Embed Size (px)
Citation preview

Molecular Phylogenetics and Evolution 44 (2007) 42–52www.elsevier.com/locate/ympev
Cryptic intercontinental colonization in water Xeas Daphnia pulicaria inferred from phylogenetic analysis of mitochondrial DNA variation
Silvia Marková a,b,c,¤, France Dufresne b, David J. Rees b, Martin Berný a, Petr Kotlík c
a Department of Ecology, Faculty of Science, Charles University, Vinibná 7, CZ-128 44, Prague 2, Czech Republicb Département de Biologie, Université du Québec à Rimouski, Que., Canada G5L 3A1
c Department of Vertebrate Evolutionary Biology and Genetics, Institute of Animal Physiology and Genetics, Academy of Sciences of the Czech Republic, Rumburská 89, CZ-27721 Lib5chov, Czech Republic
Received 20 June 2006; revised 18 December 2006; accepted 20 December 2006Available online 9 January 2007
Abstract
The water Xeas of the Daphnia pulex complex play a key role in freshwater ecosystems throughout the northern hemisphere. Despitethe fact that they have been the subject of study for numerous biological disciplines, their phylogeny and species delimitation remain con-troversial. We used DNA sequence variation of the mitochondrial ND5 gene to reconstruct the phylogenetic relationships of D. pulicariaForbes, a widespread member of this complex from North America and Europe. Populations from the two continents respectively splitinto two evolutionary lineages, Eastern Nearctic and European, which each belong to another main clade within the D. pulex complex(the pulicaria and tenebrosa groups, respectively). Unexpectedly, melanin and carotenoid pigmented D. pulicaria populations from Euro-pean high-mountain lakes were not allied with the transparent populations inhabiting the same lakes and the lowland ponds and reser-voirs throughout Europe, but were included with the samples from Canada and Greenland in the Eastern Nearctic lineage. Until nowpopulations belonging to this lineage were known only from Canada and North Atlantic islands, but not from mainland Europe. Inde-pendent data from microsatellite markers supported the genetic distinctiveness of the sympatric carotenoid pigmented and transparentpopulations and suggested that they may have undergone transition to obligate parthenogenesis, possibly as a consequence of past intro-gressive hybridization. Two diVerent taxa are therefore confused under the name D. pulicaria in Europe. The close phylogenetic relation-ships of European populations with those from Canada and Greenland suggest that the Nearctic lineage is of recent origin in Europe viaintercontinental dispersal from the North America. It has evolved melanin and carotenoid pigmentation as adaptations against the UVlight stress, which enable it to share habitat occupied by the transparent European species. The Nearctic D. pulicaria thus provides a newmodel for studying successful intercontinental invasion. In general, our study demonstrates that a considerable part of the diversityamong widespread taxa of cladoceran crustaceans has been overlooked in morphological taxonomies.© 2007 Elsevier Inc. All rights reserved.
Keywords: Arctic; Colonization; Daphnia pulex complex; mtDNA; ND5
1. Introduction
Water Xeas (Crustacea: Cladocera) of the Daphnia pulexspecies complex (sensu Colbourne and Hebert, 1996) areubiquitous residents of inland waters throughout the tem-
* Corresponding author. Present address: Department of VertebrateEvolutionary Biology and Genetics, Institute of Animal Physiology andGenetics, Academy of Sciences of the Czech Republic, Rumburská 89, CZ-27721 Lib5chov, Czech Republic. Fax: +420 315 639 510.
E-mail address: [email protected] (S. Marková).
1055-7903/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.ympev.2006.12.025
perate and arctic regions of the northern hemisphere(Hobaek and Weider, 1999). Many characteristics makethese organisms perfect subjects for ecological and evolu-tionary research (Colbourne et al., 2005). As the primaryconsumers of algae and a food of Wsh, they are key mem-bers of freshwater ecosystems and serve as model speciesfor freshwater and general ecology (Peters and de Bernardi,1987; KalV, 2002). Largely because of the wide geographicdistribution, phenotypic variation and capability of sexualand asexual reproduction they have been eVectively used in

S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52 43
evolutionary studies of speciation and adaptation (Sch-wenk et al., 2004; Paland et al., 2005), and recently becomethe subject of genomic research (Colbourne et al., 2005).However, exploration of the full potential of this emergingmodel organism is hampered by poorly resolved evolution-ary relationships and species boundaries within theD. pulex complex (Colbourne et al., 1998).
Although in recent years this species complex hasreceived considerable attention from molecular phyloge-netic studies (Colbourne et al., 1998; Weider et al., 1999a,b;Paland et al., 2005), its species delimitation remains the sub-ject of controversy. Genetic lineages within D. pulex com-plex are very distinct, suggesting that speciation has beenproceeding since the Pliocene (Colbourne and Hebert,1996). However, the slow rate of morphological evolutionrelative to the underlying molecular divergence has causedconsiderable taxonomic confusion, leading to the presenceof geographically widespread taxa with negligible morpho-logical variation that show high levels of sequence diversity(Colbourne et al., 1998; Weider and Hobaek, 2003).
Daphnia pulicaria Forbes is one such taxon. The species,as currently recognized, has a circumarctic geographic range(Hrbábek, 1959). It is widely distributed across the temper-ate and arctic regions of North America (Benzie, 2005). InEurasia it is conWned to the west of the Ural Mountains,with a center in middle Europe and extending northwards toNorway and eastwards to western Russia (Hobaek andWeider, 1999). Similar to other water Xeas, D. pulicariashows geographic variation in the breeding system (Benzie,2005), with sexual cyclical parthenogenesis predominating inlarge parts of temperate North America and Europe (Bernýand Hebert, 1993; Hebert et al., 1993), and asexual obligateparthenogenesis dominating in the western part of its NorthAmerican range in the prairies, in the Arctic, and in moun-tain habitats in Europe (Berný and Hebert, 1993; Hebertet al., 1993, 2003; Weider et al., 1999a,b).
Recent phylogenetic studies of mitochondrial DNA(mtDNA) sequence (Colbourne et al., 1998) and restrictionfragment length polymorphism data (RFLP; Weider et al.,1999a) for the members of the D. pulex complex haveshown that D. pulicaria is not a monophyletic species. TheAmerican lineages of D. pulicaria belong to a diVerent mainclade of the D. pulex complex, the pulicaria group followingthe nomenclature devised by Colbourne et al. (1998), thenthe European populations, which are a part of the highlydivergent tenebrosa group (Colbourne et al., 1998). How-ever, these studies were primarily concerned with phyloge-netic relationships and diversity within the entire speciescomplex, and their sampling coverage of D. pulicaria intemperate regions of Europe was too limited to providemuch insight into its evolutionary history.
Daphnia pulicaria has been found in a wide range of hab-itats throughout Europe, ranging from lowland Wshpondsand reservoirs to glacial lakes in high mountains and shal-low ponds in the arctic regions. Three morphotypes basedon variation in photoprotective pigmentation but lackingother prominent morphological diVerences exist in neigh-
boring sympatry in Europe. A transparent (unpigmented)morphotype inhabits lowland regions throughout Europeand mountain lakes in the High Tatra (western part of theCarpathian Mts), an orange (carotenoid pigmented) mor-photype coexists with the transparent morphotypes in theHigh Tatra (Lityjski, 1917), and a black (melanic) morpho-type occurs in glacial lakes in the Pyrenees (Ventura andCatalan, 2005).
Although there is variation in pigmentation in otherDaphnia, genetic diVerentiation between the color morphshas not been demonstrated (Schwenk et al., 2004). Pigmen-tation plays a role in protection against harmful UV-radia-tion and predation pressure, and it has been shown to havea genetic basis in isopod crustaceans (Hargeby et al., 2004).As yet no study assessed genetic diVerentiation between themorphotypes of D. pulicaria. However, ecological studiesshowed that pigmentation morphotypes are diVerentiatedin terms of their life history characteristics. The blackmorphotype and the orange morphotype start new-yeargeneration in the spring from subitaneous eggs released byover-wintering females (Gliwicz et al., 2001; Ventura andCatalan, 2005), while in the transparent morphotype thenew-year generation was recruited from diapausing eggs,which hatched in summer (Haney and Buchanan, 1987;Gliwicz et al., 2001). This synchronized reproduction isespecially pronounced in syntopic populations of the trans-parent and orange morphotypes in the High Tatra (Gliwiczet al., 2001), and its correlation with pigmentation variationsuggests that these phenotypes may be associated withevolutionary subdivision (Tarjuelo et al., 2004).
In the present study, we analyze mtDNA sequence varia-tion of D. pulicaria in North America and Europe. We forthe Wrst time include populations belonging to all three pig-mentation morphotypes distinguished in Europe and occu-pying diVerent habitats, including lowland regionsthroughout Europe and alpine lakes in the High Tatra andPyrenees. As an independent line of evidence, we analyzelength polymorphism at nuclear-encoded microsatellite locifor sympatric populations of the transparent and orangemorphotypes from the High Tatra. Our objectives were toreconstruct phylogeny of D. pulicaria from diVerent partsof the distribution range and their relationships to otherlineages in the D. pulex complex and to clarify whether thephenotypic (pigmentation and ecological) subdivision ofD. pulicaria is related to genetic divergence.
2. Materials and methods
2.1. Sample collection and DNA extraction
Daphnia individuals were collected from 50 diVerentlocalities in Europe and North America (Table 1). WaterXeas were collected from lowland Wshponds and reservoirs,high-mountain lakes, and arctic ponds and rockpoolsby horizontal sweeps on the lakeshore or by plankton nettows from an inXatable boat in the lake center, and werestored in 95% ethanol at 4 °C until analysis. To complete

44 S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52
Table 1Samples of the Daphnia pulex complex used in this study
Taxa and collection localities Sequencea Data source
pulicaria groupPanarctic D. pulex
Churchill (Site 1), Manitoba, Canada DQ779137 This studyChurchill (Site 2), Manitoba, Canada DQ779139 This studyKuujjuarapik (Site 1), Nunavik, Canada DQ779122 This studyKuujjuarapik (Site 2), Nunavik, Canada DQ779131 This studyKuujjuarapik (Site 3), Nunavik, Canada DQ779123 This studyKuujjuarapik (Site 4), Nunavik, Canada DQ792505 This studyRimouski, Quebec, Canada DQ779125 This studyWindsdor (Site 1), Ontario, Canada DQ779135 This studyWindsdor (Site 2), Ontario, Canada DQ779136 This studyWindsdor (Site 3), Ontario, Canada DQ779132 This studyWindsdor (Site 4), Ontario, Canada DQ779133 This studyWindsdor (Site 5), Ontario, Canada DQ779134 This studyKachkovsky Bay, Western Siberia WSB 1 Colbourne et al. (1998)Tuktoyaktuk, Northwest Territoris, Canada NWT 3 Colbourne et al. (1998)
D. middendorYanaBylot Island (Site 1), Nunavut, Canada DQ779126 This studyBylot Island (Site 2), Nunavut, Canada DQ779127 This studyChurchill (Site 3), Manitoba, Canada DQ779138 This studySummerset Island, District of Franklin, Canada DOF 14 Colbourne et al. (1998)Tuktoyaktuk, Northwest Territoris, Canada NWT 2 Colbourne et al. (1998)
Eastern Neartic D. pulicariaKuujjuarapik (Site 5), Nunavik, Canada DQ779119 This studyKuujjuarapik (Site 6), Nunavik, Canada DQ779128 This studyKuujjuarapik (Site 7), Nunavik, Canada DQ779129 This studyKuujjuarapik (Site 8), Nunavik, Canada DQ779130 This studyKuujjuarapik (Site 9), Nunavik, Canada DQ779124 This studyEstat Lake, Pyrenees, Catalonia, Spain DQ779112 This studySottlo Lake, Pyrenees, Catalonia, Spain DQ779113 This studyRedon Lake, Pyrenees, Catalonia, Spain DQ779113 This studyVynné Temnosmrebinské Lake, High Tatra Mts., Slovakia DQ779098 This studyCzarny Staw pod Rysmi, High Tatra Mts., Poland DQ779097 This studyZelené Krivánske Lake, High Tatra Mountain, Slovakia DQ779097 This studyGodhavn, Greenland GRL 3 Colbourne et al. (1998)Redchalk Lake, Ontario, Canada ONT 1 Colbourne et al. (1998)
Western Neartic D. pulicariaKuujjuarapik (Site 10), Nunavik, Canada DQ779120 This studyKuujjuarapik (Site 11), Nunavik, Canada DQ779121 This studyRedberry Lake, Saskatchewan, Canada SAS 1 Colbourne et al. (1998)Washington Lake, Washington, USA WAS 1 Colbourne et al. (1998)
Polar Neartic D. pulicariaSummerset Island, District of Franklin, Canada DOF 8 Colbourne et al. (1998)Western Iceland ICE 1 Colbourne et al. (1998)
D. melanicaFlorence, Oregon, USA ORE 1 Colbourne et al. (1998)Florence, Oregon, USA ORE 3 Colbourne et al. (1998)
tenebrosa groupEuropean D. pulicaria
Czarny Staw pod Rysmi, High Tatra Mts., Poland DQ779101 This studyNiqné Temnosmrebinské Lake, High Tatra Mts., Slovakia DQ779101 This studyL’adové Lake, High Tatra Mts., Slovakia DQ779104 This studyLitvorové Lake, High Tatra Mts., Slovakia DQ779105 This studyMalé Hincovo Lake, High Tatra Mts, Slovakia DQ779099 This studyNiqné Labie Bielovodské Lake, High Tatra Mts., Slovakia DQ779103 This studyVel’ké Labie Javorové Lake, High Tatra Mts., Slovakia DQ779106 This studyVynné Labie Bielovodské Lake, High Tatra Mts., Slovakia DQ779102 This studyWelki Staw Polski, High Tatra Mts., Poland DQ779100 This studyZelené Javorové Lake, High Tatra Mts., Slovakia DQ779107 This studyBohdaneb reservoir, Czech Republic DQ779108 This study

S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52 45
the dataset and provide a wider phylogenetic frameworkfor our study we included 18 homologous sequences (Table1) described by Colbourne et al. (1998) to represent allmembers of the D. pulex complex analyzed by theseauthors, including six constituent clades of the pulicariagroup (termed eastern, western and polar lineage of Nearc-tic D. pulicaria, Panarctic D. pulex, D. middendorYana andD. melanica) and D. tenebrosa of the tenebrosa group.Sequences of European D. pulex were used to root the treesbecause they form a third clade (the pulex group) within theD. pulex complex, sister to the tenebrosa and pulicariagroups, and represent an outgroup to the other lineages(Colbourne et al., 1998; Hobaek and Weider, 1999).
DNA was extracted by incubating single animals in 30�lof a 6% (w/v) suspension of Chelex 100 resin (Bio-Rad,Hercules, CA) at 55 °C for 3 h and then boiling for 9 minfollowed by cooling at room temperature, centrifuging at14,000 rpm for 2 min, and cooling at 4 °C overnight.
2.2. Mitochondrial DNA sequencing
A fragment (»850 bp) including part of the gene codingfor the NADH dehydrogenase subunit 5 (ND5) was ampli-Wed by PCR following methods described in Colbourneet al. (1998) and Weider et al. (1999a) and using primersDpuND5a and DpuND5b (Colbourne et al., 1998; Weiderand Hobaek, 2003). The resulting PCR products were puri-Wed with the QIAquick PCR PuriWcation Kit (Qiagen) andboth strands were directly sequenced with the same primersas those used for the ampliWcation and the Dye TerminatorQuick Start Kit for cycle-sequencing (Beckman Coulter,USA). The extension products were run on a BeckmanCoulter CEQ 8000 automated capillary sequencer.
Sequences were aligned manually to each other and to thesequences of the D. pulex complex described by Colbourneet al. (1998). All polymorphisms were additionally checkedby visual inspection of automated sequencer chromato-grams.
2.3. Microsatellite genotyping
Sixty-four individuals of the transparent morphotypeand 29 individuals of the orange morphotype of D. puli-caria from the High Tatra, where these morphotypes occurin sympatry, were genotyped at Wve microsatellite locidescribed by Colbourne et al. (2004). The 13 sampled lakesincluded those used in mtDNA analysis (Table 1) and rep-resented all known D. pulicaria populations in the HighTatra (Marková et al., 2006). Three microsatellite loci con-sisted of dinucleotide motifs (Dp512, Dp522, Dp523) andtwo loci (Dp514, Dp514alt) contained trinucleotide repeats.Forward primers were end-labeled with Xuorescent dyesFAM or TET (Life Technologies, Univ. of Oklahoma,USA). Conditions for PCR were as described in Colbourneet al. (2004). PCR products were electrophoresed on 6%polyacrylamide gels and scanned with a FMBio III scanner(Hitachi, Tokyo, Japan). Allele sizes were assigned using aFluorescent Ladder (Promega, USA).
2.4. Phylogenetic analyses
Estimates of nucleotide divergence between mtDNAsequences were calculated with the PAUP¤ softwarepackage, version 4.0b10 (SwoVord, 2003), using a correcteddistance based on the gamma-HKY model of sequenceevolution, which accounts for unequal nucleotide
Table 1 (continued)
ClassiWcation and terminology follows those established by Colbourne et al. (1998) and later applied by e.g., Weider et al. (1999a,b).a Sequences described by Colbourne et al. (1998), unavailable from GenBank, have been obtained from the authors and are labeled in accordance with
their study.
Taxa and collection localities Sequencea Data source
Chmelnice pond, Czech Republic DQ779109 This studyMalá Kun pond, Czech Republic DQ779110 This studyIímov reservoir, Czech Republic DQ779111 This studySlapy reservoir, Czech Republic DQ779111 This studyEmbalse Lake, San Rafael de Matallana, Spain DQ779114 This studyHortobágy pond, Hungary DQ779111 This studyKonstanz Lake, Germany DQ779111 This studyOhrid Lake, Albania DQ779115, DQ779116 This studyPetchora Delta, Western Siberia, Russia DQ779118 This studyQueen Elisabeth reservoir, London, England DQ779117 This studySt. Nolth reservoir, London, England DQ779117 This study
D. tenebrosaBelyi Island, Western Siberia WSB8 Colbourne et al. (1998)Churchill, Manitoba,Canada MAN 2 Colbourne et al. (1998)Kolyma Delta, Estern Siberia ESB9 Colbourne et al. (1998)Kolyma Delta, Estern Siberia ESB 1 Colbourne et al. (1998)
pulex groupEuropean D. pulex
Basel, Switzerland SWI 5 Colbourne et al. (1998)Basel, Switzerland SWI 3 Colbourne et al. (1998)

46 S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52
frequencies and excess transitions, and models among-siterate heterogeneity in substitution rate by the gamma distri-bution (Hasegawa et al., 1985). The gamma-HKY modelwas determined to be the appropriate model for ourdataset by the hierarchical likelihood ratio test of good-ness-of-Wt of 56 diVerent nested models to the data, asimplemented in MODELTEST program, version 3.7(Posada and Crandall, 1998).
The phylogenetic relationships among the sequenceswere reconstructed using PAUP¤ by analysis of the distancematrix by the neighbor-joining (NJ) algorithm and by usingthe maximum-parsimony (MP; heuristic search) optimalitycriterion. Phylogenetic analyses using the maximum-likeli-hood (ML) criterion were performed with the algorithmimplemented in the PHYML software, version 2.4.4, thatsimultaneously adjusts tree topology and branch lengths tomaximize tree likelihood (Guindon and Gascuel, 2003), andusing the gamma-HKY evolutionary model with the fol-lowing base frequencies: AD0.20, CD0.20, GD0.22, andTD 0.39, a transition/transversion ratio of 5.69, and agamma distribution shape parameter � equaling to 0.33. Toquantify the conWdence in the partitioning within the MLtree we performed the nonparametric bootstrap test asapplied to phylogeny by Felsenstein (1985) using 1000 rep-lications.
Bayesian phylogenetic analysis was performed withMrBayes program, version 3.1.1 (Hüelsenbeck and Ron-quist, 2001; Ronquist and Hüelsenbeck, 2003), assumingthe gamma-HKY model of sequence evolution (two sub-stitution types with gamma-distributed rates across sites)and Xat Dirichlet distributions of prior probability densi-ties on the substitution rates and nucleotide frequencies,the uniform prior distributions on the shape parameterof the gamma distribution of rate variation and on thetree topology, and the unconstrained exponential priordistribution on the branch lengths. The analysis was doneusing several independent runs starting from diVerentrandom trees, each with 4 Markov chains under theMetropolis coupling to improve mixing. Each chain wasinitiated with a burn-in period of 2500 updates and thetotal length of each analysis was 10 million updates withtrees sampled every 100 generations. The analysis wasconsidered to have converged upon a stationary distribu-tion if the independent runs generated similar posteriordistributions.
To compare competing phylogenetic hypotheses, we per-formed the test of the statistical signiWcance of diVerencesin log-likelihoods between topologies as proposed by Shi-modaira and Hasegawa (1999; SH test), which allows forsimultaneous comparison of multiple topologies and inclu-sion of the ML tree (Goldman et al., 2000; Shimodaira andHasegawa, 1999). We used the PAUP¤ implementation ofthe SH test, which generates the test distribution by a non-parametric bootstrap resampling of the sitewise log-likeli-hoods (the RELL approximation). Using the SH test, weexamined the monophyly of European D. pulicaria bytaking the ML tree estimated under the gamma-HKY
model and evaluating it against an alternative ML topologyestimated under the constraint of the European sequencesof D. pulicaria forming a clade.
Evolutionary relationships among the haplotypes withinthe Eastern Neartic D. pulicaria lineage were reconstructedusing the maximum parsimony criterion and the algorithmimplemented in the TCS program, version 1.21 (Templetonet al., 1992; Clement et al., 2000).
2.5. Microsatellite data analyses
We estimated pairwise genetic distances among the mul-tilocus genotypes using the Allele Shared Distance (DAS;Jin and Chakraborty, 1994) which counts the proportion ofshared alleles between the genotypes, and the Cavalli-Sforza’s Chord Distance (DSE; Cavalli-Sforza andEdwards, 1967) which is an Euclidean distance assumingdivergence in allele frequencies by genetic drift. Genetic dis-tances were calculated with Populations, version 1.2.28(Langella, 2003), and were used to construct unrooted NJtrees (Saitou and Nei, 1987) using PAUP¤. To quantify theconWdence in the partitioning within the trees we performedthe nonparametric bootstrap test, calculated between indi-viduals, with 1000 replications using MSAnalyzer, version2.39 (Dieringer and Schlötterer, 2003).
3. Results
3.1. Sequence variation
A nucleotide sequence of 672 bp of the ND5 gene wasobtained for each individual, which translated to 223 aminoacids. In the complete dataset, there were 217 variable char-acters, of which 177 were phylogenetically informative andrevealed 62 distinct haplotypes. Third codon positionsaccounted for most of the variation with 66.4% of variablesites located at third position, 21% at Wrst position, and12.6% at second position. Because of the absence of stopcodons and of indels, we considered it likely that thesequences represented a part of the functional mtDNAND5 gene, rather then a nuclear pseudogene (Zhang andHewitt, 1996).
3.2. Phylogenetic reconstruction
A Wfty-percentage majority-rule consensus of trees sam-pled from the posterior distribution in the Bayesian analy-sis showing posterior probabilities and ML bootstrapvalues for internal branches is presented in Fig. 1. Phyloge-netic reconstructions obtained with the ML, NJ and MPapproaches showed very similar topologies to the Bayesiantree and they consistently recovered all nine evolutionarylineages within the D. pulex complex, which fell into threemajor clades (pulicaria, tenebrosa and pulex). These groupscorroborate those previously identiWed by Colbourne et al.(1998) and Weider et al. (1999a) with a limited samplingin Europe. The haplotypes from North America were
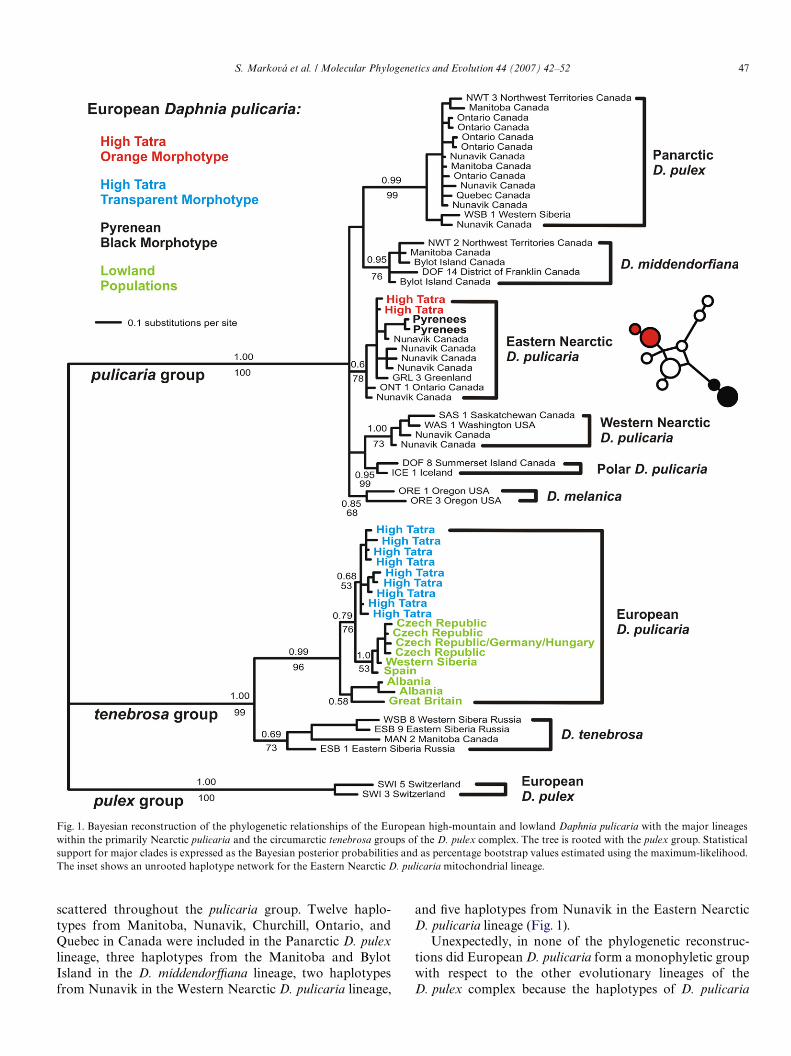
S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52 47
scattered throughout the pulicaria group. Twelve haplo-types from Manitoba, Nunavik, Churchill, Ontario, andQuebec in Canada were included in the Panarctic D. pulexlineage, three haplotypes from the Manitoba and BylotIsland in the D. middendorYana lineage, two haplotypesfrom Nunavik in the Western Nearctic D. pulicaria lineage,
and Wve haplotypes from Nunavik in the Eastern NearcticD. pulicaria lineage (Fig. 1).
Unexpectedly, in none of the phylogenetic reconstruc-tions did European D. pulicaria form a monophyletic groupwith respect to the other evolutionary lineages of theD. pulex complex because the haplotypes of D. pulicaria
Fig. 1. Bayesian reconstruction of the phylogenetic relationships of the European high-mountain and lowland Daphnia pulicaria with the major lineageswithin the primarily Nearctic pulicaria and the circumarctic tenebrosa groups of the D. pulex complex. The tree is rooted with the pulex group. Statisticalsupport for major clades is expressed as the Bayesian posterior probabilities and as percentage bootstrap values estimated using the maximum-likelihood.The inset shows an unrooted haplotype network for the Eastern Nearctic D. pulicaria mitochondrial lineage.

48 S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52
sampled in Europe were clustered within the tenebrosagroup as well as in the pulicaria group (Figs. 1 and 2). Thetransparent morphotype individuals from the High Tatrawere included in a well-supported clade with the individualssampled from the European lowland populations (meansequence divergence of 2.1%) and together they constitutedthe European D. pulicaria lineage within the tenebrosagroup (Fig. 1). The orange morphotype individuals fromthe High Tatra and black morphotype individuals from thePyrenees were phylogenetically closely related to each other(mean sequence divergence of 1.4%). However, they werehighly divergent from the High Tatra transparent morpho-type and from the lowland populations (mean sequencedivergence of 30.7–32.2%), and were unambiguously identi-Wed as members of the predominantly North Americanpulicaria group. Together with the sequences from Nuna-vik, Ontario, and Greenland (mean sequence divergence of
Fig. 2. Geographic distribution of the Eastern Neartic Daphnia pulicarialineage (pulicaria group) and the European D. pulicaria lineage (tenebrosagroup) based on the results of the present study and of Colbourne et al.(1998) and Weider et al. (1999a).
0.7–1.2%) they were included in the Eastern Nearctic D.pulicaria lineage of this group (Fig. 1). Furthermore,sequences within this lineage were phylogenetically closelyrelated to each other, with the European populations notforming a monophyletic haplotype clade with respect toNorth American and Greenland populations, and this rela-tionship was supported also by the TCS analysis (Fig. 1).
The tree partitions that rendered the European D. puli-caria polyphyletic received high posterior probability in theBayesian analyses and were well-supported by the boot-strap test (Fig. 1). The SH likelihood test indicated that thetopology where the European D. pulicaria were constrainedto form a monophyletic group (¡Ln likelihoodD 2928.3883) was signiWcantly less likely given our data (one-tailedtest, P < 0.05) than the unconstrained ML topology (¡LnlikelihoodD 2836.3344).
3.3. Microsatellite analyses
In total, 18 alleles were detected at the Wve microsatelliteloci assessed in 93 individuals of the transparent andorange morphotypes of D. pulicaria from the High Tatra.All individuals were heterozygous at the loci that showedmore than one allele in that morphotype (Table 2), support-ing the obligate parthenogenesis as the major mode ofreproduction (Berný and Hebert, 1993). We never observedmore than two alleles at locus, thus strongly suggesting thatall the individuals are diploid. This is consistent with thefact that polyploid Daphnia are restricted to artic regions(Dufresne and Hebert, 1997). Altogether six multilocusgenotypes were resolved (Table 2), four (A–D) in the trans-parent morphotype belonging to the European lineage andtwo (E and F) in the orange morphotype belonging to theEastern Nearctic lineage (Table 2). Although the morpho-types shared an allele at four of the Wve loci, no multilocusgenotype was shared between the transparent morphotypeand the orange morphotype, and 14 morphotype-speciWcprivate alleles were found, with eight private alleles of thetotal 12 (66.6%) in the transparent morphotype, and six of10 (60.0%) in orange morphotype (Table 2). Neighbor-join-ing trees constructed using DAS and DSE genetic distancesshowed identical topologies, which revealed signiWcant
Table 2Alleles detected at Wve microsatellite loci screened for variation in Daphnia pulicaria from the High Tatra and their distribution among the six multilocusgenotypes (A–F)
Numbers indicate sizes of alleles (in base pairs). x D allele was present in the genotype.
Genotype Dp512 Dp514alt Dp522 Dp523 Dp514
128 130 138 133 148 151 169 175 187 116 120 122 128 134 136 99 101 102
Transparent morphotype/European lineageA x x x x x x x x xB x x x x x x x x xC x x x x x x x x xD x x x x x x x x x
Orange morphotype/Eastern Neartic lineageE x x x x x x x x xF x x x x x x x x x

S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52 49
grouping among the multilocus genotypes with all multilo-cus genotypes of each lineage more closely related to eachother than to any multilocus genotype of the other lineage(Fig. 3).
4. Discussion
Our analyses of mtDNA variation within D. pulicariaprovide clear evidence that the European populationsbelong to two major phylogenetic groups of the D. pulexcomplex. The posterior probability distributions, nonpara-metric bootstrap test and the likelihood-based SH test con-sistently showed that the populations belonging to thetenebrosa group do not form a clade with those included inthe pulicaria group.
A widespread survey of mtDNA variation in the D.pulex complex demonstrated that pulicaria group lineageswere restricted primarily to the Nearctic, with some limitedcolonization at the northernmost fringe of Europe, whilelineages of the tenebrosa group were widespread across thecircumarctic (Colbourne et al., 1998; Weider et al., 1999a).The two groups were known to overlap only in arcticEurope and western arctic Canada (Weider et al., 1999a).Europe was thus considered to be occupied primarily bythe tenebrosa group with the European D. pulicaria lineagewidely distributed throughout the temperate zone fromcentral Europe to western Russia, and the D. tenebrosa line-age in the European arctic (Hobaek and Weider, 1999).
Consistent with this view, all our European lowlandsamples from Great Britain and Spain across centralEurope into the Balkans belonged to the European D. puli-caria lineage. High-mountain populations of the transpar-ent morphotype from the High Tatra were also included inthis lineage. However, the High Tatra populations of theorange morphotype and the Pyrenean black morphotypepopulations were not allied with the tenebrosa group butwere included in the pulicaria group. Until now this lineagewas not found on mainland Europe. It was known onlyfrom Canada and Greenland, with a few representativesfurther east on Iceland and the arctic archipelago Svalbard(Weider et al., 1999a; Hobaek and Weider, 1999). Ourresults instead demonstrate that the distribution of the
Fig. 3. Unrooted neighbor-joining tree inferred from microsatellite datausing Cavalli-Sforza’s Chord Distance (DSE), illustrating relationshipsbetween the orange morphotype of the Eastern Nearctic lineage and thetransparent morphotype of the European lineage from the High Tatra.The tree is shown rooted by the mid-point to improve clarity, branchlengths are proportional to the estimated distance, and tree topology isidentical with that based on the Allele Shared Distance (DAS; not shown).Bootstrap support is 100% for the internal branch based on 1000 repli-cates.
Eastern Nearctic lineage extends much further south, beingpresent not only on the Arctic islands but also at muchlower latitudes in the Western and Central Europeanmountains (Fig. 2).
The pulicaria and tenebrosa groups represent clearly sep-arated ancient evolutionary lineages with the time of diver-gence in the Pliocene (5–2.2 million years ago; Colbourneet al., 1998). The results of our study show that the geo-graphic distributions of these two groups overlap not onlyin the European Arctic but also in European mountainregions. The close relationships of haplotypes from theEuropean populations with haplotypes from Canada andGreenland (Fig. 1) suggest that the Eastern NearcticD. pulicaria lineage is of recent origin in Europe. Phylogeo-graphic studies by Weider and Hobaek (1997) and Weideret al. (1999a) showed that another two lineages of theD. pulex complex colonized Iceland and Svalbard acrossGreenland from the high Nearctic shortly after the last gla-cial retreat. It is therefore possible that also the EasternNearctic lineage dispersed across the North Atlantic andfurther southwards towards mainland Europe. Its distribu-tion in Europe after deglaciation thus could have beenwider than at present and only following the Holocenewarming it retreated northwards and into high altitudes. Asimilar scenario has been suggested to explain arctic–alpinedistribution of other species in Europe (Abbott and Broch-mann, 2003; Schönswetter et al., 2006; Skrede et al., 2006).As another possibility we suggest that the Eastern NearcticD. pulicaria may have crossed the Atlantic several timesand via diVerent routes. European populations of the East-ern Nearctic lineage do not form a monophyletic haplotypeclade with respect to North American populations (Fig. 1),which appears to support such a scenario. The dispersalcapabilities of water Xeas are greatly enhanced by the pas-sive transport of resistant diapausing eggs that are encasedin hard capsules (ephippia), and new populations can beestablished from ephippia that disperse over long distancesvia vectors such as migratory birds, wind and ocean cur-rents (Weider et al., 1999a,b; Havel and Medley, 2006). Wetherefore hypothesize that the colonization of mainlandEurope by the Eastern Nearctic lineage may be the result ofa direct trans-Atlantic transport from Eastern NorthAmerica rather then a southward dispersal from the Arctic.The RFLP data of Weider et al. (1999a) showed that Sval-bard is occupied by a distinct mtDNA haplotype lineagefrom those found in Greenland and Iceland, which suggestsa direct transfer from the Canadian arctic and supports theview that a long-distance dispersal from the Nearctic wasnot a unique event. Although our present data are incon-clusive in this respect, they show that the colonization ofEurope by the Nearctic D. pulicaria is recent and the foun-der populations have not had suYcient time to phylogenet-ically diverge from their source populations in NorthAmerica. Recent molecular evidence for postglacial trans-Atlantic migrations has also been provided for several Arc-tic plant species occurring on both sides of the Atlantic(Abbott and Brochmann, 2003). Consequently, the Atlantic

50 S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52
Ocean does not appear to have been an impenetrable dis-persal barrier for the freshwater and terrestrial organismsthat are passively transported as resistant propagules (eggs,seeds). Passive long-distance dispersal has been proposed asan explanation also for disjunct distributions of Daphniaspecies between North America and South America(Adamowicz et al., 2004). Recently, several instances ofintercontinental introductions have been documented ofexotic cladoceran species or subspecies to Africa (Mergeayet al., 2005) and North America (Taylor et al., 1998; Haveland Medley, 2006). Daphnia’s enormous dispersal potentialtherefore raises the intriguing possibility that some geneexchange between the two continents may have occurreduntil very recently in D. pulicaria and contributed againstthe population divergence. Further study with more exten-sive sampling should yield additional insights about the ori-gin of the Nearctic lineage in Europe and the timescale ofthe genetic exchange between the continents.
The Eastern Nearctic lineage and European lineagecoexist in the High Tatra where they even inhabit thesame lakes (Table 1). We performed microsatellite analy-sis as an independent source of molecular data to evaluatethe degree of genetic diVerentiation between the two lin-eages in this area of sympatric occurrence. Our resultsdemonstrated that the orange morphotype belonging tothe Eastern Nearctic lineage and the transparent morpho-type belonging to the European lineage shared no geno-types at Wve polymorphic microsatellite loci, and althoughthey shared alleles at some loci, they possessed a high pro-portion of alleles unique to each lineage and were wellseparated in the NJ trees (Fig. 3). Our results thus demon-strate that the two mtDNA lineages are also representedby diVerent genotypes at nuclear-encoded microsatelliteloci, with clones from each lineage sharing ancestry witheach other.
It has been shown that pigmentation allows Daphnia tostay closer to the surface under UV-radiation in contrastto unpigmented individuals that seek a refuge in deeperparts of the lake (Rhode et al., 2001). Furthermore, thework conducted by Gliwicz et al. (2001) in the High Tatrashowed that this genetic subdivision into two main lin-eages is coupled with life history divergence. Althoughboth lineages produce single generation each year, theirreproduction is temporarily separated. In the Nearcticlineage the new generation starts in spring primarily fromsubitaneous eggs released by over-wintering females,while in the European lineage it is recruited from diapaus-ing eggs that hatch in summer, which reduces resourcecompetition between the lineages (Gliwicz et al., 2001).Therefore, the ability to develop photoprotective pigmen-tation and the synchronized reproduction may representimportant adaptations that enabled the Nearctic speciesto share habitats occupied by the unpigmented Europeanspecies, and which facilitated its colonization of the Euro-pean continent.
The Wxed heterozygosity at microsatellite loci suggeststhat the High Tatra populations are composed of obligate
parthenogenetic clones (Berný and Hebert, 1993). Multipleclones from the Eastern Nearctic lineage may have there-fore colonized Europe, followed by interclonal selection fortraits facilitating the coexistence with the European lineage.However, because sexual cyclical parthenogenesis is thepredominant breeding system in large parts of NorthAmerica (Berný and Hebert, 1993; Hebert et al., 1993), it ispossible that the transition to obligate parthenogenesisoccurred after the Eastern Nearctic lineage arrived inEurope and selection for ecological and pigmentationdiVerentiation had occurred in the sexual population. Thefact that orange and transparent morphotypes share micro-satellite alleles at several loci suggests that some clones mayhave been formed by introgression between the EasternNearctic and European lineage, which could contribute toecological diVerences between the parent and hybrid taxa(Taylor et al., 2005). As there is no evidence of EuropeanmtDNA in the pigmented clones, backcrossing would haveinvolved male hybrids with female of the Eastern Nearcticlineage. Phylogeographic sequence analysis of nuclearmarkers should yield further insights about the role ofintrogressive hybridization in the ecological diVerentiationof D. pulicaria in the European mountains.
The fact that two species are confused under the nameDaphnia pulicaria Forbes in Europe has been overlooked inprevious studies, likely owing to the lack of prominent mor-phological diVerences other than pigmentation. Lityjski(1917) has noted occurrence of orange and transparentcolor morphs in the High Tatra, and Gliwicz et al. (2001)described from the same lake diVerent life history strategies,which he later matched with the two color morphs (Z.M.Gliwicz, personal communication). However, these pheno-types have been interpreted as a variation of a single speciesand attributed to environmental factors rather then geneticdivergence (Lityjski, 1917; Gliwicz et al., 2001). Our datainstead suggest that European populations belong to twophylogenetic lineages with a history of deep divergence andrecent ecological diVerentiation, possibly coupled withintrogressive hybridization.
Apparently, a considerable part of Daphnia diversityhas been missed or attributed to ecophenotypy by mor-phological taxonomies. Together with recent Wndings inNorth America (Taylor et al., 1998; Hebert et al., 2003)and arctic Europe (Schwenk et al., 2004), our study showsthat widespread and morphologically uniform Daphniaspecies are likely to harbor cryptic evolutionary subdivi-sions. Daphnia pulicaria Forbes was originally describedfrom North America and morphologically similar Daphniafrom Europe were later ascribed to it (Hrbábek, 1959;Benzie, 2005). Recent molecular phylogenetic studies dem-onstrated that the European and North American popula-tions of D. pulicaria belong to separate taxa (Berný andHebert, 1999; Hobaek and Weider, 1999). In the presentstudy, we have taken these results further and we haveshown that the current recognition of a single species inEurope is erroneous and that two taxa should be recog-nized instead, one of which appears conspeciWc with North

S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52 51
American D. pulicaria, while the other likely represents anundescribed species. A detailed taxonomic description willbe presented elsewhere. Clearly, a thorough taxonomicrevision of the species within the D. pulex complex isneeded, following the evaluation of morphological charac-ters and examination of the name-bearing types (Benzie,2005). These results have important implications for theuse of Daphnia as model organisms.
Acknowledgments
We are grateful to Spase Shumka, Litza Michaloudi,Ioanna Salvarina, Ivan Traykov, Zuzana Burdíková, PavolMarko, Marina Manca, Lorenzo Marchi, Marco Simona,Mauro Veronesi and Marco Costamagna, for their com-pany and hospitality during our collection trips to Albania,Greece and Bulgaria, Italy, Slovakia and Switzerland. Wethank Marc Ventura, Jilí Machábek and Adam Petrusekwho sent us Daphnia samples, and Catherine Simard forlaboratory assistance. Macej Gliwicz, Jozef Hrbábek, Chris-tian Jersabek, Vladimír Kolínek and Mirek Slusarczyk pro-vided helpful information about alpine populations ofDaphnia pulicaria. We thank Anders Hobaek, Piet Spaakand Vendula Klechtová for their valuable comments andsuggestions. We acknowledge John Colbourne who pro-vided sequences used in Colbourne et al. (1998) unavailablefrom GenBank. This study was supported by grants toFrance Dufresne from the Natural Sciences and Engineer-ing Research Council of Canada and the Fonds Québécoisde la Recherche sur la Nature et les technologies, and thestudent grant to Silvia Marková from the Grant Agency ofCharles University (Grant No. 197/2004–2005). Additionalsupport to Silvia Marková and Petr Kotlík was providedby the Academy of Sciences of the Czech Republic (GrantNo. IRP IAPG AV0Z 50450515), and to Martin Berný bythe Ministry of Education, Youth and Sport of the CzechRepublic (Grant No. MSM 0021620828). We thank theNorthern ScientiWc Training Grants Program and the Cen-tre d’Études Nordiques for allowing sampling in Kuujju-arapik and Churchill.
References
Abbott, R.J., Brochmann, C., 2003. History and evolution of the arcticXora: in the footsteps of Eric Hultén. Mol. Ecol. 12, 299–313.
Adamowicz, S.J., Hebert, P.D.N., Marinone, M.C., 2004. Species diversityand endemism in the Daphnia of Argentina: a genetic investigation.Zool. J. Linn. Soc. 140, 171–205.
Benzie, J.A.H., 2005. Cladocera: The Genus Daphnia (including Daphniop-sis). Backhuys, Leiden.
Cavalli-Sforza, L.L., Edwards, A.W.F., 1967. Phylogenetic analysis: modelsand estimation procedures. Evolution 21, 550–570.
Berný, M., Hebert, P.D.N., 1993. Genetic diversity and breeding systemvariation in Daphnia pulicari from North American lakes. Heredity 71,497–507.
Berný, M., Hebert, P.D.N., 1999. Intercontinental allozyme diVerentiationamong four holarctic Daphnia species. Limnol. Oceanogr. 44, 1381–1387.
Clement, M., Posada, D., Crandall, K.A., 2000. TCS: a computer programto estimate gene genealogies. Mol. Ecol. 9, 1657–1660.
Colbourne, J.K., Hebert, P.D.N., 1996. The systematics of North AmericanDaphnia (Crustacea: Anomopoda): a molecular phylogeneticapproach. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 351, 349–360.
Colbourne, J.K., Crease, T.J., Weider, L.J., Hebert, P.D.N., Dufresne, F.,Hobaek, A., 1998. Phylogenetics and evolution of a circumarcticspecies complex (Cladocera: Daphnia pulex). Biol. J. Linn. Soc. 65,347–365.
Colbourne, J.K., Robison, B., Bogart, K., Lynch, M., 2004. Five hundredand twenty-eight microsatellite markers for ecological genomic investi-gations using Daphnia. Mol. Ecol. Notes 4, 485–490.
Colbourne, J.K., Singan, V.R., Gilbert, D.G., 2005. wFleaBase: the Daph-nia genome database. BMC Bioinformatics 6, 45.
Dieringer, D., Schlötterer, C., 2003. Two distinct modes of microsatellitemutation processes: evidence from the complete genomic sequences ofnine species. Genome. Res. 13, 2242–2251.
Dufresne, F., Hebert, P.D.N., 1997. Pleistocene glaciations and polyphy-letic origins of polyploidy in an arctic cladoceran. Proc. R. Soc. Lond.B. 264, 201–206.
Felsenstein, J., 1985. ConWdence-limits on phylogenies—an approachusing the bootstrap. Evolution 39, 783–791.
Gliwicz, Z.M., Slusarczyk, A., Slusarczyk, M., 2001. Life history synchroni-zation in a long-lifespan single-cohort Daphnia population in a Wshlessalpine lake. Oecologia 128, 368–378.
Goldman, N., Anderson, J.P., Rodrigo, A.G., 2000. Likelihood-based testsof topologies in phylogenetics. Syst. Biol. 49, 652–670.
Guindon, S., Gascuel, O., 2003. A simple, fast, and accurate algorithm toestimate large phylogenies by maximum likelihood. Syst. Biol. 52,696–704.
Haney, J.F., Buchanan, C., 1987. Distribution and biogeography of Daph-nia in the arctic. Memorie dell’Instituto Italiano di Idrobiologia 45,77–105.
Hargeby, A., Johansson, J., Ahnesjö, J., 2004. Habitat-speciWc pigmenta-tion in a freshwater isopod: adaptive evolution over a small spatiotem-poral scale. Evolution 58, 81–94.
Hasegawa, M., Kishino, H., Yano, T., 1985. Dating the human-ape split-ting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 22,160–174.
Havel, J.E., Medley, K.A., 2006. Biological invasions across spatial scales:intercontinental, regional, and local dispersal of cladoceran zooplank-ton. Biological Invasions 8, 459–473.
Hebert, P.D.N., Schwartz, S.S., Ward, R.D., Finston, T.L., 1993. Macrogeo-graphic patterns of breeding system diversity in the Daphnia pulexgroup. I. Breeding systems of Canadian populations. Heredity 70,148–161.
Hebert, P.D.N., Witt, J.D.S., Adamowicz, S.J., 2003. Phylogeographicalpatterning in Daphnia ambigua: regional divergence and intercontinetalcohesion. Limnol. Oceanogr. 48, 261–268.
Hobaek, A., Weider, L.J., 1999. A circumpolar study of arctic biodiversity:phylogeographic patterns in the Daphnia pulex complex. Ambio 28,245–250.
Hrbábek, J., 1959. Über die angelibliche Variabilität von Daphnia pulexLeydig. Zool. Anz. 162, 116–126.
Hüelsenbeck, J.P., Ronquist, F., 2001. MrBayes: Bayesian inference ofphylogenetic trees. Bioinformatics 17, 754–755.
Jin, L., Chakraborty, R., 1994. Estimation of genetic distance and coeY-cient of gene diversity from single-probe multilocus DNA Wngerprint-ing data. Mol. Biol. Evol. 11, 120–127.
KalV, J., 2002. Limnology. Inland Water Ecosystems. Prentice Hall, UpperSaddle River, NJ USA.
Langella, O., 2003. Populations. Population Genetic Software (Individualsor Populations Distances, Phylogenetic Trees). Version 1.2.28. CentreNational de la Recherche ScientiWque. Paris. Available from: <http://www.pge.cnrs–gif.fr/bioinfo/populations>.
Lityjski, A., 1917. Jeziora tatrzajskie i zamieszkujaca je fauna wiomlavek.Sprawozdanie Komisyi WzyograWcznej Akademii Umiejetnomci wKrakowie 51, 1–88.
Marková, S., Berný, M., Rees, D.J., Stuchlík, E., 2006. Are they still viable?Physical conditions and abundance of Daphnia pulicaria resting eggs in

52 S. Marková et al. / Molecular Phylogenetics and Evolution 44 (2007) 42–52
sediment cores from lakes in the Tatra Mountains. Biologia 61,S135–S146.
Mergeay, J., Verschuren, D., De Meester, L., 2005. Cryptic invasion anddispersal of an American Daphnia in East Africa. Limnol. Oceanogr.50, 1278–1283.
Paland, S., Colbourne, J.K., Lynch, M., 2005. Evolutionary history of con-tagious asexuality in Daphnia pulex. Evolution 59, 800–813.
Peters, R.H., de Bernardi, R. (Eds.), 1987. Daphnia. Consiglio nazionaledelle recherché, Instituto italitano di idrobiologia, Verbania Palanza.
Posada, D., Crandall, K.A., 1998. Modeltest: testing the model of DNAsubstitution. Bioinformatics 14, 817–818.
Rhode, S.C., Pawlowski, M., Tollrian, R., 2001. The impact of ultravioletradiation on the vertical distribution of zooplankton of the genusDaphnia. Nature 412, 69–72.
Ronquist, F., Hüelsenbeck, J., 2003. MrBayes 3: Bayesian phylogeneticinference under mixed models. Bioinformatics 19, 1572–1574.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method forreconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Schönswetter, P., Popp, M., Brochmann, C., 2006. Rare arctic–alpineplants of the European Alps have diVerent immigration histories: thesnow bed species Minuartia biXora and Ranunculus pygmaeus. Mol.Ecol. 15, 709–720.
Shimodaira, H., Hasegawa, M., 1999. Multiple comparison of log-likeli-hoods with applications to phylogenetic inference. Mol. Biol. Evol. 16,1114–1116.
Skrede, I., Eidesen, P.B., Portela, R.P., Brochmann, CH., 2006. Refugia,diVerentiation and postglacial migration in artic-alpine Eurasia, exem-pliWed by the mountain avens (Dryas octopetala L). Mol. Ecol. 15,1827–1840.
Schwenk, K., Junttila, P., Rautio, M., Bastiansen, F., Knapp, A., Dove, O.,Billiones, R., Streit, B., 2004. Ecological, morphological, and geneticdiVerentiation of Daphnia (Hyalodaphnia) from the Finnish and Rus-sian subarctic. Limnol. Oceanogr. 49, 532–539.
SwoVord, D.L., 2003. PAUP¤. Phylogenetic analysis using parsimony(¤ and other methods). Version 4. Sinauer Associates, Sunderland, MA.
Tarjuelo, I., Posada, D., Crandall, K.A., Pascual, M., Turon, X., 2004. Phy-logeography and speciation of colour morphs in the colonial ascidianPseudodistoma crucigaster. Mol. Ecol. 13, 3125–3136.
Taylor, D.J., Finston, T.L., Hebert, P.D.N., 1998. Biogeography of a wide-spread freshwater crustacean: pseudocongruence and cryptic endemism inthe North American Daphnia laevis complex. Evolution 52, 1648–1670.
Taylor, D.J., Sprenger, H.L., Ishida, S., 2005. Geographic and phylogeneticevidence for dispersed nuclear introgression in a daphniid with sexualpropagules. Mol. Ecol. 14, 525–537.
Templeton, A.R., Crandall, K.A., Sing, C.F., 1992. A cladistic analysis ofphenotypic association with haplotypes inferred from restriction endo-nuclease mapping and DNA sequence data. III. Cladogram estimation.Genetics 132, 619–633.
Ventura, M., Catalan, J., 2005. Reproduction as one of the main causes oftemporal variability in the elemental composition of zooplankton.Limnol. Oceanog. 50, 2043–2056.
Weider, L.J., Hobaek, A., 1997. Postglacial dispersal, glacial refugia, andclonal structure in Russian/Siberian populations of the Arctic Daphniapulex complex. Heredity 78, 363–372.
Weider, L.J., Hobaek, A., Colbourne, J.K., Crease, T.J., Dufresne, F.,Hebert, P.D.N., 1999a. Holarctic phylogeography of an asexual speciescomplex I. Mitochondrial DNA variation in arctic Daphnia. Evolution53, 777–792.
Weider, L., Hobaek, A., Hebert, P.D.N., Crease, T.J., 1999b. Holarctic phy-logeography of an asexual species complex-II. Allozymic variation andclonal structure in arctic Daphnia. Mol. Ecol. 8, 1–13.
Weider, L.J., Hobaek, A., 2003. Glacial refugia, haplotype distributions,and clonal richness of the Daphnia pulex complex in arctic Canada.Mol. Ecol. 12, 463–473.
Zhang, D.X., Hewitt, G.M., 1996. Nuclear integrations: challenge for mito-chondrial DNA markers. Trends Ecol. Evol. 11, 247–251.





























