Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/98/$04.0010
Nov. 1998, p. 6679–6697 Vol. 18, No. 11
Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Cell Cycle-Regulated Expression of Mammalian CDC6Is Dependent on E2F
GUUS HATEBOER,1† ALBRECHT WOBST,1 BIRGIT OTZEN PETERSEN,1 LAURENT LE CAM,2
ELENA VIGO,1 CLAUDE SARDET,2 AND KRISTIAN HELIN1*
Department of Experimental Oncology, European Institute of Oncology, 20141 Milan, Italy,1 andInstitut de Genetique Moleculaire, CNRM, UMR 5535, 34033 Montpellier Cedex 1, France2
Received 20 February 1998/Returned for modification 13 April 1998/Accepted 18 August 1998
The E2F transcription factors are essential regulators of cell growth in multicellular organisms, controllingthe expression of a number of genes whose products are involved in DNA replication and cell proliferation. InSaccharomyces cerevisiae, the MBF and SBF transcription complexes have functions similar to those of E2Fproteins in higher eukaryotes, by regulating the timed expression of genes implicated in cell cycle progressionand DNA synthesis. The CDC6 gene is a target for MBF and SBF-regulated transcription. S. cerevisiae Cdc6pinduces the formation of the prereplication complex and is essential for initiation of DNA replication.Interestingly, the Cdc6p homolog in Schizosaccharomyces pombe, Cdc18p, is regulated by DSC1, the S. pombehomolog of MBF. By cloning the promoter for the human homolog of Cdc6p and Cdc18p, we demonstrate herethat the cell cycle-regulated transcription of this gene is dependent on E2F. In vivo footprinting data demon-strate that the identified E2F sites are occupied in resting cells and in exponentially growing cells, suggestingthat E2F is responsible for downregulating the promoter in early phases of the cell cycle and the subsequentupregulation when cells enter S phase. Our data also demonstrate that the human CDC6 protein (hCDC6) isessential and limiting for DNA synthesis, since microinjection of an anti-CDC6 rabbit antiserum blocks DNAsynthesis and CDC6 cooperates with cyclin E to induce entry into S phase in cotransfection experiments.Furthermore, E2F is sufficient to induce expression of the endogenous CDC6 gene even in the absence of denovo protein synthesis. In conclusion, our results provide a direct link between regulated progression throughG1 controlled by the pRB pathway and the expression of proteins essential for the initiation of DNA replication.
Although E2F was originally defined as a factor that bindsspecifically to an element in the adenovirus E2 promoter (42),it is now evident that E2F is essential for coordinating tran-scription during the mammalian cell cycle (for reviews, seereferences 12 and 72). A number of genes are found to beregulated by E2F, particularly during the transition from G1 toS phase. To date, six members of the E2F family are known:E2F-1 through E2F-5 and the recently identified E2F-6 (11).Furthermore, two heterodimerization partners of the E2Fs,DP-1 and DP-2, have been isolated. The E2F transcriptionfactors appear to be key downstream targets for the retino-blastoma protein pRB and two pRB-related proteins, p107 andp130 (reviewed in references 12 and 70). Binding of pRBfamily members (also called pocket proteins) to the E2F tran-scription factors results in transcriptional repression of E2F-regulated genes. Phosphorylation of the pocket proteins bycyclin-dependent kinases releases the pocket proteins fromE2F, leading to derepression and/or activation of E2F-depen-dent genes and subsequent entry into S phase. The demonstra-tion that deregulated E2F activity is sufficient to induce Sphase in quiescent cells has provided a model for how theinactivation of the pRB pathway in human tumors leads to thedevelopment of cancer (70).
The majority of E2F-regulated genes encode proteins thatare involved in DNA replication and in cell cycle progression(27, 72). These genes include DNA polymerase a (60), thymi-dine kinase (18), HsOrc1 (58), dihydrofolate reductase (7, 50,
71), cyclin A (35, 67), cyclin E (9, 22, 59), p107 (82), B-Myb(44), CDC2 (14, 75), E2F-1 (33, 38, 56), and E2F-2 (68). Atpresent it is not known how different E2F-DP complexes co-ordinate the expression of these genes. It is also relativelyunclear which gene is regulated by which heterodimer in whichphase of the cell cycle. However, several data suggest thatduring different phases of the cell cycle, specific subsets of E2Fand DP are functionally present. Importantly, the E2F familymembers can be divided into two subgroups based on theirability to bind pocket proteins, transactivation capacity, struc-ture, and expression during the cell cycle (for reviews, seereferences 2 and 72). E2F-1, -2, and -3 (one subgroup) arepotent transactivators and bind preferentially to pRB (29, 45),whereas E2F-4 and -5 (the other subgroup) are weak transac-tivators which are able to bind all pocket proteins (3, 23, 31, 37,51, 66). E2F-1, -2, and -3 are expressed mainly in late G1 andearly S phase, while the expression of E2F-4 is constant duringthe cell cycle. Recently, it was reported that the members ofthe two subgroups can also be distinguished functionally:whereas E2F-1, -2, and -3 are capable of inducing S phase inquiescent fibroblasts, E2F-4 and E2F-5 are not (49). The func-tional difference between the two subgroups is a result ofdifferent subcellular localization for the E2Fs, which appearsto regulate the activity of E2F-4 and E2F-5, but not the otherE2Fs that are found in the nucleus when expressed (48, 53).
Although E2Fs have been reported to regulate several geneswhose products participate in DNA replication (see above), itis clear that many of these genes are only marginally upregu-lated by deregulated E2F expression (15, 36, 76). Moreover,since most of the proteins participating in DNA replication arevery stable, it is not clear why the transcription of these genesneeds to be cell cycle regulated. None of the known geneproducts alone is able to induce S phase, suggesting that com-
* Corresponding author. Mailing address: Department of Experi-mental Oncology, European Institute of Oncology, Via Ripamonti435, 20141 Milan, Italy. Phone: 39 02 5748 9860. Fax: 39 02 5748 9851.E-mail: [email protected].
† Present address: IntroGene, 2333 AL Leiden, The Netherlands.
6679
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

binations of two or more products are needed or that theresponsible and limiting targets which can regulate the initia-tion of DNA replication have not yet been identified.
In eukaryotes, the initiation of DNA replication is a highlyregulated process which implicates a large number of proteins.These proteins are assembled at the origins of replication andform the prereplication complex (for reviews, see references 8,16, 55, and 73). Most of our knowledge regarding the regula-tion of eukaryotic DNA replication stems from the buddingyeast Saccharomyces cerevisiae, but a substantial amount ofdata has also been generated for Xenopus laevis and the fissionyeast Schizosaccharomyces pombe as experimental systems. InS. cerevisiae, the origin of replication is bound by a set ofproteins called the origin recognition complex (ORC). ORC isbound to the origin throughout the cell cycle and consists of sixpolypeptides (5) that are required for DNA replication and celldivision. The prereplication complex is formed at the end ofmitosis, and the complex formation is initiated by the associ-ation of Cdc6p with ORC (47), followed by the binding ofanother set of six related proteins, Mcm2 to Mcm7 (17, 74).After the complex formation, it is thought that components inthe complex are phosphorylated by the S-phase CDKs (Clb5por Clb6p in association with Cdc28p) and the Dbf4p/Cdc7pkinase, which leads to initiation of DNA replication (73).
Recently, it was shown that the binding of Mcm proteinsonto the origin is dependent on Cdc6p binding to ORC (17,74). Although ORC remains stably attached to the origins ofreplication during other phases of the cell cycle, Cdc6p andMcm proteins do not. When Cdc6p is overexpressed, it canbind to ORC throughout the cell cycle; however, the binding ofMcms to the ORC-Cdc6p complex during G2 and M phase isinhibited, most likely due to inhibitory effects from S-CDKprotein kinases (74). This strongly suggests that Cdc6p plays acentral and limiting role in the onset of DNA replication in S.cerevisiae. Cdc6p is a protein with a sequence similar to that ofthe large subunit of ORC, and it has a very short half-life, anumber of consensus sites for the cyclin-dependent kinases,and an ATP-binding domain (19, 63, 83). The binding of Cdc6pto ORC in G1 requires de novo synthesis of the protein (13, 62,63, 65). Similar findings were reported for Cdc18p, the S.pombe homolog of Cdc6p (40, 54, 57). S. cerevisiae Cdc6p issynthesized during the cell cycle in two peaks: in late mitosis,after anaphase where its expression is dependent on Swi5, andin late G1. This second peak is regulated by MBF-SBF and issufficient to promote DNA replication.
Interestingly, the expression of cdc181 is regulated by thetranscription factor complex DSC1, the MBF homolog inS. pombe (41). Although the E2F transcription factors arenot structural homologs of the transcription factors associatedwith the yeast MBF complex, it is believed that E2F is thefunctional homolog of MBF and DSC1 in mammalian cells.Thus, since the cell cycle-regulated expression of the yeastCDC6 and cdc181 genes is dependent on MBF and DSC1,respectively, we reasoned that the expression of the mamma-lian homolog of CDC6 would be controlled by E2Fs. There-fore, we isolated cDNAs encoding the human and mouse ho-mologs of Cdc6p or Cdc18p and the 59 regulatory region ofhuman CDC6. During the course of this work, a human cDNAfor CDC6 and a partial 59 regulatory region was published bythe laboratory of Bruce Stillman (79). Our data suggest thathCDC6 (human CDC6) protein is limiting and essential forentry into the S phase of the mammalian cell cycle and that thecell cycle-regulated expression of mammalian CDC6 is E2Fdependent.
MATERIALS AND METHODS
Isolation of human CDC6, mouse CDC6, and the 5* regulatory region ofhCDC6. A search of the human expressed sequence tag (EST) database using theS. cerevisiae Cdc6p and the S. pombe Cdc18p amino acid sequences revealedseveral EST sequences whose translation products showed homology to Cdc6pand Cdc18p. Primers were generated to one of these clones (EST identificationno. 376630), originally isolated from a Soares fetal heart NbHH19W Homosapiens cDNA library, and PCR was used to generate a 414-bp fragment of theEST clone using a human cDNA library prepared from fetal brain (61). The PCRfragment was radiolabeled, and a human embryonic fibroblast library M426(kindly provided by P. P. Di Fiore) was screened. One clone containing an insertof 3.0 kb was sequenced on both strands and was shown to contain an openreading frame of 560 amino acids with extended homology to Cdc6p and Cdc18p.This clone was named hCDC18 but later renamed hCDC6 in agreement withWilliams et al. (79) who identified a cDNA containing an identical open readingframe.
A cDNA encoding mouse CDC6 was identified in the EST database (ImageConsortium clone identification no. 477516 [46]) by searching the database withthe amino acid sequence of hCDC6. This clone was obtained from ResearchGenetic (Huntville, Ala.) and was found to contain an insert of 2,608 bp. ThecDNA was sequenced completely on both DNA strands and contained an openreading frame of 589 amino acids. Mouse CDC6 is 80% identical to humanCDC6 and contains a 27-amino-acid amino-terminal extension.
The 355-bp fragment of the human promoter (79) was generated by PCR,using human genomic DNA as a template. This PCR product was used to screena human genomic library prepared in Lambda FIX II (Stratagene). From 5 3 105
PFU, three positive phages were purified that contained three different sizedinserts: 12.8, 12.9, and 16.8 kb. The results of Southern blot analysis (data notshown) identified three different SacI fragments of 1.8, 5.5, and 7.0 kb that allcontained the 355-bp region. The 1.8-kb fragment was subcloned into pBSK2(Stratagene), and both strands were completely sequenced by using the VistraDNA Sequencer 725 (Amersham).
Plasmids. For p[2130,1225], the 355-bp PCR product described above wascloned into the NheI and HindIII sites of pGL3-Basic (Promega), a vector thatcontains a gene encoding the luciferase protein downstream of a multiple cloningsite and which lacks any promoter or enhancer elements. Site-directed mutagen-esis on p[2130,1225] was performed with the Chameleon double-stranded,site-directed mutagenesis kit (Stratagene) in accordance with the instructions ofthe manufacturer to produce pGL3-DM (double mutant, two E2F sites mutated,bp 243 to 236 and bp 28 to 21), pGL3-SM1 (single mutant 1, first E2F sitemutated, bp 243 to 236), and pGL3-SM2 (single mutant 2, second E2F sitemutated, bp 28 to 21). Except for the introduced mutations, the inserts of thesethree plasmids are identical to the one in p[2130,1225], as was confirmed byDNA sequencing. The primers used for the site-directed mutagenesis were59-GAGGCCGGGCTTTGAAGGGAGGTGGGAACG-39 and 59-CCATTCGGATTTGAAGCGAGCGC GGCTGG-39 for the E2F sites and 59-CGTGTAATTCTAGCGTCGGGGCGGCCG-39 for the unique XbaI site in pGL3-Basic.Mutated nucleotides are underlined.
p[21534,1225] was generated by cloning the 1.8-kb SacI fragment from theLambda Fix phage DNA into the SacI site of pGL3-Basic. p[2570,198] wasgenerated by subcloning the internal BamHI/BamHI fragment from p[21534,1225] into the BglII site of pGL3-Basic. p[2266,198] was generated by sub-cloning the internal NaeI/BamHI fragment into the SmaI and BglII sites ofpGL3-Basic.
pCMVHAhCDC6 was constructed by PCR amplification of the full-lengthopen reading frame of hCDC6, and the PCR product was subsequently clonedinto the BamHI site of pCMVHA (49). The entire insert was sequenced, and nomutations were introduced by PCR. pCMV-bgal was obtained from Clontech.pCMV-CD20, pCMV-E2F-1, pCMV-DP-1, and pRcCMVcyclin E have previ-ously been described (28, 30, 32, 81). pCMV-ER-E2F-1 and pCMV-ER-E2F-1-E132, which were used to generate the Rat-1 cells, stably expressing the wild-typeand a DNA-binding mutant of E2F-1 linked to the ligand-binding domain of theestrogen receptor (ER) protein, respectively, will be described elsewhere (76).
Cell culture and transfections. U2-OS, T98G, MCF7, and Rat-1 cells weremaintained in Dulbecco’s modified Eagle’s medium (DMEM) supplementedwith 10% South American (Gibco) bovine calf serum (BCS). Mouse Swiss 3T3and NIH 3T3 cells were maintained in DMEM supplemented with 10% Colo-radan BCS (Gibco).
Transfections were performed by the calcium phosphate method (24). NIH3T3 cells were transiently transfected with 1.0 mg of one of the indicated humanCDC6 promoter-luciferase constructs and 0.5 mg of pCMV-bgal to control fortransfection efficiency. Sheared salmon sperm DNA was added to 17 mg per10-cm-diameter dish as carrier. U2-OS cells were transiently transfected with thesame amounts of promoter and bgal plasmids. pCMV-E2F-1 or pCMV-DP-1 (10ng) or both were cotransfected for transactivation experiments to ascertainwhether the different human CDC6 promoter constructs responded to E2F-1 orDP-1.
For synchronization of MCF7 cells for in vivo footprinting, fluorescence-activated cell sorting (FACS), and Northern blotting experiments, cells werestarved for 36 h in DMEM lacking isoleucine and BCS. Subsequently, cells wereharvested directly (0 h) or stimulated for 12, 16, and 20 h with normal medium
6680 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

supplemented with 10% BCS. For a G1/S block, cells were kept in DMEM–10%BCS supplemented with 1 mM hydroxyurea for 20 h and subsequently harvested.
For experiments concerning the direct regulation of hCDC6 mRNA expres-sion by E2F, the Rat-1 cells, expressing an ER-E2F wild-type fusion and anER-E2F-E132 DNA-binding mutant fusion, were starved for 48 h in mediumcontaining 0.1% BCS and subsequently stimulated in the following medium: (i)fresh medium containing 10% BCS, (ii) the same starvation medium to which 1mM 4-hydroxytamoxifen (OHT) was added, (iii) the same starvation medium towhich 10 mg of cycloheximide (CHX) per ml was added, or (iv) the samestarvation medium with OHT and CHX. Cells were stimulated for the times(hours) indicated in the figures and subsequently used either for reverse tran-scriptase PCR (RT-PCR) or FACS.
For the induction of S phase by cyclin E and hCDC6, U2-OS cells weretransiently transfected with 10 mg of pCMV empty vector, 10 mg of pRcCMV-cyclinE, 10 mg of pCMVHAhCDC6, or 10 mg of both plasmids. In all cases, 1 mgof pCMV-CD20 was cotransfected and amounts were adjusted to 21 mg per10-cm-diameter dish with sheared salmon sperm DNA.
Northern blotting. Total cellular RNA was purified from human and mousecells described in the figure legends by using the Nonidet P-40 lysis method (64).RNA (20 to 30 mg) was electrophoresed per lane through a 1% formaldehydeagarose gel, transferred to nitrocellulose, and probed with 32P-labeled partialhuman or mouse CDC6 or mouse E2F-1 cDNA. Equal loading was ensured byethidium bromide staining and/or by probing the same blot with a 32P-labeled ratglyceraldehyde-3-phosphate dehydrogenase (GAPDH) probe.
RT-PCR. RNA from the Rat-1 cells stably expressing the wild-type ER-E2F-1and the mutant ER-E2F-1-E132 was prepared by the guanidium thiocynate-acidphenol method (64). RNA preparations were subsequently treated with DNase,incubated for 20 min at 37°C, and phenol extracted. RNA (1 mg) was used forcDNA preparation with RT (Superscript II; Gibco) according to the instructionsprovided by the manufacturer. Obtained cDNA was subsequently used in a PCRusing Taq polymerase (Gibco) and 32P-labeled dCTP. To be in a linear range formeasuring the level of rat CDC6 GAPDH mRNA, the following PCR conditionswere used. First 10 cycles were used; these cycles had 0.5°C decreases in anneal-ing temperature from 54 to 49°C. After this, another 13 cycles were performedat 49°C.
Gel retardation assay. Electrophoresis mobility shift assays (EMSA) wereperformed as described previously (29). To obtain a region of the human CDC6promoter that harbored both E2F sites, the following oligonucleotides were usedin a PCR: 59-AATCGAGGCCGGGCTTTG-39 and 59-GCGGCAGCAGCAAACTCCAG-39, giving rise to a fragment of 85 bp (bp 257 to 128). PCR wasperformed on p[2130,1225] and pGL3-DM as templates, and the products wereend labeled with [g-32P]ATP and analyzed for protein-binding activities. Excess(50-fold) nonlabeled wild type and mutant probes (DM fragment) were used todetermine the specificity of E2F binding. MRC5 nuclear extracts were preparedas previously described (1). The following antisera were used for shifting E2F-DP-containing complexes: Monoclonal antibodies TFD10 (anti-DP1) (80),TFE41 (anti-E2F4) (53), XZ77 and 21C9 (both anti-retinoblastoma gene prod-uct pRB) (34), SD9 (anti-p107) (20), and as a negative control, M1 (anti-E1A)(26). Polyclonal antisera were anti-E2F4 (sc-866), anti-p107 (sc-318), anti-p130(sc-317), and as a negative control, anti-farnesyl transferase (sc-137). All poly-clonal sera were obtained from Santa Cruz Biotechnology.
Luciferase and b-Gal assays. U2-OS and NIH 3T3 cells were transientlytransfected in duplicate and triplicate, respectively. One dish of the NIH 3T3transfectants or cells that were starved and then with serum stimulated was usedfor FACS analysis (see below). Forty-eight hours posttransfection, cells wereharvested by scraping in phosphate-buffered saline (PBS) and pelleted by cen-trifugation. Cell pellets were subsequently resuspended in 100 ml of 100 mMTris-HCl, pH 7.8, and cells were lysed by three freeze-thaw steps. Cell debris wasspun down, and supernatant was transferred to a new tube and used for bothluciferase and b-galactosidase (b-Gal) assays. Luciferase activity was typicallymeasured by mixing 20 ml of lysate with 30 ml of 100 mM Tris-HCl [pH 7.8] and50 ml of luciferase mix. The luciferase mix is 26 ml of luciferase buffer, 24 ml ofphosphate buffer, 0.15 ml of 1 M dithiothreitol, and 1.5 ml of 0.2 M ATP.Luciferase buffer is 25 mM glycyl glycine [pH 7.8], 15 mM MgSO4, and 4 mMEGTA, and phosphate buffer is 100 mM KH2PO4/K2HPO4, pH 7.8. Subse-quently, 50 ml of a 10 mM luciferin solution (in luciferase buffer) was added andactivity was determined in an Anthos Lucy 1 luminometer (Labtech).
b-Gal activity was measured by mixing 2 to 20 ml of lysate with Z buffer (60mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4) to a total volumeof 200 ml. To start the reaction, 40 ml of ONPG substrate (4 mg of o-nitrophenyl-b-D-galactopyranoside [Sigma] per ml of 100 mM phosphate buffer [pH 7.0]) wasadded and incubated at 30°C for 30 to 120 min. The reaction was stopped byadding 100 ml of 1 M Na2CO3, and the absorbance was read at 420 nm in a MRXplate reader (Dynatech Laboratories). All experiments involving luciferase andb-Gal activity were carried out at least three times.
FACS analysis. Cells from one 10-cm-diameter dish were washed once withPBS, trypsinized, spun, and washed once more with PBS, while samples from theelutriation fractions were spun and washed once in PBS. Cells were subsequentlyfixed in 70% methanol and either stored at 4°C or directly used for propidiumiodine (PI) staining. For PI staining, cells were washed once in PBS supple-mented with 1% BCS, spun, and subsequently resuspended in 500 ml of PI buffer(10 mM Tris-HCl [pH 7.0], 5 mM MgCl2, 50 mg of PI [Sigma] per ml, 100 mg of
RNase A per ml). After 30 min of incubation at 37°C, the samples were analyzedwith a Becton Dickinson FACScan. CD20-positive cells were selected andtreated as described previously (53).
In vivo DMS genomic footprinting. In vivo dimethyl sulfate (DMS) genomicfootprinting with the ligation-mediated PCR amplification (LMPCR) procedurewas performed essentially as previously described (25, 52) with the followingmodifications: 2 3 106 MCF7 cells per 14-cm-diameter dish that were eitherincubated with hydroxyurea or that were exponentially growing or serum starvedand then stimulated, were treated with the guanosine methylating agent DMS at0.2% for 5 min at room temperature in their respective culture medium (DMEMwith 0 or 10% BCS) buffered with HEPES (pH 7.4) (20 mM final concentration).After DMS treatment, cells were washed three times with cold PBS containing2% b-mercaptoethanol and then collected in 1 ml of lysis buffer (50 mM Tris [pH8.0], 20 mM EDTA, 1% sodium dodecyl sulfate, 2% b-mercaptoethanol).Genomic DNA was isolated by three gentle extractions with phenol (pH 8.0)followed by two precipitations in 4 M ammonium acetate with 3 volumes ofethanol. DNA was then redissolved in 1 ml of water. As a reference, MCF7naked genomic DNA (1 mg/ml in water) was methylated in vitro with 0.5% DMSfor 4 min at room temperature. Piperidine cleavage at methylated bases wasperformed in 1 N piperidine at 95°C for 30 min. Chemically cleaved samples wereprecipitated in ethanol, evaporated twice, and finally resuspended to 0.4 mg/mlin water. Portions (2 mg) of these samples were used for LMPCR.
The following primers were designed to analyze the upper strand (plus strand)and lower strand (minus strand) of the transcription start site region of thehCDC6 promoter: plus strands, S1 (59-CTCTTCCACTGGATTGGTAGC-39),S2 (59-AAAGAGGCGGTGCCCAAGGCG-39), and S3 (59-AGAGGCGGTGCCCAAGGCGAAAGCTC-39); for minus strands, AS1 (59-AAGGTGGAGGAGTCACGTCC-39), AS2 (59-TCTGCGTCGGAGAGCCTGAGTGG-39), and AS3(59-CGTCGGAGAGCCTGAGTGGTGGTGTTCGGGG-39). LMPCRs wereperformed as follows: 2 mg of cleaved genomic DNA and 0.3 pmol of S1 or AS1were denatured at 95°C for 5 min and then annealed for 30 min at 50°C in 25 mlof first-strand buffer (10 mM Tris-HCl [pH 8.9], 5 mM MgSO4, 40 mM NaCl, 20mg of bovine serum albumin [BSA] per ml, 10% dimethyl sulfoxide [DMSO]).Vent exonuclease-negative DNA polymerase (0.5 U) (New England Biolabs) anddeoxynucleoside triphosphates (dNTPs) (final concentration of each dNTP, 0.25mM) (pH 7.0) were added to the annealed DNA mixture; elongation was per-formed for 15 min at 76°C. Annealed linkers (25 pmol) LINK1 and LINK2(plus-strand LINK1, 59-GAATTCAGATC-39; minus-strand LINK2, 59-GCGGTGACCCGAGAGATCTGAATTC-39) in 45 ml of ligation mix (50 mM Tris-HCl[pH 7.5], 14 mM MgCl2, 33 mM dithiothreitol, 1.6 mM ATP [pH 7.0], 100 mg ofBSA per ml, 4 U of T4 DNA ligase [Boehringer]) were then added to theelongation reaction; ligation was performed for 1.5 h at room temperature.Ligated DNA was then precipitated in ethanol, washed in 70% ethanol, andresuspended in 100 ml of amplification buffer (65 mM Tris-HCl [pH 8.8], 40 mMNaCl, 10 mM b-mercaptoethanol, 3 mM MgCl2, 50 mg of BSA per ml, 10%DMSO, 0.4 mM dNTPs). PCR amplification was then started after addition of 10pmol of S2 or AS2 primer and 1 U of Taq DNA polymerase (Perkin Elmer). PCRwas performed as follows: 1 cycle of 4 min at 95°C followed by 30 cyclesconsisting of 40 s at 95°C, 2 min at 62°C, and 3 min at 76°C, followed by 7 minat 76°C; this procedure was then repeated. From this amplification reaction, 15ml were collected and mixed with 5 ml of labeling buffer (65 mM Tris-HCl [pH8.8], 40 mM NaCl, 10 mM b-mercaptoethanol, 2 mM MgCl2, 3.75 mM dNTPs,10% DMSO, 0.5 U of Vent exonuclease-negative DNA polymerase) containing0.15 pmol of radioactively labeled S3 or AS3 primer (primers were 59 end labeledwith T4 polynucleotide kinase (New England Biolabs) and [m-32P]ATP. Thespecific activity of the labeled primers was 3 3 106 cpm/pmol. PCR was per-formed as follows: 2 min at 95°C, followed by nine cycles consisting of 40 s at95°C, 3 min at 66°C, and 5 min at 76°C. Eighty microliters of 0.3 M sodiumacetate (pH 5.5) and 10 mg of tRNA were then added to stop the reaction.Labeled DNA was phenol extracted and ethanol precipitated. The pellet waswashed with 70% ethanol, resuspended in 8 ml of sample loading buffer (95%formamide, 10 mM EDTA, 20 mM NaOH, 0.025% bromophenol blue, 0.025%xylene cyanol), and denatured for 3 min at 95°C. Two microliters of the samplewas loaded onto a 5% sequencing gel and run at 50 W. Dried gels were analyzedwith a PhosphorImager (Molecular Dynamics Inc.).
Elutriation. A culture of 108 asynchronously growing NIH 3T3 cells washarvested after three washes in PBS by trypsinization, spun, and resuspended inPBS containing 0.3 mM EDTA, 1 mg of glucose per ml, and 1% BCS. Subse-quently, cells were separated according to cell cycle position by counter flowelutriation with Beckman equipment. Fourteen cell-containing fractions werecollected and set on ice. From these fractions, samples were taken separately forFACS analysis, while the remaining cells were used for the purification of totalRNA by the Nonidet P-40 lysis method. RNA from 10 fractions was used forRT-PCR analysis.
Microinjection and immunostaining. Human glioblastoma T98G cells, at-tached to coverslips, were starved for 48 h in DMEM–0.1% BCS and subse-quently stimulated with DMEM–10% BCS supplemented with 100 nM bromode-oxyuridine (BrdU) (Sigma). After 16 h, approximately 200 cells per coverslipwere injected with either affinity-purified rabbit immunoglobulin G (IgG) as anegative control or with affinity-purified L20 (an anti-hCDC6 rabbit polyclonalantiserum) (61) in concentrations of 0.8 mg/ml in PBS. After the cells wereinjected, coverslips were put back in the same medium. Two hours after micro-
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6681
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

injection, cells were washed twice in PBS, fixed for 10 min in 4% formaldehyde,and subsequently treated for 10 min with Triton X-100 and for 30 s in 50 mMNaOH. Incorporated BrdU was immunostained with a mouse anti-BrdU mono-clonal antibody (Becton Dickinson) and visualized with a goat anti-mouse CY3-conjugated secondary antibody. Injected cells were identified by incubation witha fluorescein isothiocyanate (FITC)-conjugated secondary goat anti-rabbit anti-body recognizing the injected antibodies. Finally, cells were treated with 49,6-diamidino-2-phenylindole (DAPI) (Sigma), rinsed in PBS and water, andmounted before analysis with a fluorescence microscope. Noninjected cells werestimulated with DMEM–10% BCS containing 100 nM BrdU, fixed after 0, 14, 16,and 18 h, and subsequently stained for BrdU incorporation as described above.
Nucleotide sequence accession numbers. The cDNA sequence of mouseCDC6 has been submitted to the DDBJ/EMBL/GenBank databases under ac-cession no. AJ009559. The sequence of the 1.8-kb human CDC6 promoter hasbeen submitted to the DDBJ/EMBL/GenBank databases under accession no.AJ009560.
RESULTS
Mammalian CDC6 is cell cycle regulated. In order to find ahuman homolog of S. cerevisiae Cdc6p and S. pombe Cdc18p,a search in the EST database was performed with the aminoacid sequences of the yeast proteins. One human cDNA wasidentified (EST identification no. 376630) that, when trans-lated, shows homology to both Cdc6p and Cdc18p. This ESTwas different from human ORC1 which also has homology toCdc6p and Cdc18p (4, 21). The cDNA coding for the homol-ogous region was used to screen a human embryonal fibroblastlibrary to obtain a potentially full-length cDNA. A clone con-taining a 3.0-kb insert was sequenced and shown to contain anopen reading frame of 560 amino acids. This clone was namedhCDC6 after a cDNA containing an identical open readingframe was described (79).
To investigate CDC6 mRNA expression, RNA was preparedfrom a number of human, rat, and mouse cell lines. Figure 1Ashows the CDC6 mRNA expression in a number of establishedhuman tumor cell lines, while Fig. 1B shows the expression ina number of fibroblast cell lines. Two CDC6 mRNA signals(approximately 3.3 and 2.5 kb) were detected in most cell linestested, which might be due to alternative splicing of themRNA. The slower-migrating mRNA appeared more abun-dant.
To investigate whether CDC6 expression is cell cycle regu-lated in immortalized murine fibroblasts, Swiss 3T3 cells wereserum starved and subsequently induced to reenter the cellcycle by addition of serum. Figure 2A shows the FACS analysisof these cells that were serum starved and then stimulated,indicating that Swiss 3T3 fibroblasts reach S phase after 16 h ofstimulation in a highly synchronous manner. A cDNA encod-ing mouse CDC6 (mCDC6) was identified in the EST database(EST identification no. 477516) by searching the database withthe amino acid sequence of hCDC6. This cDNA was se-quenced completely and shown to contain the complete openreading frame of mouse CDC6. The mouse protein is 80%identical to the human protein and contains a 27-amino-acidextension at the amino terminus (data not shown). A Northernblot containing total RNA from Swiss 3T3 fibroblasts, treatedas described above for the FACS analysis, was probed with aradiolabeled mouse CDC6 cDNA. Figure 2B shows that theexpression of CDC6 is cell cycle regulated and that the mRNAfor CDC6, like E2F-1, is upregulated in Swiss 3T3 cells inmid-to-late G1. Interestingly, the CDC6 mRNA is absent inG2/M and early G1, suggesting that the expression of CDC6 iscell cycle regulated in proliferating cells as well as in cellsentering the cell cycle from quiescence (discussed below). Asimilar expression pattern of CDC6 was observed in primaryhuman MRC5 fibroblasts and in NIH 3T3 cells, although thedownregulation of the CDC6 mRNA in G2 and early G1 wasnot as pronounced in these two cell lines (data not shown).
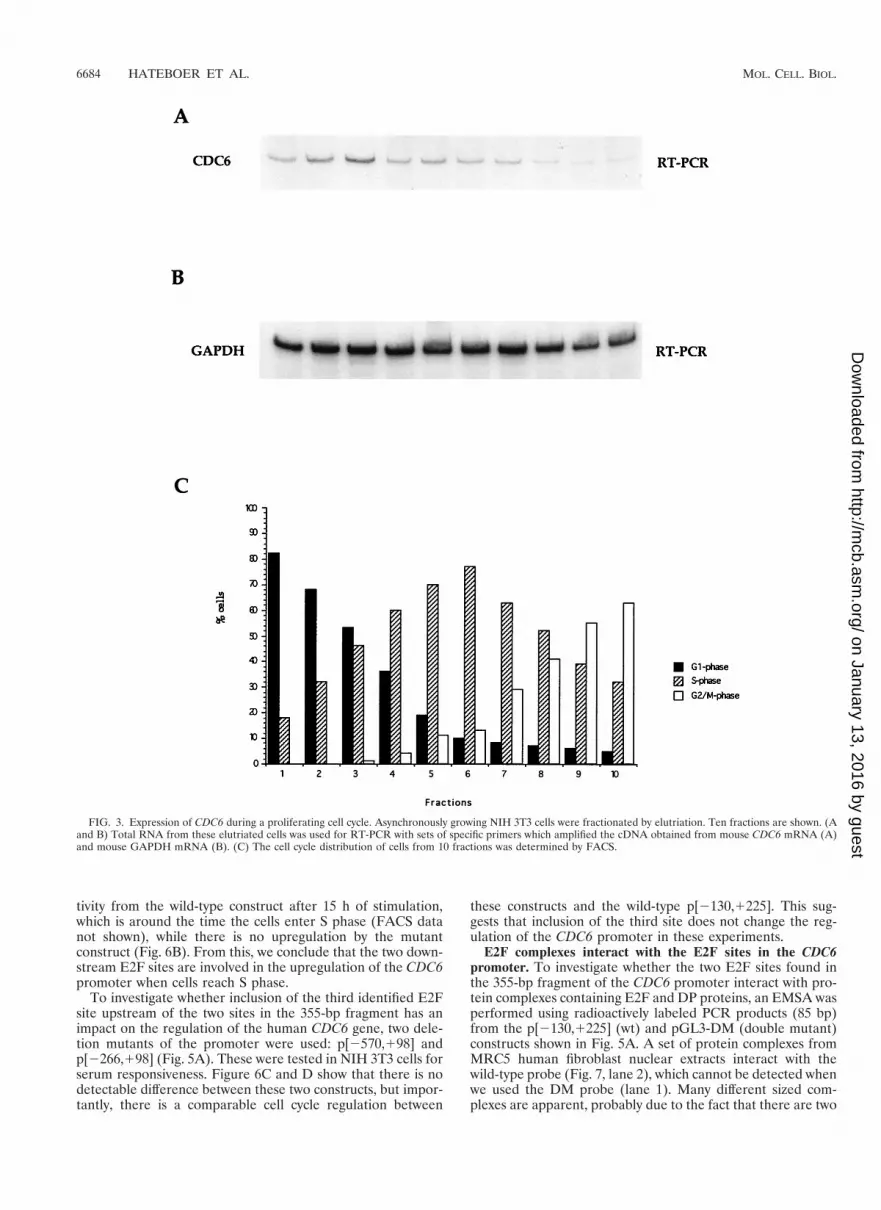
Cell cycle expression of CDC6 in proliferating cells. S. cer-evisiae CDC6 and S. pombe cdc181 are regulated differentlyduring a normal cell cycle. Data obtained from synchronouspopulations of elutriated cells have shown that both CDC6 andcdc181 transcription is periodic in proliferating cells; however,whereas CDC6 transcription is induced twice during the cellcycle (in M and late G1 [63]), the cdc181 expression is inducedonly once (in late G1 [40]). To investigate how mammalianCDC6 is regulated in proliferating cells, exponentially growingNIH 3T3 cells were elutriated, and fractions of cells were takenfor determining the cell cycle profile (Fig. 3C) and for RNApreparations. The expression of CDC6 was analyzed by RT-PCR on the obtained RNA, using primers specific for mouseCDC6 (Fig. 3A) or mouse GAPDH (Fig. 3B). The PCRs wereperformed under linear conditions in order to compare mRNAlevels. The data shown in Fig. 3A suggest that the expression ofCDC6 peaks in G1 and that the expression of CDC6 is down-regulated during S and G2/M, which is in agreement with ourfindings shown in Fig. 2B. CDC6 mRNA in these experimentsis nevertheless detectable in all analyzed fractions. This fact
FIG. 1. Expression of endogenous human, rat, and mouse CDC6 mRNA. (A)Northern blot containing total RNA from asynchronously growing human tumorcells U937 (histiocytic lymphoma), HT230 (colon adenocarcinoma), IMR32(neuroblastoma), ML1 (premyeloid leukemia), HL60 (promyeloid leukemia),NGP (neuroblastoma), SiHa (cervix carcinoma), and T98G (glioblastoma). Theblot was probed with a partial human CDC6 cDNA probe. (B) Northern blotcontaining total RNA from the human fibroblast cell lines Detroit 551 (embry-onic skin), IMR90 (fetal lung), Hs27 (newborn foreskin), and HEL 299 (embry-onal lung), the rat cell line Rat-1, and the mouse cell line NIH 3T3. Hybridiza-tion was done with a human CDC6 cDNA probe (lanes 1 to 5) and with a mouseCDC6 cDNA probe (lanes 6 and 7). Blots were subsequently probed with apartial rat GAPDH probe, as depicted (lower blots).
6682 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

can most likely be attributed to the lower degree of synchronyin the NIH 3T3 cells in G2/M in the elutriation procedurescompared to that in the Swiss 3T3 cells after serum starvationand stimulation. In conclusion, our data therefore suggest thatthe CDC6 expression is cell cycle regulated in proliferatingcells as well as in cells entering the cell cycle from a state ofquiescence.
E2F transactivates the human CDC6 promoter. In an at-tempt to find the regulatory elements controlling the expres-sion of human CDC6, the 355-bp promoter fragment of thereported gene (79) was used to screen a human genomic li-brary. Three phages containing sequences upstream from the355-bp promoter fragment, ranging in size from 1.8 to 7.0 kbwere purified. The 1.8-kb fragment was cloned and sequencedcompletely (Fig. 4A). The 1,759-bp 59 regulatory region con-tains three putative E2F DNA-binding sites upstream of thepreviously identified transcription start site, depicted in bold(bp 11). Computer analysis of the 1,759-bp sequence identi-fied several additional potential transcription factor-bindingsites (Fig. 4B). The two 39 most E2F sites match properly withthe consensus sequence TTT(G/C)(G/C)CG(G/C), while the59 most site does not.
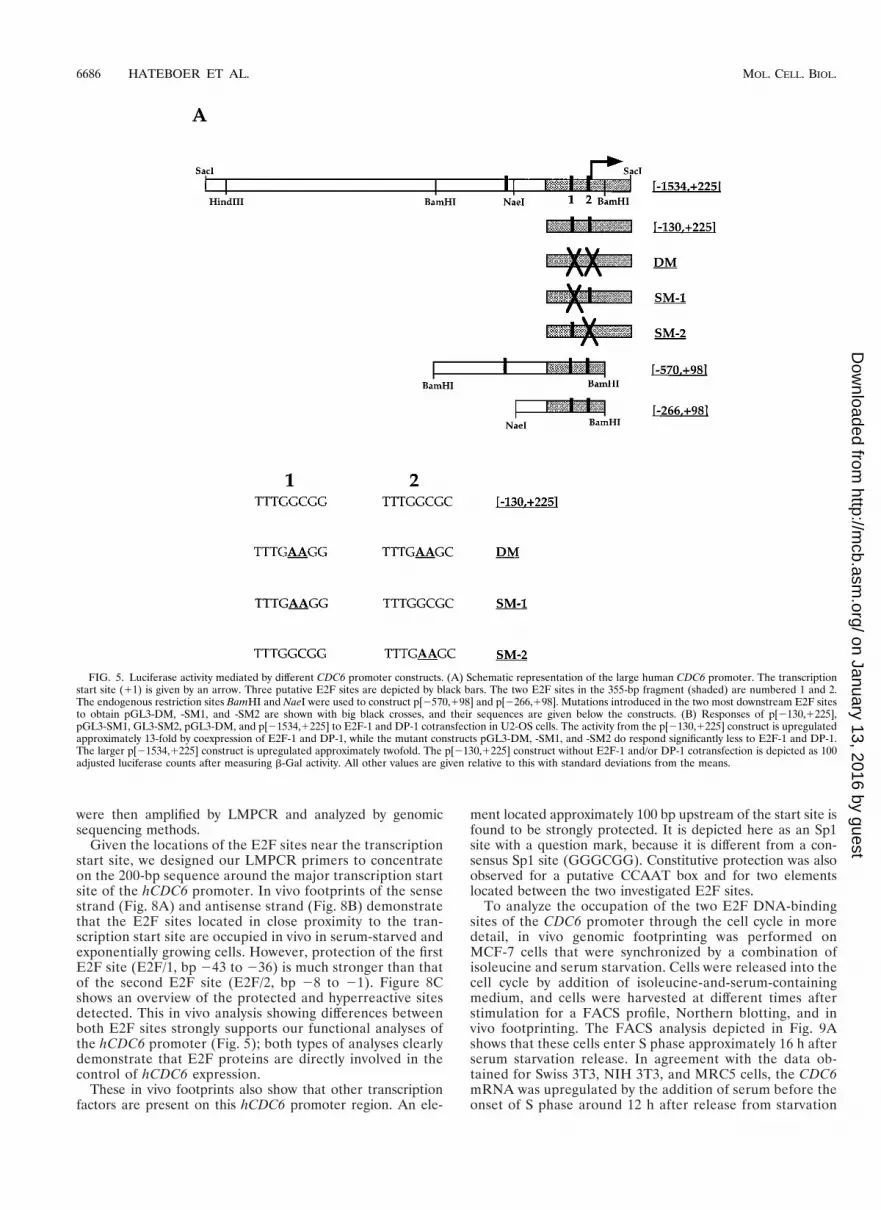
Several deletion and point mutants were generated in orderto study the roles of the E2F transcription factors in the reg-ulation of mammalian CDC6 expression. These mutants areshown in Fig. 5A. The wild-type fragments, named [21534,1225] (1,759 bp) and [2130,1225] (355 bp) were cloned up-stream of the luciferase gene. In addition, three mutants har-boring point mutations in the two E2F sites present in the
355-bp fragment were generated: SM1 carries a double-nucleo-tide mutation in the first E2F site, SM2 carries a double-nu-cleotide mutation in the second E2F site, while the double mu-tant (DM) harbors these mutations in both sites (sequences atthe bottom of Fig. 5A). Finally, two deletion mutants were gen-erated in order to measure the effect of the third (59 most) E2Fsite: (i) [2570,198], the internal BamHI/BamHI fragment(668 bp) from the 1,759-bp promoter, therefore carrying allthree putative sites, and (ii) [2266,198], the internal NaeI/BamHI fragment (364 bp), which harbors the two downstreamE2F sites but not the upstream site. All mutant fragments, likethe wild-type fragments, were cloned upstream of the lucifer-ase gene.
To investigate the roles of the E2F sites found in the humanCDC6 promoter, the activities of several of the luciferase con-structs shown in Fig. 5A were tested by transient transfectionin asynchronously growing human osteosarcoma U2-OS cells.To measure whether the two E2F sites in p[2130,1225] couldrespond to E2F and DP expression, plasmids encoding wild-type E2F-1 and DP-1 proteins were cotransfected. Figure 5Bshows the luciferase activity from the different promoters, cal-culated after adjusting for transfection efficiency. The largerp[21534,1225] construct alone is approximately sixfold moreactive than the shorter p[2130,1225] alone. It was also foundthat mutant versions of p[2130,1225] still harbor an activationcapacity in U2-OS cells, with the double mutant (DM) beingabout twofold more active than the p[2130,1225] construct.This suggests that the E2F sites in this promoter are involvedin repression of CDC6 expression, although we cannot excludethe possibility that the sites switch from positive to negativeand back during a normal cell cycle.
Luciferase activity is upregulated significantly (13-fold)when E2F-1 and DP-1 are cotransfected with p[2130,1225].Importantly, the effect of E2F and DP cotransfection is lostwhen the double mutant pGL3-DM is used, while the singlemutants pGL3-SM1 and -SM2 both still respond to E2F-1 andDP-1 coexpression, although this upregulation is significantlyless than found with the wild-type construct. The activity of thelarger promoter (p[21534,1225]) is upregulated about two- tothreefold after E2F-1 and DP-1 coexpression, while its basalactivity is much higher. A possible repressive effect by pocketproteins under more physiological conditions is lost when tran-scriptional activator proteins such as E2F and DP are tran-siently overexpressed, because endogenous pocket protein lev-els are too low to overcome the transient activity. Nevertheless,these data show that the E2F sites in the CDC6 promoter areE2F responsive. Moreover, we conclude that the first E2F sitein p[2130,1225] appears to play a more significant role in therepression of CDC6 transcription than the second site. Theseresults are in agreement with the findings from the in vivofootprints showing a strong protein binding to the upstreamelement in quiescent cells, while a weaker occupation could bedetected on the downstream site (discussed below).
Roles of the E2F sites in the G1-to-S transition. Because wefound that E2F can activate transcription from the CDC6 pro-moter, it was interesting to determine the roles of the E2F sitesin the CDC6 promoter during the cell cycle. For this, NIH3T3 cells were transiently transfected with the wild-type con-struct p[2130,1225] and the mutant construct pGL3-DM. Co-transfection was performed with a b-Gal-expressing plasmid.Next, cells were starved for 24 h in a low level of serum andsubsequently stimulated with a high level of serum. Lysateswere prepared at certain time points, and finally all lysateswere measured for luciferase activity, and compared to theactivity found in asynchronously growing cells. Figure 6Ashows that there is a significant upregulation of luciferase ac-
FIG. 2. Cell cycle-regulated expression of CDC6 mRNA. (A) Swiss 3T3 cellswere starved in low-serum-containing medium, subsequently stimulated withhigh serum, and used for FACS analysis. (B) Total RNA was isolated from Swiss3T3 cells treated similarly and used for Northern blotting. The blot was probedfirst for mouse CDC6 and E2F-1 mRNAs (upper blots). Subsequently the blotwas probed for GAPDH expression (lower blot).
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6683
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
tivity from the wild-type construct after 15 h of stimulation,which is around the time the cells enter S phase (FACS datanot shown), while there is no upregulation by the mutantconstruct (Fig. 6B). From this, we conclude that the two down-stream E2F sites are involved in the upregulation of the CDC6promoter when cells reach S phase.
To investigate whether inclusion of the third identified E2Fsite upstream of the two sites in the 355-bp fragment has animpact on the regulation of the human CDC6 gene, two dele-tion mutants of the promoter were used: p[2570,198] andp[2266,198] (Fig. 5A). These were tested in NIH 3T3 cells forserum responsiveness. Figure 6C and D show that there is nodetectable difference between these two constructs, but impor-tantly, there is a comparable cell cycle regulation between
these constructs and the wild-type p[2130,1225]. This sug-gests that inclusion of the third site does not change the reg-ulation of the CDC6 promoter in these experiments.
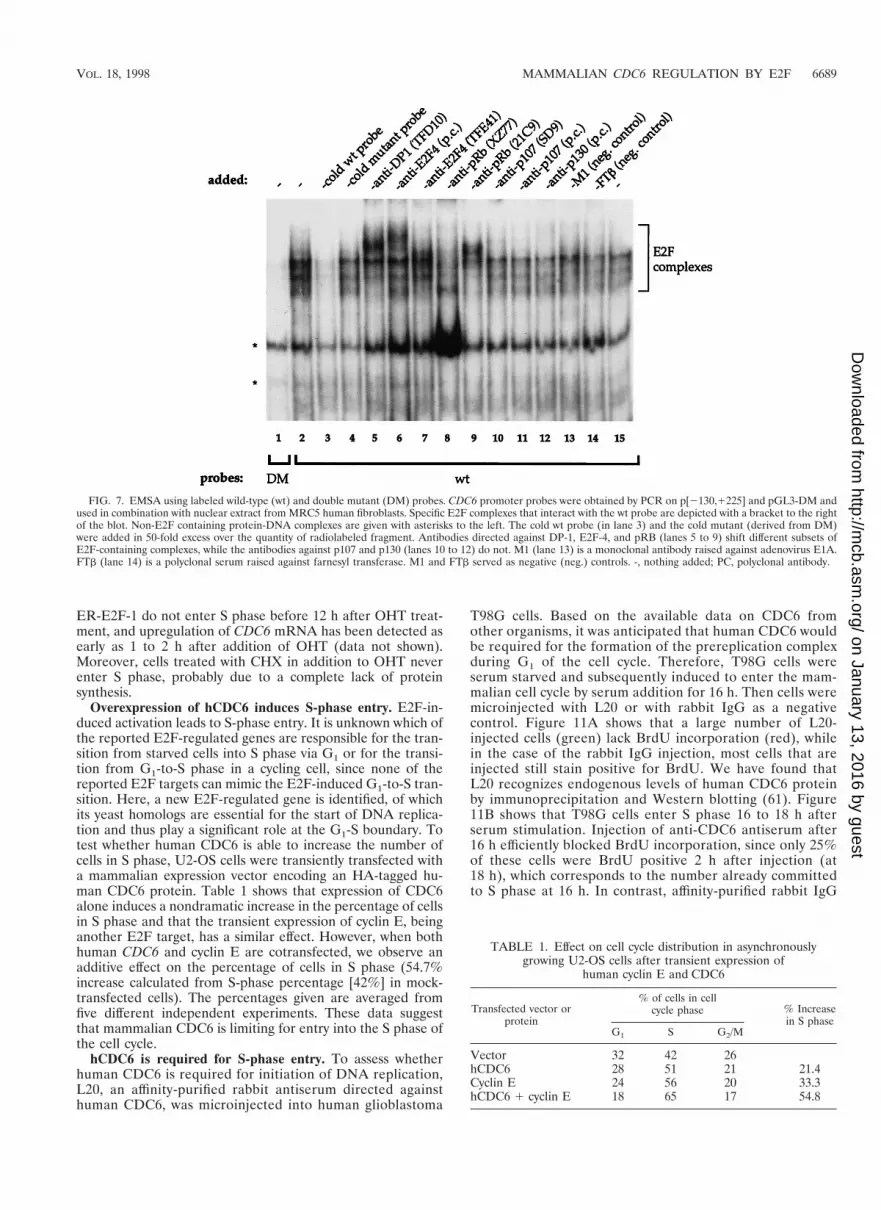
E2F complexes interact with the E2F sites in the CDC6promoter. To investigate whether the two E2F sites found inthe 355-bp fragment of the CDC6 promoter interact with pro-tein complexes containing E2F and DP proteins, an EMSA wasperformed using radioactively labeled PCR products (85 bp)from the p[2130,1225] (wt) and pGL3-DM (double mutant)constructs shown in Fig. 5A. A set of protein complexes fromMRC5 human fibroblast nuclear extracts interact with thewild-type probe (Fig. 7, lane 2), which cannot be detected whenwe used the DM probe (lane 1). Many different sized com-plexes are apparent, probably due to the fact that there are two
FIG. 3. Expression of CDC6 during a proliferating cell cycle. Asynchronously growing NIH 3T3 cells were fractionated by elutriation. Ten fractions are shown. (Aand B) Total RNA from these elutriated cells was used for RT-PCR with sets of specific primers which amplified the cDNA obtained from mouse CDC6 mRNA (A)and mouse GAPDH mRNA (B). (C) The cell cycle distribution of cells from 10 fractions was determined by FACS.
6684 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

E2F sites in the wild-type probe that are most likely bound bydifferent E2F-containing protein complexes. This makes it dif-ficult to distinguish complexes that contain only E2F-DP fromthose that contain E2F-DP bound to the different pocket pro-teins; also, the two sites might behave differently. We foundhowever that both E2F sites are independently able to bind toE2F-containing complexes (data not shown).
We detect complexes present on the wild-type and mutantprobes, suggesting the presence of protein-DNA complexesoutside (between) the E2F sites, which is in agreement withour data using in vivo footprinting (see below). Specificallybound complexes were competed by an excess of cold wild-typeE2F site probe (Fig. 7, lane 3), but not by a mutant probe (lane4). These data show that the slower-migrating bands indeedcontain protein complexes that specifically interact with theE2F sites. To see which of these complexes contain E2Fs andknown associated proteins was investigated by the addition ofantisera against DP-1 (lane 5), E2F-4 (lanes 6 and 7), and pRB(lanes 8 and 9) which all shift or disrupt one or more E2Fsite-binding complexes. Antisera against p107 and p130 did notshift any of the E2F-site bound complexes (lanes 10 to 12).
Using these antibodies alone with the radioactive wild-typeE2F and mutant probes did not result in any detectable back-ground shifts due to a specific binding of antisera to the DNA(data not shown). From these experiments, we conclude thatthe E2F sites in the 355-bp fragment of the CDC6 promotercan interact with E2F-containing complexes in vitro and thatthese proteins cannot bind when the E2F sites are mutated.
E2F site occupation in vivo. We performed a detailed anal-ysis of the DNA-protein contacts on the hCDC6 promoterduring the cell cycle by an in vivo genomic footprinting strat-egy. This technique allows the observation of DNA-proteincontacts at single-nucleotide resolution in living cells. G0-ar-rested and exponentially growing MCF7 cells were incubatedwith DMS, a base methylating agent that reacts predominantlywith the N7 position of guanines that are not protected bytranscription factors. Methylated genomic DNA was extractedfrom DMS-treated cells. As a control, naked genomic DNAwas also methylated in vitro with DMS. In vivo- or in vitro-methylated DNA was then cleaved at modified bases with hotpiperidine. Using this DNA as matrix, hCDC6-specific ladders
FIG. 4. Sequence and schematic representation of the human CDC6 pro-moter. (A) Nucleotide sequence of the 1,759-bp CDC6 promoter. The previouslyreported 355-bp fragment is underlined (79). Three putative E2F DNA-bindingsites are boxed. A putative CHR region (bp 230 to 226) and the putative INRregion (bp 216 to 211) are shown in lowercase. Restriction sites used for cloningare italic, and the transcription start site (bp 11) is bold. (B) Schematic repre-sentation of transcription factor-binding sites in the large 1,759-bp human CDC6promoter. The transcription start site is depicted with an arrow. A CHR site, aCCAAT box, and a putative Sp1 element (Sp1/?) which are found to be pro-tected in in vivo footprint assays, are located next to the two 39 E2F sites closeto the start site. Positions of consensus binding sites for AP-2, C/EBP, Ets-1, andNF-kB are also given. Putative recognition sites upstream of bp 2325 anddownstream of the start site are not shown. INR, initiator region.
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6685
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
were then amplified by LMPCR and analyzed by genomicsequencing methods.
Given the locations of the E2F sites near the transcriptionstart site, we designed our LMPCR primers to concentrateon the 200-bp sequence around the major transcription startsite of the hCDC6 promoter. In vivo footprints of the sensestrand (Fig. 8A) and antisense strand (Fig. 8B) demonstratethat the E2F sites located in close proximity to the tran-scription start site are occupied in vivo in serum-starved andexponentially growing cells. However, protection of the firstE2F site (E2F/1, bp 243 to 236) is much stronger than thatof the second E2F site (E2F/2, bp 28 to 21). Figure 8Cshows an overview of the protected and hyperreactive sitesdetected. This in vivo analysis showing differences betweenboth E2F sites strongly supports our functional analyses ofthe hCDC6 promoter (Fig. 5); both types of analyses clearlydemonstrate that E2F proteins are directly involved in thecontrol of hCDC6 expression.
These in vivo footprints also show that other transcriptionfactors are present on this hCDC6 promoter region. An ele-
ment located approximately 100 bp upstream of the start site isfound to be strongly protected. It is depicted here as an Sp1site with a question mark, because it is different from a con-sensus Sp1 site (GGGCGG). Constitutive protection was alsoobserved for a putative CCAAT box and for two elementslocated between the two investigated E2F sites.
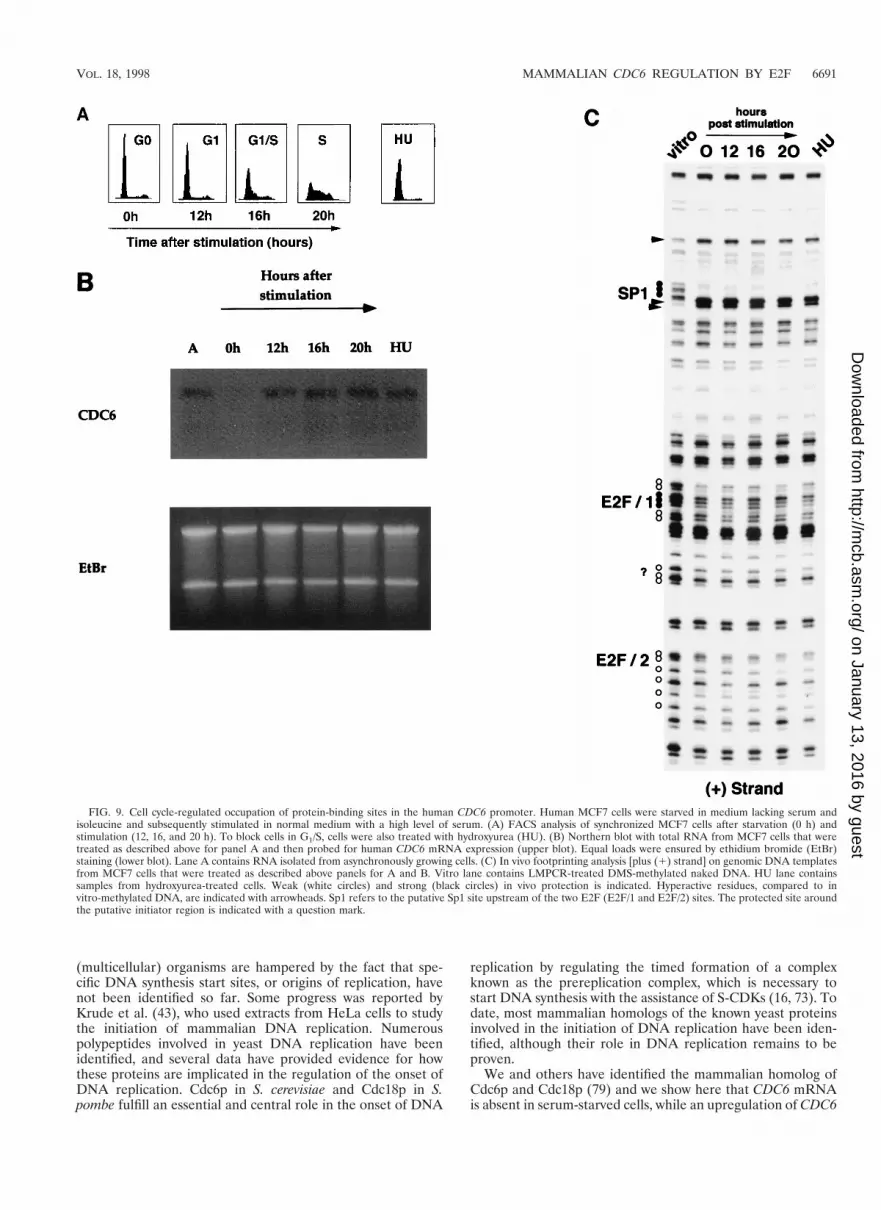
To analyze the occupation of the two E2F DNA-bindingsites of the CDC6 promoter through the cell cycle in moredetail, in vivo genomic footprinting was performed onMCF-7 cells that were synchronized by a combination ofisoleucine and serum starvation. Cells were released into thecell cycle by addition of isoleucine-and-serum-containingmedium, and cells were harvested at different times afterstimulation for a FACS profile, Northern blotting, and invivo footprinting. The FACS analysis depicted in Fig. 9Ashows that these cells enter S phase approximately 16 h afterserum starvation release. In agreement with the data ob-tained for Swiss 3T3, NIH 3T3, and MRC5 cells, the CDC6mRNA was upregulated by the addition of serum before theonset of S phase around 12 h after release from starvation
FIG. 5. Luciferase activity mediated by different CDC6 promoter constructs. (A) Schematic representation of the large human CDC6 promoter. The transcriptionstart site (11) is given by an arrow. Three putative E2F sites are depicted by black bars. The two E2F sites in the 355-bp fragment (shaded) are numbered 1 and 2.The endogenous restriction sites BamHI and NaeI were used to construct p[2570,198] and p[2266,198]. Mutations introduced in the two most downstream E2F sitesto obtain pGL3-DM, -SM1, and -SM2 are shown with big black crosses, and their sequences are given below the constructs. (B) Responses of p[2130,1225],pGL3-SM1, GL3-SM2, pGL3-DM, and p[21534,1225] to E2F-1 and DP-1 cotransfection in U2-OS cells. The activity from the p[2130,1225] construct is upregulatedapproximately 13-fold by coexpression of E2F-1 and DP-1, while the mutant constructs pGL3-DM, -SM1, and -SM2 do respond significantly less to E2F-1 and DP-1.The larger p[21534,1225] construct is upregulated approximately twofold. The p[2130,1225] construct without E2F-1 and/or DP-1 cotransfection is depicted as 100adjusted luciferase counts after measuring b-Gal activity. All other values are given relative to this with standard deviations from the means.
6686 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

(Fig. 9B). Interestingly, the in vivo footprint of the humanCDC6 promoter (Fig. 9C) shows that the two E2F DNA-binding sites are protected in quiescent cells, as well as inserum-stimulated cells after 12, 16, and 20 h of serum stim-ulation, while a similar protection is detected in hydroxyu-rea-treated samples. These findings strongly suggest thatE2Fs bound to both E2F sites present in the promoter haveboth a negative role (when bound to pocket proteins) and apositive role in the regulation of CDC6 expression.
CDC6 is directly upregulated by E2F-1 activation. It hasbeen demonstrated that the upregulation of E2F activity inserum-deprived cells can induce S phase. Although many genesare upregulated due to E2F activity, in most cases, it is unclearwhether this is a secondary effect due to the induction of Sphase or whether these genes are directly targeted by E2F. Toinvestigate this, a number of Rat-1 cell lines were generatedexpressing either a fusion protein between wild-type E2F-1 andthe modified ligand-binding domain of the ER (Rat-1-ER-E2F-1) or a fusion between a DNA-binding mutant of E2F-1with the same ligand-binding domain of the ER protein (Rat-1-ER-E2F-1-E132) as a negative control. Details concerningthe generation, selection, and characterization of both celllines will be described elsewhere (76). The addition of OHT(an estrogen antagonist) to cells expressing wild-type, but notmutant, E2F-1 as a fusion with ER is sufficient to induce S
phase in serum-starved cells that are kept in a low level ofserum (data not shown). S-phase entry occurs approximately12 h after addition of OHT, which is 4 to 6 h before S-phaseentry of serum-stimulated cells (76). Figure 10A shows thatwhen Rat-1-ER-E2F-1 cells are starved for 48 h in low-serum-containing medium and subsequently treated with OHT, asignificant upregulation of endogenous CDC6 mRNA expres-sion occurs after 4 h (lanes 2 to 4) and that the upregulation isalso detected when CHX (an inhibitor of protein synthesis) isadded (lanes 6 to 9). The addition of CHX alone leads to aslight increase of CDC6 mRNA levels, which is most likely dueto a minor stabilization of the RNA (lanes 10 to 13). The signaldrops after 8 h of OHT addition (lane 5), suggesting that theCDC6 promoter also contains negative regulatory elements orthat the stability of the ER-E2F fusion itself is regulatedthrough downstream factors.
To further investigate E2F-1-regulated expression of CDC6,an RT-PCR experiment was performed using the RNA fromthe wild-type and E132 DNA-binding mutant cells. Expressionlevels of CDC6 mRNA were determined for up to 16 h of OHTaddition and compared to levels after BCS addition. The RT-PCR in Fig. 10B confirms the Northern blot data shown in Fig.10A. There is a significant upregulation of CDC6 mRNA levelsafter 4 h of OHT-induced activation (Fig. 10B, lane 2), whichcorrelates with the upregulation of cyclin E, B-Myb, p107, and
FIG. 5—Continued.
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6687
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

other putative E2F targets which we have detected in otherexperiments (76). After this, the level is somewhat downregu-lated (lanes 3 and 4). Interestingly, another upregulation ofCDC6 after 16 h of OHT-induced activation was detected (lane5), while GAPDH levels remain stable (lower blot). At present,we have no explanation for this phenomenon, but it is mostlikely not a direct effect of E2F activation. Lanes 6 to 9 showthat the upregulation of CDC6 upon BCS addition is signifi-cantly slower.
Figure 10C shows a similar RT-PCR experiment on CDC6and GAPDH mRNA from ER-E2F-1 wild-type cells after ad-dition of CHX alone, CHX plus OHT, and OHT alone for 4 h.CDC6 is upregulated after addition of OHT and also whenCHX is present; this shows that de novo synthesis of protein isnot necessary for CDC6 transcription by ER-E2F-1. Similarresults were found when an ER-E2F-3 fusion cell line was
used, but not when an ER-E2F-2 fusion cell line was treatedwith OHT (76). These observations suggest that CDC6 expres-sion is under the control of a specific set of E2F transcriptionfactor complexes.
It was also tested whether the effect of OHT and CHXaddition on the mutant cell line Rat-1-ER-E2F-1-E132 couldinfluence CDC6 mRNA levels. Figure 10D shows that CDC6expression is not upregulated after OHT and/or CHX additionin these mutant cells, which indicates that DNA binding of E2Fis essential for the activation of the CDC6 promoter. Fromthese experiments, we conclude that the upregulation of CDC6expression in these Rat-1 cells is a rapid and direct effect ofE2F-1 activation, which does not require protein synthesis andwhich occurs earlier than upon serum addition. These experi-ments also demonstrate that the upregulation of CDC6 is in-dependent of entry into S phase, since Rat-1 cells expressing
FIG. 6. Cell cycle-regulated expression of the CDC6 promoter is dependent on E2F DNA-binding sites. NIH 3T3 cells were transiently transfected withp[2130,1225], pGL3-DM, p[2570,198], and p[2266,198]. Forty-eight hours posttransfection, cells were serum starved for 24 h and subsequently stimulated with freshmedium containing 10% BCS. Lysates were made after the depicted time spans. Asynchronous (A) samples were valued as 100 adjusted luciferase counts, while theothers were calculated in comparison to this. Transfection efficiencies were determined by pCMV-bgal cotransfection. Samples were obtained in duplicate, and thepresented data are representative for at least three independent experiments.
6688 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
ER-E2F-1 do not enter S phase before 12 h after OHT treat-ment, and upregulation of CDC6 mRNA has been detected asearly as 1 to 2 h after addition of OHT (data not shown).Moreover, cells treated with CHX in addition to OHT neverenter S phase, probably due to a complete lack of proteinsynthesis.
Overexpression of hCDC6 induces S-phase entry. E2F-in-duced activation leads to S-phase entry. It is unknown which ofthe reported E2F-regulated genes are responsible for the tran-sition from starved cells into S phase via G1 or for the transi-tion from G1-to-S phase in a cycling cell, since none of thereported E2F targets can mimic the E2F-induced G1-to-S tran-sition. Here, a new E2F-regulated gene is identified, of whichits yeast homologs are essential for the start of DNA replica-tion and thus play a significant role at the G1-S boundary. Totest whether human CDC6 is able to increase the number ofcells in S phase, U2-OS cells were transiently transfected witha mammalian expression vector encoding an HA-tagged hu-man CDC6 protein. Table 1 shows that expression of CDC6alone induces a nondramatic increase in the percentage of cellsin S phase and that the transient expression of cyclin E, beinganother E2F target, has a similar effect. However, when bothhuman CDC6 and cyclin E are cotransfected, we observe anadditive effect on the percentage of cells in S phase (54.7%increase calculated from S-phase percentage [42%] in mock-transfected cells). The percentages given are averaged fromfive different independent experiments. These data suggestthat mammalian CDC6 is limiting for entry into the S phase ofthe cell cycle.
hCDC6 is required for S-phase entry. To assess whetherhuman CDC6 is required for initiation of DNA replication,L20, an affinity-purified rabbit antiserum directed againsthuman CDC6, was microinjected into human glioblastoma
T98G cells. Based on the available data on CDC6 fromother organisms, it was anticipated that human CDC6 wouldbe required for the formation of the prereplication complexduring G1 of the cell cycle. Therefore, T98G cells wereserum starved and subsequently induced to enter the mam-malian cell cycle by serum addition for 16 h. Then cells weremicroinjected with L20 or with rabbit IgG as a negativecontrol. Figure 11A shows that a large number of L20-injected cells (green) lack BrdU incorporation (red), whilein the case of the rabbit IgG injection, most cells that areinjected still stain positive for BrdU. We have found thatL20 recognizes endogenous levels of human CDC6 proteinby immunoprecipitation and Western blotting (61). Figure11B shows that T98G cells enter S phase 16 to 18 h afterserum stimulation. Injection of anti-CDC6 antiserum after16 h efficiently blocked BrdU incorporation, since only 25%of these cells were BrdU positive 2 h after injection (at18 h), which corresponds to the number already committedto S phase at 16 h. In contrast, affinity-purified rabbit IgG
FIG. 7. EMSA using labeled wild-type (wt) and double mutant (DM) probes. CDC6 promoter probes were obtained by PCR on p[2130,1225] and pGL3-DM andused in combination with nuclear extract from MRC5 human fibroblasts. Specific E2F complexes that interact with the wt probe are depicted with a bracket to the rightof the blot. Non-E2F containing protein-DNA complexes are given with asterisks to the left. The cold wt probe (in lane 3) and the cold mutant (derived from DM)were added in 50-fold excess over the quantity of radiolabeled fragment. Antibodies directed against DP-1, E2F-4, and pRB (lanes 5 to 9) shift different subsets ofE2F-containing complexes, while the antibodies against p107 and p130 (lanes 10 to 12) do not. M1 (lane 13) is a monoclonal antibody raised against adenovirus E1A.FTb (lane 14) is a polyclonal serum raised against farnesyl transferase. M1 and FTb served as negative (neg.) controls. -, nothing added; PC, polyclonal antibody.
TABLE 1. Effect on cell cycle distribution in asynchronouslygrowing U2-OS cells after transient expression of
human cyclin E and CDC6
Transfected vector orprotein
% of cells in cellcycle phase % Increase
in S phaseG1 S G2/M
Vector 32 42 26hCDC6 28 51 21 21.4Cyclin E 24 56 20 33.3hCDC6 1 cyclin E 18 65 17 54.8
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6689
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

did not prevent initiation of DNA replication, since thesecells synthesized DNA as efficiently as noninjected cells upto 70% at 18 h. These data demonstrate that human CDC6,like the homologs of hCDC6 in lower eukaryotes, is essen-tial for initiating DNA replication.
DISCUSSION
Most of our knowledge concerning DNA replication is ob-tained from experiments using the lower eukaryotes S. cerevi-siae, S. pombe, and X. laevis, while investigations in higher
FIG. 8. In vivo footprinting analysis of the transcription start site region ofthe hCDC6 promoter. LMPCRs were performed with primer S1, S2, or [minus(2) strand] or AS1, AS2, or AS3 [plus (1) strand] on genomic DNA templatesobtained from serum-starved (G0) or exponentionally growing (expo.) MCF7cells treated in vivo with the guanosine methylating agent, DMS. SimilarLMPCRs were performed with DMS-methylated naked DNA (vitro lanes). Pro-tected residues and hyperreactive residues detected between in vitro- and invivo-methylated DNAs are indicated as circles and arrowheads, respectively.Weak (white circles) and strong (black circles) in vivo protection is indicated.The transcription start site is indicated with an arrowhead (11) to the left of theblots. Amplified DNA ladders that are visible correspond to guanines of thehCDC6 promoter. (A) Positive-sense strand. (B) Negative-sense strand. (C)Summary of DNA-protein contacts observed by in vivo footprinting on bothstrands of the hCDC6 promoter upstream of the transcription start site (blackarrow, 11). Putative consensus binding sites are indicated as open boxes. Aprotein-bound element with a sequence similar to that of an Sp1 consensus siteis depicted as Sp1/?. A protected site around the putative initiator region (INR)(bp 216 to 210) is indicated with a question mark.
6690 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from
(multicellular) organisms are hampered by the fact that spe-cific DNA synthesis start sites, or origins of replication, havenot been identified so far. Some progress was reported byKrude et al. (43), who used extracts from HeLa cells to studythe initiation of mammalian DNA replication. Numerouspolypeptides involved in yeast DNA replication have beenidentified, and several data have provided evidence for howthese proteins are implicated in the regulation of the onset ofDNA replication. Cdc6p in S. cerevisiae and Cdc18p in S.pombe fulfill an essential and central role in the onset of DNA
replication by regulating the timed formation of a complexknown as the prereplication complex, which is necessary tostart DNA synthesis with the assistance of S-CDKs (16, 73). Todate, most mammalian homologs of the known yeast proteinsinvolved in the initiation of DNA replication have been iden-tified, although their role in DNA replication remains to beproven.
We and others have identified the mammalian homolog ofCdc6p and Cdc18p (79) and we show here that CDC6 mRNAis absent in serum-starved cells, while an upregulation of CDC6
FIG. 9. Cell cycle-regulated occupation of protein-binding sites in the human CDC6 promoter. Human MCF7 cells were starved in medium lacking serum andisoleucine and subsequently stimulated in normal medium with a high level of serum. (A) FACS analysis of synchronized MCF7 cells after starvation (0 h) andstimulation (12, 16, and 20 h). To block cells in G1/S, cells were also treated with hydroxyurea (HU). (B) Northern blot with total RNA from MCF7 cells that weretreated as described above for panel A and then probed for human CDC6 mRNA expression (upper blot). Equal loads were ensured by ethidium bromide (EtBr)staining (lower blot). Lane A contains RNA isolated from asynchronously growing cells. (C) In vivo footprinting analysis [plus (1) strand] on genomic DNA templatesfrom MCF7 cells that were treated as described above panels for A and B. Vitro lane contains LMPCR-treated DMS-methylated naked DNA. HU lane containssamples from hydroxyurea-treated cells. Weak (white circles) and strong (black circles) in vivo protection is indicated. Hyperactive residues, compared to invitro-methylated DNA, are indicated with arrowheads. Sp1 refers to the putative Sp1 site upstream of the two E2F (E2F/1 and E2F/2) sites. The protected site aroundthe putative initiator region is indicated with a question mark.
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6691
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

levels is detected when cells enter S phase; these levels dropwhen cells progress through the cell cycle. Preliminary datashow that the human CDC6 protein is absent in quiescent cellsand in early G1 phase of the cell cycle, and it appears that theprotein is present throughout the remaining part of the cellcycle (61).
It has been reported that the regulation of the CDC6 andcdc181 genes in S. pombe and S. cerevisiae are under thecontrol of the transcription factor complexes MBF/SBF andDSC1, respectively (41, 62, 73), which are the functional ho-mologs of the mammalian E2F transcription factor family,although they are not structurally homologous. It is shown herethat the E2F sites found in the 59 regulatory region of humanCDC6 are essential for the induction of hCDC6 expressionduring the G1-to-S transition in cycling cells. When two E2Fsites found in close proximity to the transcription start site
were mutated, E2F protein-containing complexes were unableto interact in band shift experiments, while the promoter lostthe ability to respond to coexpression of E2F and DP proteins.Moreover, a cell cycle-regulated expression of luciferase activ-ity was lost using a mutant promoter-luciferase gene fusionconstruct transfected in mouse NIH 3T3 cells that were starvedand subsequently serum stimulated. In our experiments, weincluded a third putative E2F site (bp 2279 to 2271) next tothe two sites that were used in the mutational analysis. Ourresults demonstrate that this site is neither required nor suffi-cient for the cell cycle-regulated expression of CDC6, presum-ably because the sequence does not match well with the E2Fconsensus binding sequence TTTSSCGS in which S is either aG or C. Although we did not analyze whether this putative E2FDNA-binding site can bind E2F, our data suggest that this siteis not occupied by E2F.
FIG. 10. Direct regulation of CDC6 expression by E2F-1 transcription factor. (A) Northern blot containing total RNA purified from Rat-1 cells stably expressinga fusion between full-length wild-type E2F-1 and the ligand-binding domain of the ER (ER-E2F-1). E2F-1 activity was induced by the addition of OHT in the absence(lanes 3 to 5) or presence of CHX (lanes 7 to 9). The effect of CHX addition alone (lanes 11 to 13) was also monitored. The Northern blot was probed with a partialmouse CDC6 probe and subsequently hybridized with a partial rat GAPDH probe. Lane A contains the asynchronous cell sample. (B) RT-PCR on RNA obtained fromthe cells described above for panel A. Cells were stimulated for up to 16 h with OHT or BCS or for 4 h with OHT in the absence or presence of CHX (C). Inductionof CDC6 mRNA expression was measured by a linear range radioactive RT-PCR, and the rat endogenous GAPDH mRNA served as a control (lower blot). (D)RT-PCR on total RNA purified from Rat-1 cells stably expressing a fusion between a full-length DNA-binding mutant (E132) of E2F-1 and the ligand-binding domainof ER after the addition of CHX, OHT plus CHX, or OHT alone for 4 h. The CDC6 signal was measured by an RT-PCR, and GAPDH mRNA served as a control(lower blot).
6692 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

To investigate to what extent the two 39 E2F sites werebound by proteins during the mammalian cell cycle, we used anin vivo footprinting assay. Our results show that one (bp 243to 236) of the two E2F sites is predominantly occupied byproteins in proliferating cells and that the same site is occupiedby proteins in starved cells (Fig. 8), whereas the other E2F site(bp 28 to 21) is significantly less protected. Both sites, how-ever, match well with the E2F DNA-binding consensus se-quence (see above). Taken together with the fact that the E2Ftranscription factors are complexed with pocket proteins inG0/G1 and that pocket proteins are active repressors of tran-scription (10, 69, 77), our data suggest that the absence ofmammalian CDC6 mRNA in G0 is due to an active repressionby pRb, p107 and/or p130 bound to E2F-DP complexes viathese E2F sites. Since we also found that the same sites areoccupied throughout the cell cycle (Fig. 9), it seems as if thesesites are crucial not only for repressing CDC6 expression butalso for activating CDC6 expression in mid-to-late G1. Ourdata generated using promoter-luciferase constructs with wild-type and mutated E2F DNA-binding sites in synchronized cellsstrongly support this interpretation (Fig. 5). Similar to theCDC6 promoter, it has been reported that the E2F DNA-binding sites in the dihydrofolate reductase (78) and thymidinekinase (39) promoters are also occupied throughout the cell
cycle, suggesting that the E2F sites in these promoters are bothpositive and negative regulatory elements.
Apart from protection of the E2F sites, we also detectedprotection of a putative Sp1 site, of a CCAAT region locatedupstream of the E2F sites and two sequences between the two39 E2F sites. One of these elements has homology to a regionfound in promoters of human cyclin A, cdc25C and cdc2 andmouse B-myb genes, called the cell cycle genes’ homologyregion (CHR) see below). The other element (question mark,Fig. 8C and 9C) resembles an initiator region depicted as INRin Fig. 4B. Further mutational analyses will be required to testwhether this regulatory element is also critical for the cellcycle-regulated activation of the CDC6 promoter.
As noted above, the promoters of the genes encoding humancyclin A, CDC2, CDC25c, and mouse B-Myb, share a sequenceof homology known as CHR, located five nucleotides down-stream of a cell cycle-dependent element (CDE) (84). SomeCDE sequences are involved in E2F-dependent regulation (6,67). A CHR-homologous sequence present five nucleotidesdownstream of the first E2F site in the CDC6 promoter wasdetected (Fig. 4). It was found that nucleotide 228 in theputative CHR behaves as a hypersensitive site in in vivo foot-prints (Fig. 8 and 9). Therefore, we investigated whether asimilar situation exists for hCDC6 by mutating the putative
FIG. 10—Continued.
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6693
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

CHR region in the background of p[2130,1225], pGL3-SM1,pGL3-SM2, and pGL3-DM. We find that a reporter harboringonly mutations in the CHR and the downstream E2F site (bp28 to 21) behaves as pGL3-SM2 in E2F-DP cotransfectionand in band shifts. We also find a comparable cell cycle regu-lation with this mutant promoter in starved and proliferatingNIH 3T3 cells (data not shown). From this, we conclude thatthe putative CHR in the hCDC6 promoter does not cooperatein E2F binding and that it is not involved in E2F-dependentcell cycle regulation. However, we do not exclude the possibil-ities that proteins bind to the CHR and that this element has
a role in hCDC6 gene regulation which is independent fromE2F binding.
Next to the E2F and CHF sites we have found throughcomputer analysis, a number of other consensus sites for tran-scription factor binding (Sp1, AP-2, Ets-1, NF-kB, etc. [Fig.4B]). No E-box elements (enabling the binding of basic region–helix-loop-helix proteins) in close proximity to the transcrip-tion start site were detected. As was shown in Fig. 8 and 9, wehave evidence that the CCAAT box and the putative Sp1 siteare occupied in in vivo genomic footprints. To date, we have nofurther data indicating which of the sites present is involved in
FIG. 11. Block of DNA synthesis by microinjection of specific antibodies directed against human CDC6 protein. (A) Human T98G cells were starved for 48 h inlow-serum-containing medium and subsequently stimulated with medium with high serum plus BrdU. After 16 h, cells were injected with either an affinity-purifiedspecific anti-hCDC6 rabbit antiserum (L20) or with affinity-purified rabbit IgG. After 2 h, cells were fixed and treated with DAPI (blue; upper panels) and withantibodies for BrdU (red; middle panels) and with antirabbit antibodies to score for the injected antisera (green; lower panels). Arrowheads indicate injected cells thatare not incorporating BrdU in the case of L20 or that are synthesizing DNA in the case of rabbit IgG. (B) Noninjected cells were scored for BrdU incorporation after0, 14, 16, and 18 h following serum stimulation and compared to the total number of cells. Injected cells that were stained green because of their injected antisera werescored for BrdU incorporation and compared to the total number of injected cells. The diagram is the average of three independent experiments in which approximately200 cells per coverslip were injected. FCS, fetal calf serum; BCS, bovine calf serum; rIgG, rabbit IgG.
6694 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

hCDC6 expression. However, we have shown that the E2F sitesclose to the start site are required for upregulated expressionbefore cells reach S phase and that mutant E2F sites abolishthis effect (Fig. 5B and 6).
Although it has been extensively reported that E2F activa-tion leads to the induction of S phase, it is unclear which of theE2F regulated genes are responsible for the E2F-induced G1-to-S transition or for the entry into a new cell cycle fromquiescence. We are unable to monitor the effect of hCDC6overexpression in serum-starved cells, since the protein couldnot be detected by immunostaining after microinjection inquiescent cells (61). Therefore, we cannot ascertain whetherhCDC6 alone or in combination with cyclin E is sufficient toinduce reentry into the cell cycle. A likely explanation for theinability to detect hCDC6 in quiescent cells could be that it isan unstable protein in serum-starved cells. Nevertheless, wewere able to express hCDC6 in proliferating cells and haveshown that overexpression of hCDC6 can increase the numberof cells in S phase. We have provided evidence that the ex-pression of hCDC6, the mammalian homolog of the essentialyeast genes CDC6 and cdc181, is directly regulated by E2Fthrough the E2F sites in the promoter. Significantly, hCDC6can induce S phase in cooperation with cyclin E, another E2Ftarget. Taken together with the demonstration that hCDC6 isrequired for entry into the S phase (Fig. 11), our data providea direct link between regulation of cell cycle progression by theE2F transcription factors and the initiation of DNA replicationand suggest that the function of mammalian CDC6 is similar tothose of its yeast homologs.
ACKNOWLEDGMENTS
We thank Karin Holm, Alexandra Charlesworth, Stefania Lupo,Heiko Muller, and Emanuela Frittoli for technical assistance in se-quencing, FACS, elutriation, and microinjection. Pier Paolo di Fiore isthanked for providing reagents, and we thank the members of theLattanzio family who donated the microinjection setup. We thankPeter Cartwright and Simonetta Piatti for critical reading of the manu-script.
This work was supported in part by grants from the Human Fron-tiers Science Program and the Associazione Italiana per la Ricerca sul
Cancro (AIRC) and by fellowships from the European Community(G.H.), the Fondazione Italiana per la Ricerca sul Cancro (E.V.), theFondazione per la Formazione Oncologica (A.W.), and the DanishResearch Academy (B.O.P.).
REFERENCES
1. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A.Smith, and K. Struhl. 1988. Current protocols in molecular biology. GreenePublishing Associates & Wiley-Interscience, New York, N.Y.
2. Beijersbergen, R. L., and R. Bernards. 1996. Cell cycle regulation by theretinoblastoma family of growth inhibitory proteins. Biochim. Biophys. Acta1287:103–120.
3. Beijersbergen, R. L., R. M. Kerkhoven, L. Zhu, L. Carlee, P. M. Voorhoeve,and R. Bernards. 1994. E2F-4, a new member of the E2F gene family, hasoncogenic activity and associates with p107 in vivo. Genes Dev. 8:2680–2690.
4. Bell, S. P., J. Mitchell, J. Leber, R. Kobayashi, and B. Stillman. 1995. Themultidomain structure of Orc1p reveals similarity to regulators of DNAreplication and transcriptional silencing. Cell 83:563–568.
5. Bell, S. P., and B. Stillman. 1992. ATP-dependent recognition of eukaryoticorigins of DNA replication by a multiprotein complex. Nature 357:128–134.
6. Bennett, J. D., P. G. Farlie, and R. J. Watson. 1996. E2F binding is requiredbut not sufficient for repression of B-myb transcription in quiescent fibro-blasts. Oncogene 13:1073–1082.
7. Blake, M. C., and J. C. Azizkhan. 1989. Transcription factor E2F is requiredfor efficient expression of the hamster dihydrofolate reductase gene in vitroand in vivo. Mol. Cell. Biol. 9:4994–5002.
8. Botchan, M. 1996. Coordinating DNA replication DNA replication with celldivision: current status of the licensing concept. Proc. Natl. Acad. Sci. USA93:9997–10000.
9. Botz, J., K. Zerfass-Thome, D. Spitkovsky, H. Delius, B. Vogt, M. Eilers, A.Hatzigeorgiou, and P. Jansen-Durr. 1996. Cell cycle regulation of the murinecyclin E gene depends on an E2F binding site in the promoter. Mol. Cell.Biol. 16:3401–3409.
10. Bremner, R., B. L. Cohen, M. Sopta, P. A. Hamel, C. J. Ingles, B. L. Gallie,and R. A. Phillips. 1995. Direct transcriptional repression by pRB and itsreversal by specific cyclins. Mol. Cell. Biol. 15:3256–3265.
11. Cartwright, P., H. Muller, C. Wagener, K. Holm, and K. Helin. 1998. E2F-6:a novel member of the E2F family is an inhibitor of E2F-dependent tran-scription. Oncogene 17:611–624.
12. Cobrinik, D. 1996. Regulatory interactions among E2Fs and cell cycle con-trol proteins. Curr. Top. Microbiol. Immunol. 208:31–60.
13. Cocker, J. H., S. Piatti, C. Santocanale, K. Nasmyth, and J. F. Diffley. 1996.An essential role for the Cdc6 protein in forming prereplicative complexes ofbudding yeast. Nature 379:180–182.
14. Dalton, S. 1992. Cell cycle regulation of the human cdc2 gene. EMBO J.11:1797–1804.
15. DeGregori, J., T. Kowalik, and J. R. Nevins. 1995. Cellular targets foractivation by the E2F1 transcription factor include DNA synthesis- and
FIG. 11—Continued.
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6695
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

G1/S-regulatory genes. Mol. Cell. Biol. 15:4215–4224.16. Diffley, J. F. X. 1996. Once and only once upon a time: specifying and
regulating origins of DNA replication in eukaryotic cells. Genes Dev. 10:2819–2830.
17. Donovan, S., J. Harwood, L. S. Drury, and J. F. X. Diffley. 1997. Cdc6p-dependent loading of Mcm proteins onto pre-replicative chromatin in bud-ding yeast. Proc. Natl. Acad. Sci. USA 94:5611–5616.
18. Dou, Q.-P., P. J. Markell, and A. B. Pardee. 1992. Thymidine kinase tran-scription is regulated at G1/S phase by a complex that contains retinoblas-toma-like protein and a cdc2 kinase. Proc. Natl. Acad. Sci. USA 89:3256–3260.
19. Drury, L. S., G. Perkins, and J. F. X. Diffley. 1997. The Cdc4/34/53 pathwaytargets Cdc6p for proteolysis in budding yeast. EMBO J. 16:5966–5976.
20. Dyson, N., M. Dembski, A. Fattaey, C. Ngwu, M. Ewen, and K. Helin. 1993.Analysis of p107-associated proteins: p107 associates with a form of E2F thatdiffers from pRB-associated E2F-1. J. Virol. 67:7641–7647.
21. Gavin, K. A., M. Hidaka, and B. Stillman. 1995. Conserved initiator proteinsin eukaryotes. Science 270:1667–1671.
22. Geng, Y., E. N. Eaton, M. Picon, J. M. Roberts, A. S. Lundberg, A. Gifford,C. Sardet, and R. A. Weinberg. 1996. Regulation of cyclin E transcription byE2Fs and retinoblastoma protein. Oncogene 12:1173–1180.
23. Ginsberg, D., G. Vairo, T. Chittenden, Z.-X. Xiao, G. Cu, K. L. Wydner, J. A.DeCaprio, J. B. Lawrence, and D. M. Livingston. 1994. E2F-4, a new mem-ber of the E2F transcription factor family, interacts with p107. Genes Dev.8:2665–2679.
24. Graham, F. L., and A. J. Van der Eb. 1973. A new technique for the assay ofinfectivity of human adenovirus. Virology 52:456–467.
25. Grange, T., E. Bertrand, M. L. Espinas, M. Fromont-Racine, G. Rigaud, J.Roux, and R. Pictet. 1997. In vivo footprinting of the interaction of proteinswith DNA and RNA. Methods 11:151–163.
26. Harlow, E., B. J. Franza, and C. Schley. 1985. Monoclonal antibodies specificfor adenovirus early region 1A proteins: extensive heterogeneity in earlyregion 1A products. J. Virol. 55:533–546.
27. Helin, K. 1998. Regulation of cell proliferation by the E2F transcriptionfactors. Curr. Opin. Genet. Dev. 8:28–35.
28. Helin, K., E. Harlow, and A. R. Fattaey. 1993. Inhibition of E2F-1 transac-tivation by direct binding of the retinoblastoma protein. Mol. Cell. Biol.13:6501–6508.
29. Helin, K., J. A. Lees, M. Vidal, N. Dyson, E. Harlow, and A. Fattaey. 1992.A cDNA encoding a pRB-binding protein with properties of the transcrip-tion factor E2F. Cell 70:337–350.
30. Helin, K., C.-L. Wu, A. R. Fattaey, J. A. Lees, B. D. Dynlacht, C. Ngwu, andE. Harlow. 1993. Heterodimerization of the transcription factors E2F-1 andDP-1 leads to cooperative transactivation. Genes Dev. 7:1850–1861.
31. Hijmans, E. M., P. M. Voorhoeve, R. L. Beijersbergen, L. J. van’t Veer, andR. Bernards. 1995. E2F-5, a new E2F family member that interacts with p130in vivo. Mol. Cell. Biol. 15:3082–3089.
32. Hinds, P. W., S. Mittnacht, V. Dulic, A. Arnold, S. I. Reed, and R. A.Weinberg. 1992. Regulation of retinoblastoma protein functions by ectopicexpression of human cyclins. Cell 70:993–1006.
33. Hsiao, K.-M., S. L. McMahon, and P. J. Farnham. 1994. Multiple DNAelements are required for the growth regulation of the mouse E2F1 pro-moter. Genes Dev. 8:1526–1537.
34. Hu, Q., C. Bautista, G. Edwards, D. Defeo-Jones, R. Jones, and E. Harlow.1991. Antibodies specific for the human retinoblastoma protein identify afamily of related polypeptides. Mol. Cell. Biol. 11:5792–5799.
35. Huet, X., J. Rech, A. Plet, A. Vie, and J. M. Blanchard. 1996. Cyclin Aexpression is under negative transcriptional control during the cell cycle.Mol. Cell. Biol. 16:3789–3798.
36. Hurford, R. K. J., D. Cobrinik, M.-H. Lee and N. Dyson. 1997. pRB andp107/p130 are required for the regulated expression of different sets of E2Fresponsive genes. Genes Dev. 11:1447–1463.
37. Ikeda, M. A., L. Jakoi, and J. R. Nevins. 1996. A unique role for the Rbprotein in controlling E2F accumulation during cell growth and differentia-tion. Proc. Natl. Acad. Sci. USA 93:3215–3220.
38. Johnson, D. G., K. Ohtani, and J. R. Nevins. 1994. Autoregulatory control ofE2F1 expression in response to positive and negative regulators of cell cycleprogression. Genes Dev. 8:1514–1525.
39. Karlseder, J., H. Rothender, and E. Wintersberger. 1996. Interaction of Sp1with the growth- and cell cycle-regulated transcription factor E2F. Mol. Cell.Biol. 16:1659–1667.
40. Kelly, T. J., G. S. Martin, S. L. Forsburg, R. J. Stephen, A. Russo, and P.Nurse. 1993. The fission yeast cdc181 gene product couples S phase toSTART and mitosis. Cell 74:371–382.
41. Koch, C., and K. Nasmyth. 1994. Cell cycle regulated transcription in yeast.Curr. Opin. Cell Biol. 6:451–459.
42. Kovesdi, I., R. Reichel, and J. R. Nevins. 1986. Identification of a cellulartranscription factor involved in E1A trans-activation. Cell 45:219–228.
43. Krude, T., M. Jackman, J. Pines, and R. A. Laskey. 1997. Cyclin/Cdk-dependent initiation of DNA replication in a human cell-free system. Cell88:109–119.
44. Lam, E. W.-F., and R. J. Watson. 1993. An E2F-binding site mediates
cell-cycle regulated repression of mouse B-myb transcription. EMBO J.12:2705–2713.
45. Lees, J. A., M. Saito, M. Vidal, M. Valentine, T. Look, E. Harlow, N. Dyson,and K. Helin. 1993. The retinoblastoma protein binds to a family of E2Ftranscription factors. Mol. Cell. Biol. 13:7813–7825.
46. Lennon, G., C. Auffray, M. Polymeropoulos, and M. B. Soares. 1996. TheI.M.A.G.E. consortium: an integrated molecular analysis of genomes andtheir expression. Genomics 33:151–152.
47. Liang, C., M. Weinreich, and B. Stillman. 1995. ORC and Cdc6p interactand determine the frequency of initiation of DNA replication in the genome.Cell 81:667–676.
48. Lindeman, G. J., S. Gaubatz, D. M. Livingston, and D. Ginsberg. 1997. Thesubcellular localization of E2F-4 is cell-cycle dependent. Proc. Natl. Acad.Sci. USA 94:5095–5100.
49. Lukas, J., B. O. Petersen, K. Holm, J. Bartek, and K. Helin. 1996. Deregu-lated expression of E2F family members induces S-phase entry and over-comes p16INK4A-mediated growth suppression. Mol. Cell. Biol. 16:1047–1057.
50. Means, A. L., J. E. Slansky, S. L. McMahon, M. W. Knuth, and P. J.Farnham. 1992. The HIP binding site is required for growth regulation of thedihydrofolate reductase promoter. Mol. Cell. Biol. 12:1054–1063.
51. Moberg, K., M. A. Starz, and J. A. Lees. 1996. E2F-4 switches from p130 top107 and pRB in response to cell cycle reentry. Mol. Cell. Biol. 16:1436–1449.
52. Mueller, P. R., and B. Wold. 1989. In vivo footprinting of a muscle specificenhancer by ligation mediated PCR. Science 246:780–786.
53. Muller, H., M. C. Moroni, E. Vigo, B. O. Petersen, J. Bartek, and K. Helin.1997. Induction of S-phase entry by E2F transcription factors depends ontheir nuclear localization. Mol. Cell. Biol. 17:5508–5520.
54. Muzi-Falconi, M., G. W. Brown, and T. J. Kelly. 1996. cdc181 regulatesinitiation of DNA replication in Schizosaccharomyces pombe. Proc. Natl.Acad. Sci. USA 93:1666–1670.
55. Muzi-Falconi, M., G. W. Brown, and T. J. Kelly. 1996. DNA replication:controlling initiation during the cell cycle. Curr. Biol. 6:229–233.
56. Neuman, E., E. K. Flemington, W. R. Sellers, and W. G. Kaelin, Jr. 1994.Transcription of the E2F-1 gene is rendered cell cycle dependent by E2FDNA-binding sites within its promoter. Mol. Cell. Biol. 14:6607–6615. (Au-thor’s correction, 15:4660, 1995.)
57. Nishitani, H., and P. Nurse. 1995. p65cdc18 plays a major role controlling theinitiation of DNA replication in fission yeast. Cell 83:397–405.
58. Ohtani, K., J. DeGregori, G. Leone, D. R. Herendeen, T. J. Kelly, and J. R.Nevins. 1996. Expression of the HsOrc1 gene, a human ORC1 homolog, isregulated by cell proliferation via the E2F transcription factor. Mol. Cell.Biol. 16:6977–6984.
59. Ohtani, K., J. DeGregori, and J. R. Nevins. 1995. Regulation of the cyclin Egene by transcription factor E2F1. Proc. Natl. Acad. Sci. USA 92:12146–12150.
60. Pearson, B. E., H. P. Nasheuer, and T. S. Wang. 1991. Human DNA poly-merase alpha gene: sequences controlling expression in cycling and serum-stimulated cells. Mol. Cell. Biol. 11:2081–2095.
61. Petersen, B. O., and K. Helin. Unpublished data.62. Piatti, S., T. Bohm, J. H. Cocker, J. F. X. Diffley, and K. Nasmyth. 1996.
Activation of S-phase-promoting CDKs in late G1 defines a “point of noreturn” after which Cdc6 synthesis cannot promote DNA replication in yeast.Genes Dev. 10:1516–1531.
63. Piatti, S., C. Lengauer, and K. Nasmyth. 1995. Cdc6 is an unstable proteinwhose de novo synthesis in G1 is important for the onset of S phase and forpreventing a ‘reductional’ anaphase in the budding yeast Saccharomycescerevisiae. EMBO J. 14:3788–3799.
64. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, N.Y.
65. Santocanale, C., and J. F. X. Diffley. 1996. ORC- and Cdc6-dependentcomplexes at active and inactive chromosomal replication origins in Saccha-romyces cerevisiae. EMBO J. 15:6671–6679.
66. Sardet, C., M. Vidal, D. Cobrinik, Y. Geng, C. Onufryk, A. Chen, and R. A.Weinberg. 1995. E2F-4 and E2F-5, two members of the E2F family, areexpressed in the early phases of the cell cycle. Proc. Natl. Acad. Sci. USA92:2403–2407.
67. Schulze, A., K. Zerfass, D. Spitkovsky, S. Middendorp, J. Berges, K. Helin,P. Jansen-Durr, and B. Henglein. 1995. Cell cycle regulation of the cyclin Agene promoter is mediated by a variant E2F site. Proc. Natl. Acad. Sci. USA92:11264–11268.
68. Sears, R., K. Ohtani, and J. R. Nevins. 1997. Identification of positively andnegatively acting elements regulating expression of the E2F2 gene in re-sponse to cell growth signals. Mol. Cell. Biol. 17:5227–5235.
69. Sellers, W. R., J. W. Rodgers, and W. G. Kaelin, Jr. 1995. A potent transre-pression domain in the retinoblastoma protein induces a cell cycle arrestwhen bound to E2F sites. Proc. Natl. Acad. Sci. USA 92:11544–11548.
70. Sherr, C. J. 1996. Cancer cell cycles. Science 274:1672–1677.71. Slansky, J., Y. Li, W. G. Kaelin, and P. J. Farnham. 1993. A protein syn-
thesis-dependent increase in E2F1 mRNA correlates with growth regulation
6696 HATEBOER ET AL. MOL. CELL. BIOL.
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from

of the dihydrofolate reductase promoter. Mol. Cell. Biol. 13:1610–1618.72. Slansky, J. E., and P. J. Farnham. 1996. Introduction to the E2F family:
protein structure and gene regulation. Curr. Top. Microbiol. Immunol. 208:1–30.
73. Stillman, B. 1996. Cell cycle control of DNA replication. Science 274:1659–1664.
74. Tanaka, T., D. Knapp, and K. Nasmyth. 1997. Loading of an Mcm proteinonto DNA replication origins is regulated by Cdc6p and CDKs. Cell 90:649–660.
75. Tommasi, S., and G. P. Pfeifer. 1995. In vivo structure of the human cdc2promoter: release of a p130–E2F-4 complex from sequences immediatelyupstream of the transcription initiation site coincides with induction of cdc2expression. Mol. Cell. Biol. 15:6901–6913.
76. Vigo, E., and K. Helin. Unpublished data.77. Weintraub, S. J., C. A. Prater, and D. C. Dean. 1992. Retinoblastoma protein
switches the E2F site from positive to negative element. Nature 358:259–261.78. Wells, J., P. Held, S. Illenye, and N. H. Heintz. 1996. Protein-DNA interac-
tions at the major and minor promoters of the divergently transcribed dhfr
and rep3 genes during the Chinese hamster ovary cell cycle. Mol. Cell. Biol.16:634–647.
79. Williams, R. S., R. V. Shohet, and B. Stillman. 1997. A human proteinrelated to yeast Cdc6p. Proc. Natl. Acad. Sci. USA 94:142–147.
80. Zerfass-Thome, K., A. Schulze, W. Zwerschke, B. Vogt, K. Helin, J. Bartek,B. Henglein, and P. Jansen-Durr. 1997. p27KIP1 blocks cyclin E-dependenttransactivation of cyclin A gene expression. Mol. Cell. Biol. 17:407–415.
81. Zhu, L., S. van den Heuvel, K. Helin, A. Fattaey, M. Ewen, D. Livingston, N.Dyson, and E. Harlow. 1993. Inhibition of cell proliferation by p107, arelative of the retinoblastoma protein. Genes Dev. 7:1111–1125.
82. Zhu, L., L. Zhu, E. Xie, and L. S. Chang. 1995. Differential roles of twotandem E2F sites in repression of the human p107 promoter by retinoblas-toma and p107 proteins. Mol. Cell. Biol. 15:3552–3562.
83. Zwerschke, W., H. W. Rottjakob, and H. Kuntzel. 1994. The Saccharomycescerevisiae CDC6 gene is transcribed in late mitosis and encodes a ATP/GTPase controlling S phase initiation. J. Biol. Chem. 269:23351–23356.
84. Zwicker, J., and R. Muller. 1997. Cell-cycle regulation of gene expression bytranscriptional repression. Trends Genet. 13:3–6.
VOL. 18, 1998 MAMMALIAN CDC6 REGULATION BY E2F 6697
on January 13, 2016 by guesthttp://m
cb.asm.org/
Dow
nloaded from





























