelmaier et al. 2005: 18; Niko 2007: 66;
Hubmann and Gaetani 2007: 314; May 2008: 3; Niko and
Senzai 2010: 41
Type species: Calamopora polymorpha var. tuberosa
Goldfuss 1826. Middle Devonian, Eifel (Germany).
Diagnosis and occurrence: See Birenheide (1985: 67) and
May (1993a: 123).
Pachyfavosites vilvaensis (Sokolov, 1952)
Figs. 3a, b, 4d, e
Fig. 4 Corals. Sociophyllum semiseptatum rolfwerneri Birenheide,
1979. a External view of complete colony. b Transverse thin-section
of several corallites of the colony, SMF 21.494. c Longitudinal
section of a corallite, SMF 21.492. d, e Pachyfavosites vilvaensis(Sokolov, 1952). d Tangential thin-section, SMF 21.491. e Longitu-
dinal thin-section, SMF 21.490. f, g Coenites vermicularis (McCoy,
1850). f Branch, vertical to oblique thin-section, SMF 21.498.
g Branch longitudinal thin-section, SMF 21.498
b
Facies (2012) 58:727–758 733
123

734 Facies (2012) 58:727–758
123

*1952 Favosites (Pachyfavosites) vilvaensis n. sp.
Sokolov: 48, pl. 11, figs. 1-4
1959 Pachyfavosites vilvaensis—Dubatolov: 59-60, pl.
14, figs. 3-4
2003 Pachyfavosites ex gr. cronigerus (Orbigny, 1849)
—May: 51
Material: One large corallum (Fig. 3b) cut in pieces with
2 thin-sections (Fig. 4d, e), and several smaller coralla
(SMF 21.490–SMF 21.491).
Description: The coralla are irregularly massive, tabular,
bulbous, or encrusting. The upper surface of the corallum is
flat or with dome-like excrescences (Fig. 3a).
In transverse thin-section corallites are polygonal with
rounded corners. The corallites are 0.6–1.4 mm in diameter
(mean 1.0 mm). The common wall between the corallites is
0.15–0.50 mm thick (mean 0.28–0.30 mm) with a well-
developed dark median suture. Rarely small, short septal
spines occur. No squamulae have been observed.
In longitudinal thin-section, mural pores are round,
0.15–0.25 mm in diameter and ordered in one row in the
middle of the side. Mural pores have a distance of about
1.1–1.5 mm from each other. Among the horizontal
skeletal elements tabulae dominate, which are horizontal
or slightly inclined. Inclined or curved tabellae occur
frequently. The number of horizontal skeletal elements
amounts to 24–30 per 10 mm.
Comparison: It is a typical species of Pachyfavosites.
However, it can be distinguished by its corallite diameter
from the type species Pachyfavosites polymorphus (Gold-
fuss, 1826) and from many other Pachyfavosites species,
whose mean corallite diameters are larger than 1 mm (e.g.,
Dubatolov 1959, 1963; Birenheide 1985; May 1993a;
Stadelmaier et al. 2005; Niko 2007; May 2008; Niko and
Senzai 2010).
Pachyfavosites multiperforatus: Dubatolov, 1959 from
the Salairka Formation of the Kuznetsk Basin (Siberia) has
significantly smaller corallite diameters (0.6–0.8 mm),
thinner walls (0.1–0.3 mm) and more mural pores (Dub-
atolov 1959: 61–62, pl. 17 fig. 8).
Pachyfavosites oekentorpi: Bruhl, 1999 from the Upper
Eifelian of the Eifel (Germany) has slightly smaller
corallite diameters (0.5–1.1 mm), slightly thicker walls
(about 0.25–0.35 mm, up to 0.6 mm) (Bruhl 1999: 40–42,
pl. 25–27). Furthermore, few squamulae have been
observed in Pachyfavosites oekentorpi Bruhl, 1999.
The material from Kierspe matches well the descriptions
of Pachyfavosites vilvaensis by Sokolov (1952) and
Dubatolov (1959).
Occurrence and geological age: Until this study, Pachyf-
avosites vilvaensis was known only from the Eifelian of the
Urals and from the Salairka Formation of the Kuznetsk Basin
(Siberia) (Sokolov 1952; Dubatolov 1959). The Salairka
Formation is Early Emsian in age (Weddige 2006; Yolkin
et al. 2006). This occurrence is significant for two reasons.
Firstly, it is the first evidence of Pachyfavosites vilvaensis
from central Europe. Secondly, it is one of the oldest
occurrences of the genus Pachyfavosites in central Europe.
Genus Thamnopora Steininger, 1831
Thamnopora: Steininger, 1831: 10; Hill 1981: 584; Bire-
nheide 1985: 69; May 1993a: 125; May 1993c: 78; Lutte
1993: 57; May 1998: 142; Bruhl 1999: 42; Stadelmaier et al.
2005: 14; Niko 2005: 22; Hubmann and Gaetani 2007: 316;
May 2008: 4; Niko and Senzai 2010: 49; Blake 2010: 151
Type species: Thamnopora madreporacea Steininger,
1831. Givetian, Bensberg (Germany).
Diagnosis and occurrence: See Birenheide (1985: 69) and
May (1993c: 78–79). For lists of assigned species and their
characterization see May (1993a: 126–130) and May (1998).
Thamnopora angusta Lecompte, 1939
Fig. 3c–f
*1939 Thamnopora angusta n. sp. Lecompte: 115–116,
pl. 16, figs. 17-20
1993a Thamnopora angusta—May: 131-133, text-fig. 7,
pl. 3, figs. 3-4 [cum syn.]
1997 Thamnopora angusta—Avlar and May: 110, pl. 2,
figs. 3-5
2003 Thamnopora angusta—May: 51, pl. 1, fig. 1
Material: Two thin-sections SMF 21.488–SMF 21.489,
and several fragments of colonies.
Comparison: The material matches well with the
descriptions of Thamnopora angusta by Avlar and May
(1997), May (1993a), and Lecompte (1939). Avlar and
May (1997) gave a detailed description of material from
the Meinerzhagener Korallenkalk, there is no need for a
new description here.
Occurrence and geological age: Thamnopora angusta has
been described originally from the Lower and Upper
Fig. 5 Microfacies. a Floatstone with various fossils (bryozoans,
corals, brachiopods), SMF 21.478. b Coral-bryozoan bafflestone
(Coenites vermicularis, Acanthoclema parvula, Fenestrapora cauca-sica), SMF 21.479. Components. c Echinoid spine, SMF 21.480.
d Ostracode, SMF 21.481. e Trilobite, SMF 21.482. f–j Girvanellawetheredii Chapman, 1908. f irregular aggregation, SMF 21.483.
g Encrusting sheet on Coenites vermicularis (McCoy, 1850), SMF
21.484. h–j Globular aggregations, SMF 21.484
b
Facies (2012) 58:727–758 735
123

736 Facies (2012) 58:727–758
123

Givetian of Belgium (Lecompte 1939: 116). Furthermore,
this species occurs in the Lower and Upper Givetian of the
Eifel, Bergisches Land and Sauerland (Germany), as well
as in the Upper Emsian, Eifelian and Givetian of Poland
and the Givetian of France, the Urals and Siberia
(Birenheide 1985: 72, Tab. 2; May 1993a). Our material
from Kierspe as well as the material described by Avlar
and May (1997) and the material figured by May (2003)
originates from Lower Eifelian coral limestones of western
Sauerland and represents early occurrences of this species.
Family Alveolitidae Duncan, 1872
Genus Coenites Eichwald, 1829
Coenites: Eichwald, 1829: 179; Hill 1981: 600; Avlar
and May 1997: 113; Niko and Senzai 2010: 54
Type species: Coenites juniperinus Eichwald, 1829.
Upper Middle Silurian, Estonia.
Diagnosis and occurrence: See Hill (1981: 600).
Coenites vermicularis (M‘Coy, 1850)
Figs. 3h, i,4f-g
*1850 Alveolites vermicularis n. sp. M‘Coy: 377
1939 Thamnopora (?) vermicularis—Lecompte:
130-132, pl. 18, figs. 13-14 [cum syn.]
1978 Gracilopora vermicularis—Stasinska and Now-
inski: 205-206, pl. 18, fig. 2, pl. 19, fig. 1
1997 Coenites vermicularis—Avlar and May: 113–114,
pl. 2, figs. 6-8 [cum syn.]
1999 Thamnopora schouppei Bruhl: 44-45, pls. 30–31
2003 Coenites vermicularis—May: 50, 51, pl. 1, fig. 1
Material: Two thin-sections SMF 21.498–SMF 21.499,
and several fragments of colonies.
Comparison: The material matches the descriptions of
Coenites vermicularis provided by Avlar and May (1997)
and Lecompte (1939) as well as the original description of
Thamnopora schouppei Bruhl 1999. Thamnopora schouppei
from the Upper Eifelian of the Eifel (Germany) is not a
thamnoporid, but a branching alveolitid and without doubt
conspecific with the material described by Avlar and May
(1997) and Lecompte (1939). As Avlar and May (1997) gave
a detailed description of material from the Meinerzhagener
Korallenkalk, there is no need here to give a new description.
Coenites kamei Niko and Senzai, 2010 from the Lower
Devonian of Japan (Niko and Senzai 2010: 54–57, fig. 12)
is very similar to Coenites vermicularis (M‘Coy, 1850).
However, Coenites kamei can be distinguished by its much
smaller height-width ratios (0.3–0.4) of the calices.
Occurrence and geological age: Lecompte (1939)
described Coenites vermicularis from the Middle Eifelian
to Lower Givetian of Belgium. Furthermore, this species
occurs in the Upper Eifelian of the Eifel (Germany) (Bruhl
1999) and in the Frasnian of Poland (Stasinska and Now-
inski 1978). Our material from Kierspe as well as the
material described by Avlar and May (1997) and the
material figured by May (2003) originates from Lower
Eifelian coral limestones of the western Sauerland and
represents early occurrences of this species.
Genus Platyaxum Davis, 1887
Platyaxum: Davis, 1887: Explanations to plates 60 and
61; Birenheide 1985: 85; May 1993a: 170
Type species Platyaxum turgidum: Davis 1887. Middle
Devonian of Indiana.
Diagnosis and occurrence: See Birenheide (1985: 85–86)
and May (1993a: 170–171).
Subgenus Roseoporella Spriestersbach, 1935
Roseoporella: Spriestersbach, 1935: 485; Birenheide
1985: 88; May 1993a: 176; Lutte 1993: 58; Avlar and May
1997: 114; Bruhl 1999: 53
Type species: Roseoporella rhenana Spriestersbach,
1935. Eifelian of Oberhabbach, Germany.
Diagnosis and occurrence: See Birenheide (1985: 88) and
May (1993a: 176).
Remarks: Birenheide (1985: 88) assumed that Roseopo-
rella rhenana Spriestersbach, 1935 was a junior synonym
of Alveolites taenioformis Schluter, 1889. However, the
material described by May (1993a: 179–180, pl. 10 fig. 2)
suggests that a specific distinction between Platyaxum
(Roseoporella) rhenana (Spriestersbach, 1935) and Platy-
axum (Roseoporella) taenioformis (Schluter, 1889) is
justified.
Platyaxum (Roseoporella) gradatum (Lecompte,
1939) Fig. 3g
*1939 Coenites gradatus n. sp. Lecompte: 69–70, pl. 11,
fig. 11-17
1993a Platyaxum (Roseoporella) gradatum—May:
180-182, fig. 19, pl. 11, fig. 1 [cum syn.]
1997 Platyaxum (Roseoporella) gradatum—Avlar and
May: 114–115, pl. 3, fig. 12, pl. 4, fig. 18
1999 Platyaxum (Roseoporella) gradatum—Bruhl: 54,
pl. 38
Fig. 6 a–c Girvanella problematica Nicholson and Etheridge, 1878,
SMF 21.485. d–g Rothpletzella munthei (Rothpletz, 1913) Wood,
1948. d, e SMF 21.487. f, g SMF 21.486
b
Facies (2012) 58:727–758 737
123

738 Facies (2012) 58:727–758
123

Material: One corallum SMF 21.683 (Fig. 3g).
Comparison: The material matches well the descriptions
of Platyaxum (Roseoporella) gradatum given by Avlar and
May (1997), May (1993a), and Lecompte (1939). Avlar
and May (1997) gave a detailed description of material
from the Meinerzhagener Korallenkalk, there is no need to
provide a new description here.
Occurrence and geological age: Platyaxum (Roseoporel-
la) gradatum has been described originally from the Eifelian
and Givetian of Belgium (Lecompte 1939: 70, 74). Several
authors documented and described this species from the
Eifelian and Lower Givetian of the Eifel, Germany
(Birenheide 1985: 88–89, pl. 30 fig. 3, tab. 2; Bruhl 1999),
the Lower Givetian of the Sauerland (Germany) (May
1993a) and the Eifelian of Siberia (Dubatolov 1959:
171–172, pl. 55 fig. 1; Dubatolov 1963: 104–105, pl. 38
fig. 2). Furthermore, P. (Roseoporella) gradatum has been
mentioned or doubtful material has been described from the
Eifelian and Givetian in other parts of Europe and Asia (May
1993a; Avlar and May 1997). Our material from Kierspe, as
well as the material described from Avlar and May (1997),
originates from Lower Eifelian coral limestones of western
Sauerland and represents early occurrences of this species.
Order Auloporida Sokolov, 1947
Genus Roemerolites Dubatolov, 1963
Roemerolites: Dubatolov, 1963: 58; Hill 1981: 659;
Birenheide 1985: 117; May 1993a: 197; Lutte 1993: 63;
Stadelmaier et al. 2005: 20
Type species: Roemerolites batschatensis Dubatolov
1963. Salairka Formation of the Kuznetsk Basin (Siberia),
Lower Emsian (see Weddige 2006; Yolkin et al. 2006).
Diagnosis and occurrence: See Birenheide (1985:
117–118) and May (1993a: 198).
Roemerolites tenuis (Schluter, 1885)
Fig. 3h
*1885 Syringopora tenuis n. sp. Schluter: 12–13
1889 Syringopora tenuis—Schluter: 429, pl. 16, figs. 1-4
1983 Syringoporella tenuis—Byra: 63–64, pl. 22 fig. 64
[cum syn.]
1985 Roemerolites tenuis—Birenheide: 120, fig. 75
1993 Roemerolites tenuis—Lutte: 63, figs. 1-2
2005 Roemerolites tenuis—Stadelmaier et al.: 22, pl. 8,
figs. 7-8
Material: Single corallum SMF 21.684 (Fig. 3h).
Description: Corallum dendroid with cerioid growth
centers. At the base of the corallum the corallites are rep-
tant, resembling Aulopora. Later corallites turn abruptly to
vertical growth. Large parts of the corallum are a 4–5-mm-
thick layer of dendroid corallites. In other parts, cerioid
growth centers are developed, which form branch-like
excrescences of up to 35 mm in height.
Diameter of adult corallites 1.0–1.4 mm, mean 1.1 mm.
Single wall thickness 0.1–0.5 mm (mean about 0.35 mm).
Connecting pores not frequent, with a diameter of about
0.2–0.3 mm. Tabulae not common. No septal spines
observed.
Comparison: The corallum matches well the descriptions
of Roemerolites tenuis (Schluter 1885) published by Byra
(1983) (type material), Lutte (1993) and Stadelmaier (et al.
2005). Roemerolites tenuis can be distinguished from the
other species of Roemerolites from central Europe by its
small corallite diameter and/or its growth form (Birenheide
1985: 119–120; May 1993a: 198–201; Stadelmaier et al.
2005: 20–22).
Occurrence and geological age: All material of Roemer-
olites tenuis previously described originated from the
Givetian of the Eifel (Germany) (Schluter 1885, 1889;
Byra 1983; Lutte 1993; Stadelmaier et al. 2005). Our
material from a Lower Eifelian coral limestone is the
earliest known occurrence of this species.
Subclass Zoantharia Blainville, 1830
Superorder Rugosa Milne-Edwards and Haime, 1850
Order Stauriida Verrill, 1865
Family Stringophyllidae Wedekind, 1922
Genus Sociophyllum Birenheide, 1962
Sociophyllum: Birenheide, 1962: 53; Birenheide 1979:
202; Hill 1981: 250; Coen-Aubert 1989: 8; Coen-Aubert
1990: 24; May 1993b: 51; Coen-Aubert 1999: 33;
Brownlaw and Jell 2008: 24.
Type species: Spongophyllum elongatum Schluter 1881.
Lower Givetian, Berndorf, Eifel (Germany).
Diagnosis and occurrence: See Hill (1981: 250) and May
(1993b: 52).
Sociophyllum semiseptatum rolfwerneri Birenheide,
1979 Figs. 3i,4a-c
*1979 Sociophyllum semiseptatum rolfwerneri n. subsp.
Birenheide: 202–203, pl. 6 fig. 8, pl. 12 fig. 19
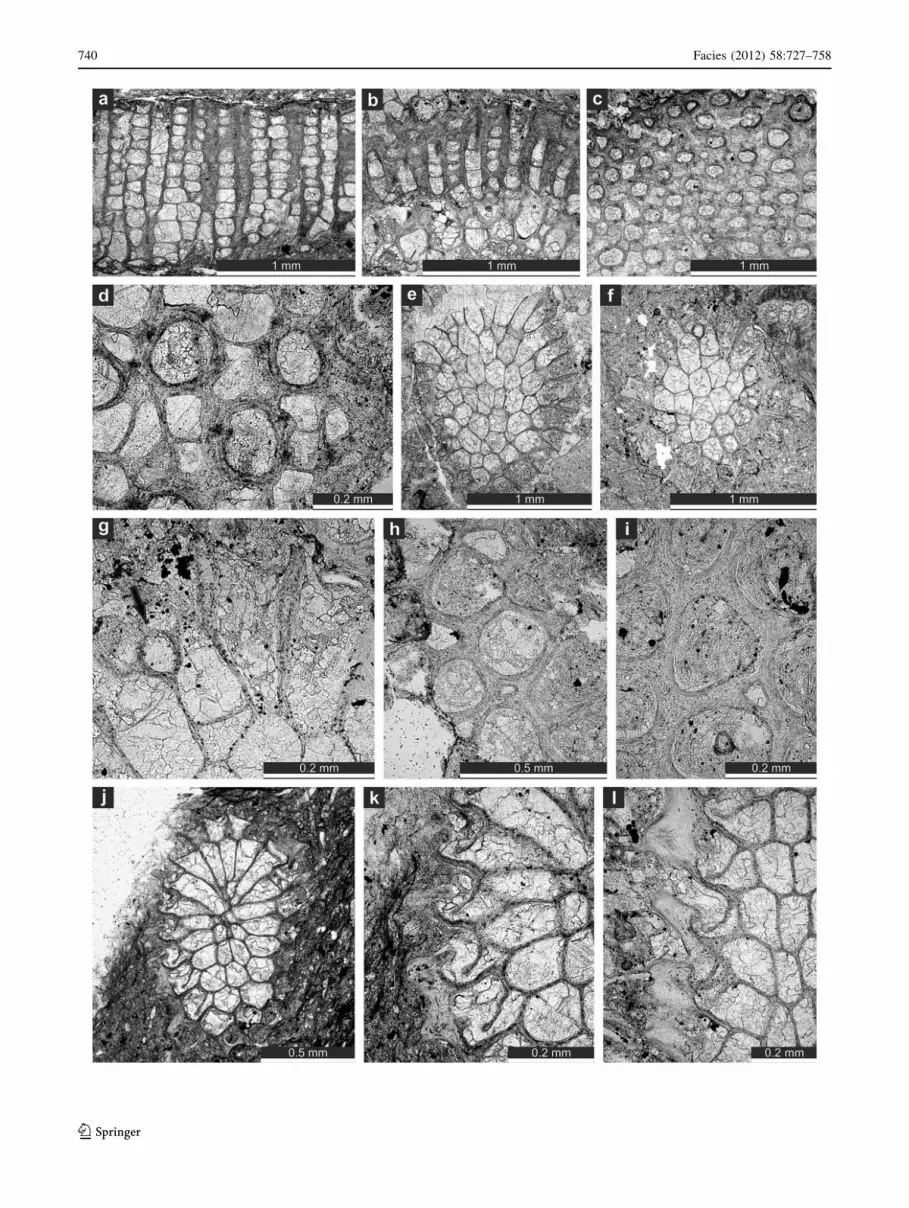
Fig. 7 a–d Fistuliphragma gracilis Ernst, 2008a, a tangential sec-
tion, SMF 21.385. b Tangential section SMF 21. 388. c Longitudinal
section SMF 21.386. d Longitudinal section, SMF 21.389.
e–k Fistuliporella kierspensis n. sp. e, f Longitudinal section,
holotype SMF 21.391. g Longitudinal section, paratype SMF
21.400. h, i Tangential section, holotype SMF 21.391
b
Facies (2012) 58:727–758 739
123
740 Facies (2012) 58:727–758
123

1990 Sociophyllum rolfwerneri—Coen-Aubert: 26–27,
pl. 4, figs. 21-27
Material: One large corallum (Fig. 4a) cut in pieces with
5 thin-sections (Fig. 4b, c) (SMF 21.492–SMF 21.493),
and some isolated corallites contained within the rock.
Description: Corallum phaceloid. Diameter of adult cor-
allites 7.5–12 mm. Diameter of juvenile corallites about
5 mm. In all juvenile corallites and a part of the adult
corallites the septal elements are reduced to irregularly
distributed spines and short septal segments. However, in
many adult corallites the septal segments are longer and
better developed. In these cases 30–36 septa can be
counted. Lonsdaleoid dissepiments occur normally in one
row, and less frequently in two rows, in vertical sections.
In longitudinal sections, the dissepiments are steeply
inclined. The tabularium is approximately half of the whole
diameter. The tabulae and tabellae are concave. In a
distance of 10 mm, there are 14–18 horizontal skeletal
elements.
Comparison: The material correlates with the descrip-
tions of Sociophyllum semiseptatum rolfwerneri by Bire-
nheide (1979) and Coen-Aubert (1990). Only the number
of septa is slightly higher in the material from Kierspe
(30–36 instead of 24–30). Taking into account the high
variability in the number of the septa, this difference can be
ignored. Sociophyllum semiseptatum rolfwerneri Birenhe-
ide 1979 can be distinguished from the other subspecies of
Sociophyllum semiseptatum (Schluter 1881) by its well-
developed lonsdaleoid dissepiments and septa. For more
detailed comparison, see Birenheide (1962, 1979) and
Coen-Aubert (1990).
Occurrence and geological age: Sociophyllum semisept-
atum rolfwerneri has been described originally from the
Lauch Formation (Lower Eifelian) of the Eifel (Birenheide
1979). The material from Kierspe is of the same age.
Furthermore, this species occurs in the Upper Eifelian of
Belgium (Coen-Aubert 1990).
Phylum Bryozoa Ehrenberg, 1831
Class Stenolaemata Borg, 1926
Order Cystoporata Astrova, 1964a
Family Fistuliporidae Ulrich, 1882
Genus Fistuliphragma Bassler, 1934
Fistulipora: Rominger, 1866: 121
Fistuliphragma: Bassler, 1934: 407; Utgaard 1983: 395;
Yang and Lu 1983: 270–271; Ernst 2008a: 326; Ernst and
Konigshof 2010: 9–19; Ernst et al. 2011a: 305
Type species: Fistulipora spinulifera Rominger, 1866.
Traverse Group (Middle Devonian); Michigan, USA.
Diagnosis: Solid, ramose and encrusting colonies, the
latter usually in form of hollow tubes. Secondary over-
growth occurring. Autozooecia originating from a thin
epitheca, bending sharply at their bases towards colony
surface, with circular apertures and large, prominent lu-
naria. Hemiphragms present, positioned in alternating
pattern in autozooecia, originating in laminated skeleton of
autozooecia, rare in endozones, rare to abundant in exoz-
ones. Basal diaphragms rare to abundant. Vesicular skele-
ton well developed. In the center of each vesicle roof a
single acanthostyle present. Monticules present.
Comparison: Fistuliphragma Bassler, 1934 is similar to
Cliotrypa Ulrich and Bassler in Bassler, 1929 and Stroto-
pora Ulrich in Miller, 1889 in the presence of hemi-
phragms, but differs from both genera in the absence of
gonozooecia.
Stratigraphic and geographic range: Middle Devonian of
USA, Germany, Western Sahara and Spain, and Upper
Carboniferous of China.
Fistuliphragma gracilis Ernst, 2008a
Fig. 7a–d; Table 1
2008a Fistuliphragma gracilis n. sp. Ernst: 329-332,
fig. 11A-G
2011a Fistuliphragma gracilis Ernst—Ernst et al.:
305-307, fig. 3e-g
Material: SMF 21. 385–SMF 21. 390.
Table 1 Descriptive statistics for Fistuliphragma gracilis Ernst,
2008a
N X SD CV MIN MAX
Aperture width (mm) 20 0.17 0.021 12.73 0.12 0.19
Autozooecial aperture
spacing (mm)
20 0.30 0.025 8.35 0.26 0.35
Lunarium length (mm) 6 0.062 0.014 23.15 0.050 0.088
Lunarium width (mm) 6 0.101 0.013 13.11 0.088 0.125
Lunarium thickness
(mm)
6 0.027 0.005 17.33 0.020 0.033
Vesicle width (mm) 10 0.085 0.025 29.78 0.050 0.125
Vesicles per aperture 10 8.3 1.252 15.08 7.0 11.0
N number of measurements, X mean, SD sample standard deviation,
CV coefficient of variation, MIN minimal value, MAX maximal value
Fig. 8 Leioclema passitabulatum Duncan, 1939. a Longitudinal
section, SMF 21.411. b Longitudinal section, SMF 21.404.
c, d Tangential section, SMF 21.410. e–i Leptotrypella sophiae n. sp.
e Branch oblique section, SMF 21.415. f, g Branch transverse section,
SMF 21.419 (arrow cyst-like structure). h, i Tangential section, SMF
21.416. Vidronovella fastigiata Gorjunova, 2006. j, k Branch oblique
section, SMF 21.430. l Branch, longitudinal section, SMF 21.428
b
Facies (2012) 58:727–758 741
123

Description: Encrusting colonies, 0.55–0.70 mm thick.
Autozooecia growing from low vertical walls on epitheca,
bending sharply at their bases towards colony surface.
Vertical walls thick, arising directly from thin epitheca,
double-layered, with distinct boundaries. Autozooecial
apertures circular to oval, having laminated, thick-walled
peristomes. Lunaria large, triangular. Long hemiphragms
positioned in alternating pattern in autozooecia, often
overlapping each other, curved proximally to distally, orig-
inating in laminated skeleton of autozooecial walls. Basal
diaphragms rare, thin, horizontal, or inclined. Vesicles large,
separating autozooecia in 1–2 rows, 7–11 surrounding each
autozooecial aperture, polygonal in tangential section, box-
like to hemispheric, with plane or concave roofs, consisting
of granular skeleton. Vesicular roofs containing acantho-
styles, 0.018–0.025 mm in diameter. Autozooecial walls
granular, 0.005–0.010 mm thick. Maculae not observed.
Comparison: Fistuliphragma gracilis Ernst, 2008a differs
from F. eifelensis Ernst, 2008a in having smaller colonies,
and smaller autozooecia (average autozooecial aperture
width 0.17 mm vs. 0.28 mm in F. eifelensis).
Occurrence and geological age: Bohnert Member of the
Freilingen Formation (Upper Eifelian); Eifel (western
Rhenish Massif), Germany. Santa Lucıa Formation,
Lower—Middle Devonian (Upper Emsian—Lower Eif-
elian); Abelgas and Paradilla, Cantabrian Mountains, NW
Spain.
Genus Fistuliporella Simpson, 1897
Type species: Lichenalia constricta Hall, 1883. Silurian-
Devonian; USA.
Diagnosis: Encrusting colonies. Autozooecia tubular,
with walls partly made by superimposed vesicle walls.
Lunaria in endozones and exozones, consisting of hyaline
material. Autozooecial diaphragms common to abundant,
straight or inclined. Vesicles usually high in endozones
becoming low in exozones, usually completely isolating
autozooecia. Elevated monticules occurring, consisting of
vesicles or dense stereom in central part, surrounded by a
ring of larger autozooecia.
Comparison: Fistuliporella Simpson, 1897 differs from
Fistuliporidra Simpson, 1897 due to the presence of
abundant diaphragms and in possessing granular auto-
zooecial walls rather than laminated walls.
Stratigraphic and geographic range: Upper Silurian—
Middle Devonian; North America. Middle Devonian;
Germany.
Remarks: Several species assigned to Fistuliporella were
described from the Upper Silurian to Middle Devonian of
North America (Rominger 1866; Hall 1883; Hall and
Simpson 1887; Ulrich and Bassler 1913; Bassler 1923).
However, most of these species were described without the
use of thin-sections. Only illustrations of thin-sections
made from the type species F. constricta (Hall, 1883) in
Utgaard (1983: 395, fig. 184, 1-d) were available for
comparison of the internal morphology of this genus.
Fistuliporella kierspensis n. sp.
Fig. 7e–k; Table 2
Etymology: The species is named after the type locality
Kierspe.
Holotype: SMF 21.391
Paratypes: SMF 21.392–SMF 21.400 Type locality: Ki-
erspe, Germany.
Type horizon: Middle Devonian (Lower Eifelian).
Diagnosis: Encrusting colonies; autozooecia with com-
mon basal diaphragms; lunaria small, triangular, thickened
to a massive node near colony surface; vesicles medium in
size, 6–10 surrounding each autozooecial aperture, arran-
ged commonly in single row between autozooecia, locally
completely isolating autozooecia; maculae not observed.
Description: Encrusting colonies, 0.9–2.1 mm thick.
Autozooecia growing from thick epitheca, bending sharply
at their bases towards colony surface. Autozooecial aper-
tures circular to oval or slightly polygonal due to surrounding
vesicles. Basal diaphragms common, locally abundant, thin,
horizontal or inclined. Lunaria small, triangular, consisting
of hyaline material, near the colony surface thickened to a
massive node. Vesicles medium in size, moderately high,
polygonal in tangential section, box-like, with concave
roofs, 6–10 surrounding each autozooecial aperture, arran-
ged commonly in single row between autozooecia, locally
completely isolating autozooecia. Autozooecial walls
0.02–0.03 mm thin, granular. Maculae not observed.
Table 2 Descriptive statistics for Fistuliporella kierspensis n. sp
N X SD CV MIN MAX
Colony thickness (mm) 5 1.4 0.493 35.32 0.9 2.1
Aperture width (mm) 30 0.39 0.052 13.42 0.30 0.49
Autozooecial aperture
spacing (mm)
30 0.49 0.068 13.99 0.36 0.60
Vesicle width (mm) 30 0.15 0.036 23.48 0.09 0.23
Vesicles per aperture 30 7.7 1.317 17.10 6.0 10.0
Vesicle spacing (mm) 30 0.17 0.049 28.57 0.10 0.27
Autozooecial diaphragm
spacing (mm)
20 0.17 0.053 31.24 0.06 0.28
Abbreviations as for Table 1
742 Facies (2012) 58:727–758
123

Comparison: Fistuliporella kierspensis n. sp. differs from
F. constricta (Hall, 1883) from the Silurian-Devonian of
USA in having larger autozooecial apertures (average
autozooecia width 0.39 mm vs. 0.25 mm in F. constricta).
Order Trepostomata Ulrich, 1882
Suborder Halloporina Astrova, 1965
Family Heterotrypidae Ulrich, 1890
Genus Leioclema Ulrich, 1882
[= Lioclema Ulrich, 1882]
Type species: Callopora punctata Hall, 1858. Lower
Carboniferous; Iowa, USA.
Diagnosis: Encrusting, branched, less commonly massive
colonies. Autozooecia with polygonal to rounded-polygo-
nal, sometimes petaloid apertures. Autozooecial diaphragms
rare. Mesozooecia abundant, with abundant diaphragms,
often beaded. Acanthostyles abundant, commonly large.
Autozooecial walls thin in endozone; laminated, regularly
thickened in exozones (modified after Astrova 1978).
Comparison: Leioclema Ulrich, 1882 differs from Het-
erotrypa Nicholson, 1879 in having rare autozooecial
diaphragms and abundant acanthostyles and mesozooecia,
and from Stigmatella Ulrich and Bassler, 1904 in having
abundant mesozooecia.
Stratigraphic and geographic range: Lower Silurian to
Upper Carboniferous; worldwide.
Leioclema passitabulatum Duncan, 1939
Fig. 8a–d; Table 3
1939 Lioclema passitabulatum n. sp. Duncan: 251, pl.
16, figs. 8-10
1964b Lioclema passitabulatum Duncan 1939—Astrova:
33, pl. 10, fig. 2a, b
2007 Leioclema passitabulatum Duncan 1939—Ernst
and Schroeder: 210, figs. 2I-K, 3A-C
2009 Leioclema passitabulatum Duncan 1939—Ernst
and May, 2009: 779, figs. 7.1-7.4
Material: SMF 21.401–SMF 21.414.
Description: Thin encrusting colonies, 0.33–1.05 mm in
thickness. Autozooecia growing from a thin epitheca,
remaining parallel to substrate for a long distance, then
bending sharply and intersecting the colony surface at right
angles. Autozooecial apertures rounded-polygonal. Auto-
zooecial diaphragms common. Mesozooecia abundant, 5–8
surrounding each aperture, polygonal in transverse section,
often beaded, containing thin diaphragms, moderate in size.
Acanthostyles moderately large, abundant, 2–4 surrounding
each aperture, originating at the base of exozone, often
indenting autozooecia, having distinct calcite cores and dark
laminated sheaths. Walls granular, 0.020–0.025 mm thick in
the endozone; distinctly laminated, 0.03–0.04 mm thick in the
exozone. Maculae indistinct, consisting of larger zooecia.
Comparison: Leioclema passitabulatum Duncan, 1939 is
similar to L. alpenense Duncan, 1939, but differs from it in
containing more abundant acanthostyles (2–5 vs. 2
acanthostyles around each aperture, respectively).
Occurrence and geological age: Bortschovski Horizon
(Lochkovian, Lower Devonian), Podolia (Ukraine). Kone-
prusy Limestone (Pragian, Lower Devonian); Czech Repub-
lic. Traverse Group (Eifelian, Middle Devonian), Michigan
(USA). Curten Formation (Lower Givetian, Middle Devo-
nian); Dollendorf Syncline, Rhenish Slate Massif (Germany).
Suborder Amplexoporina Astrova, 1965
Family Atactotoechidae Duncan, 1939
Genus Leptotrypella Vinassa de Regny, 1921
Type species: Chaetetes barrandei Nicholson, 1874.
Middle Devonian, Ontario (Canada).
Diagnosis: Branched colonies. Autozooecia with polyg-
onal to rounded-polygonal apertures. Autozooecial dia-
phragms lacking in endozones; rare to common in
exozones. Exilazooecia rare. Acanthostyles long, common
to abundant. Autozooecial walls granular, thin in endoz-
ones; laminated, irregularly thickened in exozones (modi-
fied after Astrova 1978).
Comparison: Leptotrypella Vinassa de Regny, 1921 dif-
fers from Leptotrypa Ulrich, 1883 in having a branched
Table 3 Descriptive statistics for Leioclema passitabulatum Duncan,
1939
N X SD CV MIN MAX
Colony thickness (mm) 5 0.64 0.280 44.05 0.33 1.05
Aperture width (mm) 30 0.11 0.015 14.04 0.08 0.13
Autozooecial aperture
spacing (mm)
30 0.19 0.028 14.47 0.15 0.26
Aperture width (mm)
(macular)
10 0.15 0.013 8.51 0.13 0.17
Autozooecial aperture
spacing (mm) (macular)
10 0.31 0.034 11.18 0.26 0.35
Mesozooecia width (mm) 30 0.08 0.022 26.34 0.05 0.13
Acanthostyle diameter
(mm)
30 0.04 0.008 23.48 0.03 0.06
Mesozooecia per aperture 30 6.4 0.850 13.36 5.0 8.0
Acanthostyles per
aperture
30 2.9 0.759 26.17 2.0 4.0
Mesozooecial diaphragms
spacing (mm)
30 0.08 0.015 19.65 0.05 0.11
Abbreviations as for Table 1
Facies (2012) 58:727–758 743
123

colony, and from Anomalotoechus Duncan 1939 in having
branched colonies and no diaphragms in endozones.
Stratigraphic and geographic range: Middle Silurian to
Lower Carboniferous; worldwide.
Leptotrypella sophiae n. sp.
Fig. 8e–i; Table 4
Etymology: The species is named in honor of Sophia
May Contento, third daughter of Andreas May.
Holotype: SMF 21.419.
Paratypes: SMF 21.415–SMF 21.418.
Type locality: Kierspe, Germany.
Type horizon: Middle Devonian (Lower Eifelian).
Description: Branched colonies. Branches 0.95–1.38 mm
in diameter. Exozone 0.15–0.39 mm wide, endozone
0.60–0.65 mm wide. Autozooecia long in endozones,
bending sharply in exozones, polygonal in transverse sec-
tion of endozone. Autozooecial apertures polygonal with
rounded corners. Autozooecial diaphragms rare, thin,
straight, occurring in the transition between endozone and
exozone. Exilazooecia rare, short, restricted to exozones,
rounded-polygonal in transverse section. Acanthostyles
moderate in size, rare to common, growing from the base
of the exozone, having distinct cores and laminated
sheaths. Autozooecial walls granular, 0.003–0.005 mm
thick in endozones; laminated, merged, without distinct
zooecial boundaries, 0.025–0.050 mm thick in exozones.
Secondary cingulum poorly developed. Mural spines
absent. Cyst-like structures in walls of exozone present,
0.055–0.063 mm in diameter.
Comparison: Leptotrypella sophiae n. sp. is similar to
L. inesae Ernst and May, 2012, from the Lower Devonian
(Lochkovian) of Spain. Both species have similar branch
width, rare acanthostyles and exilazooecia as well as cyst-
like structures. However, the new species has larger, but
more closely spaced autozooecial apertures (average
aperture width 0.13 mm vs. 0.09 mm in L. inesae; average
aperture spacing 0.16 mm vs. 0.20 mm in L. inesae).
Occurrence and geological age: Middle Devonian (Lower
Eifelian); Kierspe, Germany.
Order Cryptostomata Vine, 1884
Suborder Rhabdomesina Astrova and Morozova,
1956
Family Rhabdomesidae Vine, 1884
Genus Vidronovella Gorjunova, 2006
Vidronovella: Gorjunova, 2006: 48; Ernst and Konigshof
2010: 17–18; Ernst 2011: 10
Type species: Vidronovella fastigiata Gorjunova, 2006.
Late Devonian (Famennian); Afghanistan.
Diagnosis: Branched colonies. Autozooecia tubular,
short, budding from indistinct medial axis or short mesot-
heca in spiral order around the branch, oriented at high
angles to the branch axis. Autozooecial diaphragms absent.
Proximal part of autozooecia thickened in the outermost
exozone (‘‘fastigia’’ sensu Gorjunova 2006). Both superior
and inferior hemisepta present, located in the distal part of
autozooecia. Superior hemiseptum moderately long, hook-
shaped, curved distally, positioned at the base of the
exozone (‘‘fastigium’’ sensu Gorjunova 2006); inferior
hemiseptum long, slender, occupying two-thirds of body
cavity of autozooecia, positioned beneath superior hemi-
septa, curved distally. Secondary blunt hemisepta may
occur, one proximally to the superior hemiseptum, and
another one distally to the inferior hemiseptum. Auto-
zooecial apertures oval to rounded-rhombic, arranged
regularly in alternating rows on the colony surface.
Acanthostyles large and blunt, with narrow hyaline cores
and wide laminated sheaths, embedded in the skeleton.
Single or two acanthostyles positioned between two lon-
gitudinally successive autozooecial apertures. Paurostyles
occur in one species, irregularly distributed between
acanthostyles. Heterozooecia absent. Walls granular in the
endozone; laminated in exozone, becoming structureless
near the colony surface. Mural spines may occur.
Comparison: Originally the genus Vidronovella Gorju-
nova, 2006 was placed in a family of its own, Vidrono-
vellidae (Gorjunova 2006). However, the main characters
of this genus (autozooecial shape and budding mode as
well as the presence of hemisepta and acanthostyles) sug-
gest that Vidronovella Gorjunova, 2006 belongs to the
family Rhabdomesidae Vine, 1884. Vidronovella is similar
to Orthopora Hall, 1886, but differs from it by the shape of
short autozooecia and the high budding angle of auto-
zooecia in the axial area.
Table 4 Descriptive statistics for Leptotrypella sophiae n. sp
N X SD CV MIN MAX
Aperture width (mm) 25 0.13 0.017 13.51 0.10 0.16
Autozooecial Aperture
Spacing (mm)
25 0.16 0.019 11.43 0.13 0.20
Acanthostyle diameter
(mm)
4 0.030 0.003 11.24 0.025 0.033
Exilazooecia width
(mm)
9 0.050 0.019 37.81 0.018 0.068
Autozooecial wall
thickness (mm)
5 0.037 0.010 28.16 0.025 0.050
Abbreviations as for Table 1
744 Facies (2012) 58:727–758
123

Stratigraphic and geographic range: Lower Devonian of
Spain, Middle Devonian of Western Sahara and Germany,
Upper Devonian of Afghanistan.
Vidronovella fastigiata Gorjunova, 2006
Figs. 8i, j, 9a–c; Table 5
2006 Vidronovella fastigiata n. sp. Gorjunova: 48, pl. 2,
fig. 2
2010 Vidronovella fastigiata Gorjunova, 2006—Ernst
and Konigshof: 18–19, pl. 11g-h, pl. 12a-c
2010 Vidronovella fastigiata Gorjunova, 2006—Ernst
and Rodrıguez: fig. 4d
Material: SMF 21.420–SMF 21.430.
Description: Branched colonies, 0.69–0.96 mm in
diameter, with 0.35–0.54 mm wide endozones and
0.09–0.26 mm wide exozones. Branching style unknown.
Transversal sections of branches circular to oval. Auto-
zooecia tubular, short, budding from indistinct medial axis
or short mesotheca in spiral order around the branch.
Autozooecial diaphragms absent. Proximal part of auto-
zooecia thickened in the outermost exozone. Both superior
and inferior hemisepta present, located in the distal part of
autozooecia. Superior hemiseptum moderately long, hook-
shaped, curved distally, positioned at the base of the
thickening; inferior hemiseptum long, slender, occupying
two-thirds of body cavity of autozooecia, positioned
beneath superior hemisepta, curved distally. Secondary
hemisepta absent. Autozooecial apertures oval to rounded-
rhombic, arranged regularly in alternating rows on the
colony surface. Acanthostyles large and blunt, with nar-
row hyaline cores and wide laminated sheaths, embedded
in the skeleton, 0.03–0.05 mm in diameter. Single
acanthostyle positioned between two longitudinally suc-
cessive autozooecial apertures. Paurostyles absent. Het-
erozooecia absent. Walls in the endozone granular,
0.010–0.015 mm thick; laminated in exozone. Mural
spines absent.
Comparison: Vidronovella fastigiata Gorjunova, 2006
differs from V. intricata Ernst, 2011 from the Lower
Devonian of Spain in having of combination of single
superior and inferior hemisepta instead of complex doubled
superior and inferior hemisepta that is developed in
V. intricata.
Occurrence and geological age: Middle Devonian (Eif-
elian); Germany. Middle Devonian (Late Givetian);
Sabkhat Lafayrina reef complex, Tindouf Basin, Western
Sahara. Late Devonian (Famennian); Afghanistan. ? Mid-
dle Devonian; Pajarejos near Campillo de Llerena, south-
western Spain.
Family Nikiforovellidae Gorjunova, 1975
Genus Acanthoclema Hall, 1886
[= Rozanovia Gorjunova, 1992]
Type species: Trematopora alternata Hall, 1883. Lower
Devonian (Emsian); New York (USA).
Diagnosis: Branched colonies. Autozooecia diverging at
high angles from distinct median axis. Hemisepta absent.
Shallow or terminal diaphragms occurring. Autozooecial
walls laminated, without visible boundaries, containing
mural spines. Generally, single metazooecium and few
paurostyles or acanthostyles concentrated between longi-
tudinally successive autozooecial apertures.
Comparison: Acanthoclema Hall, 1886 is similar to
Streblotrypella Nikiforova, 1948, but differs in having single
metazooecium between longitudinally successive auto-
zooecial apertures, whereas metazooecia in Streblotrypella
are distributed irregularly throughout the colony. Further-
more, Acanthoclema possesses mural spines in the skeleton,
which is unique within the family Nikiforovellidae.
Stratigraphic and geographic range: Silurian of North
America, Devonian and Carboniferous of North America
and Europe.
Acanthoclema parvula Ernst et al. 2011a
Fig. 9d–i; Table 6
2011a Acanthoclema parvula n. sp. Ernst et al.: 312,
figs. 6f-j,7a
Material: SMF 21.431–SMF 21.463.
Description: Branches 0.47–1.00 mm in diameter, with
0.30–0.60 mm wide endozone and 0.09–0.26 mm wide
exozone. Tubular autozooecia growing in spiral pattern
from the distinct median axis at angles of 57�–77� in en-
dozones, abruptly bending in exozones and intersecting
colony surface at angles of 51�–77�; with a triangular to
rhombic, tear-drop shape in transverse section of endozone.
Autozooecial apertures oval, arranged in regular diagonal
rows on branches. Basal diaphragms rare to common. Single
short metazooecium between longitudinally successive
autozooecial apertures. Acanthostyles small, 2–6 irregularly
distributed between autozooecial apertures. Autozooecial
walls finely laminated, with dividing hyaline layer,
0.005–0.013 mm thick in endozone; laminated, without
distinct boundaries, with abundant mural spines in exozone.
Mural spines common, 0.010–0.015 mm in diameter,
opening into autozooecial cavities and to the colony surface.
Comparison: Acanthoclema parvula Ernst et al., 2011a
differs from A. irregularis Ernst, 2011 from the Lower
Devonian (Emsian) of NW Spain in having smaller
Facies (2012) 58:727–758 745
123

746 Facies (2012) 58:727–758
123

apertures (average aperture width 0.059 mm vs. 0.080 mm
in A. irregularis) and smaller and more abundant
acanthostyles (average acanthostyle diameter 0.025 mm vs.
0.035 mm in A. irregularis).
Occurrence and geological age: Santa Lucıa Formation,
Early—Middle Devonian (Late Emsian—Early Eifelian).
Middle Devonian (Early Eifelian); Kierspe, Germany.
Order Fenestrata Astrova and Morozova, 1956
Suborder Fenestellina Astrova and Morozova, 1956
Family Septatoporidae Engel, 1975
Genus Dissotrypa Ernst and Konigshof, 2010
Type species: Dissotrypa sincera Ernst and Konigshof,
2010. Middle Devonian (Givetian); West Sahara.
Diagnosis: Reticulate colonies in form of fans, not com-
pletely closed to cones, composed of slender branches and
relatively thin dissepiments; autozooecia arranged in two
rows on the branches; branch bifurcation without insertion of
additional autozooecia; autozooecial chambers generally
pentagonal in mid tangential section, triangular in shallower
sections; axial wall between autozooecial rows zigzag in
tangential sections; stellate structures in autozooecial aper-
tures occurring; single exozonal tube in each autozooecium
present, situated laterally between longitudinally successive
autozooecial apertures; narrow median keel with single row
of closely spaced nodes; heterozooecia in form of spherical
chambers positioned between longitudinally successive
autozooecial apertures or on dissepiments; external lami-
nated skeleton well developed, coarsely laminated, with
microstyles and larger styles.
Comparison: Dissotrypa Ernst and Konigshof, 2010 differs
from Septatopora Engel, 1975, represented by several spe-
cies from the Carboniferous of Australia, in the bifurcation
mode of branches: there is insertion of additional autozooecia
at place of bifurcation, whereas the number of autozooecial
rows in Septatopora increased up to four and was retained for
long distances. Furthermore, Dissotrypa has a median keel,
which is absent in Septatopora. Both genera display similar
internal morphology such as autozooecial shape, presence of
single exozonal tube in each autozooecium and stellate
structures, as well as similar brood chambers.
Stratigraphic and geographic range: Middle Devonian;
West Sahara, Germany, Spain.
Dissotrypa sincera Ernst and Konigshof, 2010
Figs. 9j,10a–i; Table 7
2005 Rectifenestella sp.—Scholz et al.: fig. 7f
2010 Dissotrypa sincera n. sp. Ernst and Konigshof:
25–26, pl. 21d-f, pl. 22a-h, 23a-f
Material: SMF 21.464–SMF 21.467.
External description: Reticulate colonies composed of
slender straight branches and relatively thin dissepiments.
Table 5 Descriptive statistics for Vidronovella fastigiata Gorjunova,
2006
N X SD CV MIN MAX
Branch width (mm) 7 0.81 0.100 12.35 0.69 0.96
Endozone width (mm) 7 0.46 0.066 14.15 0.35 0.54
Exozone width (mm) 7 0.17 0.031 17.58 0.14 0.21
Aperture width (mm) 3 0.09 0.010 11.11 0.08 0.10
Acanthostyle diameter
(mm)
5 0.04 0.008 19.76 0.03 0.05
Abbreviations as for Table 1
Table 6 Descriptive statistics for Acanthoclema parvula Ernst,
Dorsch and Keller, 2011
N X SD CV MIN MAX
Branch width (mm) 20 0.77 0.142 18.37 0.47 1.0
Endozone width
(mm)
20 0.42 0.088 21.04 0.30 0.60
Exozone width
(mm)
20 0.18 0.042 23.47 0.09 0.26
Aperture width
(mm)
60 0.059 0.009 15.72 0.043 0.078
Aperture spacing
along branch
(mm)
40 0.30 0.031 10.32 0.22 0.36
Aperture spacing
across branch
(mm)
40 0.18 0.012 6.71 0.16 0.20
Acanthostyle
diameter (mm)
40 0.025 0.004 15.89 0.015 0.035
Metazooecia width
(mm)
40 0.03 0.007 26.35 0.02 0.05
Autozooecial
budding angle,
endozone
7 68.7 8.200 11.93 57.0 77.0
Autozooecial
budding angle,
exozone
7 64.3 8.902 13.85 51.0 77.0
Abbreviations as for Table 1
Fig. 9 a–c Vidronovella fastigiata Gorjunova, 2006, branch, oblique
section showing hemisepta, autozooecial apertures and acanthostyles,
SMF 21.423. Acanthoclema parvula Ernst, Dorsch and Keller, 2011.
d Tangential section, SMF 21.452. e Branch, transverse section, SMF
21.454. f, g Branch, longitudinal section, SMF 21.450. h, i Tangential
section showing autozooecial apertures, metazooecia (M), acantho-
styles (A), and mural spines (MS), SMF 21.432. j Dissotrypa sinceraErnst and Konigshof, 2010, tangential section, SMF 21.467
b
Facies (2012) 58:727–758 747
123

748 Facies (2012) 58:727–758
123

Autozooecia arranged in two rows on the branches, 2–3
spaced per length of a fenestrule; branch bifurcation
without insertion of additional autozooecia. Autozooecial
apertures circular with stellate structures composed of eight
septa. Narrow and low median keel with single row of
closely spaced nodes.
Internal description: Autozooecial chambers generally
pentagonal in mid-tangential section, triangular in shallower
sections; axial wall between autozooecial rows zigzag in
tangential sections. Single exozonal tube in each autozooe-
cium present, situated laterally between longitudinally suc-
cessive autozooecial apertures, 0.02–0.05 mm in diameter.
Heterozooecia in form of spherical chambers positioned
between longitudinally successive autozooecial apertures
or on dissepiments, 0.10–0.18 mm in diameter. Internal
granular skeleton thin, continuous with obverse keel,
nodes, microstyles, peristome and across dissepiments,
0.025–0.035 mm thick on the reverse wall. External lami-
nated skeleton well developed, 0.09–0.10 mm thick on the
reverse wall, coarsely laminated, protruded by microstyles
and larger styles. Microstyles 0.010–0.015 mm and styles
0.04–0.05 mm in diameter.
Comparison: The present material differs slightly from
the West Sahara specimens in having narrower branches
(average branch width 0.29 mm vs. 0.34 mm, respectively)
and thinner dissepiments (average dissepiment width
0.12 mm vs. 0.21 mm, respectively).
Occurrence: Middle Devonian (Late Givetian); Sabkhat
Lafayrina reef complex, Tindouf Basin, Western Sahara.
Middle Devonian (Eifelian); Germany. ? Middle Devonian;
Pajarejos near Campillo de Llerena, southwestern Spain.
Family Semicosciniidae Morozova, 1987
Genus Fenestrapora Hall, 1885
Type species: Fenestrapora biperforata Hall, 1885.
Middle Devonian; New York, USA.
Diagnosis: Reticulate funnel-shaped colonies consisting
of sinusoid, relatively wide and thick branches jointed by
wide and short dissepiments. Autozooecia arranged in two
rows on the branches, opening onto outer surface of the cone.
High median keel widening apically. Vesicular skeleton
(kenozooecia) often developed. Autozooecial chambers
rectangular in mid-tangential section, short and relatively
high, with moderately short vestibules. Axial wall straight to
slightly sinusoid, projecting in the median keel. Hemisepta
absent. Heterozooecia represented by brood chambers in
form of roughly rounded voids (?enlarged zooecia), situated
preferably near dissepiments, as well as special hetero-
zooecia called aviculomorphs (McKinney 1998).
Comparison: Fenestrapora Hall, 1885 differs from
Semicoscinium Prout, 1859 in possessing aviculomorphs,
and from Isotrypa Hall, 1885 in possessing aviculomorphs
and the absence of a reticulate protecting superstructure.
Stratigraphic and geographic range: Lower—Middle
Devonian (Emsian—Givetian); USA. Middle Devonian
(Eifelian—Givetian); Germany. Middle Devonian (Eif-
elian); Transcaucasia.
Fenestrapora caucasica Morozova and Lavrentjeva,
1998
Fig. 11a–h; Table 8
1998 Fenestrapora caucasica n. sp. Morozova and
Lavrentjeva: 56–57, text-figs. 1-2, pl. 4, figs. 4-5, pl. 5,
figs. 1-2
2008b Fenestrapora caucasica Morozova and Lavrentj-
eva, 1998—Ernst: fig. 3.8-9
Table 7 Descriptive statistics for Dissotrypa sincera Ernst and
Konigshof, 2010
N X SD CV MIN MAX
Branch width (mm) 17 0.29 0.023 8.05 0.24 0.34
Dissepiment width
(mm)
25 0.12 0.012 10.40 0.10 0.14
Fenestrule width (mm) 25 0.22 0.029 13.20 0.17 0.26
Fenestrule length (mm) 25 0.36 0.016 4.31 0.34 0.40
Distance between
branch centers (mm)
25 0.46 0.054 11.76 0.37 0.54
Distance between
dissepiment centers
(mm)
25 0.50 0.029 5.79 0.42 0.54
Aperture width (mm) 25 0.078 0.004 5.55 0.070 0.085
Aperture spacing along
branch (mm)
25 0.22 0.018 8.15 0.19 0.26
Aperture spacing across
branch (mm)
25 0.21 0.018 8.43 0.18 0.25
Node diameter (mm) 20 0.051 0.007 13.75 0.040 0.075
Node spacing (mm) 10 0.28 0.030 10.90 0.23 0.32
Maximal chamber
width (mm)
15 0.12 0.011 8.93 0.11 0.14
Apertures per fenestrule
length
20 2.2 0.366 17.04 2.0 3.0
Diameter of exozonal
tubes (mm)
10 0.027 0.009 35.14 0.020 0.050
Abbreviations as for Table 1
Fig. 10 Dissotrypa sincera Ernst and Konigshof, 2010. a–e Tangen-
tial section showing autozooecia, apertures, nodes, and exozonal
tubes (arrows), SMF 21.467. f–h Tangential section, showing branch
bifurcation (f), ovicell (g, arrow), and reverse side with nodes and
microstyles (h), SMF 21.465. i Branch, transverse section showing
autozooecial chambers and exozonal tube (arrow), SMF 21.466
b
Facies (2012) 58:727–758 749
123

750 Facies (2012) 58:727–758
123

Material: SMF 21.468–SMF 21.477.
Description: Reticulate colony composed of moderately
wide, sinusoid branches jointed by wide and short dissepi-
ments. Autozooecia arranged in two rows on the branches,
3–5 spaced on the length of a fenestrule. Fenestrules oval to
lens-shaped. Narrow, high median keel present. Vesicular
skeleton locally well developed, concentrated near the base
of colonies. Reverse side smooth, containing microstyles,
styles, and large nodes. Microstyles 0.002–0.005 mm,
styles 0.025–0.075 mm, and nodes 0.10–0.20 mm in
diameter.
Internal description: Autozooecial chambers rectangular
in mid-tangential section, short and relatively high, with
moderate vestibules. Axial wall straight to slightly
sinusoid, projecting in the median keel. Hemisepta absent.
Heterozooecia represented by aviculomorphs occurring
only on reverse side of branches, 0.11–0.22 mm wide and
0.24–0.36 mm long. Internal granular skeleton thin, con-
tinuous with obverse keel, nodes, microstyles, peristome
and across dissepiments, 0.020–0.045 mm thick on the
reverse wall. External laminated skeleton well developed,
0.065–0.260 mm thick on the reverse wall, coarsely lami-
nated, with microstyles, styles and nodes.
Comparison: Fenestrapora caucasica Morozova and
Lavrentjeva, 1998 is similar to F. occidentalis Ulrich, 1890
from the Hamilton Group (Middle Devonian, Givetian) of
USA. Fenestrapora caucasica has shorter fenestrules
(average fenestrule length 0.58 mm vs. 0.70 mm in
F. occidentalis). Fenestrapora caucasica differs from
F. biperforata Hall, 1885 from the Lower-Middle Devonian
of USA in having wider branches (branch width 0.35–0.50
mm vs. 0.20–0.30 mm in F. biperforata).
Occurrence and geological age: Middle Devonian, Upper
Eifelian Mucrospirifer diluvianoides—Radiomena irregu-
laris brachiopod zone; Transcaucasia. Middle Devonian
(Eifelian—Lower Givetian); Eifel (western Rhenish Mas-
sif), Germany (unpublished data). Middle Devonian
(Lower Eifelian); Kierspe, Germany.
Discussion
Reef-building potential is generally considered to have
peaked during the Devonian (e.g., Copper 2002). Stroma-
toporoids, corals, and increasingly algae and calcimicrobes,
are considered to be the major framebuilders of that time
(e.g., Tsien 1984b; Fagerstrom 1987; Machel and Hunter
1994). Reefs in shallow areas of the Rheic Ocean were
predominantly produced by stromatoporoids and corals
(Krebs 1974; Tsien 1974; Burchette 1981; May 1987,
1988, 1991, 2003; Machel 1990; Malmsheimer et al. 1991).
These reefs show a high diversity of interactions between
organisms that were involved in their construction. Their
growth and development were controlled by different fac-
tors such as sea level fluctuations, climatic changes, and
sedimentation regimes (May 1996, 1997a, b, 2003).
A paleontological and sedimentological study of
deposits of the Meinerzhagener Korallenkalk (upper Cul-
trijugatus Beds, Lower Eifelian) at Kierspe reconstructs a
shallow marine environment just above the storm wave
base. From various literature sources (Embry and Klovan
1972; Flugel 1982: 467; Fagerstrom 1987: 42; Struve
1982a: 248–249; May 1988: 187, 1992a: 110, 1993d: 53)
the water depth can be estimated to have been about 20 m.
A large proportion of micrite in floatstones and bafflestones
(Fig. 5a, b) is indicative for quiet water conditions. The
Table 8 Descriptive statistics for Fenestrapora caucasica Morozova
and Lavrentjeva, 1998
N X SD CV MIN MAX
Branch width (mm) 25 0.40 0.038 9.49 0.35 0.50
Branch thickness (mm) 16 0.85 0.088 10.46 0.72 1.05
Dissepiment width
(mm)
25 0.40 0.053 13.33 0.29 0.52
Fenestrule width (mm) 30 0.33 0.037 11.05 0.27 0.41
Fenestrule length (mm) 30 0.58 0.086 14.71 0.45 0.78
Distance between
branch centers (mm)
30 0.70 0.099 14.15 0.52 0.90
Distance between
dissepiment centers
(mm)
25 1.02 0.068 6.63 0.90 1.15
Aperture width (mm) 30 0.10 0.007 7.07 0.09 0.12
Aperture spacing along
branch (mm)
30 0.25 0.026 10.37 0.20 0.30
Aperture spacing across
branch (mm)
20 0.25 0.025 10.02 0.20 0.28
Maximal chamber
width (mm)
25 0.156 0.020 13.13 0.125 0.200
Apertures per fenestrule
length
10 4.1 0.738 18.00 3.0 5.0
Thickness of reverse
wall laminated layer
(mm)
15 0.172 0.064 36.86 0.065 0.260
Thickness of reverse
wall granular layer
(mm)
15 0.032 0.009 28.41 0.020 0.045
Abbreviations as for Table 1
Fig. 11 Fenestrapora caucasica Morozova and Lavrentjeva, 1998.
a–c Tangential section, SMF 21.470. d Mid-tangential section
showing autozooecial chambers, SMF 21.468. e Tangential section
of the reverse side showing aviculomorh (arrow), SMF 21.473.
f, g Branch, longitudinal section showing autozooecial chambers and
aviculomorphs, SMF 21.477. h Branch, transverse section, SMF
21.47
b
Facies (2012) 58:727–758 751
123

presence of algae and spines of regular sea urchins identify
a position in the photic zone, whereas abundant bryozoans,
brachiopods, corals, and crinoids show it was at least
mesotrophic conditions.
Of worthy mention are ‘‘ragged’’ margins which occur
frequently in the coralla of Pachyfavosites vilvaensis
(Sokolov 1952) (fig. 3b). May (1992a: 112; 1993d: 56)
observed the same phenomenon commonly developed in
laminar to hemispherical stromatoporoids and alveolitids.
This feature must have been caused by episodic sedimen-
tation (Kershaw and Riding 1978; Kershaw 1984; Kershaw
1998; Tsien 1974; Tsien 1984a, b). In these skeletons a
portion of the colony died due to increasing sedimentation.
Later a reduction of the sedimentation rate allowed the
colony to grow laterally to recover the lost area. Conse-
quently the new growth phases of the skeleton detached
themselves at the margin of the colony from the lower part
of the skeleton and lie on the sediment surface.
The baffling of sediment was caused by meadows of
tabulate and rugose corals inhabited mainly by erect bry-
ozoans. Sediment was stabilized by encrusting cystoporate
and trepostome bryozoans as well as tabulate corals.
Binding by calcimicrobes was minor to moderate. Con-
trarily, there are examples of the extensive involvement of
calcimicrobes and algae in the development of Devonian
reefs (e.g., Wray 1972; Adachi et al. 2006, 2007;
Antoshkina and Konigshof 2008). Poncet (1995) showed
various complex interactions between algae and reef
organisms (stromatopores, tabulate and rugose corals) in the
Early to Middle Devonian reefs of the Armorican Massif.
It should be noted that bryozoans are usually very rare in
the Givetian to Frasnian reef complexes, but they are
important contributors in a reef complex of Pragian age in
Bohemia (May 1988; May 2005: 234; Ernst 2008c; Ernst and
May 2009). The importance of the bryozoans in the devel-
opment of the Meinerzhagener Korallenkalk lies somewhat
intermediate between both situations. Furthermore, the
Meinerzhagener Korallenkalk contains more bryozoans and
fewer stromatoporoids than what is in other aspects very
similar but younger (lowermost Givetian) limestones of the
Grunewiese Member of the Ihmert Formation (May 1992a,
1993d). One possible explanation for these differences is a
continuous increase of the water temperature from the Pra-
gian to the Upper Devonian. In fact, different observations,
amongst others the global sea-level rise (Boucot 1988;
Johnson et al. 1985, 1996; May 1996, 1997a, b), suggest a
significant increase of the global temperature from the Lower
Devonian to the Givetian-Frasnian. For a more detailed
discussion of this topic see May (2005: 234).
At the upper boundary of the Cultrijugatus Beds the
Chotec-Event is recorded. The Chotec-Event was caused by
a strong sea-level rise (May 1996; May 1997b). In the
Hercynian facies it is characterized by the deposition of
black shales (or dark limestones) and the appearance of the
ammonoid Pinacites jugleri (Roemer, 1843) (May 1997b).
In the Rhenish facies, the Chotec-Event can be recognized by
the extinction of Alatiformia alatiformis (Drevermann,
1907), all European species of Paraspirifer and some other
brachiopods (May and Avlar 1996; Avlar and May 1997;
May 1997a; May 1997b). This disappearance of Paraspirifer
(and Alatiformia alatiformis) indicates on the map sheet
Meinerzhagen the upper boundary of the Cultrijugatus Beds.
In contrast to these brachiopods, the corals found in
Kierspe show a remarkable continuity between the Lower
Eifelian and the younger Middle Devonian. This confirms
the observation of Avlar and May (1997) and May (1997a;
1997b), that in Sauerland, the rugose and tabulate corals
where not affected by the Chotec-Event. There is no evi-
dence that bryozoans were affected by the Chotec-Event
either. Fistuliphragma gracilis Ernst, 2008a, Leioclema
passitabulatum Duncan, 1939, Vidronovella fastigiata
Gorjunova, 2006, Dissotrypa sincera Ernst and Konigshof,
2010, and Fenestrapora caucasica Morozova and Lav-
rentjeva, 1998 occur also in the younger formations of the
Eifel, Morocco and Spain (Eifelian—Givetian).
The bryozoan fauna from the Meinerzhagener Koral-
lenkalk shows distinct similarities to the Lower—Middle
Devonian of Spain (Santa Lucıa Formation, Emsian—
Eifelian) and to the Middle Devonian (Eifelian) of Cau-
casus. Fistuliphragma gracilis and Acanthoclema parvula
were described from the Santa Lucıa Formation (Emsian—
Eifelian) of Abelgas and Paradilla, NW Spain (Ernst et al.
2011a, b). Fenestrapora caucasica was originally descri-
bed from the Middle Devonian of Transcaucasia (Upper
Eifelian Mucrospirifer diluvianoides—Radiomena irregu-
laris brachiopod zone). Fistuliphragma gracilis, Leioclema
passitabulatum, and Fenestrapora caucasica are known
from the Middle Devonian (Eifelian—Lower Givetian) of
the Rhenish Massif (Ernst 2008a, b). Leioclema passita-
bulatum is also known from the Lower Devonian of
Ukraine (Bortschovski Horizon, Lochkovian), Lower
Devonian of Bohemia (Koneprusy Limestone, Pragian) and
Middle Devonian (Traverse Group, Eifelian) of Michigan,
USA. Vidronovella fastigiata and Dissotrypa sincera are
known from the Givetian of Morocco.
The distribution of the corals is similar to the bryozoan
distribution. All coral genera found in the Meinerzhagener
Korallenkalk are typical representatives of the Old-World-
Realm. Except for Pachyfavosites vilvaensis (Sokolov,
1952), all coral species are known from other parts of
Central Europe. Pachyfavosites vilvaensis (Sokolov, 1952),
Thamnopora angusta Lecompte, 1939 and Platyaxum
(Roseoporella) gradatum (Lecompte, 1939) are known
from Siberia, thus documenting close paleobiogeographic
links between Central and Eastern Europe and Central
Asia. May (1993b) observed the same biogeographic
752 Facies (2012) 58:727–758
123

relationships in Lower Givetian corals from the north-
western Sauerland.
Conclusions
• The Lower Eifelian Meinerzhagener Korallenkalk
(= upper Cultrijugatus Beds) at Kierspe in the south-
western part of the Ebbe saddle (Sauerland) contains a
rich fauna of bryozoans, tabulate and rugose corals,
brachiopods, and echinoderms.
• The bryozoan fauna includes eight species as well as a
number of unidentified taxa. They show paleobiogeo-
graphic relations with the Lower-Middle Devonian of
Spain and Middle Devonian of Transcaucasia. The
majority of identified species occur also in the Middle
Devonian of the Rhenish Massif.
• The coral fauna contains five tabulate corals and one
rugose coral. Corals document paleobiogeographic rela-
tions between Central and Eastern Europe and Central
Asia.
• The accompanying fauna is represented by two bra-
chiopod species (Alatiformia alatiformis and Xystostro-
phia sp.). Furthermore, ostracods and various
fragmentary echinoderm remains were found.
• The flora is represented by three calcimicrobes. One
species (Girvanella wetheredii) is very common,
whereas two others are rare (Girvanella problematica,
Rothpletzella munthei).
• Faunal and microfacial characteristics suggest a depo-
sitional setting in shallow marine environment just
above the storm wave base, with a suggested depth of
20 m, within the photic zone and with at least a
mesotrophic conditions.
• At the top of the Cultrijugatus Beds the Chotec-Event
occurred, which strongly affected brachiopods, but not
corals and bryozoans.
Acknowledgments The authors are grateful to Wolfgang Reimers,
Kiel, Prof. Dr. Sergio Rodrıguez, Madrid, and Isabel Dıaz, Madrid, for
their help in the preparation of thin-sections used in this study.
Caroline Buttler, Cardiff, and an anonymous reviewer are thanked for
their reviews of the manuscript. Mark Wilson, Wooster, and Patrick
Wyse Jackson, Dublin, are thanked for valuable comments to the
manuscript. The present study was completed during project ER
278/4-1 and 2 supported by the Deutsche Forschungsgemeinschaft
(DFG). This paper is a contribution to UNESCO/IUGS IGCP Project
596 (Climate Change and biodiversity patterns in the Mid Palaeozoic).
References
Adachi M, Ezaki Y, Pickett JW (2006) Marked accumulation patterns
characteristic of Lower Devonian stromatoporoid bindstone:
palaeoecological interactions between skeletal organisms and
microbes. Palaeogeogr Palaeoclimatol Palaeoecol 231:331–346
Adachi M, Ezaki Y, Pickett JW (2007) Interrelations between
framework-building and encrusting skeletal organisms and
microbes: more-refined growth history of Lower Devonian
bindstones. Sedimentology 54:89–105
Anstey RL, Perry TG (1970) Biometric procedures in taxonomic
studies of Paleozoic bryozoans. J Paleont 44:383–398
Antoshkina A, Konigshof P (2008) Lower Devonian reef structures in
Russia: an example from the Urals. Facies 54:233–351
Astrova GG (1964a) New order of the Paleozoic Bryozoa. Paleont
Zhurnal 1:22–31 [in Russian]
Astrova GG (1964b) Mshanki borschovskogo i chortkovskogo
gorizontov Podolii [Bryozoa of the Borschov and Chortkov
horizons of Podolia]. Trudy Paleont Inst Akad Nauk SSSR
98:1–52 [in Russian]
Astrova GG (1965) Morphology, history of development and system
of the Ordovician and Silurian Bryozoa. Trudy Paleont Inst Akad
Nauk SSSR 106:1–432 [in Russian]
Astrova GG (1978) The history of development, system, and
phylogeny of the Bryozoa: Order Trepostomata. Trudy Paleont
Inst Akad Nauk SSSR 169:1–240 [in Russian]
Astrova GG, Morozova IP (1956) About systematics of the order
Cryptostomata. Dokl Akad Nauk SSSR 110(4):661–664 [in
Russian]
Avlar H, May A (1997) Zur Fauna und Stratigraphie der cultrijug-atus-Schichten (Wende Unter-/Mittel-Devon) im West-Sauer-
land (Rheinisches Schiefergebirge). Coral Res Bull 5:103–119
Basse M (1998) Trilobiten aus mittlerem Devon des Rhenohercynik-
ums: III. Proetida (3), Phacopida (2), Lichida (Lichoidea, Odon-
topleuroidea) und erganzende Daten. Palaeontogr A 249:1–162
Basse M (2002) Eifel-Trilobiten. 1. Proetida. Goldschneck-Verlag,
Korb, p 152
Basse M (2003) Eifel-Trilobiten. 2. Phacopida (1): Cheiruriden/
Acasten/Asteropygen. Goldschneck-Verlag, Korb, p 200
Basse M (2006) Eifel-Trilobiten. 4. Proetida (3), Phacopida (3).
Quelle and Meyer, Wiebelsheim, p 305
Basse M (2010) Neue und wenig bekannte Taxa der Scutelluinae,
Proetinae und Otarioninae (Trilobita) aus dem Mitteldevon der
Blankenheim-, Hillesheim- und Prum-Mulde (Eifel). Dortm
Beitr Landeskd Naturwiss Mitt 42:75–119
Basse M, Lemke U (1996) Trilobiten aus mittlerem Givetium (Mittel-
Devon) des nordlichen Rechtsrheinischen Schiefergebirges. Geol
Palaont Westfal 46:1–65
Bassler RS (1923) Systematic paleontology of Silurian deposits.
Bryozoa. Maryland Geological Survey, Silurian, pp 405–412
Bassler RS (1929) The Permian Bryozoa of Timor. Palaont von Timor
16:37–90
Bassler RS (1934) Notes on fossil and Recent Bryozoa. J Wash Acad
Sci 24:404–408
Bigey FP (1985) Biogeography of Devonian Bryozoa. In: Nielsen C,
Larwood GP (eds) Bryozoa: Ordovician to recent. Olsen and
Olsen, Fredensborg, pp 9–23
Birenheide R (1962) Revision der koloniebildenden Spongophyllidae
und Stringophyllidae aus dem Devon. Senckenberg leth 43:
41–99
Birenheide R (1979) Xystriphyllum- und Sociophyllum-Arten (Rug-
osa) aus dem Eifelium der Eifel. Senckenberg leth 60:189–221
Birenheide R (1985) Chaetetida und tabulate Korallen des Devon.
Leitfossilien 3:1–249
Blake PR (2010) Devonian corals of the Yarrol Province, eastern-
central Queensland. Mem Assoc Australas Palaeont 38:1–191
Borg F (1926) Studies on recent cyclostomatous Bryozoa. Zool
Bidrag Uppsala 10:181–507
Boucot AJ (1988) Devonian biogeography: an update. In: McMillan
NJ, Embry AF, Glass DJ (eds) Devonian of the world, Volume
III: Paleontology, paleoecology and biostratigraphy. Can Soc
Petrol Geol Mem, vol 14, pp 211–227
Facies (2012) 58:727–758 753
123

Brownlaw RLS, Jell JS (2008) Middle and Upper Devonian rugose
corals from the Canning Basin, Western Australia. Mem Assoc
Australas Palaeont 35:1–126
Bruhl D (1999) Stratigraphie, Fazies und Tabulaten-Fauna des oberen
Eifelium (Mittel-Devon) der Dollendorfer Mulde/Eifel (Rheinis-
ches Schiefergebirge. Kolner Forum Geol Palaont 4:1–155
Burchette TP (1981) European Devonian reefs: a review of current
concepts and models. In: Toomey DF (ed) European fossil reef
models. SEPM Special Publications, vol 30, pp 85–142
Byra H (1983) Revision der von C. L. Schluter (1880–1889)
beschriebenen Chaetetida und Tabulata aus dem Rheinischen
Devon. Cour Forsch-Inst Senckenb 59:1–127
Chapman F (1908) On the relationship of the genus Girvanella and its
occurrence in the Silurian limestones of Victoria. Rep Australas
Assoc Advanc Sci 2:377–386
Coen-Aubert M (1989) Representants des genres SociophyllumBirenheide, 1962 et Beugniesastraea n. gen. a la base du
Calcaire de Givet de Pondrome et de Resteigne (bord sud du
Bassin de Dinant, Belgique).). Bull Inst R Soc Nat Belg Sci
Terre 58:5–31
Coen-Aubert M (1990) Description de quelques Rugueux coloniaux
du Couvinien superieur de Wellin (bord sud du Bassin de Dinant,
Belgique). Bull Inst R Soc Nat Belg Sci Terre 59:15–35
Coen-Aubert M (1999) Description de quelques Rugueux coloniaux
de la Formation givetienne du Mont d’Haurs en Ardenne. Bull
Inst R Soc Nat Belg Sci Terre 69:27–46
Coen-Aubert M, Lutte B-P (1990) Massive rugose corals from the
Middle Devonian of the North Eifel Hills (Rheinisches Schi-
efergebirge, West Germany). Geol Palaeont 24:17–39
Copper P (2002) Silurian and Devonian reefs: 80 million years of
global greenhouse between two ice ages. In: Kiessling W, Flugel
E, Golonka J (eds) Phanerozoic reef patterns. SEPM Special
Publications, vol 72, pp 181–238
Cuffey RJ, McKinney FK (1979) Devonian Bryozoa. Palaeontol Spec
Pap 23:307–311
d’ Orbigny A (1849) Prodrome de paleontologie stratigraphique
universelle des animaux mollusques rayonnes, faisant suite ou
cours elementaire de paleontologie et geologie stratigraphique.
Volume 1. Victor Masson, Paris, p 392
Dana JD (1846–1849) Structure and classification of zoophytes. In:
United States exploring expedition during the years 1838, 1839,
1840, 1841, 1842 under the command of Charles Wilkes, U.S.N.,
7, Philadelphia, p x ? 740
Davis WJ (1887) Kentucky fossil corals. A monograph of the fossil
corals of the Silurian and Devonian Rocks of Kentucky. Part II.
Kentucky Geological Survey, Frankfort/Kentucky, p i–xiii ?
1-4, pls 1–139
de Blainville HMD (1830) Zoophytes. Dictionnaire des Sciences
Naturelles 60:1–546
Drevermann F (1907) Palaozoische Notizen. Ber Senckenberg
naturforsch Ges Frankfurt 1907:125–136
Dubatolov VN (1959) Tabuljaty, geliolitidy i chetetidy silura i devona
Kuzneckogo bassejna. Trudy VNIGRI 139:1–472
Dubatolov VN (1963) Pozdnesilurijskie i devonskie tabuljaty, geli-
olitidy i chetetidy Kuzneckogo bassejna. Akad Nauk SSSR,
Moscow, p 286 [in Russian]
Duncan PM (1872) Third report on the British fossil corals. In: Rep
41st Meet British Assoc Advanc Sci, vol 1871, pp 116–137
Duncan H (1939) Trepostomatous Bryozoa from the traverse group of
Michigan. Univ Mich Paleont Contrib 5:171–270
Ehrenberg CG (1831) Symbolae Physicae, seu Icones et descptiones
Corporum Naturalium novorum aut minus cognitorum, quae ex
itineribus per Libyam, Aegiptum, Nubiam, Dongalaam, Syriam,
Arabiam et Habessiniam, studia annis 1820-25, redirent. Pars
Zoologica, 4, Animalia Evertebrata exclusis Insectis. Berolini,
10 pls
Ehrenberg CG (1834) Beitrage zur physiologischen Kenntniß der
Corallenthiere im allgemeinen, und besonders des rothen
Meeres, nebst einem Versuche zur physiologischen Systematik
derselben. Abh Konigl Akad Wiss Berlin 1832:225–380
Embry AF, Klovan JE (1972) Absolute water depth limits of Late
Devonian paleoecological zones. Geol Rundsch 61:672–686
Engel B (1975) A new? Bryozoan from the Carboniferous of Eastern
Australia. Palaeontology 18:571–605
Erben H (1962) Zur Analyse und Interpretation der rheinischen und
hercynischen Magnafazies des Devons. In: Erben HK (ed) 2.
Internationale Arbeitstagung uber die Silur/Devon-Grenze und
die Stratigraphie von Silur und Devon, Bonn-Bruxelles 1960.
Symposiums-Band. Schweizerbart’sche Verlagsbuchhandlung,
Stuttgart, pp 42–61
Ernst A (2008a) Non-fenestrate bryozoans from the Middle Devonian
of the Eifel (western Rhenish Massif, Germany). N Jb Geol
Palaont Abh 250:313–379
Ernst A (2008b) Trepostome and cryptostome bryozoans from the
Koneprusy Limestone (Lower Devonian, Pragian) of Zlaty Kun
(Czech Republic). Riv Ital Paleontol Stratigr 114:329–348
Ernst A (2008b) New data on the Middle Devonian Bryozoa of
Germany. In: Hageman SJ, Key MM, Jr., Winston JE (eds)
Bryozoan Studies 2007: Proc 14th Int Bryozoology Conference,
Boone, North Carolina, July 1–8, 2007. Virginia Museum of
Natural History Spec Publ, vol 15, pp 29–36
Ernst A (2011) Cryptostome (ptilodictyine and rhabdomesine)
Bryozoa from the Lower Devonian of NW Spain. Paleontogr
A 293:147–183
Ernst A, Bohaty J (2009) Schischcatella (Fenestrata, Bryozoa) from
the Devonian of the Rhenish Massif, Germany. Palaeontology
52:1291–1310
Ernst A, Konigshof P (2010) Bryozoan fauna and microfacies from a
Middle Devonian reef complex (Western Sahara, Morocco). Abh
Senckenberg Naturforsch Ges 568:1–91
Ernst A, May A (2009) Bryozoan fauna from the Koneprusy
Limestone (Pragian, Lower Devonian) of Zlaty Kun near
Koneprusy (Czech Republic). J Paleont 83:767–782
Ernst A, May A (2012) Bryozoan fauna from the Lower Devonian
(Middle Lochkovian) of Sierra de Guadarrama, Spain. J Paleont
86:60–80
Ernst A, Rodrıguez S (2010) Bryozoan fauna from the oolitic limestone
near Pajarejos, SW Spain. Rev Espan Paleont 25:83–88
Ernst A, Schroeder S (2007) Stenolaemate bryozoans from the Middle
Devonian of the Rhenish Slate Massif (Eifel, Germany). N Jb
Geol Palaont 246:205–233
Ernst A, Dorsch T, Keller M (2011a) A bryozoan fauna from the
Santa Lucıa Formation (Lower–Middle Devonian) of Abelgas,
Cantabrian Mountains, NW-Spain. Facies 57:301–329
Ernst A, Konigshof P, Taylor P, Bohaty J (2011b) Microhabitat
complexity—an example from Middle Devonian bryozoan-rich
sediments. Palaeobiodiv Palaeoenv 91:257–284
Fagerstrom JA (1987) The evolution of reef communities. Wiley,
New York, p 618
Flugel E (1982) Microfacies analysis of limestones. Springer, Berlin,
p 633
Fuchs A (1923) Erlauterungen zur geologischen Karte von Preußen
und benachbarten Bundesstaaten. Lfg. 220, Blatt Meinerzhagen.
Geol Landesanstalt, Berlin
Gad J (1995) Neue Fossilfunde von Paraspirifer sandbergeri Solle,
1971 aus dem Unterdevon des Rheinischen Schiefergebirges und
ihre systematische und stratigraphische Bedeutung. Mainz
geowiss Mitt 24:27–46
Gad J (1997) Neue Informationen uber die Morphologie von
Alatiformia alatiformis und Alatiformia jaekeli (Brachiopoda,
Spinocyrtiidae) aus dem Ober-Emsium des Rheinischen Schi-
efergebirges. Geol Jahrb A 150:3–13
754 Facies (2012) 58:727–758
123

Goldfuss GA (1826–1833) Petrefacta Germaniae. Erster Theil. Arnz,
Dusseldorf
Gorjunova RV (1975) Permskie mshanki Pamira [Permian bryozoans
of the Pamir]. Trudy Paleont Inst Akad Nauk SSSR 148:1–127
[in Russian]
Gorjunova RV (1992) Morphology and system of the Paleozoic
bryozoans. Trudy Paleont Inst Akad Nauk SSSR 251:1–168 [in
Russian]
Gorjunova RV (2006) New bryozoans from the Devonian of
Afghanistan and the Carboniferous of Iran. Paleont J 40:626–635
Grabert H (1980) Oberbergisches Land. Zwischen Wupper und Sieg.
Samml Geol Fuhrer 68:1–178
Hageman SJ (1993) Effects of nonnormality on studies of the
morphological variation of a rhabdomesine bryozoan, Streblo-trypa (Streblascopora) prisca (Gabb and Horn). Univ Kansas
Paleont Contrib 4:1–13
Hahn HD (1990) Fazies grobklastischer Gesteine des Unterdevons
(Graue Phyllite bis Taunusquarzit) im Taunus (Rheinisches
Schiefergebirge). Thesis Univ Marburg, p 173
Hahn HD, Zankl H (1991) Sedimentation in the Lower Devonian of
the Taunus area (Graue Phyllite to Taunusquarzit). Zbl Geol
Palaont Teil I 1990:1509–1520
Hall J (1858) Report on the Geological Survey of Iowa, embracing the
results of investigations made during portions of the years 1855,
1856, 1857. Geol Surv Iowa, 1, Pt. 2, Paleontology, pp 1–724
Hall J (1883) Bryozoans of the Upper Heldelberg and Hamilton
groups. Trans Albany Inst 10:145–197
Hall J (1885) Bryozoans of the Hamilton group. Report of the State
geologist for the year 1883. Weeds, Parsons and Co., Albany,
pp 5–61
Hall J (1886) Bryozoa of the Upper Heldelberg groups; plates and
explanations. State Geologist of New York, 5th Annual Report
for 1885, pp 25–53
Hall J, Simpson GB (1887) Corals and Bryozoa from the Lower
Heldelberg, Upper Heldelberg and Hamilton groups. Geol Surv
State NY 6(98):265–288
Hatschek B (1888) Lehrbuch der Zoologie, eine morphologische
Uebersicht des Thierreiches zur Einfuhrung in das Studium
dieser Wissenschaft. Gustav Fischer, Jena, p 304
Heidelberger D (2003) Gastropoden aus dem Mittel-Eifelium der
Sotenicher Mulde (Eifel). Geol Palaeont 37:1–8
Heidelberger D (2008) Revision devonischer Gastropoden aus dem
Frettertal (Givetium; Sauerland). Geologie und Palaontologie in
Westfalen 71:5–45
Heidelberger D, Koch L (2005) Gastropoda from the Givetian
‘‘Massenkalk’’ of Schwelm and Hohenlimburg (Sauerland,
Rheinisches Schiefergebirge, Germany). Geol Palaeont Sb 4:
1–107
Hill D (1981) Rugosa and Tabulata. In: Teichert C (ed) Treatise on
invertebrate paleontology, Part F [Coelenterata] Suppl. 1.
Boulder, Colorado and Lawrence, Kansas, p xl ? 672
Hubmann B, Gaetani M (2007) Devonian calcareous algae, tabulate
corals and bioclaustrations from the Karakorum Mountains
(Northern Pakistan). Riv Ital Paleont Stratigr 113:307–328
Johnson JG, Klapper G, Sandberg CA (1985) Devonian eustatic
fluctuations in Euramerica. Geol Soc Am Bull 96:567–587
Johnson JG, Klapper G, Elrick M (1996) Devonian transgressive-
regressive cycles and biostratigraphy, northern Antelope Range,
Nevada; establishment of reference horizons for global cycles.
Palaios 11:3–14
Kershaw S (1984) Patterns of stromatoporoid growth in level-bottom
environments. Palaeontology 27:113–130
Kershaw S (1998) The applications of stromatoporoid paleobiology in
paleoenvironmental analysis. Palaeontology 41:509–544
Kershaw S, Riding R (1978) Parameterization of stromatoporoid
shape. Lethaia 11:233–242
Koch L, Lemke U, Brauckmann C (1990) Vom Ordovizium bis zum
Devon: die fossile Welt des Ebbe-Gebirges. Linnepe, Hagen, p 198
Krausel W (1953) Cyclopelta winteri (Bryozoa) aus dem Mittel-
Devon der Eifel. Senckenbergiana 34:43–52
Krausel W (1957) Loculipora Hall aus dem Mittel-Devon der Eifel.
Senckenbergiana 38:335–343
Krausel W (1981) Wuchsformen der Fenestelliden (Bryozoen) aus
dem Mitteldevon der Eifel. Aufschluß 32:29–36
Krebedunkel J (1995) Stromatoporen aus dem Givet und Frasne des
Bergischen Landes. Sonderveroff Geol Inst Univ Koln 106:
1–182
Krebs W (1974) Devonian carbonate complexes of Central Europe.
In: Laporte LF (ed) Reefs in time and space. SEPM Special
Publications, vol 18, pp 155–208
Kroner U, Hahn T, Romer RL, Linnemann U (2007) The Variscan
orogeny in the Saxo-Thuringian zone-heterogenous overprint of
Cadomian/Palaeozoic peri-Gondwana crust. In: Linnemann U,
Nance R D, Kraft P, Zulauf G (eds) The evolution of the Rheic
Ocean: From Avalonian-Cadomian active margin to Alleghe-
nian-Variscan collision. Geological Society of American Special
Papers, vol 423, pp 153–172
Langenstrassen F (1982) Sedimentologische und biofazielle Unter-
suchungen an Proben aus der Bohrung Schwarzbachtal 1
(Rheinisches Schiefergebirge, Velberter Sattel). Senckenberg
leth 63:315–333
Langenstrassen F (1983) Neritic sedimentation of the Lower and
Middle Devonian in the Rheinische Schiefergebirge East of the
River Rhine. In: Martin H, Eder FW (eds) Intracontinental Fold
Belts. Springer, Berlin, pp 43–76
Lecompte M (1939) Les tabules du Devonien moyen et superieur du
bord du bassin de Dinant. Mem Mus roy Hist natur Belg 90:1–229
Linnemann U, Pereira F, Jeffries TE, Drost K, Gerdes A (2008) The
Cadomian Orogeny and the opening of the Rheic Ocean: The
diachrony of geotectonic processes constrained by LA-ICP-MS
U–Pb zircon dating (Ossa-Morena and Saxo-Thuringian Zones,
Iberian and Bohemian Massifs). Tectonophysics 461:21–43
Lutte B-P (1987) Glossophyllum-Arten aus dem Mittel-Devon der
Eifel (Rugosa; Rheinisches Schiefergebirge). Senckenberg leth
67:433–457
Lutte B-P (1990) Horn- und kegelformige rugose Korallen aus dem
Mittel-Devon der Eifel. Senckenberg leth 70:297–395
Lutte B-P (1993) Zur stratigraphischen Verteilung tabulater Korallen
im Mittel-Devon der Sotenicher Mulde (Rheinisches Schi-
efergebirge, Nord-Eifel). Geol Palaeont 27:55–71
M‘Coy F (1850) On some new genera and species of Silurian Radiata
in the collection of the University of Cambridge. Ann Mag Nat
Hist 2:270–290
Machel H-G (1990) Faziesinterpretation des Briloner Riffs mit Hilfe
eines Faziesmodells fur devonische Riffkarbonate. Geol Jahrb D
95:43–83
Machel H-G, Hunter IG (1994) Facies models for middle to late
Devonian shallow-marine carbonates, with comparisons to
modern reefs: a guide for facies analysis. Facies 30:155–176
Malmsheimer KW, Mensink H, Stritzke R (1991) Gesteinsvielfalt im
Riffgebiet um Brilon. Geol Palaont Westf 18:67–83
Marks SJ (2010) Wiedereroffnung des Kiersper Hulloches bei
Kierspe/Meinerzhagen, Sauerland. Spelaol Jahrb 2008–2009:
45–48
May A (1987) Der Massenkalk (Devon) nordlich von Brilon
(Sauerland). Geol Palaont Westf 10:51–84
May A (1988) Fossilfuhrung und Palokologie des lagunaren Mas-
senkalkes (Devon) im Sauerland (Rheinisches Schiefergebirge).
Palaont Z 62:175–192
May A (1989) Uber Vandercammenina (Brachiopoda: Spiriferacea)
aus dem Mittel-Devon des Sauerlandes (Rheinisches Schi-
efergebirge). Hercynica 4(for 1988):15–23
Facies (2012) 58:727–758 755
123

May A (1991) Die Fossilfuhrung des westsauerlandischen Givetiums
(Devon; Rheinisches Schiefergebirge) in der Sammlung des
Stadtischen Museums Menden. Geol Palaont Westf 17:7–42
May A (1992a) Paleoecology of Upper Eifelian and Lower Givetian
coral limestones in the northwestern Sauerland (Devonian;
Rhenish Massif). Facies 26:103–116
May A (1992b) Die Kalkalgen-Flora des Ober-Eifeliums und Unter-
Givetiums (Devon) des nordwestlichen Sauerlandes (Rheinis-
ches Schiefergebirge). Palaeontogr B 228:1–28
May A (1993a) Korallen aus dem hoheren Eifelium und unteren
Givetium (Devon) des nordwestlichen Sauerlandes (Rheinisches
Schiefergebirge). Teil I: Tabulate Korallen. Palaeontogr A 227:
87–224
May A (1993b) Korallen aus dem hoheren Eifelium und unteren
Givetium (Devon) des nordwestlichen Sauerlandes (Rheinisches
Schiefergebirge). Teil II: Rugose Korallen, Chaetetiden und
spezielle Themen. Palaeontogr A 228:1–103
May A (1993c) Thamnopora und verwandte astige tabulate Korallen
aus dem Emsium bis Unter-Eifelium von Asturien (Devon;
Nord-Spanien). Geol Palaeont 27:73–101
May A (1993d) Stratigraphie, Stromatoporen-Fauna und Palokologie
von Korallenkalken aus dem Ober-Eifelium und Unter-Givetium
(Devon) des nordwestlichen Sauerlandes (Rheinisches Schi-
efergebirge). Geol Palaont Westf 24:3–93
May A (1996) Relationship among sea-level fluctuation, biogeogra-
phy, and bioevents of the Devonian: an attempt to approach
a powerful, but simple model for complex long-range control
of biotic crises. Geolines 3:38-49 (http://geolines.gli.cas.cz/
fileadmin/volumes/volume03/G3-038.pdf)
May A (1997a) Gedanken uber Zusammenhange zwischen Meeress-
piegel, Biogeographie und Bio-Events im Devon. Coral Res Bull
5:291–318
May A (1997b) Ein Modell zur Erklarung der Bio-Events und der
biogeographischen Entwicklung im Devon durch den Meeress-
piegel. Dortm Beitr Landeskd Naturwiss Mitt 31:137–174
May A (1998) Statistische Untersuchungen an der tabulaten Koralle
Thamnopora (Anthozoa; Devon). Geol Palaeont 32:141–159
May A (2003) Die Fossilfuhrung des Mitteldevons im Raum
Attendorn–Olpe (West-Sauerland; Rechtsrheinisches Schi-
efergebirge). Geol Palaont Westf 60:47–79
May A (2005) Die Stromatoporen des Devons und Silurs von Zentral-
Bohmen (Tschechische Republik) und ihre Kommensalen.
Zitteliana B25:117–250 (http://epub.ub.uni-muenchen.de/12135/)
May A (2008) Corals (Anthozoa, Tabulata and Rugosa) and chaetetids
(Porifera) from the Devonian of the Semara area (Morocco) at the
Museo Geominero (Madrid, Spain), and their biogeographic
significance. Bull Inst Sci Rabat Sect Sci terre 30:1–12
May A, Avlar H (1996) Evolution of Rhenish faunal communities during
the Late Emsian and Early Eifelian: three reviews on sedimentation,
brachiopods and bioevents. Geolines 3:50–52 (http://geolines.
gli.cas.cz/fileadmin/volumes/volume03/G3-050.pdf)
McKinney FK (1998) Avicularia-like structures in a Paleozoic
fenestrate bryozoan. J Paleont 72:819–826
Meyer W (1986) Geologie der Eifel. Schweizerbart’sche Verlags-
buchandlung, Stuttgart, p 614
Meyer W, Stets J (1980) Zur Palaogeographie von Unter- und
Mitteldevon im westlichen und zentralen Rheinischen Schi-
efergebirge. Z Dt Geol Ges 131:725–751
Meyer W, Stets J (1996) Das Rheintal zwischen Bingen und Bonn.
Samml Geol Fuhrer 89:1–386
Miller SA (1889) North American geology and paleontology.
Western Methodist Book concern, Cincinnati, p 664
Milne-Edwards H, Haime J (1850) A monograph of the British fossil
corals. First part. Introduction; corals from the Tertiary and
Cretaceous Formations. Monogr Palaeontogr Soc Lond 3(2)7:
i–lxxxv ? 1–72
Mittmeyer HG (1982) Rhenish Lower Devonian biostratigraphy. Cour
Forsch-Inst Senckenberg 55:257–270
Morozova IP (1987) Morphogenesis, system and colonial integration
in Bryozoa of the order Fenestrida. Trudy Paleont Inst Akad
Nauk SSSR 222:70–88 [in Russian]
Morozova IP, Lavrentjeva VD (1998) Novyye mshanki semeystva
Semicosciniidae [New bryozoans of the family Semicoscinii-
dae.]. Paleont Zhurnal 2:53–59 [in Russian]
Nance RD, Gutierrez-Alonso G, Keppie JD, Linnemann U, Brendan
Murphy J, Quesada C, Strachan RA, Woodcock N (2010)
Evolution of the Rheic Ocean. Gondwana Res 17:194–222
Nicholson HA (1874) Descriptions of new fossils from the Devonian
Formation of Canada West. Geol Mag 1:51–96
Nicholson HA (1879) On the structure and affinities of the ‘‘Tabulate
Corals’’ of the Paleozoic period, with critical descriptions of
illustrative species. William Blackwood and Sons, Edinburgh,
p 342
Nicholson HA, Etheridge R Jr (1878) A Monograph of the Silurian
fossils of the Girvan district, Ayrshire, with special reference to
those contained in the ‘‘Gray Collection’’. Volume 1, Pt. 1.
William Blackwood and Sons, London, pp 1–341
Nikiforova AI (1948) Lower Carboniferous bryozoans of Karatau.
Izdatelstvo Akad Nauk Kazakhskoi SSR, Alma-Ata, p 53 [in
Russian]
Niko S (2005) Devonian pachyporoidean tabulate corals from the
Fukuji Formation, Gifu Prefecture. Bull Natn Sci Mus Tokyo Ser
C 31:13–29
Niko S (2007) Stratigraphy of the Devonian Fukuji Formation in Gifu
Prefecture and its favositid coral fauna. Bull Natn Sci Mus
Tokyo Ser C 33:57–79
Niko S, Senzai Y (2010) Stratigraphy of the Devonian Kamianama
Formation in the Kuzuryu Lake-Ise River area, Fukui Prefecture
and its favositid coral fauna. Bull Natn Sci Mus Tokyo Ser C
36:31–59
Oncken O, Plesch A, Weber K, Ricken W, Schrader S (2000) Passive
margin detachment during arc-continent collision (Central
European Variscides). In: Franke W, Haak V, Oncken O, Tanner
D (eds) Orogenic processes: quantification and modelling in the
Variscan Belt. Geol Soc Lond Spec Publ 179:199–216
Poncet J (1995) Interaction organismes constructeurs, algues calcaires
et sediments dans les paleomilieux recifaux au devonien
inferieur et moyen. Rev Micropaleont 38:79–87
Prout HA (1859) Third series of descriptions of Bryozoa from the
Paleozoic rocks of western states and territories. Trans St Louis
Acad Sci Ser 3:443–452
Ribbert K-H (1998) Anmerkungen zur Devon-Korrelationstabelle,
R001dm97–R017dm97: Devon, Nordrhein-Westfalen. Sencken-
berg leth 77:308–309
Richter D (1971) Ruhrgebiet und Bergisches Land. Zwischen Ruhr
und Wupper. Samml Geol Fuhrer 55:1–166
Roemer CF (1844) Das rheinische Uebergangsgebirge. Eine palaeon-
tologisch-geologische Darstellung. Hahn0sche Verlagsbuchhand-
lung, Hannover, p 96
Romer RL, Hahne K (2010) Life of the Rheic Ocean: scrolling
through the shale record. Gondwana Res 17:408–421
Rominger C (1866) Observations on Chaetetes and some related
genera, in regard to their systematic position; with an appended
description of some new species. Proc Acad Nat Sci Philadelph
18:113–123
Rothpletz A (1913) Uber die Kalkalgen, Spongiostromen und einige
andere Fossilien aus dem Obersilur Gotlands. Sver geol Unders
Uppsala, ser C 10:1–54
Salerno C (2008) Stromatoporen-Fauna, Fazies und Palaookologie
von Plattformkarbonaten aus dem Unter-Givetium der Eifel
(Devon, Rheinisches Schiefergebirge). Zitteliana B27:1–129
(http://epub.ub.uni-muenchen.de/12002/)
756 Facies (2012) 58:727–758
123

Schlotheim EF (1820) Die Petrefactenkunde auf ihren jetzigen
Standpunkte durch die Beschreibung seiner Sammlung verstein-
erter und fossiler Uberreste des Thier- und Pflanzen-Reichs der
Vorwelt erlautert. Beckersche Buchhandlung, Gotha, p 437
Schluter C (1881) Uber einige Anthozoen des Devon. Verh Nat Ver
Preuss Rheinl Westf 38(4, 2):189–232
Schluter C (1885) Uber einige neue Anthozoen aus dem Devon. Verh
Nat Ver Preuss Rheinl Westf (Sitzungsber) 42:144–152
Schluter C (1889) Anthozoen des rheinischen Mittel-Devon. Abh geol
Spec Preuss Thuring Staaten 8:I–X, 259–465
Scholz J, Ernst A, Batson P, Konigshof P (2005) Bryozoenriffe.
Denisia 16:247–262
Schone BR, Basse M, May A (1998) Korrelationen des Eifelium/
Givetium Grenzbereichs im Rheinischen Schiefergebirge. Senc-
kenberg leth 77:233–242
Schroder S (1998) Rugose Korallen und Stratigraphie des oberen
Eifelium und unteren Givetium der Dollendorfer Mulde/Eifel
(Mittel-Devon; Rheinisches Schiefergebirge). Cour Forsch-Inst
Senckenberg 208:1–135
Schroder S, Salerno C (2001) Korallenfauna und Fazies givetischer
Kalksteinabfolgen (Curten-/Dreimuhlen-Formation) der Dollen-
dorfer Mulde (Devon, Rheinisches Schiefergebirge/Eifel). Senc-
kenberg leth 81:111–133
Simpson, GB (1897) A handbook of the genera of the North
American Paleozoic Bryozoa. 14th Annual Report of the State
Geologist (of New York) for the year 1894, pp 407–608.
Sokolov BS (1947) Novye syringoporidy Tajmyra. Bjull moskov
Obsc ispyt Prirody (Otd geol) 22(6):19–28
Sokolov BS (1952) Tabuljaty paleozoja evropejskoj casti SSSR. Cast
IV: Devon russkoj platformy i zapadnogo Urala. Trudy
vsesojuznogo neftjanogo naucno-issledovatel’skogo geologo-
razvedocnogo instituta (VNIGRI) n S 62:1–291
Solle G (1937) Hederella, eine amerikanische Bryozoen-Gattung im
rheinischen Unter-Devon. Senckenberg leth 19:15–21
Solle G (1952) Neue Untersuchungen und Arten der Bryozoen-
Gattung Hederella und eine Hernodia im rheinischen Unterdev-
on. Notzbl hess LA Bodenforsch 6:35–55
Solle G (1953) Die Spiriferen der Gruppe arduennensis-intermediusim rheinischen Devon. Abh hess LA Bodenforsch 5:1–15
Solle G (1968) Hederelloidea (Cyclostomata) und einige ctenostome
Bryozoen aus dem Rheinischen Devon. Abh hess LA Bode-
nforsch 54:1–40
Solle G (1971) Brachyspirifer und Paraspirifer im rheinischen
Devon. Abh hess LA Bodenforsch 59:1–163
Spriestersbach J (1935) Beitrag zur Kenntnis der Fauna des rheinis-
chen Devon. Jahrb preuss geol Landesanst 55[f. 1934]:475–525
Stadelmaier M, Nose M, May A, Salerno C, Schroder S, Leinfelder RR
(2005) Astige tabulate Korallen-Gemeinschaften aus dem Mittel-
devon der Sotenicher Mulde (Eifel): Faunenzusammensetzung
und fazielles Umfeld. Zitteliana B25:5–38 (http://epub.ub.uni-
muenchen.de/12133/)
Stasinska A, Nowinski A (1978) Frasnian Tabulata of the south-
eastern Poland. Acta Palaeont Polon 23:199–218
Steininger J (1831) Bemerkungen uber die Versteinerungen, welche in
dem Ubergangskalkgebirge der Eifel gefunden werden. Beilage
zum Gymnasial-Programmschrift zu Trier. Lints, Trier, p 44
Stets J, Schafer A (2002) Depositional environments in the Lower
Devonian siliciclastics of the Rhenohercynian Basin (Rheinis-
ches Schiefergebirge, W-Germany)—case studies and a model.
Contrib Sediment Geol 22:1–78
Struve W (1964) Beitrage zur Kenntnis devonischer Brachiopoden, 4:
Uber Alatiformia-Arten und andere, außerlich ahnliche Spirifer-
acea. Senckenberg leth 45:325–346
Struve W (1982a) Beitrage zur Kenntnis devonischer Brachiopoden,
23: Schaltier-Faunen aus dem Devon des Schwarzbach-Tales bei
Ratingen, Rheinland. Senckenberg leth 63:183–283
Struve W (1982b) The Eifelian within the Devonian frame, history,
boundaries, definitions. In: Ziegler W, Werner R (eds) On
Devonian stratigraphy and palaeontology of the Ardenno-
Rhenish Mountains and related Devonian matters. Cour For-
sch-Inst Senckenberg 55:401–432
Struve W (1992) Neues zur Stratigraphie und Fauna des rhenotypen
Mittel-Devon. Senckenberg leth 71:503–624
Struve W (1996) Trilobiten, Rheinisches Schiefergebirge; Mitteldev-
on. In Weddige K (ed) Devon-Korrelationstabelle. Senckenberg
leth 76:280, tab. B100dm96
Tait JA, Schatz M, Bachtadse V, Soffel H (2000) Palaeomagnetism
and Palaeozoic paleogeography of Gondwana and European
terranes. In Franke W, Haak V, Oncken O, Tanner D (eds)
Orogenic processes: quantification and modelling in the Variscan
Belt. Geol Soc London Spec Publ 179:21–34
Thormann F, Weddige K (2001) Addendum zu Struve W (1992) Neues
zur Stratigraphie und Fauna des rhenotypen Mittel-Devon:
Abbildungen der Holotypen. Senckenberg leth 81:307–327
Toots H (1951) Uber einige cryptostome Bryozoen aus dem
Mitteldevon des Rheinischen Schiefergebirges. N Jb Geol
Palaont Abh 93:233–246
Tsien HH (1974) Paleoecology of Middle Devonian and Frasnian in
Belgium. In: International symposium on Belgian micropaleon-
tological limits, Namur 1974, vol 12. Geol Survey Belgium,
Brussels, pp 1–53
Tsien HH (1984a) Organisms: their ecology and function in carbonate
construction. Palaeontogr Am 54:415–420
Tsien HH (1984b) Constructeurs de recifs devoniens: Stromatoporo-
ides, coraux tabuleux et rugueux et microorganisms. In: Geister
J, Herb R (eds) Geologie et paleoecologie des recifs. Inst Geol
Univ Berne, Bern, pp 26.1–26.21
Ulrich EO (1882) American Paleozoic Bryozoa. J Cincinnati Soc Nat
Hist 5(121–175):233–257
Ulrich EO (1883) American Paleozoic Bryozoa. J Cincinnati Soc Nat
Hist 6:245–279
Ulrich EO (1890) Paleozoic Bryozoa: III. Illinois Geol Surv 8:283–688
Ulrich EO (1896) Bryozoa. In: Eastman C (ed) Zittel’s textbook of
paleontology, vol 1. Macmillan, London, pp 257–291
Ulrich EO, Bassler RS (1904) A revision of the Paleozoic Bryozoa.
Part II: On genera and species of Trepostomata. Bull US Geol
Surv 173:15–55
Utgaard J (1983) Paleobiology and taxonomy of the Order Cystopo-
rata. In: Robison RA (ed) Treatise on Invertebrate Paleontology,
Part G (1) Bryozoa (revised). Geol Soc America and Univ
Kansas Press, Lawrence, pp G327–G439
Verrill AE (1865) Classification of polyps (Extract condensed from a
synopsis of the polypi of the North Pacific Exploring Expedition,
under captains Rinngold and Rodgers, U.S.N.). Essex Inst Proc
4:145–149
Vinassa de Regny P (1921) Sulla classificazione dei trepostomidi. Soc
Ital Sci Nat Atti 59:212–231
Vine GR (1884) Fourth report of the Committee consisting of Dr.
H. R. Sorby and Mr. G. R. Vine, appointed for the purpose of
reporting on fossil Polyzoa. Reports 53rd Mtg British Assoc
Advanc Sci, pp 161–209
von Eichwald CE (1829) Zoologia specialis quam expositis animal-
ibus tum vivis, tum fossilibus potissimum Rossiae in universum,
et Poloniae in specie, in usum, lectionum, 1. Joseph Zawadzki,
Vilna, p vi ? 314
Weddige K (ed) (2006) Devonian correlation table, Supplements
2006. Part 2. Senckenberg leth 86:327–347
Weddige K, Ziegler W (2000) Das bibliographische Lebenswerk von
Dr. Wolfgang Struve. Senckenberg leth 79:603–636
Wedekind R (1922) Zur Kenntnis der Stringophyllen des oberen
Mitteldevon. Sitzungsber Ges Beford gesamt Naturwiss Marburg
1921:1–16
Facies (2012) 58:727–758 757
123

Wedekind R (1937) Einfuhrung in die Grundlagen der historischen
Geologie. II. Mikrobiostratigraphie, Die Korallen- und Fora-
miniferenzeit. Enke, Stuttgart, p 136
Wehrmann A, Blieck A, Brocke R, Hertweck G, Jansen U, Konigshof
P, Plodowski G, Schindler E, Schultka S, Wilde V (2005)
Palaeoenvironment and palaeoecology of intertidal deposits in a
Lower Devonian siliciclastic sequence of the Mosel Region,
Germany. Palaios 20:101–120
Wierich F (1999) Orogene Prozesse im Spiegel synorogener
Sedimente; korngefugekundliche Liefergebietsanalyse siliziklas-
tischer Sedimente im Devon des Rheinischen Schiefergebirges.
Marburger Geowiss 1:1–264
Wood A (1948) ‘‘Sphaerocodium’’, a misinterpreted fossil from the
Wenlock Limestone. Proc Geol Assoc 59:9–22
Wray JL (1972) Environmental distribution of calcareous algae in
Upper Devonian reef complexes. Geol Rundsch 61:578–584
Yang J, Lu L (1983) Upper Carboniferous and Lower Permian
bryozoans from Kalpin of western Xinjiang. Paleont Cathayana
1:259–317 [In Chinese]
Yolkin EA, Izokh NG, Bakharev NK (2006) Annotations to the
Devonian correlation tables, R 175–184, H 110–111, R 608–612
di-ds 06: Devonian of West Siberia (Salair, Kuznetsk Basin,
Gorny [High] and Rudny [Ore] Altai). Senckenberg leth 86:
323–325
758 Facies (2012) 58:727–758
123





























