Embed Size (px)
Citation preview

R
C
MB
ARRAA
KCMBMAP
C
1
(
i
0h
Plant Science 210 (2013) 177– 182
Contents lists available at SciVerse ScienceDirect
Plant Science
jo ur nal home p age: www.elsev ier .com/ locate /p lantsc i
eview
hloroplast movement
asamitsu Wada ∗
iology Department, Faculty of Science, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, Fukuoka 812-8581, Japan
a r t i c l e i n f o
rticle history:eceived 18 April 2013eceived in revised form 26 May 2013ccepted 27 May 2013vailable online 7 June 2013
eywords:hloroplastovement
lue lighticrobeam irradiation
ctin filamentshototropin
a b s t r a c t
Chloroplast movement is important for plant survival under high light and for efficient photosynthesisunder low light. This review introduces recent knowledge on chloroplast movement and shows how toanalyze the responses and the moving mechanisms, potentially inspiring research in this field. Avoidancefrom the strong light is mediated by blue light receptor phototropin 2 (phot2) plausibly localized on thechloroplast envelop and accumulation at the week light-irradiated area is mediated by phot1 and phot2localized on the plasma membrane. Chloroplasts move by chloroplast actin (cp-actin) filaments that mustbe polymerized by Chloroplast Unusual Positioning1 (CHUP1) at the front side of moving chloroplast. Tounderstand the signal transduction pathways and the mechanism of chloroplast movement, that is, fromlight capture to motive force-generating mechanism, various methods should be employed based on thevarious aspects. Observation of chloroplast distribution pattern under different light condition by fixedcell sectioning is somewhat an old-fashioned technique but the most basic and important way. How-ever, most importantly, precise chloroplast behavior during and just after the induction of chloroplast
movement by partial cell irradiation using an irradiator with either low light or strong light microbeamshould be recorded by time lapse photographs under infrared light and analyzed. Recently various factorsinvolved in chloroplast movement, such as cp-actin filaments and CHUP1, could be traced in Arabidopsistransgenic lines with fluorescent protein tags under a confocal laser scanning microscope (CLSM) and/or atotal internal reflection fluorescence microscope (TIRFM). These methods are listed and their advantages and disadvantages are evaluated.© 2013 Elsevier Ireland Ltd. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1772. Fixed-cell sectioning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1783. Band assay. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1794. Light transmittance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1795. Microbeam assay . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1796. Time lapse photographic analysis of movement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1797. Confocal and TIRF microscopy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1818. Future prospects and concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 182Appendix A. Supplementary data . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 182References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 182
. Introduction
Chloroplasts move toward an area irradiated with weak lightaccumulation response) in a manner that absorbs more light
Abbreviations: cp-actin filaments, chloroplast-actin filaments; TIRFM, totalnternal reflection fluorescence microscope.∗ Tel.: +81 92 642 7252; fax: +81 92 642 7258.
E-mail address: [email protected]
168-9452/$ – see front matter © 2013 Elsevier Ireland Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.plantsci.2013.05.016
allowing efficient photosynthesis but move away from strong lightwhen irradiated directly (avoidance response), avoiding the dam-age caused by the absorption of excess light [1,2] (Fig. 1). Senninitiated experiments that explored chloroplast movement in 1900,studying basic chloroplast distribution patterns under various lightconditions in various tissues of many plant species, including
algae, mosses, ferns, herbaceous plants, and trees. His results andthe knowledge known at that time were published in 1908 [3].However, Senn was not the first to study such responses. A light-dependent chloroplast distribution pattern in a Crassulaceae plant
178 M. Wada / Plant Science 21
Fig. 1. Schematic chloroplast distribution patterns in Arabidopsis mesophyll cells.Chloroplasts are embedded in a thin layer of cytoplasm between the plasma mem-brane and the tonoplast of a large vacuole.Left. Accumulation response. Chloroplasts re-distribute to the upper and lower sidesof palisade cell under low light conditions, achieving maximal light absorption.Center. Avoidance response. Chloroplasts re-distribute on the side walls of palisadecell, minimizing the absorption of strong light.Ra
wlbapr
mpcdHblpIwtmeltpie
is to be enumerated; in this case the section should be more than
TT
Tm
ight. Dark positioning. Chloroplasts distribute on the bottom of the cell in the dark,lthough the physiological function of this distribution is not yet known.
as first reported in 1856 [4]. The induction of chloroplast accumu-ation movement by light was found in 1871 [5]. Although Senn’sook was published more than hundred years ago, it includes a largemount of detailed and extensive knowledge regarding chloro-last movement and is considered the fundamental resource foresearchers studying chloroplast movements.
Senn described the fundamental phenomena of chloroplastovement, that is, the chloroplast distribution pattern and chloro-
last behavior. He described that the effective wavelength forhloroplast movement was, in general, blue light, and he alsoescribed that red light was effective with the alga Mougeotia.e hypothesized why chloroplasts adopted distribution patternsased on the aspects of light paths through individual cells and
eaf tissues. However, the mechanisms and signal transductionathways of chloroplast movement were unknown at the time.
ndeed, the signaling factors involved in signal transduction path-ays, including photoreceptors [6,7], protein factors [8–11], and
he actin structure that underlies the motive force of the move-ent [12,13], were uncovered a hundred years after Senn’s first
xperiment not only in Arabidopsis but also in fern and moss. Weakight, which induces the accumulation response, is perceived byhe blue light receptors phototropin 1 (phot1) and phot2 [7] at the
lasma membrane, whereas phot2 that mediates the strong light-nduced avoidance response [6] is likely localized on the chloroplastnvelope [14].
able 1he merits and demerits of the chloroplast movement analysis methods.
Methods Best for
Fixed-cell-sectioning Chloroplast-distribution pattern, chloroplast number-cBand assay Rapid analysis of many samples, mutant screening
Light transmittance Time course, comparison between mutants
Microbeam assay Single chloroplast behavior, control of chloroplast movMovie analysis Time course, chloroplast behavior, speed of movementCLSF microscopy Observation of cp-actin filaments, time course of cp-ac
filament behaviorTIRF microscopy Single cp-actin filament behavior
Cryoelectron microscopy Structure of cp-actin filaments
he most appropriate method “Best for” or less appropriate method “Useless for” and (Dicroscopy: confocal laser scanning fluorescent microscopy. TIRF microscopy: total inter
0 (2013) 177– 182
The “signal” that is transmitted from the photoreceptor to thechloroplasts, the most interesting and important subject is notyet known despite the rapid advances during the last 13 years.Phosphoinositides and Ca2+, however, have been proposed ascandidates [15].
The number of groups working on chloroplast movementsand/or researchers joining this field is increasing, and variousattempts have been made to study the mechanism and/or phys-iological relevance of chloroplast movements. Due to the advancesin molecular biological techniques, many genes involved in chloro-plast movements have been identified through mutant analyses[6,8–11]. However, to fully understand them, chloroplast move-ments should be analyzed from a different perspective. Preciseobservation of chloroplast behavior and functional analyses ofthe protein factors involved using various techniques are stillneeded. As each method has its advantages and disadvantages, thepreferable method is dependent on the researcher’s experimentalpurposes and the available facilities.
The main method currently employed for the detection ofchloroplast movement is the measurement of red light trans-mittance through a leaf [16]. Transmittance decreases when thechloroplasts gather at the palisade cell surface, whereas the trans-mittance increases when the chloroplasts move from that position.Therefore, chloroplast movement can be recorded automaticallyby placing leaves in a photometer [16], and the resulting chartfacilitates the data analysis.
The merits and shortcomings of each method used to detectchloroplast movement (Table 1), i.e., observation of chloroplast dis-tribution pattern using fixed and sectioned leaves, measurement ofred light transmittance through leaf tissue, and direct observationof chloroplast movement under microscopy, is discussed below.
2. Fixed-cell sectioning
Senn [3] studied chloroplast movement in various tissues ofmany plant species mainly by sectioning plant materials that weremost likely fixed with chemicals (Fig. 2A). When we observe chloro-plast movement using a whole leaf without sectioning, it is difficultto detect the precise distribution pattern of all chloroplasts (thenumber and their position) in a long, thin palisade cell (even if theuppermost cell) of a thick leaf from the leaf surface through an epi-dermal cell layer, even when using a confocal microscope changingits focal plane from the top to the bottom of the cell (Fig. 2B). Thebest way is fixed cell observation of leaf cross sections, as Senndid (Fig. 2A). Although the chloroplast distribution pattern can beobserved with a thin section (1 �m or less), the whole cell shouldbe included in the section if the number of chloroplasts in a cell
50 �m in thickness. The sectioning of a leaf (with or without fixa-tion) embedded in an agar block with a vibrating blade microtome,such as a Leica VT1200S, is commonly used (Fig. 2C).
Useless for, (disadvantage)
ounting Time course studyTime course study, weak responseDirect observation of chloroplasts, (influenced by other factors)
ement Rapid analysis of many samples, (need special equipment) (need special equipment)tin (need transgenic line), (need costy equipment)
(need transgenic line), (need costy equipment)(need costy equipment), (time consuming)
isadvantage) for chloroplast movement analyses were listed with comments. CLSFnal reflection fluorescence microscope.

nce 21
ucmarotad
3
p[sbic1peatcdmlmttcc
4
totiatcsrltifirrllctc
tccTpe
M. Wada / Plant Scie
Chloroplasts are easily observed by chlorophyll fluorescencesing fluorescence microscopy. However, as fixed leaves are, ofourse, used for this method, live chloroplast behavior duringovement cannot be observed; indeed, only the state of movement
t the time of fixation can be observed, and the time course andoute of the moving chloroplasts cannot be analyzed. Observationf chloroplast movement in a non-fixed, live Arabidopsis leaf sec-ion is possible [12]. However, it is not realistic because persever-nce is required to accomplish this goal; moreover, the movementoes not reveal the chloroplast behavior under natural conditions.
. Band assay
The green or white band assay is a simple way to detect chloro-last movement without the requirement of expensive equipment6,9,16]. Detached leaves (Fig. 2Da) are placed on the surface ofolidified agar or wet paper, covered with a non-light-transmissibleoard that has a narrow approximately one millimeter slit, and
rradiated with strong white light through the slit. The leafolor in the region irradiated for 1–2 h with strong light (e.g.,00 �mol m−2 s−1) changes from green to pale green due to chloro-last avoidance (white band assay) (Fig. 2Db). However, if thentire leaf is pre-irradiated with strong light for 1–2 h (Fig. 2Dc)nd then irradiated with weak light through the slit, the color ofhe irradiated area changes from pale green to dark green due tohloroplast accumulation (green band assay) (Fig. 2Dd). The colorifference can be detected by the naked eye. This is the standardethod employed for detecting chloroplast movement in many
eaves at once within a short period and, thus, is very powerful inutant screening [6,8–11]. However, the greatest disadvantage of
his method is that weak phenotypic changes may be undetected,herefore weak mutant lines may be overlooked. Furthermore, wean only detect the state at the time of observation and not duringhloroplast movement.
. Light transmittance
The measurement of red light transmittance (or light absorp-ion) through a leaf is currently a sophisticated and popular methodf evaluating chloroplast movement [11,16] and can also monitorhe time course of chloroplast movement. This method was orig-nally applied for the detection of chloroplast movement as lightbsorption but not light transmission [17]. The basic mechanism ofhe equipment is simple, and photometers for this purpose can beonstructed in-lab [16]. A leaf incubated for some period of time,uch as 1 h in the dark (dark adaptation), is inserted between aed light source and a photodiode for measuring the transmittedight. During the interval between the measurements of transmit-ed light, the leaf is exposed to weak blue light (or white light) fornducing an accumulation response, and then strong light is appliedor inducing an avoidance response. Transmittance decreases dur-ng accumulation and increases during avoidance (Fig. 2E). Theesulting time course shows two important factors: the level ofesponse and the speed of chloroplast movement. The saturatedevel of transmittance is mainly the function of light strength andeaf thickness, i.e., the number of cell layers of mesophyll cells orhloroplast number (or density) throughout a leaf. The change inhe speed of transmittance mainly indicates the translocation ofhloroplasts from the cell surface to the side walls.
It should be noted that chloroplasts move specifically alonghe plasma membrane and not through vacuoles, meaning that allhloroplasts necessarily move to the edge of the cell surface (peri-
linal wall) and then move down to the side wall (anticlinal wall).he advantage of this method is that patterns can be precisely com-ared, even if the changes are small. Therefore, responses can bevaluated numerically, and mutants can be screened or analyzed0 (2013) 177– 182 179
based on the different slope patterns and/or different levels ofresponse. It is important, however, to keep in mind that the cul-ture conditions of the sample plants should be consistent becausetransmittance changes are easily affected by the diameter of thepalisade cells, which is controlled by the light fluence rate [18]. Inaddition, the chart does not show actual chloroplast movement,just the optical result of such movement.
Another important concern in the time course study performedby this and other methods is the competition between the accu-mulation and avoidance responses. Both responses are inducedsimultaneously by blue light but in opposite directions; however,the signal for the accumulation response can be transferred over along distance, whereas the signal for the avoidance response canreach only a short distance [19]. Both signals are released fromthe same irradiated area when chloroplasts are present. In thejac1 mutant, which lacks the accumulation response, the avoidanceresponse occurs at a lower fluence rate than with wild-type plants[9]. In contrast, accumulation occurs more rapidly in an avoidanceresponse-deficient phot2 mutant than in wild-type plants [20]. Con-sequently, even if the chart shows accumulation under weak lightor avoidance under strong light, both the level and speed of move-ment are under the influence of contradictory directional forces.
5. Microbeam assay
A moving chloroplast must be observed precisely to under-stand chloroplast movement. For this purpose the movement undera microscope must be controlled, an often unachievable goal.Although chloroplast movement can be induced on a microscopestage by white light irradiation, it is difficult to detect or controlthe movement using an ordinary microscope. In most of higherplant species, chloroplast movement is induced by blue light butnot red light, and chloroplasts accumulate under weak blue light(5 �mol m−2 s−1) and the avoidance response occurs under strongblue light (100 �mol m−2 s−1). Accordingly, the microscope setupneeds to cover these requirements by using red or a longer wave-length to observe chloroplast movement; this setup is facilitatedby the insertion of a red filter that prevents shorter wavelengthsof light in a portion of the light path between the light sourceand sample. Another important issue is how to induce chloroplastmovement. For this purpose, a leaf should be irradiated with weakor strong blue light. Thus, the microscope has to have two lightsources, one for red or longer wavelengths that are not functionalfor chloroplast movement and another for blue light for chloroplastmovement induction. Our group uses infrared light longer than850 nm for observation because red light is functional in chloro-plast movement of our materials (fern and moss). In this case, aninfra-red responsive camera attachment is required.
An epifluorescence microscope is ideal for this purpose becauseit has two light sources. We can use the one from the bottom forobservation with red light and the other from the top for chloro-plast movement induction with blue light. Furthermore, partial cellirradiation with blue light as a microbeam that is focused on a smallarea of a cell is very effective for precise experiments (Fig. 2F).We can observe chloroplast behavior moving toward a weak light-irradiated area or away from a strong light spot. Our laboratorydeveloped such a microbeam irradiator, whereby the color of light(blue, red, and infrared), and the size (1–100 �m), shape (roundand square), duration, and strength of the microbeam used can bechanged [21].
6. Time lapse photographic analysis of movement
Chloroplast responses are best observed under infrared lightbecause even red light can influence chloroplast movement [22].Chloroplasts can be continuously observed under infrared light
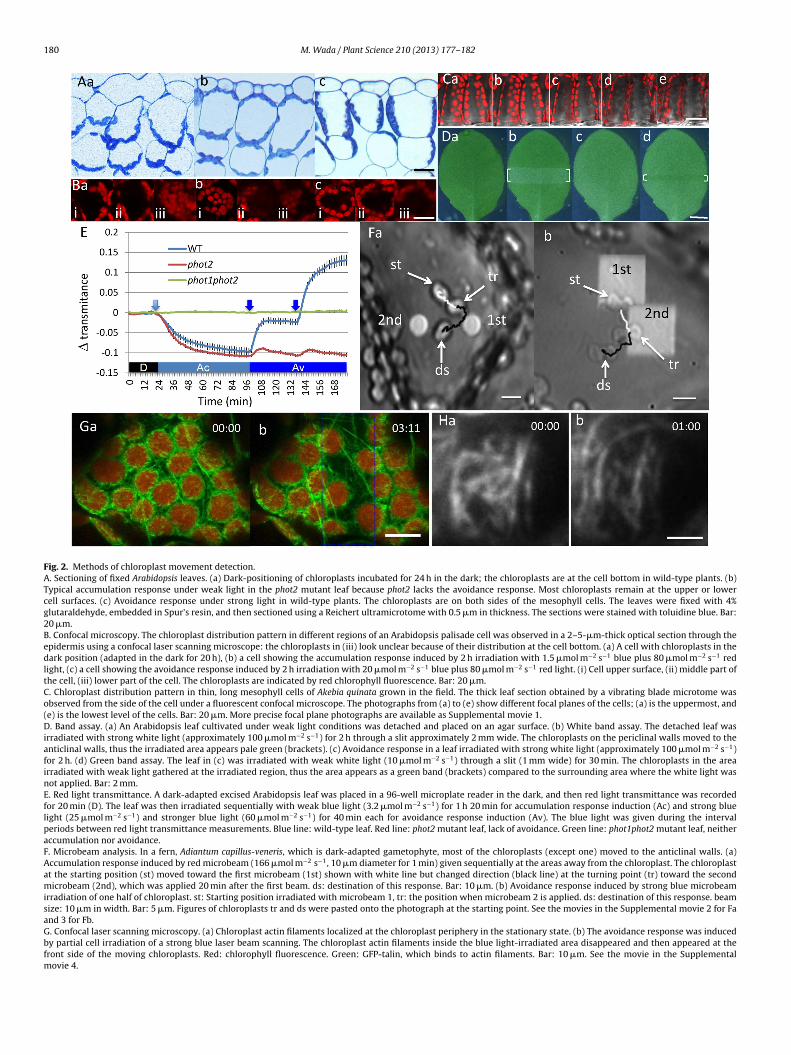
180 M. Wada / Plant Science 210 (2013) 177– 182
Fig. 2. Methods of chloroplast movement detection.A. Sectioning of fixed Arabidopsis leaves. (a) Dark-positioning of chloroplasts incubated for 24 h in the dark; the chloroplasts are at the cell bottom in wild-type plants. (b)Typical accumulation response under weak light in the phot2 mutant leaf because phot2 lacks the avoidance response. Most chloroplasts remain at the upper or lowercell surfaces. (c) Avoidance response under strong light in wild-type plants. The chloroplasts are on both sides of the mesophyll cells. The leaves were fixed with 4%glutaraldehyde, embedded in Spur’s resin, and then sectioned using a Reichert ultramicrotome with 0.5 �m in thickness. The sections were stained with toluidine blue. Bar:20 �m.B. Confocal microscopy. The chloroplast distribution pattern in different regions of an Arabidopsis palisade cell was observed in a 2–5-�m-thick optical section through theepidermis using a confocal laser scanning microscope: the chloroplasts in (iii) look unclear because of their distribution at the cell bottom. (a) A cell with chloroplasts in thedark position (adapted in the dark for 20 h), (b) a cell showing the accumulation response induced by 2 h irradiation with 1.5 �mol m−2 s−1 blue plus 80 �mol m−2 s−1 redlight, (c) a cell showing the avoidance response induced by 2 h irradiation with 20 �mol m−2 s−1 blue plus 80 �mol m−2 s−1 red light. (i) Cell upper surface, (ii) middle part ofthe cell, (iii) lower part of the cell. The chloroplasts are indicated by red chlorophyll fluorescence. Bar: 20 �m.C. Chloroplast distribution pattern in thin, long mesophyll cells of Akebia quinata grown in the field. The thick leaf section obtained by a vibrating blade microtome wasobserved from the side of the cell under a fluorescent confocal microscope. The photographs from (a) to (e) show different focal planes of the cells; (a) is the uppermost, and(e) is the lowest level of the cells. Bar: 20 �m. More precise focal plane photographs are available as Supplemental movie 1.D. Band assay. (a) An Arabidopsis leaf cultivated under weak light conditions was detached and placed on an agar surface. (b) White band assay. The detached leaf wasirradiated with strong white light (approximately 100 �mol m−2 s−1) for 2 h through a slit approximately 2 mm wide. The chloroplasts on the periclinal walls moved to theanticlinal walls, thus the irradiated area appears pale green (brackets). (c) Avoidance response in a leaf irradiated with strong white light (approximately 100 �mol m−2 s−1)for 2 h. (d) Green band assay. The leaf in (c) was irradiated with weak white light (10 �mol m−2 s−1) through a slit (1 mm wide) for 30 min. The chloroplasts in the areairradiated with weak light gathered at the irradiated region, thus the area appears as a green band (brackets) compared to the surrounding area where the white light wasnot applied. Bar: 2 mm.E. Red light transmittance. A dark-adapted excised Arabidopsis leaf was placed in a 96-well microplate reader in the dark, and then red light transmittance was recordedfor 20 min (D). The leaf was then irradiated sequentially with weak blue light (3.2 �mol m−2 s−1) for 1 h 20 min for accumulation response induction (Ac) and strong bluelight (25 �mol m−2 s−1) and stronger blue light (60 �mol m−2 s−1) for 40 min each for avoidance response induction (Av). The blue light was given during the intervalperiods between red light transmittance measurements. Blue line: wild-type leaf. Red line: phot2 mutant leaf, lack of avoidance. Green line: phot1phot2 mutant leaf, neitheraccumulation nor avoidance.F. Microbeam analysis. In a fern, Adiantum capillus-veneris, which is dark-adapted gametophyte, most of the chloroplasts (except one) moved to the anticlinal walls. (a)Accumulation response induced by red microbeam (166 �mol m−2 s−1, 10 �m diameter for 1 min) given sequentially at the areas away from the chloroplast. The chloroplastat the starting position (st) moved toward the first microbeam (1st) shown with white line but changed direction (black line) at the turning point (tr) toward the secondmicrobeam (2nd), which was applied 20 min after the first beam. ds: destination of this response. Bar: 10 �m. (b) Avoidance response induced by strong blue microbeamirradiation of one half of chloroplast. st: Starting position irradiated with microbeam 1, tr: the position when microbeam 2 is applied. ds: destination of this response. beamsize: 10 �m in width. Bar: 5 �m. Figures of chloroplasts tr and ds were pasted onto the photograph at the starting point. See the movies in the Supplemental movie 2 for Faand 3 for Fb.G. Confocal laser scanning microscopy. (a) Chloroplast actin filaments localized at the chloroplast periphery in the stationary state. (b) The avoidance response was inducedby partial cell irradiation of a strong blue laser beam scanning. The chloroplast actin filaments inside the blue light-irradiated area disappeared and then appeared at thefront side of the moving chloroplasts. Red: chlorophyll fluorescence. Green: GFP-talin, which binds to actin filaments. Bar: 10 �m. See the movie in the Supplementalmovie 4.

nce 21
waiwhimnmaitdcc
7
taa(pTppfapctc
rmmu3t
(icTpcw(aconhfwfl
8
s
Hss
M. Wada / Plant Scie
ithout modifying their behavior after partial cell irradiation with blue microbeam. However, the speed of chloroplast movements approximately 1 �m min−1 [23], which is too slow to detect
ith the naked eye. Additionally, infrared light is invisible toumans and movement must be continually recorded using an
nfrared-sensitive time-lapse camera, taking photographs at 1–5-in intervals and replayed at 100 or 600 times faster than the
ormal speed. These movies are useful for precise analysis of move-ent on a monitor (see Supplemental movie 2 for Fig. 2F). We
nalyzed chloroplast movement precisely using the microbeamrradiator combined with a time-lapse movie system and foundhat chloroplasts moved instantaneously without rotation to anyirection toward or away from a microbeam-irradiated area, indi-ating that chloroplasts do not have heads and tails in light-inducedhloroplast movement [1].
. Confocal and TIRF microscopy
Our group recently identified a novel actin filament structurehat is specifically involved in chloroplast movement: chloroplast-ctin filaments (cp-actin filaments) (Fig. 2G). This was done using
transgenic Arabidopsis line expressing green fluorescent proteinGFP)-tagged with the mouse high-molecular-weight cytoskeletonrotein talin (GFP-talin), which binds to actin filaments [12,13].he cp-actin filaments localize between the chloroplasts and thelasma membrane around the chloroplast periphery when chloro-lasts are stationary. The cp-actin filaments accumulate at theront side of the chloroplasts just prior to the start of movementnd during movement, which typically begins a few minutes afterhotoinduction [12,13]. The chloroplasts cannot move without thep-actin filaments, such as in chup1 and kac1 kac2 double mutantshat lack cp-actin filaments. However, the mechanism by which thehloroplasts move is yet unknown.
The turnover and movement of cp-actin filaments are veryapid, as carefully observed by confocal scanning fluorescenceicroscopy (Leica SP5) [13]. More importantly, the cp-actin fila-ents easily disappear under the laser-scanned, strong blue light
sed for observation. Although photographs were obtained every s, it was not possible to follow each cp-actin filament. Thus, higheremporal and spatial resolution is necessary to obtain reliable data.
Accordingly, a total internal reflection fluorescence microscopeTIRFM) is useful to obtain such resolution. Because fluorescentmages of materials at only 100 nm from the cover slip could beollected as time-lapse images every 0.2 s interval in our case [13].he structure of cp-actin filaments in Arabidopsis mesophyll cellrotoplasts and their disappearance under strong light were pre-isely analyzed using a TIRFM [13] (Fig. 2H). The cp-actin filamentsere short and zig-zag shaped and disintegrated into small pieces
0.32 �m) by the laser beam scanning for observation and then dis-ppeared. This process is different from that of long and smoothytoplasmic actin filaments which are depolymerized at the endsf severed fragments. The severed ends of cp-actin filaments areot depolymerized [13]. It will be necessary to utilize a TIRFM thatas a microbeam irradiation system and an observation system for
urther analyses of various protein factors simultaneously labeledith different marker proteins, such as cyan, green, yellow, and reduorescent proteins.
. Future prospects and concluding remarks
Chloroplast movement is a simple and short-lasting but verylow response that occurs at the plasma membrane of mesophyll
. TIRF microscopy. The process of chloroplast actin filament disappearance was observetrong blue light irradiation. The chloroplast actin filaments are shown as white lines. (b)
mall pieces. Bar: 2 �m. See the movie in the Supplemental movie 5.
0 (2013) 177– 182 181
cells. Although it is thought that gene expression is not involvedin the signal transduction pathway based on studies with enucle-ated cells [24], the second messengers or signals released fromphot1 and phot2 at both the plasma membrane and chloroplastenvelope are unknown. The physiological characteristics of thesignals for the accumulation and avoidance responses are dif-ferent: the former is long-lived and long-distance transferable,whereas the latter is short-lived and short-distance transferable.These characteristics are consistent both in Arabidopsis thalianaand fern Adiantum capillus-veneris gametophyte cells, indicatingthe establishment at the early stage of chloroplast movementevolution.
The involvement of different species of phosphatidylinositidesbetween the phot1- and phot2-induced accumulation responseand phot2-induced avoidance response has been studied usinginhibitors that function on different pathways of phosphatidyli-nositide metabolism [15]. The results suggest that the cytosolic Ca2+
signaling modulated by different species of phosphatidylinositidesmay function on chloroplast movement. If Ca2+ is the real signal forboth responses, the different characteristics between the accumu-lation and avoidance signals should be dependent on a differentsensitivity or requirement of different Ca2+ concentrations of thesignal receptors for the two responses. The signal (or signals) shouldbe emitted from the photoreceptors, and the receptors for the sig-nal(s) must be present and localized in the chloroplast envelope.However, as the identification of the signal is difficult, the searchfor the receptor proteins on the chloroplast outer envelope may bemore rapid and is more promising.
Chloroplast-actin filaments are the most important machin-ery for chloroplast movement. Although the mechanism of forcegeneration for chloroplast movement is not yet known, themodel that cp-actin polymerization itself generates the forcehas been proposed (Fig. 3). The involvement of myosin is notplausible based on the use of myosin deletion mutants in whichchloroplast movement was normal although other organelle move-ment completely stopped [25]. Chloroplast Unusual Positioning1(CHUP1) is the most important protein plausibly involved incp-actin polymerization. CHUP1 is localized on the chloroplastenvelope with its N terminus hydrophobic domain and has anactin-binding domain [1,8], so that the function of CHUP1 inactin polymerization should be studied in an in vitro system.The analysis of the precise structure of cp-actin filaments usingcryo-electron microscopy at several-angstrom resolution is alsoimportant to elucidate this mechanism, as it can show which arethe fast-growing (barbed) and the slow-growing (pointed) ends,and how many actin filaments are involved as one unit in thismovement.
Studies using model plants, such as the seed plants Arabidopsisthaliana, fern Adiantum capillus-veneris, moss Physcomitrella patens,liverwort Marchantia polymorpha, and alga Mougeotia scalaris, cul-tivated in culture rooms are highly efficient for investigating themechanisms of chloroplast movement. However, to understandthe real physiological meaning of chloroplast movement in natureunder various light conditions (plants grown under strong light,weak light, or fluctuating light conditions), we have to study howchloroplasts in wild plants move and are protected from strong lightor are oriented in a way that facilitates optimal utilization of weak
light [26]. For this purpose, analyses from various aspects and meth-ods are needed, including such highly technical methods as TIRFMand also conventional, fixed-cell sectioning. Each method has mer-its (see Table 1) that cannot be replaced by the other. Indeed,d using a total internal reflection fluorescence (TIRF) microscope. (a) Just prior toOne minute after blue light irradiation. The chloroplast actin filaments appeared as

182 M. Wada / Plant Science 21
Fig. 3. A model of chloroplast movement in Arabidopsis thaliana. Both phot1 andphot2, which mediate the accumulation response, are localized at the plasmamembrane (PM). phot2 on the chloroplast envelope likely mediates the avoidanceresponse, because the amount of phot2 on the chloroplast envelope is very highcomparing to that of phot1. Further, avoidance response was induced only whenchloroplasts were illuminated with high light. CHUP1 binds to the chloroplast enve-lope via its N-terminus and may also be anchored to the plasma membrane throughan unknown membrane-bound protein, X. CHUP1 recruits profilactin (profilin/actincomplex) and polymerizes F-actin by inserting G-actin between itself and an exist-ing F-actin filament. THRUMIN1 at the plasma membrane bundles the resultingF-actin filaments and fixes them to the plasma membrane as an anchor. Conse-quently, CHUP1 and the chloroplast are pushed by the inserted G-actin, generatingthe motive force for chloroplast photorelocation movement. Cp-actin filaments areddt
ow
A
saGC1P
A
fj
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[Cell Physiol. 29 (1988) 1227–1232.
epolymerized at the pointed end of actin filaments. The green arrow shows theirection of chloroplast movement. (For interpretation of the references to color inhis figure legend, the reader is referred to the web version of the article.)
rganisms are complicated and have their own characteristics, thuse need to observe them by various means.
cknowledgements
The author thanks Hidenori Tsuboi, Takeshi Higa, Noriyuki Suet-ugu, Eiji Gotoh and Sam-Geun Kong for preparation of figuresnd supplemental movies. This work was supported in part byrants-in-Aid for scientific research from the Ministry of Education,ulture, Sports, Science and Technology of Japan (MEXT) (Grants7084006 and 23120523 to M.W.) and the Japan Society for theromotion of Science (JSPS) (Grant 20227001 to M.W.).
ppendix A. Supplementary data
Supplementary data associated with this article can beound, in the online version, at http://dx.doi.org/10.1016/.plantsci.2013.05.016.
eferences
[1] N. Suetsugu, M. Wada, Chloroplast photorelocation movement: a sophisticatedstrategy for chloroplasts to perform efficient photosynthesis, in: M.M. Najaf-pour (Ed.), Advances in Photosynthesis—Fundamental Aspects, In Tech, Rijeka,2012, pp. 215–234.
[
[
0 (2013) 177– 182
[2] A.K. Banas, C. Aggarwal, J. Łabuz, O. Sztatelman, H. Gabrys, Blue light signallingin chloroplast movements, J. Exp. Bot. 63 (2012) 1559–1574.
[3] G. Senn, Die Gestalts- und Lageveränderung der Pflanzen-Chromatophoren,Wilhelm-Engelmann, Leipzig, 1908.
[4] J.A. Böhm, Beiträge zur näheren Kenntnis des Chlorophylls, S.B. Akad. Wiss.Wien, Math.-nat. Kl. 22 (1856) 479–498.
[5] B. Frank, Über lichtwärts sich bewegende Chlorophyllkörner, Bot. Ztg. 29 (1871)209.
[6] T. Kagawa, et al., Arabidopsis NPL1: a phototropin homolog control-ling the chloroplast high-light avoidance response, Science 291 (2001)2138–2141.
[7] T. Sakai, et al., Arabidopsis nph1 and npl1: blue light receptors that mediateboth phototropism and chloroplast relocation, Proc. Natl. Acad. Sci. USA 98(2001) 6969–6974.
[8] K. Oikawa, et al., Chloroplast unusual positioning1 is essential for proper chloro-plast positioning, Plant Cell 15 (2003) 2805–2815.
[9] N. Suetsugu, T. Kagawa, M. Wada, An auxilin-like J-domain protein, JAC1,regulates phototropin-mediated chloroplast movement in Arabidopsis, PlantPhysiol. 139 (2005) 151–162.
10] N. Suetsugu, et al., Two kinesin-like proteins mediate actin-based chloro-plast movement in Arabidopsis thaliana, Proc. Natl. Acad. Sci. USA 107 (2005)8860–8865.
11] S.L. DeBlasio, D.L. Luesse, R.P. Hangarter, A plant-specific protein essen-tial for blue-light-induced chloroplast movements, Plant Physiol. 139 (2005)101–114.
12] A. Kadota, et al., Short actin-based mechanism for light-directed chloro-plast movement in Arabidopsis, Proc. Natl Acad. Sci. USA 106 (2009)13106–13111.
13] S.-G. Kong, Y. Arai, N. Suetsugu, T. Yanagida, M. Wada, Rapid severing and motil-ity of chloroplast-actin filaments are required for the chloroplast avoidanceresponse in Arabidopsis, Plant Cell 25 (2013) 572–590.
14] S.-G. Kong, et al., Both phototropin 1 and 2 localize on the chloroplast outermembrane with distinct localization activity, Plant Cell Physiol. 54 (2013)80–92.
15] C. Aggarwal, J. Łabuz, H. Gabrys, Phosphoinositides play differential roles inregulating phototropin1- and phototropin2-mediated chloroplast movementsin Arabidopsis, PLoS ONE 8 (2013) e55393.
16] M. Wada, S.-G. Kong, Analysis of chloroplast movement and relocation in Ara-bidopsis, in: R.P. Jarvis (Ed.), Chloroplast research in Arabidopsis, Methods andProtocols, vol. 1, Methods in Molecular Biology, vol. 774, Humana Press, Totowa,NJ, USA, 2011, pp. 87–102.
17] Y. Inoue, K. Shibata, Light-induced chloroplast rearrangements and their actionspectra as measured by absorption spectrophotometry, Planta 114 (1973)341–358.
18] T. Kozuka, S.-G. Kong, M. Doi, K. Shimazaki, A. Nagatania, Tissue-autonomouspromotion of palisade cell development by phototropin 2 in Arabidopsis, PlantCell 23 (2011) 3684–3695.
19] T. Kagawa, M. Wada, Chloroplast-avoidance response induced by high-fluenceblue light in prothallial cells of the fern Adiantum capillus-veneris as analyzedby microbeam irradiation, Plant Physiol. 119 (1999) 917–923.
20] S. Ichikawa, N. Yamada, N. Suetsugu, M. Wada, A. Kadota, Red light, phot1and JAC1 modulate phot2-dependent reorganization of chloroplast actin fil-aments and chloroplast avoidance movement, Plant Cell Physiol. 52 (2011)1422–1432.
21] M. Wada, Photoresponses in fern gametophytes, in: T.A. Ranker, C.H. Haufler(Eds.), The Biology and Evolution of Ferns and Lycophytes, Cambridge Univer-sity Press, 2008, pp. 3–48.
22] D.R. Luesse, S.L. DeBlasio, R.P. Hangarter, Integration of phot1, phot2, andphyB signalling in light-induced chloroplast movements, J. Exp. Bot. 61 (2010)4387–4397.
23] H. Tsuboi, M. Wada, Speed of signal transfer in the chloroplast accumulationresponse, J. Plant Res. 123 (2010) 381–390.
24] M. Wada, Chloroplast photoorientation in enucleated fern protonemata, Plant
25] N. Suetsugu, V.V. Dolja, M. Wada, Why have chloroplasts developed a uniquemotility system? Plant Signal. Behav. 5 (2010) 1190–1196.
26] W.E. Williams, H.L. Gorton, S.M. Witiak, Chloroplast movements in the field,Plant Cell Environ. 26 (2003) 2005–2014.





























