Embed Size (px)
Citation preview

~ Pergamon 0197-0186(95)00011-9 Neurochem. Int. Vol. 27, No. 4/5, pp. 301-31 I, 1995
Copyright © 1995 Elsevier Science Ltd Printed in Great Britain. All rights reserved
0197~)186/95 $9.50+0.00
COMMENTARY
CHOLINERGIC A N D PURINERGIC SIGNALLING IN OUTER HAIR CELLS OF MAMMALIAN COCHLEA
E D I T H H E I L B R O N N , L E I F J A R L E B A R K and G R A C E L A W O K O
Department of Neurochemistry and Neurotoxicology, The Arrhenius Laboratories for Natural Sciences, Stockholm University, S-106 91 Stockholm, Sweden
(Received 6 December 1994 ; accepted 26 January 1995)
Abstract--Outer (OHC) and inner (IHC) hair cells in the organ of Corti of the mammalian cochlea process sound. OHC and their efferent synapse are part of a feedback system assumed to control and modulate information carried by afferent neurons passing from IHC to the brain. Underlying mechanisms are not well understood. This paper discusses recent progress. In vivo and in vitro information is presented on structure, pharmacology, function and localization of the pre- and postsynaptic acetylcholine receptors (AChRs) at the efferent synapse. Recent data are given on a presynaptic M3 AChR subtype, probably an autoreceptor involved in transmitter release. Data from our lab on specific binding of [3H]3-quinuclidinyl benzilate ([3H]3-QNB) to non-enzymatically isolated guinea pig OHC reveal a KD several 100 x higher than that for any known muscarinic receptor subtype, including the above-mentioned presynaptic mus- carinic AChR of the OHC efferent synapse. The extremely high concentrations of [3H]3-QNB needed for any binding at all to OHC thus rule out presynaptic membrane impurities as the cause of such binding, and also the presence of a typical mAChR subtype on OHC. The number of [JH]3-QNB binding sites ( ~ 106/OHC) we found on OHC was 1/10th of that we found for binding of nicotinic ligands to OHC, further making it questionable that an ACh-binding site on OHC binds [3H]3-QNB. Observations may instead point to the possibility of another binding site, e.g. an (allosteric) site involved with the as yet not understood 'weak' muscarinic properties of the OHC AChR. Further new data on the OHC AChR confirm reversible ~-bungarotoxin, nicotine and d-tubocurarine binding. [~H]~-Bungarotoxin and [3HI-nicotine binding sites are estimated at ~6" 107 sites/OHC. Strychnine, a glycine receptor blocker suggested to interfere with cholinergic sites of the efferent OHC synapse, was found to bind to OHC (cold strychnine for unspecific binding), This binding, not seen in the presence of high [glycine], increased in the presence of depolarizing [K+], while ACh (100 #M) had no significant effect. Results suggest strychnine binding to the outside of OHC, but also to sites accessible only after cell depolarization, possibly to the hyperpolarizing CaZ+-dependent K ÷ channel. Recent molecular cloning of the OHC AChR indicates a novel c~-subunit. An often observed ACh-activated Ca 2 +-influx close to zero into OHC leaves an unanswered question. OHC also carry P2-purinergic receptors (P2Rs), a more rapid ionotropic P2zR-like subtype and a quantitatively dominating slow metabotropic PzyR subtype coupled to a G protein-phospholipase C cascade and not desensitized, Both contribute to increased cytoplasmic [Ca2+], from respectively external and internal sources. Whether or not such receptors are part of efferent synaptic activity is unknown ; their localization on the OHC plasma membrane is so far only indirect and synaptic vesicles of the efferent nerve endings have not yet been analyzed for their ATP content. Localization, function and interaction of [Ca 2 +] increases triggered by, respectively, ATP and ACh are currently studied in this laboratory.
Two types o f mechanosensory cells are found in the organ of Cort i of the m a m m a l i a n cochlea, the inner hai r cells ( IHC) which t ransfer sound in format ion to the central nervous system (CNS) and the outer hair cells ( O H C ; Fig. 1) which are assumed to be modu- lators of sound informat ion . I H C ( ~ 3500) lie in a single row along the basi lar membrane , whi le O H C ( ~ 2 0 0 0 0 ) lie in three to five rows (Ganong , 1991). Cover ing the rows of H C is the tectorial membrane
into which their stereocilia are embedded. Afferent nerve fibres pass f rom the IHC to the CNS, while O H C synapse, at their basal pole (Ginzberg and Morest , 1983 ; L ibe rman et al., 1990) with 95% of all efferent neurons emana t ing f rom the superior olivary complex in the b ra in stem (Warr et al., 1986 ; Spoend- lin, 1986). Two majo r groups of efferent neurons have been classified according to their postsynapt ic targets in the cochlea: lateral ol ivocochlear (LOC) neurons
301

302 Edith Heilbronn et al.
Fig. I. OHC isolated from the guinea pig cochlea by microdissection (photomicrograph kindly provided by B. Canlon, Department of Physiology and Pharmacology, Laboratory of Experimental Audiology, Karolinska Institute.
Stockholm, Sweden).
which synapse axodendritically with afferent fibers beneath IHC, and medial olivocochlear (MOC) neu- rons which form large and direct synapses with OHC. The functional significance of the LOC efferent path- way is not clear. Several lines of evidence suggest that MOC efferents, by synapsing with OHC, modulate cochlear mechanical responses to peripheral sound stimulation. Presently receptors, neurotransmitters and neuromodulators of this synapse are identified and the consequences of their interactions are analyzed, in our, and in several other, laboratories. Similarly. structure and specificity of the receptors are studied, by techniques from molecular biology, biochemistry, immunochemistry, pharmacology and electro- physiology. Figure 3 summarizes issues raised throughout the text.
OHC-INVOLVING PHYSIOLOGICAL EVENTS AND THEIR UNDERLYING MECHANISMS
Activation of the MOC efferent nerve fibers ori- ginating in the brainstem suppresses the output of the
cochlea, when monitored at the afferent nerve, and further suppresses the mechanical properties of the cochlear partition, as monitored in the distortion product otoacoustic emissions (DPOAEs; Puel and Rebillard, 1990 ; Kujawa el al., 1993,1994). The mech- anism of the suppressive effects is unknown. Efferents have been suggested to decrease the resistance at the base of the OHC, thereby 'shunting' the current through the cell and decreasing its response (Wieder- hold, 1986). OHC's mechanism for tuning and modu- lation of sound information is suggested to involve interaction with IHC, via the tectorial and basilar membranes whereby changes in OHC length are expected to influence the opening of the apical IHC channels at the bases of the stereocilia, thus mod- ulating afferent signalling. Brownell et al. (1985) showed that intracellular current administration to isolated OHC of the guinea pig resulted in either an increase (hyperpolarization) or a decrease (depolar- ization) in OHC length and that the iontophoretic application of acetylcholine (ACh) to the synaptic ends of OHC causes a decrease in their length. Such changes are suggested to be part of the efferent feed- back control mechanism from the CNS (Kelly, 1991) and are known to involve cytoplasmic [Ca 2+ ] increases in OHC. A voltage-dependent Ca 2+ channel (Van den Abbeele et al., 1994) as well as one or two ligand- dependent ionotropic and at least one ligand-depen- dent metabotropic receptor seem to be involved.
THE CHOLINERGIC RECEPTORS AT THE EFFERENT SYNAPSE
Evidence confirms that ACh is the main transmitter (review: see Eybalin, 1993) released by the MOC efferent fibers. The efferent pathway is immuno- reactive to acetyl-CoA:choline O-acetyl transferase (EC 2.3.1.6; CHAT; Altschuler et al., 1985) and to acetylcholinesterase (EC 3.1.1.7: ACHE; Churchill et
a/,, 1956). Bobbin and Konishi (1971) showed that the inhibitory effects of the physiological neuro- transmitter at the auditory synapse were mimicked by ACh and the efferent stimulation was shown to be suppressed by cholinergic antagonists, suggesting blockage ofa cholinergic receptor (Fex, 1968 ; Bobbin and Konishi, 1974). The message, however, was con- fusing, as the literature described effects of both mus- carinic and nicotinic cholinergic ligands 'in the cochlea'. This might be understood by assuming the presence of different, even novel, pre- and post- synaptic acetylcholine receptor (AChR) subtypes at these locations. Current evidence indicates that this is the case : a metabotropic muscarinic AChR (mAChR)

Cochlear efferent synapse :
seems to be present at the efferent presynapse and at least one non- 'classical' nicotinic, ionotropic or mixed ionotropic and metabotropic AChR with a novel structure and unusual properties is located at the post- synapse, i.e. on OHC.
The presynaptic rnuscarinic A C h R subtype
Total RNA, extracted from mouse cochlea, reverse transcribed and amplified by the polymerase chain reaction (PCR) with primers specific for nucleotide sequences m~-m5 (mAChR subtypes) revealed ampli- fication of PCR products corresponding to the sub- types m~, m3 and m5 (Drescher et al., 1992). Corresponding mAChRs may be present in the cochlea. Data in papers on muscarinic ligand effects 'in the cochlea' or at the efferent synapse drew attention to the possibility of a mAChR subtype involvement in ACh release, as is the case at most cholinergic synapses. Using selective destruction techniques, Bar- tolami et al. (1993c) recently found, at the synapse between medial efferent axons and OHC, a carbachol- activated presynaptic M3 AChR subtype, which enhanced inositol phosphate (IP) accumulation and was inhibited by atropine. Destruction of medial chol- inergic axons by sectioning of the crossed olivo- cochlear bundle reduced the carbachol-stimulated IPs by 35% in the organ of Corti. Sites of functional cholinoceptors in the modiolus were also found at spiral ganglion neurons and/or Schwann cells. Related results come from studies on ototoxins, found to inhibit mAChR-activated polyphosphoinositide turn- over in the rat cochlea (Schacht, 1986) and, studying drug effects on [3H]3-QNB-binding to mAChRs in rat cochleas. Bartolami et al. (1994), concluded that the anticancer drug cisplatin and also HgC12, which were found to reduce dose-dependently such binding in the rat cochlea, may interfere with the phosphoinositide signalling pathway (Bartolami et aL, 1993a).
A comparison of our recent data on [3H]3-QNB binding to non-enzymatically isolated guinea pig cochlear OHC (Lawoko et al., 1995 : KD 11.5 + 5 nM, 21°C, Hill coefficient ~ 1 ; number of [3H]3-QNB binding sites/OHC about 10% of that found for the nicotinic ligands) with Bartolami's et al. (1994) data on the efferent presynaptic mAChR from adult rat (KD 30 pM, 25°C) and Whipple and Drescher's (1984) data on guinea pig 'synaptic and other membranes' (KD 30-80 pM, 37°C) rules out remaining presynaptic membrane traces as a cause of observed [3H]3-QNB binding to OHC. In our hands the binding is only barely significant even at concentrations more than two orders of magnitude higher than those considered as valid for known mAChR subtypes. Our results
receptors and signalling 303
therefore lend support to suggestions in the literature about a so far undefined interaction of muscarinic ligands with OHC. The very high concentrations needed for binding and the number of binding sites found, however, do not point to an agonist (ACh) site of a conventional mAChR. A specific and hitherto unrecognized site of the OHC A C h R may exist; at this stage the possibility that this site is allosteric and that muscarinic ligand binding at the OHC AChR may be of pharmacological relevance cannot be excluded.
Functional consequences o f postsynaptic AChR activation
Functional consequences of OHC AChR activation have been studied in vivo and in vitro with preparations of isolated hair cells. Repetitive stimulation of the crossed efferents was found to inhibit auditory nerve activity (cat), increase cochlear microphonics and evoke a slow potential (see Fex and Adams, 1978, and papers cited therein). All three effects were partially blocked, in a slowly reversible way, by 1 #M c~-bun- garotoxin (ct-bgtx). Anniko et al. (1995) found some inhibition of OHC motility when determining in vivo auditory brainstem response (ABR) thresholds after application of ~t-bgtx onto the cochlear round window membrane thereby exposing the cochlea. Minor improvements in ABR thresholds were interpreted as a facilitation of OHC, i.e. a releasing of their efferent inhibitory control. This would be consistent with the presence of an ~-bgtx-sensitive AChR on OHC and with the hypothesis that efferent influence on the coch- lea may be linked with a modulation of the mechanical functions of OHC.
The cholinergic agonists ACh and carbachol, but neither nicotine nor muscarine (Kakehata et al., 1993), were found to activate an outward K ÷ current (Art et al., 1985 ; Ashmore and Ohmori, 1990 ; Shige- moto and Ohmori, 1991; Fuchs and Murrow, 1992a,b). A small Ca: ÷ influx preceded the K ÷ current (Shigemoto and Ohmori, 1990; Fuchs and Murrow, 1992a; Nilles et al., 1994). The depolarizing inward Ca 2 + current was partially blocked by d-tubocurarine (Doi and Ohmori, 1993). Removal of external Ca 2÷ was also shown to abolish the inward current, estab- lishing its Ca :+ dependence (Housley and Ashmore, 1991 ; Fuchs and Murrow, 1992a,b ; Doi and Ohmori, 1993). The Ca 2 ÷-activated K ÷ current, found by Ash- more and Meech (1986) to influence the OHC resting potential, was sensitive to tetraethylammonium (TEA; Housley and Ashmore, 1991) and hyper- polarized the cell (Shigemoto and Ohmori, 199l; Fuchs and Murrow, 1992a,b; Kakehata et al., 1993).
304 Edith Heilbronn et al.
Finally, OHC respond to extracellularly applied ACh with slow motile activity (Brownell et al., 1985; Plin- kerr, 1989). Other data question the involvement of ACh (or GABA) in [Ca =+] increases (Dulon et al.,
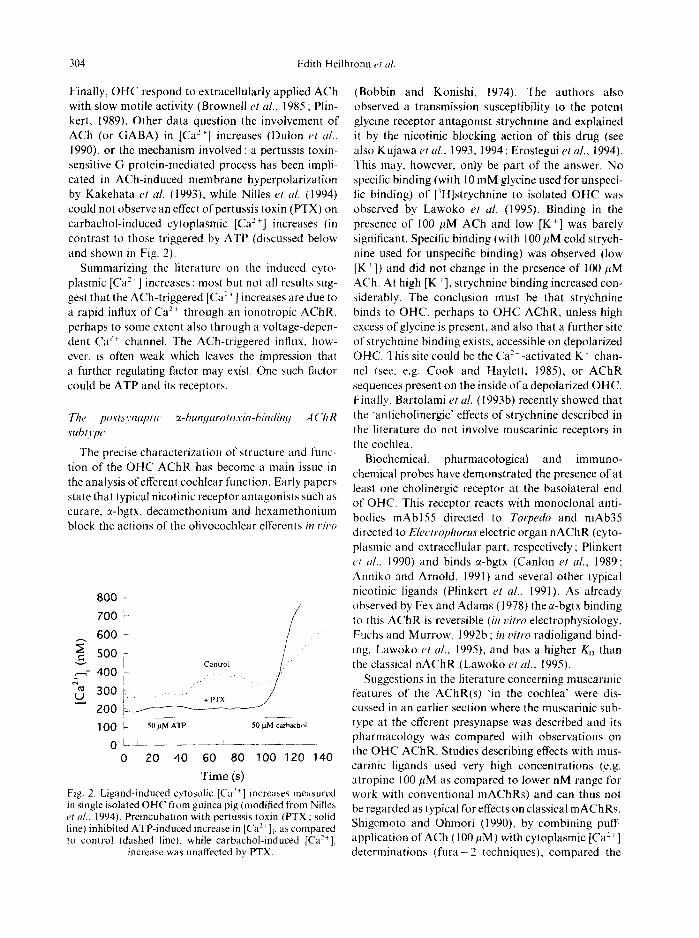
1990), or the mechanism involved : a pertussis toxin- sensitive G protein-mediated process has been impli- cated in ACh-induced membrane hyperpolarization by Kakehata et al. (1993), while Nilles et al. (1994) could not observe an effect ofpertussis toxin (PTX) on carbachol-induced cytoplasmic [Ca 2+] increases (in contrast to those triggered by ATP (discussed below and shown in Fig. 2).
Summarizing the literature on the induced cyto- plasmic (Ca z+] increases : most but not all results sug- gest that the ACh-triggered [Ca ' ' ] increases are due to a rapid influx of Ca 2+ through an ionotropic AChR, perhaps to some extent also through a voltage-depen- dent Ca 2. channel. The ACh-triggered influx, how- ever, is often weak which leaves the impression that a further regulating factor may exist. One such factor could be ATP and its receptors.
The postsvnaptic ~-bungarotoxin-hindinq A ( ' l lR
subtype
The precise characterization of structure and func- tion of the OHC AChR has become a main issue in the analysis of efferent cochlear function. Early papers state that typical nicotinic receptor antagonists such as curare, ~-bgtx, decamethonium and hexamethonium block the actions of the olivocochlear efferents in tqt~o
/ 7O0
600 ?
i / ! C o n t r o l "
,---~- 400
~ 300
200 l O 0 ~ 50 ~ An' 50 UM ~rb~hol
0 20 40 60 80 I00 120 140
Time (s) Fig. 2. Ligand-induced cytosolic [Ca :+] increases measured in single isolated OHC t¥om guinea pig (modified from Nilles el al.. 1994). Preincubation with pertussis toxin (PTX : solid line) inhibited ATP-induced increase in [Ca 2~ ],, as compared to control (dashed line), while carbachol-induced [Ca-~],
increase was unaffected by PTX.
(Bobbin and Konishi, 1974). The authors also observed a transmission susceptibility to the potent glycine receptor antagonist strychnine and explained it by the nicotinic blocking action of this drug (see also Kujawa et al., 1993, 1994 ; Erostegui et al,, 1994). This may, however, only be part of the answer. No specific binding (with l0 mM glycine used for unspeci- fic binding) of [SH]strychnine to isolated OHC was observed by Lawoko et al. (1995). Binding in the presence of 100 #M ACh and low [K +] was barely significant. Specific binding (with 100/~M cold strych- nine used for unspecific binding) was observed (low [K~]) and did not change in the presence of 100/tM ACh. At high [K ~ ], strychnine binding increased con- siderably. The conclusion must be that strychnine binds to OHC, perhaps to OHC AChR, unless high excess ofglycine is present, and also that a further site of" strychnine binding exists, accessible on depolarized OHC. This site could be the Ca2+-activated K + chan- nel (see, e.g. Cook and Haylett, 1985), or AChR sequences present on the inside of a depolarized OHC. Finally, Bartolami et al. (1993b) recently showed that the "anticholinergic' effects of strychnine described in the literature do not involve muscarinic receptors in t he cochlea.
Biochemical, pharmacological and immuno- chemical probes have demonstrated the presence of at least one cholinergic receptor at the basolateral end of OHC. This receptor reacts with monoclonal anti- bodies mAb155 directed to Torpedo and mAb35 directed to Electrophorus electric organ nAChR (cyto- plasmic and extracellular part, respectively; Plinkert et al., 1990) and binds 7-bgtx (Canlon et al., 1989; Anniko and Arnold, 1991) and several other typical nicotinic ligands (Plinkert et al., 1991). As already observed by Fex and Adams (1978) the ~-bgtx binding to this AChR is reversible (in z,itro electrophysiology, Fuchs and Murrow, 1992b; in ~gitro radioligand bind- ing, Lawoko et al., 1995), and has a higher Kf~ than the classical nAChR (Lawoko et al., 1995).
Suggestions in the literature concerning muscarinic features of the AChR(s) qn the cochlea' were dis- cussed in an earlier section where the muscarinic sub- type at the efferent presynapse was described and its pharmacology was compared with observations on the OHC AChR. Studies describing effects with mus- carinic ligands used very high concentrations (e.g. atropine 100 pM as compared to lower nM range for work with conventional mAChRs) and can thus not be regarded as typical for effects on classical mAChRs. Shigemoto and Ohmori (1990), by combining puff'- application of ACh (100 pM) with cytoplasmic [Ca ~+ ] determinations ( f u r a - 2 techniques), compared the

Cochlear efferent synapse : receptors and signalling 305
effects of a general muscarinic (atropine), an M 1 speci- fic (pirenzepine) and a nicotinic (d-tubocurarine, dTC) AChR blocker on the ACh-induced cytoplasmic Ca 2 + levels and found suppression with both atropine and dTC but not with pirenzepine. These experiments, however, may not have compared drug action at pre- cisely the same point in the sequence from receptor activation to cytoplasmic [Ca 2÷] measurements, i.e. it is not self-evident that the drugs all acted as AChR blockers. No competitive studies were performed. Housley and Ashmore (1991) approached the drug concentration question; 50 #M atropine was shown to block ACh-induced outward OHC current, i.e. the compound acted somewhere along the sequence lead- ing from ACh-induced receptor activation to induced cytoplasmic [Ca z+] increases, followed by activation of an outward-directed rapidly desensitizing Ca 2+- dependent K ÷ current that hyperpolarizes the OHC membrane. When comparing lower, equimolar, con- centrations (0.1 #M) of respectively atropine and dTC, only dTC was found to block the OHC response. Thus atropine, at high concentration, may act some- where else along the above described sequence. The largest ACh-induced responses were found near the cell base where the AChR has been demonstrated. Taken together, the above-mentioned results show characteristics of a nicotinic rather than a con- ventional muscarinic response. Another uncertainty concerns the source of the cytoplasmic [Ca 2+] in OHC. Shigemoto and Ohmori (1990, 1991) argue that the ACh-mediated activation of the outward K ÷ current involves Ca 2÷ release from intracellular stores while others (Doi and Ohmori, 1993) claim they were unable to show ACh-induced K ÷ currents in the absence of or at only low concentrations of extracellular Ca 2÷. It must be pointed out that a PTX-sensitive pathway in OHC exists, but our receptor studies (Nilles et al., 1994) indicate that this is not an ACh- but an ATP- activated receptor; more precisely a P2y-purinergic receptor subtype, coupled to a G protein-phos- pholipase C system (see below and Nilles et al., 1994). It is also of interest that Kakehata et al. (1993) pro- posed an ACh-induced mechanism which is both dependent on extracellular Ca 2+ and PTX-sensitive. This interpretation is contradicted by results from Nilles et al. (1994, and Fig. 2) where it is shown that addition of PTX cannot prevent extracellular car- bachol from increasing cytoplasmic [Ca 2÷] but blocks extracellular ATP from doing so. We interpret this as indicating that only the ATP-activated P2R uses a PTX-sensitive G protein-phospholipase C system to release Ca 2÷ from intraceUular, presumably IP3-sen- sitive stores. Data from Housley and Ashmore (1991)
also exclude the involvement of a G protein in the ACh-induced response, as GTP or GTPTS had no effect.
A further point of experimental uncertainty should be mentioned. The cochlear OHC population is not homogenous, a fact that may well contribute to some of the confusion described above. Thus, Doi and Ohmori 0993) found ACh-induced outward currents and membrane hyperpolarization, i.e. the effects of AChR activation in OHC, in only 28% of a guinea pig OHC population studied ; if, however, only OHC of less than 60 #m in length were selected ACh- induced outward currents were seen in 38% of cells studied. The short OHC are found in the basal region of the cochlea and in the medial row. Other trans- mitters, including ?-aminobutyric acid (GABA), have been proposed to act at the OHC efferent synapse, probably at OHCs from other rows of the cochlea. Whether or not these transmitters are relevant for all or only for some of the MOC efferents is not yet understood.
Currently, several laboratories are cloning the OHC AChR. An ct5 subunit has been suggested (Housley et al., 1994) to be part of a cochlear nAChR ; however, recently at least two laboratories (Heinemann's and our own) agreed that a novel ~-subunit is part of an ionotropic, largely nicotinic AChR which has novel pharmacological features. Table 1 compares results from studies on OHC with studies on 'cochlea', not unexpectedly confirming the probable presence of several AChRs in cochlea and/or in different species. Elgoyhen et al. (1994) reported the isolation and func- tional characterization of ct9, from olfactory epi- thelium. Amplification (PCR) of rat cochlear cDNA using ct9 primers suggests that this subunit gene is ex- pressed in the inner ear. Heterologous expression of the cloned cDNA in Xenopus oocytes suggests a homooligomeric receptor channel complex gated by ACh which has a mixed, but mainly nicotinic, phar- macology resembling that of OHC. Further detailed information and sequence data on this new subunit are still lacking.
In summary, the picture obtained from in vitro work with the various techniques reveals a highly protease- sensitive, unconventional AChR of (uniform or mixed?) ionotropic nature with a primarily nicotinic ligand specificity. The locations for muscarinic bind- ing at very high ligand concentrations and for strych- nine binding remain to be further analyzed. At the time this is being written, the final cloning is about to be completed. The result will surely clarify some of the unsolved problems concerning the mammalian OHC AChR.

306 Edith Heilbronn el a/.
Table I. Acetylcholine receptor subuuits detected in inner ear tissue from different species
Species/tissue Subunit(s) detected Not found Method References
Rat cochlea ~5, [34 ~2, :d, :~4, [32 PCR analysis I Rat cochlea (OHC region) (in spiral ganglion) ~2, :~3. zt4,/~2 In situ hybridization 2 Chick cochlea ~7 71, ~3, :~8, [¢2 Radioimmunoassay 3
:~7 PCR analysis Mouse cochlca :~2. ~4, :~5, [32 ~3, :~7,/J4 PCR analysis 4
([:q, c) (cd."/, 6) PCR analysis (weak amplification) Rat cochlea :~9 PCR analysis 5 Guinea pig, isolated OHCs x7 PCR analysis 6
References I : Housley et al., 1994: 2: Wackym et al., 1992: 3: Anand el al.. 1993; 4: Khan et al., 1994; 5: Elgoyhen et al,, 1994; 6 Jgrlebark et al., unpublished.
P2-PURINERGIC RECEPTORS ON O H C AND
FUNCTIONAL CONSEQUENCES OF THEIR ACTIVATION
The physiological roles of P2-purinergic receptors (P2Rs), present in many neural and non-neural systems, have been discussed for several years. In a few cases ionotropic receptors gated by extracellular ATP were shown to be responsible for rapid neuro- transmission (Edwards et al., 1992 ; Evans et al., 1992 : for subunit structure of ionotropic P2~R, see Brake et
al., 1994; Valera et al., 1994). More often, however, ATP acts extracellularly as a cotransmitter or modu- lator at P2Rs (Burnstock, 1990: Dubyak and EI- Moatassim, 1993) and activates G protein-coupled PzyR and P2uR subtypes~ which have also recently been cloned (Lustig et al., 1993 ; Webb et al., 1993 ; Parr et
al., 1994). There is evidence for at least two subtype P2Rs on OHC. The primary activity is that ofa meta- botropic P2y-purinergic subtype (agonist pattern: ATP~S > ATP = 2-methyl-S-ATP > ADP > :t,fi- methylene ATP: no activation by ADPflF or UTP) coupled to a pertussis toxin-sensitive G protein-phos- pholipase C cascade. This receptor does not desen- sitize (Heilbronn, 1993 ; Heilbronn et al., 1994; Nilles et al., 1994). An ionotropic P2R subtype of as yet unknown structure contributes marginally. Ashmore and Ohmori (1990) showed that ATP activates an inward whole cell current at resting potentials of - 60 mV and that the internal [Ca ~ ] increased during this current; current-clamped cells showed ATP-induced depolarization. This weak ATP-induced depolar- ization of the OHC membrane may further cause a small Ca 2+ influx through voltage-gated Ca 2+ chan- nels (Ashmore and Ohmori, 1990 ; Nilles et al., 1994). Ashmore and Ohmori applied ATP (5-100 /tM) to isolated and fura-2 loaded guinea pig OHC and observed two components, a rapid rise in cytoplasmic [Ca2+], peaking at 20 s and localized around the apical (endolymphatic) end of the cell, and a slower rise, peaking between 1-3 min and spread throughout the whole cell. Nilles et al. found that influx does not occur
in the absence of external Ca 2~ (Ca 2 ~-free medium. EGTA) but ATP still triggers a slow-starting rise in cytoplasmic [Ca -,+ ] without any obvious desen- sitization. This Ca 2+ is released from intracellular (most probably IP~-sensitive) sources. Nilles et al.
(1994) also found that even in the presence of extra- cellular Ca 2+, ACh (100 /~M) produced very little [Ca "-+] increase and never from intracellular sources. According to Ashmore and Ohmori (1990) ACh and ATP counteract each other; the authors suggest a model in which ACh would act as a competitive anta- gonist.
The origin of extracellular ATP in the fluids around OHC is not clear. If OHC carry P2R subtypes in the efferent synapse, the synaptic vesicles that are present in the presynaptic terminal might release it, perhaps together with ACh, but the vesicle content has not been analyzed. ATP might also be released from a depolarized postsynapse, and could be a signal to improve postsynaptic [Ca '+] regulation. So far the only observed function of the described P2R subtypes, however, is to increase cytoplasmic [Ca2+], a function of primarily the metabotropic P2R. Thus both the P2R and AChR subtypes on OHC are concerned with the control of cytoplasmic Ca 2+ levels, but may do so at different revels of OHC or, as Ashmore and Ohmori suggest, by counteracting each other.
THE SOURCE OF CYTOPLASMIC Ca ~+ INCREASES IN
OHC
Certain processes in the mammalian cochlea are believed to be related to OHC contractility (Brownell et al., 1985; Ashmore, 1987; Zenner et al., 1987; Hudspeth, 1989). Three types of motility have been described, all assumed to be Ca2+-dependent processes. Two sources exist for cytosolic [Ca 2+] increases, Ca 2+ influx from the extracellular space across the plasma membrane and release from intra- cellular storage places. Extracellular Ca 2+ enters

Cochlear efferent synapse :
through voltage-dependent Ca 2+ channels (patch clamp, Nakagawa et al., 1990; fluorescence imaging microscopy, Dulon et al., 1989; Ikeda et al., 1991) and via the activated AChR. Ca 2÷ is released from intraceUular storage sites, identified in guinea pigs by Ikeda and Takasaka (1993) using confocal laser microscopical imaging. Membrane-bound Ca 2+ was demonstrated in the subcuticular region, the infra- nuclear region and the region adjacent to the lateral wall ; in the central part of the cytoplasm and nucleus only free Ca 2+ was found, while the cuticular plate was free of both. Fluorescent clusters of mitochondria and endoplasmic reticulum were seen in the infra- cuticular and infranuclear regions, some associated with the lateral wall; these cytosolic organelles are assumed to sequester Ca 2+. The subsurface cistern was shown to be analogous to the endoplasmic reticu- lure in biological function and may be the source of Ca 2+ for the actin-mediated process in OHC. This may be a site carrying IP3-sensitive receptors thus allowing Ca 2+ release. In several experiments intra- cellularly applied IP3 was shown to increase outward directed K + currents in OHC, a proof for achieved increases in cytoplasmic [Ca2+].
DO ATP- AND ACH-ACTIVATED RECEPTORS INTERACT WHEN INCREASING CYTOPLASMIC [Ca~+]?
The OHC of mammalian cochlea are functionally polarized. As stated above, Housley et al. (1992) sug- gest that P2R and AChR actions counter each other in guinea pig OHC. The AChRs are at the basal (peri- lymphatic) region of the cells and activate a rapidly desensitizing hyperpolarizing outward-directed K + current. One type of P2R has been suggested to be closer to the apical (endolymphatic) region of OHC where it activates a depolarizing nonselective cation current which exhibits inward rectification and lacks desensitization, The largest ATP currents were found when ATP was directed toward the apical surface of the cells and these were antagonized by amiloride in a dose-dependent way. Using whole cell patch-clamp this receptor was found along the length of isolated OHC. Housley et al. (1992) suggest that ATP may mediate a humoral modulation of the mechano-elec- trical transduction process. OHC carry several types of neurotransmitter or modulator receptors which are respectively ionotropic (AChR and PzzR, both with properties of a non-selective cation channel) and metabotropic (PzyR). These receptors were found to participate to increase cytoplasmic [Ca2+]. Nilles et al. (1994) showed the main cytoplasmic Ca 2 + increase to occur after activation of the metabotropic excit-
receptors and signalling 307
atory P2yR subtype which was pharmacologically identified using several ATP derivatives. Activation of this P2yR may be suggested to increase intracellular IP3 levels via the G protein-phospholipase C cascade which would in turn result in increases in cytoplasmic [Ca 2+] thereby allowing increases in OHC motility (Schacht and Zenner, 1987). Niedzielski and Schacht (1992) showed that application of extracellular ATP to the organ of Corti increased inositol phosphate formation.
Interaction between ATP and the nAChR channel has been suggested but not yet investigated in OHC. On the other hand, both PzR and AChR activation on OHC results in increased cytoplasmic [CaZ+]. We have shown in another system (nAChR and P2R- carrying cultured chick myotubes, H~iggblad et al., 1985) that ATP- or carbachol-triggered 86Rb influx is additive; this may also be the case in OHC. On the other hand, Nakazawa (1994) discussed that a sub- population of nicotinic receptor channels might be activated even by ATP thus modulating the sensitivity to these transmitters by regulating the numbers of ATP-sensitive and non-sensitive channels. Nakazawa thus suggests that the activation by cotransmitters, of a common population of channels, may provide a modulation of neurotransmisson at the postsynaptic membrane. In the OHC system the slowness observed by us when Ca 2+ influx was measured, may speak for such a possibility. Nevertheless, we are currently studying the possibility that the cytoplasmic [Ca :+ ] increases triggered by activation of respectively AChR or P2yR activation may serve different purposes in OHC. This interpretation builds on the fact that cyto- plasmic Ca 2+ has a mobility that is very different from that of IP3. Allbritton et al. (1992) measured the diffusion coefficients (D) for IP3 and Ca 2+ in a cyto- solic extract from Xenopus oocytes. They found 283 #mZ/s for [3H]IP3 while D for Ca 2 + was between 13 and 65 /~m2/s, dependent on the free Ca 2+ concentration (physiological range). Thus there should be two fac- tors that are responsible for transient cytoplasmic [Ca z +] increases and the processes that are influenced by them : the nature and localization of the activated receptor and the origin of the cytoplasmic Ca 2+ increase.
COMMENTS AND UNSOLVED QUESTIONS
The outer hair cells of the mammalian cochlea are of crucial importance to the hearing process ; the precise analysis of their structure and function has, however, progressed slowly. Recent increased efforts have resulted in a better insight, yet much uncertainty
308
Stereocilia
Mechanosensitive i Ca 2+ c h a n n ~ . ~
Subsurface cisterna¢
Edith Heilbronn el el/.
i ~ OUTER HAIR CELL
Ca2+
2a 2+ buffering
Cuticular plate
ipa~?FMetabotropic P2yR, location? ATP
-sensitive G protein LC?
" ~ i S t i m u l u s
Ionotropic P2R *" v P2zR?
CI-
MEDIAL OLIVOCOCHLEAR NERVE ENDING
C e l l u l a r r e s p o n s e
Ca2+-activated k..._ CI- channel
Ca2+ "Novel" 41
Ca2+-activated K + channel
K +
Strychnine inhibition?
EFFERENTSYNAPSE
ACh ATP? Peptides
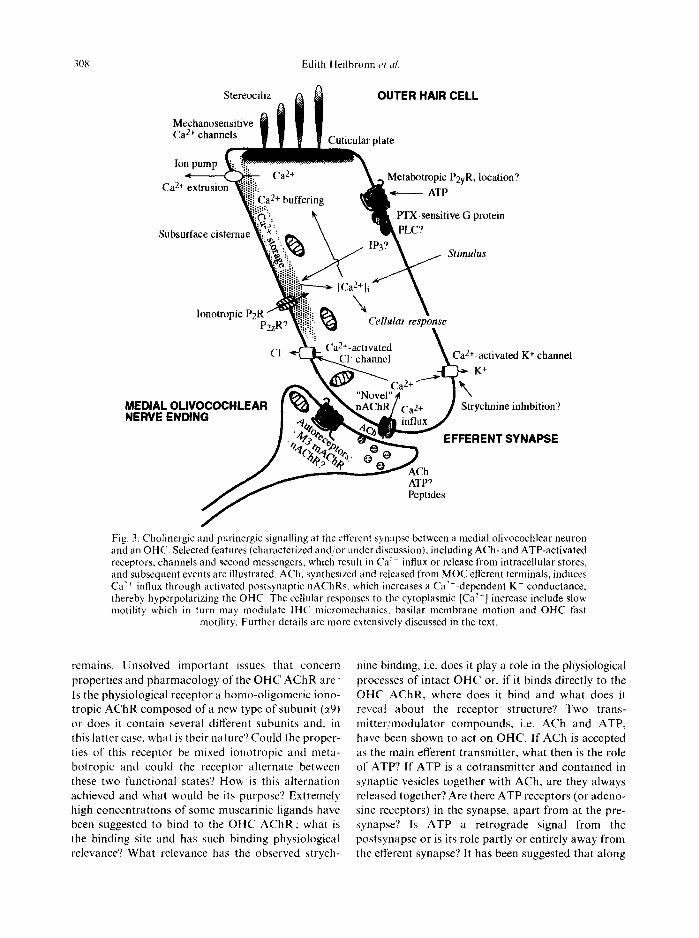
Fig. 3. Cholinergic and purinergic signalling at the efferent synapse between a medial olivocochlear neuron and an OHC. Selected features (characterized and/or under discussion), including ACh- and ATP-activated receptors, channels and second messengers, which result in Ca:~ influx or release from intracellular stores, and subsequent events are illustrated. ACh, synthesized and released from MOC efferent terminals, induces Ca -~+ influx through activated postsynaptic nAChRs, which increases a Ca-'+-dependent K + conductance, thereby hyperpolarizing the OHC. The cellular responses to the cytoplasmic [Ca 2+] increase include slow motility which in turn may modulate IHC micromechanics, basilar membrane motion and OHC fast
motility. Further details are more extensively discussed in the text.
remains. Unsolved important issues that concern properties and pharmacology of the O H C A C h R are: Is the physiological receptor a homo-oligomeric iono- tropic A C h R composed of a new type of subunit (~9) or does it contain several different subunits and, in this latter case, what is their nature'? Could the proper- ties of this receptor be mixed ionotropic and meta- botropic and could the receptor alternate between these two functional states? How is this alternation achieved and what would be its purpose'? Extremely high concentrations of some muscarinic ligands have been suggested to bind to the O H C A C h R : what is the binding site and has such binding physiological relevance? What relevance has the observed strych-
nine binding, i.e. does it play a role in the physiological processes of intact O H C or, if it binds directly to the O H C AChR, where does it bind and what does it reveal about the receptor structure? Two trans- mit ter /modulator compounds, i.e. ACh and ATP, have been shown to act on OHC. If ACh is accepted as the main efferent transmitter, what then is the role of ATP? If ATP is a cotransmitter and contained in synaptic vesicles together with ACh, are they always released together? Are there ATP receptors (or adeno- sine receptors) in the synapse, apart from at the pre- synapse? Is ATP a retrograde signal from the postsynapse or is its role partly or entirely away from the efferent synapse'? It has been suggested that along

Cochlear efferent synapse : receptors and signalling 309
the O H C m e m b r a n e it may counterac t ACh. The role of peptides (e.g. calci tonin gene-related peptide, dynorph ins and enkephal ins) in cochlear efferents and elsewhere on O H C requires elucidation. Similarly, the factors control l ing processes involving Ca 2+, i.e. the A C h R channel open t ime al lowing Ca 2÷ influx and the mechan ims involving the cytoplasmic [Ca 2+] tha t act ivates the outward-di rec ted Ca2÷-activated K + current tha t hyperpolar izes O H C need to be clarified.
Advanced insight into audi tory funct ion at the molecular level will enable pharmacologica l and molecular genetic modu la t ion o f disease states.
Note added in proof--Since this manuscript was submitted, a paper by Elgoyhen et al. (1994) has been published (Cell 79, 705-715). It describes in detail the PCR amplification of rat cochlear cDNA using ~9 primers. In situ hybridization showed ~9 gene expression exclusively over OHC and IHC, but expression in support cells could not be excluded. The primary structure of the ~9 subunit is homologous to the other cloned muscle and neuronal nAChR ct subunits, but has lower amino acid identity. The conserved amino acid residues Phe t°5 and Lys ~5° differ in the ~9 subunit and as these residues are close to the putative ACh-binding site, the switch from hydrophobic and positively charged, respectively, to polar (Ser ~°~) and non-charged (Thr ~5°) may contribute to the unconventional pharmacological properties. Heterologous expression of the ~9 subunit in Xenopus oocytes suggests a homo-oligomeric receptor channel complex gated by ACh which has a mixed nicotinic-muscarinic pharmacology resembling that of OHC. Electrophysiological data and amino acid sequence favor ionotropic (nicotinic) rather than metabotropic (muscarinic) classification of the ~9 receptor, thus confirming our data from 1991 and 1995.
REFERENCES
Allbritton N. L., Meyer T. and Stryer L. (1992) Range of messenger action of calcium ion and inositol 1,4,5-tris- phosphate. Science 258, 1812-1815.
Altschuler R. A., Kachar B., Rubio J. A., Parakkal M. A. and Fex J. (1985) Immunocytochemical localization of choline acetyltransferase-like immunoreactivity in the guinea pig cochlea. Brain Res. 338, 1-11.
Anand R., Fuchs P. A., Cooper J. F., Peng X. and Lindstrom J. (1993) Evidence that ~bungarotoxin-sensitive acetyl- choline receptors in chick's cochlea contain ~7 subunits. Soc. Neurosci. Abstr. 19, 465. (abstract).
Anniko M. and Arnold W. (1991) Acetylcholine receptor localization in human adult cochlear and vestibular hair cells. Acta Otolarynyol. (Stockh.) 111,491 499.
Anniko M., Stenqvist M., Pettersson A. and Heilbronn E. (1995) Pharmacological inhibition of outer hair cell motility in situ. Acta Otolaryn9ol. (Stockh.) 57, 105-109.
Art J. J., Crawford A. C., Fettiplace R. and Fuchs P. A. (1985) Efferent modulation of hair cell tuning in the coch- lea of the turtle. J. Physiol. (Lond.), 360, 397-421.
Ashmore J. F. (1987) A fast motile response in guinea-pig outer hair cells : the cellular basis of cochlear amplifier. J. Physiol. (Lond ) 388, 323-347.
Ashmore J. F. and Meech R. W. (1986) Ionic basis of mem-
brane potential in outer hair cells of guinea pig cochlea. Nature 322, 368-371.
Ashmore J. F. and Ohmori H. (1990) Control ofintracellular calcium by ATP in isolated outer hair cells of guinea pig cochlea. J. Physiol. (Lond.) 428, 109-131.
Bartolami S., Planche M. and Pujol R. (1993a) Inhibition of the carbachol-evoked synthesis of inositol phosphates by ototoxic drugs in the rat cochlea. Hear. Res. 67, 203-210.
Bartolami S., Planche M. and Pujol R. (1994) Effects of ototoxins on quinuclidinyl benzilate binding on the rat cochlea. Neurosci. Lett. 174, 169-172.
Bartolami S., Ripoll C. and Eybalin M. (1993b) Anti- cholinergic effects of strychnine in the cochlea do not involve muscarinic receptors. Neuroreport, 4, 1003-1006.
Bartolami S., Ripoll C., Planche M. and Pujol R, (1993c) Localisation of functional muscarinic receptors in the rat cochlea: evidence for efferent presynaptic autoreceptors. Brain Res. 626, 200-209.
Bobbin R. P. and Konishi T. (1971) Acetylcholine mimics crossed olivocochlear bundle stimulation. Nature 231, 222-223.
Bobbin R. P. and Konishi T. (1974) Action of cholinergic and anticholinergic drugs at the crossed olivo-cochlear bundle-hair cell junction. Acta OtolarynyoL (Stockh.) 77, 56-65.
Brake A. J., Wagenbach J. and Julius D. (1994) New struc- tural motif for ligand-gated ion channels defined by an ionotropic ATP receptor. Nature 371, 519-523.
Brownell W. E., Bader C. R., Bertrand D. and Ribaupierre Y. D. (1985) Evoked mechanical responses of isolated cochlear outer hair cells. Science 227, 194-196.
Burnstock G. (1990) Overview. Purinergic mechanisms. Ann. N. Y. Acad. Sci. 603, 1--18.
Canlon B., Cartaud J. and Changeux J. P. (1989) Local- ization of alpha bungarotoxin binding sites on outer hair cells from the guinea-pig cochlea. Acta Physiol. Seand. 137, 549 550.
Churchill J. A., Schuknecht H. F. and Doran R. (1956) Acetylcholinesterase activity in the cochlea. Laryngoscope 66, 1-15.
Cook N. S. and Haylett D. G. (1985) Effects of apamin, quinine and neuromuscular blockers on calcium-activated potassium channels in guinea-pig hepatocytes. J. Physiol. (Lond.) 358, 373-394.
Doi T. and Ohmori H. (1993) Acetylcholine increases intra- cellular Ca 2+ concentration and hyperpolarizes the guinea- pig outer hair cell. Hear. Res. 67, 179-188.
Drescher D. G., Upadhyay S., Wilcox E. and Fex J. (1992) Analysis of muscarinic receptor subtypes in the mouse cochlea by means of the polymerase chain reaction. J, Neurochem. 59, 765-767.
Dubyak G. R. and E1-Moatassim C. (1993) Signal trans- duction via P2-purinergic receptors for extracellular ATP and other nucleotides. Am. J. Physiol. 265, C577-606.
Dulon D., Zajic G. and Schacht J. (1990) Increasing intra- cellular free calcium induces circumferential contractions in isolated cochlear outer hair cells. J, NeuroscL 10, 1388 - 1397.
Dulon D., Zajic G., Aran J. M. and Schacht J. (1989) Amino- glycosides antibiotics impair calcium entry but not viability and motility in isolated cochlear hair cells. J. Neurosci. Res. 24, 338-346.
Edwards F. A., Gibb A. J. and Colquhoun D. (1992) ATP receptor-mediated synaptic currents in the central nervous system. Nature 371, 144-147.

310 Edith Heilbronn et al.
Elgoyhen A. B., Johnson D. S., Boulter J., Vetter D. E. and Heinemann S. (1994) Alpha 9: a new acetylcholine receptor with novel pharmacological properties (Abstract S.21). International Symposium on The Cholinergic Syn- apse : Structure, Function and Regulation. Baltimore, Md.
Erostegui C., Norris C. H. and Bobbin R. P. (1994) It, ritro pharmacologic characterization of a cholinergic receptor on outer hair cells. Hear. Res. 74, 135 147.
Evans R. J., Derkach V. and Surprenant A. (1992) ATP mediates fast synaptic transmission in mammal ian neurons. Nature 357, 503 505.
Eybalin M. (1993) Neurotransmittors and neuromodulators of the mammal ian cochlea. Physiol. Rev. 73, 309 372.
Fex J. (1968) Efferent inhibition in the cochlea by the olivo- cochlear bundle. In: Hearing Mechanisms in Vertebrates (De Reuck A. V. S. and Knight J., eds), pp. 169 181. Ciba Foundation Symposium, Churchill, London.
Fex J. and Adams J. C. (1978) :e-Bungarotoxin blocks rever- sibly cholinergic inhibition in the cochlea. Bra#l Res. 159, 440-444.
Fuchs P. A. and Murrow B. W. (1992a) Cholinergic inhi- bition of short (outer) hair cells of the chick's cochlea..I. Neurosci. 12, 800 809.
Fuchs P. A. and Murrow B. W. (1992b) A novel cholinergic receptor mediates inhibition of chick cochlear hair cells. Proe. R. Soc. Lond. (Biol.) 248, 35 40.
Ganong W. F., (1991) Hearing and equilibrium. In: Review ~?/ Medical Physiology, pp. 137 149. Prentice Hall, New York.
Ginzberg R. D. and Morest D. K. (1983) A study of cochlear innervation in the young cat with the Golgi method. Heat'. Res. 10, 227 246.
H~iggblad J., Eriksson H. and Heilbronn E. (1985) ATP- induced cation influx in myot ubes is additive to cholinergic agonist action. Acta Physiol. &'and 125, 389 393.
Heilbronn E. (1993) Extracellular ATP, acetylcholine (ACh) and the outer hair cells of the mammal ian inner ear. .I . Neurochem. Suppl. 61, S149C (Abstract).
Heilbronn E.. J'arlebark L. and Nilles R. (1994) Role of extracellular ATP in the control o1" cytoplasmic [Ca -̀ + ] of outer hair cells of the mammal ian cochlea. 5th Inter- national Adenosine Symposium Abstracts (#1287). Drt(q. Def. Res. 31,277 (Abstract).
Housley G. D. and Ashmore J. F. (I 991) Direct measurement of the action of acetylcholine on isolated outer hair cells of the guinea pig cochlea. Proc. R. Soc. Lond. (Biol.) 244, 161 167.
Housley G. D.. Greenwood D. and Ashmore J. F. (1992) Localisation of cholinergic and purinergic receptors on outer hair cells isolated from the guinea-pig cochlea. Pro(. R. So('. Lomt. (Biol.) 249, 265 273.
Housley G. D., Batcher S., Kraft M. and Ryan A. E. (1994) Nicotinic acetylcholine receptor subunits expressed in rat cochlea detected by the polymerase chain reaction. Heat'. Res. 75, 4 7 53.
Hudspeth A. J. (1989) How the ear's works work. Nature 341,397 404.
lkeda K. and Takasaka T. (1993) Confocal laser micro- scopical images of calcium distribution and intracellular organelles in the outer hair cell isolated from the guinea pig cochlea. Heat'. Res. 66, 169 176
Ikeda K., Saito Y., Nishiyama A. and Takasaka T. (1991) Effects of pH on intracellular calcium level in isolated cochlear outer hair cells of the guinea pig. Am. J. Physiol. 261, C231 C236.
Kakehata S., Nakagawa T., Takasaka T. and Akaike N. (1993) Cellular mechanism of acetylcholine-induced response in dissociated outer hair cells of guinea-pig coch- lea. J. Physiol. (Lond.) 463, 227 244.
Kelly J. P. ( 1991 ) Hearing : Vibrations of hair cells are trans- formed into electrical signals in the auditory nerve. In: Principles o f Neural Science (Kandel E. R., Schwartz J. H. and Jessell T. M., eds), pp. 481-491. Prentice-Hall, New York.
Khan K. M., Green G. E., Gordon D., Gupta A. K. and Drescher D. G. (1994) Detection of nicotinic receptor subunits in the mouse cochlea. Pro('. Ann. Meet. Assoc. Res. p. 80. Otolaryngol. 17th St Petersburg. FI. (abstract).
Kujawa S. G., Glattke T. J., Fallon M. and Bobbin R. P. (t993) Contralateral sound suppresses distortion product otoacoustic emissions through cholinergic mechanisms. Hear. Res. 68, 97 106.
Kujawa S. G., Glattke T. J.. Fallon M. and Bobbin R. P. (1994) A nicotinic-like receptor mediates suppression of distortion product otoacoustic emissions by contralateral sounds. Itear. Res. 74, 122--134.
Lawoko G., J~rlebark L. and Heilbronn E. (1995) Ligand binding properties of an unusual nicotinic acetylcholine receptor subtype on isolated outer hair cells from guinea pig cochlea. Neurosci. Lett. Submitted.
kiberman M. C., Dodds L. W. and Pierce S. (1990) Afferent and efferent innervation of the cat cochlea : quantitative analysis with light and electron microscopy. J. comp. Neurol. 301,443 460.
Lustig K. D.. Shiau A. K., Brake A. J. and Julius D. (1993) Expression cloning of an ATP receptor from mouse neuro- blastoma cells. Proc. natn. Acad. Sci. U.S.A. 90, 5[ 13 5117.
Nakagawa T., Akaike N., Kimitsuki T., Komune S. and Arima T. (1990) ATP-induced current in isolated outer hair cells of guinea pig cochlea. J. Neurophysiol. 63, 1068 1074.
Nakazawa K. (1994) ATP-activated current and its rater- action with acetylcholine-activated current in rat sym- pathetic neurons. J. Neurosei. 14, 740 750.
Niedzielski A. S. and Schacht J. (1992) P,, purinoceptors stimulate inositol phosphate release in the organ of Corti. Veuroreport 3, 273 275.
Nilles R., Jarlebark L., Zenner H. P. and Heilbronn E. (1994) ATP-induced cytoplasmic [Ca ~+] increases in isolated cochlear outer hair cells. Involved receptor and channel mechanisms. Heat'. Res. 73, 27 34.
Parr C. E.. Sullivan D. M., Paradiso A. M., Lazarowski E. R., Burch L. H., Olsen J. C., Erb g., Weisman G. A.. Boucher R. C. and Turner J. T. (1994) Cloning and expression of a human P2U nucleotide receptor, a target for cystic fibrosis pharmacotherapy. Proc. natn. Acad. Sci. l . S . A . 91, 3275~ 3279.
Plmkert P. K. (1989) Cholinerge Innervation ~usserer Haar- zellen. Eine m6gliche Bedeutung f~r den Dis- kriminationsverlust bei Perzeptionsschwerh6rigkeiten. Lawngorhinootologie 68, 450 455.
Plinkert P. K., Zenner H. P. and Heilbronn E. (1991) A nicotinic acetylcholine receptor-like c~-bungarotoxin-bind- ing site on outer hair cells. Hear. Res. 53, 123 130.
Plinkert P. K., Gitter A. H., Z immermann U., Kirchner T.. Tzartos S. and Zenner H. P. (1990) Visualization and functional testing of acetylcholine receptor-like molecules m cochlea outer hair cells. Hear. Res. 44, 25 34.

Cochlear efferent synapse : receptors and signalling 311
Puel J. L. and Rebillard G. (1990) Effect of contralateral sound stimulation on the distortion product 2F~-Fz : evi- dence that the medial efferent system is involved. J. Acoust. Soc. Am. 87, 1630-1635.
Schacht J. (1986) Molecular mechanisms of drug-induced hearing loss. Hear. Res. 22, 297 304.
Schacht J. and Zenner H. P. (1987) Evidence that phos- phoinositides mediate motility in cochlear outer hair cells. Hear. Res. 31, 155-160.
Shigemoto T. and Ohmori H. (1990) Muscarinic agonists and ATP increase the intracellular Ca 2+ concentration in chick cochlear hair cells. J. Physiol. (Lond.) 420, 127-148.
Shigemoto T. and Ohmori H. (1991) Muscarinic receptor hyperpolarizes cochlear hair cells of chick by activating Ca2+-activated K + channels. J. Physiol. (Lond.) 442, 669- 690.
Spoendlin H. (1986) Receptoneural and innervation aspects of the inner ear anatomy with respect to cochlear mech- anics. Scand. Audiol. Suppl. 25, 27-34.
Valera S., Hussy N., Evans R. J., Adami N., North R. A., Surprenant A. and Buell G. (1994) A new class of ligand- gated ion channel defined by P2 × receptor for extracellular ATP. Nature 371, 516-519.
Van den Abbeele T., Tran Ba Huy P. and Teulon J. (1994). A calcium-activated nonselective cationic channel in the
basolateral membrane of outer hair cells of the guinea-pig cochlea. Pflagers Archs 427, 56-63.
Wackym P. A., Popper P., Wada K., Wenthold R. J. and Micevych P. E. (1992) Expression of ct2, ~3, a4 and f12 neuronal nicotinic receptor subunit mRNAs in the rat auditory system Proc. Symp. Mol. Biol. Hearing and Deaf- ness, p. 49. La Jolla, Calif. (abstract).
Warr W. B., Guinan J. J. and White J. S. (1986) Organization of the efferent fibers. The lateral and medial olivocochlear systems. In: Neurobiology of Hearing : The Cochlea. (Alt- schuler R. A., Hoffmann D. W. and Bobbin R. P., eds), pp. 333-348. Raven Press, New York.
Webb T. E., Simon J., Krishek B. J., Bateson A. N., Smart T. G., King B. F., Burnstock G. and Barnard E. A. (1993) Cloning and functional expression of a brain G-protein- coupled ATP receptor. FEBS Lett. 324, 219-225.
Whipple M. R. and Drescher D. G. (1984) Muscarinic recep- tors in the cochlear nucleus and auditory nerve of the guinea pig. J. Neurochem. 43, 192 198.
Wiederhold M. L. (1986) Physiology of the olivocochlear system. In: Neurobiology of Hearing: The cochlea (Alt- schuler R. A., Hoffmann D. W. and Bobbin R. P., eds), pp. 349-370. Raven Press, New York.
Zenner H. P., Zimmermann U. and Gitter A. (1987) Fast motility of isolated mammalian auditory sensory cells. Biochem. biophys. Res. Commun. 149, 304-308.





























