Embed Size (px)
Citation preview

Comparative survey of early embryogenesis ofSecernentea (Nematoda), with phylogeneticimplications
C. Dolinski, J.G. Baldwin, and W.K. Thomas
Abstract: Insight into the evolution of class Secernentea (Nematoda) for the purpose of providing a phylogenetic con-text for the modelCaenorhabditis elegansis being gained from the use of molecular character sets. Such phylogeniesprovide a framework for mapping the evolution of diversity in some early-development characters for 70 species and19 families of Secernentea. These characters include (i) whether AB and P1 blastomeres initially develop at the same(synchronous) or different (asynchronous) rates, (ii ) whether AB and P1 are initially aligned along the linear axis ofthe embryo (tandem pattern) or obliquely (rhomboidal pattern), and (iii ) whether the founder germ cell, P4, is estab-lished early, i.e., by the sixth cleavage, or later. Evolutionary polarity of characters was evaluated through outgroupcomparisons. From our data the following inferences are made. The derived character, late establishment of P4, evolvedprimarily in the ancestor of the monophyletic groups Diplogastrina, Rhabditina, and Panagrolaimidae. Asynchronousdevelopment is convergent, defining one clade of Tylenchina as well as Cephalobina, and also arising independently inAphelenchina. The rhomboidal embryo is ancestral to the tandem-pattern embryo that defines a second clade ofTylenchina. Early-embryo characters are congruent with the polyphyly of Cephalobina and Aphelenchina, as has beendemonstrated by molecular phylogenies. Many aspects of early embryogenesis, rather than being highly conserved,evolve at a rate appropriate to defining taxa within Secernentea.
Résumé: La compréhension de l’évolution de la classe des Secernentea (Nematoda) comme contexte phylogénétiquedu modèleCaenorhabditis elegansressort des ensembles de caractéristiques moléculaires. Ces phylogénies servent decadre à l’étude de l’évolution de la diversité de certains caractères du début du développement chez 70 espèces de 19familles de Secernentea. Lors de l’étude des caractères, il a falluc déterminer (i) si, au départ, les blastomères AB etP1 se développent au même rythme (synchrones) ou à des rythmes différents (asynchrones), (ii ) si au départ AB et P1s’alignent le long de l’axe linéaire de l’embryon (arrangement en tandem) ou s’ils sont obliques à l’axe (arrangementrhomboïde) et (iii ) si la cellule germinale fondatrice, P4, est établie tôt, dès le sixième clivage, ou plus tard. La pola-rité évolutive des caractères est évaluée par comparaison à des groupes externes. De nos données, nous pouvons con-clure que l’établissement tardif de P4, un caractère dérivé, est apparu d’abord chez l’ancêtre du groupe monophylétiqueDiplogastrina, Rhabditina et Panagrolaimidae, que le développement asynchrone qui définit un clade de Tylenchina, unautre de Cephalobina est un caractère convergent qui est aussi apparu indépendamment chez les Aphelenchina, et quel’embryon rhomboïde précède l’embryon en tandem qui définit un second clade de Tylenchina. Les caractères du débutde l’embryogenèse s’accordent avec l’hypothèse du polyphylétisme des Cephalobina et des Aphelenchina, tel que dé-montré par les phylogénies moléculaires. Plusieurs aspects du début de l’embryogenèse ne sont pas des caractères stati-ques, mais ils évoluent à un rythme qui permet de définir les taxons chez les Secernentea.
[Traduit par la Rédaction]94
Dolinski et al.Introduction
Nematodes are good models for exploring the evolution ofmorphology and development (Fitch 1997; Sternberg andFélix 1997; Dolinski et al. 1998; Félix 1999, 2000; Sommeret al. 1999; Félix et al. 2000). A focus of these nematode
model systems has been the class Secernentea, which in-cludes Caenorhabditis elegans(Maupas) Dougherty. Thisnematode is unique in that its evolutionary context can beexplored and compared because it is the only metazoan forwhich complete embryonic cell lineages have been mappedand sequencing of the genome has been completed; the nem-atode also hasbeen fully reconstructed by means of transmis-sion electronmicroscopy (Sulston et al. 1983; TheC. elegansSequencing Consortium 1998; White 1988). Independent molec-ular character sets, such as those from 18s rDNA and RNAPolymerase II, as expressed by phylogenetic trees, are ex-panding insight into the evolutionary history of Secernentea(Baldwin et al. 1997; Blaxter et al. 1998; Goldstein et al.1998; Dorris et al. 1999; Félix et al. 2000). These provide aframework for mapping the evolution of early-developmentcharacters in Secernentea, and determining, for example, which
Can. J. Zool. 79: 82–94 (2001) © 2001 NRC Canada
82
DOI: 10.1139/cjz-79-1-82
Received April 20, 2000. Accepted September 20, 2000.Published on the NRC Research Press website on December 21,2000.
C. Dolinski1 and J.G. Baldwin.2 Department of Nematology,University of California, Riverside, CA 92521, U.S.A.W.K. Thomas. School of Biological Sciences, University ofMissouri, Kansas City, MO 64110, U.S.A.
1Present address: Rua Armando Ritter Viana, Campos dosGoytacazes, RJ, 28015-060, Brazil.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:20 PM
Color profile: DisabledComposite Default screen

of these characters are conserved at a level that defines ma-jor taxa and which are highly convergent. A key feature thatallows comparisons to be made among Secernentea is thatcell lineages within the class are said to be largely invariant,and blast cells are conserved to the extent that cell homologiesare readily recognizable across taxa (Félix 1999; Félix et al.2000). This invariance in these nematodes and a few otherorganisms provides a powerful tool for examining evolutionaryprocesses (Freeman and Lundelius 1992; van den Biggelaarand Haszprunar 1996). Notably, invariance does not definethe phylum Nematoda; rather, some adenophorean nematodespecies (orders Enoplida and Mononchida) demonstrably lackearly fixed cleavage patterns, and this is plausibly consid-ered to be the ancestral state for the phylum (Malakhov1994; Voronov et al. 1998; Voronov 1999; Borgonie et al.2000).
When Sulston et al. (1983) investigated cell lineages ofC. elegans, they speculated that cell fates during early embryo-genesis may be conserved throughout Secernentea. Subse-quently, Skiba and Schierenberg (1992) observed differencesin the timing of cleavages (heterochrony) of some blastomeresrelative to others when comparing a few representative speciesof Cephalobina and Rhabditidae. They also noted changes inthe spatial arrangement of blastomeres, such as differencesin the axis of cleavage. Skiba and Schierenberg (1992) ob-served that these diverse early-development patterns “flowinto one common pattern” before gastrulation; one implica-tion is that the processes of early development evolve morerapidly than the later product of these processes (Schierenberget al. 1997; Goldstein et al. 1998). More recently, Schierenberg(2000) underscored the value of recognizing those develop-mental processes that can be used as diagnostic markers forunderstanding phylogenetic relationships through comparisonswith classical morphological andrecent molecular studies.These developmental characters includethose that are directlyobserved visually (e.g., the order of cleavage of blastomeres)and those that require experimental interference (e.g., are de-tected by interfering with normal development; Schierenberg2000).
Taxon-specific differences in visible patterns of earlyembryogenesis are particularly significant for understandingevolutionary processes because there is plausible linkage ofthese patterns to mechanisms of cell fate. Embryonic- andpostembryonic-stage investigations clearly demonstrate thatcell fate often depends on induction signals from particularadjacent cells, but the details of signaling processes mayvary even among closely related taxa (Schierenberg et al.1997; Sommer 1997; Sternberg and Félix 1997; Wiegnerand Schierenberg 1998, 1999; Félix 1999). For example, inCephalobidae, includingAcrobeloides nanus, the differentorder of cell division leads to a different spatial arrangementof cells that is incompatible with certain cell-to-cell interac-tions found inC. elegans(Bossinger and Schierenberg 1992,1996; Schierenberg et al. 1997). In many cases, altered tim-ing and arrangement of blastomeres are linked to profoundchanges in cell interactions that regulate cell fate. In othercases, where cell fate is determined autonomously or regu-lated by redundant mechanisms, there may be relatively lessselection pressure against changes in timing and pattern. Aspecialized instance of cell interactions is the induction ofthe posterior pole at the site of fertilization in Rhabditina
and Diplogastrina; in Cephalobina and Tylenchida (sole repre-sentativeMeloidogyne incognita), on the other hand, axis po-larity is determined independently of fertilization (Goldstein etal. 1998).
Interpretation of the diversity and evolution of early-developmentcharacters requires that they be scored for awide range of taxa within a meaningful taxonomic frame-work for Secernentea, such as is provided by molecular char-acters. In the present study we screened early-developmentpatterns, using criteria outlined in Fig. 1, in 37 species repre-senting 15 families of Secernentea, and this is expanded us-ing accounts of early cleavage in additional taxa from theliterature.
Definition of characters
Early versus late establishment of the germ foundercell, P4
We evaluate early establishment of P4 by the sixth cleavage(the seventh blastomere stage or before) versus postpone-ment of P4 until a later cleavage (i.e., the ninth blastomerestage or later) (Figs. 1I and 1J). It has been previously shownthat in slow-developingCephalobus(= Acrobeloides) nanus,P4 is present by the 6-cell stage, in contrast to rapidly devel-oping C. elegans, where P4 is present by the 24-cell stage(Schierenberg et al. 1997). In the present study, using thesixth cleavage as the point of separation for this character isa testable hypothesis (based on preliminary observations)that this binary treatment is congruent with higher taxa, wherehigher taxa are supported by independent molecular charac-ters.
Synchronous versus asynchronous rateWe score, at the four-cell stage, whether development of
the AB and P1 lineages is preceding at a synchronous or anasynchronous rate; if it is synchronous, the first four blasto-meres are from the same (second) generation cleavage (ABx,ABx, EMS, P2) and are therefore of about the same size(Figs. 1A–1D); if it is asynchronous, the first four blasto-meres are of different generations and are therefore of verydifferent sizes (Figs. 1E–1H). Use of “synchronous” in thisstudy is not intended to suggest that AB and P1 cleaveprecisely in unison, only that they both cleave prior to anysecond-generation cleavages.
Oblique versus linear arrangement of blastomeresAt the four-cell stage, we assess whether, immediately fol-
lowing cleavages of AB and P1, the arrangement of blasto-meres is oblique (a rhomboidal embryo) or linear (anterior-to-posterior tandem) (Figs. 1A–1D). These differences inarrangement have an impact on which blastomeres are adja-cent and are relevant to specific hypotheses of cell-to-cell in-duction. InC. elegansthe rhomboidal pattern at the four-cellstage has been shown, by artificially removing the eggshell,to be the result of the combined influence of orientation ofthe cleavage furrow and constraints imposed by the ellipticalshape of the eggshell (Hyman and White 1987). Since blasto-meres may migrate into offset positions prior to the nextcleavage, a meaningful comparison at the four-cell stagedepends on scoring the arrangement immediately after cleav-age.
© 2001 NRC Canada
Dolinski et al. 83
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:21 PM
Color profile: DisabledComposite Default screen
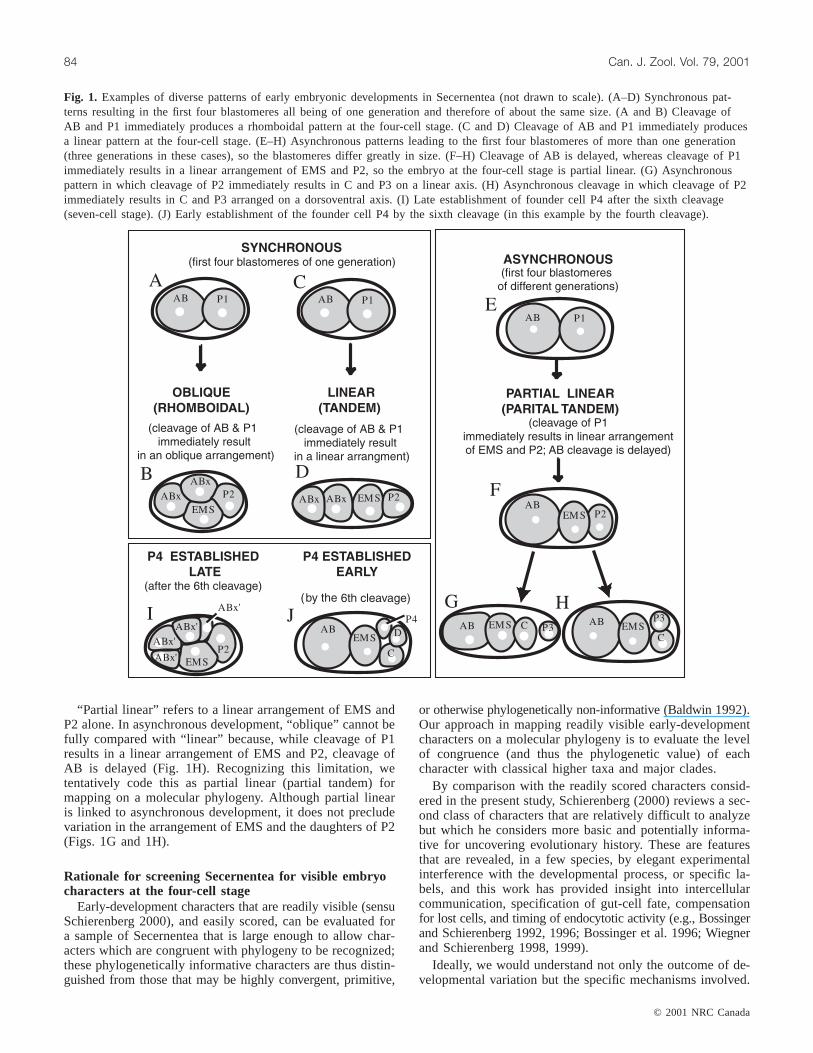
“Partial linear” refers to a linear arrangement of EMS andP2 alone. In asynchronous development, “oblique” cannot befully compared with “linear” because, while cleavage of P1results in a linear arrangement of EMS and P2, cleavage ofAB is delayed (Fig. 1H). Recognizing this limitation, wetentatively code this as partial linear (partial tandem) formapping on a molecular phylogeny. Although partial linearis linked to asynchronous development, it does not precludevariation in the arrangement of EMS and the daughters of P2(Figs. 1G and 1H).
Rationale for screening Secernentea for visible embryocharacters at the four-cell stage
Early-development characters that are readily visible (sensuSchierenberg 2000), and easily scored, can be evaluated fora sample of Secernentea that is large enough to allow char-acters which are congruent with phylogeny to be recognized;these phylogenetically informative characters are thus distin-guished from those that may be highly convergent, primitive,
or otherwise phylogenetically non-informative (Baldwin1992).Our approach in mapping readily visible early-developmentcharacters on a molecular phylogeny is to evaluate the levelof congruence (and thus the phylogenetic value) of eachcharacter with classical higher taxa and major clades.
By comparison with the readily scored characters consid-ered in the present study, Schierenberg (2000) reviews a sec-ond class of characters that are relatively difficult to analyzebut which he considers more basic and potentially informa-tive for uncovering evolutionary history. These are featuresthat are revealed, in a few species, by elegant experimentalinterference with the developmental process, or specific la-bels, and this work has provided insight into intercellularcommunication, specification of gut-cell fate, compensationfor lost cells, and timing of endocytotic activity (e.g., Bossingerand Schierenberg 1992, 1996; Bossinger et al. 1996; Wiegnerand Schierenberg 1998, 1999).
Ideally, we would understand not only the outcome of de-velopmental variation but the specific mechanisms involved.
© 2001 NRC Canada
84 Can. J. Zool. Vol. 79, 2001
Fig. 1. Examples of diverse patterns of early embryonic developments in Secernentea (not drawn to scale). (A–D) Synchronous pat-terns resulting in the first four blastomeres all being of one generation and therefore of about the same size. (A and B) Cleavage ofAB and P1 immediately produces a rhomboidal pattern at the four-cell stage. (C and D) Cleavage of AB and P1 immediately producesa linear pattern at the four-cell stage. (E–H) Asynchronous patterns leading to the first four blastomeres of more than one generation(three generations in these cases), so the blastomeres differ greatly in size. (F–H) Cleavage of AB is delayed, whereas cleavage of P1immediately results in a linear arrangement of EMS and P2, so the embryo at the four-cell stage is partial linear. (G) Asynchronouspattern in which cleavage of P2 immediately results in C and P3 on a linear axis. (H) Asynchronous cleavage in which cleavage of P2immediately results in C and P3 arranged on a dorsoventral axis. (I) Late establishment of founder cell P4 after the sixth cleavage(seven-cell stage). (J) Early establishment of the founder cell P4 by the sixth cleavage (in this example by the fourth cleavage).
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:21 PM
Color profile: DisabledComposite Default screen

The present broad survey will help to focus future work, in-cluding determining the mechanisms of development, usingrepresentatives that are appropriate to phylogenetic clades. Aphylogenetic overview of readily visible characters, such asorder of cleavage and which blastomeres are adjacent to oneanother, is basic to extending testable hypotheses concerningcell specification to phylogenetic groups that have not yetbeen targeted for more critical testing.
Materials and methods
Maintenance of nematode culture and observations ofembryos
The 37 species examined are listed in Table 1. All bacteria-feeders were grown on nematode growth medium (NGM) plates at25°C with Escherichia coli as the food source (Brenner 1974;Sulston and Hodgkin 1988). The fungus-feeders (Aphelenchina)Aphelenchoidessp. andAphelenchus avenaewere grown on potatodextrose agar (PDA) plates at 25°C withRhizoctonia solanias thefood source (S. Bartinicki-Garcia, personal communication). Theplant parasites (Tylenchida)M. incognita and Nacobbus aberranswere cultured on tomato plants as described by Souza and Baldwin(1998),Heterodera schachtiiwas grown on sugar beet plants, andBelonolaimus longicaudatuswas cultured on corn plants grown onB5 medium (Sigma) plates.Pratylenchus brachyuruswas culturedon sterilized carrot disks.
Isolate numbers were designated according to the protocol ofthe CaenorhabditisGenetics Center (CGC), with the initials of thelaboratory of origin followed by the laboratory serial number (e.g.,JB stands for James Baldwin), as shown in Table 1. Strains aredesignated as follows: PDL from P. De Ley (University of Ghent,Belgium), PS from P. Sternberg (California Technical Institute,Pasadena, California, U.S.A.), CW from P. Morgan (Case WesternReserve University, Cleveland, Ohio, U.S.A.), DF from D. Fitch(New York University, New York, U.S.A.), and SB from W. Sud-haus (Freier Universität, Berlin, Germany).Aduncospiculum halictiwas originally obtained from R. Giblin Davis (University of Florida,Fort Lauderdale, Florida, U.S.A.). The CGC providedC. elegans(N2). Living vouchers of each species are available in the respec-tive laboratories. Isolates of Aphelenchina and Tylenchina wereobtained from colleagues at the University of California (UCR)Riverside and are available through the Baldwin laboratory.
Agar pads 0.4 mm thick were prepared as described by Sulstonet al. (1983). Eggs of bacteria- and fungus-feeding nematodes, aswell the plant parasiteB. longicaudatus, were transferred fromagar plates at the one-cell stage, mounted on agar pads, coveredwith a cover glass, and sealed with petroleum jelly (Vaseline).Since some Rhabditidae do not lay eggs at the one-cell stage, theywere obtained by cutting gravid females in half under water in asmall dish. The eggs were then picked up and mounted as de-scribed above. One-cell-stage eggs of the plant parasitesM. incog-nita andN. aberranswere collected from egg masses and mountedon agar pads as described above.
For each species, three eggs were observed and photographed atthe four-cell stage with Tech pan 25 ASA film. The divisions werefollowed on the same eggs to determine the relative time of cleav-age to the founder germ cell, P4, and beyond until about the 20-cell stage. In addition, for each species the development of one ormore eggs was followed until hatching to demonstrate that theearly-development observations were not aberrant for that speciesand indeed that development of those individuals was normal.
Molecular analysisSequences of 18s rDNA were already available to this study for
a wide range for Secernentea from previous work, but this included
very few species of Tylenchida, and those included were highlyspecialized plant parasites (i.e., Blaxter et al. 1998; Goldstein et al.1998; W.K. Thomas, unpublished data). To improve the representa-tion of relevant taxa we added three species of Tylenchidae(Psilenchussp., Basiria sp., Coslenchussp.) and one species ofAnguinidae (Ditylenchus dipsaci) (Table 1, Tylenchina A inFig. 4); these taxa are generally considered basal to the order(Fortuner and Maggenti 1987; Geraert and Raski 1987; Luc et al.1987). Specimens were freshly collected and isolated by R.M.Souza from garden soil on the UCR campus, exceptD. dipsaci,which was obtained from garlic supplied by M. McKenry. Allspecimens are curated as vouchers in the UCR Nematode Collec-tion. Individual nematodes wereidentified by means of light micros-copy and one to several nematodes wereused for subsequent DNAextraction and sequence analysis. Template for the polymerasechain reaction (PCR) was prepared from nematodes, using thesingle-worm digestion buffer (Williams et al. 1992). DNA extract(2 µL) was used in subsequent amplifications of the 18s rDNAgene as in Baldwin et al. (1997). PCRproducts were purifiedfrom agarose gels using the Qiaquick gel extraction kits (QIAGEN,Valencia, Calif.). Sequencing reactions were performed usingfluorescent cycle sequencing chemistry (dRhodamine Terminators,ABI/Perkin–Elmer, Foster City, Calif.) and determined on a ABI377 automated sequencer. Alignments were performed by eye us-ing ESEE (Cabot and Beckenbach 1989). Phylogenetic analysesperformed on aligned sequences include neighbor joining asimplemented inMEGA (Kumar et al. 1993), maximum likelihood asimplemented inPHYLIP (Felsenstein 1993), andmaximum parsimonyas implemented inPAUP* (Swofford 1998).
Results
Among taxa of Secernentea, major groups vary with re-spect to the characters summarized in Fig. 1. These trendsare generally consistent within higher groups, includingsuborders and families (Table 1). The specific order of thefirst 11 cleavages for a representative of each higher group ispresented in Table 2; differences among species within thesehigher groups may include small variations in the position ofblastomeres or a reversal of the sequence of one or twocleavages, but do not impact the broader categories hereinconsidered as characters (Table 1).
CephalobinaCephalobina comprises three major families: Cephalobidae,
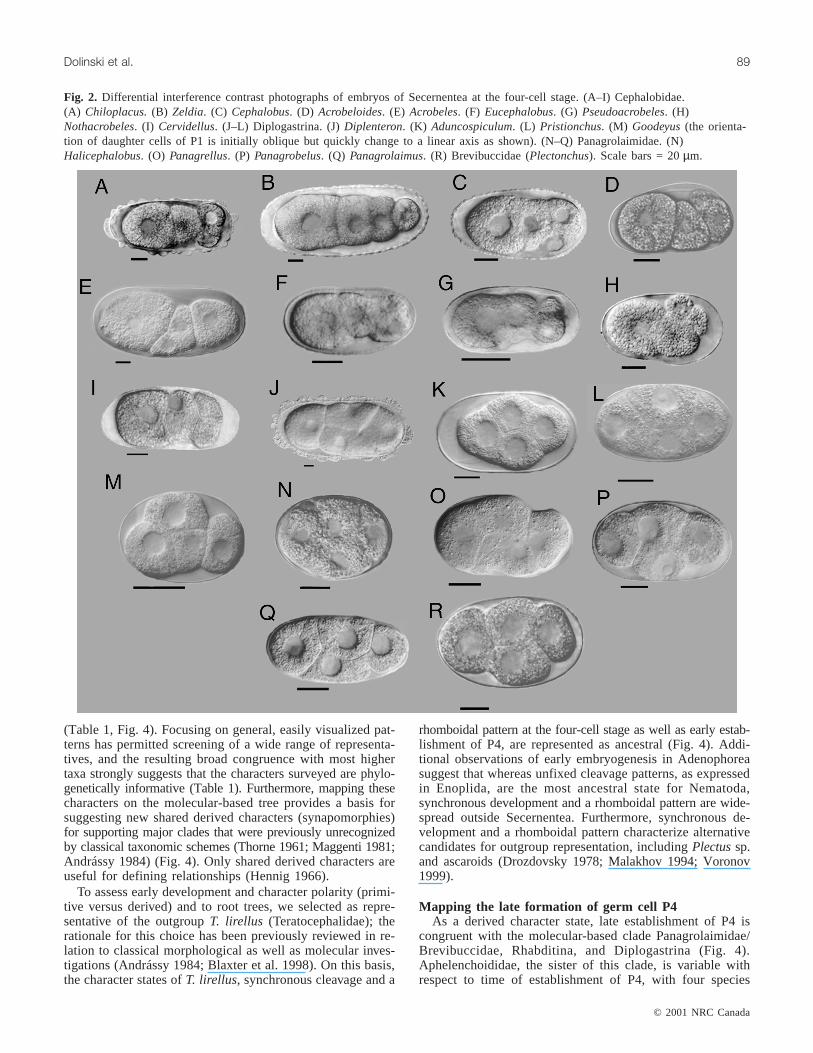
Panagrolaimidae, and Brevibuccidae (Table 1). We investigatednine species of Cephalobidae and in all of them blastomereswere unequal in size, owing to different cell generations be-ing represented at the four-cell stage, thus indicating asyn-chronous development (Figs. 2A–2I). The first cleavage ofthe fertilized egg results in a large anterior somatic foundercell (blastomere), AB, and a slightly smaller posterior germlinecell, P1. After that, P1 divides, immediately resulting in atandem arrangement of blastomere EMS and P2 (secondcleavage), and P2 divides into blastomere C and P3 (thirdcleavage). Blastomere P3 continues dividing into foundercells D and P4. Blastomere AB divides after the third cleav-age (e.g.,Cephalobus cubaensis). Therefore, at the four-cellstage the blastomeres present are AB, EMS, C, and P3(Fig. 1H, Table 2). Within Cephalobina there is variationamong species in the orientation of cells immediately aftercleavage, as well as subsequent migration of cells. Forexample, in most species examined, the daughter cells of P2are transverse or oblique to the anterior–posterior axis (Figs. 1F
© 2001 NRC Canada
Dolinski et al. 85
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:22 PM
Color profile: DisabledComposite Default screen
© 2001 NRC Canada
86 Can. J. Zool. Vol. 79, 2001
Suborder Family SpeciesbFeedinghabit/host
Early vs. lateestablishmentof P4c
Divisionorder
Blastomerearrangement Reference
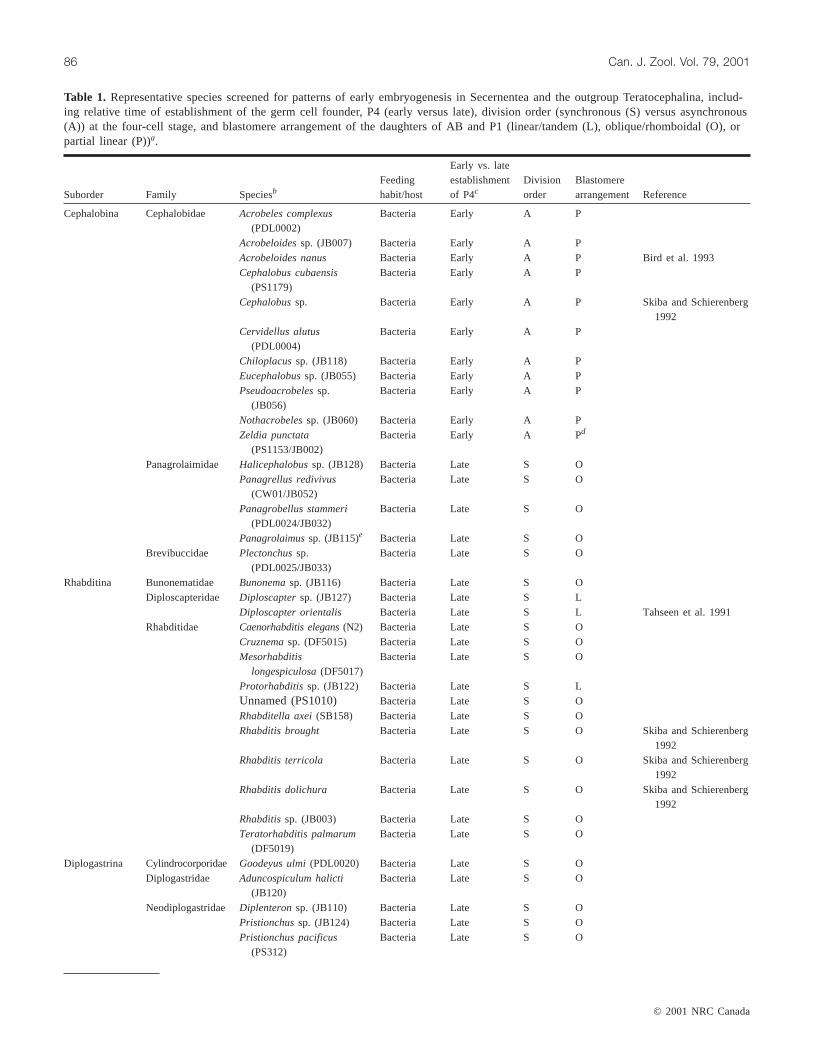
Cephalobina Cephalobidae Acrobeles complexus
(PDL0002)Bacteria Early A P
Acrobeloidessp. (JB007) Bacteria Early A PAcrobeloides nanus Bacteria Early A P Bird et al. 1993Cephalobus cubaensis
(PS1179)Bacteria Early A P
Cephalobussp. Bacteria Early A P Skiba and Schierenberg1992
Cervidellus alutus(PDL0004)
Bacteria Early A P
Chiloplacussp. (JB118) Bacteria Early A PEucephalobussp. (JB055) Bacteria Early A PPseudoacrobelessp.
(JB056)Bacteria Early A P
Nothacrobelessp. (JB060) Bacteria Early A PZeldia punctata
(PS1153/JB002)Bacteria Early A Pd
Panagrolaimidae Halicephalobussp. (JB128) Bacteria Late S OPanagrellus redivivus
(CW01/JB052)Bacteria Late S O
Panagrobellus stammeri(PDL0024/JB032)
Bacteria Late S O
Panagrolaimussp. (JB115)e Bacteria Late S OBrevibuccidae Plectonchussp.
(PDL0025/JB033)Bacteria Late S O
Rhabditina Bunonematidae Bunonemasp. (JB116) Bacteria Late S ODiploscapteridae Diploscaptersp. (JB127) Bacteria Late S L
Diploscapter orientalis Bacteria Late S L Tahseen et al. 1991Rhabditidae Caenorhabditis elegans(N2) Bacteria Late S O
Cruznemasp. (DF5015) Bacteria Late S OMesorhabditis
longespiculosa(DF5017)Bacteria Late S O
Protorhabditissp. (JB122) Bacteria Late S LUnnamed (PS1010) Bacteria Late S ORhabditella axei(SB158) Bacteria Late S ORhabditis brought Bacteria Late S O Skiba and Schierenberg
1992Rhabditis terricola Bacteria Late S O Skiba and Schierenberg
1992Rhabditis dolichura Bacteria Late S O Skiba and Schierenberg
1992Rhabditissp. (JB003) Bacteria Late S OTeratorhabditis palmarum
(DF5019)Bacteria Late S O
Diplogastrina Cylindrocorporidae Goodeyus ulmi(PDL0020) Bacteria Late S ODiplogastridae Aduncospiculum halicti
(JB120)Bacteria Late S O
Neodiplogastridae Diplenteronsp. (JB110) Bacteria Late S OPristionchussp. (JB124) Bacteria Late S OPristionchus pacificus
(PS312)Bacteria Late S O
Table 1. Representative species screened for patterns of early embryogenesis in Secernentea and the outgroup Teratocephalina, includ-ing relative time of establishment of the germ cell founder, P4 (early versus late), division order (synchronous (S) versus asynchronous(A)) at the four-cell stage, and blastomere arrangement of the daughters of AB and P1 (linear/tandem (L), oblique/rhomboidal (O), orpartial linear (P))a.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:23 PM
Color profile: DisabledComposite Default screen
and 1H). However, inZeldia punctata, the daughter cells ofP1 and P2 are oriented along the longitudinal axis. An impor-tant point is that these cells (AB, EMS, C, P3) are differentfrom those where linear cleavage is associated with synchro-nous development (ABx, ABx, EMS, P2) (Figs. 1D, 1G, and2B).
Four species of Panagrolaimidae were investigated and allshow synchronous development with blastomeres of the samegeneration, and a rhomboidal pattern (Figs. 1B, 2N–2Q).The blastomere AB divides almost at the same time as P1,giving rise to ABx, ABx, EMS, and P2. Establishment of P4is late; that is, P2 does not divide until the sixth cleavage
© 2001 NRC Canada
Dolinski et al. 87
Suborder Family SpeciesbFeedinghabit/host
Early vs. lateestablishmentof P4c
Divisionorder
Blastomerearrangement Reference
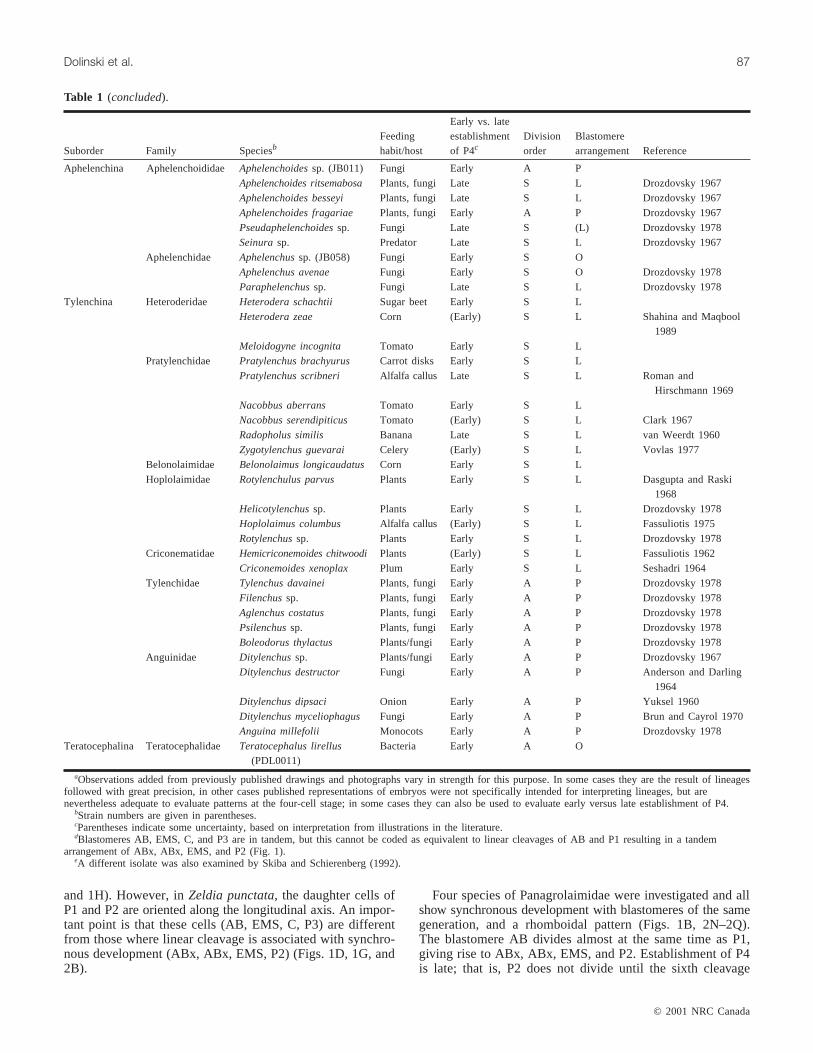
Aphelenchina Aphelenchoididae Aphelenchoidessp. (JB011) Fungi Early A PAphelenchoides ritsemabosa Plants, fungi Late S L Drozdovsky 1967Aphelenchoides besseyi Plants, fungi Late S L Drozdovsky 1967Aphelenchoides fragariae Plants, fungi Early A P Drozdovsky 1967Pseudaphelenchoidessp. Fungi Late S (L) Drozdovsky 1978Seinurasp. Predator Late S L Drozdovsky 1967
Aphelenchidae Aphelenchussp. (JB058) Fungi Early S OAphelenchus avenae Fungi Early S O Drozdovsky 1978Paraphelenchussp. Fungi Late S L Drozdovsky 1978
Tylenchina Heteroderidae Heterodera schachtii Sugar beet Early S LHeterodera zeae Corn (Early) S L Shahina and Maqbool
1989Meloidogyne incognita Tomato Early S L
Pratylenchidae Pratylenchus brachyurus Carrot disks Early S LPratylenchus scribneri Alfalfa callus Late S L Roman and
Hirschmann 1969Nacobbus aberrans Tomato Early S LNacobbus serendipiticus Tomato (Early) S L Clark 1967Radopholus similis Banana Late S L van Weerdt 1960Zygotylenchus guevarai Celery (Early) S L Vovlas 1977
Belonolaimidae Belonolaimus longicaudatus Corn Early S LHoplolaimidae Rotylenchulus parvus Plants Early S L Dasgupta and Raski
1968Helicotylenchussp. Plants Early S L Drozdovsky 1978Hoplolaimus columbus Alfalfa callus (Early) S L Fassuliotis 1975Rotylenchussp. Plants Early S L Drozdovsky 1978
Criconematidae Hemicriconemoides chitwoodi Plants (Early) S L Fassuliotis 1962Criconemoides xenoplax Plum Early S L Seshadri 1964
Tylenchidae Tylenchus davainei Plants, fungi Early A P Drozdovsky 1978Filenchussp. Plants, fungi Early A P Drozdovsky 1978Aglenchus costatus Plants, fungi Early A P Drozdovsky 1978Psilenchussp. Plants, fungi Early A P Drozdovsky 1978Boleodorus thylactus Plants/fungi Early A P Drozdovsky 1978
Anguinidae Ditylenchussp. Plants/fungi Early A P Drozdovsky 1967Ditylenchus destructor Fungi Early A P Anderson and Darling
1964Ditylenchus dipsaci Onion Early A P Yuksel 1960Ditylenchus myceliophagus Fungi Early A P Brun and Cayrol 1970Anguina millefolii Monocots Early A P Drozdovsky 1978
Teratocephalina Teratocephalidae Teratocephalus lirellus(PDL0011)
Bacteria Early A O
aObservations added from previously published drawings and photographs vary in strength for this purpose. In some cases they are the result of lineagesfollowed with great precision, in other cases published representations of embryos were not specifically intended for interpreting lineages, but arenevertheless adequate to evaluate patterns at the four-cell stage; in some cases they can also be used to evaluate early versus late establishment of P4.
bStrain numbers are given in parentheses.cParentheses indicate some uncertainty, based on interpretation from illustrations in the literature.dBlastomeres AB, EMS, C, and P3 are in tandem, but this cannot be coded as equivalent to linear cleavages of AB and P1 resulting in a tandem
arrangement of ABx, ABx, EMS, and P2 (Fig. 1).eA different isolate was also examined by Skiba and Schierenberg (1992).
Table 1 (concluded).
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:24 PM
Color profile: DisabledComposite Default screen
and P4 is not present until the eighth division (nine-cellstage) (Fig. 1I, Table 2). One species of the rare familyBrevibuccidae was examined and found to have the samepattern of cleavage as Panagrolaimidae (Fig. 2R).
DiplogastrinaRepresentatives of three families in the suborder Diplo-
gastrina were investigated: Cylindrocorporidae (one species),Diplogastridae (one species), and Neodiplogastridae (threespecies) (Table 1). All species have synchronous develop-ment as represented by four blastomeres of about the samesize and with a rhomboidal pattern (Figs. 1B and 2J–2L). InGoodeyus ulmi(Cylindrocorporidae), the orientation of thedaughter cells of P1 is initially oblique but quickly changesto along a linear axis (Fig. 2M). In all cases examined,founder cell P4 is established late, as in Panagrolaimidae(Table 2).
RhabditinaRepresentatives of three families in the suborder Rhabditina
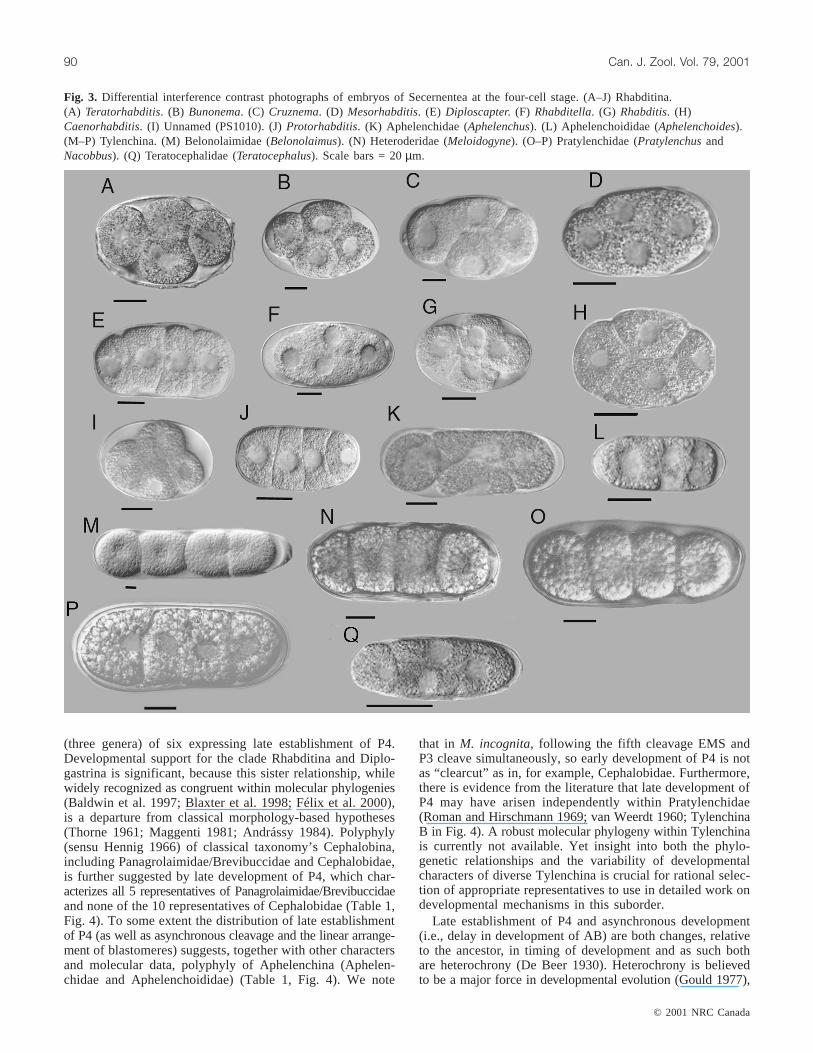
were investigated: Bunonematidae (one species), Diplo-scapteridae (one species), and Rhabditidae (eight species)(Table 1). Most of these have a synchronous pattern, withblastomeres organized in a rhomboidal pattern (Figs. 1B and3A–3I). Conversely, inDiploscaptersp. andProtorhabditissp. the blastomeres are organized linearly (Figs. 3E and 3J).In Rhabditina, founder cell P4 is established late, i.e., afterthe sixth cleavage, and in some cases much later (Table 2).Among the characters examined, the only one in whichRhabditina was observed to differ from Panagrolaimidae andBrevibuccidae was faster embryogenesis overall.
AphelenchinaRepresentatives of two families of Aphelenchina were in-
vestigated: Aphelenchidae and Aphelenchoididae (Table 1).Aphelenchussp. (Aphelenchidae) has a synchronous pattern,with blastomeres arranged in a rhomboidal pattern at thefour-cell stage (Figs. 1B and 3K); however,Aphelenchoidessp.(Aphelenchoididae) has an asynchronous pattern (Figs. 1Fand 3L). InAphelenchoidessp., P1 cleavage results in a tan-dem arrangement of EMS and P2 (second cleavage), and P2
divides into C and P3, which are organized on the dorsal–ventral axis (third cleavage). In bothAphelenchussp. andAphelenchoidessp., founder cell P4 is established early, atthe sixth cleavage (Table 2).
TylenchinaFive species in three families (Heteroderidae, Pratylenchidae,
and Belonolaimidae) were investigated. All have a synchro-nous pattern of development, with the first four blastomeresof about the same size and organized in a tandem pattern(Figs. 1D and 3M–3P). Blastomere P2 continues dividingbefore the two blastomeres ABx, thus founder cell P4 is es-tablished early, at the sixth cleavage or before (Table 2).
TeratocephalinaOne species,Teratocephalus lirellus, in the order Terato-
cephalina, was examined; it has a synchronous pattern, withthe first four blastomeres forming a rhomboidal pattern(Figs. 1B and 3Q) Founder cell P4 is established early, atthe sixth cleavage.
PhylogenyPhylogenetic analysis of Secernentea based on 18s rDNA
data is represented by a tree that divides Secernentea intotwo distinct clades, each strongly supported by bootstrapvalues (Fig. 4). Curiously, suborder Cephalobina is portrayedas polyphyletic, with the families Panagrolaimidae andBrevibuccidaedecoupled from Cephalobidae. Similarly, thesuborder Aphelenchina is represented as polyphyletic, withAphelenchidae decoupled from Aphelenchoididae. This treeprovides a basis for mapping and evaluating the evolution ofdiverse characters from early embryology.
Discussion
Diverse general patterns of early embryogenesis, definedin Fig. 1, are herein evaluated for 37 representative speciesfrom major groups (15 families) of Secernentea. Combinedwith observations from the literature, these characters arecoded for 70 species and 19 families as a basis for inferringcharacter evolution by mapping on the molecular-based tree
© 2001 NRC Canada
88 Can. J. Zool. Vol. 79, 2001
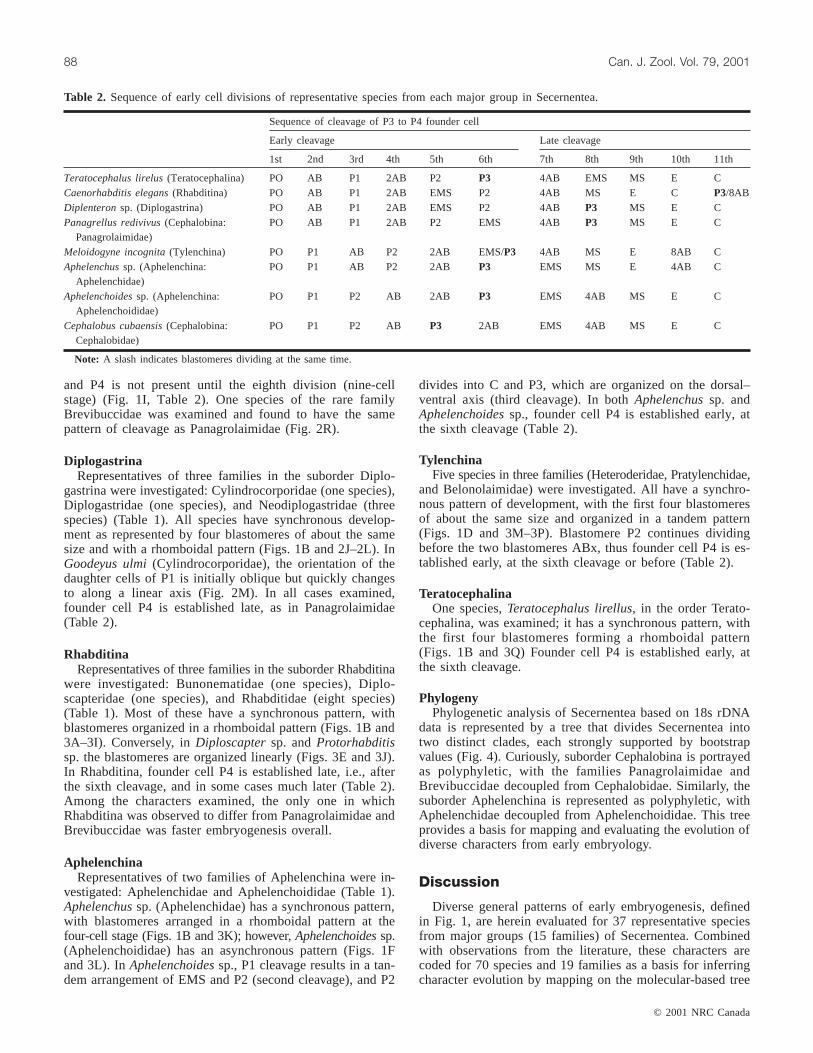
Sequence of cleavage of P3 to P4 founder cell
Early cleavage Late cleavage
1st 2nd 3rd 4th 5th 6th 7th 8th 9th 10th 11th
Teratocephalus lirelus(Teratocephalina) PO AB P1 2AB P2 P3 4AB EMS MS E CCaenorhabditis elegans(Rhabditina) PO AB P1 2AB EMS P2 4AB MS E C P3/8ABDiplenteronsp. (Diplogastrina) PO AB P1 2AB EMS P2 4AB P3 MS E CPanagrellus redivivus(Cephalobina:
Panagrolaimidae)PO AB P1 2AB P2 EMS 4AB P3 MS E C
Meloidogyne incognita(Tylenchina) PO P1 AB P2 2AB EMS/P3 4AB MS E 8AB CAphelenchussp. (Aphelenchina:
Aphelenchidae)PO P1 AB P2 2AB P3 EMS MS E 4AB C
Aphelenchoidessp. (Aphelenchina:Aphelenchoididae)
PO P1 P2 AB 2AB P3 EMS 4AB MS E C
Cephalobus cubaensis(Cephalobina:Cephalobidae)
PO P1 P2 AB P3 2AB EMS 4AB MS E C
Note: A slash indicates blastomeres dividing at the same time.
Table 2. Sequence of early cell divisions of representative species from each major group in Secernentea.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:25 PM
Color profile: DisabledComposite Default screen
(Table 1, Fig. 4). Focusing on general, easily visualized pat-terns has permitted screening of a wide range of representa-tives, and the resulting broad congruence with most highertaxa strongly suggests that the characters surveyed are phylo-genetically informative (Table 1). Furthermore, mapping thesecharacters on the molecular-based tree provides a basis forsuggesting new shared derived characters (synapomorphies)for supporting major clades that were previously unrecognizedby classical taxonomic schemes (Thorne 1961; Maggenti 1981;Andrássy 1984) (Fig. 4). Only shared derived characters areuseful for defining relationships (Hennig 1966).
To assess early development and character polarity (primi-tive versus derived) and to root trees, we selected as repre-sentative of the outgroupT. lirellus (Teratocephalidae); therationale for this choice has been previously reviewed in re-lation to classical morphological as well as molecular inves-tigations (Andrássy 1984; Blaxter et al. 1998). On this basis,the character states ofT. lirellus, synchronous cleavage and a
rhomboidal pattern at the four-cell stage as well as early estab-lishment of P4, are represented as ancestral (Fig. 4). Addi-tional observations of early embryogenesis in Adenophoreasuggest that whereas unfixed cleavage patterns, as expressedin Enoplida, are the most ancestral state for Nematoda,synchronous development and a rhomboidal pattern are wide-spread outside Secernentea. Furthermore, synchronous de-velopment and a rhomboidal pattern characterize alternativecandidates for outgroup representation, includingPlectussp.and ascaroids (Drozdovsky 1978; Malakhov 1994; Voronov1999).
Mapping the late formation of germ cell P4As a derived character state, late establishment of P4 is
congruent with the molecular-based clade Panagrolaimidae/Brevibuccidae, Rhabditina, and Diplogastrina (Fig. 4).Aphelenchoididae, the sister of this clade, is variable withrespect to time of establishment of P4, with four species
© 2001 NRC Canada
Dolinski et al. 89
Fig. 2. Differential interference contrast photographs of embryos of Secernentea at the four-cell stage. (A–I) Cephalobidae.(A) Chiloplacus.(B) Zeldia. (C) Cephalobus. (D) Acrobeloides. (E) Acrobeles. (F) Eucephalobus. (G) Pseudoacrobeles. (H)Nothacrobeles. (I) Cervidellus. (J–L) Diplogastrina. (J)Diplenteron. (K) Aduncospiculum. (L) Pristionchus. (M) Goodeyus(the orienta-tion of daughter cells of P1 is initially oblique but quickly change to a linear axis as shown). (N–Q) Panagrolaimidae. (N)Halicephalobus. (O) Panagrellus. (P) Panagrobelus. (Q) Panagrolaimus. (R) Brevibuccidae (Plectonchus). Scale bars = 20µm.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:35:52 PM
Color profile: DisabledComposite Default screen
(three genera) of six expressing late establishment of P4.Developmental support for the clade Rhabditina and Diplo-gastrina is significant, because this sister relationship, whilewidely recognized as congruent within molecular phylogenies(Baldwin et al. 1997; Blaxter et al. 1998; Félix et al. 2000),is a departure from classical morphology-based hypotheses(Thorne 1961; Maggenti 1981; Andrássy 1984). Polyphyly(sensu Hennig 1966) of classical taxonomy’s Cephalobina,including Panagrolaimidae/Brevibuccidae and Cephalobidae,is further suggested by late development of P4, which char-acterizes all 5 representatives of Panagrolaimidae/Brevibuccidaeand none of the 10 representatives of Cephalobidae (Table 1,Fig. 4). To some extent the distribution of late establishmentof P4 (as well as asynchronous cleavage and the linear arrange-ment ofblastomeres) suggests, together with other charactersand molecular data, polyphyly of Aphelenchina (Aphelen-chidae and Aphelenchoididae) (Table 1, Fig. 4). We note
that in M. incognita, following the fifth cleavage EMS andP3 cleave simultaneously, so early development of P4 is notas “clearcut” as in, for example, Cephalobidae. Furthermore,there is evidence from the literature that late development ofP4 may have arisen independently within Pratylenchidae(Roman and Hirschmann 1969; van Weerdt 1960; TylenchinaB in Fig. 4). A robust molecular phylogeny within Tylenchinais currently not available. Yet insight into both the phylo-genetic relationships and the variability of developmentalcharacters of diverse Tylenchina is crucial for rational selec-tion of appropriate representatives to use in detailed work ondevelopmental mechanisms in this suborder.
Late establishment of P4 and asynchronous development(i.e., delay in development of AB) are both changes, relativeto the ancestor, in timing of development and as such bothare heterochrony (De Beer 1930). Heterochrony is believedto be a major force in developmental evolution (Gould 1977),
© 2001 NRC Canada
90 Can. J. Zool. Vol. 79, 2001
Fig. 3. Differential interference contrast photographs of embryos of Secernentea at the four-cell stage. (A–J) Rhabditina.(A) Teratorhabditis. (B) Bunonema. (C) Cruznema. (D) Mesorhabditis. (E) Diploscapter. (F) Rhabditella. (G) Rhabditis. (H)Caenorhabditis. (I) Unnamed (PS1010). (J)Protorhabditis. (K) Aphelenchidae (Aphelenchus). (L) Aphelenchoididae (Aphelenchoides).(M–P) Tylenchina. (M) Belonolaimidae (Belonolaimus). (N) Heteroderidae (Meloidogyne). (O–P) Pratylenchidae (PratylenchusandNacobbus). (Q) Teratocephalidae (Teratocephalus). Scale bars = 20µm.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:36:23 PM
Color profile: DisabledComposite Default screen

and in nematodes, for example, lengthening of embryogenesisrelative to that ofC. elegansand other Secernenetea hasbeen shown to characterize Diplogastridae (Félix et al. 1999).In the present study, early establishment of founder P4 may,to some extent, be partially linked to asynchronous develop-ment, because in the latter case the P1 lineage (which leadsto P4) develops rapidly, whereas the AB lineage is retarded(Figs. 1G–1K). In general, many features of developmentmay be difficult to isolate as independent, and such “charac-ters” are problematic in parsimony-based cladistic phylogen-etic analysis. However, a useful approach to interpreting theevolution of interdependent characters is to map them onphylogenetic trees based on independent (e.g., molecular)character sets (Fig. 4).
Mapping asynchronous development and lineararrangement of blastomeres
The occurrence of an asynchronous early-development pat-tern, in which the P1 lineage proceeds more rapidly thanAB, is a derived state that characterizes Cephalobidae, in-cluding all 10 representatives considered. Combining the ob-servations from the present study with published illustrationsof additional representatives, mapping this character on themolecular-based tree suggests that asynchronous developmentevolved convergently in Tylenchina B (all of 10 representativespecies) and some Aphelenchoididae (Table 1, Fig. 4; An-derson and Darling 1964; Drozdovsky 1967, 1978; Yuksel1960).
The major branch of Tylenchina (Tylenchina B), includingmost plant parasites, is characterized by the derived patternof linear arrangement of the first four cells (synchronousblastomeres ABx, ABx, EMS, and P2), as demonstrated by16 representative species in five families (Table 2, Figs. 2and 4). In Pratylenchidae (Tylenchina B), some variation inthe arrangement of daughter cells of P1 is reported inRadopholus(van Werdt 1960), and inPratylenchusat thefour-cell stage the arrangement of the cells may change fromlinear to rhomboidal (Roman and Hirschmann 1969). A sec-ond branch (Tylenchina A), often considered basal to thesuborder (Fortuner and Maggenti 1987; Geraert andRaski1987; Luc et al. 1987), has asynchronous development and alinear arrangement only with respect to the cleavage of P1(P2, EMS) (i.e., partial linear) (Table 2, Fig. 4); this patternis consistent for the 10 species in two families examined.Whereas Cephalobina generally resembles Tylenchina Awith respect to an asynchronous pattern,Z. punctata(Cephalobina) appears to be an exception, in which the ar-rangement of the daughter cells of P1 and P2 results in apattern that is only superficially similar (partially linear) tothe true linear arrangement resulting from cleavages of ABand P1 in Tylenchida B (Figs. 1G, 2B, and 4).
Within Rhabditina, characterized by the primitive rhom-boidal arrangement at the four-cell stage,ProtorhabditisandDiploscapterhave a linear arrangement, as was previouslyshown withDiploscaptersp. (Drozdovsky 1978; Tahseen etal. 1991). Classical systematics varies in its classification ofProtorhabditis and Diploscapter, but generally recognizesthem as divergent and basal to most other Rhabditina; forexample, Andrássy (1984) recognizes the family Diplo-scapteridae outside Rhabditidae, and Protorhabditinae as adistinct subfamily. Sudhaus (1976), while notaddressingthe phylogenetic position ofDiploscapter, recognizesProtorhabditisas basal within “Rhabditinae sensu lato.” Re-cent 18s rDNA data and phylogenetic analysis of 31 speciesof Rhabditina suggest thatProtorhabditis and Diploscapterform an exclusive clade (Sudhaus and Fitch 2001). The rela-tionship of Protorhabditis and Diploscapter to additionalRhabditina, in conjunction with characters of early embryo-genesis, is currently being further investigated (F. Piano andD.H.A. Fitch, in preparation).
Mapping rhomboidal or linear patterns on phylogenetictrees can be refined as the mechanisms underlying thesegeneral patterns are better understood, to further explain thetrends and exceptions observed within major taxa (e.g., Ta-ble 1). These mechanisms include the relative roles of theaxis of cleavage and the influence of the eggshell (Hyman
© 2001 NRC Canada
Dolinski et al. 91
Fig. 4. Early embryonic development characters mapped on a18s rDNA-based tree. Numbers indicate bootstrap values at eachbranch. Derived states, determined on the basis of the outgroup,are depicted because shared derived states alone are useful indefining relationships. The derived state of late establishment ofP4 refers to P4 not being established until after the sixth cleav-age (Figs. 1I and 1J). The derived state of asynchronous cleav-age refers to the first four blastomeres being of differentgenerations because AB cleavage is retarded relative to P1(Figs. 1E–1H). The derived state of linear arrangement refers toABx, ABx, P2, and EMS positioned in tandem on the anterior–posterior axis of the embryo immediately following cleavage(Fig. 1D); P refers to the arrangement of EMS and P2 on theanterior–posterior axis, but AB cleavage being delayed, immedi-ately following cleavage (Figs. 1G and 1H). In the three columnsat the right, the values for each group are ratios of the numberof species representatives from Table 1 expressing the derivedstate to the number examined. “P” (partial linear) is includedseparately because, as explained in the text, its polarity is notdetermined. The outgroup representative isT. lirellus. Brokenlines define a synapomorphy based on late establishment of P4.Tylenchina B = Heteroderidae, Pratylenchidae, Belonolaimidae,Hoplolaimidae, and Criconematidae.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:36:25 PM
Color profile: DisabledComposite Default screen

and White 1987). Orientation of cells in a longitudinal con-figuration is generally linked to a relatively elongate egg,but at this point we cannot infer cause and effect; further-more, in some cases relatively short elliptical eggs alsohave a tandem arrangement of blastomeres (e.g.,Diplo-scapter, Protorhabditis; Figs. 3E and 3J).
Taxon-specific patterns of embryogenesis, including al-tered timing and arrangement of blastomeres, is particularlysignificant in relation to cell fate, based on interactions withadjacent cells (induction), versus subcellular events leadingto asymmetric divisions (autonomy), or redundant networksof interactions (Félix 1999). In Rhabditina and Diplogastrinaeven the first cleavage may be regulated as a result of a spe-cialized inductive determination of polarity (i.e., P1 versusAB), based on the point of sperm entry into the egg, and thischaracter and its alternative (maternal determination of eggpolarity) have been considered useful in defining the phylog-eny of Secernentea (Goldstein and Hird 1996; Goldstein etal. 1998). Previously, maternal determination of egg polaritywas considered a synapomorphy defining the classical taxon,Cephalobina (including Cephalobidae and Panagrolaimidae),but mapping these characters on the present tree requiresconvergent evolution of this early-development character, analternative also considered by Goldstein et al. (1998) (Fig. 4).
In conclusion, an overview of divergent patterns of earlyembryogenesis through a wide range of Secernentea pro-vides a basis for inferring the evolution of this diversity, andcan help to target where more intensive studies on specifica-tion mechanisms might be most informative for interpretingphylogeny (Félix 1999; Schierenberg 2000). With the ex-ception of axis of cleavage, most of the readily observed di-versity of early embryogenesis in Secernentea is related tochanges in timing, or heterochrony, which has been furtherconsidered to be a parameter for evolutionary classificationof nematodes (Skiba and Schierenberg 1992). Increasingly,however, it is clear that changes in early embryogenesis, anddevelopmental mechanisms in general can occur with littleimpact on morphological differentiation (Skiba and Schieren-berg 1992; Goldstein et al. 1998; Félix 1999; Schierenberg2000). Thus, early development, being much less constrainedthan wasonce believed to be the case, may prove to be in-creasingly useful for assessing evolution at lower taxonomiclevels within Secernentea, and particularly so as molecularphylogenies ofSecernentea are refined within complex sub-orders and families.
Acknowledgements
Among those that provided cultures, the authors particu-larly acknowledge the help of Mike Petrillo, Anton Ploeg,Paul De Ley, David Fitch, Ricardo Souza, and Sadia Bekal.A great friend, the late Michael Brzeski, is acknowledgedfor encouragement and supplying reprints. The authors thankRicardo Souza and Paul Sternberg for helpful comments.This research was supported by National Science Foundationgrants DEB-9318249 to J.G.B. and W.K.T. and DEB 97-12355 to S.A. Nadler and J.G.B. A full scholarship was pro-vided to the first author by the Brazilian Conselho Nacionalde Desenvolvimento Científico e Tecnológico. This paper ispart of the first author’s Ph.D. dissertation.
References
Anderson, R.V., and Darling, H. M. 1964. Embryology and repro-duction ofDitylenchus destructorThorne, with emphasis on go-nad development. Proc. Helminthol. Soc. Wash.31: 240–256.
Andrássy, I. 1984. Klasse Nematoda (Ordnungen Monhysterida,Desmoscolecida, Araeolaimida, Chromadorida, Rhabditida). GustavFischer Verlag, Stuttgart, Germany.
Baldwin, J.G. 1992. Evolution of cyst and noncyst-forming Hetero-derinae. Annu. Rev. Phytopathol.30: 271–290.
Baldwin, J.G., Giblin-Davis, R.M., Eddleman, C.D., Williams, D.S.,Vida, J.T., and Thomas, W.K. 1997. The buccal capsule ofAduncospiculum halicti(Nemata: Diplogasterina): an ultrastructuraland molecular phylogenetic study. Can. J. Zool.75: 407–423.
Bird, A.F., De Ley, P., and Bird, J. 1993. Morphology, ovopositionand embryogenesis in an Australian population ofAcrobeloidesnanus. J. Nematol.25: 607–615.
Blaxter, M.L., De Ley, P., Garey, J.R., Liu, X.L., Scheldeman, P.,Vierstraete, A., Vanfleteren, J.R., Mackey, L.Y., Dorris, M.,Frisse, L.M., Vida, J.T., and Thomas, W.K. 1998. A molecularevolutionary framework for the phylum Nematoda. Nature (Lond.),392: 71–74.
Borgonie, G., Jacobsen, K., and Coomans, A. 2000. Embryoniclineage evolution in nematodes. Nematology,2: 65–69.
Bossinger, O., Wiegner, O., and Schierenberg, E. 1996. Embryonicgut differentiation in nematodes—endocytosis of macromoleculesand its experimental inhibition. Roux’s Arch. Dev. Biol.205:494–497.
Bossinger, O., and Schierenberg, E. 1992. Cell–cell communica-tion in the embryo ofCaenorhabditis elegans. Dev. Biol. 151:401–409.
Bossinger, O., and Schierenberg, E. 1996. Cell–cell communicationin nematode embryos—differences betweenCephalobusspec andCaenorhabditis elegans. Dev. Genes Evol.206: 25–34.
Brenner, S. 1974. The genetics ofCaenorhabditis elegans. Genet-ics, 77: 71–94.
Brun, J., and Cayrol, J.C. 1970. Le développement embryonnairedu nematode mycophageDitylenchus myceliophagusGoodey,1958 (Nematoda–Tylenchida). Nematologica,l6: 523–531.
Cabot, E.L., and Beckenbach, A.T. 1989. Simultaneous editing ofmultiple nucleic acid sequences withESEE. Comput. Appl. Biosci.5: 233–234.
Clark, S.A. 1967. The development and life history of the falseroot-knot nematode,Nacobbus serendipiticus.Nematologica,13:91–101.
Dasgupta, D.R., and Raski, D.J. 1968. The biology ofRotylenchulusparvus. Nematologica,14: 429–440.
De Beer, G.R. 1930. Embryology and evolution. Clarendon Press,Oxford.
Dolinski, C., Borgonie, G., Schnabel, R., and Baldwin, J.G. 1998.Buccal capsule development as a consideration for phylogeneticanalysis of Rhabditida (Nemata). Dev. Genes Evol.208: 495–503.
Dorris, M.P., De Ley, P., and Blaxter, M.L. 1999. Molecular analysisof nematode diversity and the evolution of parasitism. Parasitol.Today,15: 188–193.
Drozdovsky, E.M. 1967. The use of embryonic development innematode taxonomy. Tr. Gel’mintol. Lab., Akad. Nauk SSSRTrans. Helminthol. Lab. Aca. Sci. U.S.S.R.18: 22–29.
Drozdovsky, E.M. 1978. The structure and formation of the pre-blastula as an indication of the phylogenetic relationships andtaxonomic position of various groups of nematodes.InFitgel’minthologicheskiye Issiedovaniya [Phytohelmintholresearch.]Edited byK.M. Ryzhikov. Nauka [Science], Moscow.pp. 14–32.
© 2001 NRC Canada
92 Can. J. Zool. Vol. 79, 2001
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:36:25 PM
Color profile: DisabledComposite Default screen

Fassuliotis, G. 1962. Life history ofHemicriconemoides chitwoodiEsser. Nematologica,8: 110–116.
Fassuliotis, G. 1975. Feeding, egg-laying, and embryology of theColumbia lance nematode,Hoplolaimus columbus. J. Nematol.7: 152–158.
Félix, M.A. 1999. Evolution of developmental mechanism in nem-atodes. J. Exp. Zool.285: 3–18.
Félix, M.A. 2000. Evolution of vulva development in the Cephalobina(Nematoda). Dev. Biol.221: 68–86.
Félix, M.A., Hill, R.J., Schwarz, H, Sternberg, P.W., Sudhaus, W.,and Sommer, R.J. 1999.Pristionchus pacificus, a nematode withonly three juvenile stages, displays major heterochronic changesrelative toCaenorhabditis elegans. Proc. R. Soc. Lond. B Biol.Sci. 266: 1617–1621.
Félix, M.A., De Ley, P., Sommer, R.J., Frisse, L, Nadler, S.A.,Thomas, W.K. Vanfleteren, J., and Sternberg, P.W. 2000. Evolu-tion of vulva development in the Cephalobina (Nematoda). Dev.Biol. 221: 68–86.
Felsenstein, J. 1993. PHYLIP (phylogeny inference package), ver-sion 3.5. University of Washington, Seattle.
Fitch, D.H.A. 1997. Evolution of male tail development in rhabditidnematodes related toCaenorhabditis elegans. Syst. Biol. 46:145–179.
Fortuner, R., and Maggenti, A.R. 1987. A reappraisal of Tylenchina(Nemata) 4. The family Anguinidae Nicoll, 1935 (1926). Rev.Nematol.10: 163–176.
Freeman, G., and Lundelius, J.W. 1992. Evolutionary implicationsof the mode of D quadrant specification in coelomates with spi-ral cleavage. J. Evol. Biol.5: 205–247.
Geraert, E., and Raski, D.J. 1987. A reappraisal of Tylenchina(Nemata) 3. The family Tylenchidae Örley, 1880. Rev. Nematol.10: 143–161.
Goldstein, B., Frisse, L.M., and Thomas, W.K. 1998. Embryonicaxis specification in nematodes: evolution of the first step in de-velopment. Curr. Biol.8: 157–160.
Goldstein B., and Hird, S.N. 1996. Specification of the anterio-posterior axis inCaenorhabditis elegans. Development,122: 1467–1474.
Gould, S.J. 1977. Ontogeny and phylogeny. The Belknap Press ofHarvard University Press, Cambridge, Mass.
Hennig, W. 1966. Phylogenetic systematics University of IllinoisPress, Urbana.
Hyman, A.A., and White, J.G. 1987. Determination of cell divisionaxes in the early embryogenesis ofCaenorhabditis elegans. J.Cell Biol. 105: 2123–2135.
Kumar, S., Tamaura, K., and Nei, M. 1993. MEGA: molecular evolu-tionary genetics analysis, version 1.01. The Pennsylvania StateUniversity, University Park.
Luc, M., Maggenti, A.R., Fortuner, R., Raski, D.J., and Geraert, E.1987. A reappraisal of Tylenchina (Nemata) 1: For a new approachto the taxonomy of Tylenchina. Rev. Nematol.10: 127–134.
Maggenti, A.R. 1981. General nematology. Springer-Verlag, NewYork.
Malakhov, V.V. 1994. Nematodes: structure development, classifi-cation, and phylogeny. Smithsonian Institution Press, Washing-ton, D.C.
Roman, J., and Hirschmann, H. 1969. Embryogenesis and postembryo-genesis in species ofPratylenchus(Nematoda: Tylenchidae).Proc. Helminthol. Soc. Wash.36: 164–174.
Schierenberg, E. 2000. Early development of nematode embryos:differences and similarities. Nematology,2: 57–64.
Schierenberg E., Wiegner, O., Bossinger, O, Skiba, F., and Kutzowitz,M. 1997. Pattern formation and cell specification in nematode
embryos: a theme with considerable variations. Zoology (Jena),100: 320–327.
Seshadri, A.R. 1964. Investigations on the biology and life cycle ofCriconemoides xenoplaxRaski, 1952 (Nematoda: Criconematidae).Nematologica,10: 540–562.
Shahina, F., and Maqbool, M.A. 1989. Embryonic and postembryonicdevelopmental stages ofHeterodera zeaeKoshy et al. 1971 onZea mays. Pak. J. Nematol.7: 3–16.
Skiba, F., and Schierenberg, E. 1992. Cell lineages, developmentaltiming, and spatial pattern formation in embryos of free-livingsoil nematodes. Dev. Biol.151: 597–610.
Sommer, R.J. 1997. Evolutionary changes of developmental mech-anisms in the absence of cell lineage alterations during vulvaformation in the Diplogastridae (Nematoda). Development,124:243–251.
Sommer, R.J., Sigrist, C.B., Grandien, K., Jungblut, B., Eizinger,A., Adamis, H., and Schlak, I. 1999. A phylogenetic interpreta-tion of nematode vulval variations. Invertebr. Reprod. Dev.36:57–65.
Souza, R.M., and Baldwin, J.G. 1998. Changes in esophageal glandactivity during the life cycle ofNacobbus aberrans(Nemata:Pratylenchidae). J. Nematol.30: 275–290.
Sternberg, P.W., and Félix, M.A. 1997. Evolution of cell lineage.Curr. Opin. Genet. Dev.7: 543–550.
Sudhaus, W. 1976. Vergleichende Untersuchengen zur Phylogenie,Systematik, Ökologie, Biologie und Ethologie der Rhabditidae(Nematoda). Zoologica (Stuttg.),43: 1–229.
Sudhaus, W., and Fitch, D. 2001. Comparative studies on the phylo-geny and systematics of the Rhabditidae (Nematoda). J. Nematol.In Press.
Sulston, J.E., and Hodgkin, J. 1988. Methods.In The nematodeCaenorhabditis elegans. Edited byW.B. Wood. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y. pp. 587–606.
Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N.1983. The embryonic cell lineage of the nematodeCaenorhabditiselegans.Dev. Biol. 100: 64–119.
Swofford, D.L. 1998. PAUP*, phylogenetic analysis using parsi-mony (*and other methods), version 4.0. Sinauer Associates,Sunderland, Mass.
Tahseen, Q., Jairapuri, M.S., and Ahmad, I. 1991. Observationson the embryonic and postembryonic development ofDiplo-scapter orientalis (Nematoda: Rhabditida). Rev. Nematol.14: 285–290.
The C. elegansSequencing Consortium (TCeSC). 1998. Genomesequence of the nematodeC. elegans: a platform for investigat-ing biology. Science (Washington, D.C.),282: 2012–2018.
Thorne, G. 1961. Principles of nematology. McGraw–Hill Inc.,New York.
van den Biggelaar, J.A.M., and Haszprunar, G. 1996. Cleavage pat-terns and mesentoblast formation in the Gastropoda: an evolu-tionary perspective. Evolution,50: 1520–1540.
van Weerdt, L.G. 1960. Studies on the biology ofRadopholussimilis (Cobb, 1893) Thorne, 1949. Part III. Embryology andpostembryonic development. Nematologica,5: 43–51.
Voronov, D.A. 1999. The embryonic development ofPontonemavulgare (Enoplida: Oncholaimidae) with a discussion of nema-tode phylogeny. Russ. J. Nematol.7: 105–114.
Voronov, D.A., Panchin, Yu.V., and Spiridonov, S.E. 1998. Nema-tode phylogeny and embryology. Nature (Lond.),395: 28.
Vovlas, N. 1977. Osservazioni sullo sviluppo embrionale diZygotylenchus guevarai. Nematol. Mediter.5: 121–124.
White, J. 1988. The anatomy.In The nematodeCaenorhabditiselegans. Edited byW.B. Wood. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y. pp. 81–122.
© 2001 NRC Canada
Dolinski et al. 93
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:36:26 PM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
94 Can. J. Zool. Vol. 79, 2001
Wiegner, O, and Schierenberg, E. 1998. Specification of gut cellfate differs significantly between the nematodesAcrobeloidesnanusand Caenorhabditis elegans. Dev. Biol. 204: 3–14.
Wiegner, O., and Schierenberg, E. 1999. Regulative developmentin a nematode embryo: a hierarchy of cell fate transformations.Dev. Biol. 215: 1–12.
Williams, B.D., Schrank, B., Huynh, C., Shownkeen, R., andWaterston, R.H. 1992. A genetic mapping system inCaenorhabditiselegansbased on polymorphic sequence-tagged sites. Genetics,131: 609–624.
Yuksel, H.S. 1960. Observations on the life cycle ofDitylenchusdipsaci on onion seedlings. Nematologica,5: 289–296.
J:\cjz\cjz79\cjz-01\Z00-179.vpSaturday, December 16, 2000 2:36:26 PM
Color profile: DisabledComposite Default screen






























