Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Arthropod Structure & Development 41 (2012) 419e433
Contents lists available
Arthropod Structure & Development
journal homepage: www.elsevier .com/locate/asd
Comparative morphology of pretarsal scopulae in eleven spider families
Jonas O. Wolff*, Stanislav N. GorbFunctional Morphology and Biomechanics, Zoological Institute, University of Kiel, Am Botanischen Garten 1-9, D-24098 Kiel, Germany
a r t i c l e i n f o
Article history:Received 10 January 2012Accepted 25 April 2012
Keywords:AraneaeArthropodaCuticleAttachmentClaw tuftLocomotionEcomorphologyScalingEvolutionAdaptation
* Corresponding author.E-mail addresses: [email protected] (J.O. Wolff
(S.N. Gorb).
1467-8039/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.asd.2012.04.004
a b s t r a c t
Many wandering spiders bear attachment pads (scopulae) on their tarsi, consisting ofhierarchically-branching adhesive setae. Amongst spider families and even species, these showremarkable differences in morphology. Using scanning electron microscopy, the scopula microstructureof sixteen spider species was described, with the focus on pretarsal scopulae (claw tufts). Area and shapeof the claw tuft, seta and setule density, as well as seta and spatula dimensions were analysed andcompared. Claw tufts of the majority of species studied show a similar gradient in size and shape fromanterior to posterior legs: the dimension of pads increases, while setal density decreases. Commonly,there is also a gradient of both the seta and spatula size within the claw tuft: Setae become larger fromthe proximal to the distal part of the pad, and spatulae size increases in the same direction at the level ofindividual seta. Often, different hierarchical levels of claw tuft organisation are differently expressed indifferent species: Species with lower setal density usually have broader setae. Smaller spatula size oftenimplicates higher setule density. Evolutionary and ecological aspects of the scopula origin are discussed.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Awide-spread morphological feature of terrestrial arthropods isthe presence of adhesive setae, cuticular structures that providedynamic attachment to various surfaces. These structures haveevolved independently several times, indicating their enormousefficiency and biological success (Scherge and Gorb, 2001; Beuteland Gorb, 2001). In wandering spiders, the complexity of theadhesive apparatus has reached a superior level among arthropods:specialized hierarchically-branched setae are organized in densearrays, called scopulae. We speak about the hierarchy, if the cuticleprotuberances are covered with smaller protuberances. Scopulaeare located on both ventral and lateral parts of the tarsus andmetatarsus. Strongly-specialised distal scopulae, so-called clawtufts situated at the tip of the tarsus, play amajor role in attachmentduring locomotion. Their microstructure was described for the firsttime by Homann (1957): The adhesive setae are flattened andcovered with hundreds of microtrichia (called setules) on theirventral side. Each setula ends in a broadened plate-like structure,the spatula. It can be assumed that these setae are modified tactile
All rights reserved.
hairs, which sometimes still are supplemented by a single sensorycell (Foelix et al., 1984).
Beside the claw tufts, scopulae are also often present on theanterio- and posterio-lateral parts of the tarsi and metatarsi in theanterior legs. In contrast to those in the claw tuft, the adhesive sidesof setae in these parts are distally-orientated (Niederegger andGorb, 2006) and can be activated through the erection of hairs,caused by increasing hemolymph pressure (Rovner, 1978; Foelixet al., 1984). Leg scopulae mainly play a role in prey capture,when large and/or dangerous prey has to be secured and heldaway from the body (Rovner, 1978; Foelix et al., 1984; Pekár et al.,2011). They also might support the spider’s body, while restingon steep surfaces and climbing over edges (Niederegger and Gorb,2006).
Depending on its physical mechanism of adhesion, the spider’sbody dimensions, the ecological and ethological demands, and itsphylogenetic origin, the scopula can be highly diverse in variousspider species.Whereas the variety of attachment devices in insectsis well covered by numerous publications (for reviews, see Beuteland Gorb, 2001; Gorb, 2001; Gorb and Beutel, 2001), there areonly a few descriptions of adhesive systems of some individualspecies of spiders (Homann, 1957; Foelix and Chu-Wang, 1975; Hill,1977; Roscoe and Walker, 1991; Kesel et al., 2003; Moon and Park,2009; Foelix and Erb, 2011). In the systematic literature claw tuftsare widely used as a character, whereas a detailed morphologicaldescription and understanding of the functional mechanism is

J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433420
lacking (Homann, 1975; Forster and Platnick, 1985; Ramírez, 1995,2003; Davila, 2003). Platnick and Lau (1975) compared claw tuftsof representatives of Anyphaenidae and Clubionidae, however,without discussing the fine structure of setae and their adhesivefunction. Ubick and Vetter (2005) demonstrated differences in theorigin of claw tufts in Dionycha and three-clawed spiders. Spatulardimensions and densities were previously compared in eight spiderspecies (Peattie and Full, 2007) without a detailed description ofthe spider attachment system and its relationship to the biology ofthe species studied.
The present paper is a comparative scanning electron micro-scopy study of scopulae in sixteen wandering spider species fromdifferent families. Morphological features are discussed accordingto their possible functional significance. The following questionswere asked. (1) Is there any correlation between different micro-morphological variables in the species studied? (2) Are there anytrends in the spatial distribution of different adhesive setae andtheir spatulae? (3) Does scopula structure correlate with thespecies’ habitat? (4) Are there similar evolutionary trends presentin different groups of spiders?
2. Material and methods
Scopulae of sixteen araneomorph species from eleven familieswere studied: Agroeca brunnea BLACKWALL 1833 and Agroeca cupreaMENGE 1873 (Liocranidae), Anyphaena accentuata WALCKENAER 1802(Anyphaenidae), Clubiona caerulescens KOCH 1867 (Clubionidae),Cupiennius salei KEYSERLING 1877 (Ctenidae), Drassyllus praeficus KOCH
1866 (Gnaphosidae), Ebrechtella tricuspidata FABRICIUS 1775 (Thomi-sidae), Evarcha arcuata CLERCK 1757 (Salticidae), Heliophanus cupreusWALCKENAER 1802 (Salticidae), Micrommata virescens CLERCK 1757(Sparassidae), Oxyopes heterophthalmus LATREILLE 1804 (Oxyopidae),Philodromus cespitum WALCKENAER 1802(Philodromidae), Thanatusformicinus CLERCK 1757 (Philodromidae), Tibellus oblongusWALCKENAER
1802 (Philodromidae), Zelotes subterraneusKOCH1833 (Gnaphosidae)and Zora spinimana SUNDEVALL 1833. Specimens were collected in theUkraine, with the exception of C. salei, which comes from a labora-tory stock (Department of Neurobiology of the University of Vienna,Austria). The specimens were fixed and stored in 70% ethanol.Mainly single female specimens were analysed, except in E. arcuata,H. cupreus, O. heterophthalmus and, T. oblongus, where both sexeswere analysed. The four walking legs on one side of the body werecut off, dehydrated in an ascending series of ethanol, critical pointdried and sputter-coated with a 15 nm thick layer of goldepalladium. Specimens were observed at 20 kV in a Hitachi S-800scanning electronmicroscope (SEM).Micrographs atmagnificationsof 80e800were used for an overview,magnifications of 2000e4000for analysis of the setae, and magnifications of 10,000e20,000 foranalysis of setules. For measurements of structures from SEM-micrographs, DatInf� Measure software (DatInf GmbH, Tübingen)was used. The following parameters were quantified: length, widthand total area of the claw tuft, number and density of setae, setalwidth at the tip, at the distal, and proximal parts of the claw tuft.Additionally, both the setule density and spatula width in the distalandproximal parts of the claw tuftweremeasured. Furthermore, thepresence of tarsal and metatarsal scopulae was analyzed.
3. Results
3.1. Scopula morphology
Clubiona caerulescens KOCH 1867 (Clubionidae)
C. caerulescens is a nocturnal wandering spider of medial size (\6e10 mm, _ 6e8 mm) that is wide-spread throughout Europe. It
lives in woods on deciduous trees and shrubs, where it searchesactively for prey and captures it using the anterior legs. Clubiona sp.can easilywalk on smooth surfaces like glass (personal observation).
Claw tufts are of an oval shape being one third longer thanwide.L1 and L2 (anterior legs) have a claw tuft of similar dimensions withabout 90 setae and an area of about 11 � 103 mm2 (Fig. 2B). Clawtufts on L3 and L4 (posterior legs) are three times larger and consistof 205 (L3) and 225 (L4) setae, respectively. L4 bears the biggestclaw tuft (Fig. 2C) with an area of 31 � 103 mm2 and double thelength (300 mm) and width (170 mm) in the anterior legs. Setaldensity is high in L1 (8.3 � 10�3 mm�2) and lower in the other legs(about 7.5 � 10�3 mm�2). L1 also differs from the others legs(4.1 mm�2) in the setule density (3.4 mm�2 in setae of the proximalpart of the claw tuft). Claw tuft setae are attached to the underlyingcuticle through the socket (Fig. 2D inset), which is shaped in sucha way that setal movement is guided by the socket in a predefined(lateral) direction. Setae are S-shaped and their sockets are directedto the claw tuft centre. These features might influence setal springproperties, which were previously assumed to be important ingenerating adhesion in spiders (Gasparetto et al., 2009). Setaebecome gradually flattened and broadened at the tip, which ends ina fringed edge (Fig. 2D). The width of the setal tip differentiatesincreasingly from the proximal to the distal part of the tuft. Thereare some differences between the anterior and posterior legs. Forexample, in L1, seta width is, on average, 10.6 mm in the proximalpart of the claw tuft and 11.6 mm in the distal part, while 7.9 mm and15.1 mm, respectively, in L4. The spatula is triangular shaped andslightly curved. Its average width is rather regular in the range of0.22 mmexcept in L4, where spatulae bear double widths (0.45 mm).
L1 and L2 have dense scopulae located at the ventro-lateral sidesof both the tarsus and metatarsus. Setae in these parts havea rounded shape. Setules coverage appears on the side that is facingthe tarsus (Fig. 2E), andmight be activated by erection, caused by anincrease in hydraulic pressure within the leg, as has been previ-ously described for lycosid and salticid scopulae (Rovner, 1978;Foelix et al., 1984). The direction and the angle of setal movement isdetermined by the kidney-shaped hair sockets (Fig. 2E inset).
Zora spinimana SUNDEVALL 1833 (Zoridae)
Z. spinimana is the most abundant zorid spider in Europe. It hasa body length of 4.5e6.5 mm. It is a ground dwelling, pursuinghunter, which can be found in leaf litter and open sites.
Claw tuft and setal structure are comparable to those ofC. caerulescens. Relative to the smaller size of Z. spinimana, its clawtufts are smaller than those in C. caerulescens: with an area of13 � 103 mm2 on L3 and 15 � 103 mm2 on L4 (Fig. 2F), and theposterior legs of Z. spinimana have claw tufts two times smallerthan those in C. caerulescens. The difference between posterior andanterior claw tufts in Z. spinimana is less expressed than in thecompared species: With an area of 10 � 103 mm2 on L1, the tuft isonly about one third smaller than that of L4. The smallest pretarsalscopula is in L2 (8 � 103 mm2). Setal density in this species is higherthan in clubionids: about 10.0 � 10�3 mm�2 on L1 and L2,9.0� 10�3 mm�2 on L3, and 7.8� 10�3 mm�2 on L4. Each seta ends ina point, but some setae aremassively broadened in their distal parts(Fig. 2F inset). Setae in the proximal part of the tuft are lessbroadened and more tapered.
Average setal width is about 10.6 mm in the distal and 7.5 mm inthe proximal parts of the claw tufts in all legs. Setule density isabout 2.5 mm�2 in distal scopula parts. Spatula width is higher thanin C. caerulescens and shows a gradient from the anterior (0.62 mmin L1) to posterior part of the scopula (0.48 mm in L4).
The claw tuft is surrounded by long (about 125 mm long) tactilehairs. Tarsal claws are relatively small. The leg scopula is not very
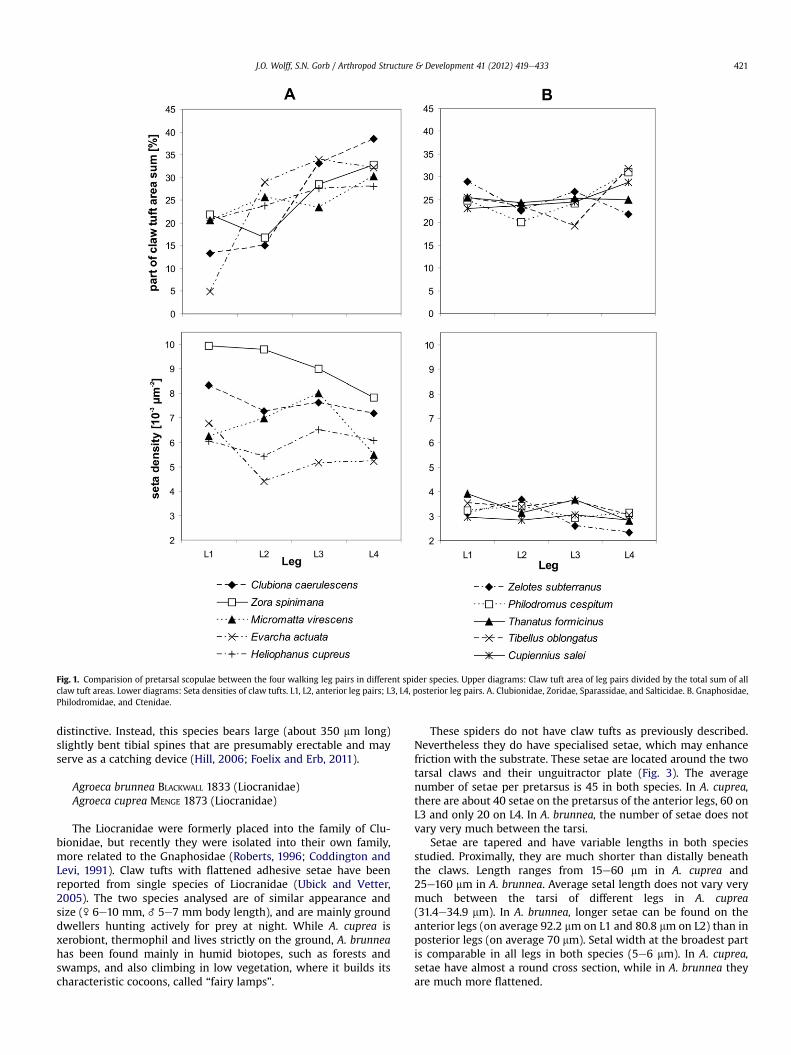
Fig. 1. Comparision of pretarsal scopulae between the four walking leg pairs in different spider species. Upper diagrams: Claw tuft area of leg pairs divided by the total sum of allclaw tuft areas. Lower diagrams: Seta densities of claw tufts. L1, L2, anterior leg pairs; L3, L4, posterior leg pairs. A. Clubionidae, Zoridae, Sparassidae, and Salticidae. B. Gnaphosidae,Philodromidae, and Ctenidae.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 421
distinctive. Instead, this species bears large (about 350 mm long)slightly bent tibial spines that are presumably erectable and mayserve as a catching device (Hill, 2006; Foelix and Erb, 2011).
Agroeca brunnea BLACKWALL 1833 (Liocranidae)Agroeca cuprea MENGE 1873 (Liocranidae)
The Liocranidae were formerly placed into the family of Clu-bionidae, but recently they were isolated into their own family,more related to the Gnaphosidae (Roberts, 1996; Coddington andLevi, 1991). Claw tufts with flattened adhesive setae have beenreported from single species of Liocranidae (Ubick and Vetter,2005). The two species analysed are of similar appearance andsize (\ 6e10 mm, _ 5e7 mm body length), and are mainly grounddwellers hunting actively for prey at night. While A. cuprea isxerobiont, thermophil and lives strictly on the ground, A. brunneahas been found mainly in humid biotopes, such as forests andswamps, and also climbing in low vegetation, where it builds itscharacteristic cocoons, called “fairy lamps”.
These spiders do not have claw tufts as previously described.Nevertheless they do have specialised setae, which may enhancefriction with the substrate. These setae are located around the twotarsal claws and their unguitractor plate (Fig. 3). The averagenumber of setae per pretarsus is 45 in both species. In A. cuprea,there are about 40 setae on the pretarsus of the anterior legs, 60 onL3 and only 20 on L4. In A. brunnea, the number of setae does notvary very much between the tarsi.
Setae are tapered and have variable lengths in both speciesstudied. Proximally, they are much shorter than distally beneaththe claws. Length ranges from 15e60 mm in A. cuprea and25e160 mm in A. brunnea. Average setal length does not vary verymuch between the tarsi of different legs in A. cuprea(31.4e34.9 mm). In A. brunnea, longer setae can be found on theanterior legs (on average 92.2 mm on L1 and 80.8 mm on L2) than inposterior legs (on average 70 mm). Setal width at the broadest partis comparable in all legs in both species (5e6 mm). In A. cuprea,setae have almost a round cross section, while in A. brunnea theyare much more flattened.

Fig. 2. Scanning electron micrographs (SEM) of tarsal structures. AeF. Clubiona caerulescens (Clubionidae), female. A. Tarsus of L2, lateral view. B. Claw tuft in L1, ventral view. C.Claw tuft in L4, ventral view. D. Claw tuft setae (type AS-III), ventral view. Inset shows sockets (same magnification). E. Leg scopula setae in L2, ventro-lateral view. Inset showssockets (same magnification). F. Zora spinimana (Zoridae), female. Claw tuft in L4, ventral view. Inset shows detailed view of setae (type AS-I). ch, chemosensory seta; cl, tarsal claw;ct, claw tuft; ms, metatarsal scopula; sb, bunch of tapered setae (type FS-II); th, tactile setae; ts, tarsal scopula.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433422
Although setule covering is present around the whole seta, notonly at its ventral part, some setal polarity can be observed: InA. brunnea, setule density is also much higher ventrally. In bothspecies, dorsal setules are two to three times longer than ventralones (Fig. 4, insets). Thus, the ability for contact formation onvarious substrates should be higher. However, setules do not havea spatulate end plate, just a blunt tip of 0.1e0.2 mm width. So themost obvious difference between these two species is in the lengthof their scopula setae, which positively correlates with the totalnumber of setules. This indicates that A. brunnea is structurallymore strongly adapted for climbing. However, the degree to whichthese setae contribute to the attachment ability remains unclearand requires further experimental evidence. Because of the lownumber of contact elements devoid of spatula, the contributionshould not be that high. For Agroeca spp., its large, well-structuredtarsal claws should have a greater relevance in attaching to veryrough and corrugated substrates, such as soil and litter.
Anyphaena accentuata WALCKENAER 1802 (Anyphaenidae)
The Anyphaenidae is a small family probably related to theClubionidae and Salticidae (Coddington and Levi, 1991), with onlysix species in Europe. A. accentuata is a typical arboreal species ofdeciduous forests. The variably-sized (\ 4.5e9 mm, _ 4e6.5 mm)spider is very common throughout Europe.
Its setae are remarkably large (with a width up to 45 mm) andless densely distributed. In the proximal part of the L3 claw tuft(Fig. 4A), the average width of the setal tip is 15.2 mm, while in thedistal part it is 31.5 mm (Fig. 4B) at an average setal density of only1.7 � 10�3 mm�2. With an area of 25 � 103 mm2, the claw tuft ismuch larger on L3 than on L4 (14 � 103 mm2) and L2 (17 � 103 mm2)(L1 was not analyzed). Setal density is highest on L4(2.2 � 10�3 mm�2). Setal width in the distal part of the claw tuft isabout 27 mmon both L2 and L4. In the proximal part of the tuft, setalwidth is higher on L4 (16.1 mm) than on L2 (13.3 mm). Variability of

Fig. 3. Agroeca spp. (Liocranidae). A. A. cuprea, male. Pretarsus of L3, frontal view. Inset shows detail of the frictional seta (type FS-I). B. A. brunnea, female. Pretarsus of L3, ventralview. Inset shows detail of the frictional seta (type FS-II). ch, chemosensory seta; cl, tarsal claw; dsl, dorsal setules; fs, frictional setae; th, tactile setae; vsl, ventral setules.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 423
setal width is also higher on L4 (width range: 6e37 mmon L4 versus10e35 mm on L2).
Average spatula width in distal setal parts is 0.50 mm. Setuledensity was measured at the distal part of the L3 claw tuft: with1.4 mm�2, it is less than one half of the value measured inC. caerulescens and Z. spinimana. Thus, in A. accentuata the contactpoints are distributed over a larger area than in the comparedspecies. It is also striking that setal planes are much more oriented
Fig. 4. Anyphaena accentuata (Anyphaenidae), female, ventral view. A. Pretarsus in L3. B. Dsetae. as, adhesive seta (type AS-II); ch, chemosensory seta; ct, claw tuft; sb, group of tape
towards the lateral sides of the tarsus than in any other speciesanalysed. Setae are slightly bent in the lateral direction: thisgeometry may increase the effectiveness of the contact area withthe substratum. In general, hierarchical organisation of the tuft isreduced in this species, as there are only a few large setae present.Presumably, this is an adaptation to less structured substrates, suchas leaves with smooth epicuticle. A lesser degree of hierarchy mayalso provide for easier detachment since structures that are less
etailed view of large adhesive setae. Inset shows spatula-bearing setules on adhesivered setae (type FS-II); th, tactile setae; ts, setae of the tarsal scopula.

J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433424
subdivided into smaller subcontacts can peel off at lower appliedforce due to a lesser degree of preventing crack propagation(Kendall, 2001; Gao et al., 2003; Chung and Chaudhury, 2005).
The tarsal scopula is sparse on the lateral sides of all legs.Adhesive setae here are relatively slender, have a rounded tip andventral setule coverage. These setae should not influence attach-ment at rest, due to their absence at the ventral site. The tarsalclaws may play an unimportant role in attachment, as they aresmall and disappear between surrounding long setae and tactilehairs.
Evarcha arcuata CLERCK 1757 (Salticidae)Heliophanus cupreus WALCKENAER 1802 (Salticidae)
Salticids are known for their excellent adhesion on smoothsurfaces (Kesel et al., 2003). These are mainly diurnal and optically-orientated hunters that are able to jump over long distances.Jumping on prey is a part of their hunting strategy. E. arcuata isa Middle European species with a body length of 6e8 mm (\\) and5e6mm (__). It lives inwet and dry meadows and heaths, where itcan be found on herbs and grasses. Evarcha spp. hunt by ambush-ing, a hunting strategy differing from the active hunting of mostspecies in the family, including H. cupreus. The latter species rea-ches a body length of from 3.5 to 6 mm in both sexes. It preferssunny sites in grasslands, ruderal areas and at forest edges.
The tarsi of both sexes in both species were analysed. InE. arcuata, claw tufts are oval with setae getting longer from ante-rior to posterior legs. The claw tuft is largest on L3 (\ 38 � 103 mm2,200 setae; _ 32 � 103 mm2, 150 setae). Females, with their largerbody size, have both a higher pad area and setal number than themales. However, in the female L1, a reduced claw tuft was found,which could be a result of leg regeneration (Fig. 5C) (\L1:6 � 103 mm2, 40 setae; _L1: 27 � 103 mm2, 100 setae). With theexception of L2, setal density is higher in females (6.8 � 10�3 mm�2
on L1, 4.4 � 10�3 mm�2 on L2 and 5.2 � 10�3 mm�2 on the posteriorlegs) than in males (3.9 � 10�3 on L1, 4.7 � 10�3 on L2 and L3,4.1 � 10�3 on L4). In H. cupreus (Fig. 5F), a similar relationship wasshown (\: 6.0 � 10�3 mm�2 on L1, 5.4 � 10�3 mm�2 on L2,6.5 � 10�3 mm�2 on L3, and 6.1 � 10�3 mm�2 on L4; _:5.1 � 10�3 mm�2 on L1, 5.3 � 10�3 mm�2 on L2, 6.7 � 10�3 mm�2 onL3, and 4.7 � 10�3 mm�2 on L4). Scopula area is about one third ofthat in E. arcuata (\: 8 � 103 mm2 on L1, 10 � 103 mm2 on L2,11 � 103 mm2 on L3, 12 � 103 mm2 on L4; _: 8 � 103 mm2 on L1,9 � 103 mm2 on L2, 10 � 103 mm2 on L3, 14 � 103 mm2 on L4).Intersexual differences are not as expressed as in the previousdescribed species, because of the similar body size of males andfemales in the latter. The largest scopula is on L4. Scopula is notreduced on the L1 of females. The shape of the pad is more rounded(nearly as wide as long).
Adhesive setae have the same shape in both species. Whilethose in representatives of Clubionidae and Anyphaenidae endrelatively straight, the tips are rounded in salticids and have a littlenotch in the middle (Fig. 5D). Setal width was measured at thebroadest part. In E. arcuata, setal width is 8e19 mm. In the distal partof the pad, it is, on average, 17 mm (9 mm on L1) in females and13 mm (14 mm on L1) in males. In the proximal part, it is on averageof 13 mm on L2 to 10 mm on L4 in females and 12 mm on L1 to 9 mmon L4 in males. In H. cupreus, the range in setal width is 5e19 mm. Inthe distal parts of the pad, it averages 13 mm in females and 14 mmin males. In the proximal parts, it is 8e10 mm, on average, in bothsexes. In E. arcuata, setule density is 1.9 mm�2 on the proximal partof the L3 claw tuft in both sexes and 2.1 mm�2 on the proximal partof the L4 claw tuft in themale. InH. cupreus, a value of 1.8 mm�2 (L3,L4, male) in the distal claw tuft part and between 1.8 mm�2 (L1,male) and 2.8 mm�2 (L3, female) in the proximal part was obtained.
Spatula in E. arcuata was measured on L3: in the male it is about0.80 mm, whereas it is only 0.50 mm in the female. In H. cupreus,spatula width is about 0.65 mm in the male, whereas in the femalespecimen, a lower value was found (about 0.50 mm on L3),comparable to that in E. arcuata. Seta sockets (Fig. 5D inset) havea bulged margin with a little thorn.
Setae in salticid claw tufts are arranged in a specific pattern. Inthe distal part of the claw tuft, setae are orientated longitudinallywhile more laterally in the proximal part (Fig. 5A,B,F). Anotherspecialisation of salticid claw tuft is in its form: while setae instraight runners, like C. caerulescens, end in a plain area, tips ofsalticid setae together form a hemispherical shape. This could beinterpreted as an adaptation to the high agility and manoeuv-rability of salticids. A hemispherical pad allows attachment anddetachment at various angles and in different directions. Suchability is definitely limited in a pad having a plain arrangement ofsetal tips. A tarsal and metatarsal scopula is absent in the jumpingspiders analysed.
Oxyopes heterophthalmus LATREILLE 1804 (Oxyopidae)
Lynx spiders have a hunting strategy similar to that of theSalticidae. They creep close to their prey and capture it by jumpingon it. However, both families are distantly related. The oxyopidspecies are distributed mainly in tropical regions. There are onlya few species known occurring in Europe. The species analysed,O. heterophthalmus, lives in western and southeastern Europe andoccupies warm and sunny sites, such as heaths and dry meadows,where it can be found on herbs and grasses.
O. heterophthalmus is 5e7 mm long. Oxyopes spp. have long stifftarsal bristles and large serrated tarsal claws. In O. heterophthalmus,the claws are about 130 mm in females and about 90 mm in males.Scopulae do not exist, but a few frictional setae are located at thepretarsus (Fig. 6). These are 50e200 mm long and 7e14 mm wide.Those, located beneath the middle hook, are flattened, while thelateral ones are not. Setules occur only at the ventral side of eachseta (Fig. 6C). Setule density is rather low (about 0.30 mm�2), but thesetules themselves are relatively long (about 8 mm on average).Spatulae are lacking: instead each setule often terminates ina slightly-bent rectangular small hook. These micro-hooks mightinterlock with asperities on rough substrates.
The female had twice the adhesive setae (9e14 per pretarsus)than the male (5e7 per pretarsus). Female setae were also longerthan those in the male. The female carried less setae on L1 (Fig. 6A)than on the other legs (Fig. 6B). In the male, there was no differencein setal number between the legs.
The contribution of these few, but obviously highly-specialisedsetae, to gripping and attaching should be evaluated in furtherexperiments. The enlarged claws of this species are presumablyvery relevant in the highly structured microhabitats, where thespiders can easily find a grip at leaf edges and trichomes.
Drassyllus praeficus KOCH 1866 (Gnaphosidae)Zelotes subterraneus KOCH 1833 (Gnaphosidae)
Most Gnaphosidae are nocturnal hunters, hiding in a silken sacduring the day. Both species studied are ground dwellers.Z. subterraneus is an euryoecious species wide-spread in middleand eastern Europe. D. praeficus prefers dry open sites. Adults ofboth species and sexes have a body length of 5e7 mm.
Distinct claw tufts are absent in both species, however,Z. subterraneus bears, in its anterior legs, a dense tarsal leg scopulareaching up to the pretarsus (Fig. 7D). Setae of the tarsal scopula inthis species are very similar to claw tuft setae previously described.Setae of the tarsal scopula are flattened with a huge number of

Fig. 5. Salticidae. AeE. Evarcha arcuata. A. Tarsus of L4, female, ventral view. B. Claw tuft in L4, male, lateral view. C. Claw tuft in L1, female, lateral view. D. Adhesive setae in L3,female, ventral view. Arrows indicate a little notch at the setal tip. Inset shows setal socket at the same magnification. E. Group of FS-II setae at the base of the L2 claw tuft. F.Heliophanus cupreus. Claw tuft in L3, female, frontal view. as, adhesive setae (type AS-II); ch, chemosensory seta; cl, tarsal claw; ct, claw tuft; fs, frictional setae (type FS-II); th, tactilesetae.
Fig. 6. Oxyopes heterophthalmus (Oxyopidae), female. A. Pretarsus of L1, ventro-lateral view. B. Pretarsus of L4, ventral-lateral view. C. Detail of the frictional seta, lateral view. ch,chemosensory seta; cl, tarsal claw; fs, frictional setae (type FS-I/II); th, tactile seta.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 425

J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433426
setules (density: 1.50 mm�2) ending in spatulae with an average tipwidth of 0.70 mm (Fig. 7F). The setal tip is rounded, with a width of10e14 mm at its broadest part. Setal density is relatively low(1.5 � 10�3 mm�2 on average), compared to that of claw scopulae.The scopula spreads over wide anterio- and posterio-lateral parts ofthe tarsus, but ventrally its setae are distributed only below theclaws. Nevertheless, these setae differ from those of the preycapture apparatus, previously described by Rovner (1978) andFoelix et al., (1984), and from those found in C. caerulescens (seeabove). Spatula-bearing setules are distributed here on the ventralside of the flattened seta, permanently facing the substrate. Lookingat the distribution of tarsal scopula setae, a ventral stripe lackingadhesive setae is obvious, indicating that this part is not primarilyinvolved in locomotion and resting. Below the claws, this hairlessstripe is absent; only the unguitractor plate remains free of setae(Fig. 7A). This pretarsal part has an area of about 13� 103 mm2 on L1
Fig. 7. Zelotes subterraneus (Gnaphosidae), female, ventral view. A. Pretarsus of L1 with adheand frictional setae (FS-I/II). C. Pretarsus of L4 with a row of frictional setae (FS-I). D. Tarsus ofseta (AS-II) with setules bearing distinct spatulae. G. Detailed view of the frictional seta (FS-I)fs, frictional setae (type FS-I/II); th, tactile seta.
and 10 � 103 mm2 on L2. Setal density is twice as high here as in therest of the scopula: 2.6 � 10�3 mm�2 on L1 and 3.2 � 10�3 mm�2 onL2. The total area of the tarsal scopula is 128 � 103 mm2 on L1 and118 � 103 mm2 on L2. Setae on all anterior legs, number about 200.Posterior legs lack such a distinct tarsal scopula, but small arrays ofspecialized setae can be found below the claws. These setae aresimilar to presumably friction-enhancing pretarsal setae, describedabove for both A. cuprea and O. heterophthalmus. These hairs occuronly in one row around the unguitractor plate and get longer inboth lateral and distal directions (from 60 mmup to 150 mm, Fig. 7C).They end in an imaginary plane orientated parallel to the ground, asin claw tufts of some other species studied. This arrangement alsoallows unhindered substrate contact by tarsal claws. A remarkablefeature demonstrated in this species is the co-occurrence of bothflattened and non-flattened setae in the pretarsus of L3 (Fig. 7B).There is a gradual transition between both types of setae: they
sive setae (AS-II). B. Pretarsus of L3 with a transition between typical adhesive (AS-I/II)L1 with ventro-lateral scopula. E. Tarsus of L4 lacking scopulae. F. Detail of the adhesivewith setules lacking spatulae. as, adhesive setae (type AS-I/II); ch, chemosensory setae;

J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 427
become more flat towards the distal parts of the pretarsus (next tothe claws). This transition correlates positively with the formationof spatulae.
The function of the described pads is assumed to be diverse.Adhesion to the prey during prey capture is presumably the mainfunction of strongly-developed tarsal scopula in anterior legs.However, the L1 claw tuft also supports locomotion on walls andceilings.
In D. praeficus, the morphology of setae is similar to those of theposterior legs in Z. subterraneus. Pretarsi bear single, non-flattened,spatula-lacking setae with a variable length of between 45 mm and210 mm, while the long ones are located on the posterior legs. Thesesetae are slightly S-shaped and covered with 3e7 mm long setules,especially on their ventral sides. Setae are irregularly distributed onanterior legs. Between 3 and 10 individual setae were found onanterior legs. On L3 and L4, the setae are arranged in a pattern,described above for L4 leg pair in Z. subterraneus, while the lateralsetae, located next to the claws, are much longer in D. praeficus.These setae are distally-bent, so that a large part of their ventralside comes into the contact with the substrate. The arrangement ofsetae leaves a wide open space for the tarsal claws. Setae are largerfrom the anterior to posterior part of the tuft. The tarsal scopula isweakly developed in the front legs. Flattened adhesive setae areabsent in pretarsi of all legs.
Claws should play a major role in attachment of these species.The claw-interlocking mechanism may be supported by adhesiveand frictional setae. In Z. subterraneus, prey gripping is presumablymainly provided by tarsal scopula, while this does not seem to bethe case in D. praeficus.
Philodromus cespitum WALCKENAER 1802(Philodromidae)Tibellus oblongus WALCKENAER 1802 (Philodromidae)Thanatus formicinus CLERCK 1757 (Philodromidae)
Philodromids are inhabitants of the upper strata that can easilyrun on both smooth and rough substrates (Wolff and Gorb, 2012b).P. cespitum is a common spider with a variable body length (\\:4.0e7.0 mm, __: 3.5e5.0 mm), that is a wide-spread inhabitant ofbushes, forest edges, and gardens. It hunts, ambushing on sunnymeadow sites, on herbs, shrubs, and trees. T. oblongus is also anambusher with a characteristic slender body supporting its camou-flage on grass leaves. It reaches a body length of up to 8e10 mm inboth sexes. T. formicinus is a holoarctic species with a body length of5e7mm, that prefers low vegetation in sunny grasslands. In contrasttomost species of this family, thisparticular spider is anactivehunter.
All three species have similar looking claw tufts and leg sco-pulae. Claw tufts are of an oval shape, having up to one third higherlength thanwidth. At the proximal base of the median cleft, there isa dense cluster of long slender tapered setae. Tips of adhesive setaeare rounded and show an arrangement previously described for theSalticidae. The comparably large size of the setae correlates wellwith the relatively low density of their distribution(2.5e3.0� 10�3 mm�2). An analogous situationwas seen at a higherhierarchical level: setule density is relatively low (0.9e1.3 mm�2),but spatulae reach widths of from 0.80 mm up to 1.20 mm.
In P. cespitum claw tufts are about 17 � 103 mm2 on L1 and L3,14�103 mm2 on L2, and 21�10�3 mm2 on L4. Setal density does notdiffer very much between legs. Claw tuft areas in the T. oblongusmale are 12 � 10�3 mm2 on L1-3 and 20 � 10�3 mm2 on L4. In thefemale, they are 13 � 103 mm2 onL1, 12 � 103 mm2 on L2,10 � 103 mm2 on L3, and 17 � 103 mm2 on L4. Setal density is about3.0 � 10�3 mm�2 on L1-3 of both sexes. On L4, it is lower(2.1 � 10�3 mm�2 in the male and 2.5 � 10�3 mm�2 in the female),but the setae are also larger there. In T. formicinus, the claw tuftareas are the largest ones (about 24 � 103 mm2), without notable
differences between the legs. Setal number is about one thirdhigher on L1 and L3 than on L2 and L4, respectively. Setal density ishigher there, and setae are more slender.
P. cespitum setae are the largest ones in the representatives ofthe family studied. Setal width at the broadest part of the setareaches up to 40 mm in the distal claw tuft part of anterior legs. Onaverage, it is 25 mm in the distal part of the claw scopula and about15 mm in the proximal part. Remarkably, setules of these large setaealso have the largest spatula width (1.02 mm on average) of allanalyzed species. Setule density is about 0.9 mm�2 in distal and1.3 mm�2 in proximal claw tuft parts. In the distal claw tuft part ofT. oblongus, setal width is,on average, 16e20 mm on L1-3 and about22 mm on L4. Setae in the proximal tuft part are, on average,12e16 mmwide. The female generally has 1 mmhigher values in thedistal claw tuft part and up to 3 mm lower values in the proximalone. Setules were analyzed only in the proximal claw scopula of thefemale L1: setule density is 1.3 mm�2, and an average spatulaewidth is 0.73 mm. In T. formicinus, setal width is, on average, 17.5 mmin the distal claw tuft part of L1 and L3, about 20 mm in that of L2,and 21 mm in that of L4. In the proximal claw tuft part, setae are, onaverage, 14 mm wide. Setule density is 1.3e1.6 mm�2, and spatulawidth is 0.6e0.9 mm.
Scopula setae cover wide parts of both tarsus and metatarsus inall three species studied (Fig. 8C). Setae in these regions havea similar structure to those of the claw tuft, but their adhesive sidesface proximally towards the leg.
Micrommata virescens CLERCK 1757 (Sparassidae)
This typically tropical spider family is represented in centralEurope by only this one species. These are mainly large arborealspiders that actively search for prey. M. virescens is a diurnal foliagerunner that reaches a body length of 12e15 mm (\\) and 7e10 mm(__). It prefers densevegetation, and is active inwarmandsunnysites.
The analysed specimen is a juvenile with not yet fully developedscopula, however, the scopula is strongly extended. The claw tufthas an area of about 40 � 103 mm2 on L1, 50 � 103 mm2 on L2,45 � 103 mm2 on L3, and 59 � 103 mm2 on L4. The shape of the clawtuft is nearly rhombic (Fig. 9A). Setal density is rather high:6.0� 10�3 mm�2 on L1 and L4, 7� 10�3 mm�2 on L2, and 8� 10�3 onL3. Setae are slender and remarkably long (up to 150 mm) and havecharacteristically-shaped ends with little clefts in the middle of thedistal edge (Fig. 9B). Setal width is 7e18 mm. In the distal part of theclaw tufts, setal tips are, on average, 13 mm wide, except on L2where they are slightly broader (15 mm). In the proximal part, theaverage widths are 10 mm (L1) and 11 mm (L2). Setule density iscomparably high with about 4 mm�2 in the proximal part of theclaw tuft, while spatulae are rather small (0.3 mm wide).
Low dense tarsal scopula occurs in all legs of M. virescens. Itssetae are long, slender, flattened, and with round tips.
Ebrechtella tricuspidata FABRICIUS 1775 (Thomisidae)
The Thomisidae (crab spiders) are typical members of theambusher feeding guild. E. tricuspidata is a small, mainly arborealthomisid (\ 6 mm, _ 3 mm) that is found in deciduous forests.Attachment is important for this spider, since surfaces of tree andshrub leaves are often smooth, and preventing falling down whilehandling struggling prey could be an important selective pressuretowards evolving effective attachment devices. Surprisingly, thisspider does not have distinct claw tufts. Compared with P. cespitumand T. oblongus, having a similar biology to E. tricuspidata, the sco-pula of the latter species differs to a high extent from the other twomentioned. Setae are flattened and covered ventrally with spatula-less setules (Fig.10B). Setae are tapered, slender, and relatively long,

Fig. 8. Philodromidae, ventral view. A. Pretarsus of Philodromus cespitum, L2, female. BeC. Thanatus formicinus, female. B. Pretarsus of L3. C. Distal parts of L1. DeE. Tibellus oblongus,female. D. Pretarsus of L1. E. Detailed view of adhesive setae of the claw tuft (type AS-II). F. Detail of setules in P. cespitum, female. ch, chemosensory seta; cl, tarsal claw; ct, claw tuft;ms, metatarsal scopula; sb, group of tapered setae (of type FS-II); th, tactile setae; ts, tarsal scopula.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433428
so they look like those described above in O. heterophthalmus.However, in E. tricuspidata, the density of setules is much higher.
E. tricuspidatahasabout30scopulahairs on theanteriorpartof thepretarsus and about 65 on the posterior one. There is a gradualtransition between setule-covered flattened setae and smooth tarsalsetae with a round cross section. Scopula setae mainly occur ina semicircle around the pretarsal edge, beneath the claws, and aregrouped together in three distinct bundles (Fig. 10A,C). Setae are60e130 mm long. The average length is 105 mm on L1 and L4, 111 mmon L3, and 86 mm on L3. Setae width is 4e7 mm (on average 5 mm).Setuleshave a lengthof about4.5mm.Setuledensity isabout1.1mm�2.
The attachment ability of thomisids on smooth surfaces is ratherpoor (personal observation). This observation corresponds well tothe presence of spatula-less setae. Spiders gain additional safety byattaching themselves to the substrate with the dragline. On roughsurfaces, spatula-less setules may presumably provide someattachment due to interlocking with substrate asperities. As setuletips are much smaller than the tips of claw appendages, they mightprovide friction on surfaces with minute asperities which claws areunable to grasp.
Cupiennius salei KEYSERLING 1877 (Ctenidae)
C. salei is a model organism in arachnology. Together withLycosidae and Oxyopidae, the family Ctenidae belongs to the superfamily of Lycosoidea. It is less related to the Dionycha group
including most spiders studied in the present paper. However,many representatives of Lycosoidea also have well-developed clawtufts (Griswold et al., 2005).
C. salei is distributed over Middle and South America. It isa nocturnal ambushing hunter (body length: 35 mm (\\) and20e25 mm (__), preferably living on bromeliad plants; Barth et al.,1988). The adhesive ability of C. salei is impressive, as it can evenattach to smooth wet surfaces (personal observation), wherea hydroplaning effect strongly reduces adhesion (Wolff and Gorb,2012a). Although it may adhere to vertical glass for hours, it candetach very fast in response to running or flying prey.
The scopula covers large parts of tarsus, metatarsus and tibia inall legs. Claw tuft setae are relatively slender (width of 7e17 mm,mean 11.6 mm). Difference in setal width between the proximaland distal part of the claw tuft is low (distal: 12.4 mm; proximal:10.6 mm). Setae are very long, about 0.3 mm on the proximal clawtuft part and up to 1.0 mm on the distal one. Setae of the claw tuftare S-shaped, while setae of both tarsal andmetatarsal scopulae arerather straight (Fig. 11B). In accordance with the large size of thisspider, the claw tuft reaches an area of about 1 mm2 with setaenumbering about 2350. Claw tuft area slightly increases fromanterior to posterior legs: 933 � 103 mm2 on L1, 959 � 103 mm2 onL2, 991 �103 mm2 on L3 and 1161 �10�3 mm2 on L4. Setal density is2.4�10�3 mm�2, on average. It does not differ significantly betweenlegs, but it is over three times higher on the distal part of the tuft(5.0 � 10�3 mm�2) than on the proximal one (1.5 � 10�3 mm�2).

Fig. 10. Ebrechtella tricuspidata (Thomisidae), female. A. Pretarsus of L2, ventral view. B. Detailed view of the FS-II seta, ventral view. C: Pretarsus of L1, frontal view. ch, chemo-sensory seta; cl, tarsal claw; fs, frictional setae (type FS-II); th, tactile seta.
Fig. 9. Micrommata virescens (Sparassidae), juvenile individual. A. Pretarsus of L3, frontal view. B. Adhesive setae in the claw tuft of L3, ventral view. ch, chemosensory seta; cl, tarsalclaw; ct, claw tuft; th, tactile setae; ts, setae of the tarsal scopula.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 429
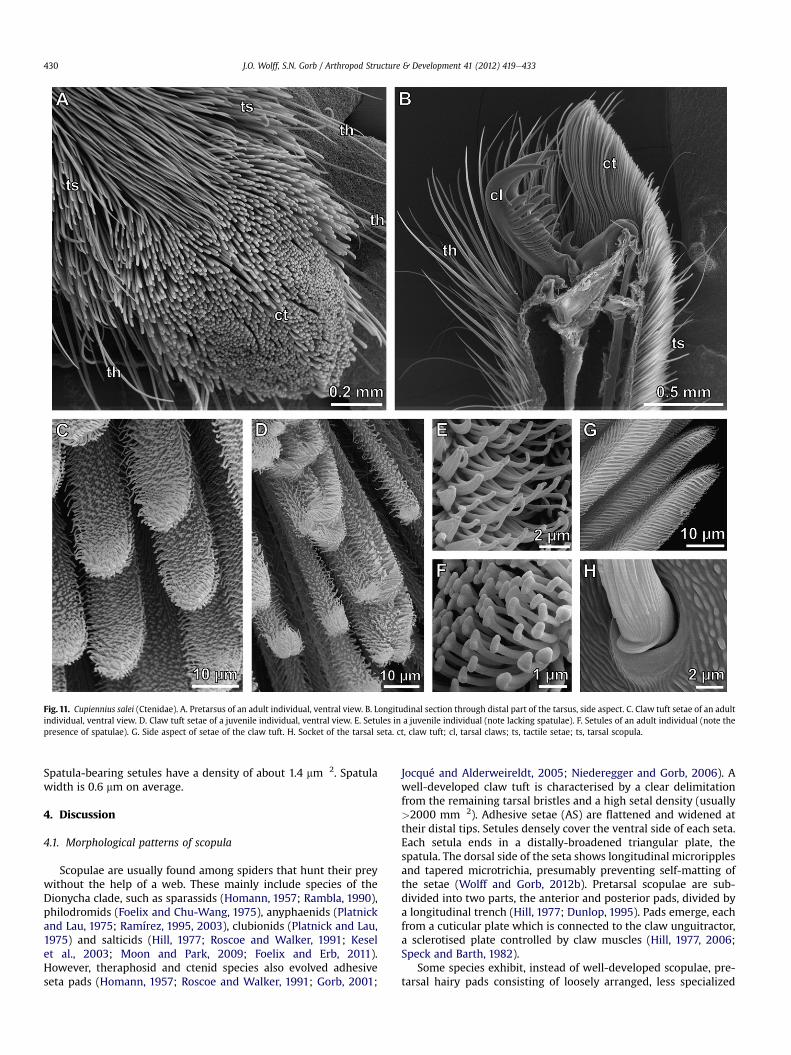
Fig. 11. Cupiennius salei (Ctenidae). A. Pretarsus of an adult individual, ventral view. B. Longitudinal section through distal part of the tarsus, side aspect. C. Claw tuft setae of an adultindividual, ventral view. D. Claw tuft setae of a juvenile individual, ventral view. E. Setules in a juvenile individual (note lacking spatulae). F. Setules of an adult individual (note thepresence of spatulae). G. Side aspect of setae of the claw tuft. H. Socket of the tarsal seta. ct, claw tuft; cl, tarsal claws; ts, tactile setae; ts, tarsal scopula.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433430
Spatula-bearing setules have a density of about 1.4 mm�2. Spatulawidth is 0.6 mm on average.
4. Discussion
4.1. Morphological patterns of scopula
Scopulae are usually found among spiders that hunt their preywithout the help of a web. These mainly include species of theDionycha clade, such as sparassids (Homann, 1957; Rambla, 1990),philodromids (Foelix and Chu-Wang, 1975), anyphaenids (Platnickand Lau, 1975; Ramírez, 1995, 2003), clubionids (Platnick and Lau,1975) and salticids (Hill, 1977; Roscoe and Walker, 1991; Keselet al., 2003; Moon and Park, 2009; Foelix and Erb, 2011).However, theraphosid and ctenid species also evolved adhesiveseta pads (Homann, 1957; Roscoe and Walker, 1991; Gorb, 2001;
Jocqué and Alderweireldt, 2005; Niederegger and Gorb, 2006). Awell-developed claw tuft is characterised by a clear delimitationfrom the remaining tarsal bristles and a high setal density (usually>2000 mm�2). Adhesive setae (AS) are flattened and widened attheir distal tips. Setules densely cover the ventral side of each seta.Each setula ends in a distally-broadened triangular plate, thespatula. The dorsal side of the seta shows longitudinal microripplesand tapered microtrichia, presumably preventing self-matting ofthe setae (Wolff and Gorb, 2012b). Pretarsal scopulae are sub-divided into two parts, the anterior and posterior pads, divided bya longitudinal trench (Hill, 1977; Dunlop, 1995). Pads emerge, eachfrom a cuticular plate which is connected to the claw unguitractor,a sclerotised plate controlled by claw muscles (Hill, 1977, 2006;Speck and Barth, 1982).
Some species exhibit, instead of well-developed scopulae, pre-tarsal hairy pads consisting of loosely arranged, less specialized
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 431
setae (FS). These are not or only slightly flattened and end ina pointed tip. They are distinguished from surrounding tactile hairs(TH) by a dense coverage of elongated microtrichia. In thesestructures, spatulae are lacking, but microtrichia often exhibita rectangular bent ending. Because of such specific tip geo-metry, these structures might increase friction on micro roughsurfaces due to interlocking with minute asperities rather thanincreasing adhesion by intermolecular forces. Presumably, speciesbearing such less-developed setules can hardly attach to smoothsurfaces.
In most spiders with claw tufts, a common distribution patternwas found. From anterior to posterior legs, total claw tuft areasincrease, while setal density is nearly constant or slightly decreases(Fig. 1). The expression of this distribution pattern is different inrepresentatives of different families: Clubionidae, Salticidae, Spar-assidae and Zoridae (Fig. 1A) show a higher differentiation betweenthe legs if compared to spiders with generally low setal density,such as Philodromidae and Gnaphosidae (Fig. 1B). During locomo-tion on a plane, L3 and L4 (posterior legs) have the function ofpushing the spider forward. In a resting position, L3 and L4 arepresumably more important for attachment to the substrate sinceL1 and L2 are often used to capture and manipulate prey.
Setae in the distal part of the claw tuft have larger dimensions(higher width and length) and lower setule density than those inthe proximal part. In addition, setal width is often higher inposterior legs, especially in species that show lower setal densityin these legs. This kind of possible compensation can also beobserved by comparing the structures between species. Forexample, setal width is often higher, if the setal density is compa-rably low. Species with lower setule density often show broaderspatulae (see Table 1). One may assume that these combinations ofdifferent features are adaptations to certain conditions of themicrohabitat.
Table 1Comparison of various characteristics of scopulae in the posterior leg (L4) in the specieapparent in all legs. (**) Setae of distal parts of the adhesive pad. (***) Average data of m
Species (family) Area[103 mm2]
Setal density[10�3 mm�2]
Setal ty(see Fig
LiocranidaeAgroeca brunnea 8.9 2.02 FS-IeIIA. cuprea 1.7 7.58 FS-I
AnyphaenidaeAnyphaena accentuata 14.1 2.21 FS-II, AS
ClubionidaeClubiona caerulescens 31.3 7.18 AS-I, AS
CtenidaeCupiennius salei 1160.7 2.34 AS-IeII
GnaphosidaeDrassyllus praeficus 7.9 1.26 FS-IZelotes subterraneus 9.8 1.84 FS-IeII,
ThomisidaeEbrechtella tricuspidata 15.6 2.88 FS-II
SalticidaeEvarcha arcuata [a] 33.9 [37.0] 5.84 (FS-II), AHeliophanus cupreus 11.5 6.07 (FS-II), A
SparassidaeMicrommata virescens 58.7 5.50 FS-II, AS
OxyopidaeOxyopes heterophthalmus 10.5 1.24 FS-I, FS-
PhilodromidaePhilodromus cespitum 21.2 2.64 FS-II, ASP. disparb 10.7 3.85 FS-II, ASThanatus formicinus 23.7 2.32 FS-II, ASTibellus oblongus 16.6 2.54 FS-II, AS
ZoridaeZora spinimana 15.1 7.82 FS-II, AS
a From Kesel et al. (2003).b From Wolff and Gorb (2012b).
4.2. Hypothetical evolutionary path of seta differentiation
An evolutionary trend of scopula seta development is shown inFig. 12. Adhesive setae are presumablymodified tactile hairs (Foelixet al., 1984). These mechanoreceptors are tapered, rather unstruc-tured setae bearing either none or only short microtrichia that arethen equally distributed around the whole setal shaft (Foelix andChu-Wang, 1973). A more derived seta type (FS-1) was found inAgroeca sp: microtrichia (setules) are asymmetrically distributedover the seta, more densely on the side facing the substrate. FS-IIsetae, found in A. brunnea, E. tricuspidata and O. heterophthalmusshow a further specialisation: seta are flattened and thus, contactarea to the substrate is enlarged.
Setae, found in Z. spinimana, (AS-I), show a hypothetical nextstep in further evolutionary development of these structures. Theystill have a tapered end on one side but are distally asymmetricallybroadened. This provides an additional area for setule coverage.Furthermore setal array can get denser as the adhesive area of theseta is shifted distally. The comparison of the setae from the distalpart of the claw tuft with setae from the proximal part shows thatbroadening resulted from an asymmetrical growth of each seta.This can also be observed in A. accentuata.
However, the most important innovation in spider adhesivesystemswas the spatulate tip of setula, responsible for tight contactformation with different substrates in different animal groups(Persson and Gorb, 2003; Varenberg et al., 2010), and therebycontributing to strong adhesion. It is already present inZ. spinimana. A further enhancement of adhesive function wasgained by symmetrical seta tips (AS-II), which are found in thewell-developed adhesive pads of salticids, philodromids, spar-assids, and clubionids. This might be due to a simultaneously fastergrowth of both lateral seta sides rather than of the median part. Anindication of this is the small median notch that occurs in some
s studied. (*) Underlined: dominating setal type in the claw tuft. In brackets: noteasured values in analysed legs. (n.a.) Not analysed.
pe. 11)
Setalwidth [mm] **
Setuledensity [mm�2] ***
Spatulawidth [mm] **
6.0 1.23 No spatula6.4 n.a. No spatula
-IeII 27.9 1.35 0.51
-III 15.1 3.48 0.28
13.1 1.43 0.60
6.3 n.a. n.a.AS-IeII 6.0 1.50 No spatula
5.1 1.13 No spatula
S-II 17.0 1.94 [2.10] 0.78S-II 13.9 2.83 0.49
-II 13.0 4.07 0.31
II 9.9 0.26 No spatula
-II 24.2 1.18 1.02-II 21.3 1.33 0.81-II 21.3 1.39 0.74-II 22.4 1.28 0.73
-I 10.6 2.47 0.56

Fig. 12. Pretarsal seta types shown in the 16 species studied. The series is presented in a sequence corresponding to a hypothetical evolutionary trend of increased complexity inscopula structure. Only the distal part of each seta is shown. TH, tactile seta (image: Evarcha arcuata); FS-I, frictional seta of type I (image: Agroeca cuprea); FS-II, frictional seta oftype II (image: Ebrechtella tricuspidata); AS-I, adhesive seta of type I (image: Zora spinimana); AS-II, adhesive seta of type II (image: Heliophanus cupreus); AS-III, adhesive seta of typeIII (image: Clubiona caerulescens). Features of setal types are listed below. Microtrichia: Seta is covered by microtrichia (setules). Anisotropy: There is an asymmetry between the sidefacing the substrate and the upside of seta according to length and distribution of microtrichia. Flattening: Seta is dorso-ventrally flattened. Spatula: Microtrichia (setules) bearspatular plates at their tips. Broad tip: Seta is distally broadened. Fringed tip: Distal edge of seta is subdivided into fringes. Triangular arrowheads indicate functional trends(increase of contact points, surface adaptability of seta, and friction and adhesion mechanisms contributing to the attachment function) superimposed by the hypotheticalevolutionary trend.
J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433432
setal tips in E. arcuata (Fig. 5D) and M. virescens (Fig. 9B). InC. caerulescens, setal tips are fringed (AS-III), which is a derivedcondition, that might further increase adhesive ability due to theaddition of a new hierarchical level. Salticids and philodromidshave evolved rounded setal tips, possibly allowing a smootherdetachment due to the gradual decrease of the peeling line width.This could support faster movements and contribute to enhancingmanoeuvrability. The extremely broadened setae of A. accentuatafollow another structural strategy that leads to the reduction ofhierarchy, presumably contributing to easier detachment.
Finally, both analysed gnaphosid species bear very primitivesetae that are not even flattened (FS-I). However, the tarsal scopulaof Z. subterraneus includes adhesive setae with a typical structure.Intermediate forms of both setal types provide an insight to howthe adhesive setae could have evolved. This clearly shows that thelack of adhesive setae must not necessarily mean an incapacity forbuilding these structures, but rather can be a structural reductiondue to the ground-living type of habitat.
4.3. Phylogenetic significance of the morphology of adhesive setae
Adhesive setae occur in spider taxa that are not necessarilyclosely-related, such as representatives of the families Ther-aphosidae, Ctenidae, and Salticidae. Nevertheless, they exhibit
rather similar morphological features (flattened setal tip with ananisotropic setule coverage and the presence of spatulae on thesetule tips). On the other hand, the present comparative study onsetal morphology has shown that the shape and size of thesestructures differs greatly indifferent spider taxa.While spiders livingin the upper strata are oftenwell equipped with claw tufts, ground-living species usually lack them or bear strongly reduced ones.
The Dionycha is a heavily discussed clade, with the Gnapho-soidea as the only more or less reliably-resolved super family(Coddington and Levi, 1991; Coddington, 2005). For instance, therelationship between Philodromidae and Thomisidae has beencontroversially discussed (Homann, 1975; Roberts, 1996; Benjaminet al., 2008). Homann (1975) used structural differences of scopulahairs between these two families as an argument against their closerelationship. However, a closer look reveals that philodromids bearthomisid-like setae (FS-II) forming a bundle at the base of the clawtuft (Fig. 8A,B,D), that could be a synapomorphic character for bothgroups. Such a bundle is also present in Z. spinimana (Fig. 2F),A. accentuata (Fig. 4A), and E. arcuata (Fig. 5E), although it is not asdistinct as in the philodromids analysed. In some legs it is totallyreduced. We assume that the morphology of the thomisid scopulasetae is a plesiomorphic character state, neither derived nor lost.The same might be true for oxyopid setae, which could demon-strate a plesiomorphic level of adhesive seta evolvement among the

J.O. Wolff, S.N. Gorb / Arthropod Structure & Development 41 (2012) 419e433 433
Lycosoidea. There is some evidence that thomisids are more closelyrelated to the Lycosoidea than the other dionychans (Homann,1975; Corronca and Terán, 2000). The similarity of pretarsal setaebetween thomisids and oxyopids supports this hypothesis.
Generally, characters of the setal morphology of the scopulahave previously been judged to be relatively insignificant forphylogenetics (Homann, 1975) due to their strong adaptive char-acter. In spite of this statement, they are often applied to phylo-genetic studies (Ramírez, 1995, 2003; Davila, 2003). Platnick andLau (1975) used the difference in the claw tuft structures todelimit Anyphaenidae from Clubionidae. Furthermore, theseauthors based their statement about the polyphyletic state of theClubionidae on the use of these characters. Our study has providedsome examples showing that functional adhesive setae could be animportant character in resolving spider phylogeny.
5. Conclusions
Specific morphology of hairy adhesive pads (scopulae) inspiders evolved in different evolutionary contexts under theinfluence of particular body size and as an adaptation to specificecological niches. Whereas pretarsal scopulae (claw tufts) mainlyoccur in speciesmoving fast on smooth surfaces of vegetation, well-developed leg scopulae can also be found in strictly ground-livingspecies, presumably as a function for capturing and manipulatingprey.
Acknowledgements
Dr. Valentina E. Gurjanova (Schmalhausen Institute of Zoologyof National Academy of Sciences of Ukraine, Kiev, Ukraine) hasprovided ethanol-fixated material for this study. We also thankProf. Dr. F.G. Barth and Clemens Schaber (Department of Neurobio-logy of the University of Vienna, Austria) for providing specimens ofC. salei. Victoria Kastner (Max Planck Institute for IntelligentSystems, Stuttgart, Germany) is acknowledged for improving thelanguage of an early version of the manuscript. This study wassupported by a grant from the German Science Foundation, DFG(project GO 995/10-1), to S.N.G.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.asd.2012.04.004.
References
Barth, F.G., Seyfarth, E.A., Bleckmann, H., Schüch, W., 1988. Spiders of the genusCupiennius Simon 1891 (Araneae, Ctenidae). I. Range distribution, dwellingplants, and climatic characteristics of the habitats. Oecologia 77, 187e193.
Benjamin, S.P., Dimitrov, D., Gillespie, R.G., Hormiga, G., 2008. Family ties: molecularphylogeny of crab spiders (Araneae: Thomisidae). Cladistics 24, 708e722.
Beutel, R.G., Gorb, S.N., 2001. Ultrastructure of attachment specializations ofhexapods (Arthropoda): evolutionary patterns inferred from a revised ordinalphylogeny. J. Zool. Syst. Evol. Res. 39, 177e207.
Chung, J.Y., Chaudhury, M.K., 2005. Roles of discontinuities in bio-inspired adhesivepads. J. R. Soc. Interface 2, 55e61.
Coddington, J.A., Levi, H.W., 1991. Systematics and evolution of spiders. Ann. Rev.Ecol. Syst. 22, 565e592.
Coddington, J., 2005. Phylogeny and classification of spiders. In: Ubick, D., Paquin, P.,Cushing, P.E., Roth, V. (Eds.), Spiders of North America. American ArachnologicalSociety, pp. 18e24.
Corronca, J.A., Terán, H.R., 2000. Optical structure of the crab spider Misumenopspallens (Araneae, Thomisidae). J. Arachn. 28, 16e22.
Davila, D.S., 2003. Higher-level relationships of the spider family Ctenidae(Araneae: Ctenoidea). Bull. AMNH 274, 1e86.
Dunlop, J.A., 1995. Movements of scopulate claw tufts at the tip of a tarantula spider.Netherlands J. Zool. 45, 513e520.
Foelix, R.F., Chu-Wang, I.-W., 1973. The morphology of spider sensilla. I.Mechanoreceptors. Tissue Cell 5, 451e460.
Foelix, R.F., Chu-Wang, I.-W., 1975. The structure of scopula hairs in spiders. In: Proc.6th Int. Arachnol. Congr. 1974. Free University Amsterdam, pp. 156e157.
Foelix, R.F., Jackson, R.R., Henksmeyer, A., Hallas, S., 1984. Tarsal hairs specialized forprey capture in the salticid Portia. Rev. Arachnol. 5, 329e334.
Foelix, R.F., Erb, B., 2011. Microscopical studies on exuviae of the jumping spiderPhidippus regius. Peckhamia 90, 1e15.
Forster, R.R., Platnick, N.I., 1985. A review of the austral spider family Orsolobidae(Arachnida, Araneae), with notes on the superfamily Dysderoidea. Bull. AMNH181, 1e230.
Gao, H., Ji, B., Jäger, I.L., Arzt, E., Fratzl, P., 2003. Materials become insensitiveto flaws at nanoscale: lessons from nature. Proc. Natl. Acad. Sci. 100,5597e5600.
Gasparetto, A., Seidl, T., Vidoni, R., 2009. A mechanical model for the adhesion ofspiders to nominally flat surfaces. J. Bionic. Engin. 6, 135e142.
Gorb, S.N., 2001. Attachment Devices of Insect Cuticle. Kluwer Academic Publishers,Dordrecht.
Gorb, S.N., Beutel, R.G., 2001. Evolution of locomotory attachment pads of hexapods.Naturwissenschaften 88, 530e534.
Griswold, C.E., Ramírez, M.J., Coddington, J.A., Platnick, N.I., 2005. Atlas ofphylogenetic data for entelegyne spiders (Araneae, Araneomorphae,Entelegynae) with comments on their phylogeny. Proc. Calif. Acad. Sci. 56,1e324.
Hill, D.E., 1977. The pretarsus of salticid spiders. Zool. J. Linn. Soc. 60, 319e338.Hill, D.E., 2006. The Pretarsus of Salticid Spiders. Version 3. Peckhamia
Epublications, pp. 1e41.Homann, H., 1957. Haften Spinnen an einer Wasserhaut? Naturwissenschaften 44,
318e319.Homann, H., 1975. The systematic position of the spider families Thomisidae and
Philodromidae (Chelicerata, Arachnida). Z. Morph. Tiere 80, 181e202.Jocqué, R., Alderweireldt, M., 2005. Lycosidae: the grassland spiders. Acta Zoologica
Bulgarica S1, 125e130.Kendall, K., 2001. Molecular Adhesion and Its Applications. Kluwer, New York.Kesel, A.B., Martin, A., Seidl, T., 2003. Adhesion measurements on the attachment
devices of the jumping spider Evarcha arcuata. J. Exp. Biol. 206, 2733e2738.Moon, M.J., Park, J.G., 2009. Fine structural analysis on the dry adhesion system of
the jumping spider Plexippus setipes (Araneae: Salticidae). Anim. Cells Syst. 13,161e167.
Niederegger, S., Gorb, S.N., 2006. Friction and adhesion in the tarsal and metatarsalscopulae of spiders. J. Comp. Physiol. A 192, 1223e1232.
Peattie, A.M., Full, R.J., 2007. Phylogenetic analysis of the scaling of wet and drybiological fibrillar adhesives. Proc. Natl. Acad. Sci. USA 104, 18595e18600.
Pekár, S., �Sobotník, J., Lubin, Y., 2011. Armoured spiderman: morphological andbehavioural adaptations of a specialised araneophagous predator (Araneae:Palpimanidae). Naturwissenschaften 98, 593e603.
Persson, B.N.J., Gorb, S.N., 2003. The effect of surface roughness on the adhesion ofelastic plates with application to biological systems. J. Chem. Phys. 119,11437e11444.
Platnick, N.I., Lau, A., 1975. A revision of the celer group of the spider genusAnyphaena (Araneae, Anyphaenidae) in Mexico and Central America. Am. Mus.Novit. 2575, 1e36.
Rambla, M., 1990. Les scopula des Opilions, differences avec les scopula desAraignées (Arachnida, Opiliones, Araneae). Bull. Soc. Europ. Arachnol. 1,293e298.
Ramírez, M.J., 1995. A phylogenetic analysis of the subfamilies of Anyphanidae(Arachnida, Araneae). Ent. Scand. 26, 361e384.
Ramírez, M.J., 2003. The spider subfamily Amaurobioidinae (Araneae,Anyphaenidae): a phylogenetic revision at the generic level. Bull. AMNH 277,1e262.
Roberts, M.J., 1996. Spiders of Britain and Northern Europe. HarperCollinsPublishers, London.
Roscoe, D.T., Walker, G., 1991. The adhesion of spiders to smooth surfaces. Bull. Br.Arachnol. Soc. 8, 224e226.
Rovner, J.S., 1978. Adhesive hairs in spiders: behavioral functions and hydraulicallymediated movement. Symp. Zool. Soc. Lond. 42, 99e108.
Scherge, M., Gorb, S.N., 2001. Biological Micro- and Nanotribology: Nature’s Solu-tions. Springer, Berlin Heidelberg New York.
Speck, J., Barth, F.G., 1982. Vibration sensitivity of pretarsal slit sensilla in the spiderleg. J. Comp. Physiol. 148, 187e194.
Ubick, D., Vetter, R.S., 2005. A new species of Apostenus from California, with noteson the genus (Araneae, Liocranidae). J. Arachn. 33, 63e75.
Varenberg, M., Pugno, N.M., Gorb, S.N., 2010. Spatulate structures in biologicalfibrillar adhesion. Soft Matter 6, 3269e3272.
Wolff, J.O., Gorb, S.N., 2012a. The influence of humidity on the attachment ability ofthe spider Philodromus dispar (Araneae, Philodromidae). Proc. R. Soc. B 279,139e143.
Wolff, J.O., Gorb, S.N., 2012b. Surface roughness effects on attachment ability of thespider Philodromus dispar (Araneae, Philodromidae). J. Exp. Biol. 215, 179e184.





























