Embed Size (px)
Citation preview

Protoplasma (1993) 175:1-8
�9 Springer-Vertag 1993 Printed in Austria
Chromatin structure variation in successful and unsuccessful arbuscular mycorrhizas of pea
S. Sgorbati 1'*, G. Berta 2, A. Trotta 2, L. Sehellenbaum 3, S. Citterio 1, M. Dela Pierre 3, V. Gianinazzi -Pearson 3, and S. Scannerini 2
~Dipartimento di Biologia, Sezione Botanica Generate, Universit/t di Milano, Milano, z Dipartimento di Biologia Vegetale, Universitfi di Torino, Torino, and 3 Laboratoire de Phytoparasitologie INRA-CNRS, Station de G6n6tique et d'Am61ioration des Plantes, INRA, Dijon
Received December 30, 1992 Accepted March 29, 1993
Summary, Lincoln and Frisson varieties of endomycorrhiza-forming pea plants and isogenic mycorrhiza-resistant Frisson mutant ~ 2) plants were inoculated ~:ith Gtomus mosseae. Nuclei released from inoculated and non-inoculated (control) roots were analysed for chromatin structure and activity using flow cytometric techniques. Chromatin accessibility to the specific DNA fluorochrome DAPI at saturating and non-saturating concentrations was measured. DNA fluorescence of nuclei of mycorrhizal Lincoln and wild genotype Frisson plants was significantly increased, compared to the controls, at saturating and, more strongly, at non-saturating DAPI concen- trations. In contrast, the nuclei of inoculated P2 mutant roots showed a much lower increase in fluorescence, compared to unin- oculated controls. Nuclei released from mycorrhiza-infected Lincoln roots were more sensitive to DNase I than those of uninfected ones. These results indicate a dramatic increase in that portion of the genome which can be transcribed in response to AM infection.
Keywords: Chromatin structure; DAPI; DNase I; Flow cytometry; Pea arbuscular mycorrhizas.
Abbreviations: AM arbuscular mycorrhizas; CRBCs chicken red blood cells; CV coefficient of variation; DAPI 4",6-diamidino-2- phenylindole; DNase I deoxyribonuclease I; EDTA ethylenediamine tetraacetic acid; FCM flow cytometry; TMN Tris MgC12 NaC1 buffer.
Introduction
Much recent work has investigated the chromatin struc- ture of various organisms, especially in relation to its active, partially active, or inactive state (van Holde 1989). Plant research is lagging behind in this area
* Correspondence and reprints: Dipartimento di Biologia, Sezione Botanica Generale, Universit~t di Milano, Via Celoria 26, 1-20133 Milano, Italy.
(Slatter and Gray 1991), although there have been many studies of changes in chromat in structure during the cell cycle and differentiation using optical and elec- tron microscopy (Nagl 1985).
Flow cytometry (FCM) appears to be an ideal tech- nique for studying chromatin structure "in situ" at the supranucleosomal level, since biochemical procedures to unravel chromatin superstructure destroy its native conf igurat ion (Darzynkiewicz 1990). Many D N A fluo- rochromes can be used as probes for chromatin struc- ture in different cell systems (Cowden and Curtis 1981). In relation to cell differentiation, there are two clear examples of variat ion in D N A accessibility to various fluorochromes detected by flow cytometry in animal
systems, i.e., erythroid differentiation and spermato- genesis (Darzynkiewicz et al. 1984, Evenson et al. 1986). Differential D N A staining of leukocyte populations
with propidium iodide has also been detected at non- saturating fluorochrome concentrations (Mazzini et al. 1983). D N A sensitivity to nuclease digestion, measured by flow cytometry, has also proved useful in assessing chromatin structure and activity in different cell types (Prentice etal. 1985, Roti Roti etat. 1985, PelIicciari etal. 1987). In plant material, FCM using various fluorochromes has been employed to show changes in chromatin struc- ture during the cell cycle ofEuglena (Bonaly et al. 1987), or in root tissue after arbuscular mycorrhiza (.AM) infect ion at non-saturating dye concentrations (Berta etal. 1990). By measuring the D N A accessibility of

2 S. Sgorbati etal.: Chromatin in arbuscular mycorrhizas of pea
nuclei isolated f rom leek roots to non-sa tu ra t ing D A P I
staining, Berta etal . (1990) showed that nuclear hy-
per t rophy in A M is related to var ia t ion in ch romat in
structure (and activity), as a response of host cells to
the fungal infection.
In the present work, we used non-sa tu ra t ing D A P I
concentra t ions and a DNase I digestion protocol on
nuclei released from infected and uninfected pea roots,
together with electron microscopy to conf i rm the hy-
pothesis that the more dispersed and active chromat in
produced in response to A M infection is similar in
different, unrelated plants, and therefore a general fea-
ture of A M infection. We also report var ia t ion in chro-
mat in structure between different pea genotypes which
are resistant or susceptible to A M fungal infection.
Materials and methods
Plant growth cultivation and infection
Seeds of Pisum sativum L. cv. Lincoln, Frisson, and an isogenic mycorrhiza-resistant Frisson mutant (P 2; Duc et al. 1989) were ster- ilized in 1% NaC10 and sown in quartz sand. Some were inoculated with chopped roots from pot plants colonized by GIomus mosseae (Nicot and Gerd) Gerdmann and Trappe; non-inoculated pea plants served as controls. On alternate days, a low phosphorus solution was apNied (Berta et al. 1990). The seedlings were maintained in a growth chamber (20 ~ 16 h photoperiod, 150 gE/ma/s of irradiance at pot height). Mycorrhizal and control roots were harvested 1 month after sowing, at the time of maximal growth response in mycorrhizal plants. The degree of root infection was evaluated microscopically using staining techniques (Phillips and Hayman 1970) and coloni- zation percentage estimates of the root system were made using a grid intersect method (Giovannetti and Mosse 1980).
Electron microscopy
Small pieces (1-2ram) of mycorrhizal and non-mycorrhizal 7-8 cm long roots of P. sativum cv. Lincoln were cut 5 cm behind the apex. The samples were fixed in 3% glutaraldehyde (pH 7.2) at room tem- perature for 3h, rinsed, postfixed in 1% osmium tetroxide in the same buffer for 2h at room temperature, washed, dehydrated in an ethanol series and embedded according to Spurt (1969). Semithin (1-2 gin) and thin sections were cut with diamond knives on a Reich- ert Ultracnt ultramicrotome. Sections were stained with 1% toluidine blue in 1% borax for examination by light microscopy, or with uranyl acetate and lead citrate for transmission electron microscopy (Philips CM 10).
Sample preparation for flow cytometry
About 2 g of infected and control pea roots without apices were fixed in 4% (w/v) formaldehyde for 1 h at 4 ~ in Tris buffer and then washed thoroughly in the same cold buffer (Berta etal. 1990). Em- bryo roots growing from ev. Lincoln germinating seeds t5h after wetting were also fixed as above. Root samples were crushed with a glass rod to release nuclei in a few milliliters of buffer. Nuclear suspensions were filtered through a nylon mesh of 60 and 15 gm pore size (Sgorbati et al. 1986). All the operations were carried out on ice. The concentration of the nuclear suspensions was adjusted
with buffer in order to obtain 3 x 105 pea nuclei per ml in each sample.
Staining conditions for accessibility measurements
4',6-diamidino-2-phenylindole (DAPI) has previously been found to be superior to other fluorochromes as a probe for plant chromatin structure in root meristems using flow cytometry, with an arc lamp instrument (Berta et at. 1990). Coefficients of variation (CVs) of 2 C nuclei did not exceed 1 I%, even at non-saturating dye concentra- tions. Saturating and non-saturating dye concentrations were estab- lished from saturating curves for each pea genotype. To test DNA accessibility at saturating and non-saturating fluorochrome concen- trations it is necessary to control instrument and staining variability; chicken red blood cells (CRBCs), prepared as previously described (Sgorbati et al. 1986), were added as an internal biological standard to each sampte before staining (proportion not exceeding 1/3 chicken to sample nuclei). Nuclear suspensions obtained from roots of Lin- coln, P 2, and Frisson peas were, stained with DAPI at saturating (5.6, 28, and 56~tM) and non-saturating (0.14, 2.8, and 5.6gM) concentrations, respectively, for 30rain before FCM analysis.
DNase I digestions
Nuclei of control and mycorrhizal roots of the Lincoln cultivar were subjected to DNase I digestion. For comparison, nuclei of embryo roots of germinating seeds (after 15h seed imbibition) were also subjected to DNase I digestion following the same protocol. Nuclei were released from control, mycorrhizal and embryo roots in a TMN digestion buffer (10mM Tris, 5raM MgC12, 10raM NaC1, pH 7.4) similar to that used by Roti Roti etat. (1985), but without phenyl- methylsulfonylfluoride. 2 lal DNase I solution (23 Kunitz units/gI, Boehringer, Mannheim, Federal Republic of Germany) was added to each milliliter (3 x t05 nuclei) of sample, for various lengths of time at 25 ~ At each subsequent time, 0.I mI of stock staining solution (56 ~tM DAPI and 100raM EDTA in TMN buffer, to stop DNA digestion) was added to 0.9 ml of the digested nuclear sus- pensions, mixed and immediately analysed in the flow cytometer.
Flow eytometry
The Partec PAS II system (Sonnenweg, Switzerland) was equipped with a HBO 100 W/2 are mercury lamp. UV excitation employed a UG 1 filter and TK420 dichroic mirror. DAPI fluorescence was detected using a K435 barrier filter. The data acquisition system from the same company was used to store and print data.
Results
Saturating and non-saturating D A P I staining
Mycorrhizat roots of Fr isson and Lincoln plants were
highly infected (77% and 71%, respectively), while
roots of mutants showed only appressoria formation.
The sa tura t ion curves of the D N A - D A P I complex of
nuclei released f rom non-mycor rh iza l roots of Lincoln,
Frisson, and P 2 pea genotypes are shown in Fig. t.
The b ind ing characteristics of D A P I to D N A varied
between the different cultivars, so that dye concentra-
t ions 10-40 times lower than the corresponding satu-
ra t ing values were used for the chromat in structure
analysis.

S. Sgorbati et al.: Chromatin in arbuscular mycorrhizas of pea 3
200 -
o ~ 150-
111 ... . . . . 2c ,.,,coL'N'
1oo-
"~ z 50"
0 200 400
Relative Fluorescence
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . i ~ . . . . . i" 14 28 42 d 6
DAPI Concentrat ion (,uM)
Fig. 1. Relative fluorescence (channel number) of 2C nuclei released from uninfected (control) roots of Lincoln (El), Frisson (~) , and P2 (O) pea genotypes plotted as function of DAPI concentration (~tM). Inset Example of a flow cytometric histogram of 2 C and 4 C nuclei released from control' roots of Lincoln and stained at the saturating DAPI concentration (5.6 gM). About 20,000 nuclei were analyzed per histogram. Nuclei with a 4 C DNA content have about double the fluorescence intensity of 2 C nuclei, The relative fluorescence of 2 C peak nuclei at different DAPI concentrations was taken for the saturation curves
" ' ' t F.,.so. 800- 2C 2C CONTROL I~ CONTROL CONTROL
6 0 0 ,
E, 200- "6
8 0 0 -
Z
6 0 0 -
CRBCs
2 C
4C
4C 4C
INOCULATED I C 2 i INOCULATED II ~ INOCULATED
I--C= 8% RBCs I - C = 12% 2C I-- C "" 10%
4 0 0 -
2 C
,oo.~ll 1 ILl �9 "? I i l ,c 4 C
0 100 2o0 3o0 40o 0 100 20o 300 4o0 0 100 2o0 30o 4oo
Fluorescence intensity of DAPI (sa tura t ing)
Fig. 2. Fluorescence intensity distribution of nuclei released from uninfected (controls; top) and mycorrhiza-inoculated (bottom) roots of Lincoln, Frisson, and P 2 pea genotypes. Nuclei were stained at saturating (5.6, 56, and 28 gM, respectively) DAPI concentrations for 30 rain before FCM analysis. Chicken red blood cells (CRBCs) were added as an internal biological standard. The percent increase in fluorescence (relative to the internal standard) of 2 C peak nuclei of inoculated compared to control roots (I-C) is indicated

4 S. Sgorbati etal.: Chromatin in arbuscular mycorrhizas of pea
.~176 I 6001
400/CRBCs
ti 200 2C
"~ O J .......
" 800- E Z
6 0 0 -
400- ,RBCs
200-
0
LINCOLN
CONTROL
FRISSON i 2C P2 CONTROL ; =~ CONTROL
2C I I.
4 C 4C
4C
INOCULATED
I --C = 30%
,CRBCs INOCULATED
I--C=39%
2C
A 4C
100 260 "l . . . . 300 4(~0 0 100 200 300 400
I C i C INOCULATED
I--C=20%
RBCs~ 4C
0 100 200 300 400
F l u o r e s c e n c e i n t e n s i t y of DAP1 ( u n d e r s a t u r a t i n g )
Fig. 3. Fluorescence intensity distribution of nuclei released from uninfected (controls; top) and mycorrhiza-inoculated (bottom) roots of Lincoln, Frisson, and P 2 pea genotypes. Nuclei were stained at non-saturating (0.14, 5.6, and 2.8 gM, respectively) DAPI concentrations for 30rain before FCM analysis. Chicken red blood cells (CRBCs) were added as an internal biological standard. The percent increase of fluorescence (relative to the internal standard) of 2 C peak nuclei of mycorrhizal compared to control roots (I-C) is indicated
~o 200 ~J
ul
2 150 LL O~
.>_
n" 100
50
@ _ Controllt .~Mycorrhizal 1 ~ Embryo root '~ 200 :'c 0 mln 0 min I~ 2c 0 rain
= .., . . . . ,~k. t ~ i ~ 4c
oo,. Z 200 ~" Ill 132r l, 2c oo , ik Ik
\ 0 , ,~,mmm,,l_~ . . . . . , _ _ ~ ' ~ 0 200 400 0 v 200 400 0 200 400
I I I I - 1 . . . . . . . . 5 10 115 20 30 D i g e s t i o n t i m e ( ra in )
Fig, 4. Digestion kinetics of nuclear DNA of Lincoln pea roots with DNase I at dif- ferent times. The average fluorescence in- tensity of 2C nuclei of control (D) and mycorrhizal (0) roots was plotted along the ordinate, Nuclei released from pea em- bryo roots (~) after 15h of seed imbi- bition were also subjected to DNase I digestion following the same protocol. The initial digestion rate was much faster in mycorrhizal and embryo nuclei than in controls. After 30 min the digestion limit of mycorrhizal nuclear DNA (25.5% of the initial fluorescence) was about 34% lower than that of control roots (38.8%), and very close to that of germinating pea embryo nuclei. Inset Examples of DNA histograms of control, mycorrhizal, and pea embryo root nuclei at time 0 and after 30rain DNase I digestion. The mean flu- orescence intensity of the 2 C peak at dif- ferent times was taken as a reference to draw the digestion curves
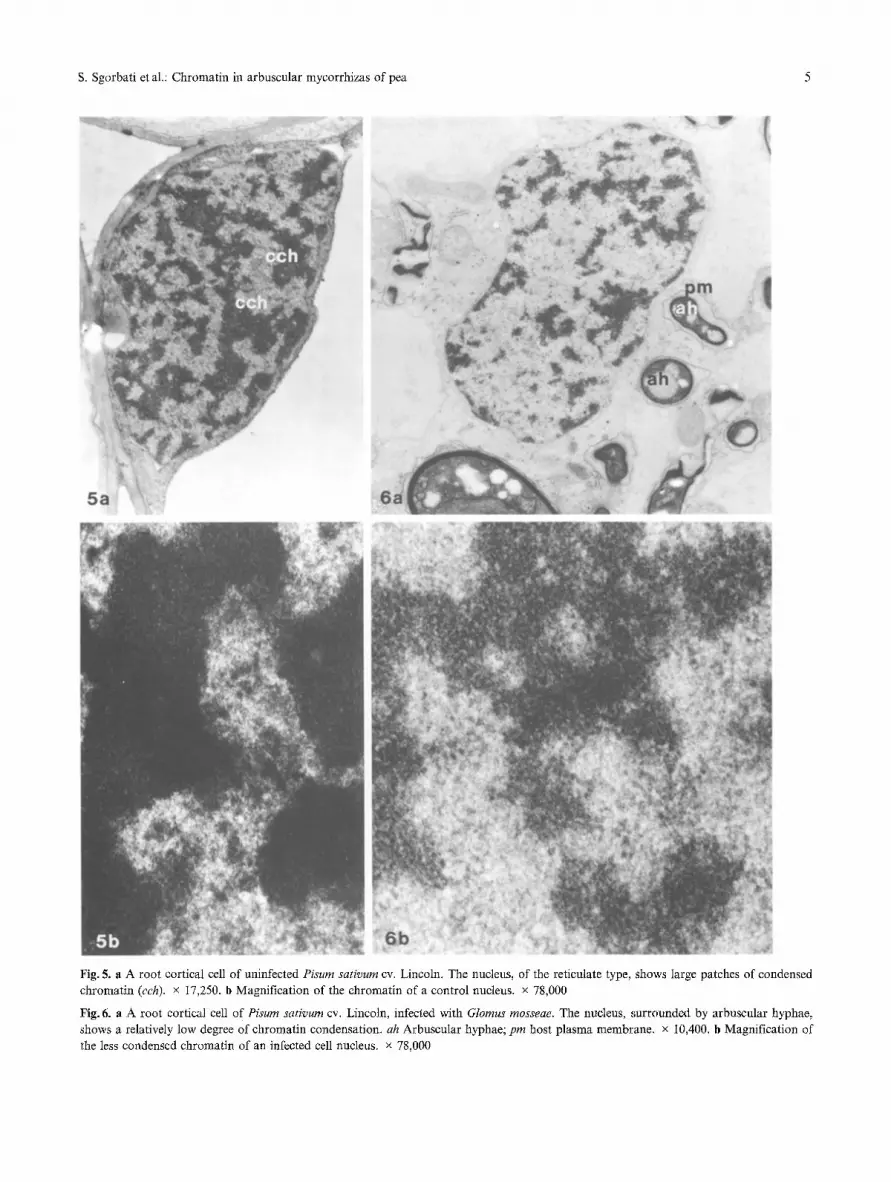
S. Sgorbati et aL: Chromatin in arbuscular mycorrhizas of pea 5
Fig. 5. a A root cortical cell of uninfected Pisum sativum cv. Lincoln. The nucleus, of the reticulate type, shows large patches of condensed chromatin (cch). x 17,250. b Magnification of the chromatin of a control nucleus, x 78,000
Fig, 6. a A root cortical cell of Pisum sativum cv. Lincoln, infected with Glomus mosseae. The nucleus, surrounded by arbuscular hyphae, shows a relatively low degree of chromatin condensation, ah Arbuscular hyphae; pin host plasma membrane, x 10,400. b Magnification of the less condensed chromatin of an infected cell nucleus, x 78,000

6 s. Sgorbati et al.: Chromatin in arbuscular mycorrhizas of pea
The staining capacity of nuclei released from roots of mycorrhiza-inoculated and control plants of the dif- ferent genotypes at saturating DAPI concentrations is shown in Fig. 2. With respect to controls, the increase in fluorescence (measured as ratio to that of the internal standard) of nuclei released from inoculated roots of Lincoln and Frisson genotypes ranged from 8 to 12%, respectively. Figure 3 shows the staining capacity of nuclei released from mycorrhiza-inoculated and control roots of the different genotypes at non-saturating DAPI concen- trations. An increase in fluorescence (measured as a ratio to the internal standard) of about 30 and 39%, compared to controls, was observed in the mycorrhizal Lincoln and Frisson genotypes, respectively. Interest- ingly, the increase in fluorescence over the control of 2 C nuclei of P 2 inoculated roots, which because of their genetic resistance to AM fungi block the infection at appressorium formation, is considerably lower (20%) than that observed in the Frisson wild genotype susceptible plants (39%).
DNase I digestion
Sensitivity of active genes to DNase I seems to be a general phenomenon, reflecting the altered confor- mation of nucleosome particles (Weintraub and Grou- dine 1976, Gross and Garrad 1988). Figure 4 shows the digestion curves of nuclei from control and mycorrhizaI roots of the Lincoln cultivar, characterized by a marked decrease in DNA fluorescence. The initial decrease was faster in mycorrhizal than in control roots, reaching digestion limits of about 25 and 39% of the initial fluorescence, respectively, after 30 rain of digestion. In the same figure, the digestion curve of nuclei released from mycorrhizal roots was very close to that obtained by digestion of nuclei released from embryo roots of the same pea cultivar during seed germination, when the DNA of meristematic cells is thought to be actively transcribing (Grellet et al. 1977, Caers et al. 1979). The inset shows the corresponding DNA histograms at time 0 and alter 30 rain DNase I digestion. During the diges- tion time the histograms maintained their normal shape, with an unchanged fluorescence ratio between 4 C and 2 C peaks.
Electron microscopy
Ultrastructural observations of root cortical cell nuclei of P. sativum cv. Lincoln showed a chromonematic or reticulate type structure, with large patches of con- densed chromatin (Fig. 5 a, b). Nuclei of infected roots,
however, were in a less condensed state, with smaller zones of condensed chromatin interspersed with wide zones of diffuse chromatin, a structure that could there- fore be defined as chromomeric-chromonematic or semi-reticulate (Fig. 6 a, b).
Discussion
The modes of interaction of fluorochromes with DNA are various, including external binding and intercala- tion (Latt and Langlois 1990). For these reasons, DNA fluorochromes can be used as probes of chromatin structure using microfluorimetry and flow cytometry (Coweden and Curtis 1981, Darzynkiewicz 1990). We found non-saturating concentrations of DAPI partic- ularly useful for probing chromatin structure variation which accompanies size and ultrastructural changes in nuclei released from leek roots after AM infection (Berta et al. 1990). In this study, the approach was extended to two cup tivars (Lincoln and Frisson) of a dicotyledon, P. sa- tivum and a P 2 isogenic mutant of cv. Frisson. The saturation kinetics of root nuclei of these plants with DAPI staining were quite different and may reflect the developmental stage of root and cell differentiation. The increase in nuclear fluorescence of Lincoln and Frisson pea cultivars one month after AM inoculation, compared to controls, was evident at saturating DAPI concentrations. The accessibility of DNA to DAPI at staining equilibrium was previously reported to be sig- nificantly increased in exponentially growing compared to differentiated animN cells (Darzynkiewicz etal. 1984). In our endomycorrhizal system, it cannot be excludext that the different DNA staining capacity at saturating DAPI concentration was enhanced by- for- maldehyde fixation. This is reported to alter DNA staining with different fluorochromes in relation to chromatin structure (Larsen etal. 1986). Increase in nuclear DNA fluorescence observed in inoculated roots can be explained by variation in chromatin structure, confirmed by analysis at non-saturating DAPI con- centrations and (for the Lincoln cultivar) electron mi- croscopy. However, even if polyploidization can be excluded (Berta etal. 1986, 1990; Blair and Peterson 1988), quantitative differences in DNA content, i.e., DNA amplification, cannot be completely ruled out. Moreover, DNA amplification is very frequent in plants at different developmental phases, especially un- der environmental stress (Nagl 1992) and/or during cell differentiation (Bassi 1990). Biochemical and cytoge- netic data, together with molecular biology techniques,

s. Sgorbati et a1.: Chromatin in arbuscular mycorrhizas of pea 7
could clarify whether there is variation in chromatin
structure only, or if DNA amplification is also occur- ring in these endomycorrhizal systems. At non-saturating DAPI concentration (Fig. 3), the in- crease in nuclear fluorescence of mycorrhizal Lincoln and Frisson cultivars (30 and 39% compared to con- trols, respectively) was much higher than at saturating DAPI concentrations, indicating variation in chro- matin structure of the host nuclei and confirming that reported previously from AM of leek (Berta et al. 1990), In addition, at least for the Lincoln cuttivar, there is ultrastructural evidence that variation in chromatin structure is induced by the fungal symbiont in pea mycorrhizae. All these observations suggest that this kind of response, although differing in intensity, could be a normal host plant reaction to fungal infection in endomycorrhizal systems. However, the DNA staining capacity of host nuclei seems to be related to the degree of infection and the development of arbuscules in the cortical parenchyma cells (Berta et N. 1989, Balestrini et al. i992). In fact, in AM resistant P 2 plants, where the presence of the fungus is limited to the surface of root epidermal cells (Duc et al. 1989), increase in DNA staining was considerably lower (20%) than in Frisson (39 %) whose cortical parenchyma cells were, contrast- ingly, highly infected. Although the increase in DNA staining capacity of nuclei in infected roots corresponds to increased ac- cessibility of chromatin to fluorochromes, in relation to its more dispersed structure (Berta etal. 1990), it does not itself demonstrate higher transcriptional ac- tivity. On the other hand, it is known that DNA sen- sitivity to nuclease digestion is well correlated with the transcriptional potential of genes (Weisbrod 1982, Zared and Yamamoto 1984). The DNase I digestion of nuclei released from mycorrhizal roots of the Lincoln cultivar reveals a dramatic increase in sensitivity to the enzyme relative to the controls, both as rate of digestion and digestion limit. The DNA sensitivity of nuclei of mycorrhizal roots is also very close to that of nuclei released from pea embryo roots during germination, when cells are considered transcriptionally very active (Grellet et al. 1977). Work is in progress to elucidate the role of histories and other chromosomal proteins in sustaining the exceptional transcriptional activity of host cell chromatin. The possibility that AM-infected cells are transcriptionally much more active than con- trol cells is indirectly supported by ultrastructural data (see, e.g., Toth and Miller 1984). In addition, some evidence exists that modifications in host gene expres- sion occur during endomycorrhizal infection, since new
polypeptides or proteins (endomycorrhizins) have been reported in infected roots of tobacco, soybean (Dumas et al. 1990, Wyss et al. 1990) and pea cv. Frisson, which are not detected in the pea mutant resistant to AM infection (Schetlenbaum et al. 1992). In conclusion, we are convinced that AM mycorrhizae could provide an interesting model to study chromatin structure variation and function accompanying root differentiation due to the host-fungus interaction, and that FCM analysis could be a rapid and precise tool for assessing the functional condition of the infected roots.
A c k n o w l e d g e m e n t s
This work has been partially supported by Italian MURST (60%) and CNR, INRA (France) and Bridge (EEC) grants.
R e f e r e n c e s
Balestrini R, Berta G, Bonfante P (1992) The plant nucleus in my- corrhizal roots: positional and structural modifications. Biol Cell 75:235-243
Bassi P (1990) Quantitative variations of nuclear DNA during plant development: a critical analysis. Biol Rev 65:185-225
Berta G, Fusconi A, Sgorbati S, Trotta A, Scannerini S (1986) Pre- liminary results on the ploidy and fine structure of the nuclei of the host cells in a VA mycorrhiza. Giorn Bot Ital I20:84-86
- Sgorbati S, Trotta A, Fusconi A, Scannerini S (1989) Correlations between host-endophyte interactions and structural changes in host cell chromatin in a VA mycorrhiza. In: Nardon P, Gian- inazzi-Pearson V, Grenier AM, Margulis L, Smith DC (eds) Endocytobiology IV. INRA Press, Paris, pp 153-157
- Sgorbati S, Soler V, Fusconi A, Trotta A, Citterio A, Bottone MG, Sparvoli E, Scannerini S (1990) Variations in chromatin structure in host nuclei of a vesicular arbuscular mycorrhiza. New Phytol 114:199-205
Blair DA, Peterson RL, Bowley SR (1988) Nuclear DNA content in root cells of Lotus and Trifoliurn colonized by the VAM fungus, Glomus versiforme. New Phytol 109:167-170
Bonaly J, Bre MH, Lefort-Tran M, Mestre JC (1987) A flow cy- tometric study of DNA staining in situ in exponentially growing and stationary Euglena gracilis. Cytometry 8:42--45
Caers LI, Peumans WJ, Carlier AR (1979) Preformed and newly s~athesized messenger RNA in germinating wheat embryos. Planta 144:491-496
Cowden RR, Curtis SK (1981) Microfluorimetric investigation of chromatin structure I. Evaluation of nine DNA-specific fluo- rochromes as probes of chromatin organization. Histochemistry 72:1 t-23
DarzTnkiewicz Z (1990) Probing nuclear chromatin by flow cyto- metry. In: Melamed MR, Lindmo T, Mendelsohn ML (eds) Flow cytometry and sorting, 2nd edn. Wiley-Liss, New York, pp 315- 340 Traganos F, Kapuscinski J, Staiano-Coico L, Melamed MR (1984) Accessibility of DNA in situ to various fluorochromes: relationship to chromatin changes during erythroid differentia- tion of Friend leukemia cells. Cytometry 5:355-363
Duc G, Trouvelot A, Gianinazzi-Pearson V, Gianinazzi S (1989) First report of non-mycorrhizal plant mutants (Myc-) obtained

8 S. Sgorbati et al.: Chromatin in arbuscular mycorrhizas of pea
in pea (Pisum sativum L.) and fababean (Vicia faba L.). Plant Sci 60:215-222
Dumas E, Tahiri-Alaoui A, Gianinazzi S., Gianinazzi-Pearson V (1990) Observations on modifications in gene expression with VA endomycorrhiza development in tobacco: qualitative and quantitative changes in protein profiles. In: Nardon P, Gianin- azzi-Pearson V, Grenier AM, Margulis L, Smith DC (eds) En- docytobiology IV. INRA Press, Paris, pp 153-157
Evenson D, Darzynkiewicz Z, Jost L, Janca F, Ballachey B (1986) Changes in accessibility of DNA to various fluorochromes during spermatogenesis. Cytometry 7:45-53
Giovannetti M, Mosse B (1980) An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol 84:489-500
Grellet F, Delseny M, Guitton Y (1977) Histone content of ger- minating pea embryo chromatin decreases as DNA replicates. Nature 267:724-726
Gross DS, Garrard WT (1988) Nuclease hypersensitive sites in chro- matin. Annu Rev Biochem 57:159-197
Larsen JK, Munch-Peterson B, Christiansen J Jorgensen K (1986) Flow cytometric discrimination of mitotic ceils: resolution of M, as well as G 1, S, and G2 phase nuclei with mitramycin, pro- pidium iodide, and ethidium bromide after fixation with form- aldehyde. Cytometry 7:54-63
Latt SA, Langlois RG (1990) Fluorescent probes of DNA micro- structure and DNA synthesis. In: Melamed MR, Lindmo T, Mendelsohn ML (eds) Flow cytometry and sorting, 2nd edn. Wiley-Liss, New York, pp 249-290
Mazzini G, Giordano P, Riccardi A, Montecucco CM (1983) A flow cytometric study of the propidium iodide staining kinetics of human leukocytes and its relationship with chromatin structure. Cytometry 3:443-448
Nagl W (1985) Chromatin organization and control of gene activity. Int Rev Cytol 94:21-56
- (1992) I. Replication. In: Behnke HD, Esser K, Kubitzki K, Runge M, Ziegler H (eds) Progress in botany, vo153. Springer, Berlin Heidelberg New York Tokyo, pp 166-180
Pellicciari C, Marchese G, Bottone MG, Manfredi Romanini MG (1987) Kinetics of DNase I digestion of interphase chromatin in
differentiated cell nuclei of the mouse: a flow cytometric study. Basic Appl Histochem 31:501-509
Phillips JM, Hayman DS (1970) Improved procedures for clearing roots and staining parasitic and vesicular arbuscular fungi for rapid assessment of infection. Trans Br Mycol Soc 55:158-161
Prentice DA, Tobey RA, Gurley LR (1985) Cell cycle variations in chromatin structure detected by DNase I. Exp Cell Res 157: 242- 252
Roti Roti JL, Wright WD, Higashikubo R, Dethlefsen LA (1985) DNase I sensitivity of nuclear DNA measured by flow cytometry. Cytometry 6:101-108
Schellenbaum L, Gianinazzi S, Gianinazzi-Pearson V (1993) Com- parison of acid soluble protein synthesis in roots of endomy- corrhizal wild type Pisum sativum and corresponding isogenic mutants. J Plant Physiol 141:2-6
Sgorbati S, Levi M, Sparvoli E, Trezzi F, Lucchini G (1986) Cy- tometry and flow cytometry of 4',6-diamidino-2-phenylindole (DAPI)-stained suspensions of nuclei released from fresh and fixed tissues of plants. Physiol Plant 68:471-476
Slatter RE, Gray JC (1991) Chromatin structure of plant genes. Oxford Surv Plant Mol Cell Biol 7:115-142
Spurr AR (1969) A low viscosity epoxy resin embedding medium for electron microscopy. J Ultrastruct Res 26:31-43
Toth R, Miller RM (1984) Dynamics of arbuscule development and degeneration in a Zea mays mycorrhiza. Amer J Bot 71: 449- 460
van Holde KE (1989) Chromatin structure and transcription. In: Rich A (ed) Chromatin. Springer, Berlin Heidelberg New York Tokyo, pp 355-408
Weintraub H, Groudine M (1976) Chromosomal subunits in active genes have an altered conformation. Science 193:848-856
Weisbrod S (1982) Active chromatin. Nature 297:289-295 Wyss P, Mellor RB, Wiemken A (1990) Vesicular-arbuscular my-
corrhizas of wild-type soybean and non-nodulating mutants with Glomus mosseae contain symbiosis-specific polypeptides (my- corrhizins), immunologically cross-reactive with nodulins. Planta 182:22-26
Zared KS, Yamamoto KR (1984) Reversible and persistent changes in chromatin structure accompany activation of a glucocorticoid dependent enhancer element. Cell 38:29-38





























