Embed Size (px)
Citation preview

After online publication, subscribers (personal/institutional) to this journal will haveaccess to the complete article via the DOI using the URL:
If you would like to know when your article has been published online, take advantageof our free alert service. For registration and further information, go to:
.
Due to the electronic nature of the procedure, the manuscript and the original figureswill only be returned to you on special request. When you return your corrections,please inform us, if you would like to have these documents returned.
Dear Author
Here are the proofs of your article.
• You can submit your corrections online, via e-mail or by fax.
• For online submission please insert your corrections in the online correction form.
Always indicate the line number to which the correction refers.
• You can also insert your corrections in the proof PDF and email the annotated PDF.
• For fax submission, please ensure that your corrections are clearly legible. Use a fine
black pen and write the correction in the margin, not too close to the edge of the page.
• Remember to note the journal title, article number, and your name when sending your
response via e-mail or fax.
• Check the metadata sheet to make sure that the header information, especially author
names and the corresponding affiliations are correctly shown.
• Check the questions that may have arisen during copy editing and insert your
answers/corrections.
• Check that the text is complete and that all figures, tables and their legends are included.
Also check the accuracy of special characters, equations, and electronic supplementary
material if applicable. If necessary refer to the Edited manuscript.
• The publication of inaccurate data such as dosages and units can have serious
consequences. Please take particular care that all such details are correct.
• Please do not make changes that involve only matters of style. We have generally
introduced forms that follow the journal’s style.
• Substantial changes in content, e.g., new results, corrected values, title and authorship are
not allowed without the approval of the responsible editor. In such a case, please contact
the Editorial Office and return his/her consent together with the proof.
• If we do not receive your corrections within 48 hours, we will send you a reminder.
• Your article will be published Online First approximately one week after receipt of your
corrected proofs. This is the official first publication citable with the DOI. Further
changes are, therefore, not possible.
• The printed version will follow in a forthcoming issue.
Please note
http://www.link.springer.com
http://dx.doi.org/10.1007/s13277-014-1966-1

AUTHOR'S PROOF
Metadata of the article that will be visualized in OnlineFirst
1 Article Title Concomitant upregulation of nucleostemin and downregulation
of Sox2 and Klf4 in gastric adenocarcinoma
2 Article Sub- Title
3 Article Copyright -Year
International Society of Oncology and BioMarkers (ISOBM) 2014(This will be the copyright line in the final PDF)
4 Journal Name Tumor Biology
5
Corresponding
Author
Family Name Mowla
6 Particle
7 Given Name Seyed Jav ad
8 Suffix
9 Organization Tarbiat Modares University
10 Division Department of Molecular Genetics, Faculty ofBiological Sciences
11 Address Tehran, Iran
12 e-mail [email protected]
13
Author
Family Name Asadi
14 Particle
15 Given Name Malek Hossein
16 Suffix
17 Organization Graduate University of Advanced Technology
18 Division Department of Biotechnology, Institute ofEnvironmental Science
19 Address Kerman, Iran
20 Organization Tarbiat Modares University
21 Division Department of Molecular Genetics, Faculty ofBiological Sciences
22 Address Tehran, Iran
23 e-mail
24
Author
Family Name Derakhshani
25 Particle
26 Given Name Ali
27 Suffix
28 Organization Graduate University of Advanced Technology

AUTHOR'S PROOF
29 Division Department of Biotechnology, Institute ofEnvironmental Science
30 Address Kerman, Iran
31 e-mail
32
Schedule
Received 8 February 2014
33 Revised
34 Accepted 11 April 2014
35 Abstract Nucleostemin (NS) is a nucleolar protein involved in stem cell (SC)self-renewal by controll ing cell cycle progression. In addition toSCs, NS is also expressed in some highly proliferating cellsincluding several adult stem cells and cancer cell l ines. NSknock-down in different cell l ines demonstrated its celltype-dependent function in arresting cell cycle in either G1 or G2/Mphases. Here, we have evaluated the expression of NS and iPSgenes in 36 gastric cancer and their matched marginal nontumortissues by means of real-time polymerase chain reaction (RT-PCR).We have also examined a potential causative role of NS in gastrictumorigenesis by suppressing its expression in a gastric cancer celll ine, AGS. Our data revealed that NS expression level is much
higher in tumor tissues (p = 0.046), especially in high-grade ones (p < 0.001), whereas the expression of Klf4 and Sox2 is downregulated
in tumor tissues compared to marginal nontumor samples (p < 0.001). Furthermore, NS suppression in the AGS cell l ine causedsome morphological alterations, a cell cycle arrest at G1 phase,and an upregulation of iPS genes: Nanog, Sox2, and Klf4. Basedon our results, NS overexpression seems to have a causative role ingastric tumorigenesis and/or progression, and it could beconsidered as a potential tumor marker for diagnosis, molecularclassification, and molecular therapy of gastric adenocarcinoma.
36 Keywordsseparated by ' - '
Nucleostemin - Gastric cancer - RNAi - Cancer stem cell - iPS
37 Foot noteinformation

AUTHOR'S PROOF
UNCORRECTEDPROOF
1
23 RESEARCH ARTICLE
4 Concomitant upregulation of nucleostemin and downregulation5 of Sox2 and Klf4 in gastric adenocarcinoma
7 Malek Hossein Asadi & Ali Derakhshani &8Q2 Seyed Javad Mowla9
10
11 Received: 8 February 2014 /Accepted: 11 April 201412 # International Society of Oncology and BioMarkers (ISOBM) 2014
13 Abstract Nucleostemin (NS) is a nucleolar protein involved14 in stem cell (SC) self-renewal by controlling cell cycle pro-15 gression. In addition to SCs, NS is also expressed in some16 highly proliferating cells including several adult stem cells17 and cancer cell lines. NS knock-down in different cell lines18 demonstrated its cell type-dependent function in arresting cell19 cycle in either G1 or G2/M phases. Here, we have evaluated20 the expression of NS and iPS genes in 36 gastric cancer and21 their matched marginal nontumor tissues by means of real-22 time polymerase chain reaction (RT-PCR). We have also23 examined a potential causative role of NS in gastric tumori-24 genesis by suppressing its expression in a gastric cancer cell25 line, AGS. Our data revealed that NS expression level is much26 higher in tumor tissues (p=0.046), especially in high-grade27 ones (p<0.001), whereas the expression of Klf4 and Sox2 is28 downregulated in tumor tissues compared to marginal29 nontumor samples (p<0.001). Furthermore, NS suppression30 in the AGS cell line caused some morphological alterations, a31 cell cycle arrest at G1 phase, and an upregulation of iPS genes:32 Nanog, Sox2, and Klf4. Based on our results, NS overexpres-33 sion seems to have a causative role in gastric tumorigenesis34 and/or progression, and it could be considered as a potential35 tumor marker for diagnosis, molecular classification, and36 molecular therapy of gastric adenocarcinoma.
37 Keywords Nucleostemin . Gastric cancer . RNAi . Cancer38 stem cell . iPS
39Introduction
40Nucleostemin (NS) is a nucleolar protein primarily expressed41in embryonic and adult stem cells (SCs) [1–5]. NS has two42GTP-binding domains that control the dynamic shuttling of43the protein between the nucleolus and the nucleoplasm [6].44Based on early observations, NS had been reported to regulate45cell proliferation and growth by controlling ribosome biosyn-46thesis and suppressing of P53 [1, 7–9]. Moreover, expression47of NS has been shown to delay cellular senescence by nega-48tively regulating telomeric repeat-binding factor1 (TRF1) sta-49bility [10]. However, other reports suggest that NS may also50contribute to cell proliferation in a P53-independent manner51[4, 11].52In addition to stem cells, NS is also known to be expressed53in several cancer cell lines, as well as many types of normal54proliferating cells. For instance, NS is expressed in normal55kidney cells and renal carcinoma cells at comparable levels56[12–14]. NS knock-down experiments in several stem cells57and cancer cell lines have revealed a cell type-dependent58function of NS in arresting cell proliferation in either G1 or59G2/M phases of cell cycle [1, 4, 14–19]. In a recent study,60tumor cells forced to overexpress NS exhibiting increased61expression of ES-specific genes and enhanced tumorigenesis62[20]. In another report, nucleophosmin was shown to directly63interact with NS in the nucleoli of human tumor cells, where64they individually form complexes with ES-specific transcrip-65tion factors OCT4, Nanog and Sox2 [21].66Despite the vital role of NS in stem and tumor cells, the67molecular links between NS and stem cell self-renewal,68multipotency, embryogenesis, and/or tumorigenesis is not69well understood. Thus, to further elucidate the expression70and function of NS in tumor cells and tissues, we have71investigated here the potential expression of NS and iPS genes72in a series of gastric cancer samples. We further evaluated a73potential causative role of NS in gastric tumorigenesis by
M. H. Asadi :A. DerakhshaniDepartment of Biotechnology, Institute of Environmental Science,Graduate University of Advanced Technology, Kerman, Iran
M. H. Asadi : S. J. Mowla (*)Department of Molecular Genetics, Faculty of Biological Sciences,Tarbiat Modares University, Tehran, Irane-mail:Q1 [email protected]
Tumor Biol.DOI 10.1007/s13277-014-1966-1
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
74 suppressing its expression in a gastric adenocarcinoma cell75 line, AGS, by means of RNAi strategy.
76 Methods and materials
77 Clinical samples collection
78 Tumor and nontumor (apparently normal tissues from the79 margin of the same tumors, as control) surgical specimens80 from 36 patients with gastric adenocarcinoma were provided81 by the Iran National Tumor Bank, funded by Cancer Institute82 of Tehran University, for Cancer Research. Briefly, the ob-83 tained samples had been immediately snap-frozen in liquid84 nitrogen and had been stored in −70 °C until being used for85 RNA extraction. A record of clinicopathological parameters86 including grading and staging for each patient were received87 along with each tissue sample (Table 1). The experiment88 design had been initially approved by the Ethics Committee89 of Tarbiat Modares University.
90 Cell culture
91 The gastric adenocarcinoma cell line, AGS, was obtained92 from the national cell bank of Iran (Pasteur Institute of Iran,93 Tehran) and was cultured at 37 °C in a humidified atmosphere94 of 5 % CO2 in RPMI1640 medium (Gibco, USA), supple-95 mented with penicillin/streptomycin (100 U/ml and96 100 μg/ml, respectively), and 10 % fetal bovine serum. The97 human embryonic carcinoma cell line NTERA2 (NT2) was98 cultured in Dulbecco’s modified eagle medium (DMEM) with99 high concentration of glucose (4,500 mg/l), supplemented100 with 10 % fetal bovine serum at 37 °C (humidified) and 5 %101 CO2.
102 Immunocytochemistry
103 AGS cells were fixed in 4 % paraformaldehyde for 15 min at104 room temperature. Cells were then permeabilized in 0.25 %105 Triton X-100 [0.1 % Tween-20 in phosphate-buffered saline106 (PBS)] for 10 min and blocked in 1 % BSA in PBST for107 30 min. The cells were then incubated with 1:50 diluted first108 antibody (an affinity-purified goat polyclonal anti-NS
109antibody, sc-46219, Santa Cruz Biotechnology) in PBST con-110taining 1 % BSA for 1 h at room temperature. After washing111the cells three times with PBS, cells were incubated with the112secondary antibody (an anti-goat IgG conjugated with FITC;113Sigma-Aldrich) at dilutions 1:100 in 1 % BSA for 1 h at room114temperature. The cells were then washed three times with PBS115and visualized under a fluorescent microscope (Nikon, Ger-116many). For the negative control sample, all conditions were117kept the same, except that the primary antibody was omitted.
118Immunohistochemistry
119Formalin-fixed paraffin-embedded (FFPE) gastric tissue sec-120tions (5 μm) were deparaffinized with xylene, rehydrated in121descending concentrations of ethanol and boiled for 10 min in122citrate buffer (10 mM, pH 9.0). Endogenous peroxidase ac-123tivity was suppressed with 1.5 % H2O2 for 30 min. Slides124were then serum-blocked with normal goat serum and incu-125bated with anti-NS polyclonal antibody (R&D Systems,126Abingdon, UK; 1:100 dilution) followed by incubation with127secondary HRP-conjugated anti-goat antibody (Abcam, Cam-128bridge, UK; 1:500), both for overnight at 48 °C. In negative129controls, all the conditions were kept the same, except for the130omission of NS antibody.
131RNA extraction, reverse transcription, semiquantitative132RT-PCR, and real-time PCR
133Total RNAwas isolated using RNX plus solution (CinnaGen,134Tehran, Iran) according to the manufacturer’s instructions.135Purity and integrity of the extracted RNA were measured by136optical density (260/280 nm ratio) as well as visual observa-137tion of the samples on agarose gel electrophoresis. RNase-free138DNase (Fermentase, Lithuania) treatment of the total RNA139was performed according to the manufacturer’s instructions.140The first strand of cDNAwas synthesized at 42 °C for 60 min141in the presence of 1 μg RNA and 200 U/μl MMLV reverse142transcriptase (Fermentase), 20 U RNase inhibitor, dNTP mix143(final concentration of 1 mM) with oligo(dT)18 priming in a14420 μl reaction.145Specific primers and probes were designed for NS,146GAPDH, Klf4, Nanog, Sox2, Oct4, and P21 using AlleleID1474.0 and Genrunner softwares (Table 2). The PCR
t1:1 Table 1 Characteristics of pa-tients with gastric cancert1:2 Tumor characteristics
t1:3 Grade differentiation Tumor staging Gender
t1:4 I II III IV T1 T2 T3 T4 Male Female
t1:5 9 9 14 4 2 5 26 3 25 11
t1:6 25 % 25 % 39 % 11 % 6 % 14 % 72 % 8 % 69 % 31 %
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
148 amplification of cDNAs were carried out by using 2 μl of the149 first cDNA strand, 1 unit of Taq DNA polymerase, 2.5 μl of150 10× PCR buffer, 0.5 μl of 20 mM dNTPs, 0.8 μl of 20 pmol151 forward primer, 0.8 μl of 20 pmol reverse primer, and added152 water to reach the volume of 25μl. The PCR carried out under153 the following conditions: initial denaturation at 94 °C for154 2 min, a 30 cycle amplification (94 °C for 30 s, 58.5 °C for155 30 s, 72 °C for 45 s), and a final extension at 72 °C for 5 min.156 Real-time PCR reactions were performed using Taqman157 method with premix Ex TaqTM (Takara, Japan) on an ABI158 7500 real-time quantitative PCR system. PCR reaction mix-159 ture (12.5 μl premix Ex Taq™, 0.2 μM forward primer,160 0.2 μM reverse primer, 0.1 μM TaqMan probe, 0.5 μl ROX161 reference dye II, 2 μl template, and 8 μl dH2O) was added,162 and PCR was carried out with the following cycling condi-163 tions: initiation at 94 °C for 30 s, amplification for 45 cycles164 with denaturation at 94 °C for 5 s, annealing and extending at165 60 °C for 34 s. To determine the reaction efficiencies for each
166primer pair and the corresponding probe, standard curves were167plotted by using serial dilutions of NT2 cDNA. All reaction168efficiencies were measured to be close to 100 %.169All experiments were conducted in duplicate or triplicate.170Groupwise comparison and statistical analysis of relative ex-171pression results of real-time PCR were carried out by REST1722008 (Relative Expression Software Tool, V2.0.7, Corbett173Research).
174Small interfering RNA (siRNA)
175The siRNA sequences used for the suppression of human NS176were the same as the ones used by Tsai and McKay (2002).177Moreover, an irrelevant siRNA was designed by the178Selection Program (Whitehead Institute for Biomedical179Research; http://jura.wi.mit.edu/) and synthesized by180MWG Company (Germany) with the following sense and181antisense sequences:
182& NS–siRNA
183– Sense: GAACUAAAACAGCAGCAGdTdT184– Antisense: UCUGCUGCUGUUUUAGUUCdTdT
185& IR–siRNA
186– Sense: GCGGAGAGGCUUAGGUGUAdTdT187– Antisense: UACACCUAAGCCUCUCCGCdTdT
188One day before siRNAs transfection, 2×105 cells per well189were cultured on a six-well plate in growth medium without190antibiotic, such that they were 30–50 % confluent at the time191of transfection. siRNAs were transfected into the cells using192the Lipofectamine™ RNAiMAX Transfection Reagent193(Invitrogen, UK). Briefly, 5 μl siRNA (20 μM solution) and1944.5 μl RNAiMAX reagent were diluted in 250 μl Opti-MEM195(Invitrogen, UK) medium. The combined diluted RNAi du-196plex with diluted Lipofectamine™ RNAiMAXwas incubated197for 15 min in room temperature. The mixture was then added198to the cells in a final volume of 2.5 ml. Cells were incubated1993 days at 37 °C in a CO2 incubator.
200Cell cycle analysis
201For cell cycle analysis, AGS cells were washed with202phosphate-buffered saline (PBS) and carefully dissociated to203yield single cell suspensions with 0.025 % trypsin–EDTA,204followed by two washes in growth medium. Then, the cells205were stained with 50 μg/ml propidium iodide solution con-206taining 0.1 % Triton X-100 and sodium citrate as described207elsewhere [21], and the single cell suspensions were used for208flow cytometric analysis (Partec, Germany). Cell cycle pro-209files were analyzed using Partec Flomax software.
t2:1 Table 2 Designed primers and probes
t2:2 Genes Amplicanlength
Sequence Designed oligo
t2:3 Nucleostemin 118 ATAGCATCCTTTTCCAGTCTTCCG
F
t2:4 TCTTTGTCATCCTCCCCTCTCC
R
t2:5 CTTCCTTTCTTTCCGTTTTGGCA
P
t2:6 GAPDH 123 GTGACCATGAGAAGTATGACAAC
F
t2:7 CATGAGTCCTTCCACGATACC
R
t2:8 CCTCAAGATCATCAGCAATGCCTCCTG
P
t2:9 Nucleostemin 646 CAGAGATCCTCTTGGTTGCAG
F
t2:10 AATGAGGCACCTGTCCACTC
R
t2:11 Klf4 410 GGATGGAAATTCCGCCCGCTCAG
F
t2:12 TTGGCTTCCTTGTTTGGTACC
R
t2:13 P21 385 GGGCGGCCAGGGTATGTACA
F
t2:14 TTCGGCCCAGTGGACAGCGA
R
t2:15 Oct4 470 GAGAATTTGTTCCTGCAGTGC
F
t2:16 GTTCCCAATTCCTTCCTTAGTG
R
t2:17 Nanog 470 ACCTATGCCTGTGATTTGTGG
F
t2:18 AAGAGTAGAGGCTGGGGTAGG
R
t2:19 Sox2 426 ATGGGTTCGGTGGTCAAGTC
F
t2:20 GTGGATGGGATTGGTGTTCTC
R
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
210 Results
211 NS is significantly overexpressed in high-grade gastric212 adenocarcinoma
213 We used a real-time RT-PCR approach to measure the expres-214 sion level of NS in 36 tumor samples obtained from patients215 with gastric adenocarcinoma. We also used apparently normal216 tissues from the margins of the same tumors, as controls. The217 data were also normalized to the expression of GAPDH, as an218 internal house-keeping gene. The expression ratios of NS/219 GAPDH were compared between tumors vs. nontumors, as220 well as between high-grade vs. low-grade subgroups of tumor221 samples. NS expression was detectable in both tumor and222 nontumor samples of stomach. Moreover, as it is evident in223 Fig. 1a, the expression of NS was noticeably higher in tumor224 group in comparison to that of nontumor (marginal; p=0.046).225 Among the tumor samples, NS expression was significantly226 higher in high-grade tumors compared to the low-grade ones227 (p<0.001; Fig. 1b).
228 Downregulation of Sox2 and Klf4 in gastric adenocarcinoma
229 To study the possibility of iPS genes expression in gastric230 adenocarcinoma, primers and probes were carefully designed231 for specific amplification of Nanog, Klf4, Sox2, and Oct4232 genes. From the list of iPS genes, we already reported [21]233 the expression level of Oct4 variants in gastric adenocarcino-234 ma, where our results revealed that the expression of all Oct4235 variants were significantly elevated in tumor groups, com-236 pared to that of nontumor ones.
237We observed no expression of Nanog, neither in tumor nor238in nontumor samples of the stomach. In contrast, Sox2 and239Klf4 were significantly downregulated in tumor group com-240pared to that of nontumor ones (p<0.001, Fig. 2a and b). Our241result showed that no significant difference between the ex-242pression level of Klf4 and Sox2 in high-grade vs. low-grade of243gastric adenocarcinoma.
244Tissue and subcellular distribution of NS protein in gastric245adenocarcinoma samples
246Using a polyclonal anti-NS antibody, we examined the ex-247pression, tissue distribution, as well as subcellular localization248of NS protein in gastric adenocarcinoma tissues by means of249immunohistochemistry (IHC). As expected, NS was primarily250localized within the nucleoli of the tumor cells, with no251staining signal within the cytoplasm of the positive cells252(Fig. 3a, c). We also observed the expression of NS in a group253of normal cells located at the bottom of tubular structures of254the epithelium adjacent to the basal lamina (nontumor). The255latter finding was in agreement with our results obtained from256real-time PCR (Fig. 3b, d). No immunoreactivity was ob-257served in negative controls, which obtained from the same258specimen and were similarly processed, except for the omis-259sion of the first antibody.
260Morphological and functional alterations following NS261suppression in AGS cells
262To evaluate a potential causative role of NS in tumorigenesis263of gastric adenocarcinoma, we suppressed the expression of
Fig. 1 Histograms comparing therelative gene expression of NS toGAPDH (the internal control) asdetermined by quantitative RT-PCR. a The relative expression ofNS in 36 tumor and their matchednontumor samples. b The relativeexpression of NS in high-gradevs. low-grade tumors. Duplicateexperiments were performed foreach sample and the values areshown as the mean ± SD
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014
AUTHOR'S PROOF
UNCORRECTEDPROOF
264 NS in a well-characterized cell line of gastric adenocarcinoma,265 AGS. The expression of NS at mRNA level was dramatically266 reduced after the cells were transfected with NS–siRNA. As267 expected, the transfection of cells with an irrelevant siRNA/IR268 siRNA, with no specific target within the human genome,
269showed no obvious change in the level of endogenous NS270(Fig. 4a). Compared to the AGS cells that were treated with271IR–siRNA, the NS–siRNA-treated cells demonstrated a272prominent alteration in the cell cycle distribution, as measured273by flow cytometry. As it is evident in Fig. 4b and c, NS
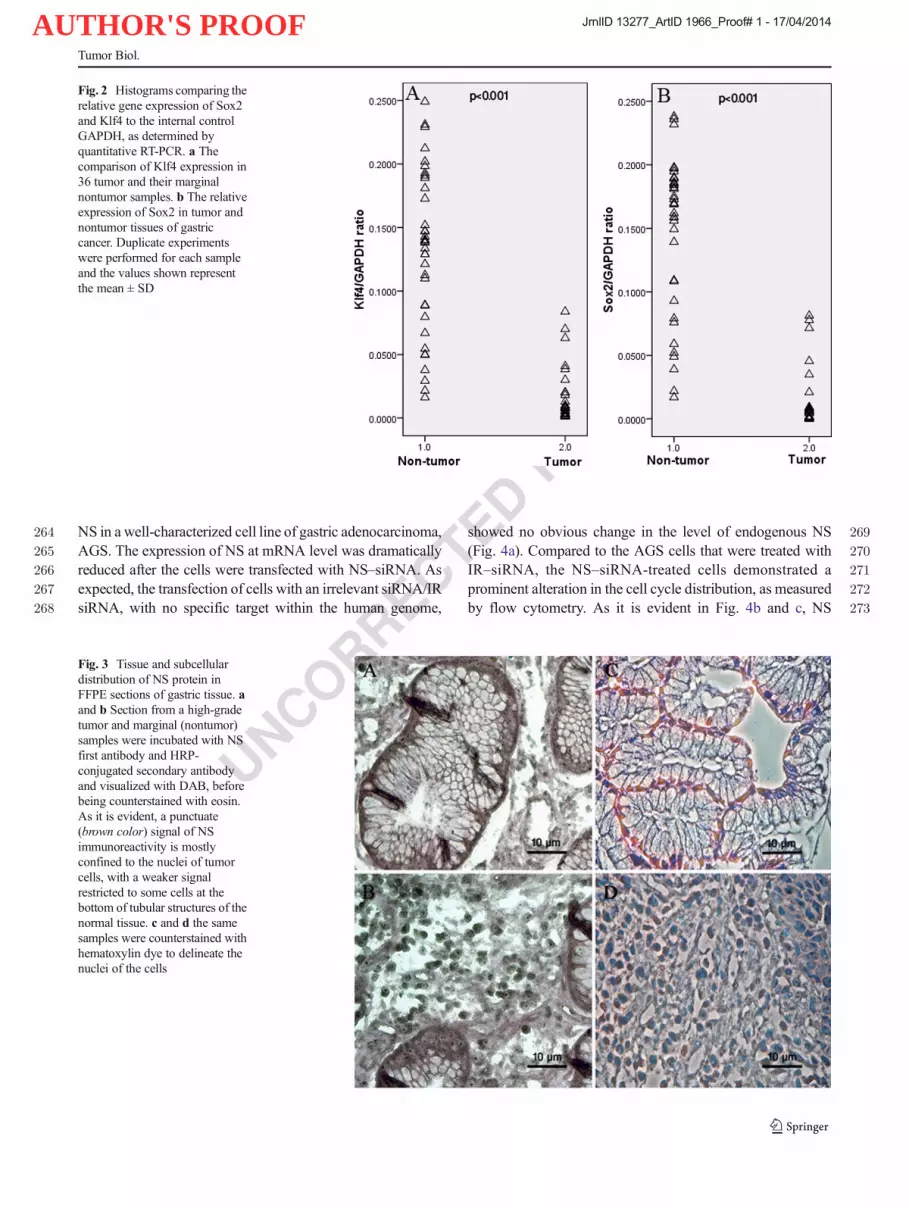
Fig. 2 Histograms comparing therelative gene expression of Sox2and Klf4 to the internal controlGAPDH, as determined byquantitative RT-PCR. a Thecomparison of Klf4 expression in36 tumor and their marginalnontumor samples. b The relativeexpression of Sox2 in tumor andnontumor tissues of gastriccancer. Duplicate experimentswere performed for each sampleand the values shown representthe mean ± SD
Fig. 3 Tissue and subcellulardistribution of NS protein inFFPE sections of gastric tissue. aand b Section from a high-gradetumor and marginal (nontumor)samples were incubated with NSfirst antibody and HRP-conjugated secondary antibodyand visualized with DAB, beforebeing counterstained with eosin.As it is evident, a punctuate(brown color) signal of NSimmunoreactivity is mostlyconfined to the nuclei of tumorcells, with a weaker signalrestricted to some cells at thebottom of tubular structures of thenormal tissue. c and d the samesamples were counterstained withhematoxylin dye to delineate thenuclei of the cells
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
274 suppression in AGS cells resulted in an elevation of cell275 distribution within the G1 phase (77±0.1 vs. 54±0.2 % in276 IR–siRNA-treated cells). Consequently, the percentages277 of the cells within S and G2+M phases were decreased278 (S phase: 18±0.1 vs. 30±0.2 %, G2: 16±0.2 vs. 5±279 0.01 % in IR–siRNA-treated cells).280 The expression of NS in AGS cell line was further con-281 firmed at the protein level (Fig. 5c), by using a polyclonal282 antibody against a N-terminal segment of NS. The immuno-283 cytochemistry (ICC) data confirmed the expression and284 localization of NS protein in nucleoli of the IR–siRNA-285 treated cells. In contrast, there were some apparent286 morphological changes in the AGS cells, 2–3 days after287 transfection with NS–siRNA. The almost round shape of288 the untreated and/or IR–siRNA-treated AGS cells
289changed into a hummingbird-like appearance in NS–290siRNA-treated cells (Fig. 5a, b). Furthermore, there291existed no or very low immunoreactivity for NS protein292in NS-suppressed cells, confirming the accuracy of NS293suppression at protein level.
294Upregulation of iPS genes and P21 after NS suppression
295A likely key to understanding the actions of NS in tumorigen-296esis would be to define the proteins with which it interacts. An297interaction between N-terminal basic region of NS and tumor298suppressor gene P53 leads to P53 inactivation. For this reason,299we examined the expression alteration of P21, the main300downstream target of P53, as well as those of Klf4, a suppres-301sor of P53 and inducer of P21, following the inhibition of NS
Fig. 4 NS suppression in AGScells. a NS knock-down in AGScells by using specific siRNAagainst NS. A significantdownregulation of NS expressionis confirmed by asemiquantitative RT-PCR atmRNA level. GAPDH was usedas an internal control. b and cEffect of NS knock-down on cellcycle distribution of AGS cellline. NS–siRNA treatment ofAGS cells resulted in a significantelevation in the percentage of thecells in the G1 phase (77±0.1 %vs. 54±0.2 % in IR–siRNA-treated cells). Consequently, adecreased number of distributedcells within S and G2+M phasesare also evident
Fig. 5 aMorphological appearance of AGS cells stained with Geimsa. bThe altered morphology of the cells after treatment with NS–siRNA.Notethe altered, hummingbird, morphology of the cells after NS–siRNAtreatment. c Immunocytochemistry of NS expression in AGS cells, using
an affinity-purified goat polyclonal anti-NS antibody. NS immunoreac-tivity gave punctuate signals corresponding to the nucleolar localizationof the protein within AGS cells treated with IR-siRNA
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
302 in AGS cells. In addition, based on a recent report claiming303 that NS expression in human cancer cells confers a tumori-304 genic phenotype by expressing iPS factors [20] and also the305 interaction between these factors and nucleophosmin, a mul-306 tifunctional nucleolar phosphoprotein that has been reported307 directly interact with NS, motivated us to evaluate a possible308 correlation between iPS genes and NS. In doing so, we309 assessed the changes in the expression level of these genes310 after NS suppression in AGS cells. Our data revealed that311 Klf4, Sox2, and Nanog as well as p21 were upregulated after312 NS suppression in AGS cells (p<0.001, Fig. 6). However, the313 expression of Oct4 was not significantly altered by NS314 suppression.
315 Discussion
316 In the current study, we have initially quantified the expres-317 sion alteration of NS and iPS genes in a series of gastric318 adenocarcinoma specimens. Compared to the apparently nor-319 mal tissues obtained from the margin of same tumors, NS320 showed a significant upregulation in tumor samples. More-321 over, while neither Nanog nor lin28 expressed in tumor and322 nontumor samples, Klf4 and Sox2 showed a significant down-323 regulation in tumor samples. As we reported before [22],324 similar to what we observed for NS, Oct4 variants (Oct4A,325 Oct4B, and Oct4B1) are overexpressed in gastric adenocarci-326 noma. Therefore, the expression of NS seems to have a327 negative correlation with those of iPS genes, except Oct4, in328 gastric adenocarcinoma. While we observed a moderate over-329 expression of NS in tumor samples vs. nontumor ones, the330 level of upregulation was much higher in high-grade tumors in331 comparison to that of low-grade group. The latter observation332 suggests that NS has a better suitability, as a potential tumor
333marker, for discriminating high-grade tumors from low-grade334and/or nontumor ones. The same finding has been already335reported for human esophageal squamous cell carcinoma [23]336and prostate cancer [24]. In both reports, the expression of NS337was found to be negatively correlated with the degree of cell338differentiation. The elevated overexpression of NS in high-339grade tumors, with higher proliferation rate and lower differ-340entiation state, is an expected finding especially with the new341reports which state the involvement of NS in ribosome bio-342synthesis [7, 8].343We further performed immunohistochemistry to examine344the expression of NS at the protein level and also its localiza-345tion on gastric samples. In agreement with our real-time PCR346data, we determined the nucleolar localization of NS in almost347all of cancerous cells. Moreover, the restricted localization of348NS in some cells located at the bottom of tubular structures,349where the highly dividing cells are located, is in agreement350with lower expression of NS transcript in normal gastric351tissue.352To further explore the functional role of NS in gastric tumor353initiation/progression and its correlation with iPS genes, we354suppressed its expression in a gastric adenocarcinoma cell355line, AGS. As expected, following NS suppression a signifi-356cant change in the morphology and cell cycle distribution of357the cells was noticed. The treated cells showed hummingbird358morphology, suggesting that their morphology has been359changed from an undifferentiated into a more differentiated360state. A similar conclusionwas obtained on comparing the cell361cycle distribution of the NS–siRNA-transfected cells with362those transfected with IR–siRNA.363Our cell cycle analysis of the NS–siRNA-transfected AGS364(a p53-positive cell line) cells by means of flow cytometry365revealed a G1 arrest of the treated cells. A similar finding has366been already reported for most of the NS knocked-down in367stem/cancer cells [4, 15, 16]. Moreover, a sub-G1 and G2+M368arrest of NS+/−mouse embryonic fibroblasts (MEFs) has been369also reported [10]. Interestingly, in a report by Nikpour et al.370(2009), a differential mechanism of action for NS in two371bladder cancer cell lines was elucidated, which suggest NS372may not only depend on p53 but also on the status of RB1. In373the latter work, NS–siRNA treated 5,637 cells accumulated374mainly in G2+M, whereas SW1710 cells arrested in G0/G1375phase of cell cycle.376In the next phase of our study, we assessed the expression377of iPS genes and P21 after NS suppression in AGS cells. In378agreement with our aforementioned data on expression of NS379and iPS genes in gastric adenocarcinoma tissues, the suppres-380sion of NS leads to a significant upregulation of Klf4, Sox2,381and Nanog in the treated cells.382Based on original observations, NS was identified as a383nucleolar protein functioning via controlling the proliferation384of stem/cancer cells [1]. Subsequently, it was realized that NS385is expressed in highly proliferating cells, binding, and
Fig. 6 The RT-PCR analysis of P21, Klf4, and iPS genes in AGS cellstreated with NS–siRNA, compared to the cells treated with IR–siRNA.Relative (normalized to GAPDH) expression of these genes after sup-pression of NS in AGS cells compared to the cells treated with IR–siRNA. P21 and iPS genes, but not Oct4, are significantly overexpressedin the cells after NS inhibition
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
386 inactivating P53. Accordingly, upon induction of differentia-387 tion the expression of NSwas rapidly turned off [10], which in388 turn caused a rapid elevation in P53 level, and consequently, a389 G1 cell cycle arrest in P53-positive cells [16]. However, our390 data demonstrated that after suppressing NS in AGS cells, the391 expression of P21 was elevated, suggesting that NS could act392 in an apparently P53-dependent manner causing a G1 cell393 cycle arrest. This is in contrast to our previous report in bone394 marrow stromal stem cells [4], where the suppression of NS395 had no effects on the expression of P21, a finding that chal-396 lenged the claimed P53-dependent mechanism of action for397 NS.398 Based on our data, the expression of Klf4 is increased399 following NS suppression. Klf4 is a zinc finger transcription400 factor that is highly expressed in the gastrointestinal tract and401 other epithelial tissues [25]. Furthermore, accumulating clin-402 ical evidence suggest that Klf4 is a potential tumor suppressor,403 and that overexpression of Klf4 in AGS cell line causes a G1404 cell cycle arrest [26, 27]. It is believed that the suppressive role405 of Klf4 is mediated by P21 [28]. Following NS suppression in406 AGS cells, a G1 cell cycle arrest occurs which coincides with407 Klf4 upregulation. Our data suggest that a dual overexpression408 of both Klf4 and P21 might be an indirect consequence of NS409 suppression in AGS cells. In other words, Klf4 could be410 considered as a downstream regulator of NS, and that Klf4411 overexpression might cause a G1 cell cycle arrest via P21-412 dependent pathway. However, the proposed claim still needs413 to be experimentally validated under different conditions in-414 cluding in the present or absent of functional P53.415 Altogether, our data suggest that NS overexpression have a416 causative role in gastric tumorigenesis and/or progression, and417 therefore, it could be considered as a potential molecular418 marker for diagnosis, molecular classification, and gene-419 based therapy of gastric adenocarcinoma.
420 Acknowledgments We thank Dr. Hamid-Reza Kalhor for his valuable421 comments. This work was supported, in part, by a research grant from the422 Iranian stem cell network.
423424 Conflicts of interest None.
425 References426
427 1. Tsai RY, McKay RD. A nucleolar mechanism controlling cell prolif-428 eration in stem cells and cancer cells. Genes Dev. 2002;16:429 2991–3003.430 2. Baddoo M, Hill K, Wilkinson R, Gaupp D, Hughes C, Kopen GC,431 et al. Characterization of mesenchymal stem cells isolated from432 murine bone marrow by negative selection. J Cell Biochem.433 2003;89:1235–49.434 3. Politz JC, Polena I, Trask I, Bazett-Jones DP, Pederson T. A435 nonribosomal landscape in the nucleolus revealed by the stem cell436 protein nucleostemin. Mol Biol Cell. 2005;16:3401–10.
4374. Jafarnejad SM, Mowla SJ, Matin MM. Knocking-down the expres-438sion of nucleostemin significantly decreases rate of proliferation of439rat bone marrow stromal stem cells in an apparently p53-independent440manner. Cell Prolif. 2008;41:28–35.4415. Kermani AJ, Fathi F, Mowla SJ. Characterization and genetic ma-442nipulation of human umbilical cord vein mesenchymal stem cells:443potential application in cell-based gene therapy. Rejuvenation Res.4442008;11:379–86.4456. Tsai RY, McKay RD. A multistep, GTP-driven mechanism control-446ling the dynamic cycling of nucleostemin. J Cell Biol. 2005;168:447179–84.4487. Du X, RaoMR, Chen XQ,WuW,Mahalingam S, Bala-sundaram D.449The homologous putative GTPases Grn1p from fission yeast and the450human GNL3L are required for growth and play a role in processing451of nucleolar pre-rRNA. Mol Biol Cell. 2006;17:460–74.4528. Romanova L, Grand A, Zhang L, Rayner S, Katoku-Kikyo N,453Kellner S, et al. Critical role of nucleostemin in pre-rRNA process-454ing. J Biol Chem. 2009;284(8):4968–77.4559. Zhu Q, Yasumoto H, Tsai RY. Nucleostemin delays cellular senes-456cence and negatively regulates TRF1 protein stability. Mol Cell Biol.4572006;26:9279–90.45810. Beckman C, Nichane M, Clercq SD, Maetens M, Floss T, Wurst W,459et al. Evolutionary conserved role of nucleostemin: controlling pro-460liferation of stem/progenitor cells during early vertebrate develop-461ment. Mol Cell Biol. 2006;26:9291–301.46211. Kafienah W, Mistry S, Williams C, Hollander P. Nucleostemin is a463marker of proliferating stromal stem cells in adult human bone464marrow. Stem Cells. 2006;24:1113–20.46512. Liu SJ, Cai ZW, Liu YJ, Dong MY, Sun LQ, Hu GF, et al. Role of466nucleostemin in growth regulation of gastric cancer, liver cancer and467other malignancies. World J Gastroenterol. 2004;10:1246–9.46813. Fan Y, Liu Z, Zhao S, Lou F, Nilsson S, Ekman P, et al. Nucleostemin469mRNA is expressed in both normal and malignant renal tissues. Br J470Cancer. 2006;94:1658–62.47114. Malakootian M, Mowla SJ, Saberi H, Asadi MH, Atlasi Y,472Shafaroudi AM. Differential expression of nucleostemin, a stem cell473marker, and its variants in different types of brain tumors. Mol474Carcinog. 2010;49(9):818–45.47515. Sijin L, Ziwei C, Yajun L, Meiyu D, Hongwei Z, Guofa H, et al. The476effect of nocking-down nucleostemin gene expression on the in vitro477proliferation and in vivo tumorigenesis of HeLa cells. J Exp Clin478Cancer Res. 2004;23:529–38.47916. Ma H, Pederson T. Depletion of the nucleolar protein nucleostemin480causes G1 cell cycle arrest via the p53 pathway. Mol Biol Cell.4812007;18:2630–5.48217. Dai MS, Sun XX, Lu H. Aberrant expression of nucleostemin acti-483vates p53 and induces cell cycle arrest via inhibition of MDM2. Mol484Cell Biol. 2008;28:4365–76.48518. Nikpour P, Mowla SJ, Jafarnejad SM, Fischer U, Schulz WA.486Differential effects of Nucleostemin suppression on cell cycle arrest487and apoptosis in the bladder cancer cell lines 5637 and SW1710. Cell488Prolif. 2009;42:762–9.48919. Okamoto N, YasukawaM, Nguyen C, Kasim V,Maida Y, Possemato490R, et al. Maintenance of tumor initiating cells of defined genetic491composition by nucleostemin. PNAS. 2011;108:20388–93.49220. Johansson H, Simonsson S. Core transcription factors, Oct4,493Sox2 and Nanog, individually form complexes with494nucleophosmin(Npm1) to control embryonic stem (ES) cell495fate determination. Aging. 2010;2(11):815–22.49621. Janssen K, Pohlmann S, Janicke RU, Schulze-Osthoff K, Fischer U.497Apaf-1 and caspase-9 deficiency prevents apoptosis in a Bax-498controlled pathway and promotes clonogenic survival during pacli-499taxel treatment. Blood. 2007;110:3662–72.50022. Asadi MH, Mowla SJ, Fathi F, Asadzadeh J, Atlasi Y. OCT4B1, a501novel spliced variant of OCT4, is highly expressed in gastric cancer502and acts as antiapoptotic factor. Int J Cancer. 2011;128:2645–52.
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
503 23. Zhang G, Zhang Q, Zhang Q, Yin L, Li S, Cheng K, et al. Expression504 of Nucleostemin, epidermal growth factor and epidermal growth505 factor receptor in human esophageal squamous cell carcinoma tis-506 sues. J Cancer Res Clin Oncol. 2013;136:587–94.507 24. Liu RL, Zhang ZH, Zhao WM, Wang M, Qi SY, Li J, et al.508 Expression of nucleostemin in prostate cancer and its effect on the509 proliferation of PC-3 cells. Chin Med J. 2008;121(4):299–304.510 25. Wei D, GongW, Kanai M, Schlunk C,Wang L, Yao JL, et al. Drastic511 downregulation of Krüppel-Like Factor 4 expression is critical in512 human gastric cancer development and progression. Cancer Res.513 2005;65:2746–54.
51426. Wang N, Liu ZH, Ding F, Wang XQ, Zhou CN, Wu M. Down-515regulation of gut-enriched Krüppel-like factor expression in esopha-516geal cancer. World J Gastroenterol. 2002;8:966–70.51727. Masashi K, Daoyan W, Qiang L, Zhiliang J, Jaffer A,518Xiangdong L, et al. Loss of krüppel-like factor 4 expression519contributes to Sp1Overexpression and human gastric cancer520development and progression. Hum Cancer Biol. 2006;12:5216395–402.52228. Rowland BD, Bernards R, Peeper DS. The KLF4 tumor suppressor is523a transcriptional repressor of p53 that acts as a context-dependent524oncogene. Nat Cell Biol. 2005;7(11):1074–82.
525
Tumor Biol.
JrnlID 13277_ArtID 1966_Proof# 1 - 17/04/2014

AUTHOR'S PROOF
UNCORRECTEDPROOF
AUTHOR QUERIES
AUTHOR PLEASE ANSWER ALL QUERIES.
Q1. Please check captured Email address if correct.Q2. (Seyed Javad Mowla) has been set as the corresponding author. Please check and advise if correct.Q3. Please check provided Volume number, first range and year in reference 23 if correct.





























