Embed Size (px)
Citation preview

CpG oligodeoxynucleotide-enhanced humoral immuneresponse and production of antibodies to prionprotein PrPSc in mice immunized with 139Ascrapie-associated fibrils
Daryl S. Spinner,*,1 Regina B. Kascsak,* Giuseppe LaFauci,* Harry C. Meeker,* Xuemin Ye,*,2
Michael J. Flory,* Jae Il Kim,*,3 Georgia B. Schuller-Levis,* William R. Levis,†
Thomas Wisniewski,† Richard I. Carp,* and Richard J. Kascsak**New York State Institute for Basic Research in Developmental Disabilities, Staten Island, New York, USA;and †New York University School of Medicine, New York, New York, USA
Abstract: Prion diseases are characterized byconversion of the cellular prion protein (PrP) to aprotease-resistant conformer, the srapie form ofPrP (PrPSc). Humoral immune responses to non-denatured forms of PrPSc have never been fullycharacterized. We investigated whether produc-tion of antibodies to PrPSc could occur in PrP null(Prnp�/�) mice and further, whether innate im-mune stimulation with the TLR9 agonist CpG oli-godeoxynucleotide (ODN) 1826 could enhancethis process. Whether such stimulation could raiseanti-PrPSc antibody levels in wild-type (Prnp�/�)mice was also investigated. Prnp�/� and Prnp�/�
mice were immunized with nondenatured 139Ascrapie-associated fibrils (SAF), with or withoutODN 1826, and were tested for titers of PrP-specific antibodies. In Prnp�/� mice, inclusion ofODN 1826 in the immunization regime increasedanti-PrP titers more than 13-fold after two immu-nizations and induced, among others, antibodies,which were only present in the immune repertoireof mice receiving ODN 1826, to an N-terminalepitope. mAb 6D11, derived from such a mouse,reacted with the N-terminal epitope QWNK in na-tive and denatured forms of PrPSc and recombi-nant PrP and exhibited a Kd in the 10�11 M range.In Prnp�/� mice, ODN 1826 increased anti-PrPlevels as much as 84% after a single immunization.Thus, ODN 1826 potentiates adaptive immune re-sponses to PrPSc in 139A SAF-immunized mice.These results represent the first characterization ofhumoral immune responses to nondenatured, in-fectious PrPSc and suggest methods for optimizingthe generation of mAb to PrPSc, many of whichcould be used for diagnosis and treatment of priondiseases. J. Leukoc. Biol. 81: 000–000; 2007.
Key Words: transmissible spongiform encephalopathy (TSE)� monoclonal antibodies � innate immunity � Toll-like receptor 9(TLR9 or TLR-9)
INTRODUCTION
The transmissible spongiform encephalopathies (TSEs), orprion diseases, are a group of infectious, invariably fatal,neurodegenerative diseases that affect the CNS. These dis-eases, which include scrapie in sheep and goats, bovine spon-giform encephalopathy in cattle, chronic wasting disease incervids, and Creutzfeldt-Jakob disease in humans, are charac-terized by the development of spongiform degeneration in thebrain and the conversion of the normal cellular prion protein(PrPC) to a pathologic, �-rich conformer, which is designatedthe scrapie form of PrP (PrPSc; reviewed in ref. [1]). As PrPSc
is highly resistant to proteolytic degradation, its clearance fromthe body is slow. Deposition of PrPSc occurs throughout thelymphoreticular tissues and the brain in patterns that dependon the TSE strain, the host genotype, and the route of infection(reviewed in ref. [2]).
The abnormal PrP, PrPSc, is a notoriously poor immunogen.This may partly be a result of its high degree of proteaseresistance, rendering it resistant to proteolytic degradationwithin phagocytic APCs, such as dendritic cells (DCs), mac-rophages, and microglia. As presentation of PrPSc fragments byAPCs to T cells is so impaired, initiation of adaptive immuneresponses is blocked. Although this may be true in PrPC-expressing and nonexpressing hosts, those hosts that expressPrPC are further prevented from developing humoral immunityto PrPSc by immune tolerance [3–7], which to these agents,composed largely or entirely of PrPSc, is avoided in PrP null(Prnp�/�) mice that support the production of anti-PrP anti-bodies [8–10].
1 Correspondence: New York State Institute for Basic Research in Devel-opmental Disabilities, 1050 Forest Hill Rd., Staten Island, NY 10314, USA.E-mail: [email protected]
2 Current address: Department of Ophthalmology, Mount Sinai School ofMedicine, One Gustave L. Levy Place, New York, NY 10029, USA.
3 Current address: Department of Physiology and Biophysics, Case WesternReserve University, 2085 Adelbert Road, Cleveland, OH 44106, USA.
Received October 23, 2006; revised December 8, 2006; accepted February23, 2007.
doi: 10.1189/jlb.1106665
0741-5400/07/0081-0001 © Society for Leukocyte Biology Journal of Leukocyte Biology Volume 81, June 2007 1
Uncorrected Version. Published on March 22, 2007 as DOI:10.1189/jlb.1106665
Copyright 2007 by The Society for Leukocyte Biology.

Stimulation of innate immune signaling through members ofthe TLR family (TLR1�13) potentiates immune activation andthe destruction of foreign pathogens such as bacteria, viruses,and fungi and a variety of cancers [11–14]. The function ofTLRs is to recognize pathogen-associated macromolecules andsubsequently, to activate immune cells on which the TLRsreside [11–14]. TLR ligation also plays a role in adaptiveimmunity to certain pathogen-associated proteins by leading toa strong antibody response [15]. Bacterial DNA, which con-tains unmethylated CpG dinucleotides, or similar nucleic acidsequences, e.g., synthetic CpG oligodeoxynucleotides (ODNs),signal through TLR9 [16, 17]. CpG ODNs have been shown toheighten humoral immunity to proteins such as the hepatitis Bvirus (HBV) surface antigen [18, 19] and Leishmania majorantigens [20]. This effect is attributable to several factors,including enhanced activation and mobilization of immunesystem components such as macrophages, DCs, and B cells[16, 17], as well as reduction of CD4�CD25� regulatory T cell(Treg)-mediated immune suppression [21–24].
In the present study, we investigated whether humoral im-mune responses to nondenatured, infectious murine (m)PrPSc
could be developed in Prnp�/� mice and whether stimulatinginnate immunity through TLR9 signaling increases this re-sponse. These studies were prompted in part by results show-ing effective prevention of disease in scrapie-infected micetreated with the CpG ODN 1826 [25]. Therefore, we alsoinvestigated the effect of TLR9 stimulation on humoral immu-nity to mPrPSc in wild-type (WT; Prnp�/�) mice. To addressthese issues, mice were immunized with nondenatured, infec-tious 139A scrapie-associated fibrils (SAF) in a standard ad-juvant (TiterMax), with or without CpG ODN 1826 (also calledODN 1826). 139A SAF were chosen as the immunogen, asrelative to other mouse-adapted scrapie strains, 139A PrPSc isone of the more protease-sensitive PrPSc conformers [26] and istherefore more amenable to proteolytic processing and antigenpresentation in vivo. Our data indicate that immunizedPrnp�/� and Prnp�/� mice developed measurable titers ofantibodies to 139A SAF, and the levels of humoral immunitywere heightened in the mice treated with ODN 1826. Theseresults represent the first characterization of humoral immuneresponses to a nondenatured, infectious PrPSc immunogen inPrnp�/� or Prnp�/� mice and further indicate how such re-sponses are enhanced by CpG ODNs.
MATERIALS AND METHODS
Antibodies
mAb 3F4 and rabbit polyclonal antibody (pAb) 78295 have been describedpreviously [5, 6, 27]. mAb 4B4 was derived from a BALB/c mouse immunizedwith formic acid-denatured, proteinase K (PK)-treated sheep SAF in TiterMaxadjuvant (CytRx, Norcross, GA, USA); the corresponding hybridoma line wasproduced by standard splenocyte fusion protocols using the P3/X63-Ag8.653myeloma line [American Type Culture Collection (ATCC), Manassas, VA,USA]. mAb 3F4 and 4B4 are available from Signet Laboratories (Dedham, MA,USA). mAb 7A12 [28, 29] was a gift from Man-Sun Sy (Case Western ReserveUniversity, Cleveland, OH, USA).
mAb were purified from concentrated tissue-culture supernatant producedin Integra CL bioreactor flasks (Integra Biosciences, Chur, Switzerland) or fromascites, using Pierce UltraLink Immobilized Protein A/G Plus affinity resin
(Pierce, Rockford, IL, USA). Concentrations of antibodies were determined bymouse IgG radial immunodiffusion assay kits (MP Biomedicals, Irvine, CA,USA). Isotypes of antibodies were determined with ImmunoPure mAb isotyp-ing kits (Pierce). Determination of Kd values for antibody-antigen binding wasperformed by ELISA as described previously [30, 31] with modifications.Briefly, PolySorp immunomodule (Nalge Nunc, Rochester, NY, USA) wellswere coated with varying concentrations of recombinant mPrP (rmPrp; 5, 2.5,1.25, 0.63, 0.31, or 0.16 �g/ml in PBS), and each concentration of PrP wastitrated with varying concentrations of mAb representing a titration series.Apparent Kd (Kdapp) values were obtained from each titration of mAb againsta given PrP concentration. Finally, data were plotted as graphs of Kdapp for agiven PrP concentration (ordinate) versus PrP concentration (abscissa), and theextrapolated y-intercept of the resulting linear regression represented the trueKd value for the mAb-PrP interaction. Pepscan epitope analyses were con-ducted by Pepscan Systems (Lelystad, The Netherlands) with overlapping 15-or 30-mer PrP peptides, which were shifted by one residue at a time from theN-terminal to the C-terminal and together, spanned the 1�254 or the 61�240region of mPrP, respectively. Plasma was analyzed at dilutions ranging from125- to 1000-fold. Peaks representing antibody epitopes were defined byextended regions of OD values exceeding twice the background for a givenantibody.
Preparation of proteins
SAF
Isolation and purification of 139A SAF were performed from brains of clini-cally positive, 139A scrapie-infected C57BL/6J or CD-1 mice, according to theprotocol of Hilmert and Diringer [32], with modifications as described previ-ously [6]. The final isolated pellets were resuspended in water (�0.5 ml/25 gstarting brain material final concentration) and dialyzed into water. The SAFpreparations were sonicated prior to analyses or immunizations, as this treat-ment was shown to disaggregate and increase the number of infectious particlespresent [33]. For formic acid denaturation of SAF, preparations were treated for2 h at 4°C with 3 vol 1.0 M formic acid, lyophilized, resuspended in water,relyophilized, and finally resuspended in a volume of PBS equivalent to thevolume of the starting SAF preparation.
r-PrPs
Full-length rmPrP23–230 (recPrP), unmodified with affinity tags, includingResidues 23�230, was prepared in Escherichia coli as described previously[34]. Syrian hamster recPrP was a gift from Ilia V. Baskakov (University ofMaryland, Baltimore, MD, USA) and Robert G. Rohwer and Luisa Gregori(Department of Veterans Affairs Medical Center, Baltimore, MD, USA).
Protein concentrations
Protein concentrations were determined by bicinchoninic protein assay(Pierce).
Mice
Prnp�/� mice, crossed into the FVB/N genetic background, were originallyobtained from Charles Weissmann (Scripps Research Institute, Jupiter, FL,USA) [35].
Transgenic sheep (TgShp), Tg666 [Tg human (TgHu)], TgElk, and Tg muledeer (TgMuD) mice, exclusively expressing sheep (Q171 genotype), human(M129 genotype), elk (eGMSE allele) [36], or mule deer PrPC, respectively,were developed at the New York State Institute for Basic Research in Devel-opmental Disabilities (NYS IBRDD; Staten Island, NY, USA; G. LaFauci,personal communication). All TgPrP mice were homozygous with respect to thetransgene, except the TgMuD mice, which were heterozygous. The TgElk micehave been described elsewhere [37].
All of the strains of mice above were bred and maintained at the NYSIBRDD Animal Facility. Female BALB/cJ and C57BL/6J mice were purchasedfrom the Jackson Laboratory (Bar Harbor, ME, USA). Mice expressing bovinePrPC (BoTg 3204), of which only brains were used, were developed by LarisaCervenakova at the American Red Cross (Rockville, MD, USA) and were notbred or maintained on site.
All protocols involving animals were approved by the NYS IBRDD or theNew York University Institutional Animal Care and Use Committee (NewYork, NY, USA) prior to beginning experiments.
2 Journal of Leukocyte Biology Volume 81, June 2007 http://www.jleukbio.org

ELISA analyses
For ELISA analyses against PrP, PolySorp immunomodule wells were coatedwith �5 �g/ml protein. For determinations of plasma antibody titers, eachplasma sample was run in triplicate at three sequential tenfold dilutions. Thesecondary reagent was an alkaline phosphatase conjugate of goat antimouseIgG � M (Biosource, Camarillo, CA, USA) used at 0.4 �g/ml. Plasma Igisotype titer determination was done similarly to the IgG � M determinationsdescribed above but using ImmunoPure mAb isotyping kits (Pierce), accordingto the manufacturer’s protocol. The ELISA buffer used for blocking and forantibody binding steps, when appropriate, contained 0.2% Tween-20 and 1%normal goat serum (Vector Laboratories, Burlingame, CA, USA) in PBS, andwash buffer contained 0.05% Tween-20 in PBS. p-Nitrophenyl phosphate wasused for ELISA colorimetric development. All ELISA analyses were done onan ELx800 microplate reader from BioTek Instruments (Winooski, VT, USA).In determining endpoint dilution (E.D.), a threshold value for considering ameasurable response was set at 0.200 OD405 units. For determining antibodytiters in plasma, OD405 was plotted against plasma dilution for each dilution inthe series on a log-log transform, and the plasma dilution, at which theresulting regression line crossed OD405 � 0.200, was taken as the E.D., whichfor each mouse plasma sample, was derived by averaging the results of threeseparate assays of each sample. For statistical analyses, two-tailed Student’st-tests were used. Differences between groups of data were considered statis-tically significant for P values �0.05.
SDS-PAGE/Western blotting analyses
Protein samples in sample loading buffer containing SDS and 2-ME wereheated at 100°C for 5 min and loaded to and run on SDS-PAGE gels with 12%polyacrylamide (Bio-Rad, Hercules, CA, USA), blotted onto nitrocellulosemembranes of 0.45 �m pore size (Bio-Rad) using premade buffers (Bio-Rad),blocked with 5% nonfat dry milk in 0.2% Tween-20/TBS, and stained withantibodies in 1% normal goat serum/0.2% Tween-20/TBS, according to stan-dard methods. Secondary antibodies were antimouse IgG � M or antirabbitIgG alkaline phosphatase conjugates at 0.1 �g/ml (Biosource). The Mr valuesof protein bands were identified by running gels with Kaleidoscope prestainedstandards (Bio-Rad). Brain homogenates were 10% or 20% (w/v) in 1%sarcosyl/TBS. PK treatment of brain homogenates and purified proteins wasdone with 50 �g/ml PK at 37°C for 1 h, followed by treatment with PefablocSC (Roche, Indianapolis, IN, USA) and Complete protease inhibitor cocktail(Roche). Protein concentrations in brain homogenates were quantified asdescribed above under Preparation of proteins.
Immunohistochemistry
As described previously [38, 39], clinical, 87V mouse-adapted, scrapie-in-fected MB/Dk mice [40] were lethally anesthetized with sodium pentobarbitali.p. and perfused transaortically with PBS, followed by 4% paraformaldehyde/PBS. Brains were subsequently removed, incubated for 2 days in 4% parafor-maldehyde/PBS at 4°C, and then transferred to a preservation solution con-taining 30% glycerol/10% DMSO in PBS, also at 4°C until sectioning. Serialcoronal brain sections of 40 �m thickness were prepared by microtome (Leica,Nussloch, Germany) and subsequently stained with mAb 6D11 at 3 �g/ml,with or without prior PK treatment (50 �g/ml for 1 h). Tissue staining,including peroxidase quenching and nonspecific signal blocking, was com-
pleted with a Vector Elite kit (Vector Laboratories), according to the manu-facturer’s directions.
Mouse immunizations
Six-week-old female mice were immunized i.p. with semipurified, nondena-tured, PK-treated 139A SAF (25 �g) in PBS (25 �l), emulsified with 10%AlCl3 (50 �l) in TiterMax (150 �l) with 25 �l water or ODN 1826 (63 �g, 10nmol). After the first immunization only, mice were administered daily doses ofODN 1826 (63 �g) or vehicle (PBS) for the subsequent 4 (Prnp�/� andBALB/cJ) or 2 (C57BL/6J) days. C57BL/6J mice were administered ODN 1826for only 2 days, as further dosing with these levels of ODN 1826 was found tobe lethal for these mice. Mice were retro-orbitally bled under Avertin-inducedanesthesia 3 days prior to the first immunization (for preimmune controls) andat 2-week intervals postimmunization.
ODN 1826 [5�-TCC ATG ACG TTC CTG ACG TT-3� (CpG motifs under-lined)], with a complete phosphorothioate backbone, was purchased fromIntegrated DNA Technologies (Coralville, IA, USA).
RESULTS
ODN 1826 greatly potentiates the humoralimmune response to PrPSc in PrPC null mice
Preliminary studies had been conducted to examine the im-munogenicity of nondenatured PrPSc in Prnp�/� mice. Plasmafrom Prnp�/� mice, which were immunized four times with139A SAF, displayed moderate immunoreactivity to the im-munogen (E.D., �20�103; pepscan results, see Fig. 5B). Thisfinding indicated that antibodies to nondenatured PrPSc couldbe generated in Prnp�/� mice and suggested that stimulationof the innate immune response using CpG ODNs may result ina significantly heightened response to PrPSc after fewer immu-nizations.
Prnp�/� mice were immunized with inocula containing139A SAF alone (identical to that described for the miceimmunized four times) or 139A SAF and ODN 1826 to stim-ulate innate immunity, henceforth referred to as Inocula A(139A only) and B (139A�1826), respectively. Plasma PrPSc
immunoreactivity (specific IgG�M) was then monitored overtime. After an initial spike in plasma anti-PrPSc levels follow-ing immunization (i.e., antibodies capable of reacting to PrPSc),titers in all mice decreased to relatively low levels by Week 10.Mice were reimmunized at Week 11 and monitored for anti-PrPSc levels for an additional 11 weeks. As reported in Table1 (see also Fig. 1), immunization with Inoculum B resulted inantigen-specific antibody titers, which were 18.9-fold greater
TABLE 1. Ratios for Analyzing ODN 1826 Effect in 139A SAF-Immunized Prnp�/� Mice
139A � 1826(E.D. SEM � 103)
139A (E.D. SEM � 103)
t-TestP value
139A � 1826/139Aratio Derivation of ratios
134.5 12.6 7.1 3.2 0.05 18.9 Week 12/Week 12134.5 12.6 10.2 5.1 0.05 13.2 Week 12/Week 14147.7 0.6 11.2 4.3 0.001 13.2 peak E.D./peak E.D.
Comparative analyses of anti-PrPSc humoral immunity for immunized Prnp�/� mice. Week 12 is the highest mean E.D. time-point for the 139A � 1826 group;Week 14 is the highest mean E.D. for the 139A group; for the peak E.D.:peak E.D. ratio, peak E.D. is the mean for the pooled, highest E.D. for each mouse ina group (n�3), regardless of time-point. E.D. values for each mouse were the average of three separate determinations. Significance was measured as probabilityby two-tailed Student’s t-test.
Spinner et al. CpG DNA enhances humoral immunity to PrPSc in immunized mice 3
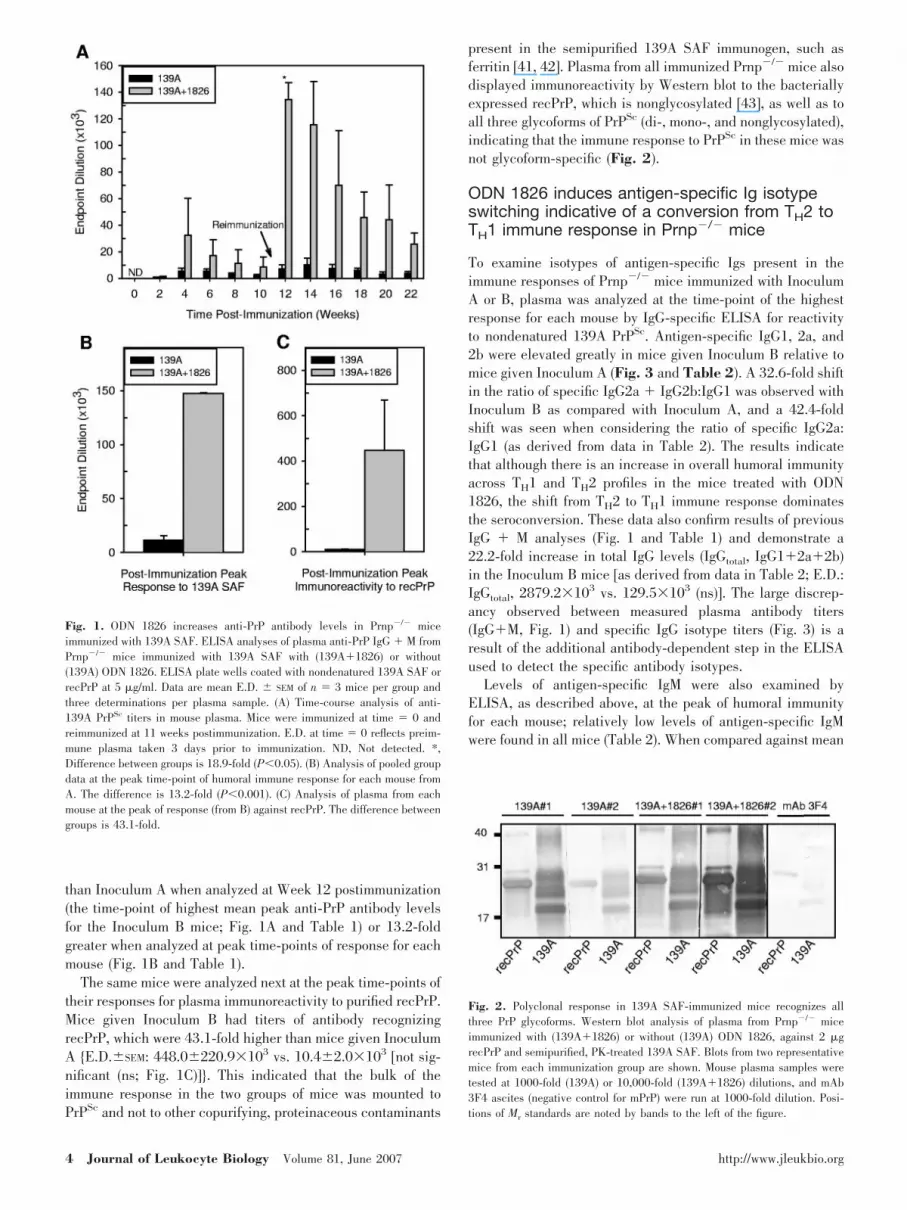
than Inoculum A when analyzed at Week 12 postimmunization(the time-point of highest mean peak anti-PrP antibody levelsfor the Inoculum B mice; Fig. 1A and Table 1) or 13.2-foldgreater when analyzed at peak time-points of response for eachmouse (Fig. 1B and Table 1).
The same mice were analyzed next at the peak time-points oftheir responses for plasma immunoreactivity to purified recPrP.Mice given Inoculum B had titers of antibody recognizingrecPrP, which were 43.1-fold higher than mice given InoculumA {E.D.SEM: 448.0220.9�103 vs. 10.42.0�103 [not sig-nificant (ns; Fig. 1C)]}. This indicated that the bulk of theimmune response in the two groups of mice was mounted toPrPSc and not to other copurifying, proteinaceous contaminants
present in the semipurified 139A SAF immunogen, such asferritin [41, 42]. Plasma from all immunized Prnp�/� mice alsodisplayed immunoreactivity by Western blot to the bacteriallyexpressed recPrP, which is nonglycosylated [43], as well as toall three glycoforms of PrPSc (di-, mono-, and nonglycosylated),indicating that the immune response to PrPSc in these mice wasnot glycoform-specific (Fig. 2).
ODN 1826 induces antigen-specific Ig isotypeswitching indicative of a conversion from TH2 toTH1 immune response in Prnp�/� mice
To examine isotypes of antigen-specific Igs present in theimmune responses of Prnp�/� mice immunized with InoculumA or B, plasma was analyzed at the time-point of the highestresponse for each mouse by IgG-specific ELISA for reactivityto nondenatured 139A PrPSc. Antigen-specific IgG1, 2a, and2b were elevated greatly in mice given Inoculum B relative tomice given Inoculum A (Fig. 3 and Table 2). A 32.6-fold shiftin the ratio of specific IgG2a � IgG2b:IgG1 was observed withInoculum B as compared with Inoculum A, and a 42.4-foldshift was seen when considering the ratio of specific IgG2a:IgG1 (as derived from data in Table 2). The results indicatethat although there is an increase in overall humoral immunityacross TH1 and TH2 profiles in the mice treated with ODN1826, the shift from TH2 to TH1 immune response dominatesthe seroconversion. These data also confirm results of previousIgG � M analyses (Fig. 1 and Table 1) and demonstrate a22.2-fold increase in total IgG levels (IgGtotal, IgG1�2a�2b)in the Inoculum B mice [as derived from data in Table 2; E.D.:IgGtotal, 2879.2�103 vs. 129.5�103 (ns)]. The large discrep-ancy observed between measured plasma antibody titers(IgG�M, Fig. 1) and specific IgG isotype titers (Fig. 3) is aresult of the additional antibody-dependent step in the ELISAused to detect the specific antibody isotypes.
Levels of antigen-specific IgM were also examined byELISA, as described above, at the peak of humoral immunityfor each mouse; relatively low levels of antigen-specific IgMwere found in all mice (Table 2). When compared against mean
Fig. 2. Polyclonal response in 139A SAF-immunized mice recognizes allthree PrP glycoforms. Western blot analysis of plasma from Prnp�/� miceimmunized with (139A�1826) or without (139A) ODN 1826, against 2 �grecPrP and semipurified, PK-treated 139A SAF. Blots from two representativemice from each immunization group are shown. Mouse plasma samples weretested at 1000-fold (139A) or 10,000-fold (139A�1826) dilutions, and mAb3F4 ascites (negative control for mPrP) were run at 1000-fold dilution. Posi-tions of Mr standards are noted by bands to the left of the figure.
Fig. 1. ODN 1826 increases anti-PrP antibody levels in Prnp�/� miceimmunized with 139A SAF. ELISA analyses of plasma anti-PrP IgG � M fromPrnp�/� mice immunized with 139A SAF with (139A�1826) or without(139A) ODN 1826. ELISA plate wells coated with nondenatured 139A SAF orrecPrP at 5 �g/ml. Data are mean E.D. SEM of n � 3 mice per group andthree determinations per plasma sample. (A) Time-course analysis of anti-139A PrPSc titers in mouse plasma. Mice were immunized at time � 0 andreimmunized at 11 weeks postimmunization. E.D. at time � 0 reflects preim-mune plasma taken 3 days prior to immunization. ND, Not detected. *,Difference between groups is 18.9-fold (P0.05). (B) Analysis of pooled groupdata at the peak time-point of humoral immune response for each mouse fromA. The difference is 13.2-fold (P0.001). (C) Analysis of plasma from eachmouse at the peak of response (from B) against recPrP. The difference betweengroups is 43.1-fold.
4 Journal of Leukocyte Biology Volume 81, June 2007 http://www.jleukbio.org
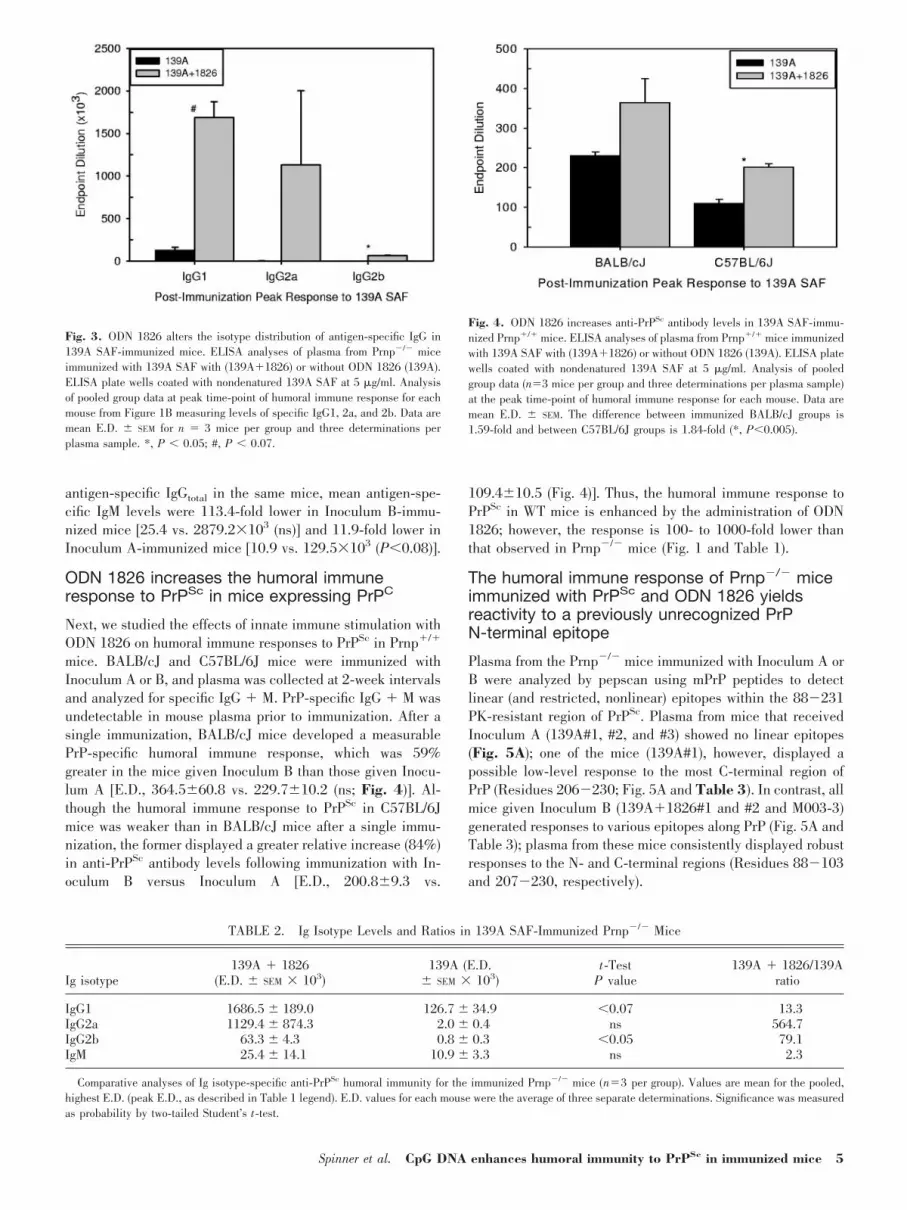
antigen-specific IgGtotal in the same mice, mean antigen-spe-cific IgM levels were 113.4-fold lower in Inoculum B-immu-nized mice [25.4 vs. 2879.2�103 (ns)] and 11.9-fold lower inInoculum A-immunized mice [10.9 vs. 129.5�103 (P0.08)].
ODN 1826 increases the humoral immuneresponse to PrPSc in mice expressing PrPC
Next, we studied the effects of innate immune stimulation withODN 1826 on humoral immune responses to PrPSc in Prnp�/�
mice. BALB/cJ and C57BL/6J mice were immunized withInoculum A or B, and plasma was collected at 2-week intervalsand analyzed for specific IgG � M. PrP-specific IgG � M wasundetectable in mouse plasma prior to immunization. After asingle immunization, BALB/cJ mice developed a measurablePrP-specific humoral immune response, which was 59%greater in the mice given Inoculum B than those given Inocu-lum A [E.D., 364.560.8 vs. 229.710.2 (ns; Fig. 4)]. Al-though the humoral immune response to PrPSc in C57BL/6Jmice was weaker than in BALB/cJ mice after a single immu-nization, the former displayed a greater relative increase (84%)in anti-PrPSc antibody levels following immunization with In-oculum B versus Inoculum A [E.D., 200.89.3 vs.
109.410.5 (Fig. 4)]. Thus, the humoral immune response toPrPSc in WT mice is enhanced by the administration of ODN1826; however, the response is 100- to 1000-fold lower thanthat observed in Prnp�/� mice (Fig. 1 and Table 1).
The humoral immune response of Prnp�/� miceimmunized with PrPSc and ODN 1826 yieldsreactivity to a previously unrecognized PrPN-terminal epitope
Plasma from the Prnp�/� mice immunized with Inoculum A orB were analyzed by pepscan using mPrP peptides to detectlinear (and restricted, nonlinear) epitopes within the 88�231PK-resistant region of PrPSc. Plasma from mice that receivedInoculum A (139A#1, #2, and #3) showed no linear epitopes(Fig. 5A); one of the mice (139A#1), however, displayed apossible low-level response to the most C-terminal region ofPrP (Residues 206�230; Fig. 5A and Table 3). In contrast, allmice given Inoculum B (139A�1826#1 and #2 and M003-3)generated responses to various epitopes along PrP (Fig. 5A andTable 3); plasma from these mice consistently displayed robustresponses to the N- and C-terminal regions (Residues 88�103and 207�230, respectively).
Fig. 3. ODN 1826 alters the isotype distribution of antigen-specific IgG in139A SAF-immunized mice. ELISA analyses of plasma from Prnp�/� miceimmunized with 139A SAF with (139A�1826) or without ODN 1826 (139A).ELISA plate wells coated with nondenatured 139A SAF at 5 �g/ml. Analysisof pooled group data at peak time-point of humoral immune response for eachmouse from Figure 1B measuring levels of specific IgG1, 2a, and 2b. Data aremean E.D. SEM for n � 3 mice per group and three determinations perplasma sample. *, P 0.05; #, P 0.07.
TABLE 2. Ig Isotype Levels and Ratios in 139A SAF-Immunized Prnp�/� Mice
Ig isotype139A � 1826
(E.D. SEM � 103)139A (E.D.
SEM � 103)t-Test
P value139A � 1826/139A
ratio
IgG1 1686.5 189.0 126.7 34.9 0.07 13.3IgG2a 1129.4 874.3 2.0 0.4 ns 564.7IgG2b 63.3 4.3 0.8 0.3 0.05 79.1IgM 25.4 14.1 10.9 3.3 ns 2.3
Comparative analyses of Ig isotype-specific anti-PrPSc humoral immunity for the immunized Prnp�/� mice (n�3 per group). Values are mean for the pooled,highest E.D. (peak E.D., as described in Table 1 legend). E.D. values for each mouse were the average of three separate determinations. Significance was measuredas probability by two-tailed Student’s t-test.
Fig. 4. ODN 1826 increases anti-PrPSc antibody levels in 139A SAF-immu-nized Prnp�/� mice. ELISA analyses of plasma from Prnp�/� mice immunizedwith 139A SAF with (139A�1826) or without ODN 1826 (139A). ELISA platewells coated with nondenatured 139A SAF at 5 �g/ml. Analysis of pooledgroup data (n�3 mice per group and three determinations per plasma sample)at the peak time-point of humoral immune response for each mouse. Data aremean E.D. SEM. The difference between immunized BALB/cJ groups is1.59-fold and between C57BL/6J groups is 1.84-fold (*, P0.005).
Spinner et al. CpG DNA enhances humoral immunity to PrPSc in immunized mice 5

To better characterize the epitope specificity of the humoralimmune response induced by Inoculum A, two mice that hadbeen immunized four times with this inoculum (describedpreviously in Results; M001-9 and M002-1) were analyzed.Mice immunized four times with Inoculum A generated detect-able responses to epitopes at the C-terminal region of PrPbetween Residues 205 and 229 (Fig. 5B and Table 3). Al-though one of these mice exhibited an antibody response withepitopes as close to the N-terminal as Residues 120�126,none exhibited any in the 88�103 region, as found in theInoculum B-immunized mice. Thus, the absence of detectableepitopes in the plasma of mice immunized twice with InoculumA was probably a reflection of the low level of humoral immu-nity in these mice. In addition, these results indicate that thepresence of ODN 1826 in Inoculum B allowed for the gener-ation of antibodies to the N-terminal region of PrP, which werenotably absent from the mice treated with Inoculum A (Fig. 5Aand Table 3). Although pepscan-identifiable epitopes in the
N-terminal domain of PrP were displayed only in InoculumB-treated mice, it cannot be excluded that nonlinear epitopesto this region of PrP were present but not detected in micegiven Inoculum A.
Characterization of mAb 6D11 derived from aPrnp�/� mouse immunized with PrPSc
and ODN 1826
The significantly elevated antibody response and unique N-terminal epitope observed in Inoculum B-treated miceprompted us to use these mice to generate mAb. The InoculumB-immunized Prnp�/� mouse with the greatest level of hu-moral immunity to PrPSc (M003-3; see Fig. 5A) was used togenerate hybridomas (fusion with myeloma line P3/NSI/1-Ag4-1 from ATCC), and an array of hybridoma clones wasgenerated; the anti-PrP mAb secreted by the hybridoma line6D11 was fully characterized in this study.
Fig. 5. ODN 1826 induces antibodies to a unique N-terminal epitope in139A SAF-immunized Prnp�/� mice. Pepscan epitope analyses of plasmafrom immunized Prnp�/� mice. Number ranges above pepscan peaks repre-sent PrP epitope residues in the corresponding peak. The shade of thenumbers (black or gray) corresponds to that of the plot that they describe. (A,upper panel) Analyses with 30-mer peptides spanning mPrP 61�240 frommice immunized twice with 139A SAF alone. (A, lower panel) Analyses with30-mer peptides (as described for A) from mice immunized twice with 139A� 1826. (B) Analyses with 15-mer peptides spanning PrP 1�254 from miceimmunized four times with 139A alone.
TABLE 3. mPrP Epitopes Represented in Pepscan Analyses of Plasma from 139A SAF-Immunized Prnp�/� Mice
Mouse ID ImmunogenNumber of
immunizationsMurine PrP epitope(residue numbers) PrP epitope sequence
139A#1 139A SAF 2 – –139A#2 139A SAF 2 – –139A#3 139A SAF 2 206–230 ERVVEQMCVTQYGKESQAYYDGRRS139A � 1826#1 139A SAF �
ODN 18262 96–100
206–211NQWNKERVVEQ
139A � 1826#2 139A SAF �ODN 1826
2 96–100206–211
NQWNKERVVEQ
M003-3 139A SAF �ODN 1826
2 88–103132–149211–230
WGQGGGTHNQWNKPSKAMSRPMIHFGNDWEDRYY
QMCVTQYQKESQAYYDGRRSSM001-9 139A SAF 4 204–211 MMERVVEQM002-1 139A SAF 4 120–126
140–149220–229
VVGGLGGFGNDWEDRYYESQAYYDGRR
Linear epitopes identified in Pepscan analyses of mice described in the text.
6 Journal of Leukocyte Biology Volume 81, June 2007 http://www.jleukbio.org

Binding and reactivity
mAb 6D11 is of the IgG2a� isotype, which is the subclass(IgG2a) that exhibited the greatest increase (�560-fold; seeTable 2) in the Prnp�/� mice treated with ODN 1826. mAb6D11 exhibits a broad species specificity by Western blot,which includes PrP of bovine, cervid (elk and mule deer),human, ovine, and rodent (hamster and mouse; Fig. 6). Reac-tivity of 6D11 was also tested by ELISA against nondenaturedforms of 139A and hamster-adapted scrapie 263K PrPSc, semi-denatured (formic acid-treated) 139A PrPSc, and recPrP andwas found to bind very well to these antigens (data not shown).The binding affinity/avidity of 6D11 for recPrP, as measured byKd, was determined by ELISA to be 40 28 pM (KdSD,n�2). This Kd is comparable with that of the mAb 3F4 forSyrian hamster rPrP and that of 7A12 for recPrP (Kd values of�80 and �190 pM, respectively; D. S. Spinner and R. J.Kascsak, unpublished data). In addition, 6D11 reactedstrongly with nondenatured PrPSc deposits in PK- and non-PK-treated brain sections taken during the clinical phase of miceinfected with the plaque-forming, 87V mouse-adapted scrapiestrain (Fig. 7); immunostaining of intraneuronal and extracel-lular PrPSc, but not PrPC, was observed. The recognition ofPrPC and nondenatured and denatured forms of PrPSc in avariety of formats indicates that mAb 6D11 recognizes anepitope whose structure is conserved in PrPC and PrPSc andwhich is readily accessible in the two PrP isoforms.
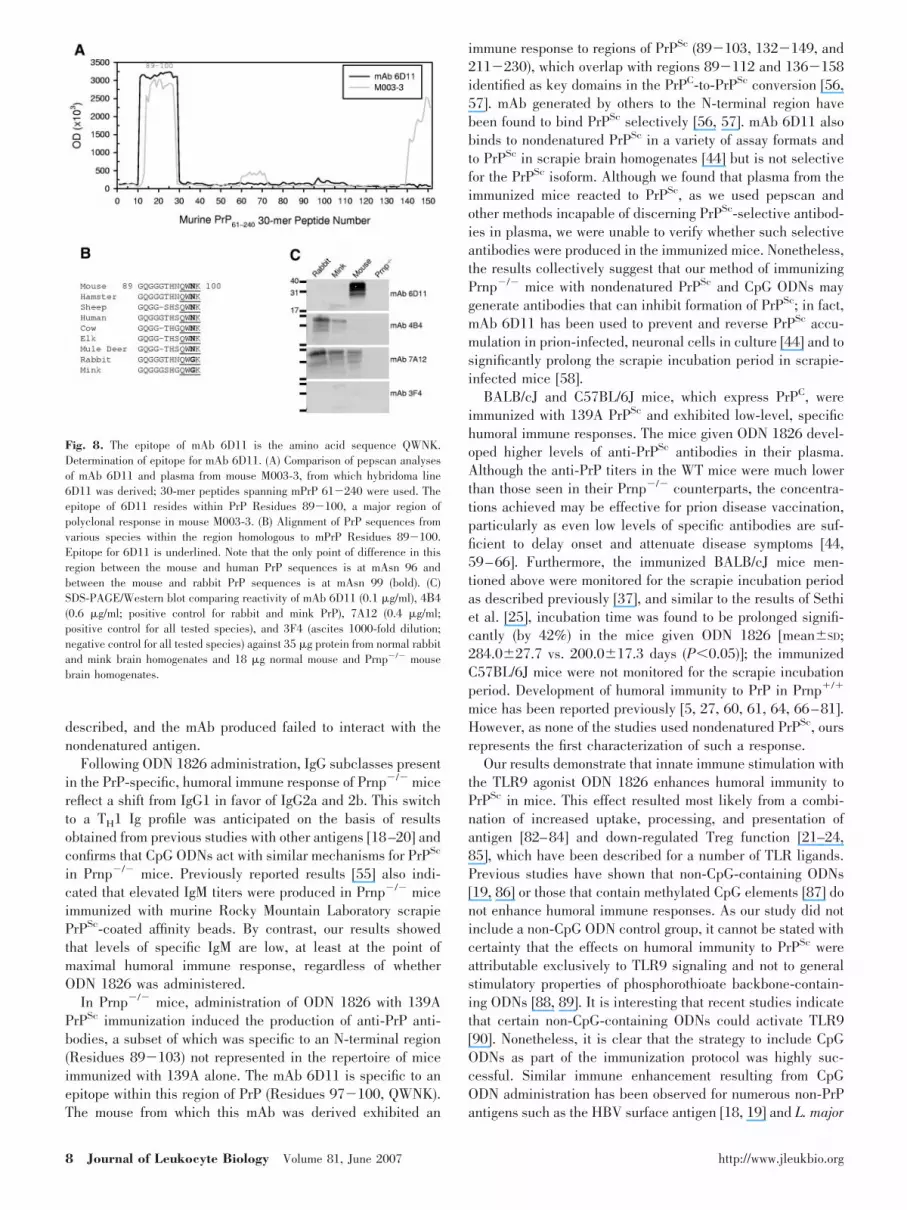
Epitope determination
The epitope of 6D11 was mapped by pepscan analysis to withinthe N-terminal region of PrP, Residues 89�100 (GQGGGTH-NQWNK; Fig. 8A). This region of PrP also displayed thestrongest immunoreactivity in the polyclonal response of themouse, from which 6D11 was derived (M003-3; Figs. 5A and8A). On the basis of the species specificity exhibited in West-ern blot, peptide reactivity determined previously in dot-blot[44], and knowledge of the PrP sequences in the various
species, the 6D11 epitope was further narrowed to Residues97�100 (QWNK; Fig. 8B). The asparagine of QWNK (murinePosition 99) is replaced by a glycine in the rabbit and mink PrPsequences, which we had not examined previously by Westernblotting; it is notable that in the region of 89�100, the PrPsequences of rabbit and mouse differ only at Residue 99 (Fig.8B). To confirm the epitope of 6D11 to be QWNK, Westernblotting was performed against normal rabbit and mink brainhomogenates (Fig. 8C). As expected, the reactivity of 6D11 inthe Western blot excludes PrPC of rabbit and mink, whereasmAb 4B4 and 7A12 stain these proteins, confirming the se-quence QWNK as the epitope for 6D11. In addition, moreextensive, higher resolution pepscan analyses on 6D11 usingtruncated peptides further confirmed QWNK as the epitope(data not shown). This epitope imparts upon 6D11 unique andhighly desirable diagnostic and therapeutic properties (seeDiscussion).
DISCUSSION
We have shown that Prnp�/� mice are capable of generating ahumoral immune response to nondenatured 139A PrPSc, whichis increased significantly by the TLR9 agonist ODN 1826.Several other studies have demonstrated humoral immunity toPrP in Prnp–/– mice and the ability to develop anti-PrP, mAb-secreting hybridomas from them [6, 7, 10, 27, 28, 45–54]. Ofthese studies, one reported the generation of mAb followingimmunization with nondenatured PrPSc [51]; however, in thatstudy, the nature of the humoral immune response was not
Fig. 6. mAb 6D11 recognizes PrP isoforms from multiple species. Analysis ofmAb 6D11 reactivity to PrP by Western blotting. SDS-PAGE/Western blot of6D11 (0.4 �g/ml), pAb 78295 (positive control for homogenate PrP levels;5000-fold dilution), and secondary antibodies only (0.1 �g/ml) tested against8 �g protein (measured prior to PK treatment) loaded to gel from brainhomogenates of a normal mouse (C57BL/6J), 139A scrapie-infected mouse(139A in C57BL/6J), Prnp�/� mouse [knockout (KO)], Syrian hamster (SHa),or TgPrP mice expressing TgShp, TgHu, BoTg, TgElk, or TgMuD PrP.
Fig. 7. mAb 6D11 reacts to PrPSc deposits in PK- and non-PK-treated brainsections from plaque-forming, scrapie-infected mice. Immunohistochemistryon fixed brain sections from clinical, 87V scrapie-infected MB/Dk mice. Fixedtissue sections were treated with (�) or without (�) PK prior to immunostain-ing with mAb 6D11 (3 �g/ml). Original magnifications, 200�. Closed andopen arrowheads mark select mAb 6D11-stained extracellular and intraneu-ronal PrPSc deposits, respectively. (A) Coronal brain section of cortical regionoverlying the hippocampus. (B) PK-treated coronal brain section.
Spinner et al. CpG DNA enhances humoral immunity to PrPSc in immunized mice 7
described, and the mAb produced failed to interact with thenondenatured antigen.
Following ODN 1826 administration, IgG subclasses presentin the PrP-specific, humoral immune response of Prnp�/� micereflect a shift from IgG1 in favor of IgG2a and 2b. This switchto a TH1 Ig profile was anticipated on the basis of resultsobtained from previous studies with other antigens [18–20] andconfirms that CpG ODNs act with similar mechanisms for PrPSc
in Prnp�/� mice. Previously reported results [55] also indi-cated that elevated IgM titers were produced in Prnp�/� miceimmunized with murine Rocky Mountain Laboratory scrapiePrPSc-coated affinity beads. By contrast, our results showedthat levels of specific IgM are low, at least at the point ofmaximal humoral immune response, regardless of whetherODN 1826 was administered.
In Prnp�/� mice, administration of ODN 1826 with 139APrPSc immunization induced the production of anti-PrP anti-bodies, a subset of which was specific to an N-terminal region(Residues 89�103) not represented in the repertoire of miceimmunized with 139A alone. The mAb 6D11 is specific to anepitope within this region of PrP (Residues 97�100, QWNK).The mouse from which this mAb was derived exhibited an
immune response to regions of PrPSc (89�103, 132�149, and211�230), which overlap with regions 89�112 and 136�158identified as key domains in the PrPC-to-PrPSc conversion [56,57]. mAb generated by others to the N-terminal region havebeen found to bind PrPSc selectively [56, 57]. mAb 6D11 alsobinds to nondenatured PrPSc in a variety of assay formats andto PrPSc in scrapie brain homogenates [44] but is not selectivefor the PrPSc isoform. Although we found that plasma from theimmunized mice reacted to PrPSc, as we used pepscan andother methods incapable of discerning PrPSc-selective antibod-ies in plasma, we were unable to verify whether such selectiveantibodies were produced in the immunized mice. Nonetheless,the results collectively suggest that our method of immunizingPrnp�/� mice with nondenatured PrPSc and CpG ODNs maygenerate antibodies that can inhibit formation of PrPSc; in fact,mAb 6D11 has been used to prevent and reverse PrPSc accu-mulation in prion-infected, neuronal cells in culture [44] and tosignificantly prolong the scrapie incubation period in scrapie-infected mice [58].
BALB/cJ and C57BL/6J mice, which express PrPC, wereimmunized with 139A PrPSc and exhibited low-level, specifichumoral immune responses. The mice given ODN 1826 devel-oped higher levels of anti-PrPSc antibodies in their plasma.Although the anti-PrP titers in the WT mice were much lowerthan those seen in their Prnp�/� counterparts, the concentra-tions achieved may be effective for prion disease vaccination,particularly as even low levels of specific antibodies are suf-ficient to delay onset and attenuate disease symptoms [44,59–66]. Furthermore, the immunized BALB/cJ mice men-tioned above were monitored for the scrapie incubation periodas described previously [37], and similar to the results of Sethiet al. [25], incubation time was found to be prolonged signifi-cantly (by 42%) in the mice given ODN 1826 [meanSD;284.027.7 vs. 200.017.3 days (P0.05)]; the immunizedC57BL/6J mice were not monitored for the scrapie incubationperiod. Development of humoral immunity to PrP in Prnp�/�
mice has been reported previously [5, 27, 60, 61, 64, 66–81].However, as none of the studies used nondenatured PrPSc, oursrepresents the first characterization of such a response.
Our results demonstrate that innate immune stimulation withthe TLR9 agonist ODN 1826 enhances humoral immunity toPrPSc in mice. This effect resulted most likely from a combi-nation of increased uptake, processing, and presentation ofantigen [82–84] and down-regulated Treg function [21–24,85], which have been described for a number of TLR ligands.Previous studies have shown that non-CpG-containing ODNs[19, 86] or those that contain methylated CpG elements [87] donot enhance humoral immune responses. As our study did notinclude a non-CpG ODN control group, it cannot be stated withcertainty that the effects on humoral immunity to PrPSc wereattributable exclusively to TLR9 signaling and not to generalstimulatory properties of phosphorothioate backbone-contain-ing ODNs [88, 89]. It is interesting that recent studies indicatethat certain non-CpG-containing ODNs could activate TLR9[90]. Nonetheless, it is clear that the strategy to include CpGODNs as part of the immunization protocol was highly suc-cessful. Similar immune enhancement resulting from CpGODN administration has been observed for numerous non-PrPantigens such as the HBV surface antigen [18, 19] and L. major
Fig. 8. The epitope of mAb 6D11 is the amino acid sequence QWNK.Determination of epitope for mAb 6D11. (A) Comparison of pepscan analysesof mAb 6D11 and plasma from mouse M003-3, from which hybridoma line6D11 was derived; 30-mer peptides spanning mPrP 61�240 were used. Theepitope of 6D11 resides within PrP Residues 89�100, a major region ofpolyclonal response in mouse M003-3. (B) Alignment of PrP sequences fromvarious species within the region homologous to mPrP Residues 89�100.Epitope for 6D11 is underlined. Note that the only point of difference in thisregion between the mouse and human PrP sequences is at mAsn 96 andbetween the mouse and rabbit PrP sequences is at mAsn 99 (bold). (C)SDS-PAGE/Western blot comparing reactivity of mAb 6D11 (0.1 �g/ml), 4 4(0.6 �g/ml; positive control for rabbit and mink PrP), 7A12 (0.4 �g/ml;positive control for all tested species), and 3F4 (ascites 1000-fold dilution;negative control for all tested species) against 35 �g protein from normal rabbitand mink brain homogenates and 18 �g normal mouse and Prnp�/� mousebrain homogenates.
8 Journal of Leukocyte Biology Volume 81, June 2007 http://www.jleukbio.org

antigens [20]. Obviously, mice lacking the gene encoding PrPC
possess a greater capacity to respond to PrP than do theirPrnp�/� counterparts. Primarily, this effect is believed to befrom tolerance induced by endogenous PrPC, which does notoccur in Prnp�/� mice. However, some of the differencesobserved between Prnp�/� and Prnp�/� mice may be attrib-utable to a lower capacity of Prnp�/� immune cells for phago-cytosis [91], a process crucial for antigen presentation andhence, initiation of adaptive immunity. It is also possible thatantibody titers in WT mice may appear artificially low as aresult of circulating blood cells and other cell types thatexpress high levels of surface PrP [92, 93], which can adsorbanti-PrP antibodies and mask their presence. Similar antibodyadsorption effects have been seen in amyloid-�-immunizedTgAPP mice [94].
A recent study by Heikenwalder and colleagues [95] re-ported ODN 1826-dependent induction of lymphoreticulardamage in WT mice after just 2 consecutive days of adminis-tration in doses similar to those used in the present study. Theydemonstrated that severe damage to the lymphoreticular sys-tem occurred by Day 7 of treatment and grew progressivelyworse with increasing numbers of successive daily doses. Onewould expect increasing lymphoreticular destruction to se-verely diminish the capacity of mice to initiate adaptive im-mune responses. Results obtained in the present study (andthose of others including Sethi and colleagues [96]) differ fromthose of Heikenwalder et al. [95] in that we found repeatedtreatments with ODN 1826 to increase humoral immune re-sponses in Prnp�/� and Prnp�/� mice. In our study, after 3 or5 consecutive daily doses of ODN 1826, significant potentia-tion of anti-PrP antibody production occurred. In addition, totalplasma IgG levels in ODN 1826-treated Prnp�/� mice were inthe normal range after the first and second immunizations atthe peaks of humoral immune response for each mouse whenmeasured by radial immunodiffusion assay. After one immu-nization, Prnp�/� mice inoculated with 139A � 1826 hadlevels of total plasma IgG 1.8-fold higher (P0.05) than miceimmunized with 139A alone (meanSD; 27.10.9 vs.15.41.5 mg/ml; n�3 mice per group, with two determinationsper plasma sample). Levels of total plasma IgG in mice of bothimmunization groups were identical after the second immuni-zation (139A 14.02.0 vs. 139�1826, 14.11.8 mg/ml).
Heikenwalder et al. [95] reported mouse strain-dependentdifferences in toxicity, which we also observed. In the currentstudy, C57BL/6J mice were administered ODN 1826 for only 3consecutive days (including the immunization day; vs. 5), asthe levels of ODN 1826 used were found to be lethal for someof these mice when administered over a longer period. ThePrnp�/� (FVB/N strain) and BALB/cJ mice received five dosesof ODN 1826, which may be expected to result in toxicity anddiminished immune response, yet we observed no overt symp-toms. It may be noted that one difference between our studyand that reported by others [95] was the use of the TiterMaxadjuvant in the present study.
Our data for Prnp�/� mice suggest that CpG ODN-inducedprotection against prion disease in mice [25] may be partlyattributable to therapeutic antibody production, along withincreased clearance of the scrapie agent by activated phago-cytes. The antigen-specific IgGs present and particularly those
of the IgG2a or 2b isotype, which we found to be increasedsubstantially following ODN 1826 treatment in the Prnp�/�
mice, are likely to be highly effective opsonins for PrPSc [97].In addition, T cell-based mechanisms, including down-regula-tion of Treg function [21–24, 85], enhanced levels of antigencross-presentation [98–101], and increased activation of CD8�
T cells [86, 102–104] are likely to play important roles in thetherapeutic effect of CpG ODNs on scrapie; these effects maybe especially pertinent, as T cell effector functions are dimin-ished in scrapie-infected mice despite their ability to bindmajor histocompatibility complex:PrP peptide tetramers [105].Recently, additional mechanisms have been proposed for phos-phorothioate backbone-containing ODNs in scrapie prophy-laxis involving the binding of these molecules to PrPC andpreventing its conversion to PrPSc [106].
Our studies demonstrate that the ODN 1826 enhances adap-tive/humoral immune responses to PrPSc in mice and therefore,also may greatly improve the efficacy of vaccination [60, 61,64–66] and passive immunization [44, 59, 62, 63] against TSEinfection. mAb 6D11, induced in the presence of ODN 1826,and similar mAb possessing high affinities and unique epitopespecificities derived from PrPSc appear to be ideal candidatesthat offer considerable diagnostic [27, 107, 108] and therapeu-tic [44] promise. As improved therapy at later stages of diseaseand diagnosis at earlier stages of prion disease remain majorgoals of this field, it is clear that further studies pertinent to thedevelopment of such antibodies to PrP are important.
ACKNOWLEDGMENTS
This work was supported in part by the New York State Officeof Mental Retardation and Developmental Disabilities, Na-tional Institutes of Health, Contract N01-NS-0-2327 (R. J. K.,Subcontract P.I.; Robert G. Rohwer, Contract P.I.) and GrantNS047433 (to T. W.), and Alzheimer’s Association/StranahanFoundation Grant NIRG-04-1162 (to D. S. S.). The authorsgratefully acknowledge Larisa Cervenakova (American RedCross, Rockville, MD, USA) for the gift of bovine PrP Tg mouse(BoTg 3204) brains, Man-Sun Sy (Case Western Reserve Uni-versity, Cleveland, OH, USA) for providing mAb 7A12, andIlia V. Baskakov (University of Maryland, Baltimore, MD,USA), Robert G. Rohwer, and Luisa Gregori (Department ofVeterans Affairs Medical Center, Baltimore, MD, USA) forSyrian hamster rPrP used for mAb Kd determination. We alsothank Jerry Slootstra (Pepscan Systems, Lelystad, The Nether-lands) for help in interpreting pepscan analyses; MichaelNatelli and Marisol Cedeno for maintaining Tg mouse lines;Victor Sapienza, Heni Hong, and Chengmo James Chen fortechnical assistance; Patricia A. Merz for helpful discussions;Mary Ellen Cafaro and Robert L. Freedland for graphicalassistance; and Elaine J. Marchi for editorial assistance. Con-flict of Interest Disclosure: D. S. S., R. B. K., H. C. M., R. I. C.,and/or R. J. K. may receive royalties from the sale of mAb 3F4,4B4, and 6D11 described in the text. These mAb are soldthrough Signet Laboratories (Dedham, MA, USA).
Spinner et al. CpG DNA enhances humoral immunity to PrPSc in immunized mice 9

REFERENCES
1. Aguzzi, A., Heikenwalder, M. (2006) Pathogenesis of prion diseases:status and outlook. Nat. Rev. Microbiol. 4, 765–775.
2. Spinner, D. S., Ye, X., Carp, R. I., Kascsak, R. J. (2003) The role ofcomponents of the lymphoreticular system in transmissible spongiformencephalopathy pathogenesis and neuroinvasion. Curr. Med. Chem. Im-mun. Endocrinol. Metab. Agents 3, 119–135.
3. Porter, D. D., Porter, H. G., Cox, N. A. (1973) Failure to demonstrate ahumoral immune response to scrapie infection in mice. J. Immunol.111, 1407–1410.
4. Kasper, K. C., Stites, D. P., Bowman, K. A., Panitch, H., Prusiner, S. B.(1982) Immunological studies of scrapie infection. J. Neuroimmunol. 3,187–201.
5. Kascsak, R. J., Rubenstein, R., Merz, P. A., Tonna-DeMasi, M., Fersko,R., Carp, R. I., Wisniewski, H. M., Diringer, H. (1987) Mouse polyclonaland monoclonal antibody to scrapie-associated fibril proteins. J. Virol.61, 3688–3693.
6. Rubenstein, R., Kascsak, R. J., Papini, M., Kascsak, R., Carp, R. I.,LaFauci, G., Meloen, R., Langeveld, J. (1999) Immune surveillance andantigen conformation determines humoral immune response to the prionprotein immunogen. J. Neurovirol. 5, 401–413.
7. Rubenstein, R., Kascsak, R. J., Papini, M., Kascsak, R., Carp, R. I.,Langeveld, J. (1999) Identification of a highly immunogenic site on themurine prion protein. Alzheimer’s Dis. Rev. 4, 13–18.
8. Prusiner, S. B., Groth, D., Serban, A., Koehler, R., Foster, D., Torchia,M., Burton, D., Yang, S. L., DeArmond, S. J. (1993) Ablation of the prionprotein (PrP) gene in mice prevents scrapie and facilitates production ofanti-PrP antibodies. Proc. Natl. Acad. Sci. USA 90, 10608–10612.
9. Krasemann, S., Groschup, M. H., Harmeyer, S., Hunsmann, G., Bode-mer, W. (1996) Generation of monoclonal antibodies against humanprion proteins in PrP0/0 mice. Mol. Med. 2, 725–734.
10. Williamson, R. A., Peretz, D., Smorodinsky, N., Bastidas, R., Serban, H.,Mehlhorn, I., DeArmond, S. J., Prusiner, S. B., Burton, D. R. (1996)Circumventing tolerance to generate autologous monoclonal antibodies tothe prion protein. Proc. Natl. Acad. Sci. USA 93, 7279–7282.
11. Janeway Jr., C. A., Medzhitov, R. (2002) Innate immune recognition.Annu. Rev. Immunol. 20, 197–216.
12. Sieling, P. A., Modlin, R. L. (2002) Toll-like receptors: mammalian“taste receptors” for a smorgasbord of microbial invaders. Curr. Opin.Microbiol. 5, 70–75.
13. van Eden, W., Koets, A., van Kooten, P., Prakken, B., van der Zee, R.(2003) Immunopotentiating heat shock proteins: negotiators betweeninnate danger and control of autoimmunity. Vaccine 21, 897–901.
14. Tsan, M. F., Gao, B. (2004) Endogenous ligands of Toll-like receptors.J. Leukoc. Biol. 76, 514–519.
15. Schnare, M., Barton, G. M., Holt, A. C., Takeda, K., Akira, S., Medzhitov,R. (2001) Toll-like receptors control activation of adaptive immuneresponses. Nat. Immunol. 2, 947–950.
16. Krieg, A. M. (2002) CpG motifs in bacterial DNA and their immuneeffects. Annu. Rev. Immunol. 20, 709–760.
17. Krieg, A. M. (2006) Therapeutic potential of Toll-like receptor 9 activa-tion. Nat. Rev. Drug Discov. 5, 471–484.
18. Brazolot Millan, C. L., Weeratna, R., Krieg, A. M., Siegrist, C. A., Davis,H. L. (1998) CpG DNA can induce strong Th1 humoral and cell-mediated immune responses against hepatitis B surface antigen in youngmice. Proc. Natl. Acad. Sci. USA 95, 15553–15558.
19. Davis, H. L., Weeratna, R., Waldschmidt, T. J., Tygrett, L., Schorr, J.,Krieg, A. M. (1998) CpG DNA is a potent enhancer of specific immunityin mice immunized with recombinant hepatitis B surface antigen. J. Im-munol. 160, 870–876.
20. Zimmermann, S., Egeter, O., Hausmann, S., Lipford, G. B., Rocken, M.,Wagner, H., Heeg, K. (1998) CpG oligodeoxynucleotides trigger protec-tive and curative Th1 responses in lethal murine leishmaniasis. J. Im-munol. 160, 3627–3630.
21. Pasare, C., Medzhitov, R. (2003) Toll pathway-dependent blockade ofCD4�CD25� T cell-mediated suppression by dendritic cells. Science299, 1033–1036.
22. Wiegard, C., Frenzel, C., Herkel, J., Kallen, K. J., Schmitt, E., Lohse,A. W. (2005) Murine liver antigen presenting cells control suppressoractivity of CD4�CD25� regulatory T cells. Hepatology 42, 193–199.
23. El Andaloussi, A., Sonabend, A. M., Han, Y., Lesniak, M. S. (2006)Stimulation of TLR9 with CpG ODN enhances apoptosis of glioma andprolongs the survival of mice with experimental brain tumors. Glia 54,526–535.
24. Wu, W., Weigand, L., Belkaid, Y., Mendez, S. (2006) Immunomodulatoryeffects associated with a live vaccine against Leishmania major contain-ing CpG oligodeoxynucleotides. Eur. J. Immunol. 36, 3238–3247.
25. Sethi, S., Lipford, G., Wagner, H., Kretzschmar, H. (2002) Postexposureprophylaxis against prion disease with a stimulator of innate immunity.Lancet 360, 229–230.
26. Kascsak, R. J., Rubenstein, R., Carp, R. I. (1991) Evidence for biologicaland structural diversity among scrapie strains. Curr. Top. Microbiol.Immunol. 172, 139–152.
27. Kascsak, R. J., Fersko, R., Pulgiano, D., Rubenstein, R., Carp, R. I.(1997) Immunodiagnosis of prion disease. Immunol. Invest. 26, 259–268.
28. Zanusso, G., Liu, D., Ferrari, S., Hegyi, I., Yin, X., Aguzzi, A., Horne-mann, S., Liemann, S., Glockshuber, R., Manson, J. C., Brown, P.,Petersen, R. B., Gambetti, P., Sy, M. S. (1998) Prion protein expressionin different species: analysis with a panel of new mAbs. Proc. Natl. Acad.Sci. USA 95, 8812–8816.
29. Li, R., Liu, T., Wong, B. S., Pan, T., Morillas, M., Swietnicki, W.,O’Rourke, K., Gambetti, P., Surewicz, W. K., Sy, M. S. (2000) Identifi-cation of an epitope in the C terminus of normal prion protein whoseexpression is modulated by binding events in the N terminus. J. Mol.Biol. 301, 567–573.
30. Hoylaerts, M. F., Bollen, A., De Broe, M. E. (1990) The application ofenzyme kinetics to the determination of dissociation constants for anti-gen-antibody interactions in solution. J. Immunol. Methods 126, 253–261.
31. Glaser, R. W. (1993) Determination of antibody affinity by ELISA with anon-linear regression program. Evaluation of linearized approximations.J. Immunol. Methods 160, 129–133.
32. Hilmert, H., Diringer, H. (1984) A rapid and efficient method to enrichSAF-protein from scrapie brains of hamsters. Biosci. Rep. 4, 165–170.
33. Rohwer, R. G., Gajdusek, D. C. (1979) Scrapie-virus or viroid: the casefor a virus, In Search for the Cause of Multiple Sclerosis and Other ChronicDiseases of the Central Nervous System. The First International Sympo-sium of Hertie Foundation, Weinheim, Federal Republic of Germany.
34. Wong, B. S., Wang, H., Brown, D. R., Jones, I. M. (1999) Selectiveoxidation of methionine residues in prion proteins. Biochem. Biophys.Res. Commun. 259, 352–355.
35. Bueler, H., Aguzzi, A., Sailer, A., Greiner, R. A., Autenried, P., Aguet,M., Weissmann, C. (1993) Mice devoid of PrP are resistant to scrapie.Cell 73, 1339–1347.
36. Raymond, G. J., Bossers, A., Raymond, L. D., O’Rourke, K. I., McHol-land, L. E., Bryant III, P. K., Miller, M. W., Williams, E. S., Smits, M.,Caughey, B. (2000) Evidence of a molecular barrier limiting suscepti-bility of humans, cattle and sheep to chronic wasting disease. EMBO J.19, 4425–4430.
37. LaFauci, G., Carp, R. I., Meeker, H. C., Ye, X., Kim, J. I., Natelli, M.,Cedeno, M., Petersen, R. B., Kascsak, R., Rubenstein, R. (2006) Passageof chronic wasting disease prion into transgenic mice expressing RockyMountain elk (Cervus elaphus nelsoni) PrPC. J. Gen. Virol. 87, 3773–3780.
38. Sigurdsson, E. M., Brown, D. R., Alim, M. A., Scholtzova, H., Carp, R.,Meeker, H. C., Prelli, F., Frangione, B., Wisniewski, T. (2003) Copperchelation delays the onset of prion disease. J. Biol. Chem. 278, 46199–46202.
39. Sadowski, M., Pankiewicz, J., Scholtzova, H., Tsai, J., Li, Y., Carp, R. I.,Meeker, H. C., Gambetti, P., Debnath, M., Mathis, C. A., Shao, L., Gan,W. B., Klunk, W. E., Wisniewski, T. (2004) Targeting prion amyloiddeposits in vivo. J. Neuropathol. Exp. Neurol. 63, 775–784.
40. Bruce, M. E., Dickinson, A. G. (1985) Genetic control of amyloid plaqueproduction and incubation period in scrapie-infected mice. J. Neuro-pathol. Exp. Neurol. 44, 285–294.
41. Cho, H. J., Greig, A. S., Corp, C. R., Kimberlin, R. H., Chandler, R. L.,Millson, G. C. (1977) Virus-like particles from both control and scrapie-affected mouse brain. Nature 267, 459–460.
42. Mishra, R. S., Basu, S., Gu, Y., Luo, X., Zou, W. Q., Mishra, R., Li, R.,Chen, S. G., Gambetti, P., Fujioka, H., Singh, N. (2004) Protease-resistant human prion protein and ferritin are cotransported acrossCaco-2 epithelial cells: implications for species barrier in prion uptakefrom the intestine. J. Neurosci. 24, 11280–11290.
43. Kirby, L., Birkett, C. R., Rudyk, H., Gilbert, I. H., Hope, J. (2003) Invitro cell-free conversion of bacterial recombinant PrP to PrPres as amodel for conversion. J. Gen. Virol. 84, 1013–1020.
44. Pankiewicz, J., Prelli, F., Sy, M-S., Kascsak, R. J., Kascsak, R. B.,Spinner, D. S., Carp, R. I., Meeker, H. C., Sadowski, M., Wisniewski, T.(2006) Clearance and prevention of prion infection in cell culture byanti-PrP antibodies. Eur. J. Neurosci. 23, 2635–2647.
45. Demart, S., Fournier, J. G., Creminon, C., Frobert, Y., Lamoury, F.,Marce, D., Lasmezas, C., Dormont, D., Grassi, J., Deslys, J. P. (1999)New insight into abnormal prion protein using monoclonal antibodies.Biochem. Biophys. Res. Commun. 265, 652–657.
10 Journal of Leukocyte Biology Volume 81, June 2007 http://www.jleukbio.org

46. Krasemann, S., Jurgens, T., Bodemer, W. (1999) Generation of mono-clonal antibodies against prion proteins with an unconventional nucleicacid-based immunization strategy. J. Biotechnol. 73, 119–129.
47. Betemps, D., Baron, T. (2001) Molecular specificities of antibodiesagainst ovine and murine recombinant prion proteins. Biochem. Biophys.Res. Commun. 281, 101–108.
48. Yokoyama, T., Kimura, K. M., Ushiki, Y., Yamada, S., Morooka, A.,Nakashiba, T., Sassa, T., Itohara, S. (2001) In vivo conversion of cellularprion protein to pathogenic isoforms, as monitored by conformation-specific antibodies. J. Biol. Chem. 276, 11265–11271.
49. Nakamura, N., Miyamoto, K., Shimokawa, M., Nishida, N., Mohri, S.,Kitamoto, T., Horiuchi, H., Furusawa, S., Matsuda, H. (2003) Generationof antibodies against prion protein by scrapie-infected cell immunizationof PrP(0/0) mice. Hybrid. Hybridomics 22, 263–266.
50. Brun, A., Castilla, J., Ramirez, M. A., Prager, K., Parra, B., Salguero,F. J., Shiveral, D., Sanchez, C., Sanchez-Vizcaino, J. M., Douglas, A.,Torres, J. M. (2004) Proteinase K enhanced immunoreactivity of theprion protein-specific monoclonal antibody 2A11. Neurosci. Res. 48,75–83.
51. Kim, C. L., Umetani, A., Matsui, T., Ishiguro, N., Shinagawa, M.,Horiuchi, M. (2004) Antigenic characterization of an abnormal isoform ofprion protein using a new diverse panel of monoclonal antibodies.Virology 320, 40–51.
52. Khalili-Shirazi, A., Quaratino, S., Londei, M., Summers, L., Tayebi, M.,Clarke, A. R., Hawke, S. H., Jackson, G. S., Collinge, J. (2005) Proteinconformation significantly influences immune responses to prion protein.J. Immunol. 174, 3256–3263.
53. Matucci, A., Zanusso, G., Gelati, M., Farinazzo, A., Fiorini, M., Ferrari,S., Andrighetto, G., Cestari, T., Caramelli, M., Negro, A., Morbin, M.,Chiesa, R., Monaco, S., Tridente, G. (2005) Analysis of mammalianscrapie protein by novel monoclonal antibodies recognizing distinct prionprotein glycoforms: an immunoblot and immunohistochemical study atthe light and electron microscopic levels. Brain Res. Bull. 65, 155–162.
54. Yuan, F. F., Biffin, S., Brazier, M. W., Suarez, M., Cappai, R., Hill, A. F.,Collins, S. J., Sullivan, J. S., Middleton, D., Multhaup, G., Geczy, A. F.,Masters, C. L. (2005) Detection of prion epitopes on PrP and PrP oftransmissible spongiform encephalopathies using specific monoclonalantibodies to PrP. Immunol. Cell Biol. 83, 632–637.
55. Tayebi, M., Enever, P., Sattar, Z., Collinge, J., Hawke, S. (2004) Disease-associated prion protein elicits immunoglobulin M responses in vivo.Mol. Med. 10, 104–111.
56. Moroncini, G., Kanu, N., Solforosi, L., Abalos, G., Telling, G. C., Head,M., Ironside, J., Brockes, J. P., Burton, D. R., Williamson, R. A. (2004)Motif-grafted antibodies containing the replicative interface of cellularPrP are specific for PrPSc. Proc. Natl. Acad. Sci. USA 101, 10404–10409.
57. Moroncini, G., Mangieri, M., Morbin, M., Mazzoleni, G., Ghetti, B.,Gabrielli, A., Williamson, R. A., Giaccone, G., Tagliavini, F. (2006)Pathologic prion protein is specifically recognized in situ by a novel PrPconformational antibody. Neurobiol. Dis. 23, 717–724.
58. Pankiewicz, J., Prelli, F., Sy, M-S., Kascsak, R., Kascsak, R., Spinner,D. S., Carp, R., Meeker, H., Sadowski, M., Wisniewski, T. (2006)P4–433: therapeutically effective anti-prion protein monoclonal anti-bodies. Alzheimer’s Dement. 2, S645–S646.
59. Heppner, F. L., Musahl, C., Arrighi, I., Klein, M. A., Rulicke, T., Oesch,B., Zinkernagel, R. M., Kalinke, U., Aguzzi, A. (2001) Prevention ofscrapie pathogenesis by transgenic expression of anti-prion protein an-tibodies. Science 294, 178–182.
60. Sigurdsson, E. M., Brown, D. R., Daniels, M., Kascsak, R. J., Kascsak,R., Carp, R., Meeker, H. C., Frangione, B., Wisniewski, T. (2002)Immunization delays the onset of prion disease in mice. Am. J. Pathol.161, 13–17.
61. Schwarz, A., Kratke, O., Burwinkel, M., Riemer, C., Schultz, J., Hen-klein, P., Bamme, T., Baier, M. (2003) Immunization with a syntheticprion protein-derived peptide prolongs survival times of mice orallyexposed to the scrapie agent. Neurosci. Lett. 350, 187–189.
62. Sigurdsson, E. M., Sy, M. S., Li, R., Scholtzova, H., Kascsak, R. J.,Kascsak, R., Carp, R., Meeker, H. C., Frangione, B., Wisniewski, T.(2003) Anti-prion antibodies for prophylaxis following prion exposure inmice. Neurosci. Lett. 336, 185–187.
63. White, A. R., Enever, P., Tayebi, M., Mushens, R., Linehan, J., Brand-ner, S., Anstee, D., Collinge, J., Hawke, S. (2003) Monoclonal antibodiesinhibit prion replication and delay the development of prion disease.Nature 422, 80–83.
64. Goni, F., Knudsen, E., Schreiber, F., Scholtzova, H., Pankiewicz, J.,Carp, R., Meeker, H. C., Rubenstein, R., Brown, D. R., Sy, M. S.,Chabalgoity, J. A., Sigurdsson, E. M., Wisniewski, T. (2005) Mucosal
vaccination delays or prevents prion infection via an oral route. Neuro-science 133, 413–421.
65. Magri, G., Clerici, M., Dall’Ara, P., Biasin, M., Caramelli, M., Casalone,C., Giannino, M. L., Longhi, R., Piacentini, L., Della Bella, S., Pollera,C., Puricelli, M., Servida, F., Crescio, I., Boasso, A., Ponti, W., Poli, G.(2005) Decrease in pathology and progression of scrapie after immuni-zation with synthetic prion protein peptides in hamsters. Vaccine 23,2862–2868.
66. Bade, S., Baier, M., Boetel, T., Frey, A. (2006) Intranasal immunizationof Balb/c mice against prion protein attenuates orally acquired transmis-sible spongiform encephalopathy. Vaccine 24, 1242–1253.
67. O’Rourke, K. I., Baszler, T. V., Miller, J. M., Spraker, T. R., Sadler-Riggleman, I., Knowles, D. P. (1998) Monoclonal antibody F89/160.1.5defines a conserved epitope on the ruminant prion protein. J. Clin.Microbiol. 36, 1750–1755.
68. Souan, L., Tal, Y., Felling, Y., Cohen, I. R., Taraboulos, A., Mor, F.(2001) Modulation of proteinase-K resistant prion protein by prion pep-tide immunization. Eur. J. Immunol. 31, 2338–2346.
69. Koller, M. F., Grau, T., Christen, P. (2002) Induction of antibodiesagainst murine full-length prion protein in wild-type mice. J. Neuroim-munol. 132, 113–116.
70. Arbel, M., Lavie, V., Solomon, B. (2003) Generation of antibodies againstprion protein in wild-type mice via helix 1 peptide immunization. J. Neu-roimmunol. 144, 38–45.
71. Gilch, S., Wopfner, F., Renner-Muller, I., Kremmer, E., Bauer, C., Wolf,E., Brem, G., Groschup, M. H., Schatzl, H. M. (2003) Polyclonal anti-PrPauto-antibodies induced with dimeric PrP interfere efficiently with PrPScpropagation in prion-infected cells. J. Biol. Chem. 278, 18524–18531.
72. Stoltze, L., Rezaei, H., Jung, G., Grosclaude, J., Debey, P., Schild, H.,Rammensee, H. G. (2003) CD4� T cell-mediated immunity againstprion proteins. Cell. Mol. Life Sci. 60, 629–638.
73. Curin Serbec, V., Bresjanac, M., Popovic, M., Pretnar Hartman, K.,Galvani, V., Rupreht, R., Cernilec, M., Vranac, T., Hafner, I., Jerala, R.(2004) Monoclonal antibody against a peptide of human prion proteindiscriminates between Creutzfeldt-Jacob’s disease-affected and normalbrain tissue. J. Biol. Chem. 279, 3694–3698.
74. Gregoire, S., Logre, C., Metharom, P., Loing, E., Chomilier, J., Rosset,M. B., Aucouturier, P., Carnaud, C. (2004) Identification of two immu-nogenic domains of the prion protein—PrP—which activate class II-restricted T cells and elicit antibody responses against the native mole-cule. J. Leukoc. Biol. 76, 125–134.
75. Polymenidou, M., Heppner, F. L., Pellicioli, E. C., Urich, E., Miele, G.,Braun, N., Wopfner, F., Schatzl, H. M., Becher, B., Aguzzi, A. (2004)Humoral immune response to native eukaryotic prion protein correlateswith anti-prion protection. Proc. Natl. Acad. Sci. USA 101 (Suppl, 2),14670–14676.
76. Rosset, M. B., Ballerini, C., Gregoire, S., Metharom, P., Carnaud, C.,Aucouturier, P. (2004) Breaking immune tolerance to the prion proteinusing prion protein peptides plus oligodeoxynucleotide-CpG in mice.J. Immunol. 172, 5168–5174.
77. Gregoire, S., Bergot, A. S., Feraudet, C., Carnaud, C., Aucouturier, P.,Rosset, M. B. (2005) The murine B cell repertoire is severely selectedagainst endogenous cellular prion protein. J. Immunol. 175, 6443–6449.
78. Nikles, D., Bach, P., Boller, K., Merten, C. A., Montrasio, F., Heppner,F. L., Aguzzi, A., Cichutek, K., Kalinke, U., Buchholz, C. J. (2005)Circumventing tolerance to the prion protein (PrP): vaccination withPrP-displaying retrovirus particles induces humoral immune responsesagainst the native form of cellular PrP. J. Virol. 79, 4033–4042.
79. Xiao, X. L., Jiang, H. Y., Zhang, J., Han, J., Nie, K., Zhou, X. B., Huang,Y. X., Chen, L., Zhou, W., Zhang, B. Y., Liu, Y., Dong, X. P. (2005)Preparation of monoclonal antibodies against prion proteins with full-length hamster PrP. Biomed. Environ. Sci. 18, 273–280.
80. Andrievskaia, O., McRae, H., Elmgren, C., Huang, H., Balachandran, A.,Nielsen, K. (2006) Generation of antibodies against bovine receombinantprion protein in various strains of mice. Clin. Vaccine Immunol. 13,98–105.
81. Yamanaka, H., Ishibashi, D., Yamaguchi, N., Yoshikawa, D., Nakamura,R., Okimura, N., Arakawa, T., Tsuji, T., Katamine, S., Sakaguchi, S.(2006) Enhanced mucosal immunogenicity of prion protein followingfusion with B subunit of Eschericia coli heat-labile enterotoxin. Vaccine24, 2815–2823.
82. Blander, J. M., Medzhitov, R. (2004) Regulation of phagosome matura-tion by signals from Toll-like receptors. Science 304, 1014–1018.
83. Doyle, S. E., O’Connell, R. M., Miranda, G. A., Vaidya, S. A., Chow,E. K., Liu, P. T., Suzuki, S., Suzuki, N., Modlin, R. L., Yeh, W. C., Lane,T. F., Cheng, G. (2004) Toll-like receptors induce a phagocytic geneprogram through p38. J. Exp. Med. 199, 81–90.
Spinner et al. CpG DNA enhances humoral immunity to PrPSc in immunized mice 11

84. Blander, J. M., Medzhitov, R. (2006) Toll-dependent selection of micro-bial antigens for presentation by dendritic cells. Nature 440, 808–812.
85. Kabelitz, D., Wesch, D., Oberg, H. H. (2006) Regulation of regulatory Tcells: role of dendritic cells and Toll-like receptors. Crit. Rev. Immunol.26, 291–306.
86. Kraft, A. R., Arndt, T., Hasenkrug, K. J., Dittmer, U. (2005) Effectivetreatment of retrovirus-induced suppression of antibody responses withCpG oligodeoxynucleotides. J. Gen. Virol. 86, 3365–3368.
87. Branda, R. F., Moore, A. L., Lafayette, A. R., Mathews, L., Hong, R.,Zon, G., Brown, T., McCormack, J. J. (1996) Amplification of antibodyproduction by phosphorothioate oligodeoxynucleotides. J. Lab. Clin.Med. 128, 329–338.
88. Baek, K. H., Ha, S. J., Sung, Y. C. (2001) A novel function of phospho-rothioate oligodeoxynucleotides as chemoattractants for primary macro-phages. J. Immunol. 167, 2847–2854.
89. Vollmer, J., Janosch, A., Laucht, M., Ballas, Z. K., Schetter, C., Krieg,A. M. (2002) Highly immunostimulatory CpG-free oligodeoxynucleotidesfor activation of human leukocytes. Antisense Nucleic Acid Drug Dev. 12,165–175.
90. Vollmer, J., Weeratna, R. D., Jurk, M., Samulowitz, U., McCluskie, M. J.,Payette, P., Davis, H. L., Schetter, C., Krieg, A. M. (2004) Oligode-oxynucleotides lacking CpG dinucleotides mediate Toll-like receptor 9dependent T helper type 2 biased immune stimulation. Immunology113, 212–223.
91. de Almeida, C. J., Chiarini, L. B., da Silva, J. P., E. Silva, P. M., Martins,M.A., Linden, R. (2005) The cellular prion protein modulates phagocy-tosis and inflammatory response. J. Leukoc. Biol. 77, 238–246.
92. Holada, K., Vostal, J. G. (2000) Different levels of prion protein (PrPc)expression on hamster, mouse and human blood cells. Br. J. Haematol.110, 472–480.
93. Simak, J., Holada, K., D’Agnillo, F., Janota, J., Vostal, J. G. (2002)Cellular prion protein is expressed on endothelial cells and is releasedduring apoptosis on membrane microparticles found in human plasma.Transfusion 42, 334–342.
94. Li, Q., Cao, C., Chackerian, B., Schiller, J., Gordon, M., Ugen, K. E.,Morgan, D. (2004) Overcoming antigen masking of anti-amyloid� anti-bodies reveals breaking of B cell tolerance by virus-like particles inamyloid � immunized amyloid precursor protein transgenic mice. BMCNeurosci. 5, 21.
95. Heikenwalder, M., Polymenidou, M., Junt, T., Sigurdson, C., Wagner, H.,Akira, S., Zinkernagel, R., Aguzzi, A. (2004) Lymphoid follicle destruc-tion and immunosuppression after repeated CpG oligodeoxynucleotideadministration. Nat. Med. 10, 187–192.
96. Sethi, S., Ebner, S., Hinske, C., Kretzschmar, H. (2005) Multiple ad-ministrations of oligodeoxynucleotides containing CpG motifs influenceIg isotype production. Immunopharmacol. Immunotoxicol. 27, 447–460.
97. Lefeber, D. J., Benaissa-Trouw, B., Vliegenthart, J. F., Kamerling, J. P.,Jansen, W. T., Kraaijeveld, K., Snippe, H. (2003) Th1-directing adju-
vants increase the immunogenicity of oligosaccharide-protein conjugatevaccines related to Streptococcus pneumoniae type 3. Infect. Immun. 71,6915–6920.
98. Maurer, T., Heit, A., Hochrein, H., Ampenberger, F., O’Keeffe, M.,Bauer, S., Lipford, G. B., Vabulas, R. M., Wagner, H. (2002) CpG-DNAaided cross-presentation of soluble antigens by dendritic cells. Eur.J. Immunol. 32, 2356–2364.
99. Datta, S. K., Redecke, V., Prilliman, K. R., Takabayashi, K., Corr, M.,Tallant, T., DiDonato, J., Dziarski, R., Akira, S., Schoenberger, S. P.,Raz, E. (2003) A subset of Toll-like receptor ligands induces cross-presentation by bone marrow-derived dendritic cells. J. Immunol. 170,4102–4110.
100. Bevaart, L., Van Ojik, H. H., Sun, A. W., Sulahian, T. H., Leusen, J. H.,Weiner, G. J., Van De Winkel, J. G., Van Vugt, M. J. (2004) CpGoligodeoxynucleotides enhance Fc � RI-mediated cross presentation bydendritic cells. Int. Immunol. 16, 1091–1098.
101. Heit, A., Huster, K. M., Schmitz, F., Schiemann, M., Busch, D. H.,Wagner, H. (2004) CpG-DNA aided cross priming by cross-presenting Bcells. J. Immunol. 172, 1501–1507.
102. Suvas, S., Kumaraguru, U., Pack, C. D., Lee, S., Rouse, B. T. (2003)CD4�CD25� T cells regulate virus-specific primary and memoryCD8� T cell responses. J. Exp. Med. 198, 889–901.
103. Rao, M., Matyas, G. R., Vancott, T. C., Birx, D. L., Alving, C. R. (2004)Immunostimulatory CpG motifs induce CTL responses to HIV type Ioligomeric gp140 envelope protein. Immunol. Cell Biol. 82, 523–530.
104. Belyakov, I. M., Isakov, D., Zhu, Q., Dzutsev, A., Klinman, D., Berzofsky,J. A. (2006) Enhancement of CD8� T cell immunity in the lung by CpGoligodeoxynucleotides increases protective efficacy of a modified vac-cinia Ankara vaccine against lethal poxvirus infection even in a CD4-deficient host. J. Immunol. 177, 6336–6343.
105. Lewicki, H., Tishon, A., Homann, D., Mazarguil, H., Laval, F., Asensio,V. C., Campbell, I. L., DeArmond, S., Coon, B., Teng, C., Gairin, J. E.,Oldstone, M. B. (2003) T cells infiltrate the brain in murine and humantransmissible spongiform encephalopathies. J. Virol. 77, 3799–3808.
106. Kocisko, D. A., Vaillant, A., Lee, K. S., Arnold, K. M., Bertholet, N.,Race, R. E., Olsen, E. A., Juteau, J-M., Caughey, B. (2006) Potentantiscrapie activities of degenerate phosphorothioate oligonucleotides.Antimicrob. Agents Chemother. 50, 1034–1044.
107. Oberdieck, U., Xi, Y. G., Pocchiari, M., Diringer, H. (1994) Character-ization of antisera raised against species-specific peptide sequences fromscrapie-associated fibril protein and their application for post-mortemimmunodiagnosis of spongiform encephalopathies. Arch. Virol. 136,99–110.
108. Safar, J., Wille, H., Itri, V., Groth, D., Serban, H., Torchia, M., Cohen,F. E., Prusiner, S. B. (1998) Eight prion strains have PrP (Sc) moleculeswith different conformations. Nat. Med. 4, 1157–1165.
12 Journal of Leukocyte Biology Volume 81, June 2007 http://www.jleukbio.org





























