Embed Size (px)
Citation preview

Complete tumor prevention by engineered tumor cell vaccinesemploying nonviral vectors
Ines Moret-Tatay,1 Joaquın Dıaz,2 Francisco M Marco,2 Antonio Crespo,1 and SalvadorF Alino*,1
1Grupo de Terapia Genica, Departamento de Farmacologıa, Facultad de Medicina, Universitat de Valencia,Avda de Blasco Ibanez no. 15, 46010 Valencia, Spain; and 2ASAC Pharmaceutical International AIE Alicante,Spain.
We report that 100% mice survival after tumor challenge is achieved with cytokine-engineered cells employing nonviral lipoplexes
and without using viral vectors. We describe this effect with cytokine-secreting tumor cell vaccines, based on cell clones or fresh
transfected cells. Tumor cells were transfected with murine granulocyte–macrophage colony-stimulating factor (GM-CSF) or IL-4
plasmids employing the cationic lipid DOTAP, were irradiated (150 Gy) and kept frozen until use. The transfection efficacy was
analyzed by qRT-PCR and flow cytometry. Vaccination induced potent antitumor rejection, resulting in 100% mice survival.
Furthermore, the antitumor immunity was long lasting, since a two-fold survival delay was observed in mice after tumor rechallenge
(6 months later). While cell clones secreting GM-CSF were the most effective in wild-type tumor cell rejection, little or no effect was
observed with clones secreting IL-4. We found similar antitumor efficacy employing fresh transfected cells by nonviral procedures,
demonstrating that cells genetically modified by nonviral vectors (both clones and fresh transfected cells) are a safe and efficient
tool for antitumor vaccines. These vaccines allow us to achieve the highest antitumor efficacy based on nonviral gene therapy
techniques. In addition, the vaccination success with fresh transfected cells simplifies the procedure and provides new insights into
the clinical application of nonviral gene therapy procedures.
Cancer Gene Therapy (2003) 10, 887–897. doi:10.1038/sj.cgt.7700646
Keywords: vaccine; melanoma; GM-CSF; IL-4; nonviral vectors
Tumor cells can be genetically modified for differentpurposes, for instance to express immunostimulatory
substances (cytokines) that can stimulate the immunesystem against tumor cells.1,2 This approach is based onthe fact that in many cases tumor cells do not mediateimmune system activation, although they do presenttumor-associated antigens at the surface level.3 Theexpression of certain cytokines by these tumor cells canstimulate the immune system, resulting in cell eliminationby enhancing tumor immunogenicity with minimalsystemic toxicity.4 Certain immune reactions are involvedin tumor rejection, such as ‘‘cross-priming’’ processes,1,5
or direct antigen presentation by vaccine cells,6,7 resultingin the activation of CD8+ cytotoxic T (CTL) and CD4+helper T (Th) lymphocyte responses.8 Cross-primingprocesses imply cellular antigen transfer from tumor cellsto host antigen-presenting cells (APC) for processing andpresentation.9 This event has been shown to require T-cellhelp,10 the presence of transporters associated withantigen presentation (TAP) in tumor cells11 and
CD8a+ dendritic cells (DC).12,13 Another importantpoint to be considered is that the immune responseachieved by the vaccine depends on the immunostimula-tory protein secreted by the cell vaccines.1,14,15 Thus, thegranulocyte–macrophage colony-stimulating factor (GM-CSF), interleukin 4 (IL-4) and others cytokines1,5 havebeen demonstrated to exert antitumor effects, but IL-8has been described to act as a growth factor in humanmelanoma.16 In the present study, we have chosen GM-CSF and IL-4 for their purported different roles inimmune system stimulation.17–19 Accordingly, GM-CSFinduces high antibody titers and T-cell proliferation,20
even after allogeneic T-cell-depleted bone marrow trans-plantation.21 Cytokine IL-4 in turn promotes the devel-opment of Th 2 cells and increases the production ofantibodies and MHC class I and ICAM-1 expression intransfected cells,22 although it could suppress CTLactivity.8,23 In addition, the combination of GM-CSFand IL-4 would be particularly beneficial, since both GM-CSF and IL-4 induce growth expansions of phenotypi-cally and functionally different subpopulations of den-dritic cells.24
We expect the efficacy of vaccination to depend on theefficacy of gene transfer to cells.25 Since nonviral vectorsare growing in gene transfer efficiency, we hope that suchvectors could be appropriate systems for transfecting cells
Received March 2, 2003.
Address correspondence and reprint requests to: Dr Salvador F
Alino, PhD, Departamento de Farmacologıa, Facultad de Medicina,Universitat de Valencia, Avda. Blasco Ibanez 15, 46010 Valencia,
Spain. E-mail: [email protected]
Cancer Gene Therapy (2003) 10, 887–897r 2003 Nature Publishing Group All rights reserved 0929-1903/03 $25.00
www.nature.com/cgt

mediating an effective antitumor response. It has pre-viously been demonstrated that effective antitumorresponse against melanoma and other tumors can beobserved employing viral vectors targeted to modifiedtumor cells.26,27 However, nonviral gene therapy hasshown fewer approaches to cell vaccine applications. Theaim of achieving similar antitumor responses withnonviral vectors is very important, since it allows thechoice of using this safer antitumor therapy.28 Wetherefore studied the effect of vaccination on syngeneicmice survival employing tumor cell clones and freshlytransfected tumor cells producing GM-CSF or IL-4.Different doses of irradiated vaccines were employed(from 0.2 to 2 million cells per mouse), resulting in 80–100% mice survival with GM-CSF cytokine-producingcells, but not with IL-4. Our results suggest that vaccinesbased on nonviral gene therapy can be an efficient andsafe procedure with possible future clinical applications.
Materials and methods
Plasmids
Plasmids containing murine GM-CSF or IL-4 genes,controlled by a cytomegalovirus promoter, were obtainedfrom Dr A Koenig (Mologen, Germany). The mGM-CSF(425 bp) was inserted into a 3853 kb pRL-CMV-modifiedplasmid with resistance to Kanamycin, and mIL-4(438 bp) in a pCDNA3 (5414 kb) with Neomycin- andAmpicillin-resistance genes.The plasmids were amplified in Escherichia coli
(DH5a) and purified with the Qiagen Maxi protocol kit(Izasa SA, Spain).
Animals
Male and female C57BL/6 mice (7–10 weeks old) wereobtained from Harlan (France). The animals were housedfour to five per cage under standard laboratory condi-tions.
B16 melanoma cells
B16 melanoma cells, selected by IJ Fidler from a murinemelanoma, are syngeneic in C57BL/6 mice. Cells weregrown in cell culture Dulbecco’s modified Eagle’s medium(DMEM), supplemented with 10% heat-inactivated fetalbovine serum (FBS, Biomedia, France) and with penicillin(100U/ml) and streptomycin (100mg/ml) in a humidifiedincubator with 5% CO2 at 371C. This adherent cell linewas detached from the flasks with phosphate-bufferedsaline (PBS) and ethylenediaminetetraacetic acid (EDTA)2mM.Cells were washed with PBS and inoculated in DMEM
without antibiotics or FBS.
Cell transfection
The plasmid DNAs encoding for GM-CSF or IL-4 weretransfected to cells using the cationic lipid vector DOTAP(Boehringer Mannheim-Roche, Diagnostics Roche,
Spain). We employed a mass ratio of 4 : 1 (DOTAP:D-NA), which implies a molar ratio of 1.7 : 1 (N/P,N¼moles of nitrogen atoms in DOTAP and P¼molesof DNA phosphate groups). The DNA solution wasprepared in PBS, and the DOTAP was prepared inendotoxin-free ultrapure water (MilliQ). An equal volumeof each solution was mixed (500 mL final volume) andincubated 15 minutes before addition. In all cases, thelipoplex characterization was performed as previouslydescribed.29 B16 cells were detached and seeded (3 millioncells) in flasks (75 cm3). When 70% confluence wasreached (about 24 hours later), the cells were transfectedwith DOTAP/DNA complexes at 5 mg of DNA plasmidper ml of medium without serum. After 30 minutes ofincubation at 371C, 10mL of medium supplemented with15% FBS was added (10% final FBS in DMEM). Toselect clones expressing GM-CSF or IL-4, 48 hours aftertransfection, cells were cultured in the presence of G418(2mg/mL), and 3 weeks later the resistant cells to G418(Calbiochem, Spain) were harvested with PBS EDTA andseeded as 104 cells per Petri dish. Colonies were ring-cloned, picked up, expanded and tested for cytokineproduction by quantitative RT-PCR and flow cytometry.For fresh transfection vaccination (transient expression),cells were irradiated 24 hours after transfection and frozen(�801C). For in vivo use, cells were warmed and two timeswashed in DMEM and in PBS to eliminate the freezingmedium. Cells were counted and administered in DMEM/PBS. Other samples (nontransfected) were also treatedsimilarly. Aliquots were withdrawn to be analyzed byqRT-PCR and flow cytometry, employing GolgiStopsolution (PharMingen, BD Biociencias, Spain). Otheraliquots were used to evaluate the cell viability employingCalceine-AM (Live/Deads viability/cytotoxicity fromMolecular Probes, The Netherlands). Accordingly, cellswere seeded (7,000 cells/microwell) in wells (96 microwellsper plate) and cultured. At different times, the cellmedium was removed to add 50mL of a Calceine dilutionin PBS (0.5mL/mL PBS). Cells were incubated at 371C for40min, and the fluorescence (as a result of the cell esteraseactivity) was measured with a microplate cytofluorimeter(CytofluorTM 2350 Millipore, Ex¼ 485 nm, Em¼ 530 nm).
Quantitative RT-PCR (qRT-PCR)
Primers for the cytokines as well as for the housekeepinggene glyceraldehyde-3-phosphate dehydrogenase(GAPDH) were designed using the computer programPrimer Express (Applied Biosystems, Spain). The meltingtemperature (Tm) was close to 601C, and the ampliconlengths were under 100 bp; primers for murine GM-CSF,forward: 50-30 TGGGCATTGTGGTCTACAGC, re-verse: AAGGCCGGGTGACAGTGAT. For murine IL-4, forward: 50-30 ACAGGAGAAGGGACGCCAT andreverse: CTGTGGTGTTCTTCGTTGCTG.RNA was extracted from the cell samples employing
the RNeasy purification Kit (Qiagen, Izasa SA, Spain).Reverse transcription of RNA to generate cDNA wascarried out using the SYBR Green RT-PCR reagents kit(P/N 4310179, Applied Biosystems, Spain). The condi-
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
888
Cancer Gene Therapy

tions for each 30 mL of final volume were: 3 mL 10� roomtemperature buffer, 6.6 mL 25mM MgCl2, 6mL 2.5mMNTPs, 1.5 mL 50 mM Random Hexamers, 0.6 mL 20U/mLRNase inhibitor, 0.75 mL50U/mL MultiScribe reversetranscriptase and 300 ng of total RNA. Samples wereincubated at 251C for 10 minutes, at 481C for 30 minutes,and for 5 minutes at 951C to heat inactivate the reversetranscriptase.PCR was performed using the ABI PRISM 7700
Sequence Detection System (Applied Biosystems). Theassay conditions for quantitative PCR (final volume of50 mL) were: 2mL of primer forward (10 mM), 2mL ofprimer reverse (10mM), 5mL cDNA and 25mL of 2�SYBR Green PCR master mix. The thermal cyclingconditions included an initial step at 951C for 10 minutes,followed by 40 cycles at 951C for 15 seconds and 601C for1 min.30,31 The method used for obtaining quantitativedata of relative gene expression is the comparative Ctmethod (DDCt method) as described by the manufacturer.Ct is measured as the PCR cycle numbers at which thereporter fluorescent emission increases beyond a thresh-old level that is based on the background fluorescence ofthe system. The standard curves in qPCR were generatedby cytokine plasmid 10-fold dilution from 10�1 to10�6 ng. The Ct results were input in the standard curvewith the quantity log.An alternative approach for expressing qRT-PCR
results is the use of the formula 2�DDCt, where:DDCt¼ (CtGM-CSF or IL-4�CtGAPDH)samples�(CtGM-CSF or
IL-4�CtGAPDH)controls.
Flow cytometry
For the detection of GM-CSF in transfected cells, weemployed an immunostaining protocol. Briefly, cells weredetached from the flasks and cultured under a solution (inDMEM 10% FBS) of GolgiStop (PharMingen, BDBiociencias, Spain), adding 4 mL per 6� 106 cells (1mil-lion/mL) following the manufacturer’s instructions. Cellswere incubated at 371C for 4 hours under continuousagitation, followed by the IntraPrept Permeation Re-agent protocol (PharMingen) employing 0.5 million cells,and 5mL of GM-CSF or IL-4 antibodies coupled tofluorescein isothiocyanate (FITC) molecules (PharMin-gen). Cells were kept overnight in darkness untilmeasurement was carried out using an FACScan flowcytometer (Becton-Dickinson).
Vaccination protocol
B16 cells were irradiated (150Gy) before administrationin vivo to prevent tumor proliferation of the vaccines. Forcell clones vaccination, cells were immediately adminis-tered to mice after irradiation. For fresh transfection, cellswere irradiated 24 hours after transfection, and after-wards were frozen at �801C until use.Mice were vaccinated on days �21,�7 and +7, and the
tumorigenic dose of 100,000 wild-type B16 cells wasinjected on day 0 in the opposite leg. The doses of vaccineemployed in cell clones vaccination were 0.2 and 1 millioncells, via an intradermal injection in the leg. A similar
procedure was adopted (with 0.2 and 2 million cells) in thecase of fresh transfected cells. Tumor development in vivowas monitored visually and analyzed for volume with acaliper in two dimensions on different days. The volumewas calculated by the formula V¼ (A�B2)/2, whereV¼ volume (mm3), and A and B are the long and shortdiameters, respectively. Two blood extractions wereperformed in mice on days �1 and +15.
Radial immunodiffusion
For the measurement of IgM, IgG and IgG subtypelevels, pooled sera from animals of each experimentalgroup were used. Mice were bled by the tail vein intoheparinized capillaries on days �1 and +15, followed bycentrifugation at 160 g for 5 minutes and plasmacollection. The assay consisted of a commercial radialimmunodiffusion technique (the Binding Site, Birming-ham, UK) with agarose plates containing monospecificantisera to mouse immunoglobulin isotypes and IgGsubclasses. Serum samples were diluted according to theinstructions of the manufacturer (1/2 for IgM and IgG3;1/10 for IgG1, IgG2a and IgG2b), and an appropriatevolume was dispensed into the precut wells. Immunoglo-bulins were allowed to diffuse for 96 hours, andprecipitate ring diameters were measured by means ofan eyepiece. The concentration was established on thebasis of a correspondence table included in the kit.Controls were run in each assay to test for accuracy. In allinstances, immunoglobulin levels in control samples werewithin the admissible range as stated by the manufacturer.
ELISA
The GM-CSF in the serum was measured with a murineGM-CSF kit following the instructions of the manufac-turer (ParMingen, BD Biociencias, Spain). Serum-specificantibodies to tumor antigens were also measured byELISA. Briefly, flat-bottom 96-well plates (Maxisorp,NUNC, USA) were coated with tumor membrane protein(TMP) 100 mL per well, obtained from B16 tumor cells aspreviously described.32 The coating concentration of thisELISA antigen employed was 1 mg/mL in 50mM CO3Na/HCO3Na pH 9.6. The plates were sealed and incubatedovernight at 41C. On the following day, the plates werewashed with buffer (PBS containing 0.1% Tween 20), andincubated at room temperature (RT) for 1 hour withblocking buffer (PBS with 1% BSA). Mice sera werediluted (1 : 20) in PBS containing 1% BSA and 0.1%Tween 20, added to the plates and incubated at 371C for 2hours. To detect bound IgG, plates were incubated at RTfor 1 hour with the secondary antibody (goat-anti mouseIgG-peroxidase, Caltag, USA), diluted 1 : 3000 in thesame solution employed with mice sera. Finally, thesubstrate solution (30mg/mL O-phenylenediamine dihy-drochloride (OPD) with 0.03% H2O2 in citrate buffer)was added to each well. The reaction was stopped by theaddition of 1N HCl. The results are expressed as themean of the optical density (OD) at 492 nm.
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
889
Cancer Gene Therapy
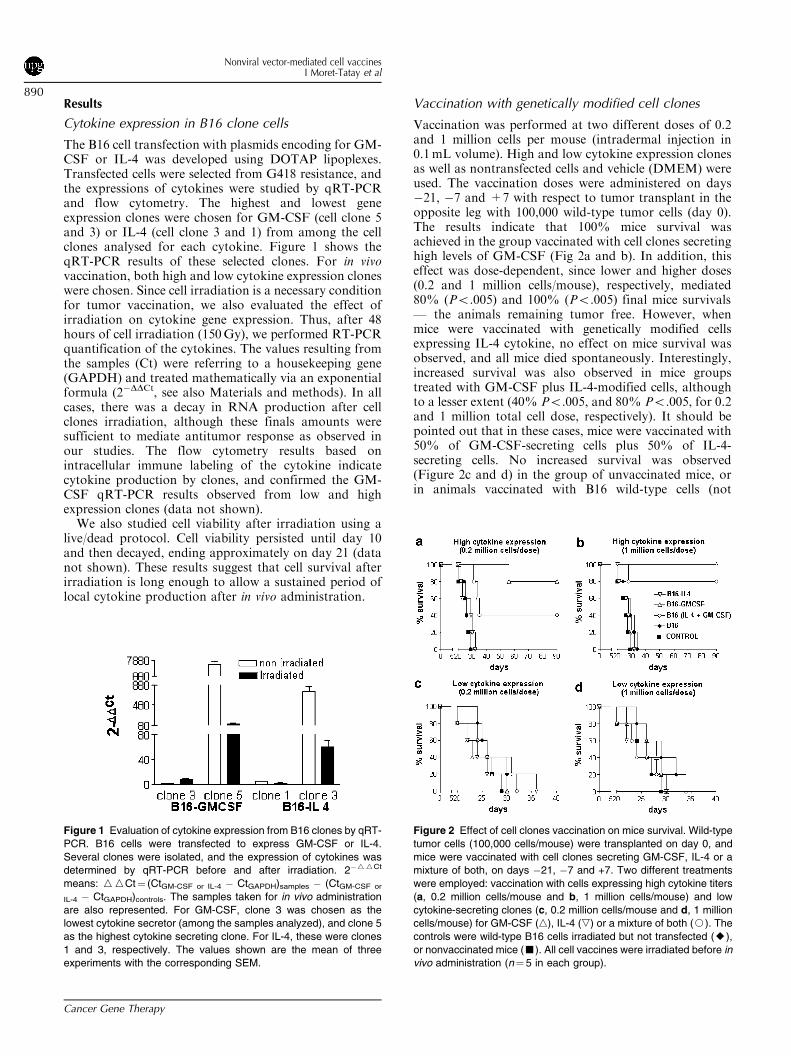
Results
Cytokine expression in B16 clone cells
The B16 cell transfection with plasmids encoding for GM-CSF or IL-4 was developed using DOTAP lipoplexes.Transfected cells were selected from G418 resistance, andthe expressions of cytokines were studied by qRT-PCRand flow cytometry. The highest and lowest geneexpression clones were chosen for GM-CSF (cell clone 5and 3) or IL-4 (cell clone 3 and 1) from among the cellclones analysed for each cytokine. Figure 1 shows theqRT-PCR results of these selected clones. For in vivovaccination, both high and low cytokine expression cloneswere chosen. Since cell irradiation is a necessary conditionfor tumor vaccination, we also evaluated the effect ofirradiation on cytokine gene expression. Thus, after 48hours of cell irradiation (150Gy), we performed RT-PCRquantification of the cytokines. The values resulting fromthe samples (Ct) were referring to a housekeeping gene(GAPDH) and treated mathematically via an exponentialformula (2�DDCt, see also Materials and methods). In allcases, there was a decay in RNA production after cellclones irradiation, although these finals amounts weresufficient to mediate antitumor response as observed inour studies. The flow cytometry results based onintracellular immune labeling of the cytokine indicatecytokine production by clones, and confirmed the GM-CSF qRT-PCR results observed from low and highexpression clones (data not shown).We also studied cell viability after irradiation using a
live/dead protocol. Cell viability persisted until day 10and then decayed, ending approximately on day 21 (datanot shown). These results suggest that cell survival afterirradiation is long enough to allow a sustained period oflocal cytokine production after in vivo administration.
Vaccination with genetically modified cell clones
Vaccination was performed at two different doses of 0.2and 1 million cells per mouse (intradermal injection in0.1mL volume). High and low cytokine expression clonesas well as nontransfected cells and vehicle (DMEM) wereused. The vaccination doses were administered on days�21, �7 and +7 with respect to tumor transplant in theopposite leg with 100,000 wild-type tumor cells (day 0).The results indicate that 100% mice survival wasachieved in the group vaccinated with cell clones secretinghigh levels of GM-CSF (Fig 2a and b). In addition, thiseffect was dose-dependent, since lower and higher doses(0.2 and 1 million cells/mouse), respectively, mediated80% (Po.005) and 100% (Po.005) final mice survivals— the animals remaining tumor free. However, whenmice were vaccinated with genetically modified cellsexpressing IL-4 cytokine, no effect on mice survival wasobserved, and all mice died spontaneously. Interestingly,increased survival was also observed in mice groupstreated with GM-CSF plus IL-4-modified cells, althoughto a lesser extent (40% Po.005, and 80% Po.005, for 0.2and 1 million total cell dose, respectively). It should bepointed out that in these cases, mice were vaccinated with50% of GM-CSF-secreting cells plus 50% of IL-4-secreting cells. No increased survival was observed(Figure 2c and d) in the group of unvaccinated mice, orin animals vaccinated with B16 wild-type cells (not
Figure 1 Evaluation of cytokine expression from B16 clones by qRT-
PCR. B16 cells were transfected to express GM-CSF or IL-4.
Several clones were isolated, and the expression of cytokines was
determined by qRT-PCR before and after irradiation. 2�WWCt
means: WWCt¼ (CtGM-CSF or IL-4 � CtGAPDH)samples � (CtGM-CSF or
IL-4 � CtGAPDH)controls. The samples taken for in vivo administration
are also represented. For GM-CSF, clone 3 was chosen as the
lowest cytokine secretor (among the samples analyzed), and clone 5
as the highest cytokine secreting clone. For IL-4, these were clones
1 and 3, respectively. The values shown are the mean of three
experiments with the corresponding SEM.
Figure 2 Effect of cell clones vaccination on mice survival. Wild-type
tumor cells (100,000 cells/mouse) were transplanted on day 0, and
mice were vaccinated with cell clones secreting GM-CSF, IL-4 or a
mixture of both, on days �21, �7 and +7. Two different treatments
were employed: vaccination with cells expressing high cytokine titers
(a, 0.2 million cells/mouse and b, 1 million cells/mouse) and low
cytokine-secreting clones (c, 0.2 million cells/mouse and d, 1 million
cells/mouse) for GM-CSF (n), IL-4 (,) or a mixture of both (J). The
controls were wild-type B16 cells irradiated but not transfected (E),
or nonvaccinated mice (’). All cell vaccines were irradiated before in
vivo administration (n¼ 5 in each group).
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
890
Cancer Gene Therapy
genetically modified) or with cells exhibiting low GM-CSF or IL-4 gene expression.Regardless of the cell vaccine used, many vaccinated
animals developed a vitiligo reaction, although not all ofthem remained tumor free until the end of the observationperiod. In most cases, vitiligo was located on the snout,although some animals exhibited extended reactions overthe body.
Second exposure to tumor challenge in mice vaccinatedwith cell clones
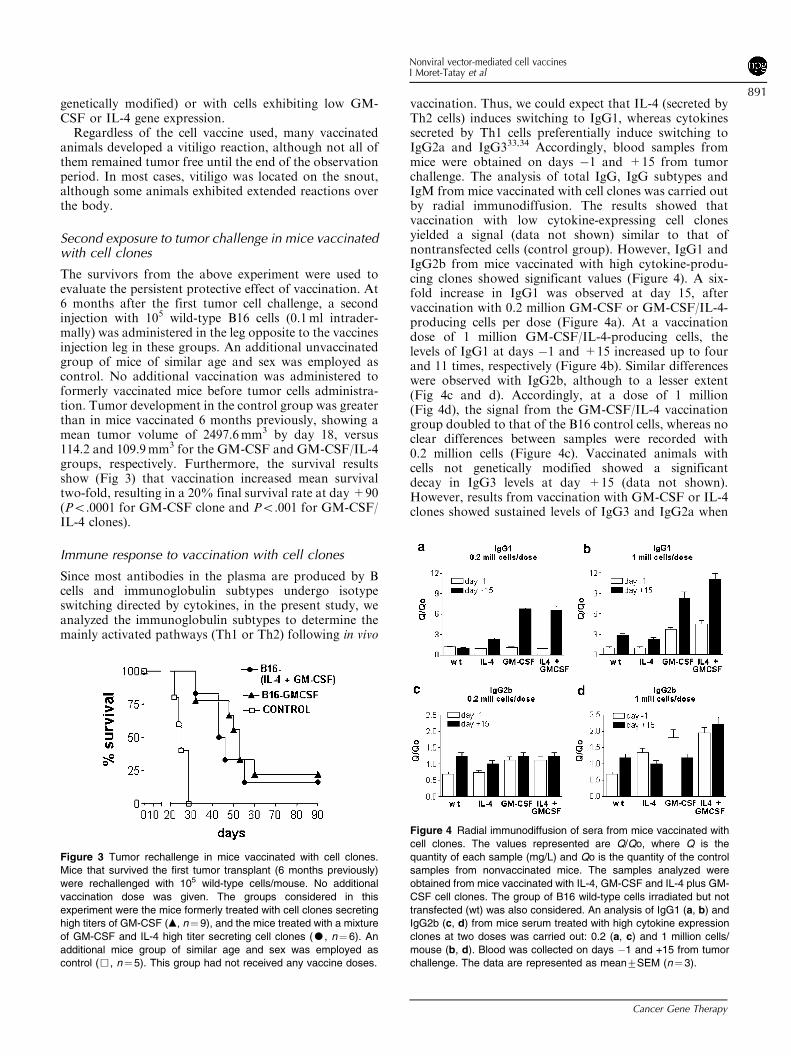
The survivors from the above experiment were used toevaluate the persistent protective effect of vaccination. At6 months after the first tumor cell challenge, a secondinjection with 105 wild-type B16 cells (0.1ml intrader-mally) was administered in the leg opposite to the vaccinesinjection leg in these groups. An additional unvaccinatedgroup of mice of similar age and sex was employed ascontrol. No additional vaccination was administered toformerly vaccinated mice before tumor cells administra-tion. Tumor development in the control group was greaterthan in mice vaccinated 6 months previously, showing amean tumor volume of 2497.6mm3 by day 18, versus114.2 and 109.9mm3 for the GM-CSF and GM-CSF/IL-4groups, respectively. Furthermore, the survival resultsshow (Fig 3) that vaccination increased mean survivaltwo-fold, resulting in a 20% final survival rate at day +90(Po.0001 for GM-CSF clone and Po.001 for GM-CSF/IL-4 clones).
Immune response to vaccination with cell clones
Since most antibodies in the plasma are produced by Bcells and immunoglobulin subtypes undergo isotypeswitching directed by cytokines, in the present study, weanalyzed the immunoglobulin subtypes to determine themainly activated pathways (Th1 or Th2) following in vivo
vaccination. Thus, we could expect that IL-4 (secreted byTh2 cells) induces switching to IgG1, whereas cytokinessecreted by Th1 cells preferentially induce switching toIgG2a and IgG333,34 Accordingly, blood samples frommice were obtained on days �1 and +15 from tumorchallenge. The analysis of total IgG, IgG subtypes andIgM from mice vaccinated with cell clones was carried outby radial immunodiffusion. The results showed thatvaccination with low cytokine-expressing cell clonesyielded a signal (data not shown) similar to that ofnontransfected cells (control group). However, IgG1 andIgG2b from mice vaccinated with high cytokine-produ-cing clones showed significant values (Figure 4). A six-fold increase in IgG1 was observed at day 15, aftervaccination with 0.2 million GM-CSF or GM-CSF/IL-4-producing cells per dose (Figure 4a). At a vaccinationdose of 1 million GM-CSF/IL-4-producing cells, thelevels of IgG1 at days �1 and +15 increased up to fourand 11 times, respectively (Figure 4b). Similar differenceswere observed with IgG2b, although to a lesser extent(Fig 4c and d). Accordingly, at a dose of 1 million(Fig 4d), the signal from the GM-CSF/IL-4 vaccinationgroup doubled to that of the B16 control cells, whereas noclear differences between samples were recorded with0.2 million cells (Figure 4c). Vaccinated animals withcells not genetically modified showed a significantdecay in IgG3 levels at day +15 (data not shown).However, results from vaccination with GM-CSF or IL-4clones showed sustained levels of IgG3 and IgG2a when
Figure 3 Tumor rechallenge in mice vaccinated with cell clones.
Mice that survived the first tumor transplant (6 months previously)
were rechallenged with 105 wild-type cells/mouse. No additional
vaccination dose was given. The groups considered in this
experiment were the mice formerly treated with cell clones secreting
high titers of GM-CSF (m, n¼ 9), and the mice treated with a mixture
of GM-CSF and IL-4 high titer secreting cell clones (K, n¼ 6). An
additional mice group of similar age and sex was employed as
control (&, n¼5). This group had not received any vaccine doses.
Figure 4 Radial immunodiffusion of sera from mice vaccinated with
cell clones. The values represented are Q/Qo, where Q is the
quantity of each sample (mg/L) and Qo is the quantity of the control
samples from nonvaccinated mice. The samples analyzed were
obtained from mice vaccinated with IL-4, GM-CSF and IL-4 plus GM-
CSF cell clones. The group of B16 wild-type cells irradiated but not
transfected (wt) was also considered. An analysis of IgG1 (a, b) and
IgG2b (c, d) from mice serum treated with high cytokine expression
clones at two doses was carried out: 0.2 (a, c) and 1 million cells/
mouse (b, d). Blood was collected on days �1 and +15 from tumor
challenge. The data are represented as mean7SEM (n¼3).
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
891
Cancer Gene Therapy
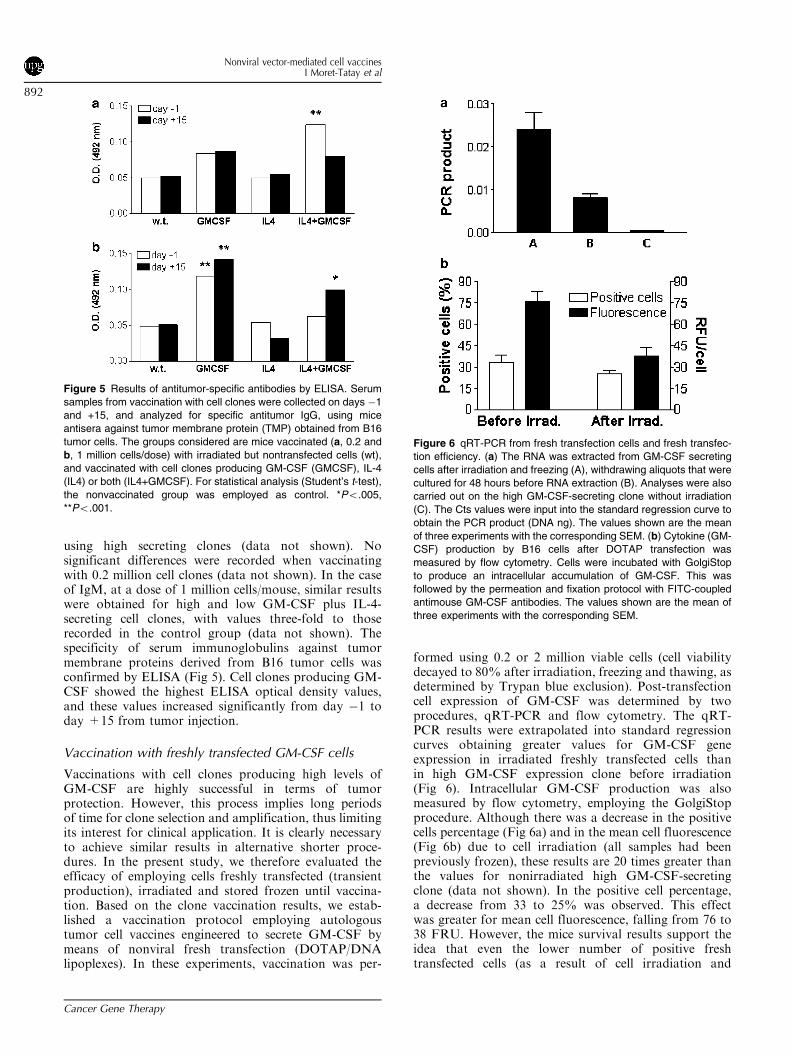
using high secreting clones (data not shown). Nosignificant differences were recorded when vaccinatingwith 0.2 million cell clones (data not shown). In the caseof IgM, at a dose of 1 million cells/mouse, similar resultswere obtained for high and low GM-CSF plus IL-4-secreting cell clones, with values three-fold to thoserecorded in the control group (data not shown). Thespecificity of serum immunoglobulins against tumormembrane proteins derived from B16 tumor cells wasconfirmed by ELISA (Fig 5). Cell clones producing GM-CSF showed the highest ELISA optical density values,and these values increased significantly from day �1 today +15 from tumor injection.
Vaccination with freshly transfected GM-CSF cells
Vaccinations with cell clones producing high levels ofGM-CSF are highly successful in terms of tumorprotection. However, this process implies long periodsof time for clone selection and amplification, thus limitingits interest for clinical application. It is clearly necessaryto achieve similar results in alternative shorter proce-dures. In the present study, we therefore evaluated theefficacy of employing cells freshly transfected (transientproduction), irradiated and stored frozen until vaccina-tion. Based on the clone vaccination results, we estab-lished a vaccination protocol employing autologoustumor cell vaccines engineered to secrete GM-CSF bymeans of nonviral fresh transfection (DOTAP/DNAlipoplexes). In these experiments, vaccination was per-
formed using 0.2 or 2 million viable cells (cell viabilitydecayed to 80% after irradiation, freezing and thawing, asdetermined by Trypan blue exclusion). Post-transfectioncell expression of GM-CSF was determined by twoprocedures, qRT-PCR and flow cytometry. The qRT-PCR results were extrapolated into standard regressioncurves obtaining greater values for GM-CSF geneexpression in irradiated freshly transfected cells thanin high GM-CSF expression clone before irradiation(Fig 6). Intracellular GM-CSF production was alsomeasured by flow cytometry, employing the GolgiStopprocedure. Although there was a decrease in the positivecells percentage (Fig 6a) and in the mean cell fluorescence(Fig 6b) due to cell irradiation (all samples had beenpreviously frozen), these results are 20 times greater thanthe values for nonirradiated high GM-CSF-secretingclone (data not shown). In the positive cell percentage,a decrease from 33 to 25% was observed. This effectwas greater for mean cell fluorescence, falling from 76 to38 FRU. However, the mice survival results support theidea that even the lower number of positive freshtransfected cells (as a result of cell irradiation and
Figure 5 Results of antitumor-specific antibodies by ELISA. Serum
samples from vaccination with cell clones were collected on days �1
and +15, and analyzed for specific antitumor IgG, using mice
antisera against tumor membrane protein (TMP) obtained from B16
tumor cells. The groups considered are mice vaccinated (a, 0.2 and
b, 1 million cells/dose) with irradiated but nontransfected cells (wt),
and vaccinated with cell clones producing GM-CSF (GMCSF), IL-4
(IL4) or both (IL4+GMCSF). For statistical analysis (Student’s t-test),
the nonvaccinated group was employed as control. *Po.005,
**Po.001.
Figure 6 qRT-PCR from fresh transfection cells and fresh transfec-
tion efficiency. (a) The RNA was extracted from GM-CSF secreting
cells after irradiation and freezing (A), withdrawing aliquots that were
cultured for 48 hours before RNA extraction (B). Analyses were also
carried out on the high GM-CSF-secreting clone without irradiation
(C). The Cts values were input into the standard regression curve to
obtain the PCR product (DNA ng). The values shown are the mean
of three experiments with the corresponding SEM. (b) Cytokine (GM-
CSF) production by B16 cells after DOTAP transfection was
measured by flow cytometry. Cells were incubated with GolgiStop
to produce an intracellular accumulation of GM-CSF. This was
followed by the permeation and fixation protocol with FITC-coupled
antimouse GM-CSF antibodies. The values shown are the mean of
three experiments with the corresponding SEM.
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
892
Cancer Gene Therapy
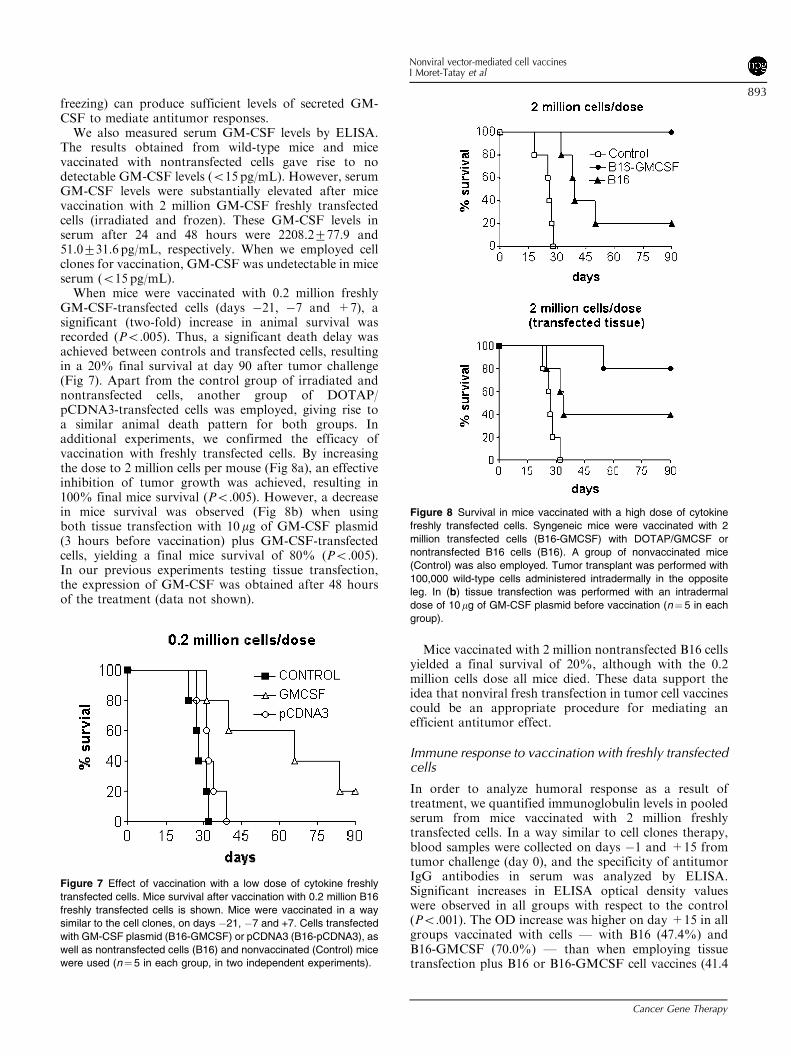
freezing) can produce sufficient levels of secreted GM-CSF to mediate antitumor responses.We also measured serum GM-CSF levels by ELISA.
The results obtained from wild-type mice and micevaccinated with nontransfected cells gave rise to nodetectable GM-CSF levels (o15 pg/mL). However, serumGM-CSF levels were substantially elevated after micevaccination with 2 million GM-CSF freshly transfectedcells (irradiated and frozen). These GM-CSF levels inserum after 24 and 48 hours were 2208.2777.9 and51.0731.6 pg/mL, respectively. When we employed cellclones for vaccination, GM-CSF was undetectable in miceserum (o15 pg/mL).When mice were vaccinated with 0.2 million freshly
GM-CSF-transfected cells (days �21, �7 and +7), asignificant (two-fold) increase in animal survival wasrecorded (Po.005). Thus, a significant death delay wasachieved between controls and transfected cells, resultingin a 20% final survival at day 90 after tumor challenge(Fig 7). Apart from the control group of irradiated andnontransfected cells, another group of DOTAP/pCDNA3-transfected cells was employed, giving rise toa similar animal death pattern for both groups. Inadditional experiments, we confirmed the efficacy ofvaccination with freshly transfected cells. By increasingthe dose to 2 million cells per mouse (Fig 8a), an effectiveinhibition of tumor growth was achieved, resulting in100% final mice survival (Po.005). However, a decreasein mice survival was observed (Fig 8b) when usingboth tissue transfection with 10mg of GM-CSF plasmid(3 hours before vaccination) plus GM-CSF-transfectedcells, yielding a final mice survival of 80% (Po.005).In our previous experiments testing tissue transfection,the expression of GM-CSF was obtained after 48 hoursof the treatment (data not shown).
Mice vaccinated with 2 million nontransfected B16 cellsyielded a final survival of 20%, although with the 0.2million cells dose all mice died. These data support theidea that nonviral fresh transfection in tumor cell vaccinescould be an appropriate procedure for mediating anefficient antitumor effect.
Immune response to vaccination with freshly transfectedcells
In order to analyze humoral response as a result oftreatment, we quantified immunoglobulin levels in pooledserum from mice vaccinated with 2 million freshlytransfected cells. In a way similar to cell clones therapy,blood samples were collected on days �1 and +15 fromtumor challenge (day 0), and the specificity of antitumorIgG antibodies in serum was analyzed by ELISA.Significant increases in ELISA optical density valueswere observed in all groups with respect to the control(Po.001). The OD increase was higher on day +15 in allgroups vaccinated with cells — with B16 (47.4%) andB16-GMCSF (70.0%) — than when employing tissuetransfection plus B16 or B16-GMCSF cell vaccines (41.4
Figure 7 Effect of vaccination with a low dose of cytokine freshly
transfected cells. Mice survival after vaccination with 0.2 million B16
freshly transfected cells is shown. Mice were vaccinated in a way
similar to the cell clones, on days �21, �7 and +7. Cells transfected
with GM-CSF plasmid (B16-GMCSF) or pCDNA3 (B16-pCDNA3), as
well as nontransfected cells (B16) and nonvaccinated (Control) mice
were used (n¼5 in each group, in two independent experiments).
Figure 8 Survival in mice vaccinated with a high dose of cytokine
freshly transfected cells. Syngeneic mice were vaccinated with 2
million transfected cells (B16-GMCSF) with DOTAP/GMCSF or
nontransfected B16 cells (B16). A group of nonvaccinated mice
(Control) was also employed. Tumor transplant was performed with
100,000 wild-type cells administered intradermally in the opposite
leg. In (b) tissue transfection was performed with an intradermal
dose of 10mg of GM-CSF plasmid before vaccination (n¼ 5 in each
group).
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
893
Cancer Gene Therapy

and 56.9%, respectively). All these values were greaterthan those yielded by cell clones (Fig 5). In the cases oftotal IgG, IgG1 and IgG3 subtypes (Fig 9) analyzed byradial immunodiffusion, increased levels were seen in themice group vaccinated with GM-CSF freshly transfectedcells, showing a six-fold increase on day +15 for the IgG3subtype compared with the controls (Fig 9). The greatestdifferences between groups were recorded for IgG2a(Table 1), the highest levels being obtained in the micegroup vaccinated with GM-CSF-transfected cells. How-ever, the immunoglobulin levels decreased when tissuewas transfected before GM-CSF cell vaccine injection.These results agree with the decrease in mice survivalobserved when we employed both tissue pretreatment andGM-CSF cell vaccines (Fig 8).
Discussion
In the present study, we report that vaccination withnonviral vector-transfected cells can afford up to 100%survival in mice after tumor challenge. The tumor vaccinetransfected with DOTAP lipoplexes to express GM-CSFexerted a potent antitumor effect, preventing tumorgrowth and greatly increasing mice survival (80–100%).This effect was observed employing both stable geneti-cally modified cells (clones) or recently transfected cells(fresh transfection), with a transfection efficiency ofX25% using DOTAP/DNA lipoplexes. The antitumorsuccess of these vaccines was similar in all cases involvingthe use of tumor cells genetically modified to produceGM-CSF. In contrast, similar vaccination strategies withIL-4 genetically modified cells failed to mediate tumorprotection. Finally, no synergy was observed when bothIL-4- and GM-CSF-modified cells were employed forvaccination.Referring to the updated results in laboratory studies,
the technique of inducing the expression of immunestimulatory genes by tumor cells is being widely appliedusing viral vectors.1,15,35–37 In contrast, little work hasbeen reported thus far in the case of cell vaccinesengineered by nonviral vectors to secrete cytokines.38
Our results of vaccination with cells producing GM-CSFoffer expectations regarding the promising results ob-tained by applying nonviral gene therapy, which aims toafford safer therapy despite comparatively lower efficacy.The present study was designed to establish the antitumorefficacy after vaccination of two different cytokines, GM-CSF and IL-4, in genetically modified tumor cells, as cellclones or freshly transfected cells. We chose thesecytokines because of their different roles in immuneresponse. GM-CSF seems to afford protective immunity(T-cell memory) and an intense infiltration of immaturemononuclear cells.3,39 IL-4 is a cytokine that stimulatesthe proliferation of T cells, B cells and mast cells.40 It alsocooperates with GM-CSF in activating/differentiatingdendritic cells.7 In our experiments, the results show thatvaccination with cell clones producing IL-4 did notprotect mice from tumor growth, affording tumorprotection similar to that obtained with nontransfectedB16 wild-type cells. Nevertheless, IL-4 is a cytokine thathas been employed in studies with genetically modifiedcells as vaccines, with disparate results. In this sense, ourfindings agree with those observed by other authorsemploying retrovirus-transduced cells, who found a B16-producing GM-CSF-mediated antitumor effect in a B16brain tumor model, although this effect was not observedwhen mice were vaccinated with B16 cells secreting IL-4or with nontransfected B16 cells.15 However, in othertumors it has been described that vaccination with IL-4-producing cells (viral transduction) could activate tumor-killing mechanisms.19,41,42
The use of cell clones secreting IL-4 did not protectmice from tumor challenge, but cell clones secreting GM-CSF mediated a dose-dependent antitumor response,since 0.2 and 1 million cells yielded a final survival of80 and 100%, respectively. Cross-priming through the
Figure 9 Radial immunodiffusion of serum from mice vaccinated
with freshly transfected cells. Two blood extractions were carried out
on days �1 and +15 in mice vaccinated with 2 million cells (B16 wild
type, or B16 transfected to express GM-CSF). Determinations of IgG
and IgG1 and IgG3 subtypes were performed by radial immunodiffu-
sion. The values are represented as Q/Qo, where Q is the quantity
(mg/L) of each sample (B16 wild-type and B16-secreting GM-CSF)
and Qo represents the values of the control group (nonvaccinated
mice). The data are represented as the mean7SEM (n¼ 3).
Table 1 Results of radial immunodiffusion from vaccination with 2million freshly transfected cells.
Groups IgG2a (mg/L) IgG2b (mg/L)
Day �1
Control 1090 537
B16 12,960 730
B16+T.T. 9470 730
B16-GMCSF 14,360 705
B16-GMCSF+T.T. 10,340 705
Day+15
Control 11,970 405
B16 13,300 655
B16+T.T. 10,960 537
B16-GMCSF 14,716 943
B16-GMCSF+T.T. 10,674 756
Analyses of IgG2a and IgG2b from mice vaccinated with B16 wildtype (nontransfected) or B16 freshly transfected with DOTAP/GM-CSF were carried out. Also represented are the results obtained froman additional experiment of transfected tissue (T.T.) in mouse legwith 10 mg GM-CSF plasmid before vaccination. The control groupcomprises the nonvaccinated mice. In all cases, the relative SD didnot exceed 10%.
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
894
Cancer Gene Therapy

host APCs or direct antigen presentation by the tumorcells is postulated to play an important role in vaccinationwith GM-CSF, since this cytokine exerts an effect on thematuration of APCs.6 Since B16 cells do not express H-2molecules, direct antigen presentation by B16 cells mustbe a limit mechanism; we therefore speculate that peptidesreleased from tumor cells, employed for vaccination,could prime antitumor response by a cross-primingmechanism.Since the GM-CSF and IL-4 cytokines have been
previously described to play an antineoplastic role,1,24 wecould hope to increase tumor cell rejection by thecombination of genetically modified cells producing IL-4and GM-CSF. However, we found the greatest antitumoreffect when using only GM-CSF-secreting cells, and lowerefficacy was observed when both GM-CSF- and IL-4-producing cells were combined for vaccination. Thisadverse effect could be produced not only by the reducednumber of cells employed, but also by a dysregulatoryeffect of IL-4 on Th1/Th2-activated balance mediated byGM-CSF, resulting in an inhibitory effect on Th1 andreduced CTL response. On evaluating the possibleadvantages of employing both cytokines, it has beenobserved that in an intracerebral tumor model, 80% ofmice immunized with GM-CSF plus IL-4 transduced B16cells (viral vectors) rejected an intracerebral tumor cellchallenge.24 These results indicate that when employing adifferent vaccination model, IL-4 is able to produce atherapeutic effect against tumor challenge (5,000 wild-type cells/mouse).To confirm that GM-CSF vaccines could produce potent
long-lasting antitumor immunity,1,43 even through nonviralgene therapy, the cell clone-vaccinated mice that survivedwild-type tumor transplantation (6 months previously)were challenged by a second tumor transplant. The latterresulted in a significant reduction in tumor progression anda final survival of 20%. These results can be compared withthose of other authors who have obtained similar patternsof response employing viral vectors to transduce cells, andcan be interpreted as the establishment of an immunologicmemory affording long-term protection against tumorrecurrence or metastasis.1,4,17
GM-CSF has been found to be helpful in not onlygenetically modified tumor cells (clones) but also in freshcytokine-secreting cells. It is important to point out that itis difficult to keep patient cells in vitro for long periods oftime. Thus, evaluation of the vaccination effect of recentlytransfected cells that are irradiated and kept frozen untilvaccination constitutes an important step towards clinicaltrial application of nonviral gene therapy. With avaccination dose of 0.2 million cells per mouse, survivalin mice was 20%, and increased to 100% with 2 millioncells. The modification of the vaccination protocol,including pretreatment of the tissue with 10 mg of GM-CSF plasmid, did not offer advantages — yielding a lesserfinal survival (80%) in mice when employing 2 millioncells. This plasmid tissue pretreatment mediates GM-CSFproduction by local cells, as determined in previousexperiments by qRT-PCR of the tissue (data not shown),but our results indicate that GM-CSF tissue transfection
does not increase mice survival. This might reflect thatimmune stimulation was overexpressed when using GM-CSF-secreting cells plus GM-CSF tissue transfection,resulting in a decrease in tumor rejection. Indeed, it hasbeen demonstrated that the amount of GM-CSF secretedby cell vaccines is critical to the magnitude of the immuneresponse generated.17,18 Thus, it might be inferred that iflower levels fail to induce strong antitumor immunity,excessively high levels might produce adverse effects.24,44
In addition, we should also consider that the transfectedcell types after tissue treatment as well as the time courseof cytokine production with respect to antigen presenta-tion could play a role in mediating tolerance rather thanimmune response stimulation.45
Different attempts to immunize with irradiated butnontransfected tumor cells have had little success.1,46 Inour study, the protection of vaccination with wild-typecells had a slight antitumor effect, with a survival ofnearly 20% when employing 2 million cells per mouse.This observation agrees with the idea of B16 melanoma asan immunogenic tumor cell line47 that can producevitiligo reactions in immunized animals.48 Almost all theanimals in our experiment had vitiligo on the snout, butsome also developed a high vitiligo reaction over the restof the body. With respect to the humoral response, it wasshown that GM-CSF elicits an increase in IgG productionwith the ability to recognize membrane proteins derivedfrom B16 tumor cells. The more significant increases wereobserved in the cases of IgG3 and IgG2a in freshlytransfected cells versus B16 clones, which could explainthe success in mice survival of cells secreting GM-CSF.Both immunoglobulin subtypes result from IgG switchinginduced by IFN-g, mainly produced by Th1-activatedcells. In turn, the Th1response is mainly involved in CTLantitumor activity, resulting in increased animal survival.Some authors have observed that the production ofIgG2a is associated with a therapeutic response againstexperimental metastases, although not so in the case ofIgG1 induced by IL-4.14 Cells genetically modified toproduce IL-4 had a very limited effect on immunoglobulinproduction, including the IgG1 subtype. Similar resultsare reported by other authors, as no differences have beenfound in terms of an increase in each IgG in serum aftertreatment with cells producing IL-4.7 Vaccination withIL-4 was not effective in suppressing tumors, although IL-4 should be necessary to show therapeutic antitumorresponses, since on avoiding IL-4 production the numberof tumor metastases seem to increase.49
In conclusion, to our knowledge, this is the first reportshowing that cell vaccines prepared by nonviral genetherapy procedures can mediate 100% tumor growthprevention. The antitumor efficacy has been observedemploying either genetically modified cell clones or freshtransfected cells. Since fresh transfection is a rapidprocedure for obtaining genetically modified cells forvaccination, it offers an easy and versatile opportunity tostudy the antitumor efficacy of different cytokines andtheir possible synergies. Thus, vaccines based on nonviralgene therapy could be an interesting procedure for clinicalapplications in human cancer immunotherapy.
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
895
Cancer Gene Therapy

Acknowledgments
We thank all those who have collaborated in the presentstudy, particularly Dr Rosa Algas and Dr Diego Dualdefrom the Service of Radiotherapy (University ClinicHospital, Valencia, Spain) for their help and kindness,and our colleagues Vicent Guillem and Isaias Sanmartınfor their collaboration in preparing the vaccines and inthe in vivo administration. The plasmids employed werekindly donated by Dr Andres Koenig (MOLOGEN,Germany). The present work has been supported by theGeneralitat Valenciana, IMPIVA.
References
1. Dranoff G, Jaffee E, Lazenby A, et al. Vaccination withirradiated tumor cells engineered to secrete murine granu-locyte–macrophage colony-stimulating factor stimulatespotent, specific, and long-lasting anti-tumor immunity. ProcNatl Acad Sci USA. 1993;90:3539–3543.
2. Hogge GS, Burkholder JK, Culp J, et al. Development ofhuman granulocyte–macrophage colony-stimulating factor-transfected tumor cells vaccines for the treatment ofspontaneous canine cancer. Hum Gen Ther. 1998;9:1851–1861.
3. Simons JW, Bahar M. Ex vivo gene therapy using cytokine-transduced tumor vaccines: molecular and clinical pharma-cology. Semin Oncol. 1998;25:661–676.
4. Schadendorf D, Paschen A, Sun Y. Autologous allogeneictumor cells or genetically engineered cells as cancer vaccineagainst melanoma. Immunol Lett. 2000;74:67–74.
5. Todryk SM, Birchall LJ, Erlich R, et al. Efficacy of cytokinegene transfection may differ for autologous and allogeneictumour cell vaccines. Immunology. 2001;102:190–198.
6. Thomas MC, Greten TF, Pardoll DM, et al. Enhancedtumor protection by granulocyte–macrophage colony-sti-mulating factor expression at the site of an allogeneicvaccine. Hum Gene Ther. 1998;9:835–843.
7. Arienti F, Belli F, Napolitano F, et al. Vaccination ofmelanoma patients with interleukin 4 gene-transducedallogeneic melanoma cells. Hum Gene Ther. 1998;9:835–843.
8. Pan CH, Chen HW, Tao MH. Modulation of immuneresponses to DNA vaccines by codelivery of cytokines genes.J Formos Med Assoc. 1999;98:722–729.
9. Bevan MJ. Cross-priming for a secondary cytotoxicresponse to minor H antigens with H-2 congenic cells whichdo not cross-react in the cytotoxic assay. J Exp Med.1976;143:1283–1288.
10. Bennett SR, Carbone FR, Karamalis F, et al. Induction of aCD8+ cytotoxic T lymphocyte response by cross-primingrequires cognate CD4+ T cell help. J Exp Med.1997;186:65–70.
11. Huang AY, Bruce AT, Pardoll DM, et al. In vivo cross-priming of MHC class I-restricted antigens requires the TAPtransporter. Immunity. 1996;4:349–355.
12. den Haan JM, Lehar SM, Bevan MJ. CD8(+) but notCD8(�) dendritic cells cross-prime cytotoxic T cells in vivo.J Exp Med. 2000;192:1685–1696.
13. Pooley JL, Heath WR, Shortman K. Cutting edge:intravenous soluble antigen is presented to CD4 T cells byCD8-dendritic cells, but cross-presented to CD8 T cells byCD8+ dendritic cells. J Immunol. 2001;166:5327–5330.
14. Rodolfo M, Melani C, Zilocchi C, et al. IgG2a induced byinterleukin (IL) 12-producing tumor cell vaccines but notIgG1 induced by IL-4 vaccine is associated with theeradication of experimental metastases. Cancer Res.1998;58:5812–5817.
15. Sampson JH, Archer GE, Ashley DM, et al. Subcutaneousvaccination with irradiated, cytokine-producing tumorcells stimulates CD8+ cell-mediated immunity againsttumors located in the ‘‘immunologically privileged’’ centralnervous system. Proc Natl Acad Sci USA. 1996;93:10399–10404.
16. Hensley C, Spitzler S, McAlpine BE, et al. In vivo humanmelanoma cytokine production: inverse correlation of GM-CSF production with tumor depth Exp Dermatol.1998;7:335–341.
17. Kayaga J, Souberbielle BE, Sheikh N, et al. Anti-tumouractivity against B16-F10 melanoma with a GM-CSFsecreting allogeneic tumour cell vaccine. Gene Therapy1999;6:1475–1481.
18. Jaffee EM, Abrams R, Cameron J, et al. A phase I clinicaltrial of lethally irradiated allogeneic pancreatic tumor cellstransfected with the GM-CSF gene for the treatment ofpancreatic adenocarcinoma. Hum Gene Ther. 1998;9:1951–1971.
19. Rodolfo M, Zilocchi C, Accornero P, et al. IL-4-transducedtumor cell vaccine induces immunoregulatory type 2 CD8 Tlymphocytes that cure lung metastases upon adoptivetransfer. J Immunol. 1999;163:1923–1928.
20. Nakazaki Y, Tani K, Lin ZT, et al. Vaccine effect ofgranulocyte–macrophage colony-stimulating factor orCD80 gene-transduced murine hematopoietic tumor cellsand their cooperative enhancement of antitumor immunity.Gene Therapy 1998;5:1355–1362.
21. Teshima T, Mach N, Hill GR, et al. Tumor cell vaccineelicits potent antitumor immunity after allogeneic T-cell-depleted bone marrow transplantation. Cancer Res.2001;61:162–171.
22. Cao X, Chen G, He L, et al. Involvement of MHC class Imolecule and ICAM-1 in the enhancement of adhesion andcytotoxic susceptibility to immune effector cells of tumorcells transfected with the interleukin (IL)-2, IL-4 or IL-6gene. J Cancer Res Clin Oncol. 1997;123:602–608.
23. Song K, Chang Y, Prud’homme GJ. Regulation of T-helper-1 versus T-helper-2 activity and enhancement of tumorimmunity by combined DNA-based vaccination andnonviral cytokine gene transfer. Gene Therapy 2000;7:481–492.
24. Wakimoto H, Abe J, Tsunoda R, et al. Intensifiedantitumor immunity by a cancer vaccine that producesgranulocyte–macrophage colony-stimulating factor plusinterleukin 4. Cancer Res. 1996;56:1828–1833.
25. Parmiani G, Rodolfo M, Melani C. Immunological genetherapy with ex vivo gene-modified tumor cells: a critiqueand a reappraisal. Hum Gene Ther. 2000;11:1269–1275.
26. Dong Z, Yoneda J, Kumar R, et al. Angiostatin-mediatedsuppression of cancer metastases by primary neoplasmsengineered to produce granulocyte/macrophage colony-stimulating factor. J Exp Med. 1998;188:755–763.
27. Boyer MW, Waller EK, Bray RA, et al. Cytokineupregulation of the antigen presenting function of acutemyeloid leukemia cells. Leukemia. 2000;14:412–418.
28. Botella-Estrada R, Malet G, Revert F, et al. Antitumoreffect of B16 melanoma cells genetically modified with theangiogenesis inhibitor rnasin. Cancer Gene Ther.2001;8:278–284.
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
896
Cancer Gene Therapy

29. Moret I, Peris JE, Guillem V, et al. Stability of PEI-DNAand DOTAP-DNA complexes: effect of alkaline pH,heparin and serum. J Control Release. 2001;76:169–181.
30. Dasi F, Benet M, Crespo J, et al. Asialofetuin liposome-mediated human alpha1-antitrypsin gene transfer in vivoresults in stationary long-term gene expression. J Mol Med.2001;79:205–212.
31. Overbergh L, Valckx D, Waer M, Mathieu C. Quantifica-tion of murine cytokine mRNAs using real time quantitativereverse transcriptase PCR. Cytokine. 1999;11:305–312.
32. Alino SF, Lejarreta M, Alfaro J, et al. Antimetastatic effectof immunization with liposome-encapsulated tumor cell-membrane proteins obtained from experimental tumors.Immunopharmacol Immunotoxicol. 1995;17:419–436
33. Lorenz M, Jung S, Radbruch A. Switch transcripts inimmunoglobulin class switching. Science. 1995;267:1825–1828.
34. Stavnezer J. Immunoglobulin class switching. Curr OpinImmunol. 1996;8:199–205.
35. Simons JW, Mikhak B, Chang JF, et al. Induction ofimmunity to prostate cancer antigens: results of a clinicaltrial of vaccination with irradiated autologous prostatetumor cells engineered to secrete granulocyte–macrophagecolony-stimulating factor using ex vivo gene transfer. CancerRes. 1999;59:5160–5168.
36. Wittig B, Marten A, Dorbic T, et al. Therapeutic vaccina-tion against metastatic carcinoma by expression-modulatedand immunomodified autologous tumor cells: a first clinicalphase I/II trial. Hum Gene Ther. 2001;12:267–278.
37. Soiffer R, Lynch T, Mihm M, et al. Vaccination withirradiated autologous melanoma cells engineered to secretehuman granulocyte–macrophage colony-stimulating factorgenerates potent antitumor immunity in patients withmetastatic melanoma. Proc Natl Acad Sci. USA.1998;95:13141–13146.
38. Asada H, Kishida T, Hirai H, et al. Significant antitumoreffects obtained by autologous tumor cell vaccine engineeredto secrete interleukin (IL)-12 and IL-18 by means of theEBV/Lipoplex. Mol Ther. 2002;5:609–616.
39. Mach N, Dranoff G. Cytokine-secreting tumor cell vaccines.Curr Opin Immunol. 2000;12:571–575.
40. Huang LR, Chen FL, Chen Y, et al. Potent induction oflong-term CD8+ T cell memory by short-term IL-4exposure during T cell receptor stimulation. Proc Natl AcadSci USA. 2000;97:3406–3411.
41. Dunussi-Joannopoulos K, Dranoff G, Weinstein HJ, et al.Gene immunotherapy in murine acute myeloid leukemia:granulocyte–macrophage colony-stimulating factor tumorcell vaccines elicit more potent antitumor immunitycompared with B7 family and other cytokine vaccines.Blood. 1998;91:222–230.
42. Okada H, Villa L, Attanucci J, et al. Cytokine gene therapyof gliomas: effective induction of therapeutic immunity tointracranial tumors by peripheral immunization with inter-leukin-4 transduced glioma cells. Gene Therapy2001;8:1157–1166.
43. Heller L, Pottinger C, Jaroszeski M, et al. In vivoelectroporation of plasmids encoding GM-CSF or inter-leukin-2 into existing B16 melanomas combined withelectrochemotherapy induces long-term antitumour immu-nity. Melanoma Res. 2000;10:577–583.
44. Abe J, Wakimoto H, Yoshida Y, et al. Antitumor effectinduced by GM-CSF gene-modified tumor vaccination:comparison of adenovirus- and retrovirus-mediated genetictransduction. J Cancer Res Clin Oncol. 1995;121:587–592.
45. Borrello I, Pardoll D. GM-CSF-based cellular vaccines: areview of the clinical experience. Cytokine Growth FactorRev. 2002;13:185–193.
46. Toes RE, Blom RJ, van der Voort E, et al. Protectiveantitumor immunity induced by immunization with com-pletely allogeneic tumor cells. Cancer Res. 1996;56:3782–3787.
47. Souberbielle BE, Westby M, Ganz S, et al. Comparison offour strategies for tumour vaccination in the B16-F10melanoma model. Gene Therapy 1998;5:1447–1454.
48. van Slooten ML, Storm G, Zoephel A, et al. Liposomescontaining interferon-gamma as adjuvant in tumor cellvaccines. Pharm Res. 2000;17:42–48.
49. Blaya C, Crespo J, Crespo A, et al. Anti-interleukin 4antibody and indomethacin synergistic effect on B16melanoma tumor progression. J Pharmacol Exp Ther.1996;279:472–477.
Nonviral vector-mediated cell vaccinesI Moret-Tatay et al
897
Cancer Gene Therapy





























