Embed Size (px)
Citation preview

www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 333 (2005) 858–867
BBRC
Conserved autophosphorylation pattern in activation loopsand juxtamembrane regions of Mycobacterium tuberculosis
Ser/Thr protein kinases
Rosario Duran a, Andrea Villarino b, Marco Bellinzoni b, Annemarie Wehenkel b,Pablo Fernandez b, Brigitte Boitel b, Stewart T. Cole c, Pedro M. Alzari b,*,
Carlos Cervenansky a,*
a Laboratorio de Bioquımica Analıtica, Instituto de Investigaciones Biologicas Clemente Estable and Facultad de Ciencias, Montevideo, Uruguayb Unite de Biochimie Structurale (URA 2185 CNRS), Institut Pasteur, 25/28 rue du Docteur Roux, 75724 Paris Cedex 15, France
c Unite de Genetique Moleculaire Bacterienne, Institut Pasteur, 25/28 rue du Docteur Roux, 75724 Paris Cedex 15, France
Received 23 May 2005Available online 13 June 2005
Abstract
The identification of phosphorylation sites in proteins provides a powerful tool to study signal transduction pathways and toestablish interaction networks involving signaling elements. Using different strategies to identify phosphorylated residues, we reporthere mass spectrometry studies of the entire intracellular regions of four �receptor-like� protein kinases from Mycobacterium tuber-
culosis (PknB, PknD, PknE, and PknF), each consisting of an N-terminal kinase domain and a juxtamembrane region of varyinglength (26–100 residues). The enzymes were observed to incorporate different numbers of phosphates, from five in PknB up to 11 inPknD or PknE, and all detected sites were dephosphorylated by the cognate mycobacterial phosphatase PstP. Comparison of thephosphorylation patterns reveals two recurrent clusters of pThr/pSer residues, respectively, in their activation loops and juxtamem-brane regions, which have a distinct effect on kinase activity. All studied kinases have at least two conserved phosphorylated residuesin their activation loop and mutations of these residues in PknB significantly decreased the kinase activity, whereas deletion of theentire juxtamembrane regions in PknB and PknF had little effect on their activities. These results reinforce the hypothesis that myco-bacterial kinase regulation includes a conserved activation loop mechanism, and suggest that phosphorylation sites in the juxtamem-brane region might be involved in putative kinase-mediated signaling cascades.� 2005 Elsevier Inc. All rights reserved.
Keywords: Ser/Thr kinase autophosphorylation; Mycobacterium tuberculosis; Activation loop; Juxtamembrane region; Phosphopeptide identifica-tion; Mass spectrometry; Bacterial signaling
Protein modification by covalent phosphate attach-ment to specific serine, threonine or tyrosine residuesis a prominent mechanism in signal transduction pro-cesses triggered by environmental stimuli [1]. Long timethought to be restricted to eukaryotic organisms, a massof evidence now indicates that Ser/Thr/Tyr phosphory-
0006-291X/$ - see front matter � 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2005.05.173
* Corresponding authors. Fax: +33 145688604 (P.M. Alzari).E-mail addresses: [email protected] (P.M. Alzari), [email protected].
uy (C. Cervenansky).
lation also takes place in prokaryotes [2], in parallel to(and possibly interacting with) the prevailing two-com-ponent His phosphorylation systems [3]. It is nowknown that these �eukaryotic-like� signaling systems,relying on Ser/Thr- or Tyr-protein kinases, are directlyinvolved in bacterial processes such as developmentalregulation, control of cell growth, stress responses, reg-ulation of secondary metabolite production, infectivity,and survival in host cells [4–8]. One relevant exampleis the human pathogen Mycobacterium tuberculosis,

R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867 859
whose genome includes genes encoding as many as 11functional Ser/Thr protein kinases (compared to a sim-ilar number of functional two-component systems), oneprotein Ser/Thr phosphatase (PstP), and two proteinTyr phosphatases [9]. Most of these enzymes have al-ready been produced as recombinant proteins and foundto have biochemical properties similar to their eukaryot-ic homologues, including autophosphorylation as a pre-sumptive mechanism of activity modulation [10–18].
Ser/Thr protein kinases catalyze protein phosphory-lation by transferring a phosphate group from the c-po-sition of ATP to the side-chain hydroxyl group of Ser orThr residues in protein substrates. While these enzymesshare a well-conserved catalytic scaffold and active sitemachinery, extensive studies of eukaryotic enzymesstressed the remarkable conformational plasticity ofthe kinase domain, which gives rise to quite differentmechanisms of activity regulation in response to auto-phosphorylation and/or interactions with specific regu-latory domains [19,20]. However, except for thecatalytic domain of PknB, whose activity was shownto be regulated by phosphorylation in the activationloop [14,15], no information is currently available onthe autophosphorylation pattern and possible regulationmechanisms of mycobacterial kinases.
In the present work, we combined matrix-assisted la-ser desorption/ionization time of flight mass spectrome-try (MALDI-TOF MS) with reverse-phase HPLCseparations, enzymatic treatments, and specific chemicalmodifications to identify the autophosphorylation sitesin the entire intracellular regions of four receptor-likeSer/Thr protein kinases from M. tuberculosis. Theemerging pattern of phosphorylation and the biochemi-cal and biological implications of protein kinase auto-phosphorylation are discussed; data supporting a
Table 1Oligonucleotides used in this study
Gene
PknD
PknE
PknF
Mutation
PknB K40fi A
PknB E280fi Stop
PknD S279fi Stop
PknE A278fi Stop
PknF A281fi Stop
a Underlined letters indicate restriction sites (NdeI for forward primers anb Bold and underlined letters indicate modified nucleotides.
regulatory role for the cognate Ser/Thr phosphoproteinphosphatase (PstP) are also presented.
Materials and methods
Cloning and mutagenesis. Subcloning of the intracellular region ofPknB (residues 1–331) was carried out as described [14] and the samestrategy was used for the three other Ser/Thr kinases used in this work.The cosmid MTCY10H4, including the genes pknD, pknE, and pknF,was used as PCR template in subcloning experiments. PCR fragmentsencoding the putative cytoplasmic regions of PknD (residues 1–378),PknE (residues 1–336), and PknF (residues 1–306) were amplified usingforward primers including a NdeI site and reverse primers containing astop codon and a HindIII site (Table 1). The digested and purifiedPCR products were ligated into the pET-28a expression vector usingthe engineered NdeI and HindIII sites. All constructs were verified byDNA sequencing.
In order to express the catalytic domains only of the above kinases,a stop codon was introduced in each construct by site-directed muta-genesis using the Quikchange Site-Directed Mutagenesis Kit (Strata-gene), according to the manufacturer�s instructions. Stop codons wereintroduced in the coding sequences in positions corresponding to thefollowing amino acid residues: 280 (PknB), 279 (PknD), 278 (PknE),and 281 (PknF). The construct expressing the single PknB mutantLys40Ala was also obtained by site-directed mutagenesis from theplasmid with the wild-type pknB gene. The sequences of all themutagenic primers are listed in Table 1.
Protein expression and purification. Protein expression and purifi-cation was performed as described before for PknB [14]. Briefly,Escherichia coli BL21(DE3) strain transformed with the appropriateplasmid was grown at 37 �C until late log phase in Luria–Bertani (LB)medium with 50 lg/ml kanamycin. Induction of expression was con-ducted for 12–16 h at 15 �C, after addition of 1 mM isopropyl-b-thiogalactopyranoside (IPTG). Bacterial pellet was resuspended in50 mM Hepes buffer, pH 7.0, containing 0.2 M NaCl, in the presenceof protease inhibitors, and sonicated. The recombinant proteins werepurified from the supernatant by immobilized Ni-affinity chromatog-raphy followed by a gel filtration step (Hi-Trap Sepharose and Su-perdex 75 columns, Amersham Biosciences). Proteins weresubsequently concentrated and purity of the samples was checked by
Primer sequence (50 fi 3 0)a
FOR: AATTCGCATATGAGCGATGCCGTTCREV: CGACGGAAGCTTACTTCCGTTTGTTFOR: AGTCGGCATATGGATGGCACCGCREV: AAATTAAAGCTTAGGGCTGGCGGGCFOR: GATGTTCATATGCCGCTCGCGGAAREV: AAACGAAAGCTTAGGTCCGCAGCAG
Primer sequence (50 fi 3 0)b
FOR: CGCGACGTTGCGGTCGCGGTGCTGCGCGCTGATCREV: GATCAGCGCGCAGCACCGCGACCGCAACGTCGCGFOR: CGTGCACAACGGTTAACCGCCCGAGGCGCREV: GCGCCTCGGGCGGTTAACCGTTGTGCACGFOR: GCACTCACCACATAAGAGCAACACCAGGCCACGREV: CGTGGCCTGGTGTTGCTCTTATGTGGTGAGTGCFOR: GCCCTGGCCACCTAAGATCAGGATCGTGCCACCREV: GGTGGCACGATCCTGATCTTAGGTGGCCAGGGCFOR: CGTCTGGGCGGCTAAGGTGATCTGACGACACGREV: CGTGTCGTCAGGATCACCTTAGCCGCCCAGACG
d HindIII for reverse primers, respectively).

Fig. 1. Myelin-basic protein (MBP) as a substrate of mycobacterialkinases. (A) Autoradiographies showing that the intracellular regionsof the four kinases studied in this work (PknB, PknD, PknE, andPknF) are able to phosphorylate MBP. The kinase assays were carriedout as described in Materials and methods. (B) Mass spectra of a MBPcontrol tryptic digest before and after kinase incubation in the presenceof ATP and Mn2+. Arrows indicate the tryptic peptide 30–41 fromnative MBP and its monophosphorylated species. Thr 33 was identifiedas the phosphorylated residue by PSD sequencing.
860 R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867
SDS–PAGE. Except for the constructions PknE1–277 and PknD1–278, all other proteins were produced in soluble and active form.
Protein kinase assays. Measurements of protein kinase activity werecarried out in 50 mM Hepes buffer, pH 7.0, containing 1 mM DTT,2 mM MnCl2, 0.01% Brij35, 5% glycerol, 50 lM ATP, and 0.05 lCi/llof [c-33P]ATP (3000 Ci/mmol; Amersham Biosciences). Myelin-basicprotein (MBP) was used as substrate at a concentration of 10 lM, withkinase–substrate ratios of 1:10 or 1:20, for variable times at 30 �C. Thereaction was stopped by adding SDS–PAGE loading buffer supple-mented with 5 mM EDTA (final concentration) and incubating thesamples at 95 �C for 5 min. The reaction mixtures were resolved by12% SDS–PAGE; gels were Coomassie-stained, dried, and analyzedwith a Storm860 PhosphorImager (Molecular Dynamics). MBPphosphorylation was also followed measuring changes in molecularmass after incubation with different kinases. In some experiments,increased sensitivity was achieved by monitoring phosphorylation ofMBP at peptide 30–41 by MS measurements after tryptic digestion.
Protein dephosphorylation. Dephosphorylation assays were carriedout by incubation of recombinant protein kinases (5–20 lM) withpurified PstP (phosphatase–substrate ratios 1:5 to 1:10) in 50 mMHepes buffer, pH 7.0, containing 1 mM DTT, 2 mM MnCl2 for 1 h at35 �C. In some experiments, alkaline phosphatase from calf intestine(Roche Diagnostics) was used for enzymatic dephosphorylation, fol-lowing instructions supplied by the manufacturer.
Sample preparation for mass spectrometry. Proteolytic digestionswere carried out by incubating recombinant proteins with trypsin(Promega) in 50 mM ammonium bicarbonate, pH 8.3, overnight at35 �C (enzyme–substrate ratios 1:10 to 1:50). HPLC separations wereperformed on a reverse-phase column (Vydac C18, 150 · 2.1 mm)equilibrated with 0.1% trifluoroacetic acid and eluted with a lineargradient of 0.07% trifluoroacetic acid in acetonitrile from 0% to 50% in100 min. Eluate was monitored at 220 nm and fractions were manuallycollected for mass spectrometry analysis. The b-elimination reactionsat phosphoresidues were performed by treatment of 0.2–1 lg of pep-tide with saturated solution of Ba(OH)2 at room temperature for 12 has previously reported [21]. Samples were then acidified with acetic acidand desalted using a home-made micro-column filled with POROS-R250 lm chromatography media (Applied Biosystem) prior to MSanalysis.
Carboxylates were modified following published procedures [22]. Inbrief, digestion mixtures (obtained from approximately 2 lg of pro-tein) were lyophilized, resuspended in 50 ll of 2 M methanolic HCl,and incubated at room temperature for 2 h. Solvent was removed bylyophilization. The sample was resuspended in 0.01% acetic acid andphosphopeptides were isolated from peptide mixtures using IMACSepharose gel with chelated Fe3+. After equilibration and extensivewashing with 0.01% acetic acid, retained peptides were eluted from thegel containing 50 mM Na2HPO4, pH 9, and desalted with a POROS-R2 micro-column as described above prior to MS analysis. Alterna-tively, elution was performed with 0.2% trifluoroacetic acid in aceto-nitrile–H2O (50%, v/v).
Mass spectrometry. Mass spectrometry measurements were carriedout in a Voyager DE-PROMALDI-TOF system (Applied Biosystems)equipped with a N2 laser source (337 nm). Molecular mass of re-combinant Ser/Thr protein kinases was determined before and aftertreatment with PstP using sinapinic acid as matrix (10 mg/ml in ace-tonitrile–H2O 50%, 0.2% trifluoroacetic acid) and enolase from Baker�syeast as external standard. Protein samples (1–3 lg) were desalted byincubation in batch with POROS-R2 media and followed by extensivewashing with 0.1% trifluoroacetic acid. POROS beads were mixed withmatrix solution and directly applied onto the MALDI-TOF sampleplate.
Mass spectra of digestion mixtures or isolated peptides were ac-quired on linear and reflector modes using a matrix solution of a-cy-ano-4-hydroxycinnamic acid in 0.2% trifluoroacetic acid inacetonitrile–H2O (50%, v/v) and were externally calibrated using amixture of peptide standards (Applied Biosystem). Phosphopeptides
were analyzed by post-source decay (PSD) MS, in order to determinethe number of phosphate groups and the identity of phosphorylatedresidues.
Results and discussion
Recombinant protein kinases are phosphorylated active
enzymes
The whole intracellular regions of four membrane-bound Ser/Thr protein kinases from M. tuberculosis,namely PknB, PknD, PknE, and PknF, were producedas recombinant proteins in E. coli. All the enzymes showautophosphorylation activity and can use MBP as a sur-rogate substrate (Fig. 1A). Each kinase can phosphory-late MBP in multiple sites, as mass spectrometryanalysis revealed the incorporation of five to nine

R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867 861
phosphates depending on the protein kinase used. Aparticular site (Thr 33) was found to be systematicallyand strongly phosphorylated by all enzymes (Fig. 1B),and thus its detection by mass spectrometry was usedas a sensitive marker to confirm kinase activity.
The overall phosphorylation state of the recombinantprotein kinases was tested by MALDI-TOF mass mea-surements of whole molecules before and after incuba-tion with the protein phosphatase PstP. This approachconfirmed the phosphorylated state of all the enzymesunder investigation. The most abundant molecular spe-cies for each protein kinase contains five (PknB), nine(PknF) or 11 (PknD, PknE) phosphorylated residues(data not shown). The phosphorylation pattern wasmore homogeneous for PknB, as three amino acid posi-tions appeared fully phosphorylated in all samples stud-ied, corresponding to Thr171 and Thr173 in theactivation loop (as previously shown [14]) and Thr294in the juxtamembrane region. Traces of a third modifiedresidue in the activation loop could only be detected insome preparations upon chemical treatment withBa(OH)2 and PSD sequencing (see below). In contrast,phosphate incorporation was more heterogeneous forthe other three enzymes, since multiple species with dif-ferent degrees of phosphorylation were clearly present inall studied samples. Thus, the phosphorylation patternsof PknD, PknE, and PknF appear to be more complexthan in the case of PknB, regarding both the degreeand heterogeneity of the modification.
Kinase phosphorylation results from autocatalytic activity
The observed phosphorylation pattern in the four re-combinant kinases can be assumed to be the conse-quence of an autocatalytic activity, since all proteinswere expressed in E. coli, where no endogenous Ser/Thr kinase activity has been reported. To confirm thishypothesis, we produced an inactive point mutant ofthe catalytic domain of PknB (which contains two phos-phorylation sites in its activation loop) by mutating theessential catalytic residue Lys40 to Ala. As expected, thispoint mutant had no kinase activity on MBP and MSstudies of the recombinant protein expressed in E. coli
revealed that its activation loop was completely unphos-phorylated (data not shown), demonstrating that thephosphorylation pattern observed in the wild-type pro-teins is the result of an autocatalytic process.
Identification of phosphorylation sites
Our first approach to localize phosphorylation sitesincluded MS measurements, in linear and reflectormodes, of tryptic digests before and after treatment withphosphatases (mycobacterial PstP or alkaline phospha-tase). Results obtained from comparing spectra takenwith both digests have a predictive value for the pres-
ence of pThr- and pSer-containing peptides, indicatedby mass values shifting 80 Da or multiples thereof. Arepresentative experiment is shown for PknD in Fig.2A, where three different phosphopeptides could beidentified. However, a full coverage of phosphorylationsites is quite difficult to achieve in this way, due toanalytical problems associated with the detection ofphosphopeptides. Mass spectrometric signals of phos-phopeptides present in complex mixtures are usuallylow due to poor desorption/ionization properties ofnegatively charged species, the tendency to undergofragmentation, and ion suppression effects induced bythe presence of dominant unphosphorylated peptides.To circumvent these problems, a set of strategies was ap-plied in order to attain the broadest coverage of phos-phorylation sites for a given molecule, includingphosphopeptide enrichment steps such as affinity chro-matography or reversed-phase HPLC. For instance,after HPLC separation of tryptic PknD digest, phospho-peptide coverage can be significantly improved (Figs. 2Band C), allowing the identification of 13 phosphorylatedpeptides compared to only three in the non-fractionatedtryptic mixture (Fig. 2A). Improvement in phosphositecoverage for the three other kinases was attained aftersimilar phosphopeptide enrichment steps.
Affinity methods for phosphopeptide isolation onimmobilized Fe3+ ions bound to a chelating gel werefound particularly useful for large multiphosphorylatedspecies previously derivatized by methylation of carbox-ylate groups [22,23], as shown in Fig. 3. Such chemicalmodification increased IMAC selectivity for phospho-peptides while avoiding non-specific binding of negative-ly charged peptides. In particular, the diphosphorylatedsequence 345–377 from PknD (Table 2) could only bedetected in this way.
The phosphorylation state and the number of phos-phate groups in each phosphopeptide was further con-firmed by PSD MS experiments (Fig. 4). Generationof mass differences of 80 Da (loss of HPO3) or 98 Da(loss of H3PO4), or combinations thereof in multiplephosphorylated species (as shown in Fig. 4A), arestrongly indicative of phosphorylation at serine or thre-onine residue(s) [24]. Further fragmentation andsequencing by PSD MS (Fig. 4B) was performed toidentify the phosphorylation sites, which are reportedin Table 2. Even if PSD MALDI-TOF MS is not gener-ally considered the most appropriate MS-based tech-nique to get sequence data from phosphopeptides, theabove results point to its usefulness for a rapid identifi-cation of multiple phosphorylated peptides in complexmixtures. In addition, it is generally reliable for anunequivocal localization of the modified amino acids.
In some cases, the fragmentation properties of phos-phopeptides still preclude the detection of sequence-spe-cific ion signals, thus rendering the localization ofphosphorylated sites difficult. When that was found to
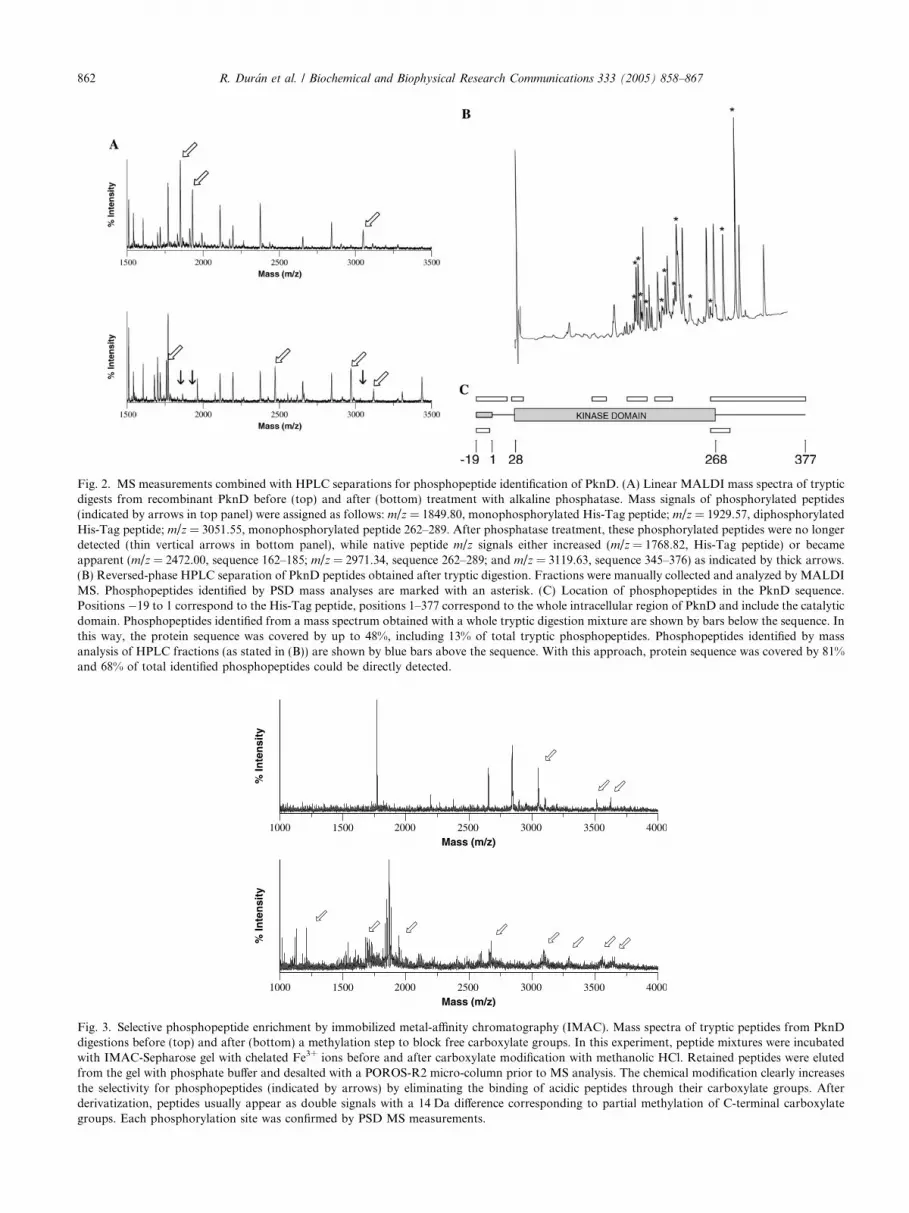
Fig. 2. MS measurements combined with HPLC separations for phosphopeptide identification of PknD. (A) Linear MALDI mass spectra of trypticdigests from recombinant PknD before (top) and after (bottom) treatment with alkaline phosphatase. Mass signals of phosphorylated peptides(indicated by arrows in top panel) were assigned as follows: m/z = 1849.80, monophosphorylated His-Tag peptide; m/z = 1929.57, diphosphorylatedHis-Tag peptide; m/z = 3051.55, monophosphorylated peptide 262–289. After phosphatase treatment, these phosphorylated peptides were no longerdetected (thin vertical arrows in bottom panel), while native peptide m/z signals either increased (m/z = 1768.82, His-Tag peptide) or becameapparent (m/z = 2472.00, sequence 162–185; m/z = 2971.34, sequence 262–289; and m/z = 3119.63, sequence 345–376) as indicated by thick arrows.(B) Reversed-phase HPLC separation of PknD peptides obtained after tryptic digestion. Fractions were manually collected and analyzed by MALDIMS. Phosphopeptides identified by PSD mass analyses are marked with an asterisk. (C) Location of phosphopeptides in the PknD sequence.Positions �19 to 1 correspond to the His-Tag peptide, positions 1–377 correspond to the whole intracellular region of PknD and include the catalyticdomain. Phosphopeptides identified from a mass spectrum obtained with a whole tryptic digestion mixture are shown by bars below the sequence. Inthis way, the protein sequence was covered by up to 48%, including 13% of total tryptic phosphopeptides. Phosphopeptides identified by massanalysis of HPLC fractions (as stated in (B)) are shown by blue bars above the sequence. With this approach, protein sequence was covered by 81%and 68% of total identified phosphopeptides could be directly detected.
Fig. 3. Selective phosphopeptide enrichment by immobilized metal-affinity chromatography (IMAC). Mass spectra of tryptic peptides from PknDdigestions before (top) and after (bottom) a methylation step to block free carboxylate groups. In this experiment, peptide mixtures were incubatedwith IMAC-Sepharose gel with chelated Fe3+ ions before and after carboxylate modification with methanolic HCl. Retained peptides were elutedfrom the gel with phosphate buffer and desalted with a POROS-R2 micro-column prior to MS analysis. The chemical modification clearly increasesthe selectivity for phosphopeptides (indicated by arrows) by eliminating the binding of acidic peptides through their carboxylate groups. Afterderivatization, peptides usually appear as double signals with a 14 Da difference corresponding to partial methylation of C-terminal carboxylategroups. Each phosphorylation site was confirmed by PSD MS measurements.
862 R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867

Table 2Phosphorylation status of recombinant protein kinases
Phosphorylated tryptic peptide sequence -P04 groupsa Phosphorylated residueb
PknB
His-Tag GSSHHHHHHSSGLVPR 0,1,2162–189 AIADSGNSVTQTAAVIGTAQYLSPEQAR 2,3c S166, T171, T173287–308 VLTDAERTSLLSSAAGNLSGPR 1,2 T294287–314 VLTDAERTSLLSSAAGNLSGPRTDPLPR 2d T294, T309
PknD
His-Tag GSSHHHHHHSSGLVPR 0,1,2,3(�3)–19e SHMSDAVPQVGSQFGPYQLLR 0,124–36 GGMGEVYEAEDTR 0,1121–137 QIAAALDAAHANGVTHR 0,1 T135162–185 AASDPGLTQTGTAVGTYNYMAPER 2,3 T169, T171, T173186–194 FTGDEVTYR 0,1195–215 ADIYALACVLGECLTGAPPYR 0,1 T209262–289 FMSAGDLAIAAHDALTTSEQHQATTILR 1290–326 RGDNATLLATPADTGLSQSESGIAGAGTGPPTPGAAR 0,1,2327–344 WSPGDSATVAGPLAADSR 0,1,2345–377 GGNWPSQTGHSPAVPNALQASLGHAVPPAGNKR 2
PknE
His-Tag GSSHHHHHHSSGLVPR 0,1,2,3(�3)–17e GSHMDGTAESREGTQFGPYR 2d T11, S79–17 EGTQFGPYR 0, 1 T1146–58 LMSETLSSDPVFR 0,1 T5046–60 LMSETLSSDPVFRTR 0,2 T50, T59169–186 LTQLGNTVGTLYYMAPER 1,2,3 T170, T175, T178f
187–195 FSESHATYR 0,1261–288 YVTCGDLSAAAHAALATADQDRATDILR 0,1295–331 LPVPSTHPVSPGTRWPQPTPWAGGAPPWGPPSSPLPR 1,2,3,4309–331 WPQPTPWAGGAPPWGPPSSPLPR 1,2
PknF
His-Tag GSSHHHHHHSSGLVPR 1,2,3(�3)–16e GSHMPLAEGSTFAGFTIVR 0,1,2 T8, T13155–198 IMLADFGIAGWVDDPSGLTATNMTVGTVSYAAPEQLMGNELDGR 2 T173, T175199–241 ADQYALAATAFHLLTGSPPFQHANPAVVISQHLSASPPAIGDR 1,2278–300 LGGAGDPDDTRVSQPVAVAAPAK 1, 2 T287, S290
a Each peptide sequence was usually found with different degree of phosphorylation.b These residues were identified by PSD sequencing.c Sequence 162–189 appears mostly diphosphorylated at T171 and T173. Trace amounts of a triphosphorylated peptide includes pS166, identified
by PSD sequencing after treatment with Ba(OH)2 as stated in the text.d In this case, a phosphorylated residue adjacent to R or K precludes tryptic cleavage, then this peptide is only detected when phosphorylated.e This peptide includes three amino acids from the His-Tag at the N terminus.f Phosphorylated residues in mono- and diphosphorylated peptides were not identified.
R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867 863
be the case, a pre-treatment of the phosphorylated pep-tides with Ba(OH)2 was included to generate the corre-sponding dehydroamino acid from pThr and pSerresidues by b-elimination of H3PO4, thus rendering amore appropriate derivative for MS studies [21]. As anexample, using the above approach S166 was identifiedto be a minor, third phosphorylated residue in the acti-vation loop of PknB (Table 2).
Data summarizing phosphosite mapping of the intra-cellular regions from the four mycobacterial Ser/Thrprotein kinases are presented in Fig. 5 and Table 2. To-tal sequence coverage using the different approachesoutlined above was 90% for PknB, 86% for PknD,82% for PknE, and 84% for PknF. This extensivesequence coverage left aside only a small number ofundetected peptides containing Ser or Thr residues
whose phosphorylation status could not be determined:PknB (Ser129), PknD (Thr98, Ser99, Ser218, andSer235), PknE (Thr67, Ser148, Ser162, Thr164,Thr165, and Ser332), and PknF (Ser302 and Thr306).Since specific methods were used to enrich the peptidemixtures in phosphorylated species, it is unlikely thatundetected peptides included pThr or pSer residues.
The four protein kinases are substrates of the cognate
mycobacterial phosphatase PstP
Since M. tuberculosis contains only one gene, pstP,coding for a Ser/Thr protein phosphatase, we wonderedwhether PstP was active on the different kinases, asmight be expected if the kinase activity is dependenton (or regulated by) reversible phosphorylation
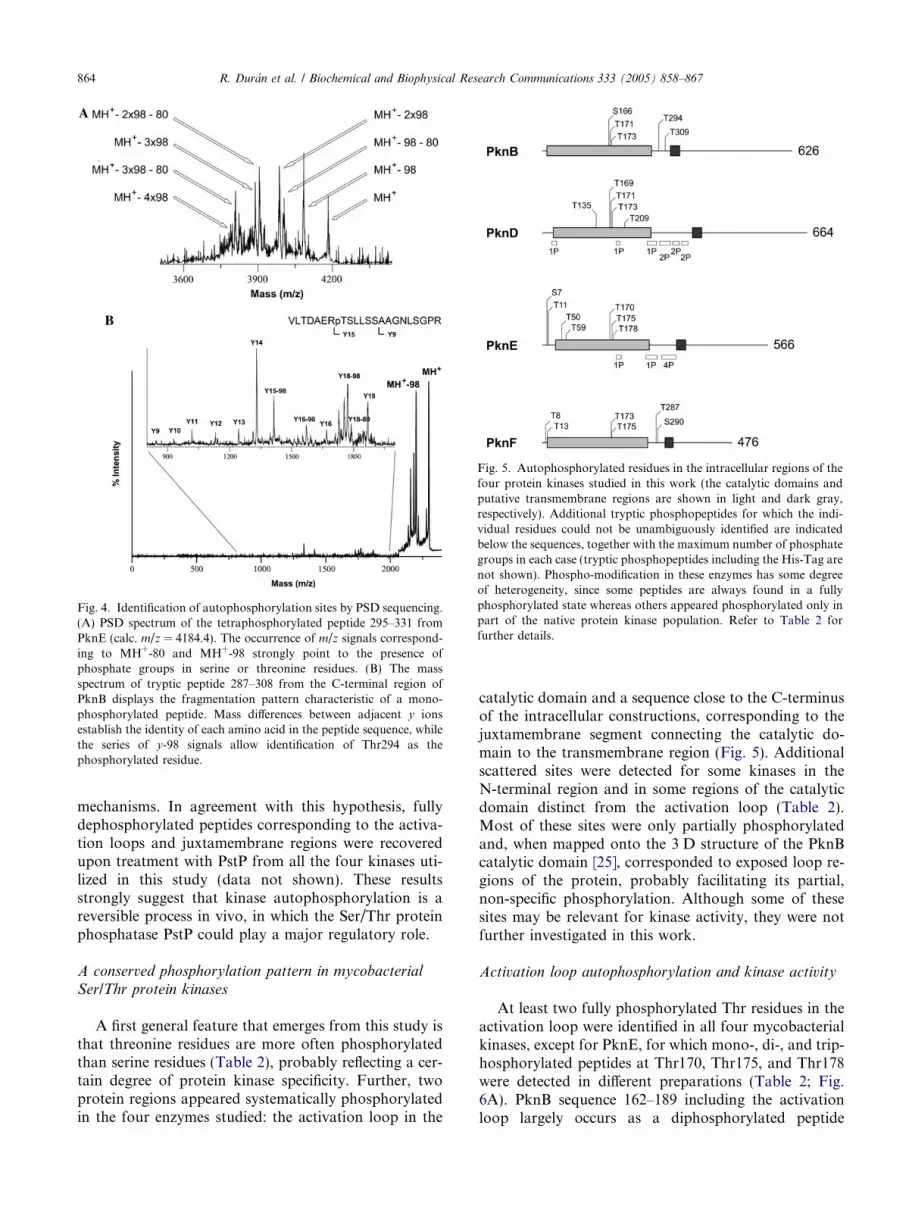
Fig. 4. Identification of autophosphorylation sites by PSD sequencing.(A) PSD spectrum of the tetraphosphorylated peptide 295–331 fromPknE (calc. m/z = 4184.4). The occurrence of m/z signals correspond-ing to MH+-80 and MH+-98 strongly point to the presence ofphosphate groups in serine or threonine residues. (B) The massspectrum of tryptic peptide 287–308 from the C-terminal region ofPknB displays the fragmentation pattern characteristic of a mono-phosphorylated peptide. Mass differences between adjacent y ionsestablish the identity of each amino acid in the peptide sequence, whilethe series of y-98 signals allow identification of Thr294 as thephosphorylated residue.
Fig. 5. Autophosphorylated residues in the intracellular regions of thefour protein kinases studied in this work (the catalytic domains andputative transmembrane regions are shown in light and dark gray,respectively). Additional tryptic phosphopeptides for which the indi-vidual residues could not be unambiguously identified are indicatedbelow the sequences, together with the maximum number of phosphategroups in each case (tryptic phosphopeptides including the His-Tag arenot shown). Phospho-modification in these enzymes has some degreeof heterogeneity, since some peptides are always found in a fullyphosphorylated state whereas others appeared phosphorylated only inpart of the native protein kinase population. Refer to Table 2 forfurther details.
864 R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867
mechanisms. In agreement with this hypothesis, fullydephosphorylated peptides corresponding to the activa-tion loops and juxtamembrane regions were recoveredupon treatment with PstP from all the four kinases uti-lized in this study (data not shown). These resultsstrongly suggest that kinase autophosphorylation is areversible process in vivo, in which the Ser/Thr proteinphosphatase PstP could play a major regulatory role.
A conserved phosphorylation pattern in mycobacterial
Ser/Thr protein kinases
A first general feature that emerges from this study isthat threonine residues are more often phosphorylatedthan serine residues (Table 2), probably reflecting a cer-tain degree of protein kinase specificity. Further, twoprotein regions appeared systematically phosphorylatedin the four enzymes studied: the activation loop in the
catalytic domain and a sequence close to the C-terminusof the intracellular constructions, corresponding to thejuxtamembrane segment connecting the catalytic do-main to the transmembrane region (Fig. 5). Additionalscattered sites were detected for some kinases in theN-terminal region and in some regions of the catalyticdomain distinct from the activation loop (Table 2).Most of these sites were only partially phosphorylatedand, when mapped onto the 3 D structure of the PknBcatalytic domain [25], corresponded to exposed loop re-gions of the protein, probably facilitating its partial,non-specific phosphorylation. Although some of thesesites may be relevant for kinase activity, they were notfurther investigated in this work.
Activation loop autophosphorylation and kinase activity
At least two fully phosphorylated Thr residues in theactivation loop were identified in all four mycobacterialkinases, except for PknE, for which mono-, di-, and trip-hosphorylated peptides at Thr170, Thr175, and Thr178were detected in different preparations (Table 2; Fig.6A). PknB sequence 162–189 including the activationloop largely occurs as a diphosphorylated peptide

Fig. 6. Phosphorylation of kinase activation loops. (A) Amino acidsequences of the activation loops of M. tuberculosis STPKs. Thephosphorylated residues found in the four kinases studied areindicated, as well as the conservation of these residues in the otherenzymes. (B) Protein kinase activity of PknB (wild-type, PknBT171A,PknBT173A, and PknBT171A,T173A). The enzymes, expressed and puri-fied from E. coli, were incubated with MBP in vitro as described inMaterials and methods. The MBP phosphorylation levels of themutants are given as relative values with respect to those obtained forwild-type PknB, which was defined as 100%.
R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867 865
(Thr171 and Thr173). For PknD, residues Thr169 andThr173 are always phosphorylated, while Thr171 is onlypresent in a minor triphosphorylated peptide fraction.The corresponding sequence for PknF was always pres-ent as a diphosphorylated peptide at Thr173 and Thr175(Fig. 6A). These phosphorylation patterns were not sig-nificantly affected after incubation of the purifiedproteins with ATP, indicating that the kinase autophos-phorylation reactions mostly occur during proteinexpression in E. coli.
To investigate whether phosphorylation of the activa-tion loop affects kinase activity, we selected PknB (whichhas the more homogeneous pattern of phosphorylationamong the four kinases studied here) and substitutedthe two pThr residues (Thr171 and Thr173) by alanine[14]. As shown in Fig. 6B, each single mutant had 10-to 20-fold less activity on MBP than the wild-type en-zyme, whereas the double mutant had near 300-fold lessactivity. These results confirm that phosphorylation ofthe activation loop has a strong effect on PknB kinaseactivity. Similar results have been observed for PrkCfrom Bacillus subtilis [26], the only other bacterial STPK
whose autophosphorylation pattern has been studied indetail, suggesting a conserved activation loop mecha-nism to regulate kinase activity.
In eukaryotic kinases, the activation loop is known toundergo large conformational changes associated withenzyme activation [20]. In particular, the phosphategroup from one of the central loop residues usuallyforms an ion pair with a conserved arginine residue,Arg165 in the eukaryotic cAMP-dependent protein ki-nase [27] and Arg137 in mycobacterial PknB [15,25].This conserved arginine is situated in the catalytic loopof the kinase domain, adjacent to a critical aspartic acidresidue, and its interaction with pThr (pSer) residuesfrom the activation loop is thought to be important toachieve the proper conformation of the active site forcatalysis. Indeed, we have shown that this is the casefor PknB, where the mutation of Thr171 and Thr173resulted in a significantly decreased activity towardsMBP (Fig. 6B) [14]. Similar results were obtained forB. subtilis PrkC, where the replacement of four auto-phosphorylated Thr residues in the activation loop withAla substantially reduced the enzymatic activity withadditive effect [26], lending strong support to a regulato-ry role of the activation loop in these receptor-like bac-terial kinases. This may also be the case for the otherthree analyzed mycobacterial kinases, given the presenceof conserved pThr in the activation loop (Fig. 6A) andthe conservation of the RD motif too (data not shown).Moreover, recent results from our laboratory indicate adirect involvement of Thr171 and Thr173 of PknB notonly in the regulation of kinase activity, but also in aprocess of activation loop-mediated recruitment of itssubstrate [28].
Phosphorylation sites in the juxtamembrane segments
The second region displaying a systematic pattern ofautophosphorylation in all four kinases is the juxta-membrane segment (Fig. 5). Two phosphoresidues couldbe unambiguously identified for PknB (Thr294, Thr309)and PknF (Thr287, Ser290), while the covalent modifi-cations in PknD and PknE appear more complex, withup to six sites diversely modified (Table 2). This com-plexity might be due, at least in part, to a higher acces-sibility of the juxtamembrane region in the recombinant(soluble) protein as compared to the membrane-boundenzyme in vivo.
To investigate whether phosphorylated residues inthis region could affect kinase activity, we constructeddeletion mutants of the four protein kinases that includ-ed only the catalytic domain, without the juxtamem-brane region. Soluble and active proteins wereobtained for PknB and PknF. In both the cases, theautophosphorylation and the activity against MBP werelittle or not affected (Fig. 7), strongly suggesting thatjuxtamembrane phosphorylation is not required for

Fig. 7. Autoradiography showing the kinase activity of the wholeintracellular regions (left) and the catalytic domains (right) of PknBand PknF on the surrogate substrate MBP.
866 R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867
the catalytic activity, as also observed for B. subtilis
PrkC [26]. However, the consistent pattern of modifica-tions observed in this region argues for a role of juxta-membrane phosphorylation in activity regulation orprotein stability (Fig. 5). A possible mechanism mightresemble those reported for eukaryotic receptors suchas the type I TGF-b Ser/Thr kinase [29] or the ephrinTyr kinase EphB2 [30], in which phosphorylation ofthe juxtamembrane region modulates its interactionwith the catalytic domain. Alternatively, juxtamem-brane phosphorylation may be important for therecruitment of other proteins involved in a putative sig-naling cascade and/or coupling to other signaling path-ways, as it has been suggested for B. subtilis PrkC [26].Future work will unravel the gating signals that switchthe autophosphorylation activity in vivo, and will clarifythe role of these transmembrane protein kinases inmycobacterial signaling.
Acknowledgments
This work was supported by grants from the Europe-an Union (X-TB, Contract No. QLK2-CT-2001-02018),Institut Pasteur (GPH-5), Ministere de la Recherche,France (Contract No. 01-B-0095), PEDECIBA andDINACYT PDT Project No. 160/29 (Montevideo,Uruguay).
References
[1] T. Hunter, Signaling—2000 and beyond, Cell 100 (2000) 113–127.[2] C.J. Bakal, J.E. Davies, No longer an exclusive club: eukaryotic
signalling domains in bacteria, Trends Cell Biol. 10 (2000) 32–38.[3] A.M. Stock, V.L. Robinson, P.N. Goudreau, Two-component
signal transduction, Annu. Rev. Biochem. 69 (2000) 183–215.[4] S. Hakansson, E.E. Galyov, R. Rosqvist, H. Wolf-Watz, The
Yersinia YpkA Ser/Thr kinase is translocated and subsequently
targeted to the inner surface of the HeLa cell plasma membrane,Mol. Microbiol. 20 (1996) 593–603.
[5] J. Wang, C. Li, H. Yang, A. Mushegian, S. Jin, A novel serine/threonine protein kinase homologue of Pseudomonas aeruginosa isspecifically inducible within the host infection site and is requiredfor full virulence in neutropenic mice, J. Bacteriol. 180 (1998)6764–6768.
[6] E. Madec, A. Laszkiewicz, A. Iwanicki, M. Obuchowski, S.Seror, Characterization of a membrane-linked Ser/Thr proteinkinase in Bacillus subtilis, implicated in developmental processes,Mol. Microbiol. 46 (2002) 571–586.
[7] K. Petrıckova, M. Petricek, Eukaryotic-type protein kinases inStreptomyces coelicolor: variations on a common theme, Micro-biology 149 (2003) 1609–1621.
[8] A. Walburger, A. Koul, G. Ferrari, L. Nguyen, C. Prescianotto-Baschong, K. Huygen, B. Klebl, C. Thompson, G. Bacher, J.Pieters, Protein kinase G from pathogenic mycobacteria promotessurvival within macrophages, Science 304 (2004) 1800–1804.
[9] S.T. Cole, R. Brosch, J. Parkhill, T. Garnier, et al., Decipheringthe biology of Mycobacterium tuberculosis from the completegenome sequence, Nature 393 (1998) 537–544.
[10] P. Peirs, L. De Wit, M. Braibant, K. Huygen, J. Content, Aserine/threonine protein.kinase from Mycobacterium tuberculosis,Eur. J. Biochem. 244 (1997) 604–612.
[11] Y. Av-Gay, S. Jamil, S.J. Drews, Expression and characterizationof the Mycobacterium tuberculosis serine/threonine protein kinasePknB, Infect. Immun. 67 (1999) 5676–5682.
[12] A. Koul, A. Choidas, A.K. Tyagi, K. Drlica, Y. Singh, A.Ullrich, Serine/threonine protein kinases PknF and PknG of
Mycobacterium tuberculosis: characterization and localization,Microbiology 147 (2001) 2307–2314.
[13] R. Chaba, M. Raje, P.K. Chakraborti, Evidence that a eukary-otic-type serine/threonine protein kinase from Mycobacterium
tuberculosis regulates morphological changes associated with celldivision, Eur. J. Biochem. 269 (2002) 1078–1085.
[14] B. Boitel, M. Ortiz-Lombardıa, R. Duran, F. Pompeo, S.T.Cole, C. Cervenansky, P.M. Alzari, PknB kinase activity isregulated by phosphorylation in two Thr residues and dephos-phorylation by PstP, the cognate phospho-Ser/Thr phosphatase,in Mycobacterium tuberculosis, Mol. Microbiol. 49 (2003) 1493–1508.
[15] T.A. Young, B. Delagoutte, J.A. Endrizzi, A.M. Falick, T. Alber,Structure of Mycobacterium tuberculosis PknB supports auniversal activation mechanism for Ser/Thr protein kinases,Nat. Struct. Biol. 10 (2003) 168–174.
[16] V. Molle, C. Girard-Blanc, L. Kremer, P. Doublet, A.J. Cozzone,J.F. Prost, Protein PknE, a novel transmembrane eukaryotic-likeserine/threonine kinase from Mycobacterium tuberculosis, Bio-chem. Biophys. Res. Commun. 308 (2003) 820–825.
[17] P. Chopra, B. Singh, R. Singh, R. Vohra, A. Koul, L.S. Meena,H. Koduri, M. Ghildiyal, P. Deol, T.K. Das, A.K. Tyagi, Y.Singh, Phosphoprotein phosphatase of Mycobacterium tuberculo-
sis dephosphorylates serine-threonine kinases PknA and PknB,Biochem. Biophys. Res. Commun. 311 (2003) 112–120.
[18] R. Gopalaswamy, P.R. Narayanan, S. Narayanan, Cloning,overexpression, and characterization of a serine/threonine proteinkinase pknI from Mycobacterium tuberculosis H37Rv, ProteinExpr. Purif. 36 (2004) 82–89.
[19] L.N. Johnson, M.E. Noble, D.J. Owen, Active and inactiveprotein kinases: structural basis for regulation, Cell 85 (1996) 149–158.
[20] M. Huse, J. Kuriyan, The conformational plasticity of proteinkinases, Cell 109 (2002) 275–282.
[21] M. Mann, S.-E. Ong, M. Gronborg, H. Steen, O.N. Jensen, A.Pandey, Analysis of protein phosphorylation using mass spec-trometry: deciphering the phosphoproteome, Trends Biotechnol.20 (2002) 261–268.

R. Duran et al. / Biochemical and Biophysical Research Communications 333 (2005) 858–867 867
[22] S.B. Ficarro, M.L. McCleland, P.T. Stukenberg, D.J. Burke,M.M. Ross, J. Shabanowitz, D.F. Hunt, F.M. White, Phos-phoproteome analysis by mass spectrometry and its applicationto Saccharomyces cerevisiae, Nat. Biotechnol. 20 (2002) 301–305.
[23] K.A. Resing, R.S. Johnson, K.A. Walsh, Mass spectrometricanalysis of 21 phosphorylation sites in the internal repeat of ratprofilaggrin, precursor of an intermediate filament associatedprotein, Biochemistry 34 (1995) 9477–9487.
[24] R.S. Annan, S.A. Carr, Phosphopeptide analysis by matrix-assisted laser desorption time-of-flight mass spectrometry, Anal.Chem. 68 (1996) 3413–3421.
[25] M. Ortiz-Lombardıa, F. Pompeo, B. Boitel, P.M. Alzari, Crystalstructure of the catalytic domain of the PknB serine/threoninekinase fromMycobacterium tuberculosis, J. Biol. Chem. 278 (2003)13094–13100.
[26] E. Madec, A. Stensballe, S. Kjellstrom, L. Cladiere, M. Obu-chowski, O.N. Jensen, S.J. Seror, Mass spectrometry and site-directed mutagenesis identify several autophosphorylated residues
required for the activity of PrkC, a Ser/Thr kinase from Bacillus
subtilis, J. Mol. Biol. 330 (2003) 459–472.[27] D.R. Knighton, J.H. Zheng, L.F. Ten Eyck, V.A. Ashford, N.H.
Xuong, S.S. Taylor, J.M. Sowadski, Crystal structure of thecatalytic subunit of cyclic adenosine monophosphate-dependentprotein kinase, Science 253 (1991) 407–414.
[28] A. Villarino, R. Duran, A. Wehenkel, P. Fernandez, P. England,P. Brodin, S.T. Cole, U. Zimny-Arnat, P.R. Jungblut, C.Cervenansky, P.M. Alzari, Proteomic identification of M. tuber-
culosis protein kinase substrates: PknB recruits GarA, a FHA-containing protein, through activation loop-mediated interac-tions, J. Mol. Biol. (2005) in press.
[29] M. Huse, Y.-G. Chen, J. Massague, J. Kuriyan, Crystal structureof the cytoplasmic domain of the type I TGF beta receptor incomplex with FKBP12, Cell 96 (1999) 425–436.
[30] L.E Wybenga-Groot, B. Baskin, S.H. Ong, J. Tong, T. Pawson,F. Sicheri, Structural basis for autoinhibition of the Ephb2receptor tyrosine kinase by the unphosphorylated juxtamembraneregion, Cell 106 (2001) 745–757.




























