Embed Size (px)
Citation preview

Kaupia • Darmstadter Beitrage zur Naturgeschichte Heft 2 133 -144 Darmstadt, 15. Marz 1993
Xavier Martínez-Delclòs & Jordi Martinell
Insect Taphonomy Experiments. Their Application to theCretaceous Outcrops of Lithographic Limestones
from SpainAuthors' address: Xavier Martinez-Delclos & Jordi Martinell, Laboratori de Paleontologia, Facultat de Geologia,
Universitat de Barcelona, 08071 Barcelona, Spain.
ZusammenfassungUntersuchungen necrobiotischer und biostratinomischer Prozesse bei Insekten auf Wasseroberflachen wurden inspeziell entwickelten Aquarien durchgeführt. Die Ergebnisse der aktualistischen Arbeiten dienen zur Interpretation derFossilisationsprozesse die zur Insektenerhaltung in lithographischen Kalksteinen der Unterkreide Spaniens führten.
AbstractThe study of necrobiotic and biostratinomic processes affecting insects on being trapped in water are described. Speciallyconditioned aquariums were used for the laboratory experiments. The results obtained have been used to interpret thefossilization processes of insects found in the lithographic limestones of the Lower Cretaceous in Spain.
IntroductionThe "post-mortem" floating of fossil remains (ma-rine or terrestrial) has been studied by severalauthors (DENTON & GILPIN-BROWN 1966, CHAMBER-LAIN et al. 1981, FERGUSON 1985, SAUNDERS & SHAPIRO1986, HAMMANN et al. 1986, REYMENT 1986, etc.).
The agony and death of insects is not a subjectthat would normally interest the biologist. The onlystudies that can be cited for a description of actu-alistic observations on the behavior of insects trap-ped on the water surface are those of LUTZ (1984,1990).
Our biostratinomic observations help to explainthe fossilization process of insects and the environ-mental conditions.The present experiments wereinitiated to determine the nature of the taphonomicprocesses which had caused the excellent state ofpreservation of the insects of the Lower Cretaceousof Spain (MARTÍNEZ-DELCLÒS 1991).
These insects are found in lithographic lime-stone outcrops of lacustrine origin. They are lo-cated in two distinct areas. The first being at "LaSerra del Montsec, Lleida province" in the outcopsof "La Pedrera de Rúbies" and "La Cabrua" ofBerriasian-Valanginian age (MARTÍNEZ-DELCLÒS etal. 1991a) and the other being at "La Serranía deCuenca, Cuenca province" in the outcrop of "LasHoyas" of Barremian age (SANZ et al. 1988) (Fig. 1).
Insect BuoyancyThe present investigation has studied the behav-iour of different kinds of terrestrial insects on beingtrapped on the water surface by the surface tensionand the physical conditions that permit submersi-on and thus the settling on the bottom and later
fossilization. Therefore, the necrobiotic and bio-stratinomic processes have been studied exclusively.
Observations of the same characteristics havealso been made on aquatic insects (principally He-teroptera) or on those which during a period oftheir life may live in this habitat (odonate andmayflies nymphs).
For these studies a series of experiments werecarried out in the laboratory with aquariums ofvarious sizes, the largest being of 250 litres and thesmallest being of 25 litres (MARTINELL & MARTÍNEZ-DELCLÒS 1990).
In some of these, natural conditions were simu-lated by the incorporation of species from riversand lakes of different life habits (fish, amphibia,crustaceans, insect nymphs, etc.). Evidently, thefauna and flora which populated the lacustrineecozones during the Cretaceous are not at onesdisposal. Yet, the performance may be consideredcomparable in global terms, bearing in mind thatthe direct application of actualism must in somecases be done with caution.
In other tanks no disturbing agent of organic norinorganic nature was introduced, except themicroorganisms that the water may have containedor which developed therein.
A tank of 1500 litres was also installed (surfacearea of 3 m2) in the open air to facilitate the study ofthose insects which through natural causes fell intothe experimental tank. All experiments were car-ried out with still, oxygenated freshwater.
These experiments were carried out during thespring, summer and autumn of 1990 and 1991, and

134 Xavier Martínez-Delclòs & Jordi Martinell
Fig. 1: Geographical location of the lithographic limestones outcrops of Spain (Lower Cretaceous) and briefdescription of the evolution of the different lithofacies.
were suspended in winter due to the decrease in thenumber of insects.
The effects of rain, wind and waves on the insectremains were also observed. At times these ele-ments were created artificially. Both dead and liv-ing insects were introduced into the tanks. Thenecrobiotic behaviour of these and also the beha-viour of the other individual inhabitants of thetanks was observed.
Experimental ObservationsObservations were made periodically on each ofthe aquariums and in the large tank.
With respect to the terrestrial insects, observa-tions were made constantly from the beginning ofagony until death and afterwards until the definitesinking. This method was used to study thoseorganisms that died in the water and also those
introduced into the water already dead.With respect to the aquatic species, observations
were made from the time of definite death on-wards.
Insects of the following orders were controlledduring the experiment: Blattodea, Orthoptera, Hom-optera, Heteroptera, Ephemeroptera, Diptera, Hy-menoptera, Coleoptera, and Neuroptera. All thesefossilized in the lithographic limestones at "LasHoyas" and / or Montsec. The behaviour of Lepido-ptera and Dermaptera was also observed even thoughthey were not present in the deposits studied.
To explain the observations made on terrestrialinsects three stages were differentiated with all theprocesses that function in each one of them:- On the water surface.- During the sinking process.- On the tank bottom.

Insect Taphonomy Experiments 135
On the water surface
On falling into the water the insect is trapped by thesurface tension. The struggle to reach a solid sur-face and thus to be able to leave the water nowbegins. Behaviour varies according to insect group.
Two types of behaviour patterns are demon-strated by wingless insects when falling into thewater:
A) The first group includes those organismswith a low specific weight, inferior to that of water.The behaviour of individuals of this type has beenstudied solely with worker ants. On falling into thewater they move around freely on the surface. Asthey go forward the abdomen is raised in a rythmicmovement which may be interpreted as a means toachieve respiration. They arrive at the edges of thetanks without difficulty and leave without anyproblem.
B) The second group includes those organismswith a high specific weight, greater than that ofwater. Tests here were carried out on earwigs (Pl.l,Fig. A) and/or cockroaches. On falling into the wa-ter they overcome the surface tension without dif-ficulty and descend directly to the bottom stillalive.
Winged individuals on falling into the waterremain with the wings stuck to the surface. These,also are classified in two groups according to theircapacity to return to land or not.
1) The first group includes those of a small size(less than 5 mm wingspan): Homoptera, Dipteraand Hymenoptera; and those of a greater wingspanlike the Lepidoptera. When the organisms aretrapped by the surface tension they are unable toescape because of lack of sufficient strength to takeoff from the surface. Firstly they move their feet inan attempt to move forward, yet without success.Exhaustion arrives within a few moments and theonly signs of life observed are a weak movement ofantennae and palps. Death occurs within a timeperiod not surpassing one hour.
2) The second group consists of Orthoptera. Theypresent a behaviour markedly different. When fall-ing into the water they remain in a lateral positionwith the wings folded and it is impossible for themto move on the surface (PL 1, Fig. B). This observa-tion is very significant because it is in this very sameposition that the winged Orthoptera are generallyfound fossilized (outcrops at Solnhofen-EichstattUpper Jurassic, Germany), "Las Hoyas" and Mont-sec (Lower Cretaceous, Spain). In this group onecan also include some Hymenoptera, Diptera,Neuroptera, Coleoptera and Heteroptera.
The initial behaviour of these organisms is tomaintain the wings folded dorsally and move alongthe water surface yet with difficulty and withoutmuch advancement, afterwards the wings are un-folded and a period of repose takes place. Each time
the period of repose is longer in comparison to thatof activity. Finally, exhausted, wing movement isnow impossible and the wings remain in an open orunfolded position. Due to movement the insectsare soaked even more quickly. One can still observesigns of life because of antennae and palp move-ment. The time of agony does not last longer thantwo hours. It was also observed that at the momentof death the wasps direct the abdomen forward.This is a very typical posture found among some ofthe fossil Hymenoptera at Montsec (PL 1, Fig. C).
The representatives of the blatid group are thosethat endure the longest period of agony. The beha-viour pattern here is the same as for the abovesecond group, but the difference being that theperiod of agony can last up to one and a half days.Yet if on the water, the organism remains stillduring the time necessary to recuperate and thenraises itself and begins to move around normally.The organism moves with a great speed around thesurface yet without a fixed direction. If it is capableof arriving at one of the edges of the tank, it canleave the water. Others, due to the speed of themovement will find themselves drifting more andmore into the centre of the lake (PL 1, Fig. D).
The cause of death in all these organisms isasphyxia. According to BAUDOIN (1976) the terres-trial insects which fall into water remain stuck tothe water by their wings and death results fromasphyxia after obturation of the stigmata by thewater. Yet during the present experiments it wasobserved that many individuals are able to savethemselves because they are able to arrive at a solidsurface, raise themselves and thus continue withtheir normal activity. Others, in contrast, once falleninto the water are incapable of getting up and aretherefore condemned to death like, for example,butterflies.
Buoyancy periodThe floating time varies greatly according to thespecies under observation. As explained above allthe winged forms die on the water surface. Thefollowing stage of the experiment therefore was tocontrol the time that passes from the death of theinsect until the beginning of the sinking of theremains and to study the biostratinomic processesinvolved in this stage.
To complete the experiment at this stage, organ-isms which had died in the air were introduced intothe aquariums.
To explain the behaviour with respect to buoy-ancy, the insects are distributed into groups cha-racterized by similar responses.
A) Types which die in the air and then fall intothe water.
B) Types which die through asphyxia in the water.B.I) Small forms (wingspan smaller than 5 mm)
and Lepidoptera.

136 Xavier Martínez-Delclòs & Jordi Martinell
Plate 1:Fig. A: Fossil earwigspecimen (Dermaptera) ofthe Miocene from Rubielosde Mora, Teruel province).Fig. B: Domenechia marti-nelli, Orthoptera, fossilizedin a lateral position inwhich one can see theantennae in a fully arti-culated state (105 mmlong). Lower Cretaceousfrom "La Cabrua" Montsec.Fig. C: Hymenopter withabdomen bent forward.Lower Cretaceous from "LaPedrera de Rubies",Montsec.Fig. D: Mesoblattinacolominasi, Blattodea.Lower Cretaceous from "LaCabrua", Montsec.Fig. E: An image of a beearriving at the bottom ofthe aquarium. It still hasnot adopted the position ofmaximum stability.Observe the developmentof an fungus covering onthe abdomen.Fig. F: Aquatic Heteroptera(Balostomatidae). This isone of the predatoryinsects found in abundanceat the outcrop of "LasHoyas" (Cuenca).Fig. G: Wasp in the mo-ment of sinking.
B.2) Forms with a wingspan greater than 5 mmexcept Lepidoptera. Here all the winged formsclassified above in the second group plus theOrthoptera are included.
Wingless forms are not included in this classifi-cation mentioned before, no observations have beenmade of wingless forms dying on the water surface.
The behaviour with respect to floating time isvery different for forms of the same species depen-ding on whether the insect has been introducedinto the tank already dead or if it has fallen in alive.
The floating time is much longer in the first casethan in the second. In a very still medium, such asan aquarium, into which no living organism has
been introduced (bearing in mind that some micro-organisms may have developed through naturalprocesses), it was observed that an insect that diedin the air is capable of remaining on the surface fora period exceeding to six months without disarticu-lation.
The floating time of the remains of small insectsand Lepidoptera (under observation in the tank) isvariable, yet the factor that characterizes thesegroups is the inability to sink. They remain on thesurface where disarticulation, fragmentation anddecomposition takes place.
Lepidoptera have scaled wings which becomesoaked easily in the water (PL 2, Fig. A-C). On

Insect Taphonomy Experiments 137
Plate 2: Lepidoptera decomposition sequence in 15 days, in lentic water, without any physical disturbating factor(wind or rain).Fig. A: Early killed Lepidoptera remain over the water surface.Fig. B: 5th day: wings and abdomen are affected by decompositon.Fig. C: 10th day: wing scales disappear.Fig. D: 15th day: wings are practically missing.
falling into the water it is impossible for them tocounteract the surface tension, even having at theirdisposition a solid object. The adhesion surface isvery large in comparison with the total volume ofthe organism. Also, the adhesion force (the forceopposing the separation of two bodies in contact) isso strong that it cannot overcome the force of thesurface tension, neither in a positive nor negativesense. After two days in the floating state the abdo-men shows an advanced state of decompositionand a lack of scales is observed on areas of thewings. Whatever biological or physical factor (rain,wind, waves, etc.) that disturbs the medium pro-vokes the fragmentation of the remains, it is impos-sible to identify the parts that have rapidly decom-posed (PL 2, Fig. D). It is not surprising thereforethat the fossil record of Lepidoptera is so poorbecause the taphonomic processes that permit theirpreservation must be of a special nature (amberdeposits, cinerites). However such fossils have beenfound in lacustrine sediment, for example at Cerestein France (LUTZ 1984, NEL 1991). The conservation in
these deposits can be explained by the activity of asuperficial algal mat that may collaborate in thepreservation and burial of the remains.
With respect to the small insects a similar behav-iour is observed. Sporadically their remains arefound on the bottom of the tanks, while on thesurface the quantity is very high. For every twentyforms with a wingspan over 5 mm that may becounted on a 3 m2 surface area up to 500 of the smallforms may appear. The quantities were controlledevery two days over a two week period and theratio of large and small forms found decreasesmarkedly after a day of rain or wind. These causewaves in the tank which in turn are responsible forthe disarticulation and fragmentation of the re-mains of the very small forms, similar to thatoccuring with the Lepidoptera. The small frag-ments that could correspond to the remains ofsmall insects cannot be fully identified. At theMiocene outcrops at "Rubielos de Mora" (Teruel,Spain) important accumulations of small insectshave appeared (MARTÍNEZ-DELCLÒS et al. l991b)com-

138 Xavier Martínez-Delclòs & Jordi Martinell
Plate 3: Wasp sinking sequence in 10 days.Fig. A: The wasp is trapped by the water surface tension, and it produces waves.Fig. B: 3th day: fungus and algal development mainly in the abdomen.Fig. C: 5th day: the fungus covering spreads to the rest of the body.Fig. D: 8th day: the wasp is totally embedded by the fungi.Fig. E: 10th day: the ensemble fungi plus wasp overpasses the water surface tention and sinks to the bottom.
pletely intact. They have been conserved, such asthe Lepidoptera, due to the action of algal mats onthe surface.
The remains with a wingspan greater than 5 mm,in conditions of complete stillness are capable ofremaining on the surface for 4 to 14 days. The mostimportant factor causing the increase in specificweight of the remains is the degree of decomposi-tion of the soft parts and the content of watercontained internally. It has been stated above thatthe cause of death in the insect is asphyxia. This isvery interesting because the trachea remain full ofwater. While the insect is still alive it attempts tobreathe and it can be deduced that the same processthat permits air to enter the trachea also allowswater to enter there. This system is very well devel-oped in some insects (through air bags, complextracheal systems, etc.), and on allowing the body tobecome partially filled with water the total weightof the insect increases considerably. To prove theentry of water in the trachea, living insects wereintroduced into water dyed with black chlorazolindicator. When the insect had died various micro-scopic smears were made; it was observed thatwater had entered the system. This process alsoserves to explain the different buoyancy times forthose organisms that die in the water and those thatdie in the exterior.
In other cases sinking is induced due to someexternal biological or physical agent which pro-
duces a change in the surface tension and at thesame time produces a force that pushes the remainsto the bottom.
A wasp that had been floating on the watersurface for one and a half weeks was used as asupport by another wasp that flew into the tank.This second wasp remained trapped on the surface,but the first wasp was then able to overcome thesurface tension and sink to the bottom (PI. 3, Fig. E).
The insects or parts of insects that remain float-ing on the surface for various days also cause thedevelopment of fungi (Saprolegnia sp.) and/or al-gae in their immediate surroundings (PL 3, Fig. B-D). That increases their weight and thus helps theinsect to overcome the surface tension and causesinking (PL 3, Fig.E). These fungi and / or algae alsoact as a type of cohesion mechanism for the remainsthat impedes fragmentation and disarticulation (PL1, Fig. E).
The physical factor mentioned could be lightwaves produced by wind or rain. It is normal, forexample, that after a heavy rainfall practically noinsects are left in the experimental tank.
The relationship between volume and body sur-face area of the insect is an important factor thatdetermines the ability to float. So, for example,wingless individuals are able to sink almost instan-taneously, in some cases even dying on the bottom(for example, earwigs). Those with wings of a largesurface area are capable of decomposing on the

Insect Taphonomy Experiments 139
water surface without ever sinking, especially ifthey remain there with wings extended; for exam-ple Lepidoptera. As decomposition progresses, thesoft parts of the body slowly become filled withwater increasing therefore the insects weight andcausing, in some cases, sinking.
Both the tank and the aquariums may be re-garded as still environments, in which the biologi-cal activity is seen as the most important factorprovoking disarticulation and/or fragmentation.Diverse groups of aquatic organisms exist that mayattack the insects during the agony time or afterdeath (other insects, fish, amphibia, etc.). The move-ments made during the agony stage in the waterproduce waves that are captivated by aerial oraquatic organisms. Among the aerial groups thatare important here are other insects and smallbirds. Aquatic organisms of importance here inclu-de the nektonics, principally fish and aquatic in-sects. It is known that for example during theLower Cretaceous there was an important numberof fish in the lake at "La Cabrua" (Lleida province,Spain). The absence of insects with a predatoryfeeding habit (as for example the present Notonectaand Nepa) transfers importance to the Leptolepides(small and very numerous fish in the deposit). Theaquatic Heteroptera were also important predatorsin the deposit at "Las Hoyas" (Cuenca province,Spain) (PL 1, Fig. F).
It was observed that the attack in some casesoccured at random and was selective in other cases.One can cite here the case of attacks on the abdomenand other soft parts of the body and the rejection ofthe wings and thorax. This, however, does notimply that all the wings found in the fossil recordwere caused by this process. One must bear in mindthat the wing-thorax joints are especially fragile. Inwhatever case the attack produces the fragmentsthat potentially may sink. It was also observed inthe aquariums that traumatisms of the Triops typeof lacustrian crustaceans caused the sinking of onepart of the body of the insect while the other partremained floating.
During sinkingThe descent in a still medium is slow by the major-ity of the individual corpses observed and alwaysin a straight line, without modification of the initialsinking position. It was possible to observe this invarious groups, principally in Diptera and Hy-menoptera (PI. 1, Fig. G).
During this period of time, the insects are sus-ceptible to the same biological processes whichthey may have encountered on being attacked byaquatic organisms on the water surface.
In natural aquatic environments the thermoclina,which separates the warm water of the surfacefrom the colder waters at the bottom, is not only aboundary of different temperatures but also of
density. Many insects and other organic material,which are less dense than the lower cold layer, aretrapped here (MARGALEF 1983) inhibiting their po-tential fossilization.
On the bottomIt was observed in the laboratory that the insectswhen reaching the oxygenated bottom are capableof remaining there for at least one year withoutbecoming disarticulated. This, of course, alwayswith the premise that no biological or physicaldisturbing agents exist. On the contrary, the activ-ity will be more intense when the degree of decom-position of the insect is higher. Moreover, even inthe case of the remains being on the bottom withoutthe presence of the disturbing agents mentionedabove or of chitinofag bacteria, with the passing oftime disarticulation occurs around the joint zones,given that the skeleton in these areas is less sclerotic.
Crustaceans, molluscs and fish are the principalbiological agents that are capable of causing disar-ticulation and/or fragmentation; currents are theprincipal physical agent.
The sediments where the insect fossils are gener-ally found have certain characteristics which indi-cate that they were deposited in an anoxic environ-ment (absence of bentonic fauna, traces of displace-ment, etc.). This leads one to believe that the disar-ticulation and/or fragmentation takes place dur-ing the floating and sinking periods in the oxygen-ated area.
The decomposition of the insect tegument iscaused by the activity of chitinofag bacteria thatlive almost exclusively in oxygenated zones, yetsome of these species are capable of anaerobicmetabolism. Up to the present, no type of destruc-tive process of the tegument that may be attributedto bacteria of this kind has been observed withcertainty in the remains of insects from the lithogra-phic limestones of "Montsec" and "Las Hoyas"(Spain).
Taphonomic observations in the litho-graphic limestones from the SpanishLower CretaceousIn this section preferential treatment is given to theLower Cretaceous outcrop at "La Cabrua" (Lleidaprovince, Spain) as it is the one that has beenstudied systematically for the longest time andconsequently provides most taphonomic field data.
References of a taphonomic nature are made tothe Cretaceous outcrops at "La Pedrera de Rubies"(Lleida province, Spain) and "Las Hoyas" (Cuencaprovince, Spain) when considering specific groups.
Causes of DeathThe death of the terrestrial insects is caused, gener-ally, by asphyxia which in turn is caused by the

140 Xavier Martínez-Delclòs & Jordi Martinell
water lodged in the trachea of the insect which istrapped on the water surface by the surface tension.The current experiments have shown that the in-sects which die in sub-aerial conditions and aresubsequently transported into the water possessfew possibilities of overcoming the surface tensionand sink.
Direct evidence of death is not available and it isvery difficult to deduce such proofs from compari-sons with actualistic experiments. Whatever inter-pretation proposed would be of pure speculation.The death of an aquatic insect may result fromvarious causes (a change in environmental condi-tions, depredation and causes pertinent to the or-ganism itself), but these causes condition the fos-silization to a quite lesser degree than in the case ofterrestrial insects. In the case of the latter the prin-cipal condition affecting possible preservation isdeath by asphyxia in water.
Place of deathTwo possibilities exist:
A) The insects die in the same lacustrine environ-ment in which they were buried.
B) The remains were transferred immediatelyafter death or during the agony from the palustrineenvironment that surrounded the lake.
At any rate the first option is the most probabledue to the high degree of preservation of delicatestructures (antennae, ovipositor, etc.) which evenon being moved very lightly, are probable to break(PL 4, Fig. A).
For the remains of aquatic insects it is difficult todetermine the place of death by taphonomic crite-ria. The high degree of intact joints and of conserva-tion in general is not a direct indicator of absence oftransport. In any case the proximal fades of thelacustrine environment seems to be positive for thepreservation of some taxa, as for example Meso-palingea and Palaeaeschna, both found in high num-bers in the deposits at Montsec (MARTINEZ-DELCLÒS
1991), or for example Iberonepa from "Las Hoyas".These three genera are represented in the depositsby a long series of ontogenetic stages, apart fromthe two types of remains: those obtained by thedeath of the individual or by its biological activity(moulted skin). It can be deduced from these consi-derations that at least these three genera lived nearthe place of death. For the other aquatic forms it ismore difficult and at present impossible to deter-mine the place of death.
Degree of articulationThe insect fossils of the lithographic limestones out-crops at Montsec and "Las Hoyas" are representedin two ways according to the degree of articulation.
The more common are complete forms with alljoint areas connected as for example many speci-mens of: Mesopalingea leridae, Palaeaeschna vidali (PI.
4, Fig. B), Huergoneta ciervaensis and Iberoneparomerali, among other aquatic forms; Chresmodaaquatica as a semi-aquatic form (PI. 4, Fig. C);Meiatermes bertrani, Mesoblatina colominasi (PI. 1,Fig. D), Pachypsyche vidali, Ilerdocossus villaltai, Iler-daegomphus torcae, among other terrestrial forms.
It is less common to find incomplete forms. Iso-lated parts of the body are present or only certainjoints remain intact with the body, such as: man-dibular structures of Mesopalingea leridae; head andwings of Meiatermes bertrani (Pl. 4, Fig. D); wing ofIlerdocossus villaltai; an abundance of elytra of Col-eoptera; abdomens, feet and wings of blattids; wingsof odonats: Condalia woottoni, Iberoaeschnidiumconquensis, Gigantoaeschnidium ibericus; wings ofIberonepa romerali sporadically and wings ofOrthoptera among others. On the basis of the iso-lated remains one can, at times, only indicate that itis an insect that is under discussion but in otheroccasions, due to the general knowledge of theinsect fossils from the deposits, it is possible toclassify the to orders and, on the basis of feet andabdomens of blattids or wings of Orthoptera, forinstance finally to the species. Some of the incom-plete remains are very characteristic of the outcropsand demonstrate that some forms are disarticu-lated easily in determined zones; for example themandibular structure of Mesopalingea leridae; elytraof Coleoptera; wings of Odonata; feet, wings andabdomens of Blattodea and wings of Kalligram-matidae (fossil Neuroptera).
Degree of fragmentationThe cases of fragmentation of wings from the out-crops studied are irregular. This process is the onlyone that does have a bibliographical reference (LUTZ
1984). It is observed that the fragmentation of wingsof blatids has a constant pattern: they break alongthe back posterior cubital vein (CuP). This veinpasses through a very defined ridge, that is to saythrough a zone where a considerable narrowing ofthe cuticle occurs which limits zones of differentconsistency, therefore a mechanically weak zonemust exist. It has also been observed in Wonnacottel-la pulcherrima (PL 4, Fig. E), Pachypsyche vidali and inisolated wings of Iberonepa romerali.
Resulting fossilization positionIn each group of insects a characteristic fossiliza-tion process is observed.
The majority of the groups studied remain in adorsal or ventral position when fossilized: nymphsof Ephemeroptera and Plecoptera and adults andnymphs of Odonata, adults of Heteroptera, Homo-ptera, Isoptera, Blattodea, Paraplecoptera (Chres-moda aquatica) and some Coleoptera. The Orthopteraare found in a lateral position. The Hymenopteravary their position and both lateral and dorsalposition forms are found.

Insect Taphonomy Experiments 141
Plate 4:Fig. A: Ephialtites jurassicus,Hymenoptera in which theovipositor has been conser-ved. Lower Cretaceous from"La Pedrera de Rubies"Montsec.Fig. B: Palaeaeschna vidali, aperfectly conserved odonatnymph. Lower Cretaceousfrom "La Pedrera de Rubies"Montsec.Fig. C: Chresmoda aquatica,Paraplecoptera, a semi-aquatic adult form. LowerCretaceous from "La Pedrerade Rubies" Montsec.Fig. D: Meiatermes bertrani,Isoptera, with the presenceof two wings joined to athorax fragment. LowerCretaceous from "La Pedrerade Rubies" Montsec.Fig. E: Wonnacottellapulcherrima, Homoptera.Observe how the clavusremains separated from therest of the wing (arrow).Lower Cretaceous from "LaPedrera de Rubies" Montsec.Fig. F: Torcagryllus apexre-ditus, Orthoptera. Observethe dorsal position and thegeneral dismembered stateof the individual. It ischaracteristic among thisspecies to find wings woundup at the apex.Fig. G: Chresmoda aquatica.A detail from the distal partof the abdomen in which onecan see the dismembering ofone of the segments (arrow).Fig. H: Ilerdocossus villaltai,Homoptera showing tracesof the original stripedcolouring on the thorax andwings. Lower Cretaceousfrom "La Pedrera deRubies", Montsec.
G
It has been observed in the laboratory experi-ments that some Hymenoptera (wasps) bend theabdomen forward on dying, (PL 1, Fig. G) and thisis the position acquired at the moment of death, andis the one that conditions mainly the stability on thebottom. During the sinking time this floating posi-tion is maintained, that is to say with the wingsunfolded above the superior part of the body; theremains may stay in this position for some days andonly the feet and abdomen repose almost imper-ceptibly on the sediment. Later, the remains turnlaterally and the result is that a greater surface
comes into contact with the sediment; in this posi-tion it will remain buried. It would be very interest-ing to verify this behaviour in the Hymenoptera atMontsec and "Las Hoyas" in addition to the actu-alistic comparisons, to find the traces made in theinitial moment of contact with the bottom; unfortu-nately such traces have never been seen at thelithographic limestones from the Spanish Creta-
ceous.The Orthoptera are also fossilized in a lateral
position, yet this position of maximum stability,unlike in the case of the Hymenoptera, is acquired

142 Xavier Martínez-Delclòs & Jordi Martinell
during the buoyancy time on the surface.The fossilized forms with a dorsal position also
maintained this position from the moment of beingtrapped by the surface tension.
The constancy in the fossilization position (inaddition to other taphonomic observations) informsus of the lack of strong currents in the basin thatcould have varied the position of maximum stabil-ity that the different insect show.
Isolated cases exist in which the fossilizationposition is not representative of the group, forexample an Orthoptera and some specimens whichone would have expected to find in a dorsal posi-tion are in fact found in a ventral position. In thesecases the sinking of the remains or their arrival atthe deepest parts of the basin were carried out, forcertain, by sedimentary occurences that altered thenormal conditions of sedimentation, for exampleturbiditic flows (PI. 4, Fig. F).
DecayThe insect fossils of the lithographic limestone de-posits at Montsec and "Las Hoyas" may conservethe cuticle but remains of musculature or soft partshave never been observed. Decay progressed untilthe destruction of these structures, before the earlymineralization process was able to induce theirconservation.
In some specimens the cuticle was conserved,yet it is probable that the composition of this wasnot the original one. The techniques used to deter-mine the composition have not given results. Thereare specimens such as casts in which there is notrace of cuticle found. In these cases it can beconsidered that the cuticle disappeared due to asecondary process not connected with the chemi-stry nor the bacteria of the environment. This de-struction process took place later than the sedimentcompactation process, moreover the remains wouldnot be conserved in mould form.
A clear example of decomposition that has beenseen is that of Chresmoda aquatica. The remains arevery well articulated yet one of the segments of theabdomen is displaced slightly from the originalposition (PL 4, Fig. G). The remains, before burial,surely spent a period of time on the bottom and thedecay of the abdomen began with the zones ofconnection with the front and back parts and aweak force (light current on the bottom) moved it.Decay did not progress, probably because the re-mains were buried and the mineralization processbegan.
Another example observed was one specimenfrom "Las Hoyas". This refers to an adult aquaticHeteroptera that has not been classified with greaterdetail because the state of conservation of the speci-men does not allow it. However, the mandibularstructure is observed to be displaced from the origi-nal position. It is a stage of decomposition of the
incipient cuticle that does not progress, probablydue to the burial of the remains and the beginningof the mineralization process.
The insect fossils of the lithographic limestonedeposits of Montsec and "Las Hoyas" stand out forthe lack of decomposition of the cuticle. It is diffi-cult to cite more examples of this taphonomic pro-cess because it is sporadic in the forms at ourdisposal.
ConclusionsThe experiments carried out in aquariums andtanks with insects permitted us to study what hap-pened when they were trapped on the surface bythe surface tension. All actualistic experiments werecarried out with still, oxygenated freshwater.
A first series of experiments were carried out ina tank that lacked any kind of activity of otherorganisms. The insects that fell into the water diedby drowning and floated on the surface due to thesurface tension. They were able to float for periodsof time superior to one month conserving them-selves totally articulated and with no loss of colour(PI. 4, Fig. H).
If the insect is capable of overcoming the super-ficial tension, due to its own weight or due to somephysical agent (rain, wind) it will begin to descend.It has been observed that those insects with a largewingspan (butterflies for example) find it impossi-ble to descend to the bottom due to the effects of thephenomenon explained before. The decomposi-tion takes place therefore on the water surface. Thisprobably explains why those insects are rarely foundin fossil deposits. The same phenomenon was alsoobserved in individuals of small size (less than5 mm), these float and do not overcome the surfacepressure. This is not caused by its weight (ex-tremely small), nor by any outside pressure. It isnecessary to think of some other phenomenon thatpermits sinking. The explanation in these casesmay be found in the formation of floating algalmats which when found on the surface of the watermay lump together and so help in the preservationof the remains.
The fossilization of terrestrial insects is highly con-ditioned by the type of death undergone, by their sizeand alar surface and the relationships of all thisfactors. It has been observed experimentally that theinsects that die by asphyxia in the water and that havedetermined sizes are potentially capable of overcom-ing the surface tension and sinking.
Moreover, very different results are observedwhen the insects are introduced into aquariumswith other living organisms (fish, crustaceans, otherinsects). The activity of these organisms aids thedisarticulation and fragmentation of the insects,(terrestrial or aquatic) that have died in the water.
All the remains of fossil insects from the LowerCretaceous lithographic limestones from Spain
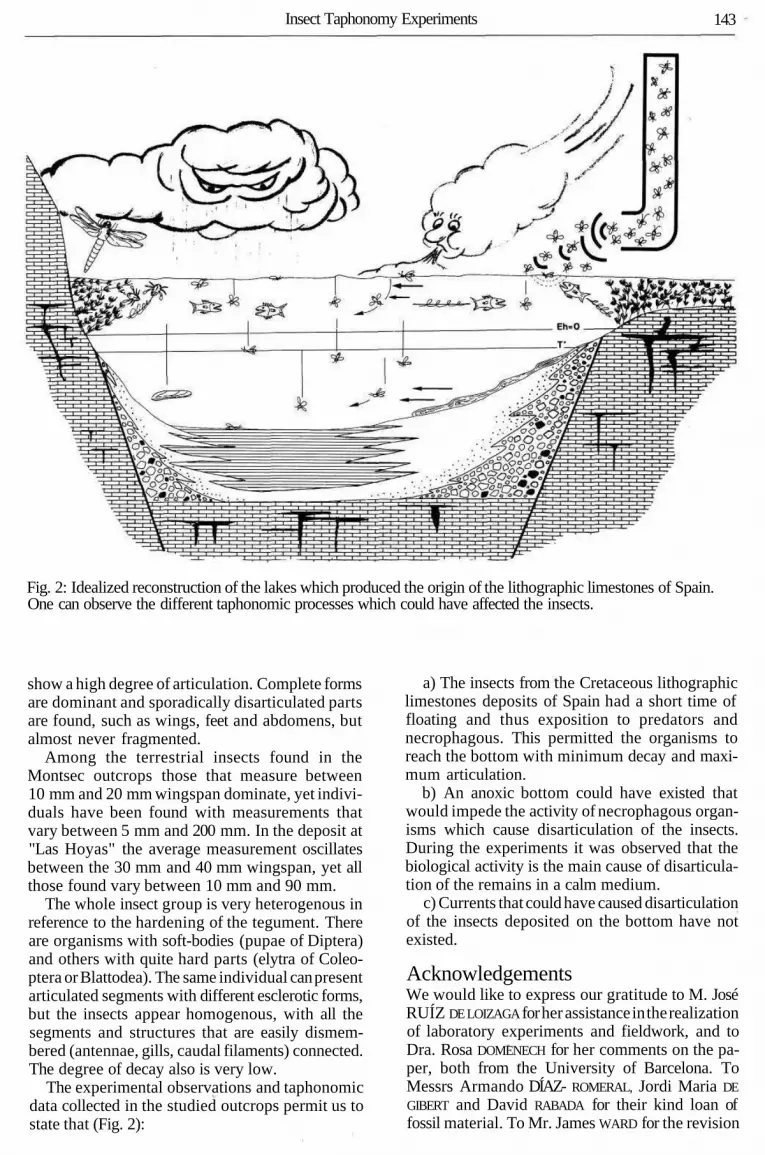
Insect Taphonomy Experiments 143
Fig. 2: Idealized reconstruction of the lakes which produced the origin of the lithographic limestones of Spain.One can observe the different taphonomic processes which could have affected the insects.
show a high degree of articulation. Complete formsare dominant and sporadically disarticulated partsare found, such as wings, feet and abdomens, butalmost never fragmented.
Among the terrestrial insects found in theMontsec outcrops those that measure between10 mm and 20 mm wingspan dominate, yet indivi-duals have been found with measurements thatvary between 5 mm and 200 mm. In the deposit at"Las Hoyas" the average measurement oscillatesbetween the 30 mm and 40 mm wingspan, yet allthose found vary between 10 mm and 90 mm.
The whole insect group is very heterogenous inreference to the hardening of the tegument. Thereare organisms with soft-bodies (pupae of Diptera)and others with quite hard parts (elytra of Coleo-ptera or Blattodea). The same individual can presentarticulated segments with different esclerotic forms,but the insects appear homogenous, with all thesegments and structures that are easily dismem-bered (antennae, gills, caudal filaments) connected.The degree of decay also is very low.
The experimental observations and taphonomicdata collected in the studied outcrops permit us tostate that (Fig. 2):
a) The insects from the Cretaceous lithographiclimestones deposits of Spain had a short time offloating and thus exposition to predators andnecrophagous. This permitted the organisms toreach the bottom with minimum decay and maxi-mum articulation.
b) An anoxic bottom could have existed thatwould impede the activity of necrophagous organ-isms which cause disarticulation of the insects.During the experiments it was observed that thebiological activity is the main cause of disarticula-tion of the remains in a calm medium.
c) Currents that could have caused disarticulationof the insects deposited on the bottom have notexisted.
AcknowledgementsWe would like to express our gratitude to M. JoséRUÍZ DE LOIZAGA for her assistance in the realizationof laboratory experiments and fieldwork, and toDra. Rosa DOMENECH for her comments on the pa-per, both from the University of Barcelona. ToMessrs Armando DÍAZ- ROMERAL, Jordi Maria DEGIBERT and David RABADA for their kind loan offossil material. To Mr. James WARD for the revision

144 Xavier Martínez-Delclòs & Jordi Martinell
of the English text. This study has been partiallyfunded by the "Institut d'Estudis Ilerdencs"through an investigation grant, awarded in 1990. Itis part of the projects DGICYT PB-88 and IntegratedAction HF-84A.
ReferencesBAUDOIN, R. (1976): Les insectes vivant a la surface et
au sein des eaux. -In: GRASSE, P.P. [ed.]: Traite deZoologie, T. VIII, Insectes IV, Splanchnologie,phonation, vie aquatique, rapports avec les plan-tes: 843-926; Masson, Paris.
CHAMBERLAIN, J.A. jr., WARD, P.D. & WEAVER, J.S.
(1981): Post-mortem ascent of Nautilus shells:implications for cephalopod paleobiogeography.- Paleobiology, 7(4): 494-509.
DENTON, E.J. & GILPIN-BROWN, J.B. (1966): On thebuoyancy of the pearly Nautilus. - Journal ofMarine Biologists Association, 46: 723-759.
FERGUSON, D.K. (1985): The origin of leaf-assem-blages. New light on an old problem. - Review ofPaleobotany and Palynology, 46:117-188.
FREGENAL, M.A. (1990): El Sistema lacustre de LasHoyas (Cretacico inferior, Serrania de Cuenca):Estratigraf ia y Sedimentologia: 226 pp. -Tesis deLicenciatura, Universidad Complutense de Ma-drid.
HAMMANN, W., RABANO, I. & GUTIERREZ-MARCO, J.C.
(1986): Morfologia funcional del exoesqueletodel genero Selnopeltis HAWLE & CORDA, 1847
(Trilobita, Odontopleurida; Ordovicico). - Pa-leontologia i Evolucio, 20: 203-211.
LUTZ, H. (1984): Parallelophoridae -isolierte Anal-felder eozaner Schaben (Insecta: Blattodea). -Palaontologische Zeitschrift, 58 (1/2): 145-147.
LUTZ, H. (1990): Systematische und palokologischeUntersuchungen an Insekten aus dem Mittel-Eozan der Grube Messel bei Darmstadt. - Cou-rier Forschungsinstitut Senckenberg, 124: 165pp.; Frankfurt am Main.
MARGALEF, R. (1983): Limnologia. - Ed. Omega:1010 pp.; Barcelona.
MARTINELL, J. & MARTINEZ-DELCLOS, X. (1990):Observaciones sobre la flotabilidad de losinsectos. - Comunicaciones de la Reunion deTafonomia y Fosilizacion: 201-210; Madrid.
MARTINEZ-DELCLOS,X. (1991): Insectes hemimetabolsdel Cretaci inferior d'Espanya. Tafonomia i Pa-leoautoecologia: 784 pp..-Tesi Doctoral Univ. deBarcelona.
MARTINEZ-DELCLOS, X., BARALE, G., WENZ, S., DOME-
NECH, R., MARTINELL, J., MERCADE, L. & Ruiz DE
LOIZAGA, M.J. (1991a): The outcrops of lithogra-phical limestones of Montsec (Catalonia, Spain).Current state of knowledge. - In: MARTINEZ-
DELCLOS, X [coord]: The Lower Cretaceous litho-graphic limestones of Montsec. Ten years ofpaleontological expeditions. - Institut d'EstudisIlerdencs Ed: 101-106; Lleida, Spain.
MARTINEZ-DELCLOS, X., PENALVER, E. & BELINCH6N, M.
(1991b): Primeras aportaciones al estudio de losinsectos del Mioceno de Rubielos de Mora, Teruel(Espana). - Revista Espafiola de Paleontologia,Volumen Extraordinario: 125-137.
NEL, A. (1991): Analysies d'entomofaunes cenozo-iques interets de la paleoentomologie pour lessciences de la Terre et de la Vie: 882 pp. - Thesede Doctorat; Univ. Reims-Champagne-Ardenne.
REYMENT, R.A. (1986): Nekroplanktonic dispersal ofechinoid tests. - Palaeogeography, Palaeoclima-tology, Palaeoecology, 52: 347-349.
SANZ, J.L., WENZ, S., YEBENES, A., ESTES, R., MARTINEZ-
DELCLOS, X., JlMENEZ-FUENTES, E., DlEGUEZ, C,
BUSCALIONI, A.D., BARBADILLO, L.J. & VIA, L. (1988):An Early Cretaceous faunal and floral continen-tal assemblage: Las Hoyas fossil-site (Cuenca,Spain). - Geobios, 21(5): 611-631.
SAUNDERS, N.B. & SHAPIRO, E.A. (1986): Calculationand simulation of ammonoid hydrostatics. - Pa-leobiology, 12 (1): 64-79.





























