Embed Size (px)
Citation preview

DOI: 10.1634/stemcells.2005-0315 2006;24;531-546; originally published online Nov 17, 2005; Stem Cells
Thomas C. Schulz, Anirban Maitra, Mahendra S. Rao and Jonathan M. Auerbach Toumadje, Sandii N. Brimble, Eric S. Sherrer, Elizabeth W. Uhl, William J. Freed,
Todd W. Plaia, Richard Josephson, Ying Liu, Xianmin Zeng, Carol Ording, Arazdordi Line, BG01V: A Tool for Human Embryonic Stem Cell Research
Characterization of a New NIH-Registered Variant Human Embryonic Stem Cell
This information is current as of April 19, 2007
http://www.StemCells.com/cgi/content/full/24/3/531located on the World Wide Web at:
The online version of this article, along with updated information and services, is
1066-5099. Online ISSN: 1549-4918. 260, Durham, North Carolina, 27701. © 2006 by AlphaMed Press, all rights reserved. Print ISSN:Journal is owned, published, and trademarked by AlphaMed Press, 318 Blackwell Street, Suite STEM CELLS® is a monthly publication, it has been published continuously since 1983. The
genetics and genomics; translational and clinical research; technology development.embryonic stem cells; tissue-specific stem cells; cancer stem cells; the stem cell niche; stem cell STEM CELLS®, an international peer-reviewed journal, covers all aspects of stem cell research:
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

Characterization of a New NIH-Registered Variant HumanEmbryonic Stem Cell Line, BG01V: A Tool for HumanEmbryonic Stem Cell Research
TODD W. PLAIA,a RICHARD JOSEPHSON,a YING LIU,b XIANMIN ZENG,c CAROL ORDING,a
ARAZDORDI TOUMADJE,a SANDII N. BRIMBLE,d ERIC S. SHERRER,d ELIZABETH W. UHL,e WILLIAM J. FREED,c
THOMAS C. SCHULZ,d ANIRBAN MAITRA,f MAHENDRA S. RAO,b JONATHAN M. AUERBACHa
aStem Cell Center, American Type Culture Collection, Manassas, Virginia, USA; bLaboratory of Neuroscience,
National Institute on Aging, National Institutes of Health, Baltimore, Maryland, USA; cCellular Neurobiology
Branch, National Institute on Drug Abuse, National Institutes of Health, Baltimore, Maryland, USA; dBresaGen,
Inc., Athens, Georgia, USA; eDepartment of Veterinary Pathology, College of Veterinary Medicine, The University
of Georgia, Athens, Georgia, USA; fDepartment of Pathology, Oncology, and Genetic Medicine, Johns Hopkins
University School of Medicine, Baltimore, Maryland, USA
Key Words. Embryonic stem cell • hESC karyotype • Characterization • BG01V • NTERA-2
ABSTRACT
Human embryonic stem cells (hESCs) offer a renewablesource of a wide range of cell types for use in research andcell-based therapies. Characterizing these cells provides im-portant information about their current state and affordsrelevant details for subsequent manipulations. For example,identifying genes expressed during culture, as well as theirtemporal expression order after passaging and conditionsinfluencing the formation of all three germ layers may behelpful for the production of functional beta islet cells usedin treating type I diabetes. Although several hESC lines havedemonstrated karyotypic instability during extended time inculture, select variant lines exhibit characteristics similar totheir normal parental lines. Such variant lines may be ex-cellent tools and abundant sources of cells for pilot studiesand in vitro differentiation research in which chromosome
number is not a concern, similar to the role currently playedby embryonal carcinoma cell lines. It is crucial that the cellsbe surveyed at a genetic and proteomic level during exten-sive propagation, expansion, and manipulation in vitro.Here we describe a comprehensive characterization of thevariant hESC line BG01V, which was derived from thekaryotypically normal, parental hESC line BG01. Our char-acterization process employs cytogenetic analysis, short tan-dem repeat and HLA typing, mitochondrial DNA sequenc-ing, gene expression analysis using quantitative reversetranscription-polymerase chain reaction and microarray,assessment of telomerase activity, methylation analysis, andimmunophenotyping and teratoma formation, in addition toscreening for bacterial, fungal, mycoplasma, and humanpathogen contamination. STEM CELLS 2006;24:531–546
INTRODUCTIONSince the first successful derivations of human embryonic stemcells (hESCs) from isolated inner cell mass cells of surplus invitro fertilization embryos, rapid progress has been achieved inthe characterization and manipulation of undifferentiated hESCsin vitro [1, 2]. However, the state of the art for working withhESCs is far from ideal. Much of the current knowledge regard-ing conditions for deriving and culturing the undifferentiatedcells was based on work performed on murine and nonhumanprimate ESCs, cells derived from embryonal carcinomas (ECs),
and more recently, on the work accomplished on a relativelylimited number of isolated hESC lines.
The availability of hESC lines in the rapidly advancing fieldof stem cell biology is currently limited in the United States.Restricted hESC access is primarily due to high cost, limitedgovernment funding, and complicated intellectual property is-sues. A temporary and imperfect solution to these conditions hasbeen the use of EC cells in place of hESC lines for certainapplications. As the number of potential therapies and differentresearch applications increases, so does the demand for the
Correspondence: Jonathan Auerbach, Ph.D., Stem Cell Center, American Type Culture Collection, 10801 University Boulevard,Manassas, Virginia 20110, USA. Telephone: 703-365-2809; Fax: 703-365-2790; e-mail: [email protected] Received July 12,2005; accepted for publication November 11, 2005; first published online in STEM CELLS EXPRESS November 17, 2005. ©AlphaMedPress 1066-5099/2006/$20.00/0 doi: 10.1634/stemcells.2005-0315
EMBRYONIC STEM CELLS: CHARACTERIZATION SERIES
STEM CELLS 2006;24:531–546 www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

production and characterization of additional hESC lines grownin conditions more suitable for clinical use. Moreover, theavailability of additional cell lines for comparative studies willenable the establishment of standardized methods of propaga-tion and better refinement of characterization criteria, a prereq-uisite to testing for therapeutic efficacy. Although karyotypi-cally stable relative to other cell lines, hESCs occasionallyundergo spontaneous trisomies during prolonged periods in cul-ture [3]. Here, we propose the establishment and use of suchvariant hESC lines (e.g., BG01V) to better address the imposedrestrictions on normal hESC lines. Data presented here suggestthat stable karyotypic variant hESC lines may more closelyparallel normal hESC lines than EC lines.
The BG01 hESC line was established and characterized byBresaGen, Inc. (Athens, GA, http://www.bresagen.com.au) in2001 and is listed on the National Institutes of Health (NIH)Stem Cell Registry (http://stemcells.nih.gov/research/registry/index.asp) [4, 5]. A stable variant hESC line, designatedBG01V, was derived from the karyotypically normal BG01hESC through routine enzymatic passaging [6, 7]. This cell linecontains known and stable chromosomal aberrations (XXY,�12, �17) yet possesses characteristics similar to its normalparental line. Although karyotypically abnormal, the cell lineproliferates in culture, expresses markers of pluripotency, andretains the ability to differentiate into cell types representative ofthe three germ layers. Variant hESC lines, relative to theirnormal parental lines, are easier to manipulate in vitro andrecover more rapidly after passaging and cryopreservation[3, 6].
Historically, pluripotent human testicular EC cells, such asthe NTERA-2 cl.D1 cell line, have provided a convenient meansand a reasonable model for investigation of the molecular mech-anisms by which stem cells commit to specific lineages duringembryonic development. The NTERA-2 cl.D1 cells were de-rived from the NTERA-2 cell line, which was established froma nude mouse xenograft of TERA2. The TERA2 cell line wasoriginally derived from a metastasis of a human testicular car-cinoma [8, 9]. The NTERA-2 line has been shown to differentiateto neuroectodermal lineages after exposure to retinoic acid orhexamethylene bisacetamide [10, 11]. Although EC cells can pro-vide a convenient and robust experimental system, their differen-tiation capacity is often limited, unlike ESC differentiation [12].
To gain a better understanding of the differentiative poten-tial of the BG01V cell, several genetic and protein-based assayswere used to fully characterize the line. The resulting data werecompared with NTERA-2 cl.D1 (NTERA-2) cells. We proposethat BG01V cells can be useful as an additional or alternative hESCresource for selected investigations in stem cell biology.
MATERIALS AND METHODS
In Vitro Propagation of BG01V and NTERA-2 CellsBG01V hESCs (P10) were obtained from BresaGen, Inc., as afrozen stock. Approximately 1 � 106 BG01V cells were platedinto each of two 9.5-cm2 wells of a six-well culture dish (Corn-ing Life Sciences, Acton, MA, http://www.corning.com/lifesciences) containing a feeder layer of Mitomycin C-treatedCF-1 mouse embryonic fibroblasts (MEFs) (American Type Cul-ture Collection [ATCC] SCRC-1040.2; Manassas, VA, http://www.atcc.org). Cells were cultured at 37°C, 5% CO2. The growth
medium used was Dulbecco’s modified Eagle’s medium (DMEM)/F12 (ATCC 30-2006) supplemented with ESC-Qualified fetal bo-vine serum (FBS) (15%) (ATCC SCRC-30-2020), knockout serumreplacement (KSR) (5%) (Invitrogen Corporation), L-alanyl-L-glu-tamine (2.0 mM) (ATCC 30-2115), MEM nonessential aminoacids (1X) (ATCC 30-2116), �-mercaptoethanol (0.1 mM) (Sig-ma-Aldrich, St. Louis, http://www.sigmaaldrich.com), penicillin(100 IU/ml)/streptomycin (100 �g/ml) (ATCC 30-2300), and basicfibroblast growth factor (bFGF) (4 ng/ml) (R&D Systems, Inc.,Minneapolis, http://www.rndsystems.com). Daily medium changesbegan after the first 48 hours in culture. Colony formation wasvisible within 2 to 3 days. Cells were passaged every 4 to 5 daysusing collagenase IV (200 Units/ml) (Invitrogen, Carlsbad, CA,http://www.invitrogen.com) for 15 passages (P25).
The hESC line BG01 was obtained from BresaGen, Inc., andcultured as described previously [6]. Briefly, hESCs were main-tained on MEF in DMEM/F12 supplemented with 15% FBS,5% KSR, 2 mM nonessential amino acids, 2 mM L-glutamine,50 �g/ml Penn-Strep (all from Invitrogen), 0.1 mM �-mercap-toethanol (Specialty Media, Phillipsburg, NJ, http://www.specialtymedia.com), and 4 ng/ml bFGF (Sigma-Aldrich). Cellswere passaged by incubation in Cell Dissociation Buffer (In-vitrogen), dissociated, and then seeded at approximately 20,000cells per cm2. Under such culture condition, the ESCs werepassaged every 4 to 5 days.
NTERA-2 cells were obtained from the ATCC (ATCCCRL-1973). Cells were thawed and plated into six-well culturedishes (Corning Life Sciences) without a feeder layer. Thegrowth medium used was DMEM (ATCC 30-2002) supple-mented as described above. No bFGF was used to suppressNTERA-2 differentiation. Cells were cultured at 37°C, 5% CO2,and the medium was changed every 24 hours. Cells were pas-saged every 4 days by mechanical scraping.
Karyotype AnalysisThe karyotype analysis was performed using a standard G-banding technique. Cells cultured in a T75 culture flask weretreated with 0.05 �g/ml Colcemid (Invitrogen Corporation15210-040) for 1 hour, followed by dissociation using 0.25%trypsin/0.53 mM EDTA in Hanks’ Balanced Salt Solution(HBSS) without calcium or magnesium (ATCC 30-2101). Thecells were then collected by centrifugation (5 minutes at 240 g)and gently resuspended in a 0.06 M KCl hypotonic solution andplaced in an incubator at 37oC for 25 minutes. The hypotoniceffect was halted by the addition of 3:1 Carnoy’s Fixative(methanol/glacial acetic acid). The cells were collected by cen-trifugation and resuspended by gentle mixing and run through aseries of fixes prior to slide preparation. Metaphase spreadswere prepared on glass microscope slides, exposed briefly to a2% Enzar-T trypsin 40X (7000-65; Mediatech, Inc., Herndon,VA, http://www.cellgro.com) in HBSS (20-021-CV; Mediatech,Inc.) solution, and stained using a 2:1 Gurr/Giemsa stain. A totalof 43 metaphase spreads were analyzed by microscopy.
Short Tandem Repeat AnalysisFrozen BG01V cells grown on MEF feeders were resuspendedin phosphate-buffered saline (PBS) (ATCC SCRR-2201). A20-�l aliquot was spotted on a labeled FTA card (Whatman,Brentford, Middlesex, U.K., http://www.whatman.com) and al-lowed to dry. The FTA card lyses the cells on contact and binds
532 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

the DNA to the paper surface. Prior to polymerase chain reac-tion (PCR), a portion of the dried spot was removed with aHarris punch, washed three times with Purification Reagent(Whatman), washed once with TE Buffer (Tris-EDTA, pH 8.0),and allowed to dry. Short tandem repeat (STR) analysis wasconducted using a multiplex PCR-based PowerPlex 1.2 kit (Pro-mega, Madison, WI, http://www.promega.com). Loci analyzedinclude D5S818, D13S317, D7S820, D16S539, vWA, TH01,Amelogenin, TP0X, and CSF1P0. Electropherogram data werecollected on an ABI 310 Genetic Analyzer. Data were analyzedusing Genescan 3.1 and Genotyper 2.0 (Applied BioSystems,Foster City, CA, http://www.appliedbiosystems.com). The re-sulting profiles were imported into an in-house database andscreened against all other baseline profiles of all samples testedby the ATCC. STR analysis of the NTERA-2 cell line wasperformed similarly using isolated genomic DNA.
HLA TypingGenomic DNA was isolated from BG01V cells grown onMEFs using the GenElute Mammalian Genomic DNA Mini-prep Kit (Sigma-Aldrich). HLA DNA typing was performedby using hybridization of PCR amplified DNA with sequencespecific oligonucleotide probes (Tepnel Lifecodes Corpora-tion, Stamford, CT, http://www.tepnel.com). The target DNAis amplified by PCR and then allowed to denature and rehy-bridize to complementary DNA probes conjugated to fluo-rescently coded microspheres. A flow analyzer identifies thefluorescent intensity on each microsphere, and the deter-mined HLA type is based on the reaction pattern comparedwith patterns associated with public HLA gene sequences.Assays were performed to determine the HLA-A, HLA-B,HLA-C, HLA-DRB, and HLA-DQB loci.
Mitochondrial DNA AnalysisThe entire coding sequence of the human mitochondrial genomeof BG01V was sequenced using an automated sequencing mi-croarray (Human MitoChip; Affymetrix, Santa Clara, CA,http://www.affymetrix.com) as previously described [13].Briefly, the MitoChip can sequence 29,366 base pairs (bp) ofdouble stranded DNA, which includes 980 bp of plasmid DNAsequence as a control for chip hybridization. Both strands of theentire mitochondrial coding region (nucleotides 573–16,024;15,451 bp) are tiled once on the array. The forward and reversestrands of an additional 12,935 bp of the mitochondrial DNA(the coding region minus 12S and 16S RNA sequences) are tiledon the remaining features and thus provide an inbuilt duplicationof sequence data for approximately 84% of the mitochondrialcoding region. Automated sequence analysis of hybridized mi-croarrays was performed using Affymetrix GeneChip DNAAnalysis Software (GDAS) version 3.0, using a modification ofa previously described ABACUS (adaptive background geno-type-calling scheme) [14]. Once a batch analysis is completed,the GDAS software generates a report containing individual,total number, and percentages of base calls within the batch anda detailed case-by-case list of genotype variations vis-a-vis thereference human mitochondrial DNA sequence (revised Cam-bridge reference sequence [RCRS]). The MitoChip has previ-ously been demonstrated [13] to have more than 99.99% repro-ducibility of base calls in replicate experiments and is thus ahighly automated and sensitive microarray tool for human mi-
tochondrial DNA sequencing and mutation detection. To com-pare the mitochondrial sequence of BG01V to its parent cell line(BG01), we also sequenced the entire coding sequence from thelatter, using DNA obtained from an early cell passage.
Methylation AnalysisMethylation-specific PCR (MSP) was used to determine themethylation status of a select sample of specific developmentaland imprinted genes in BG01V and NTERA-2 bisulfite-treatedgenomic DNA. MSP exploits sequence differences existingbetween methylated and unmethylated alleles after treatmentwith sodium bisulfite. The frequencies of the CpG dinucleotideswithin the target sequences help emphasize the sequence differ-ences. Oligonucleotide primers for a given locus are designed todirectly discriminate methylated from unmethylated sequencesin bisulfite-modified DNA. The results are obtained immedi-ately after PCR amplification and gel electrophoresis withoutthe need for further restriction or sequencing analysis. Selectedimprinted gene targets examined included the PWS/AS (snrpn)locus [15], the H19 promoter region (GenBank AF125183) [16,17], and the DLK1/MEG3 differentially methylated region(DMR) [18]. Developmentally important genes included the Xistpromoter region (GenBank U50908 [gi:1575006]) [19, 20], theOct-3/4 promoter region (GenBank AJ297527) [21], and theNotch1 promoter region (GenBank NT_024000 [gi:29793214])[22].
Genomic DNA from the BG01V hESCs grown on feederlayers and cultured NTERA-2 cells was isolated and purifiedusing the GenElute Mammalian Genomic DNA Miniprep Kitand protocol (Sigma-Aldrich) as provided by the manufacturer.Bisulfite conversion of the genomic DNA was accomplishedusing the EZ DNA Methylation Kit (Zymo Research, Orange,CA, http://www.zymoresearch.com) and protocol as provided.If necessary, promoter regions of genes were verified usingPromoterInspector software (Genomatix Software GmbH,Munchen, Germany, http://www.genomatix.de). MSP primerswere designed using MethPrimer software (http://itsa.ucsf.edu/�urolab/methprimer/index1.html) [23] with the exception ofthose already published for PWS/AS (snrpn) [15] and DLK1/MEG3 [16] (Table 1). Amplification and gel analysis wereperformed as described [15]. Bisulfite-converted CpGenomeUniversal Methylated DNA (Chemicon, Temecula, CA, http://www.chemicon.com) and bisulfite-converted male human pla-cental DNA (Sigma-Aldrich) were used as positive controls.Negative controls consisted of amplification reactions withouttemplate.
Telomerase ActivityTelomerase activity in the cultured BG01V cells and NTERA-2cells was examined using the TRAPeze Telomerase DetectionKit (Chemicon). The BG01V cells were separated from feedercells either by sorting SSEA-4 expressing BG01V cells using aBD FACS Aria Cell Sorter (BD Biosciences, San Diego, http://www.bdbiosciences.com) or by mechanical scraping of the ESCcolonies from the culture dish. The lysate preparation and thetelomeric repeat amplification protocol (TRAP) assay were per-formed according to the manufacturer’s instructions, except forthe PCR amplification steps. The PCR amplification was per-formed in three steps, instead of two steps suggested by thevendor: 32 cycles of 94°C for 30 seconds, 50°C for 30 seconds,
533Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

and 72°C for 90 seconds with one cycle of 72°C for 10 minutes.The TRAP products, along with appropriate size markers, wereelectrophoresed on a 10% nondenaturing polyacrylamide gelunder 50 mM Tris-borate buffer and visualized by SYBR Green(Molecular Probes, Inc., Eugene, OR, http://probes.invitrogen.com) fluorescence using a gel documentation imaging system(UVP, Inc., Upland, CA, http://www.uvp.com).
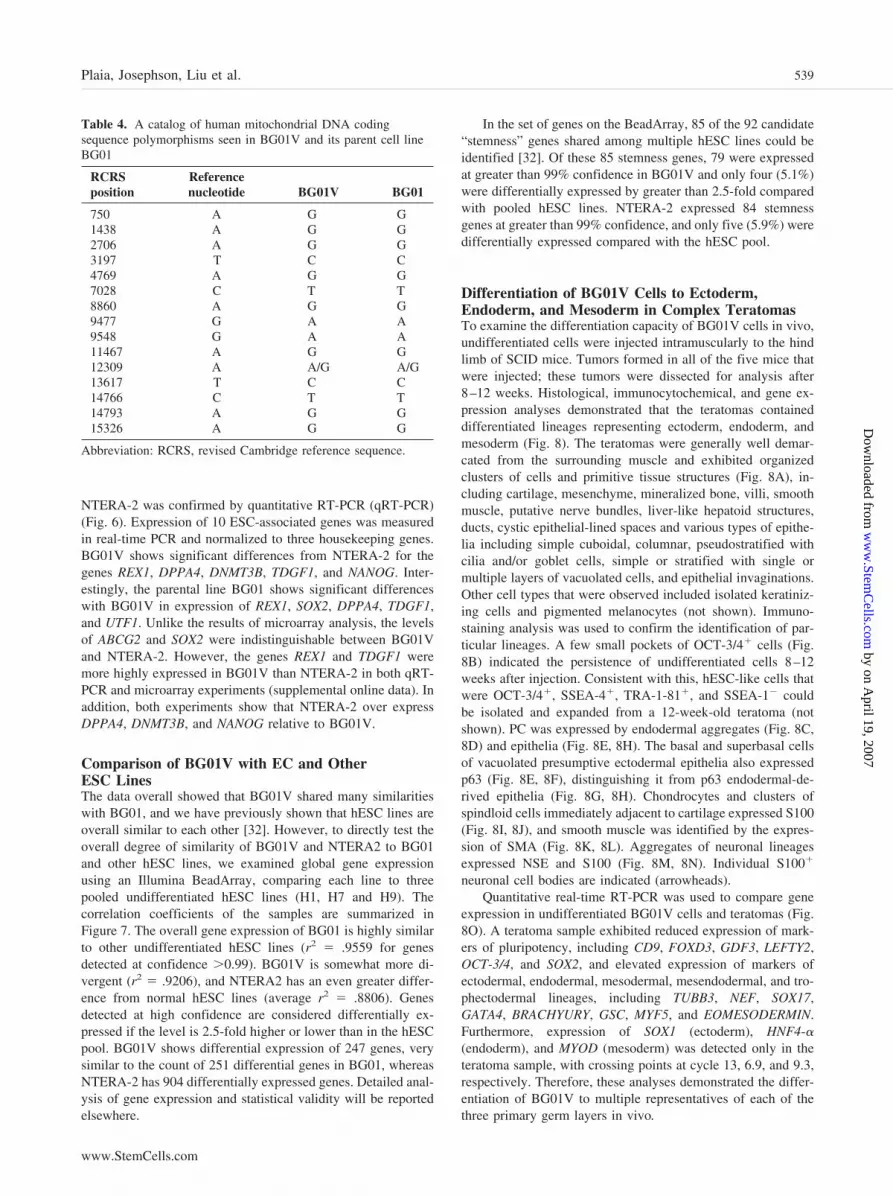
Immunophenotyping of Undifferentiated CellsUndifferentiated BG01V ESCs and NTERA-2 EC cells wereexamined for expression of pluripotent hESC markers usingimmunocytochemistry [24]. Cells were fixed in 2% paraformal-dehyde for at least 20 minutes at room temperature. They werewashed in PBS and then incubated in 3% normal goat serum(NGS) to inhibit nonspecific binding. Saponin detergent (0.5%)was used to permeablize cell membranes on samples stained forintracellular markers. Cells were then assayed with monoclonalantibodies specific for Oct-3/4 (1:250) (BD Biosciences Trans-duction Laboratories, Lexington, KY, http://www.bdbiosciences.com), SSEA-1 (1:100), SSEA-4 (1:50), TRA-1-60 (1:100), andTRA-1-81 (1:100) (all four from Chemicon), washed usingPBS/1% NGS to remove any unbound protein, and then incubatedwith an Alexa Fluor 488 conjugated goat anti-mouse IgG (H�L)antibody (1:750) (Molecular Probes, Inc.). Positively stained cellswere visualized using an epifluorescence microscope.
Endogenous alkaline phosphatase activity in BG01V andNTERA-2 cells was detected using the ELF 97 EndogenousAlkaline Phosphatase Detection Kit (Molecular Probes, Inc.)according to the manufacturer’s instructions. Cells cultured on12-mm round glass cover slips in 24-well plates (Corning LifeSciences) were treated with 2% paraformaldehyde for 20 min-utes at room temperature. The cells were washed with PBS,treated with 0.2% Tween-20 for 20 minutes at room tempera-ture, and rinsed with PBS. Fixed cells were then incubated witha filtered 1:20 dilution of the phosphatase substrate in situ. Theimmunoassayed colonies and the reaction were monitored usingan epifluorescence microscope. The reaction was terminatedusing a stop solution consisting of PBS, 25 mM EDTA, and 5mM levamisole, pH 8.0. Cells were rinsed with PBS beforemounting on glass microscope slides.
Gene Expression of Undifferentiated Cells UsingQuantitative RT-PCRTotal RNA was isolated by lysing cells in Trizol LS (Invitro-gen) according to instructions. Two micrograms of total RNAwas treated with DNAseI (Promega) 25°C for 15 minutes,65°C for 10 minutes, then reverse-transcribed using theiScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, http://www.bio-rad.com). One fortieth of the cDNA synthesis re-action was used as template for each real-time PCR using iQSYBR Green Supermix (Bio-Rad). For each primer set, Ta isthe annealing temperature determined empirically using tem-plate cDNA from NTERA-2 cells (Table 2). Tr is the tem-perature at which the SYBR Green fluorescence is read,chosen by examining the melt curves of the PCR products.PCR was run in an iCycler iQ Real Time Detection System(Bio-Rad) for 50 cycles of 95°C for 15 seconds, Ta 30seconds, 72°C for 45 seconds, and Tr for 15 seconds. Therelative amounts of PCR product were quantified using therelative threshold cycle (��Ct) method corrected for effi-ciency of each amplification [25]. The gene quantities foreach sample were normalized against the geometric mean ofexpression of the housekeeping genes GAPDH, �-actin, andtata-binding protein (tbp). Statistically significant (p � .05)differences in gene expression were determined by the Stu-dent’s t test.
Gene Expression of Undifferentiated CellsUsing MicroarraysOligo microarray experiments for undifferentiated BG01V andNTERA-2 cells were conducted with procedures of G4110Bfrom Agilent Technologies, Inc. (Palo Alto, CA, http://www.agilent.com). The chip contains 20,170 human probes. RNAextracted from BG01V and NTERA-2 was labeled with Cy5,and human universal RNA (huURNA) (Clontech, Palo Alto,CA, http://www.clontech.com) was labeled with Cy3. Microar-ray data were analyzed with the Agilent G2567AA FeatureExtraction Software (version 7.5). The huURNA is a high-quality reference standard for microarray experiments whichpermits multiple laboratories working on different microarrayplatforms to compare and share data. The output data were used
Table 1. Oligonucleotide MSP primer sequences
Target L-Unmethylated primer L-Methylated primer
PWS/AS (snrpn) locus 5�-GTAGGTTGGTGTGTATGTAGGT-3� 5�-TAAATAAGTACGTTTGCGCGGTC-3�H19 promoter region 5�-TTTTAGGAATGTGAGGTTTGAGTT-3� 5�-TTTAGGAACGTGAGGTTTGAGTC-3�Xist promoter region 5�-GGGTTGGAGTGTAGTGGTATTATTTT-3� 5�-GGTGGAGCGTAGTGGTATTATTTC-3�Oct-4 promoter region 5�-GGGGGAAAAGTTTTAAAAGATTAT-3� 5�-GGGGGAAAAGTTTTAAAAGATTTAC-3�Notch1 promoter region 5�-GTTTGGTTTAGTGTATGGTGTATAT-3� 5�-CGGTTTAGCGTACGGTGTATAC-3�DLK1/MEG3 region 5�-GAGGATGGTTAGTTATTGGGGT-3� 5�-GTTAGTAATCGGGTTTGTCGG-3�
Target R-Unmethylated primer R-Methylated primer
PWS/AS (snrpn) locus 5�-ACATCAAAATCTCCAACAACCA-3� 5�-AACCTTACCCGCTCCATCGCG-3�H19 promoter region 5�-CCTACTACTCCCTACCTACCAACAC-3� 5�-CCTACTACTCCCTACCTACCAACG-3�Xist promoter region 5�-AAACCAACCTAACCAACATAACAAA-3� 5�-AAACCAACCTAACCAACATAACG-3�Oct-4 promoter region 5�-ACTAAACACACTAACCTCTATCAAC-3� 5�-ACTAAACACGCTAACCTCTATCGAC-3�Notch1 promoter region 5�-TACAAAAAAATCCAACTCTAACAAT-3� 5�-TACGAAAAAATCCGACTCTAACG-3�DLK1/MEG3 region 5�-CCACCATAACCAACACCCTATAATCACA-3� 5�-AATCATAACTCCGAACACCCGCG-3�
Abbreviation: MSP, methylation-specific polymerase chain reaction.
534 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

for further computation. UniGene Build no. 176 was used toretrieve UniGene IDs. There are two replicates of microarraydata. The z score method [26] was used to analyze data andidentify significantly elevated genes.
For Illumina BeadArray, sample amplification was per-formed using 100 ng of total RNA as input material by themethod of Van Gelder et al. [27] using the Illumina RNAAmplification kit (Ambion, Inc., Austin, TX, http://www.ambi-on.com) following the Manufacturer’s instructions; labeling wasachieved by use of the incorporation of biotin-16-UTP(PerkinElmer Life and Analytical Sciences, Boston, http://www.perkinelmer.com) present at a ratio of 1:1 with unlabeled UTP.Labeled, amplified material (700 ng per array) was hybridized toa pilot version of the Illumina Ref-8 BeadChip according to themanufacturer’s instructions (Illumina, Inc., San Diego, http://www.illumina.com). Arrays were washed and then stained withAmersham fluorolink streptavidin-Cy3 (GE Healthcare Bio-Sciences, Little Chalfont, U.K., http://www.gehealthcare.com)in accordance with the BeadChip manual. Arrays were scannedwith an Illumina BeadArray Reader confocal scanner accordingto the manufacturer’s instructions. Array data processing andanalysis were performed using Illumina BeadStudio software.
Generation and Analysis of Teratomasin SCID MiceBG01V hESC were maintained and harvested as described. Five6-week-old male Nod CB17-Prkdc (severe combined immuno-deficient [SCID])/J mice (Jackson Laboratory, Bar Harbor, ME,http://www.jax.org) were each injected with a cell suspension of4 � 106 cells using a 28-gauge half-inch needle in the gastro-cnemius muscle of the hind limb. Palpable tumors formed in allmice and were dissected after 8 to 12 weeks. The tumors werefixed in 10% formalin (Fisher Scientific International, Hampton,NH, http://www.fisherscientific.com), dehydrated, and embed-ded in paraffin. Sections were cut at 4 �M and examinedhistologically after hematoxylin and eosin staining or immuno-histochemical staining. Sections were processed for immuno-histochemical analysis using antibodies to pancytokeratin (PC,catalog no. MU181-UC; BioGenex, San Ramon, CA, http://www.biogenex.com), neuron-specific enolase (NSE, catalog no.MUO55-UC; Invitrogen), smooth muscle actin (SMA, catalogno. MO851; DakoCytomation, Glostrup, Denmark, http://www.
dakocytomation.com), p63 (catalog no. 8431; Santa Cruz Bio-technology, Inc., Santa Cruz, CA, http://www.scbt.com), Oct-3/4 (catalog no. 5279; Santa Cruz Biotechnology, Inc.), andS-100 (catalog no. Z0311; DakoCytomation). All antibodieswere raised in mice against human targets and used at 1:100,except for antibodies against S-100, which were raised in rabbitand used at 1:1000. Immunohistochemistry protocols were asfollows: for p63 and Oct-3/4, sections were microwaved for 15minutes at 400 W in 10 mM sodium citrate buffer (pH 6.0) andstained using Universal DAKO LSAB � Kit, Peroxidase(K0679, DakoCytomation) according to the manufacturer’s in-structions. Staining for PC, S-100, SMA, and NSE was per-formed using a DakoCytomation Autostainer Plus according tomanufacturer’s instructions, with no antigen retrieval (SMA,S-100), pre-treatment with 1% proteinase K for 5 minutes (PC),or with steam for 15 minutes (NSE). All slides were counter-stained with Harris’ hematoxylin. Negative controls were incu-bated with mouse immunoglobulin G reagent grade (Lyophi-lized; Sigma-Aldrich) in a 1:100 dilution.
Gene Expression of Teratomas Using Real-TimeRT-PCRTotal RNA was prepared from a harvested tumor using theTrizol reagent (Invitrogen) and was treated with DNase I (Pro-mega) prior to first-strand cDNA synthesis using 2 �g RNA andthe iScript system (Bio-Rad). Assays-On-Demand (AppliedBioSystems), which are prequalified TaqMan quantitative PCRsets, were used in Micro-Fluidic Cards in the ABI PRISM7900HT Sequence Detection System (Applied BioSystems).Duplicate reaction sets, each containing cDNA generated from0.25 �g RNA, were used to compare the relative gene expres-sion in undifferentiated BG01V and BG01V teratoma popula-tions. The �Ct values were averaged and normalized withGAPDH using the ��Ct method. Fold changes were calculatedas 2-Ct, and the expression ratio of the teratoma sample relativeto the BG01V sample indicated.
Sterility and Pathogen TestingExtensive bacterial and fungal tests were performed on theBG01V and NTERA-2 cell cultures that include incubations inthe following media: HTYE, TSB aerobic, Sabouraud, DMEMwith 10% FBS, Blood Agar aerobic, Blood Agar anaerobic, and
Table 2. qRT-PCR primers for gene expression analysis
Gene Forward primer 5� to 3� Reverse primer 5� to 3� Ta (°C) Tr (°C)
pou5f1 tgcccgaaaccgacactg cttgatcgcttgcccttctg 60.7 84dppa4 aaaagcctccaatgggaatag gcagaggaccagagttcac 57 78dnmt3b cacagacgacacagaggacac atcctgatactctgaactgtctcc 57 78tdgf1 ctgcaacctccgcatccg attagctgggcatggtgacac 60.7 80sox2 agtagtttgctgcctctttaagac cgcttccctcctcctctgg 57 78abcg2 gcgacctgccaatttcaaatg gaagagctgctgagaactgtaag 61.9 78tert cgagggtgaaggcactgttc agtacagctcaggcggcg 60.7 84foxd3 agttccccgcctggcag ggtccagtagttgcccttgc 60.7 84rex1 tgcactaggaagttgaggaatag ggaaatgtctctttagttttgagc 57 78nanog ctccatgaacatgcaacctg ctcgctgattaggctccaac 57 78gapdh tgtccccactgccaacg ccaggatgcccttgaggg 60.7 80�-actin ctggaacggtgaaggtgaca aagggacttcctgtaacaatgca 80 59.2tbp cggctgtttaacttcgcttc ttcttggcaaaccagaaacc 59.2 80
Abbreviation: qRT-PCR, quantitative reverse transcription-polymerase chain reaction.
535Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

THIO anaerobic. The cultures were routinely monitored andreported at 48-hour, 14-day, and 21-day post incubations. Inaddition, mycoplasma testing was performed using a HoechstAssay for each culture.
Pathogen testing was performed by a Clinical LaboratoryImprovement Amendments-approved laboratory (Genetics andIVF Institute, Fairfax, VA, http://www.givf.com). All pathogenassays use “nested” multiplex PCR or reverse transcription(RT)-PCR formats and have a detection threshold of 10 copiesof target DNA or RNA per assay (with an analytical sensitivityof 100%), with a limit of detection (LOD) of two to five copiesper assay. Nucleic acids are extracted using the Qiagen QIAmpViral RNA Kit that yields both intact RNA and DNA. One fifthof each extract is used per 50-�l “nested” multiplex PCR(Platinum Taq DNA Polymerase; Invitrogen). Amplificationproducts are visualized by high-resolution PAGE and SybrGreen staining. For RNA targets, 10-�l of extract is used forOne Step RT-PCR (Invitrogen) followed by second-round PCRprior to the PAGE analysis. For all DNA PCR assays, a humangenomic sequence is amplified as an internal positive control.For RNA viruses (such as hepatitis C virus [HCV]), an endog-enous human mRNA is targeted in all assays. These sample-derived targets must be scored as “detected,” indicating propernucleic acid extraction and amplification, prior to scoring anypathogenic target.
RESULTS
Karyotype and Propagation In VitroAlthough early work on hESC lines supported the conclusionthat these cells are karyotypically normal and stable [4, 5], thereare cases in which the culture conditions employed for theirpropagation apparently do not comprehensively maintain chro-mosomal stability [3, 6]. Parental BG01 hESCs are male (XY)and have a normal complement of 46 chromosomes (data notshown). The BG01V hESC line evolved from routine enzymaticpassaging by BresaGen, Inc., and exhibits a karyotype of 49chromosomes (XXY, �12, �17). Through 25 passages, BG01Vmaintained the karyotype 49XXY, �12, �17 (data not shown). Incomparison, the NTERA-2 cell line is hypotriploid and contains an
average of 12 marker chromosomes (Fig. 1A). Despite the abnor-mal karyotype, BG01V cell colonies grown on MEFs exhibiteduniform flat colonies. These colonies are tightly compacted, and thecells have a high nuclear/cytoplasmic ratio, as do undifferentiatedcells of the parental line BG01 [5]. The BG01V cells exhibitprominent nucleoli, and cell edges are not phase-bright.
The cells had a predictable growth rate and were easy tomaintain in culture (data not shown). After thawing and plating,the BG01V cells recovered rapidly and colony formation wasvisible after 2 days. Enzymatic passaging was possible every4–5 days, enabling efficient expansion of the cell line. In ourhands, the normal parental hESC line, BG01, was much moredifficult to successfully maintain in culture. BG01 had a slightlyslower growth rate, and colony formation was not evident until3–4 days after passaging manually every 5 to 7 days (data notshown). BresaGen, Inc., the originator of this cell line, recom-mends manual passage to avoid differentiation and mutation.Manual passaging by scraping and aspirating individual colo-nies under a microscope greatly increases “hands on” time for invitro maintenance of this cell line, is very labor intensive, andreduces expansion capacity. Like BG01V, NTERA-2 was alsoeasily maintained in vitro. However, this cell line did not exhibituniform colony formation (Fig. 1B).
Identification AssaysTo ensure that work was actually performed on the appropriatecell lines, identification assays were performed and comparedwith previously available data. These assays included STRanalysis and HLA typing. The STR profiles of the BG01V andNTERA-2 were determined as described in Materials and Meth-ods (Fig. 2). Loci analyzed for tetra nucleotide STR analysisincluded D5S818, D13S317, D7S820, D16S539, vWA, TH01,TP0X, CSF1P0, and Amelogenin for gender determination. Theamplicons were separated by capillary electrophoresis and an-alyzed using Genescan 3.1 and Genotyper 2.0 software fromApplied BioSystems. Each peak in the resulting electrophero-gram represented an allele that is alphanumerically scored andwas then entered into a database. As expected, the number ofrepeats and the location and peak height in the BG01V STRprofile was identical to the STR profile to BG01. The NTERA-2
Figure 1. Karyotype of the BG01V hESC line and the NTERA-2 EC cell line. (A): Karyotype of the variant (BG01V: 49XXY, �12, �17) cellsand karyotype of the NTERA-2 cells. All BG01V cells examined (43 metaphase spreads) had triploidy for chromosomes 12, 17, and X. A gain ofchromosome 9 or loss of chromosome 11 was found at low frequencies (2.5% each). The human NTERA-2 EC line is hypotriploid human cell linewith the modal chromosome number of 63 in 48% of cells examined. However, cells with 62 chromosome counts also occurred at a rather highfrequency (24%). The rate of polyploidy was 1.6%. Approximately 12 marker chromosomes are constantly found in most NTERA-2 cells. Theyinclude der(9)t(1;9)(q25;q34.3), del(1)(q25), der(13)t(11;13)(q13;q34), t(Xq1q), and eight others. At least two markers are found only in some cells.The normal Y chromosome was found in all cells. Only single copies of normal chromosomes 1, 10, 11, and 13 were present. Others were mostlyin two or three copies per cell. (B): Phase-contrast microscopy of undifferentiated BG01V cells grown on an MEF feeder layer showing distinct colonyformation and NTERA-2 cells after 4 days in culture. Abbreviations: EC, embryonal carcinoma; hESC, human embryonic stem cell; MEF, mouseembryonic fibroblast.
536 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

STR profile was identical to original NTERA-2 material (CRL-1973) deposited with ATCC.
The HLA profiles of BG01V and NTERA-2 were generatedas described (Table 3). For purposes of comparison and ofinterest to the user community, we also performed HLA typingon the parental hESC line, BG01. HLA typing at the allelic levelincluded HLA-A, HLA-B, HLA-C, HLA-DRB, and HLA-DQBloci. As can be seen from Table 3, BG01V had an identical HLAprofile to BG01. The NTERA-2 HLA profile was interesting inits homozygosity at each locus.
Mitochondrial DNA SequencingMitochondrial DNA anomalies apparently contribute to, and arecorrelated to, various human diseases [13, 14]. MitochondrialDNA sequence analysis was performed on both BG01V andBG01. MitoChip sequencing yielded an approximately 96%base call rate in both samples (28,244 bp/29,366 bp for BG01Vand 28,134 bp/29,366 bp for BG01); the remaining calls weredeemed by GDAS version 3.0 (Affymetrix) to be of unsatisfac-tory quality. There were no homoplasmic sequence variationsbetween the two cell lines (i.e., the mitochondrial coding se-quence was identical across nearly 28 kb of DNA sequencebetween the two cell lines). These findings suggest an extremelyhigh degree of fidelity being retained by BG01V relative to itsparental cell line (BG01), at least at the mitochondrial DNAsequence level. However, it should be noted that both cell linesdemonstrated identical sequence variations compared with theRCRS, consistent with the presence of coding sequence poly-morphisms in the mitochondrial DNA; coding sequence poly-morphisms for BG01V and BG01 were tabulated (Table 4).
MethylationMSP was employed to monitor epigenetic stability and analyzethe methylation status of six specific genes for both BG01V andNTERA-2. The results are shown in Figure 3. The maternal,methylated allele and paternal, unmethylated allele of thePWS/AS (snrpn) gene locus were detected, indicating preserva-tion of the normal imprinting pattern in BG01V cells. In con-trast, only the paternal, unmethylated allele of the PWS/AS(snrpn) gene was detected in the NTERA-2 sample (Fig. 3A).MSP analysis of DLK1/MEG3 DMR and the H19 locus dem-onstrated the expected maternal, unmethylated allele productand the paternal, methylated allele product, indicating a normalimprint pattern in these regions in both cell lines (Fig. 3B, 3C).Both unmethylated and methylated alleles of the Xist genepromoter region were detected in BG01V and NTERA-2 cells(Fig. 3D). Only the unmethylated homologue of the Notch1promoter region was generated in the analysis of the BG01Vand NTERA-2 DNAs (Fig. 3E). Both unmethylated and meth-ylated alleles of the Oct-3/4 promoter region were detected inBG01V, whereas only the unmethylated allele of the Oct-3/4promoter region was detected in the NTERA-2 sample (Fig. 3F).
Telomerase ActivityGermline and stem cells are known to contain enzymatic activ-ity associated with the maintenance of chromosomal caps thatprevent terminal fusion [28, 29]. Assay of telomerase activitywas employed to validate the presence of hTERT activity inBG01V and NTERA-2 cells. BG01V and NTERA-2 cells testedpositive for telomerase activity (Fig. 4). Heat-inactivated,RNase-treated samples of BG01V, NTERA-2, and IMR-90were included as negative controls. The telomerase-positivecontrol is shown in lane 8.
ImmunophenotypingImmunocytochemistry and alkaline phosphatase activity assayswere used to assess the presence of stem cell markers charac-teristic of the undifferentiated state. BG01V and NTERA-2 cellsstained positively for Oct-3/4, SSEA-4, TRA-1-60, and TRA-1-81. As originally identified in EC cells, the stage-specificembryonic antigen, SSEA-4, and the keratin sulfate-associatedepitopes, TRA-1-60 and TRA-1-81, were found to be expressedin ESCs later on [30, 31]. Both cell lines also demonstratedendogenous alkaline phosphatase activity as evidenced by flu-orescence microscopy (Fig. 5). Phase-contrast and fluorescentimages of the cells showed the expected morphology and stemcell markers previously observed on hESCs [6, 31]. BG01Vcells formed well-defined, compact colonies, whereasNTERA-2 cells formed a diffuse monolayer. Although a quan-titative analysis of the immunophenotyping in situ was notperformed, the majority of BG01V and NTERA-2 coloniesstained positive for all the markers tested. Within the positivecolonies, a large majority of the individual cells stained positive.The parent line BG01 has been described as strongly and uni-formly positive for the markers SSEA-4, TRA-1-60, TRA-1-81,and Oct-3/4 [5], and the same phenotype was observed here forBG01V. The results from quantitative immunophenotyping us-ing flow cytometry were in agreement with the immunocyto-chemical staining (data not shown).
Figure 2. STR profile of the BG01V, BG01, and NTERA-2 cell lines.Loci analyzed include D5S818, D13S317, D7S820, D16S539, vWA,TH01, TP0X, CSF1P0, and sex chromosomal marker Amelogenin; theywere collected on an ABI 310 Genetic Analyzer. Data were analyzedusing Genescan 3.1 and Genotyper 2.0 (Applied BioSystems). STRanalysis of the D16S539 loci for NTERA-2 cells revealed three repet-itive elements for chromosome 16, which is consistent with its predom-inant karyotype. Abbreviation: STR, short tandem repeat.
537Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

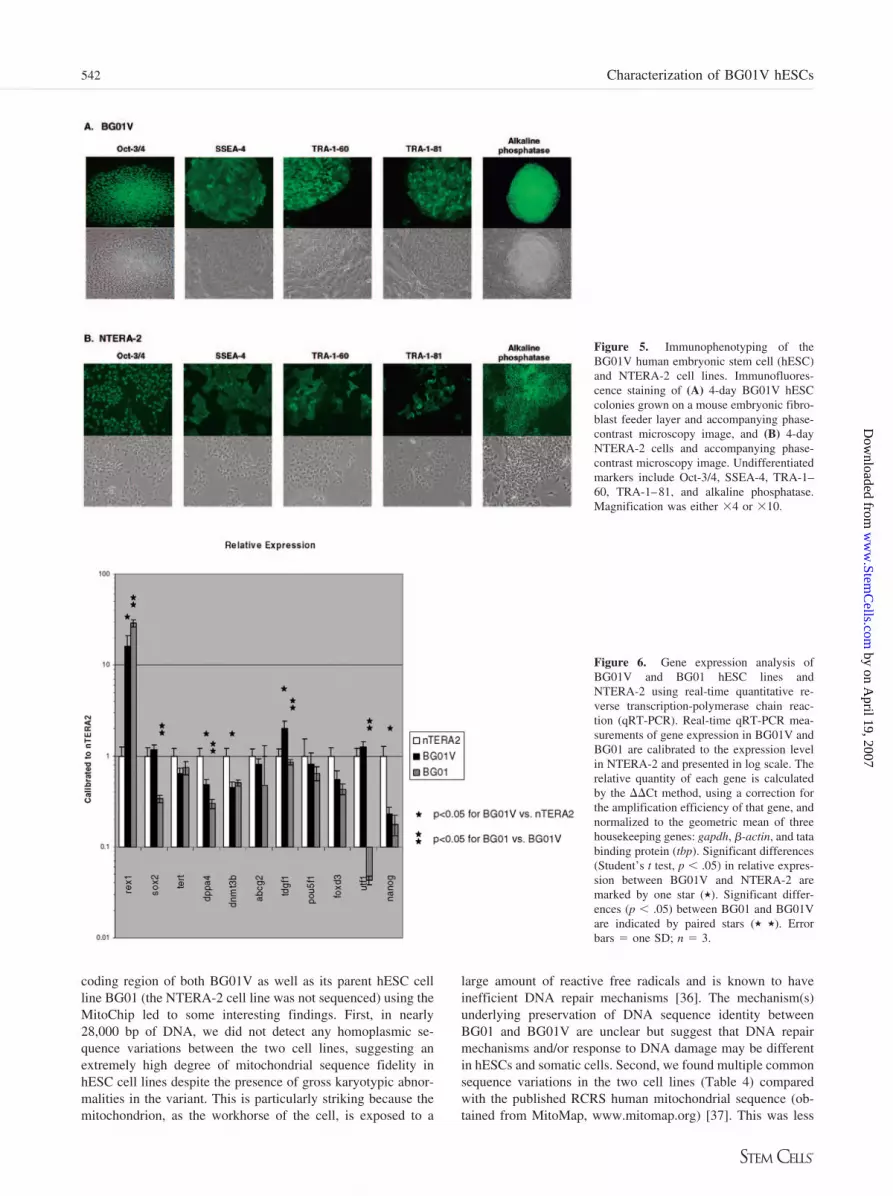
Gene ExpressionSeveral hundred genes were found to be significantly expressed(greater than threefold higher than huURNA) in BG01V orNTERA-2 as assayed by gene expression analysis with microar-rays. The list of significant genes is available as supplementalonline Tables 1 through 3. Genes including POU5F1 (Oct-3/4),DNMT3B, TDGF1 (Cripto), DPPA4 (FLJ10713), REX-1(NM_174900.1), and NANOG (FLJ12581) were significantlyexpressed and in common to both cell lines. Gene ABCG2, anATP-binding cassette subfamily G member 2, was significantlyexpressed in the BG01V hESC line, and a transcription factor
gene SOX2 was significantly expressed in the NTERA-2 cellline only. In addition, germ line-specific genes, including SCP3,VASA, IFITM1 and IFTM2, were not present at significant levelsin either cell line.
Of 264 genes highly expressed in BG01V, 22 (8.3%) were alsohighly expressed in both BG02 and the parental line BG01 [5] (datanot shown). For NTERA-2, 32 (9.6%) of 334 highly expressedgenes were shared with BG01 and BG02. Both of these shared setsinclude the known hESC markers TDGF1, POU5F1, and NANOG.
Confirmation of gene expression for select targets in undif-ferentiated BG01V, the undifferentiated parental line BG01, and
Table 3. HLA typing profile of the BG01V, BG01, and NTERA-2 cell lines
Locus AG1 AG2
A. BG01V HLA profileHLA-A 02XX 03XX
Serological equivalent A2 A3HLA-B 35XX 49XX
Serological equivalent B35 B49HLA-C 04XX 07XX
Serological equivalent Cw4 Cw7DRB1 03VEW 04DAZ
Serological equivalent DR3 DR4Allelic results DRB1*0301/19/22 DRB1*0405/10/28/29/30
DRB3 01ANEXSerological equivalent DR52Allelic results DRB3*0101/02/04/05/11
DQB1 02XX 03XXSerological equivalent DQ2 DQ3
B. BG01 HLA profileHLA-A 02XX 03XX
Serological equivalent A2 A3HLA-B 35XX 49XX
Serological equivalent B35 B49HLA-C 04XX 07XX
Serological equivalent Cw4 Cw7DRB1 03VEW 04DAZ
Serological equivalent DR3 DR4Allelic results DRB1*0301/19/22 DRB1*0405/10/28/29/30
DRB3 01ANEXSerological equivalent DR52Allelic results DRB3*0101/02/04/05/11
DQB1 02XX 03XXSerological equivalent DQ2 DQ3
C. NTERA-2 HLA profileHLA-A 01ADJA
Serological equivalent A1Allelic results A*0101/03/04N/08/09
HLA-B 08AZMJSerological equivalent B8Allelic results B*0801/05/08N/10/11/14/15/18/19N
HLA-C 07XXSerological equivalent Cw7
DRB1 03ASDUSerological equivalent DR3Allelic results DRB1*0301/104/05/06/09/11/13/14/15/16/18/19/22/23/25
DRB3 01ASFCSerological equivalent DR52Allelic results DRB3*0101/02/03/04/05/07/08/10/11
DQB1 02XXSerological equivalent DQ2
HLA class I and class II loci were analyzed using hybridization of polymerase chain reaction (PCR)-amplified DNA with sequencespecific oligonucleotide probes as the primary technology for HLA typing.
538 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from
NTERA-2 was confirmed by quantitative RT-PCR (qRT-PCR)(Fig. 6). Expression of 10 ESC-associated genes was measuredin real-time PCR and normalized to three housekeeping genes.BG01V shows significant differences from NTERA-2 for thegenes REX1, DPPA4, DNMT3B, TDGF1, and NANOG. Inter-estingly, the parental line BG01 shows significant differenceswith BG01V in expression of REX1, SOX2, DPPA4, TDGF1,and UTF1. Unlike the results of microarray analysis, the levelsof ABCG2 and SOX2 were indistinguishable between BG01Vand NTERA-2. However, the genes REX1 and TDGF1 weremore highly expressed in BG01V than NTERA-2 in both qRT-PCR and microarray experiments (supplemental online data). Inaddition, both experiments show that NTERA-2 over expressDPPA4, DNMT3B, and NANOG relative to BG01V.
Comparison of BG01V with EC and OtherESC LinesThe data overall showed that BG01V shared many similaritieswith BG01, and we have previously shown that hESC lines areoverall similar to each other [32]. However, to directly test theoverall degree of similarity of BG01V and NTERA2 to BG01and other hESC lines, we examined global gene expressionusing an Illumina BeadArray, comparing each line to threepooled undifferentiated hESC lines (H1, H7 and H9). Thecorrelation coefficients of the samples are summarized inFigure 7. The overall gene expression of BG01 is highly similarto other undifferentiated hESC lines (r2 � .9559 for genesdetected at confidence �0.99). BG01V is somewhat more di-vergent (r2 � .9206), and NTERA2 has an even greater differ-ence from normal hESC lines (average r2 � .8806). Genesdetected at high confidence are considered differentially ex-pressed if the level is 2.5-fold higher or lower than in the hESCpool. BG01V shows differential expression of 247 genes, verysimilar to the count of 251 differential genes in BG01, whereasNTERA-2 has 904 differentially expressed genes. Detailed anal-ysis of gene expression and statistical validity will be reportedelsewhere.
In the set of genes on the BeadArray, 85 of the 92 candidate“stemness” genes shared among multiple hESC lines could beidentified [32]. Of these 85 stemness genes, 79 were expressedat greater than 99% confidence in BG01V and only four (5.1%)were differentially expressed by greater than 2.5-fold comparedwith pooled hESC lines. NTERA-2 expressed 84 stemnessgenes at greater than 99% confidence, and only five (5.9%) weredifferentially expressed compared with the hESC pool.
Differentiation of BG01V Cells to Ectoderm,Endoderm, and Mesoderm in Complex TeratomasTo examine the differentiation capacity of BG01V cells in vivo,undifferentiated cells were injected intramuscularly to the hindlimb of SCID mice. Tumors formed in all of the five mice thatwere injected; these tumors were dissected for analysis after8–12 weeks. Histological, immunocytochemical, and gene ex-pression analyses demonstrated that the teratomas containeddifferentiated lineages representing ectoderm, endoderm, andmesoderm (Fig. 8). The teratomas were generally well demar-cated from the surrounding muscle and exhibited organizedclusters of cells and primitive tissue structures (Fig. 8A), in-cluding cartilage, mesenchyme, mineralized bone, villi, smoothmuscle, putative nerve bundles, liver-like hepatoid structures,ducts, cystic epithelial-lined spaces and various types of epithe-lia including simple cuboidal, columnar, pseudostratified withcilia and/or goblet cells, simple or stratified with single ormultiple layers of vacuolated cells, and epithelial invaginations.Other cell types that were observed included isolated keratiniz-ing cells and pigmented melanocytes (not shown). Immuno-staining analysis was used to confirm the identification of par-ticular lineages. A few small pockets of OCT-3/4� cells (Fig.8B) indicated the persistence of undifferentiated cells 8–12weeks after injection. Consistent with this, hESC-like cells thatwere OCT-3/4�, SSEA-4�, TRA-1-81�, and SSEA-1 couldbe isolated and expanded from a 12-week-old teratoma (notshown). PC was expressed by endodermal aggregates (Fig. 8C,8D) and epithelia (Fig. 8E, 8H). The basal and superbasal cellsof vacuolated presumptive ectodermal epithelia also expressedp63 (Fig. 8E, 8F), distinguishing it from p63 endodermal-de-rived epithelia (Fig. 8G, 8H). Chondrocytes and clusters ofspindloid cells immediately adjacent to cartilage expressed S100(Fig. 8I, 8J), and smooth muscle was identified by the expres-sion of SMA (Fig. 8K, 8L). Aggregates of neuronal lineagesexpressed NSE and S100 (Fig. 8M, 8N). Individual S100�
neuronal cell bodies are indicated (arrowheads).Quantitative real-time RT-PCR was used to compare gene
expression in undifferentiated BG01V cells and teratomas (Fig.8O). A teratoma sample exhibited reduced expression of mark-ers of pluripotency, including CD9, FOXD3, GDF3, LEFTY2,OCT-3/4, and SOX2, and elevated expression of markers ofectodermal, endodermal, mesodermal, mesendodermal, and tro-phectodermal lineages, including TUBB3, NEF, SOX17,GATA4, BRACHYURY, GSC, MYF5, and EOMESODERMIN.Furthermore, expression of SOX1 (ectoderm), HNF4-�(endoderm), and MYOD (mesoderm) was detected only in theteratoma sample, with crossing points at cycle 13, 6.9, and 9.3,respectively. Therefore, these analyses demonstrated the differ-entiation of BG01V to multiple representatives of each of thethree primary germ layers in vivo.
Table 4. A catalog of human mitochondrial DNA codingsequence polymorphisms seen in BG01V and its parent cell lineBG01
RCRSposition
Referencenucleotide BG01V BG01
750 A G G1438 A G G2706 A G G3197 T C C4769 A G G7028 C T T8860 A G G9477 G A A9548 G A A11467 A G G12309 A A/G A/G13617 T C C14766 C T T14793 A G G15326 A G G
Abbreviation: RCRS, revised Cambridge reference sequence.
539Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

Pathogen TestingPathogen testing was performed by a CLIA-approved labo-ratory (Genetics and IVF Institute), using “nested” multiplexPCR or RT-PCR formats and had a detection threshold of 10copies of target DNA or RNA per assay (with an analyticalsensitivity of 100%), with an LOD of two to five copies perassay. Nucleic acids were extracted from BG01V, and sam-ples were processed for RNA and DNA viral detection asdescribed in Materials and Methods. For all DNA PCR as-says, a human genomic sequence is amplified as an internalpositive control. For RNA viruses (such as HCV), an endog-enous human mRNA is targeted in all assays. These sample-derived targets had to be scored as “detected,” indicatingproper nucleic acid extraction and amplification, prior toscoring any pathogenic target. The list of viruses tested issummarized in supplemental online Table 4. No viral con-tamination was detected in the BG01 or BG01V samples thathave been maintained at ATCC. Thus, detailed pathogentesting as required for any master bank preparation or large-
scale use of cells can be performed from a small amount ofsample using commercially available reagents or validatedclinical laboratories.
DISCUSSIONAlthough it was shown after establishment that the BG01 hESCline is karyotypically normal [4, 5], its continued propagation invitro has given rise to a reported stable variant, BG01V [6, 7],which has been characterized and compared with hESC lines asdescribed herein. We also analyzed various features of thehuman EC cell line NTERA-2 for comparative reasons. Al-though BG01V and NTERA-2 have disparate origins, and bothare cytogenetically abnormal, they display many common cel-lular and molecular traits previously found in pluripotenthESCs. Our goal was to present an initial, in-depth character-ization of the BG01V cell line and propose it as a viable adjunctin stem cell biology research, as well as to propose a standardscheme for characterizing and analyzing hESC lines, therebytyping them for large-scale production.
Figure 3. Methylation-specific polymerasechain reaction analysis of the BG01V humanembryonic stem cell and NTERA-2 celllines. (A): PWS/AS (snrpn): paternal (P)(100 bp) unmethylated allele and maternal(M) (174 bp) methylated allele. (B): DLK1/MEG3 DMR: maternal (M) (120 bp) un-methylated allele and paternal, P (160 bp),methylated allele. (C): H19 promoter: ma-ternal (M) (137 bp) unmethylated allele andthe paternal (P) (136 bp) methylated allele.(D): Xist promoter: unmethylated (U) (176bp) product and methylated (M) (174 bp)product. (E): Notch1 promoter: the unmeth-ylated MSP product (U) (143 bp) and meth-ylated MSP product (M) (140 bp) of Notch1.(F): Oct-3/4 promoter: unmethylated (U)(181 bp) product and methylated (M) (181bp) product. Abbreviations: DMR, differen-tially methylated region; MSP, methylation-specific polymerase chain reaction.
540 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

Frequent morphological analysis, along with routine karyo-typic analysis, of cell lines is critical to assess the genetic andepigenetic stability of hESCs during extended periods in culture.Although both lines examined possess chromosomal aberra-tions, it is reasonable that karyotypic analysis of the varianthESC line should prove less demanding than routine cytogeneticanalysis of the teratocarcinoma cell line because it contains farfewer abnormalities to monitor. In addition, the morphology ofBG01V more closely resembles that of normal hESC lines [4,5], whereas the EC morphology is less defined. Furthermore, asevidenced through 25 passages, BG01V cells maintain karyo-typic stability with the use of enzymatic passaging techniquesrather than mechanical passaging. This also facilitates efficientexpansion of the cells to generate adequate quantities of materialfor research purposes and comparative studies.
The BG01V and NTERA-2 cells both possess trisomy 12.Duplication of the p arm of chromosome 12 has been reportedin male germ cell tumors, including ECs [33]. This region ofchromosome 12 is known to contain genes such as STELLAR,NANOG, and GDF3 that influence proliferation in pluripotentcells [34]. In addition, BG01V cells also carry trisomy 17.Anomalies involving chromosome 17 have been reported inother hESC sublines, including H7 [3]. STAT3 and GRB2 arelocated on chromosome 17; interestingly, the homologues ofthese genes help regulate self-renewal in mouse ESCs. Collec-tively, such gains in gene copy number could afford a selectiveadvantage for propagation in vitro.
As part of the continuing efforts of the Stem Cell Center atATCC and the general international stem cell community to
fully characterize and authenticate hESC lines, we are develop-ing a comprehensive database of DNA profiles based on STRloci. The exploitation of tandemly repeated elements in thegenome has become important in several fields, including ge-netic mapping, linkage analysis, and human identity testing.STR loci are among the most informative polymorphic markersin the genome. The profiling process involves simultaneouslyamplifying eight STR loci and the amelogenin gene for sexdetermination in a multiplex PCR reaction (Promega PowerPlex1.2 system). This test allows for discrimination of at least 1 in108 individuals [35]. STR analysis can be useful in confirmingand clarifying some of the anomalies identified through cyto-genetic analysis. For example, the triple-repeat element forlocus D16S539 correlates with a triosomy 16 or a homozygouspeak for locus D13S317 may indicate a monosomy 13 as seen inthe NTERA-2 cell line. The data from this analysis were enteredinto our database. Such data will help ensure the quality andconsistency of large-scale cell culture expansions and can beperiodically queried as they are maintained. This informationmay become the standard for identifying human embryonic celllines. STR analysis is vital for the correct identification andverification of cells in culture. Furthermore, STR data willensure the quality and integrity of work performed with thesecells.
HLAs are a family of cell proteins found on the surface ofwhite blood cells and other nucleated cells in the body. Theseproteins vary from one person to another and are critical for theactivation of immune responses. HLA typing is used for tissueanalysis before organ and/or cell transplantation. PerformingHLA matching minimizes the possibility of rejection becausetransplantation of tissues and organs between genetically unre-lated people usually results in rejection of the donor graft, tissue,or cell by the recipient. HLA typing is increasingly performedusing DNA techniques. Such typing analyses will be critical asstem cell-based therapies evolve. In addition, the need to collect,compare, and use the typing data over long periods of time willpermit stored sequence polymorphism data to be reinterpreted inthe future as new alleles are discovered and newly derived hESClines are authenticated. The Stem Cell Center at ATCC isestablishing such a database to store HLA typing information.Interestingly, our assessment of the BG01V cell line revealed aheterozygous HLA genotype identical to its parental BG01originator, whereas the profile for the NTERA-2 cell line washomozygous for all HLA alleles examined. The HLA class I (A,B, and C) and class II (DR, DQ, and DP) genes are located onthe short arm of chromosome 6. Although there were two copiesof chromosome 6 identified in the NTERA-2 karyotype, it wouldbe intriguing to determine whether they were fully identical chro-mosomes attributable to a partial meiotic event occurring earlier inits lineage, a loss of heterozygosity, or other complex chromosomalrearrangements originating in the germinal tumor.
The mitochondrion is the only organelle outside of thenucleus to harbor its own DNA. The role of mitochondrial DNAanomalies in a variety of human conditions, including old ageand cancer, has brought to the fore the importance of a sensitiveand accurate, yet high-throughput, platform for sequencing the16.5 kb of mitochondrial DNA. The Human MitoChip wasrecently developed to fulfill this need, and extensive character-ization of this microarray has been described previously [13]. Inthe current study, microarray-based sequencing of the entire
Figure 4. Telomerase detection of the BG01V human embryonic stemcell and NTERA-2 cell lines. Both the BG01V and NTERA-2 cell linesdemonstrate telomerase activity associated with the pluripotent state.Telomerase activity in extracts from 500 cells of BG01V and NTERA-2.Lane 1: 50-bp ladder; lane 2: 123-bp ladder; lane 3: heat-treatedNTERA-2 extract; lane 4: RNase-treated NTERA-2 extracts; lane 5:NTERA-2; lane 6: mechanically scraped BG01V cells; lane 7: flow-sorted SSEA-4� BG01V; lane 8: kit telomerase-positive; lane 9:IMR-90 (telomerase-negative control); lane 10: lysis buffer (primer-dimer control).
541Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from
coding region of both BG01V as well as its parent hESC cellline BG01 (the NTERA-2 cell line was not sequenced) using theMitoChip led to some interesting findings. First, in nearly28,000 bp of DNA, we did not detect any homoplasmic se-quence variations between the two cell lines, suggesting anextremely high degree of mitochondrial sequence fidelity inhESC cell lines despite the presence of gross karyotypic abnor-malities in the variant. This is particularly striking because themitochondrion, as the workhorse of the cell, is exposed to a
large amount of reactive free radicals and is known to haveinefficient DNA repair mechanisms [36]. The mechanism(s)underlying preservation of DNA sequence identity betweenBG01 and BG01V are unclear but suggest that DNA repairmechanisms and/or response to DNA damage may be differentin hESCs and somatic cells. Second, we found multiple commonsequence variations in the two cell lines (Table 4) comparedwith the published RCRS human mitochondrial sequence (ob-tained from MitoMap, www.mitomap.org) [37]. This was less
Figure 5. Immunophenotyping of theBG01V human embryonic stem cell (hESC)and NTERA-2 cell lines. Immunofluores-cence staining of (A) 4-day BG01V hESCcolonies grown on a mouse embryonic fibro-blast feeder layer and accompanying phase-contrast microscopy image, and (B) 4-dayNTERA-2 cells and accompanying phase-contrast microscopy image. Undifferentiatedmarkers include Oct-3/4, SSEA-4, TRA-1–60, TRA-1–81, and alkaline phosphatase.Magnification was either �4 or �10.
Figure 6. Gene expression analysis ofBG01V and BG01 hESC lines andNTERA-2 using real-time quantitative re-verse transcription-polymerase chain reac-tion (qRT-PCR). Real-time qRT-PCR mea-surements of gene expression in BG01V andBG01 are calibrated to the expression levelin NTERA-2 and presented in log scale. Therelative quantity of each gene is calculatedby the ��Ct method, using a correction forthe amplification efficiency of that gene, andnormalized to the geometric mean of threehousekeeping genes: gapdh, �-actin, and tatabinding protein (tbp). Significant differences(Student’s t test, p � .05) in relative expres-sion between BG01V and NTERA-2 aremarked by one star (�). Significant differ-ences (p � .05) between BG01 and BG01Vare indicated by paired stars (� �). Errorbars � one SD; n � 3.
542 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

surprising because there are multiple known mitochondrialDNA haplotypes, and the RCRS is only one common represen-tative haplotype. Interestingly, in a recent study, six heteroplas-mic sequence alterations occurring in two of nine (22%) laterpassage hESC lines were identified and confirmed by conven-tional dideoxy sequencing. Five of these alterations occurred inthe coding region, three resulting in missense mutations in ND1,ND2, and ND4, and one causing a nonsense mutation in AT-Pase6 [38]. Thus, although there are changes in the mitochon-drial genome, these are at a lower frequency than what has beenreported in human cancer cell lines (80–100%) [39]. Neverthe-less, the detailed cataloging of these sequence variations inBG01 and BG01V should serve as a precedent for “mitochon-drial fingerprinting” of additional hESC lines, which (like STRpolymorphisms) can serve as an identity test for researchers whodeal with multiple cell lines in their laboratories.
DNA methylation plays a key role in tissue- and stage-specific gene regulation, genomic imprinting, and X-chromo-some inactivation. It has been demonstrated to be essential fornormal mammalian development [40]. Although much efforthas focused on the mechanism of CpG hypomethylation andhypermethylation of gene-specific promoter regions in tumori-genesis [41], little is known about its role and patterns during thedifferentiation of pluripotent stem cells. Moreover, less is un-derstood concerning the potential epigenetic changes that mayresult throughout extensive in vitro cell culture. Thus, we pro-posed assaying for methylation status via MSP to examineepigenetic stability during cell culture and to indirectly deter-mine the expression status of certain temporally regulated de-velopmental genes important in maintaining the pluripotent stateand initiating the differentiation process.
Three imprinted genetic regions, PWS/AS (Prader-Willi/Angelman) (snrpn), H19, and DLK1/MEG3, were examined formethylation status using MSP. Differential DNA methylationexists at several sites in the PWS/AS (snrpn) critical region suchthat the maternal allele is methylated and the paternal allele isunmethylated and transcriptionally active. In contrast, the dif-
ferentially methylated regions of the H19 promoter region andDLK1/MEG3 are heavily methylated on the paternal homologueand unmethylated on the maternal homologue. MSP analysis ofthese regions demonstrated this to be the case in the BG01VhESCs, thereby implying epigenetic stability of the cells inculture. Although both the unmethylated and methylated allelesof the H19 and DLK1/MEG3 locus were detected in theNTERA-2 cells, only the paternal, unmethylated allele of thesnrpn promoter region was present. Absence of the methylatedand inactive maternal allele in this region indicates a maternaldeletion of the locus through microdeletion or a paternal unipa-rental disomy of chromosome 15. In either case, it is evident thatthe NTERA-2 cell line demonstrated an imprinting anomaly atthis locus.
Both the unmethylated and methylated products for the Xistpromoter region were also detected in the analysis. The Xistgene is expressed exclusively from the inactive X chromosomeand is required for X-chromosome inactivation to occur in earlydevelopment. Interestingly, the inactivation process is randomand can occur on either the paternal or the maternal X chromo-some in females. In certain males who have inherited an extra Xchromosome (i.e., XXY males), one of the X chromosomes isinactivated as in normal XX females. Given the XXY karyotypeof BG01V, it is plausible that unmethylated and methylatedMSP products are generated from one of each of the X chro-mosomes. Furthermore, the presence of both MSP homologuesin the hESCs may be reflective of the cell state and their originfrom the blastocyst. The time during development when mam-mals, including humans, address dosage compensation by inac-tivating one of the X chromosomes begins to initiate just afterthe blastocyst stage. The presence of the additional X chromo-some in BG01V could also promote this hESC line as an in vitromodel to study random X inactivation. Occurrence of bothmethylation products in the teratocarcinoma cell line NTERA-2may be attributed to its abnormal cytogenetic features, includingevidence of a translocation involving the X chromosome,t(Xq1q), and its molecular-level instability in general.
Figure 7. Correlation of overall gene ex-pression among BG01V, NTERA-2, and hu-man embryonic stem cell (hESC) lines byBeadArray. Samples of the karyotypicallyabnormal variant BG01V (A), the normalhESC line BG01 (B), and embryonic carci-noma line NTERA-2 (C) were comparedwith a pool of three normal hESC lines (H1,H7, and H9) by Illumina BeadArray. Corre-lation coefficients and scatter plots were gen-erated using all genes with expression levelmore than 0 (black or blue spots), or allgenes detected at more than 0.99 confidence(blue spots); the correlation coefficientshown (r2) is for the latter group. In eachscatter plot, the central red line representsequivalent expression in the two samples; theouter red lines represent 2.5-fold differencein expression.
543Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

In a pure population of undifferentiated, pluripotent ESCs,the Oct-3/4 gene is highly expressed. This would suggest thatthe promoter is unmethylated. Whereas our MSP analysis de-tected only the unmethylated promoter region of the Oct-3/4promoter in the NTERA-2 cells, both the unmethylated pro-moter and slight evidence of the methylated Oct-3/4 promoterwere visible in the BG01V cells. In a culture of ESCs whichcontains a small percentage of cells differentiating, or beginningto differentiate, we can argue that both hyper- and hypomethy-lated regions of promoter may exist transiently as the cells beginto downregulate expression in committed cell types. Interest-ingly, the robustness of the amplicons in this assay may reflectthe overall status and level of undifferentiation in the cellpopulation and afford a means of quantitative analysis. Theanalysis of the Notch1 promoter generated only the unmethyl-ated allele in both cell lines, indicating a fully unmethylated andtranscriptionally active gene, reflective of its crucial role indetermining cell fate.
It is well known that the cells of the human germline,including ESCs, highly express the catalytic activity of humanTelomere Reverse Transcriptase, or hTERT [24, 42]. Accord-ingly, telomerase activity was detected in BG01V andNTERA-2 cells with similar activity levels based on a qualita-
tive assessment of the data. Likewise, ESCs are known toexhibit elevated levels of alkaline phosphatase activity. This wasevident in both cell lines by the intense staining observed usingthe Enzyme-Labeled Fluorescence (ELF 97) substrate system.
Undifferentiated BG01V cells were immunoreactive forOct-3/4, SSEA-4, TRA-1-60, and TRA-1-81. These pluripo-tency markers were similarly expressed in BG01 and NTERA-2cells by immunocytochemistry. Our assessment of gene expres-sion analysis of the undifferentiated cells using microarray andqRT-PCR revealed that the three cell lines appear similar toeach other with respect to the expression of a select number ofgenes. Using the Agilent array, 28 genes were significantlyexpressed above background and in common between BG01Vand NTERA-2. These included upregulated markers represen-tative of the undifferentiated state, such as POU5F1 (Oct-3/4),Cripto/TDGF1, and NANOG (FLJ12581). Expression data forBG01V were also compared with the previously publishedexpression profiles of the parental line, BG01 and the line BG02[5]; 22 genes were expressed in common between the BG01V,BG01, and BG02 cell lines, including upregulated expression ofPOU5F1, NANOG, and TDGF1 (data not shown). The intersec-tion of BG01V and NTERA-2 was also notable for elevatedlevels of gene expression for DNA methyltransferase DNMT3�,
Figure 8. Differentiation of BG01V to endoderm, ectoderm, and mesoderm in vivo. (A): General structure of a BG01V teratoma exhibitingcomplex-differentiated regions, representative lineages of the three primary germ layers, and a cyst (c). (B): A few small nests of OCT-3/4� cells werepresent in the teratomas, indicating residual pluirpotent cells 8–12 weeks after injection. (C, D): Pancytokeratin� (PC), p63 (not shown)-solidaggregates of endodermal cells. (E): The basal and superbasal cells of vacuolated presumptive ectodermal epithelia-expressed p63 and (F) PC. (G)p63- and (H) PC� endodermal epithelia-containing goblet cells (arrowheads). (I): Cartilage (c), mesenchyme (m), and mineralized bone (b). (J):Chondrocytes expressed S100. (K, L): Smooth muscle was identified by the presence of smooth muscle actin. (M): Neural cell aggregates expressedneuron specific enolase and (N) S100. Individual S100� cell bodies are indicated (arrowheads). (O): Quantitative real-time polymerase chain reactionanalysis of gene expression in BG01V teratomas. The ��Ct method was used to determine relative gene expression, and the ratio of expression inthe teratomas relative to undifferentiated cells is depicted in log scale. The teratoma sample exhibited reduced expression of markers of pluripotencyand increased expression of markers of differentiated lineages, including ectoderm (Ect.), endoderm (End.), mesoderm (Mes.), mesendoderm (M/En),and trophectoderm (Tr).
544 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

developmental pluripotency-associated (DPPA4) (FLJ10713),and a zinc finger protein gene REX-1 (NM_174900.1) and nosignificant expression of markers indicating differentiation.
NTERA-2 shows less similarity to BG01V and BG01 whenglobal gene expression is compared with a pool of undifferen-tiated hESC lines by bead array. The correlation coefficient forall reliably discriminated genes is quite high when BG01 iscompared with the pooled hESC, significantly lower forNTERA-2, and intermediate for BG01V. In addition, the spreadof the data is similar for BG01V and BG01, which both haveabout 250 genes differentially expressed compared with thepool. The number of differentially expressed genes in NTERA-2cells is significantly higher at 904. Thus, gene expression innormal hESCs is modeled better by BG01V than by the EC line.
qRT-PCR analysis was used to confirm and further analyzethe expression of a subset of genes, including POU5F1,DNMT3�, ABCG2, TDGF1, and REX-1. These genes wereupregulated in all cell lines examined, although not always withthe same fold changes as observed using microarray. As manysignificant differences were detected between BG01V andBG01 as between BG01V and NTERA-2. A more in-depthanalysis of the expression profiles of these cell lines usingMPSS (Massively Parallel Signature Sequencing) technology(Lynx Therapeutics, Inc., Hayward, CA, http://www.lynxgen.com) is in progress.
Clusters of BG01V hESC colonies xenotransplanted toSCID mice formed teratomas consisting of cell types derivedfrom ectoderm, endoderm, and mesoderm. Antibodies specificfor human PC, NSE, SMA, p63, Oct-3/4, and S-100 were usedto discriminate the human from mouse cells. Highly differenti-ated cells and tissues derived from all three germ layers, includ-ing identified primitive muscle, cartilage and mesenchyme (me-soderm), ciliated goblet cells (gut endoderm), and neuralepithelia (ectoderm) formed from a majority of teratomas. A fewresidual nests of pluripotent Oct-3/4 cells remained severalweeks after injection. Quantitative real-time PCR demonstrateddownregulation of multiple hESC markers in the tumor (CD9,
Oct-3/4, SOX2, LEFTY2). Upregulation of NEF was detected inthe teratoma, indicating ectoderm differentiation. Multipleendoderm and mesodermal markers were upregulated in theteratomas (i.e., Sox17, brachyury, and GSC).
To establish a baseline of characteristics of BG01V hESCs,we examined karyotype, morphology, STR and HLA profiles,mitochondrial DNA sequence, telomerase activity, methylationstatus, immunocytochemistry, gene expression analysis, andteratoma formation. These characteristics have been retainedthrough 25 passages. In all measures of cell line identity andundifferentiated quality, except karyotype, BG01V is identicalto its parent line BG01. Research models of early embryogen-esis employing variant cell lines such as the BG01V hESC inaddition to, or in lieu of, human EC cells may prove useful andvaluable. Yet despite their independent advantages and disad-vantages, it is likely that both systems will coexist to benefitexperimental stem cell research.
ACKNOWLEDGMENTSWe thank members of ATCC, including Jason D. Dinella forassistance with karyology, Deyun Pan for help with microarrayanalysis, Gregory R. Sykes for performing STR analysis, Wein-ing Xu, Helen K. Josephson, Tahereh Tavakoli for work on cellculture and immunocytochemistry, and Ken Wasserman fortechnical discussion and editorial assistance. In addition, wethank Clifton Baile and Diane Hartzel and the Animal Facility,Animal and Dairy Science Department, University of Georgia,for assisting with generating teratomas. This work was sup-ported by the National Institute on Aging, NIH (N01AG40002).BresaGen, Inc. was supported by NIH grant 9R24RR021313-04(T.S.). The BG01V hESC line was deposited by BresaGen, Inc.with the Stem Cell Center at ATCC and is available for distri-bution as item ATCC SCRC-2002. T.W.P. and R.J. contributedequally to this work.
DISCLOSURESThe authors indicate no potential conflicts of interest.
REFERENCES
1 Thomson JA, Itskovitz-Eldor J, Shapiro SS et al. Embryonic stem celllines derived from human blastocysts. Science 1998;282:1145–1147.
2 Reubinoff BE, Pera MF, Fong C et al. Embryonic stem cell lines fromhuman blastocysts: Somatic differentiation in vitro. Nat Biotechnology2000;18:399–404.
3 Draper JS, Smith K, Gokhale P et al. Recurrent gain of chromosomes 17qand 12 in cultured human embryonic stem cells. Nat Biotechnol 2004;22:53–54.
4 Mitalipova M, Calhoun J, Shin S et al. Human embryonic stem celllines derived from discarded embryos. STEM CELLS 2003;21:521–526.
5 Zeng X, Miura T, Luo Y et al. Properties of pluripotent human embry-onic stem cells BG01 and BG02. STEM CELLS 2004;22:292–312.
6 Zeng X, Chen J, Liu Y et al. BG01V: A variant human embryonic stemcell line which exhibits rapid growth after passaging and reliable dopa-minergic differentiation. Restor Neurol Neurosci 2004;22:421–428.
7 Zeng X, Cai J, Chen J et al. Dopaminergic differentiation of humanembryonic stem cells. STEM CELLS 2004;22:925–940.
8 Andrews PW, Damjanov I, Simon D et al. Pluripotent embryonal carci-noma clones derived from the human teratocarcinoma cell line Tera-2.Differentiation in vivo and in vitro. Lab Invest 1984;50:147–162.
9 Dewji NN, Singer SJ. Cell surface expression of the Alzheimer disease-related presenilin proteins. Proc Natl Acad Sci U S A 1997;94:9926–9931.
10 Andrews PW. Retinoic acid induces neuronal differentiation of a clonedhuman embryonal carcinoma cell line in vitro. Dev Biol 1984;103:285–293.
11 Andrews PW, Nudelman E, Hakomori S et al. Different patterns ofglycolipid antigens are expressed following differentiation of TERA-2human embryonal carcinoma cells induced by retinoic acid, hexameth-ylene bisacetamide (HMBA) or bromodeoxyuridine (BUdR). Differen-tiation 1990;43:131–138.
12 Przyborski SA, Christie VB, Hayman MW et al. Human embryonalcarcinoma stem cells: Models of embryonic development in humans.Stem Cells Dev 2004;13:400–408.
13 Maitra A, Cohen Y, Gillespie SE et al. The Human MitoChip: Ahigh-throughput sequencing microarray for mitochondrial mutation de-tection. Genome Res 2004;14:812–819.
14 Cutler DJ, Zwick ME, Carrasquillo MM et al. High-throughput variationdetection and genotyping using microarrays. Genome Res 2001;11:1913–1925.
15 Kubota T, Das S, Christian SL et al. Methylation-specific PCR simplifiesimprinting analysis. Nat Genet 1997;16:16–17.
545Plaia, Josephson, Liu et al.
www.StemCells.com
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

16 Gao ZH, Suppola S, Liu J et al. Association of H19 promoter methylationwith the expression of H19 and IGF-II genes in adrenocortical tumors.J Clin Endocrinol Metab 2002;87:1170–1176.
17 Poon LL, Leung TN, Lau TK et al. Differential DNA methylationbetween fetus and mother as a strategy for detecting fetal DNA inmaternal plasma. Clin Chem 2002;48:35–41.
18 Murphy SK, Wylie AA, Coveler KJ et al. Epigenetic detection of humanchromosome 14 uniparental disomy. Hum Mutat 2003;22:92–97.
19 Johnston CM, Nesterova TB, Formstone EJ et al. Developmentallyregulated Xist promoter switch mediates initiation of X inactivation. Cell1998;94:809–817.
20 Hendrich BD, Plenge RM, Willard HF. Identification and characteriza-tion of the human XIST gene promoter: Implications for models of Xchromosome inactivation. Nucleic Acids Res 1997;25:2661–2671.
21 Hattori N, Nishino K, Ko YG et al. Epigenetic control of mouse Oct-4gene expression in embryonic stem cells and trophoblast stem cells.J Biol Chem 2004;279:17063–17069.
22 Mitsiadis TA, Gayet O, Zhang N et al. Expression of Deltex1 duringmouse embryogenesis: Comparison with Notch1, 2 and 3 expression.Mech Dev 2001;109:399–403.
23 Li LC, Dahiya R. MethPrimer: Designing primers for methylation PCRs.Bioinformatics 2002;18:1427–1431.
24 Heins N, Englund MC, Sjoblom C et al. Derivation, characterization, anddifferentiation of human embryonic stem cells. STEM CELLS 2004;22:367–376.
25 Vandesompele J, De Preter K, Pattyn F et al. Accurate normalization ofreal-time quantitative RT-PCR data by geometric averaging of multipleinternal control genes. Genome Biol 2002;3:RESEARCH0034.
26 Cheadle C, Vawter MP, Freed WJ et al. Analysis of microarray datausing Z score transformation. J Mol Diagn 2003;5:73–81.
27 Van Gelder RN, von Zastrow ME, Yool A et al. Amplified RNAsynthesized from limited quantities of heterogeneous cDNA. Proc NatlAcad Sci U S A 1990;87:1663–1667.
28 Greider CW, Blackburn EH. Identification of a specific telomere termi-nal transferase activity in Tetrahymena extracts. Cell 1985;43:405–413.
29 Wright WE, Piatyszek MA, Rainey WE et al. Telomerase activity inhuman germline and embryonic tissues and cells. Dev Genet 1996;18:173–179.
30 Draper JS, Pigott C, Thomson JA et al. Surface antigens of humanembryonic stem cells: Changes upon differentiation in culture. J Anat2002;200:249–258.
31 Henderson JK, Draper JS, Baillie HS et al. Preimplantation humanembryos and embryonic stem cells show comparable expression ofstage-specific embryonic antigens. STEM CELLS 2002;20:329–337.
32 Bhattacharya B, Miura T, Brandenberger R et al. Gene expression inhuman embryonic stem cell lines: Unique molecular signature. Blood2004;103:2956–2964.
33 Samaniego F, Rodriguez E, Houldsworth J et al. Cytogenetic and mo-lecular analysis of human male germ cell tumors: Chromosome 12abnormalities and gene amplification. Genes Chromosomes Cancer1990;1:289–300.
34 Clark AT, Rodriguez RT, Bodnar MS et al. Human STELLAR,NANOG, and GDF3 genes are expressed in pluripotent cells and map tochromosome 12p13, a hotspot for teratocarcinoma. STEM CELLS 2004;22:169–179.
35 Lins AM, Micka KA, Sprecher CJ et al. Development and populationstudy of an eight-locus short tandem repeat (STR) multiplex system. JForensic Sci 1998;43:1168–1180.
36 Mambo E, Gao X, Cohen Y et al. Electrophile and oxidant damage ofmitochondrial DNA leading to rapid evolution of homoplasmic muta-tions. Proc Natl Acad Sci U S A 2003;100:1838–1843.
37 MitoMap. Available at: http://www.mitomap.org. Accessed December2004.
38 Maitra A, Arking DE, Shivapurkar N et al. Genomic alterations incultured human embryonic stem cells. Nat Genet 2005;37:1099–1103.
39 Polyak K, Li Y, Zhu H et al. Somatic mutations of the mitochondrialgenome in human colorectal tumours. Nat Genet 1998;20:291–293.
40 Herman JG, Graff JR, Myohanen S et al. Methylation-specific PCR: Anovel PCR assay for methylation status of CpG islands. Proc Natl AcadSci U S A 1996;93:9821–9826.
41 Nuovo GJ, Plaia TW, Belinsky SA et al. In situ detection of thehypermethylation-induced inactivation of the p16 gene as an early eventin oncogenesis. Proc Natl Acad Sci U S A 1999;96:12754–12759.
42 Stojkovic M, Lako M, Strachan T et al. Derivation, growth and appli-cations of human embryonic stem cells. Reproduction 2004;128:259–267.
See www.StemCells.com for supplemental material available online.
546 Characterization of BG01V hESCs
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from

DOI: 10.1634/stemcells.2005-0315 2006;24;531-546; originally published online Nov 17, 2005; Stem Cells
Thomas C. Schulz, Anirban Maitra, Mahendra S. Rao and Jonathan M. Auerbach Toumadje, Sandii N. Brimble, Eric S. Sherrer, Elizabeth W. Uhl, William J. Freed,
Todd W. Plaia, Richard Josephson, Ying Liu, Xianmin Zeng, Carol Ording, Arazdordi Line, BG01V: A Tool for Human Embryonic Stem Cell Research
Characterization of a New NIH-Registered Variant Human Embryonic Stem Cell
This information is current as of April 19, 2007
& ServicesUpdated Information
http://www.StemCells.com/cgi/content/full/24/3/531including high-resolution figures, can be found at:
Supplementary Material http://www.StemCells.com/cgi/content/full/24/3/531/DC1
Supplementary material can be found at:
by on April 19, 2007
ww
w.Stem
Cells.com
Dow
nloaded from





























