Embed Size (px)
Citation preview

Heredrtas 113: 277-2x9 (19901
Chromosomal localization and copy number of 18s + 28s ribosomal RNA genes in evolutionarily diverse mosquitoes (Diptera, Culicidae) A. KUMAR and KARAMJIT S. RAI
Department of Biological Sciences, University of Notre Dame, Notre Dame, Indiana 46556, U.S.A.
KUMAR, A. and RAI, K. S. 1990. Chromosomal localization and copy number of 18s + 28s ribosomal RNA genes in evolutionarily diverse mosquitoes (Diptera, Culicidae). - Hereditus 113: 277-289. Lund, Sweden. ISSN 0018-0661. Received June 25, 1990. Accepted October 17, 1990
In situ hybridization using 3H-labeled 18s and 28s ribosomal DNA @DNA) probes from Aedes albopictus was performed on the mitotic chromosomes of 20 species of mosquitoes belonging to 8 genera of subfami- lies Culicinae and Anophelinae. In all but one species examined, the rDNA family was localized to a single chromosome per haploid genome. Aedes triseriatus was the only exception, with the rDNA cistrons present on chromosome 1 and on chromosome 3. The ribosomal RNA genes were located on chromosome I in Ae. albopictus, Ae. aegypti, Ae. fzavopictus, Ae. seatoi, A. polynesiensis, Ae. alcasidi, Ae. annandalei. Ae. mascarensis, Ae. hendersoni, Ae. atropalpus, Ae. epactius, Culex pipiens quinquefasciatus. Wyeomyia smithii, and Sabethes cyaneus; chromosome 2 in Ae. mediovittatus and Haemugogus equinus; chromosome 3 in Armigeres subalbatus and Tripteroides bambusa; and the heteromorphic X and Y chromosomes in Anopheles quadrimaculatus. The variation in the location of ribosomal RNA genes on the different chromosomes and at different positions on the chromosome arm among the mosquito species examined is suggestive of considerable chromosome repatterning through translocations and inversions in the karyo- typic evolution of mosquitoes. Dot-blot hybridization was used to estimate copy number of rRNA genes; the copy number per haploid genome ranged from 39 3.27 in So. cyaneus to 1023 f 68.14 in Ae. flavopictus.
A . Kumar, Department of Biological Sciences, University of Notre Dame, Notre Dame, Indiana 46556, USA.
The ribosomal RNA gene family is composed of multiple tandemly repeated units. Each unit con- tains an intergenic ‘nontranscribed’ spacer (IGS, formerly NTS) and a transcribed portion consisting of an external transcribed spacer (ETS), 18s rRNA gene, internal transcribed spacer (ITS) containing the 5.8s rRNA gene, and 28s rRNA gene (BECKING- HAM 1982). The copy number of these units ranges from 45 per haploid genome in Sciara coprophila (GEnBI and CnousE 1976) to over 3000 in the grass- hopper, Locusta migratoria (OISHI et al. 1985). RI- TOSSA and SPIEGELMAN (1965) were the first to es- tablish a simple relationship between the number of nucleolus organizer regions (NORs) (secondary constrictions) and the number of sites of ribosomal gene cluster coding for 18s and 28s ribosomal RNA molecules in Drosophila melanogaster. In general, this relationship has been found to be con- sistent throughout eukaryotic genomes (PARDUE et al. 1970; PARDUE 1974; HENDERSON et al. 1972, 1974a, b; EVANS et al. 1974; PARDUE and Hsu 1975;
Hsu et al. 1975; SCHAFER and SCHAFER 1980; VITELLI et al. 1982).
The mosquito family Culicidae is divided into three subfamilies: Toxorhynchitinae, Culicinae and Anophelinae. Based on their cytological studies on four nematocerous families, RAO and RAI (1987a) proposed that the Mochlonyx-like ancestor of the family Chaoboridae with 2 n = 8 may be the an- cestral stock from which Culicidae evolved. They also proposed that two main evolutionary lines arose from this stock-one that led to Anophelinae through Chagasia-like ancestor with 2n = 8, and the other splitting into two independent lines leading to Toxorhynchitinae and Culicinae. Little is known on the phylogenetic relationships between different genera or taxonomic groupings of these subfamilies (KNIGHT and STONE 1977; RAO 1985). These three subfamilies together include 34 genera and approxi- mately 3100 species. The karyotypes of about 200 species are known to date (WHITE 1980). The dip- loid number of chromosomes of all but one species

7uh/e i Taxonom) and source of genera and species examined ___I .___.
Subfamil? T r i k Genus Subsenus Species Strain (year of collection), source G'
Culicinae Aedini A edc Sr~qomrin urg\prr Rockefeller Inst.. USA (1959). 74 __.~._____.______
D. W. Jenkins: Memphis. USA (1986). S. Jones
Malaysia (1973). B. Knudson 45 frai opicrus Nagasaki. Japan (1988). Y. Eshita 5 sea I1 11 Bangkok, Thailand (1972), D. Gould 43 pfyircsreiisis Suva. Fiji (1982). D. Gubler 19 ulcusrdi Taiwan (1981). J. Lien 21 atiiiairdulei Taiwan (1963, G. A. H. McClelland 60 mu.wurensr .r Mauritius (1965). R. Marnet 60
Pr-oroniar~leciju rr-iser-iarits St. Joseph Co., USA (19691, R. Beach; 50 2
(1990). D. Wesson Iiender-soil I St. Joseph Co., USA (1981). S. Paulson; 21
Eddy St., St. Joseph Co.. USA (1990). 2 S. Hanson
9 alhoprcrirs Mauritius (1974). C. Courtois; 46
40 Oahu. Hawaii (1971). L. Rosen;
Potato Creek, St. Joseph Co., USA
Ochler~~rari~s arropa1pu.r Coosa. USA (1972), R. Brust 43
Gymnonietopu medioi.irtatus Puerto Rico (1981) R. Novak 21 epar'trus Sansal, El Salvador (1968). S. Breeland 53
Sabethini Tripteroides Tripteroides hambusa Kyushu Island, Japan (1987), W. Hawley 36 Sahethes Sahethes cxanrus Balboa heights, Panama (1983), 52
J. L. Peterson Wyeompia Wjeomyia smithii UNDERC* (1989) 0
Culicini Culer Culer pipiens quinquefasciarus Gainesville, USA (l969), R. Lowe 125 Aedini At-nii,qere> Arnrigeres suhaibarrrs Japan (1971). M. Sasa 1 I4
Huema,qo,qrrs Hacmagogits eqitinus Panama (1979). J. L. Peterson 26 Anophelinae Anripheler Anoplirles qiradrimacrrlariis Savannah, USA (1971), H. Schoof 40
* Approximate generations in colon) * L'nivervty of Notre Dame Environmental Re\earch Center at Land O'Lakes. Wirconsin
is 6 (2n = 6). The only exception, Chagasia bathana of the subfamily Anophelinae, possesses 4 pairs of chromosomes (KREUTZER 1978). In general, species of the subfamilies Toxorhynchitinae and Culicinae demonstrate a considerable uniformity with respect to their karyotypes. A typical karyotype comprises 3 pairs of homomorphic chromosomes: a pair of small chromosomes (chromosome I ) , a pair of large chromosomes (chromosome 2) and a pair of inter- mediate-sized chromosomes (chromosome 3) (RAI 1963, 1966; WHITE 1980: RAO and RAI 1987a). In a karyotype, all three pairs of chromosomes may be metacentric, or one or two chromosome pairs may be slightly submetacentric (RAI 1963, 1966:
RAO and RAI 1987a). The lengths of all three pairs of chromosomes often vary in different species (RAI 1963. 1966; BAKER and ASLAMKHAN 1969: R ~ ~ a n d RAI 1987a). In contrast, the species of the subfami- lies Anophelinae possess 2 pairs of metacentric chromosomes of unequal size and one pair of het- eromorphic sex chromosomes, X and Y (KITZ- MILLF;R 1967). Using standard chromosome tech-
WW1T.E 1980: MUNSTERMANN and MARCHI 1986:
niques, the mosquito karyotypes do not demonstrate any linear differentiation for NORs along the length of chromosomes. A different approach, using silver staining technique of GOODPASTURE and BLOOM (1975), has also failed to locate ribosomal RNA genes on the mitotic mosquito chromosomes (RAO and RAI 1987a).
In situ hybridization has been extensively used in the localization of ribosomal RNA gene cluster on the chromosomes of both animals and plants (PARDUE et al. 1970; HENDERSON et al. 1972, 1974a, b; EVANS et al. 1974; PARDUE and Hsu 1975: Hsu et al. 1975; SCHAFER and SCHAWR 1980; VITELLI et al. 1982: DELUCCHINI et al. 1988; VISSER et al. 1988). In insects such as Drosophila and the grass- hoppers, in situ hybridization has been used to loc- ate the rRNA genes on their chromosomes (SPEAR 1974; HENNIG et al. 1975; SCHAFER and SCHAFER 1980: SINIBALDI and CUMMINGS 1981; WHITE et al. 1982). However, in situ hybridization has not so far been used to locate the ribosomal RNA genes on the chromosomes of mosquitoes. In this paper we report the successful in situ localization of ribo-

Hrieditus 113 llY90) 18s + zxs RIBOSOMAL R Y A G E ~ E S I N MOSQUIIOES 279
soma1 RNA genes on the mitotic chromosomes of 20 species of mosquitoes belonging to 8 genera of subfamilies Culicinae and Anophelinae. In addition, the copy number of rRNA genes has been deter- mined in 13 species, using dot-blot hybridization.
at 35°C. The slides were dehydrated in 70 % ethanol (2x10 min), 95 % ethanol (2x5 min) and air-dried. The slides were coated with Kodak NTB-2 emul- sion diluted 1:l with distilled water at 43°C and stored at 4°C for 11-25 days. After exposure, the slides were developed, fixed and stained in S 5% Giemsa (30 min) as described in PARDUE (1985).
Materials and methods Mosquito species and chromosome preparation. -The details of genera and species of mosquitoes employed during the present study are given in Table 1. All mosquito species used were from la- boratory stocks, except Wyeomyia smithii. The lar- vae of Wy. smithii were collected from the pitcher plants (Sarracenia purpurea) at the University of Notre Dame Environmental Research Center at Land O'Lakes, Wisconsin. Mitotic chromosome preparations were obtained from colchicine-treated brain ganglia of fourth instar larvae, following the method of RAO and RAI (1987a).
DNA probes and nick-translation. - Three recom- binant plasmids: pP11, pP1 and PE2 were used as DNA probes (BLACK et al. 1989). Plasmid pP l l contains the 28Sp gene plus about 800 bp of in- tergeriic spacer (IGS). Plasmid pP1 contains the complete 28Sa gene and some internal transcribed spacer (ITS) of the ribosomal cistron. Plasmid pE2 contains the complete 1.8 kb 18s gene. These re- combinant plasmids are pUC 1 18 subclones derived from a lambda EMBL3 rDNA clone from a ge- nomic library of Ae. albopictus (BLACK et al. 1989). The subcloned DNAs were purified according to MANIATIS et al. (1982). Each subcloned DNA was nick-translation with 3H-dCTP (18 Ci/mmole) and ?H-dGTP (10 Ci/mmole) using a BRL (Bethesda Research Laboratory USA) nick-translation kit ac- cording to a standard protocol of MANIATIS et d. (1982). For dot-blot hybridization, pE2 DNA was nick-translated with "P-dCTP (3000 Ci/mmole) as described above.
In situ hybridization. - Pretreatment of the slides and in situ hybridization were carried out according to PARDUE (1985). In situ hybridization was con- ducted overnight at 40°C using 1 6 ~ 1 0 ~ 4 0 ~ 1 0 ~ cpm per slide in 50 p1 hybridization buffer (~xSSC, SO % formamide, 5 % dextran sulphate, 1 % Den- hardt's solution, 0.037 pg probe DNAs, 7 pg calf- thymus DNA). The three rDNA probes (pP1, pP11 and pE2) were mixed for each in situ hybridization reaction. The non-specific bound probes were re- moved by incubating slides in 2xSSC (3x10 min)
DNA extraction and dot-blot hybridization. - DNAs were extracted from honey-fed adult female mosquitoes according to BLIN and STAFFORD ( 1976) with minor modifications as described in BRAD- FIELD and WYATT (1983). A typical yield from 10 adults was 8-15 pg DNA, depending on the size of mosquito species used.
Since the number of 18s rRNA genes equals the number of 28s genes or the ribosomal repeat units, 18s DNA was used as a standard and the nitrocel- lulose membrane was hybridized with '*P-labeled pE2 DNA. To isolate the 18s rDNA insert, plasmid pE2 (20 pg) was digested with EcoRI to excise the complete 1.8 kb 18s DNA under the conditions recommended by the supplier. The digest was run in 0.8 % agarose, and the fragments were purified from the agarose, using a Geneclean kit (Bio 101 Inc. USA).
Standard DNA and species DNA were blotted onto a nitrocellulose membrane, using a Schleicher and Schuell manifold. The nitrocellulose membrane was vacuum dried for 2 h at 80"C, and prehybrid- ized in 50 % deionized formamide, SxSSC, 0.5 R SDS, 5 % dextran sulphate, 5 x Denhardt's solution and 100 pg/ml denatured calf-thymus DNA at 42°C overnight. 32P-labeled pE2 DNA was directly added to the prehybridization mixture and the hybridiza- tion was carried out for 16-24 h at 42°C (SAMBROOK et al. 1989). Following hybridization, the membrane was washed twice in 1 x wash buffer (10 x wash buffer = 3M NaCl, 0.6 M Tris-HCI, pH 8.0, 0.02 M EDTA, pH 8.0) for 10 rnin each at room tem- perature, twice in 1 x wash buffer and 1 % SDS for 30 min each at 60°C, and finally twice in 0.01 x wash buffer for 30 min each at room temperature. The membrane was dried at room temperature, wrapped in the Saran wrapTM and exposed to Kodak X-OMAT X-ray film with a Dupont intensifying screen at -70°C for a range of time. The copy number of rRNA repeat units was determined by comparing the dot intensity of the species DNA with the appropriate standard DNA of the autora- diograms that were in linear range of the film re- sponse (ZIMMERMAN et al. 1989) using a EC- densi- tometer and a Zeineh software 1-D scanner (Fisher Scientific Inc. USA).
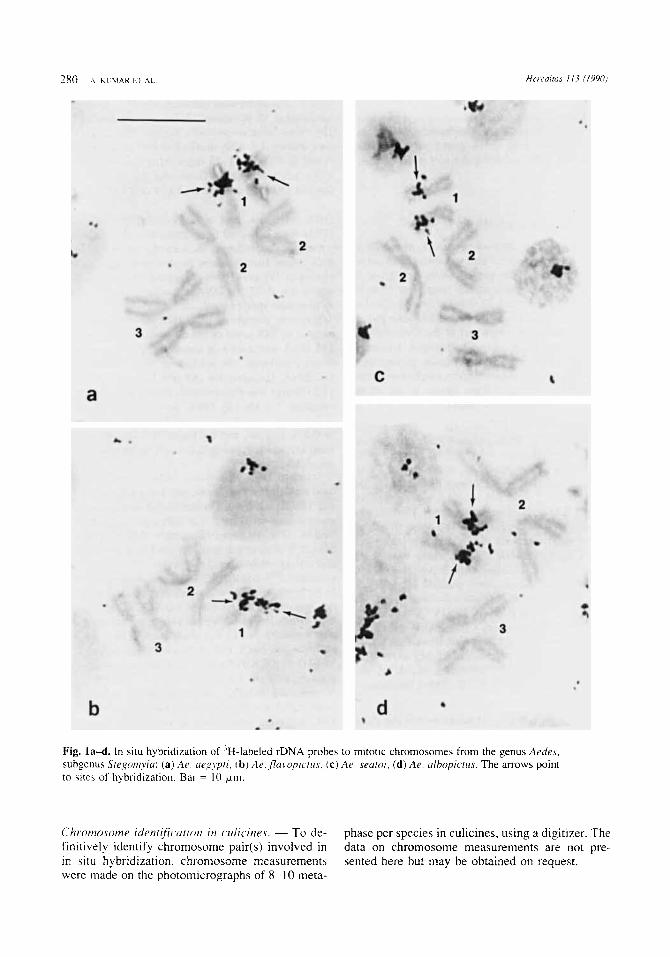
Fig. l a d . In situ hybridization of "-labeled rDNA probes to mitotic chromosomes from the genus Aede.5, subgenus Stegoniyia: (a) Ae. oexyptr, (b) Ae.,fluwpicri(.s, ( c ) A e searoi. (d) Ae. alhopictus. The arrows point to sites of hybridization. Bar = 10 pn.
Chr~onzosome ider~ificatiori i r i czrliciires. - To de- finitively identify chromosome pair(s) involved in in situ hybridization. chromosome measurements were made on the photomicrographs of 8-1 0 meta-
phase per species in culicines, using a digitizer. The data on chromosome measurements are not pre- sented here but may be obtained on request.
I X S + ?XS RIBOSOMAL RNA GENES IN MOSQUITOES 28 1
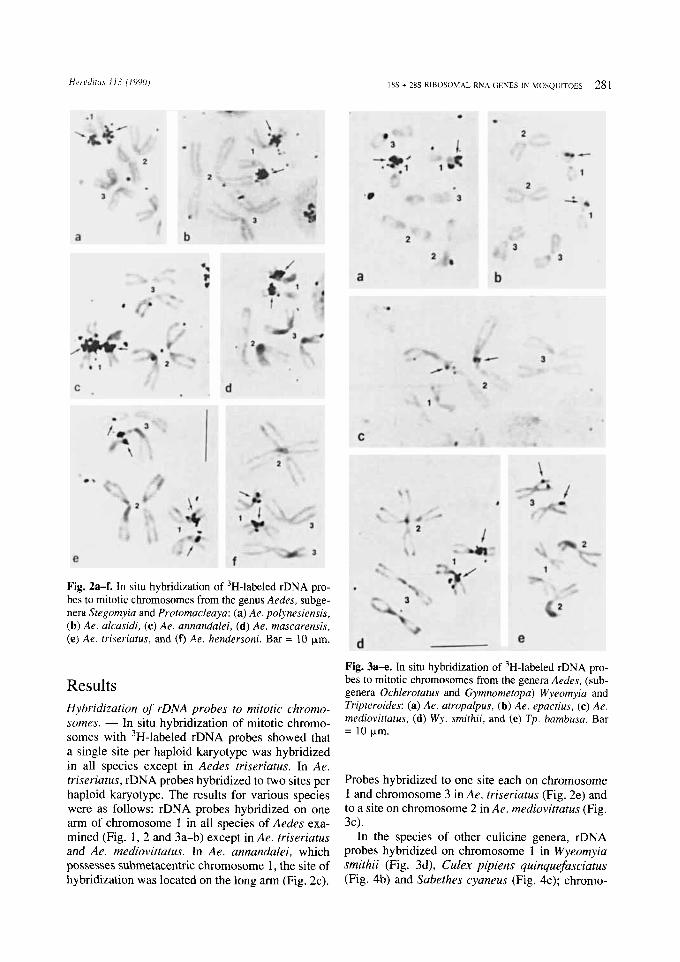
Fig. 2a-f. In situ hybridization of 3H-labeled rDNA pro- bes to mitotic chromosomes from the genus Aedes, subge- nera Stegomyia and Protomacleaya: (a) Ae. polynesiensis, (b ) Ae. alcasidi, (c) Ae. annandalei, ( d ) Ae. mascarensis, (e) Ae. triseriatus, and (f) Ae. hendersoni. Bar = 10 pm.
Results Hybridization of rDNA probes to mitotic chromo- somes. - In situ hybridization of mitotic chromo- somes with 3H-labeled rDNA probes showed that a single site per haploid karyotype was hybridized in all species except in Aedes triseriatus. In Ae. triseriatus, rDNA probes hybridized to two sites per haploid karyotype. The results for various species were as follows: rDNA probes hybridized on one arm of chromosome 1 in all species of Aedes exa- mined (Fig. 1, 2 and 3a-b) except in Ae. triseriatus and Ae. mediovittatus. In Ae. annandalei, which possesses submetacentric chromosome 1, the site of hybridization was located on the long arm (Fig. 2c).
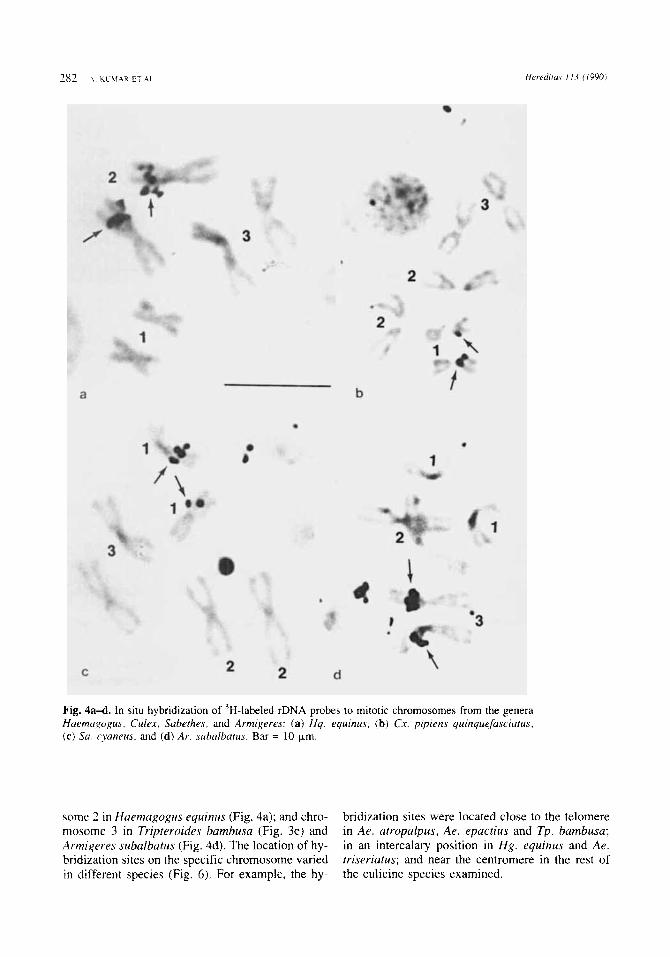
Fig. 3a-e. In situ hybridization of 3H-labeled rDNA pro- bes to mitotic chromosomes from the genera Aedes, (sub- genera Ochlerotatus and Gymnometopa) Wyeomyia and Tripteroides: (a) Ae. atropalpus, (b ) Ae. epactius, ( c ) Ae. mediovittatus, (d) W y . smithii, and (e) Tp. bambusa. Bar = 10 p m .
Probes hybridized to one site each on chromosome 1 and chromosome 3 in Ae. triseriatus (Fig. 2e) and to a site on chromosome 2 in Ae. mediovittatus (Fig.
In the species of other culicine genera, rDNA probes hybridized on chromosome 1 in Wyeomyia smithii (Fig. 3d), Culex pipiens quinquefasciatus (Fig. 4b) and Sabetkes cyaneus (Fig. 4c); chromo-
3c).
Her-rdtrus 113 (1990)
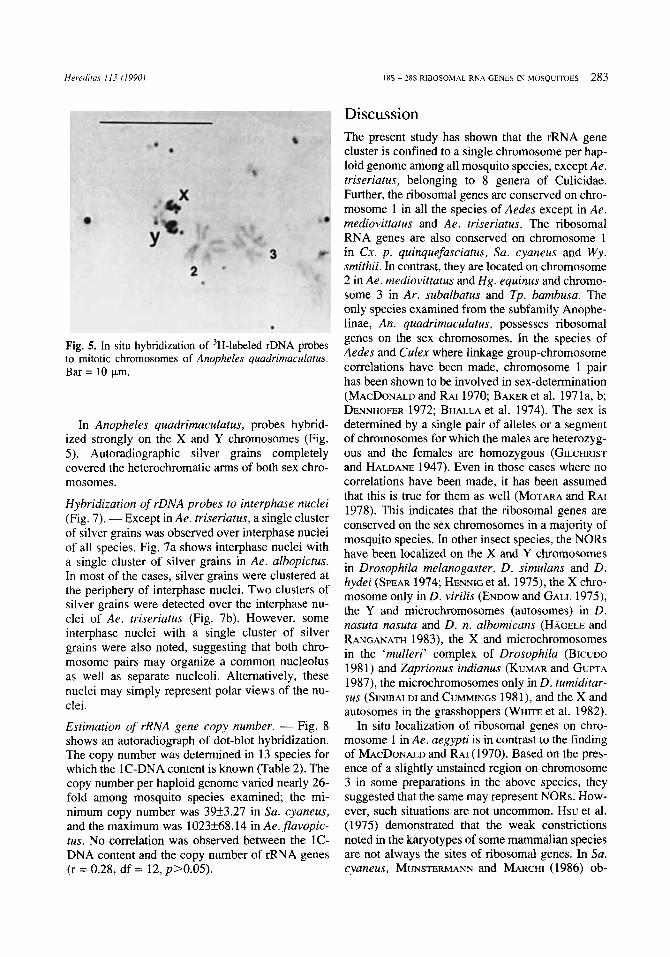
Fig. 4a-d. In situ hybridization of 3H-labeled rDNA probes to mitotic chromosomes from the genera Haemagogus. Culex, Sabethes, and Armigeres: (a) Hq. equinus, (b) C.r. pipiens quinquefasciutus, (c) Su. ryatieus, and ( d ) Ar. subalbatus. Bar = 10 +m.
some 2 in Haemagogus equinus (Fig. 4a); and chro- mosome 3 in Tripteroides bamhusa (Fig. 3e) and Armigeres slcbalbatus (Fig. 4d). The location of hy- bridization sites on the specific chromosome varied in different species (Fig. 6). For example, the hy-
bridization sites were located close to the telomere in Ae. atropalpus, Ae. epactius and Tp. bambusa; in an intercalary position in Hg. equinus and Ae. triseriatus; and near the centromere in the rest of the culicine species examined.
Heredifas 113 (19901 18s + 2RS RIBOSOMAL RNA GENES IN MOSQUITOES 283
Discussion The present study has shown that the rRNA gene cluster is confined to a single chromosome per hap- loid genome among all mosquito species, except Ae. triseriatus, belonging to 8 genera of Culicidae. Further, the ribosomal genes are conserved on chro- mosome 1 in all the species of Aedes except in Ae. mediovittatus and Ae. triseriatus. The ribosomal RNA genes are also conserved on chromosome 1 in Cx. p . quinquefasciatus, Sa. cyaneus and Wy. smithii. In contrast, they are located on chromosome 2 in Ae. mediovittatus and H g . equinus and chromo- some 3 in Ar. subalbatus and Tp. bambusa. The only species examined from the subfamily Anophe- linae, An. quadrimaculatus, possesses ribosomal genes on the sex chromosomes. In the species of Aedes and Culex where linkage group-chromosome correlations have been made, chromosome 1 pair has been shown to be involved in sex-determination ( M A C D O N A L D ~ ~ RAI 1970; BAKER^^ al. 1971a, b; DENNHOFER 1972; BHALLA et al. 1974). The sex is determined by a single pair of alleles or a segment of chromosomes for which the males are heterozyg- ous and the females are homozygous (GILCHRIST and HALDANE 1947). Even in those cases where no correlations have been made, it has been assumed that this is true for them as well (MOTARA and RAI 1978). This indicates that the ribosomal genes are conserved on the sex chromosomes in a majority of mosquito species. In other insect species, the NORs have been localized on the X and Y chromosomes in Drosophila melanogaster, D. simuhns and D . hydei (SPEAR 1974; H E N N I C ~ ~ al. 1975), the X chro- mosome only in D . virilis (ENDOW and GALL 1975), the Y and microchromosomes (autosomes) in D . nasuta nasuta and D . n. albomicans (HAGELE and RANGANATH 1983), the X and microchromosomes in the ‘mulleri’ complex of Drosophila (BICUDO 1981) and Zaprionus indianus (KUMAR and GUITA 1987), the microchromosomes only in D. tumiditar- sus (SINIBALDI and CUMMINCS 1981), and the X and autosomes in the grasshoppers (WHITE et al. 1982).
In situ localization of ribosomal genes on chro- mosome 1 in Ae. aegypti is in contrast to the finding of MACDONALD and RAI (1970). Based on the pres- ence of a slightly unstained region on chromosome 3 in some preparations in the above species, they suggested that the same may represent NORs. How- ever, such situations are not uncommon. Hsu et al. (1975) demonstrated that the weak constrictions noted in the karyotypes of some mammalian species are not always the sites of ribosomal genes. In Sa. cyaneus, MUNSTERMANN and MARCHI (1986) ob-
Fig. 5. In situ hybridization of 3H-labeled rDNA probes to mitotic chromosomes of Anopheles quadrimaculatus. Bar = 10 pm.
In Anopheles quadrimaculatus, probes hybrid- ized strongly on the X and Y chromosomes (Fig. 5). Autoradiographic silver grains completely covered the heterochromatic arms of both sex chro- mosomes.
Hybridization of rDNA probes to interphase nuclei (Fig. 7). -Except in Ae. triseriatus, a single cluster of silver grains was observed over interphase nuclei of all species. Fig. 7a shows interphase nuclei with a single cluster of silver grains in Ae. albopictus. In most of the cases, silver grains were clustered at the periphery of interphase nuclei. Two clusters of silver grains were detected over the interphase nu- clei of Ae. triseriatus (Fig. 7b). However, some interphase nuclei with a single cluster of silver grains were also noted, suggesting that both chro- mosome pairs may organize a common nucleolus as well as separate nucleoli. Alternatively, these nuclei may simply represent polar views of the nu- clei.
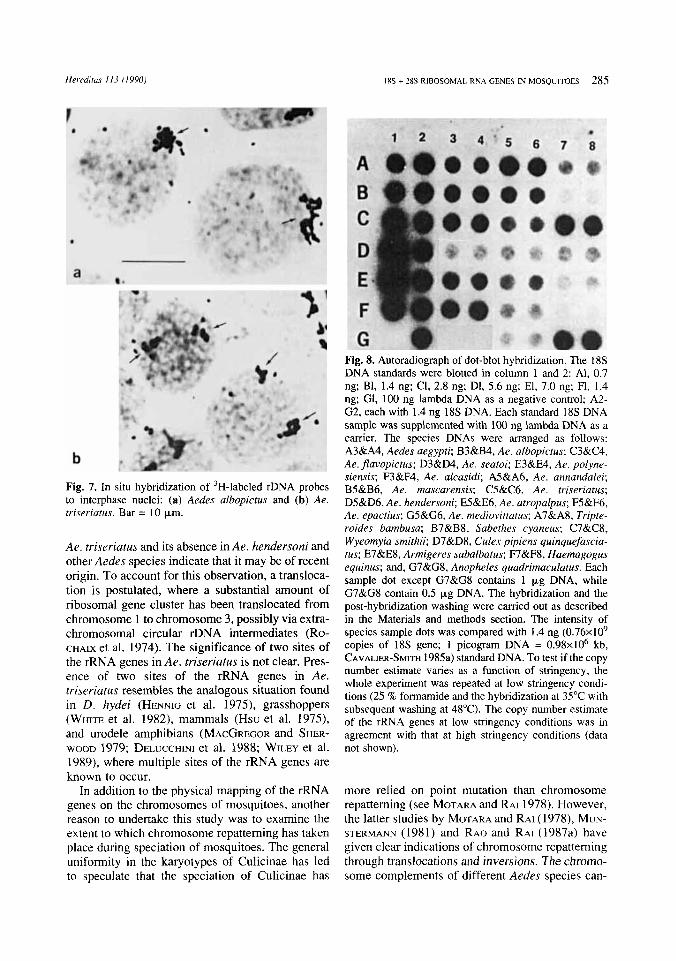
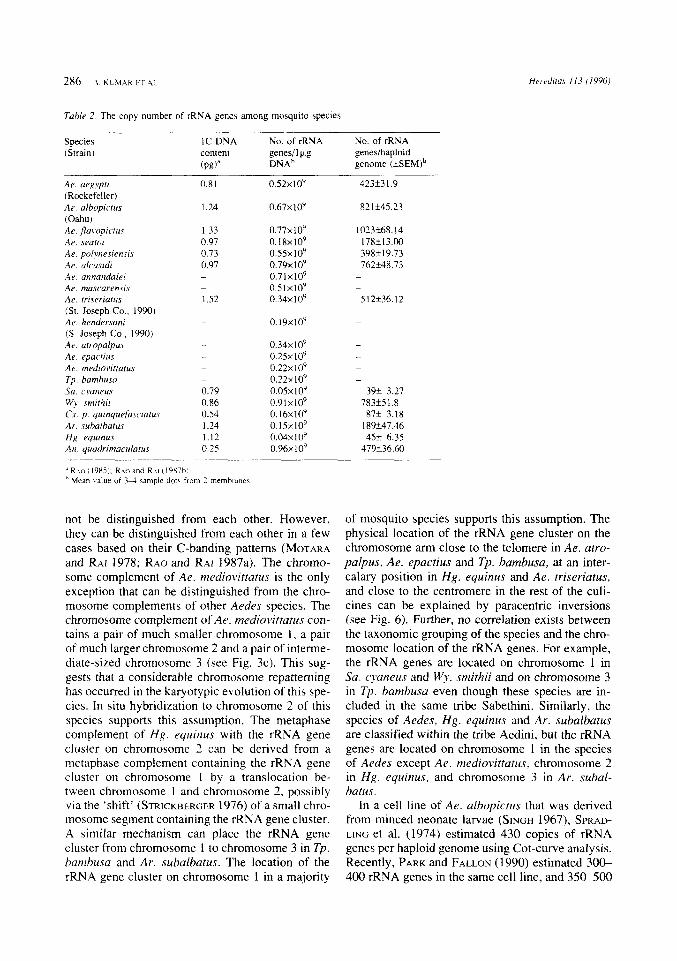
Estimation of rRNA gene copy number. - Fig. 8 shows an autoradiograph of dot-blot hybridization. The copy number was determined in 13 species for which the 1C-DNA content is known (Table 2). The copy number per haploid genome varied nearly 26- fold among mosquito species examined the mi- nimum copy number was 39f3.27 in Sa. cyaneus, and the maximum was 1023k68.14 in Ae.flavopic- tus. No correlation was observed between the 1C- DNA content and the copy number of rRNA genes (r = 0.28, df = 12, p>0.05).

CHROMOSOME 1 C H R O M O S O M E 2 CHROMOSOME 3 S P E C I E S
Aedes aegypti, Ae. albopictus, Ae. flavo pictus. Ae. seatoi, Ae. polynesiensis, Ae. alcasidi. Ae. mascarensis. Ae. hen- dersoni, Sabethes cyaneus, Wyeomyia smithii, Culex pipiens quinquefasciatus
Aedes annandalei
Aedes triseriatus
Aedes atropalpus, Ae. epactius
IXI IXI Aedes mediovittatus
Ix1 Tripteroides bambusa
Ixi Arrn/geres subalbatus
Haemagogus equinus
IXI Anopheles quadrimawlatus IXI
Fig. 6. Diagrammatic repre5entation of mitotic chromosomes of mosquito species in situ hybridized with 3H-labeled rDN.4 probes.
served recurrent association of the nucleolus with chromosome 1 in polytene nuclei. They suggested that the NORs are probably located on chromosome 1 in this species. Our result of in situ hybridization confirms their finding.
In addition to chromosome 1 . ribosomal genes are also located on chromosome 3 in Ae. rriseriurus. Sometimes i t was difficult to determine whether the large pair of chromosomes with hybridization is chroinosome 2 or chromosome 3. The difference between the lengths of these chromosomes is only 0.17 Kni (data not shown). It appeared that in some inetaphase plates hybridization sites are located on chromosome 2 and in others on chromosome 3 from the \ame individual. Since in majority of the cases, 4gnal is located o n chromosome 3. we concluded
that the second site of ribosomal gene is on chromo- some 3. Recently, DELUCCHINI et al. (1988) reported extra ribosomal sites in the genome of Triturus vul- guris meridiotiulis. These sites contain only IGS and are devoid of 18s and 28s genes. Unlike the Triturirs situation, both chromosome 1 and chromo- some 3 of Ae. [riser-iatus still hybridize to rDNA probes that lack pP11, and therefore are devoid of IGS sequences (data not shown). Thus the second site of hybridization in Ae. tt.iser-iatus appears to be a bona fide rDNA site. and not just transposed IGS sequences.
Ae. triseriuflts and Ae. henrlersoni are sibling spe- cies and crosses between them are compatible bi- directionally (TRLIMAN and CRAIG 1968). Presence of an additional site of ribosomal gene cluster in
Hereditus 113 (1990) IRS + 28s RIBOSOMAL RNA GENES IN MOSQUITOES 285
Fig. 7. In situ hybridization of 3H-labeled rDNA probes to interphase nuclei: (a) Aedes albopictus and (b) Ae. triseriatus. Bar = 10 pm.
Ae. triseriatus and its absence in Ae. hendersoni and other Aedes species indicate that it may be of recent origin. To account for this observation, a transloca- tion is postulated, where a substantial amount of ribosomal gene cluster has been translocated from chromosome 1 to chromosome 3, possibly via extra- chromosomal circular rDNA intermediates (Ro- CHAIX et al. 1974). The significance of two sites of the rRNA genes in Ae. triseriatus is not clear. Pres- ence of two sites of the rRNA genes in Ae. triseriatus resembles the analogous situation found in D. hydei (HENNIG et a]. 1975), grasshoppers (WHITE et al. 1982), mammals (Hsu et al. 1975), and urodele amphibians (MACGREGOR and SHER- WOOD 1979; DELUCCHINI et al. 1988; WILEY et al. 1989), where multiple sites of the rRNA genes are known to occur.
In addition to the physical mapping of the rRNA genes on the chromosomes of mosquitoes, another reason to undertake this study was to examine the extent to which chromosome repatteming has taken place during speciation of mosquitoes. The general uniformity in the karyotypes of Culicinae has led to speculate that the speciation of Culicinae has
Fig. 8. Autoradiograph of dot-blot hybridization. The 18s DNA standards were blotted in column 1 and 2: Al, 0.7 ng; B1, 1.4 ng; CI, 2.8 ng; D1, 5.6 ng; El, 7.0 ng; F1, 1.4 ng; GI, 100 ng lambda DNA as a negative control; A2- G2, each with 1.4 ng 18s DNA. Each standard 18s DNA sample was supplemented with 100 ng lambda DNA as a carrier. The species DNAs were arranged as follows: A3&A4, Aedes aegypti; B3&B4, Ae. albopictus; C3&C4, Ae. flavopictus; D3&D4, Ae. seatoi; E3&E4, Ae. polyne- siensis; F3&F4, Ae. alcasidi; A5&A6, Ae. annandalei; B5&B6, Ae. mascarensis; C5&C6, Ae. triseriatus; D5&D6, Ae. hendersoni; E5&E6, Ae. atropalpus; F5&F6, Ae. epactius; G5&G6, Ae. mediovittatus; A7&A8, Tripte- roides bambusa; B7&B8, Sabethes cyaneus; C7&C8, Wyeomyia smithii; D7&D8, Culex pipiens quinquefascia- tus; E7&E8, Armigeres subalbatus; F7&F8, Haemagogus equinus; and, G7&G8, Anopheles quadrimaculatus. Each sample dot except G7&G8 contains 1 p g DNA, while G7&G8 contain 0.5 pg DNA. The hybridization and the post-hybridization washing were carried out as described in the Materials and methods section. The intensity of species sample dots was compared with 1.4 ng (0.76~10’ copies of 18s gene; 1 picogram DNA = 0 . 9 8 ~ 1 0 ~ kb, CAVALIER-SMITH 1985a) standard DNA. To test if the copy number estimate varies as a function of stringency, the whole experiment was repeated at low stringency condi- tions (25 % formamide and the hybridization at 35°C with subsequent washing at 48°C). The copy number estimate of the rRNA genes at low stringency conditions was in agreement with that at high stringency conditions (data not shown).
more relied on point mutation than chromosome repatteming (see MOTARA and RAI 1978). However, the latter studies by MOTARA and RAI (1978), MUN- STERMANN (1981) and RAO and RAI (1987a) have given clear indications of chromosome repatteming through translocations and inversions. The chromo- some complements of different Aedes species can-
286 A KLMAR FT 4L Her-editus 113 ( IYYUJ
Table 2 The copy number of rRXA gene5 among mosquito species
Species (Strain)
IC-DNA content (P€)"
No. of rRNA genesilkg D N A ~
No. of rRNA geneshaploid genome ( ~ s E M ) ~
~
Ae. aegypri (Rockefeller) Ae. albopicrus (Oahu) Ae. flaropictus Ae. seator Ae. polvnesiensis Ae. ulcmidi Ae. annandaler Ae mascarensis Ae. triwr-iarus (St. Joseph Co., 1990) Ae. hendersoni ( S . Joseph Co., 1990) Ae atrnpalpus Ae. epuctius Ae. medrovirrarus Tp bamhusu So. cymeus W?. smirhii Cx. p yuinquejasciatus Ar. .suhalhatus Hg. equinus An. quadrrmaculatus
0.81
1.24
1.33 0.97 0.73 0.97 - -
I .52
-
- - - ~
0.79 0.86 0.54 1.24 1.12 0.2s
o.s2x10y
0.67~10'
0 . 7 7 ~ 1 0 ~ 0 . 1 8 ~ 10' 0.55~10' 0.79~10'
0.5 1x10' 0 . 3 4 ~ 10'
0.71x109
o.19x109
0.34~10' 0.2Sx1OY 0 . 2 2 ~ 1 0 ~ 0.22x1OY O.OSx10' 0.91~10' 0 . 1 6 ~ 1 0' 0. 15x10'
0 . 9 6 ~ 1 0 ~ 0 . 0 4 ~ 1 0 ~
423k3 1.9
82 1545.23
1023568.14 1785 l3.M) 398119.73 762k48.73 - -
512k36.12
-
-
- -
39f 3.27 783+S 1.8
87f 3.18 189k47.46 45C 6.35
47W36.60
not be distinguished from each other. However, they can be distinguished from each other in a few cases based on their C-banding patterns (MOTARA and RAI 1978; RAO and R.41 1987a). The chromo- some complement of Ae. mediovittatus is the only exception that can be distinguished from the chro- mosome complements of other Aedes species. The chromosome complement of Ae. mediotittatus con- tains a pair of much smaller chromosome 1 , a pair of much larger chromosome 2 and a pair of interme- diate-sized chromosome 3 (see Fig. 3c). This sug- gests that a considerable chromosome repatterning has occurred in the karyotypic evolution of this spe- cies. In situ hybridization to chromosome 2 of this species supports this assumption. The metaphase complement of Hg. equinus with the rRNA gene cluster on chromosome 2 can be derived from a metaphase complement containing the rRNA gene cluster on chromosome 1 by a translocation be- tween chromosome I and chromosome 2 , possibly via the 'shift' (STRICKBERGER 1976) of a small chro- mosome segment containing the rRNA gene cluster. A similar mechanism can place the rRNA gene cluster from chromosome 1 to chromosome 3 in Tp. hurnhusa and Ar. subalhatus. The location of the rRNA gene cluster on chromosome 1 in a majority
of mosquito species supports this assumption. The physical location of the rRNA gene cluster on the chromosome arm close to the telomere in Ae. atro- palpus, Ae. epactius and Tp. bambusa, at an inter- calary position in Hg. equinus and Ae. triseriatus, and close to the centromere in the rest of the culi- cines can be explained by paracentric inversions (see Fig. 6). Further, no correlation exists between the taxonomic grouping of the species and the chro- mosome location of the rRNA genes. For example, the rRNA genes are located on chromosome 1 in Sa. cyaneus and Wy. smithii and on chromosome 3 in Tp. bamhusa even though these species are in- cluded in the same tribe Sabethini. Similarly, the species of Aedes, H g . equinus and Ar. suhalbatus are classified within the tribe Aedini, but the rRNA genes are located on chromosome 1 in the species of Aedes except Ae. mediovittatus, chromosome 2 in Hg. equinus, and chromosome 3 in Ar. subal- hatus.
In a cell line of Ae. albopipictus that was derived from minced neonate larvae (SINGH 1967), SPRAD- LING et al. (1974) estimated 430 copies of rRNA genes per haploid genome using Cot-curve analysis. Recently, PARK and FALLON (1990) estimated 30C 400 rRNA genes in the same cell line, and 35C.500

Hereditas 113 (1990) 18s + 28s RIBOSOMAL R N A GENES I N MOSQUITOES 287
in larvae, and about 1200 copies in the pupae and adults of Ae. aegypti, using dot-blot hybridization. However, the interpretation of their data was based on the 1C-DNA content of 1.45 pg for Ae. albopic- tus cell line (SPRADLING et al. 1974) and 1.30 pg for Ae. aegypti (DITTMANN et al. 1990). During the pre- sent study, we estimated 423k31.9 and 821k45.23 copies of rRNA genes per haploid genome in the adults of Ae. aegypti and Ae. albopictus respec- tively. However, our interpretation was based on the 1C-DNA content of 0.81 pg in Ae. aegypti and 1.24 pg in Ae. albopictus (RAO and RAI 1987b). The haploid nuclear DNA content has been shown to vary nearly 3-fold among world strains of Ae. albo- pictus (KUMAR and RAI 1990). Our preliminary study has shown a 5-fold variation in rRNA gene copy number between two strains of Ae. albopictus that differed 3-fold in their 1C-DNA contents; the Savannah strain from the USA with 1.65 pg 1C- DNA content has 1083 copies of rRNA genes per haploid genome and the Koh Samui strain from Thailand with 0.62 pg 1C-DNA content has 218f12.5 copies (KUMAR and RAI, unpublished). In three species examined from the Ae. albopictus sub- group (viz.: Ae. albopictus, Ae. flavopictus and Ae. seatoi), 1.4-fold difference in the haploid DNA con- tent is accompanied by over 5-fold difference in the copy number of rRNA genes. Among other species of mosquitoes examined, the copy number spanned with a minimum value of 39k3.27 in Sa. cyaneus to a maximum value of 783k51.8 in Wy. smithii. It has been suggested that there is a positive correla- tion between the copy number of rRNA genes and 1C-DNA content (BIRNSTIEL et al. 1971; CAVALIER- SMITH 1985b). In contrast, no such correlation was observed during the present study similar to the observation made in the flax genotrophs by CULLIS (1 976). As with the chromosome localization of the rRNA genes, no correlation is observed between the taxonomic position of the species and the copy number of the rRNA genes in mosquitoes exam- ined. For example, the difference in the copy number of the rRNA genes between Wy. smithii and Sa. cyaneus with nearly the same 1C-DNA content is as much as 20-fold, although both species are included in the tribe Sabethini. At present, the fac- tors controlling the copy number of the rRNA genes among different species of mosquitoes are unknown and remain to be investigated.
On an average rDNA constitutes only about 1 %, or at maximum only 3.7 % of total genome for eu- karyotes (see CAVALIER-SMITH 1985b) with more values ranging around 1 % (SCHAFER and KUNZ 1987). Based on the average ribosomal repeat unit
length of 9.0 kb in Ae. aegypti (GALE and CRAMP- TON 1989) and 15.6 kb in Ae. albopictus (PARK and FALLON 1990), the proportion of haploid genome represented by the rDNA in these species is 0.96 and 2.10 % respectively.
In conclusion, the present study has shown that the rRNA gene cluster is located on a single chro- mosome per haploid genome in all species of mos- quitoes examined except in Ae. triseriatus. How- ever, the species differed nearly 26-fold in the abundance of rRNA genes. The presence of the ribosomal RNA gene cluster on different chromo- somes and at different positions on the chromosome arm among mosquito species indicates that a con- siderable chromosome repatterning through translo- cations and inversions has occurred in the karyo- typic evolution of the members of Culicinae and corroborates earlier findings of RAI et al. (1982) and MUNSTERMANN (1981).
Acknowledgements. -We thank Drs S. Kambhampati and L. E. Munstermann for critically reading the manuscript; Mr Brian Turco, Mrs Peggy Hodges and Miss Dawn Verleye for rearing the mosquitoes in the insectary; and Mrs Tracy Frost for help in typ- ing. This work was supported by NIH research grant 5R01 A1 21443 to KSR. We also thank the reviewers for suggesting changes in the earlier draft of this paper.
References BAKER, R. H. and ASLAMKHAN, M. 1969. Karyotypes of some
Asian mosquitoes of the subfamily Culicinae (Diptera Culici. dae). - J . Med. Entomol. 6: 44-52
BAKER, R. H., SAKAI, R. K. and MIAN, A. 1971a. Linkage group- chromosome correlation in a mosquito: inversions in Culex tritaeniorhynchus. - J . Hered. 62: 31-36
BAKER, R. H., SAKAI, R. K. and MIAN, A. 1971b. Linkage group- chromosome correlation in Culex tritaeniorhynchus. - Science 171: 585-587
BECKINGHAM, B. 1982. Insect rDNA. -In: The Cell Nucleus (eds H. BUSCH and L. ROTHBLUM), Academic Press, New York, p. 205-263
BHALLA, S. C., CAIAIBA, A. C. I., CARVALHO, W. M. P. and SAN- TOS, J. M. 1974. Translocations, inversions and correlations of linkage groups to chromosomes in the mosquito Culex pipiens fatigans. - Can. J . Genet. Cytol. 16: 337-850
BICUDO, H. E. M. C. 1981. Nucleolar organizer activity and its regulatory mechanisms in Drosophila species of the ‘mulleri’ complex and their hybrids. - Caryologia 34: 231-253
BIRNSTIEL, M. L., CHIPCHASE, M. and SPIERS, J. 1971. The ribo- somal RNA cistrons. - Progr. Nucleic Acid Res. Mol. Biol. 11: 351-389
BLACK, W. C. IV, MCLAIN, D. K. and &I, K. S. 1989. Patterns of variation in the rDNA cistron within and among world popu- lations of a mosquito, Aedes albopictus (Skuse). - Genetics
BLIN, N. and STAFFORD, D. W. 1976. A general method for isola- tion of high molecular weight DNA from eukaryotes. - Nu- cleic Acids Res. 3: 2303-2308
BRADFIELD, J. Y. and WYATT, G. R. 1983. X-linkage of a vitelloge- nin gene in Locusta migratoria. - Chromosoma 88: 19&193
121: 539-550

CAvmfiK-Smii. T. 19XSa. Preface. - In: The Ewlrrtinri o/ Ge- nome Size (ed T. C4\ w u - S w T i i ) , Wi!ey. N ~ M Ynrk . p. x
C.AvAi.im-Smrti. 1'. 198% Eukaryotic gene number$, non-coding DNA, and genome s i ~ e . - In: The E\-c~/rctiori of Geriome Sire
CcLi.iY. C. A. 1976. Environmentally induced changes in ribo- somal RNA cistron number in tlax. - Her-edit? 34. 73-75,
M.. S ~ ~ I U O . R. and U ~ K I I I . I 1988. Extra ribo\omal \pacer sequences in Trititrir\ - J ,Wid B~IJ/ 204: K05-813
D t \ \ i i o ~ i x L. 1972. Die Zuordnung der Koppelunp\griippen IU
den Chromojornen hei der Stechmiicke Cirle.~ / i r p i e r i \ L. - Chron i~~~on io 37: 43-52
. F.. KIKI-\. P.. 5 I i \ i ~ ~ i i . K. and HV.ZDOK\. H . H . 1990. Ploidy level\ and DNA nthe\i\ in fat body cells of the adult moyuito. A e d r t O P Q ~ I the rok of ju\enilr hormone
( ~ d T. CA\'.4i.i~K-SMiItlJ, W'ilr'?. . V ~ M Yr1r.k. p. 69-103
D E L ~ ! C ~ . I I ~ X I . S. D.. A\I)HO\i(i). F., .4\L>KE.\Z%OLI. hl. . GlLl i - \ \ i ,
t \ i x ~ a . S A. and G \ i i ~ J . G. 1975. Ditferential replication of \atellite DNA iii polyploid t i \we o fDr~ i .~op / i i / u i . i r i / rs . ~ Clrr~i- nin\ornir 5iJ: 175-195
F\ q \ \ , H J.. B% c k i A \ I L R A. dnd P4mi I.. 51. L. 1974. Location ot gcnw coding for I X S and l X S ribo\omal R N A in the human
riiniowmiu 48: 405426 + \ i P i o \ . J. IOXY. The riho\omal gene\ of the .\ ' l l ~ q \ / l f l ~ Elll. .I Rrrx h n 185: 3 I 1-3 17
GI K i i i . S. A. and C w i 55 . H. V. 1976. Further studie> on the rihwamal R h A cistroii\ ot Sc iura ~op in iph i lu [Diptsra). - G c ~ n t , f i r . r 83: 8 1-90
G i i t I I H ~ C I , H. M. and Hxi.i>.&\i. J . B. S. 1347. Se\-lmkaee and \ex detemiinatioii i n a mosquito. Ci1le.1 niole.srrrr - Herc,rlirii.\ .?.i: !75-190
( h ) i w , % \ i ~ K k . c. and Bi oov. s. E. 1975. Vi\ualr7ation (it n u - cleolu\ organirer in mammalian chromoromer wing \il\er mining. - C / ~ r ~ ~ n i o \ ~ ~ n r ~ i S.+ 37-50
H \ c ~ ~ . i . i . K. and R, \v- . \ \* r t i . H . A. 1983. The chromoionie\ of t h o 1 h o w p t i i l i i race\: I l r ~ ~ w p / r r / ~ ~ n u ( i i r t i i i u \ i t r ~ i and D n u/hi)niicum 111. Localintion of nucleolar organiier region\. - ~ ; e n ~ t o u hi): 123-1?X
IWk. Thc chromororne location of ribomnal DN.4 i n the mou\r. - C'hroirioriniit 4Y: 155-16C)
I-ocation of ribowma1 DNA i n thc. human chromosome comple- ment. P ~ o c . .Vuri. , -kud. Sl: I LrSA 69: 339.1-3398
t I f \ t ~ ~ ~ i i ~ \ . A . S. . W.\RHLRIO\ . D. and ;\r\\cjou. K. C. 1971b. Localiution of rDNA i n the chimpanzee (Puri rro,y/odvre\) chromosome complenient. - C/i r~~ni~~.soni i~ 46: 4 3 5 3 4 I
t-iEv\ic.. W., LnK. B. and L~o\c l \ i , 0. 1975. The location of nucleoluE organizer regions 111 D~o\r:plrilu Inder - Chronio- wmo S l : 57-63
H v . T C.. SPIRI.I.(I. S . E. and PARDL 1.. M. L. 1975. Distribution of IXSc28S riboromal genes rn mammalian genomes. - C / I ~ ( I . niouJmo 53: 25-36
KITZMIL.LEH. J . H. 1967. Mosquito c)togenetic\. - In: Gerirrrc I I J ~ Inrrc.tr Irc tors of Ilrseose ( e d s J. W WRIGHT m l f R. P.\L I .
Khiwi-r. K . L. and STO\I:. A. 1977. - A Catalog of the Mmqui- toes of the World (Diptera: Culicidae). (2nd edition). - Zlie (20 W. King Conipuiiy. Bultirnow. p. 2
K R I L ' I L L K . R. D. 1978. A inocquito hith eight chromosome\: Cliu- qmii: hurhunu Dyar. - Mo54 new^ 38: 551558
K L v m A . and G ~ , P T ~ . J . P. 1987. Cytogenetic5 o f Zt~pi-~oriut indiunii\ Gupta f Diptera: Drosophilidae): Nucleolar organizer region\. mitotic and polytene chromosonies and inversion poly- morphivn. - Gerierim 74: 19-25
KL \I.AR. A. and R4i. K. S. 1990. lntraspecific variation in nuclear DNA content among uorld populationc of a mosquito. Aedes
t { l \ i l F v b ~ ) \ , A . s . E I ~ ~ ~ ~ ~ . E. 51.. y L . IT. 'r. a l l d ~ ~ i \ \ ( j ~ l t ~ . K . c.
. 4. s.. WARHI R I O \ . D and .Al \%<X)i>. K. c. 1Y77.
E/tci'/<'l- PUh/ C O , . j ' L W Y1Jr-k. p. 133-146
olbopicri~s (Skuse). - Theor. Appl. Genet. 79: 748-752 M,ACDOSALD, P. T. and RAI, K. S. 1970. Correlation of linkage
groups with chromo\omes in the mosquito Aedes aegypfi L. - - Gerretics 66: 475485
M ~ C G R E C ~ O R , H. C. and SHERWOOD, S. 1979. The nucleolus orga- nirers of Pletkodon and Aiieides located by in situ nucleic acid hybridization with Xenopits 'H-ribosomal RNA. - Chromo- .sonlo 72: 271-280
M~\I .ATIS. T., FRnwti. E. F. and SAMBROOK, J. 1982. Molecular Cloning: A Laboratory Manual. - Cold Spyin,? Hurhor La-
M O T A R ~ , M. A. and R.u. K. S. 1978. Giernsa C-handing patterns iii A d e \ (Stegoniyiu) mosquitoes. - Chroniosoma 70: 51-58
h l L \ S T E R > 1 4 U \ . L. E. 1981. Enzyme linkage maps for tracing chromosomal evolution in Ardes mosquitoes. -In: Application of Generic..s 011 Cito/o,q? i n Insec~t .Yj.sremoric.s und Evolution ( e d M. W. SIOCK). Unw Oflduho. Moscon8. p. 129-140
ML\STERMAY\. L. E. and MAR('ii1. A. 1986. Cytogenetic and iso- ryme profile of Soherhe.\ cymeus: a mosquito of the neotropical canop). - J . tiered 77: 241-248
OISHI. M.. LWKF. J. and W v ~ r r . G. R. 1985. The ribosomal DNA gene\ of Locu.\tu migraror-ia: copy number and evidence for underreplication in a polyploid tissue. - Cun. J . Bioc.lieni. Cell. Biol. hi: 106&1070
P \RDI E. 51. L. 1974. Localization of repeated DNA sequences in Xenopiis chromosomes. - Cold Spring Harbor Synip. Quunr.
. In situ hybridization. - In : N d e i c . Acid H>hi-idirurion: A Pi-uc r i d Approach (eds B. D. HAMES (2nd S. J . I+IGGI\S!. I.K.L. Press. Ovjord, p. 179-203
P 4 ~ i ) i . t . 41. L.. GERBI. S. A,. ECKHARDT, R. A. and GALL. J. G. 1970. Cytological localimion of DNA complementary to ribo- somal RNA in polytene chromosomes of Diptera. - Chromo- uiniu 29: 268-290
P ~ K I X 1.. hl. L. and H ~ v , T. C. 1975. Localization of 18s and 28s ribowrnal genes i i i the chromosome$ of the lndian Muntjac. --
P ~ R K . Y.\\G-JA and F.xi.i.cn, A. M. 1990. Mosquito ribosomal RNA genes: characterization of gene structure and evidence for change, in copy numher during development. - Insect Bio- 1 licni. 3011): 1-1 I
R4i. K . S. 1963. A comparative \tudy of mosquito karyotypes. - .Arm Entomol. Sor Anr 56: 160-170
R\i. K. S. 1966. Funher observations on the somatic chromosome cyto!og} of some mosquitoes (Diptera: Culicidae). -Ann . En- ronrol Soc . A m . 59: 242-246
R.M. K. S . , PASHLEY, D. P. and ML,NSTERMANN, L. E. 1982. Genet- ics of speciation in aedine mosquitoes. -- In: Rec,ent Derelop- firenis i n Genetics if Insect Disease Vectors (ed,s W. M. M.
Ckunipoign. 111. p. 83-1 29 RAO. P. N. 1985. Nuclear DNA and chromosomal evolution in
mosquitoes. - Ph.D Thesis. Unii3. of Notre Dame. US4 R m . P. N. and R4i. K. S. 1987a. Comparative karyotypes and
chrommomal evolution in some genera of nematocerous (Dip- tera: Nematocera! families. -Ann. Entomol. Soc. h i . 80: 321- 332
R ~ o . P. N. and RAI . K. S. 1987b. Inter- and intraspecific variation in nuclear DNA content in Aedes mosquitoes. - Heredity 59: 253-258
RrTcisSj\. F. M. and SPIEGELMAN, S. 1965 Localization of DNA complementary to ribosomal RNA in the nucleolus organizer region of Drosophila nielanogaster. - Proc. Nnrl. A [ m i Sci. USA 53: 737-74s
ROCHAIX, J . D.. BIRD. A. and BAKKEN A. 1974. Ribosomal RNA gene amplification by rolling circle . -.I. Mol. El,Ill. x 7 473- 487
boriltol~\.. .vt'\+' Y#rk
./. ( E l / BlIJl 64: 25 1-254
S T E l \ E K . W. J. T~BACHNICK. K. s. RAI ond s. NAKANF), Stipes,

H<vedrtas 113 (19901 18s + 2RS RIBOSOMAL RNA GENES IN MOSQUITOES 289
SAMBROOK, J., FRITSCH, E. F. and MANIATIS, T. 1989. Molecular Cloning: A Laboratory manual. Vol. 2. - Cold Spring Harbor Laboratory, New York
SciiAFER, M. and KUNZ, W. 1987. Gene amplification does not occur in the oocytes of Locusta migratoria. - Dev. Biol. 120: 4 1 8 4 2 4
SCHAFER, U. and SCHAFER, M. 1980. Localization of the ribosomal RNA genes in Drosophila simulans. - Chromosoma 79: 287- 29 1
SIKH, K. R. P. 1967. Cell cultures derived from larvae of Aedes albonictu., (Skuse) and Aedes aepvati (L.). - Curr. Sci. 36: 506- 508
SiwALDI, R. M. and CUMMINGS, M. R. 1981. Localization and characterization of rDNA in Drosophila rumiditarsus. - Chro- momma 81: 655471
SPEAK, B. 1974. The genes for ribosomal RNA in diploid and polytene chromosomes of Drosophilu melanogaster. - Chro- mosoma 48: 159-179
Repetitious and unique sequences in the heterogeneous nuclear and cytoplasmic messenger RNA of mammalian and insect cells. - Cell 3: 23-30
STRICKBERGER, M. W. 1976. Genetics, (2nd edition). -Macmillan Publishing Company, New York, p. 5 12
TRUMAN, J . W. and CRAIG, G. B. 1968. Hybridization between Aedes hendersoni and Aedes triseriatus. -Ann. Entomol. Soc.
SPRADLING, A,, PENMAN, s., CAMPO, M. s. and BISHOP, J. 0. 1974.
Am. 61: 10261025
L. P., WITXOLT, B. and FEENSTRA, W. J. 1988. In situ hybridiza- tion to somatic metaphase chromosomes of potato. - Theor. Appl. Genet. 76: 4 2 M 2 4
CHI-PILONE, G . 1982. Chromosome localization of 18S+28S and 5 s genes in evolutionarily diverse anuran amphibians. - Chro- mosoma 84: 47-91
WHITE, G. B. 1980. Academic and applied aspects of mosquito cytogenetics. - In: Insects Cytogenetics. R . Entomol. S w p . 10 (eds R. L. BLACKMAN, G. M. HEWITT and M. ASHBLIR~EK) , Blackwell Scientific, Oxford. p. 245-274
N. 1982. Cytogenetics of the parthenogenetic grasshopper W m - ramaba virgo and its bisexual relatives. IX. The ribosomal RNA cistrons. - Chromosoma 85: 181- 199
WILEY, J. E., LITTLE, M. L., ROMANO, M. A,, BLOUNT, D. A. and CLINE, G. R. 1989. Polymorphism in the location of the 18s and 28s rRNA genes on the chromosomes of the diploid-tetra- ploid treefrogs Hyla chrysoscelis and H . ~~ersicolor - Chronro- soma 97: 481487
ZIMMERMAN, P. A., LANC-UNNASCH. N . and CULLIS, C. A. 1989. Polymorphic regions in plant genomes detected by an M13 probe. - Genome 32: 824828
VISSER, R. G. F., HOEKSTRA, R., VAN DER LEJI, F. R., PIJNACKER,
VITELLI, L., BATISTONI, R., ANDRONICO, F., NARDI, 1. and BARSAC-
WHITE, M. J. D., DENNIS, E. S., HONEYCUTT, R. L. and CONTRERAS.





























