Embed Size (px)
Citation preview

Infection, Genetics and Evolution 16 (2013) 191–199
Contents lists available at SciVerse ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsevier .com/locate /meegid
Current status of equine piroplasmosis in the Sudan
Bashir Salim a,e,⇑, Mohammed Ahmed Bakheit a,c, Joseph Kamau b,d, Chihiro Sugimoto b
a Department of Parasitology, Faculty of Veterinary Medicine, University of Khartoum, 13314 Khartoum-North, Sudanb Department of Collaboration and Education, Research Center for Zoonosis Control, Hokkaido University, Sapporo 001-0020, Japanc Veterinary Infection Biology and Immunology, Research Center Borstel, D-23845 Borstel, Germanyd Department of Biochemistry, School of Medicine, University of Nairobi, 30197-00100 Nairobi, Kenyae School of Biology, Centre for Genetics and Genomics, The University of Nottingham, University Park, Nottingham NG7 2RD, UK
a r t i c l e i n f o
Article history:Received 21 September 2012Received in revised form 6 February 2013Accepted 13 February 2013Available online 26 February 2013
Keywords:Equine piroplasmosisTrypanosomiasisSudan
1567-1348/$ - see front matter � 2013 Elsevier B.V. Ahttp://dx.doi.org/10.1016/j.meegid.2013.02.008
⇑ Corresponding author at: Department of ParasitMedicine, University of Khartoum, 13314 Khartoum-N
E-mail address: [email protected] (B. Salim)
a b s t r a c t
This is a cross-sectional molecular epidemiological study on equine piroplasmosis (EP) affecting horsesand donkeys in the Sudan. The study evaluated 499 samples from geographically distinct regions in east-ern, central and western parts of the country. PCR amplification of the 18S rRNA gene of both Thelieriaequi and Babesia caballi was carried out. Horses from all sampled areas were found positive to T. equiDNA but no B. caballi was detected. Absence of B. caballi infection was confirmed by another PCR targetingthe B. caballi 48-kDa merozoite antigen. The overall prevalence was found to be 35.95%. The highest prev-alence was detected in Showak 13 (81.3%) and the lowest was in Shearia locality in South Darfur 1 (5.6%).In another experiment, capillary electrophoresis was used to detect and differentiate between T. equi andB. caballi using one set of primers designed to amplify the 18S rRNA gene in a single PCR. Capillary elec-trophoresis method was found to be powerful in detecting mixed infections in artificially mixed controlssamples. The data obtained in this study would contribute to the development of a national control strat-egy of EP in the Sudan.
� 2013 Elsevier B.V. All rights reserved.
1. Introduction simultaneous detection and identification of Theileria and Babesia
Equine piroplasmosis (EP) is a tick-borne protozoan disease ofhorses, mules, donkeys and zebra caused by Theileria equi and Babe-sia caballi. EP is a notifiable disease (OIE, 2010) that has a wide dis-tribution and occurs in southern Europe, Asia, Africa, South andCentral America, and certain parts of the southern United States ofAmerica. Infected animals may remain carriers of these blood para-sites for long periods and act as sources of infection for tick vectors.
Diagnosis of equine piroplasmosis in carrier animals by meansof blood smear examination is difficult and subjective. Serologicaldiagnostic methods have been widely adopted because of absenceof cross-reaction between the antibodies produced against parasitespecies. However, false-negative or false-positive reactions couldbe encountered in serological tests (Donnelly et al., 1980; Tenterand Freidhoff, 1986). Several molecular techniques for the detec-tion of T. equi and B. caballi have been described. These methodsare based on species-specific PCR amplification of mainly the 18SrRNA gene (Tenter and Freidhoff, 1986; Bashiruddin et al., 1999;Criado-fornelio et al., 2003). Further refinements to PCR-basedtechniques included nested PCR (Rampersad et al., 2003) andloop-mediated isothermal amplification (LAMP) (Alhassan et al.,2007). Reverse line blot (RLB) and multiplex PCR were used for
ll rights reserved.
ology, Faculty of Veterinaryorth, Sudan.
.
species in horses (Alhassan et al., 2005; Nagore et al., 2004).In the last five years we have been monitoring EP in Sudan by
using serological and molecular tools to investigate prevalenceand genetic diversity of T. equi detected from different regions (Salimet al., 2008, 2010). These studies demonstrated that the disease ishighly prevalent with main genotypes being identified. However,more information on the distribution of EP and its enzootic situationis required to build a national control strategy against the disease.
To that end, the current study aimed to give a comprehensivepicture on the prevalence of EP covering whole Sudan. The studyincluded new geographical locations, where horse populationsare high in the country, thus complementing the epidemiologicalpicture of the disease in the country.
This paper also reports, the development of a rapid sensitiveand specific diagnostic test for detection and identification T. equiand B. caballi. The new test based on the amplification of parasite18S rRNA gene and the subsequent capillary electrophoresis canbe a useful tool for future investigation on the prevalence of EP.
2. Materials and methods
2.1. Sample collection
In a surveillance conducted in 2010 (October 1st–November15th), 509 blood samples were collected from horses and donkeysin FTA cards (Whatman FTA� elute, Whatman, UK). Samples were
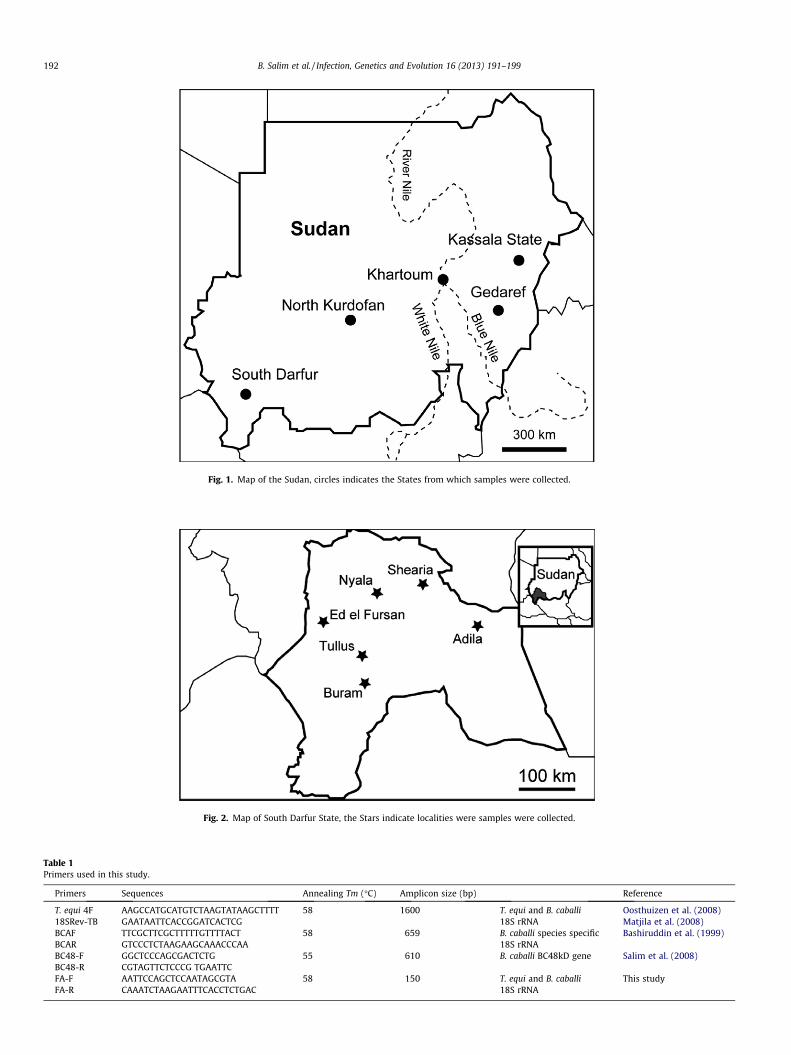
Fig. 1. Map of the Sudan, circles indicates the States from which samples were collected.
Fig. 2. Map of South Darfur State, the Stars indicate localities were samples were collected.
Table 1Primers used in this study.
Primers Sequences Annealing Tm (�C) Amplicon size (bp) Reference
T. equi 4F AAGCCATGCATGTCTAAGTATAAGCTTTT 58 1600 T. equi and B. caballi Oosthuizen et al. (2008)18SRev-TB GAATAATTCACCGGATCACTCG 18S rRNA Matjila et al. (2008)BCAF TTCGCTTCGCTTTTTGTTTTACT 58 659 B. caballi species specific Bashiruddin et al. (1999)BCAR GTCCCTCTAAGAAGCAAACCCAA 18S rRNABC48-F GGCTCCCAGCGACTCTG 55 610 B. caballi BC48kD gene Salim et al. (2008)BC48-R CGTAGTTCTCCCG TGAATTCFA-F AATTCCAGCTCCAATAGCGTA 58 150 T. equi and B. caballi This studyFA-R CAAATCTAAGAATTTCACCTCTGAC 18S rRNA
192 B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199
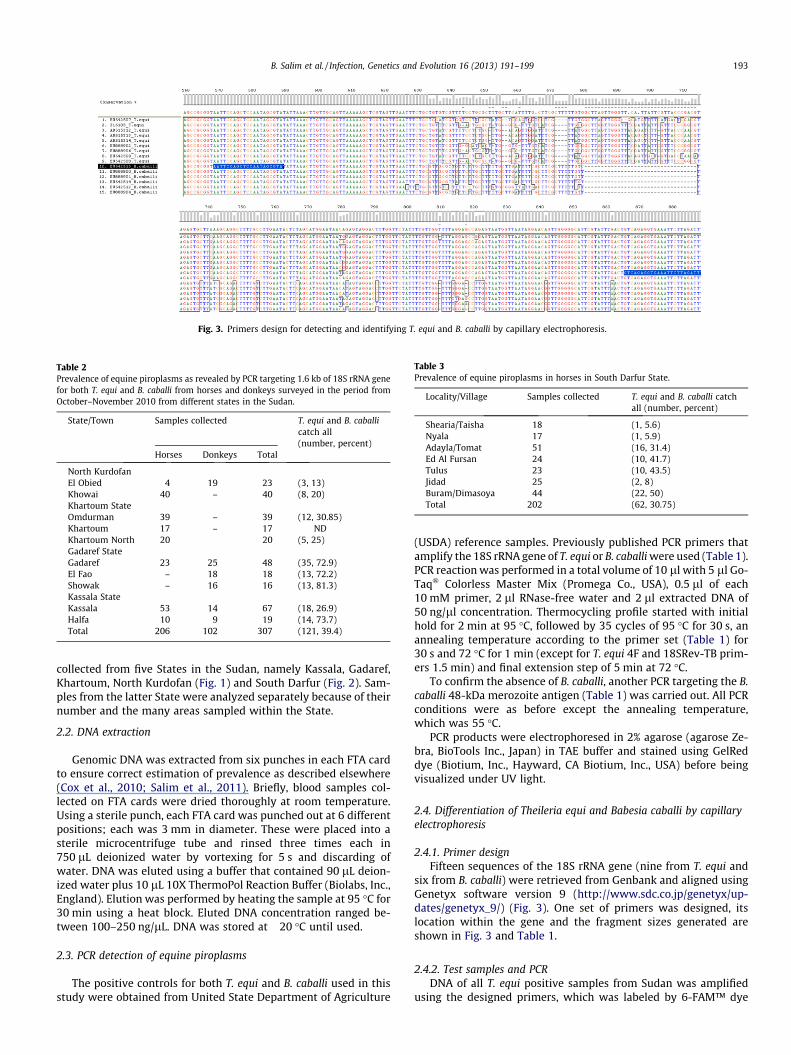
Fig. 3. Primers design for detecting and identifying T. equi and B. caballi by capillary electrophoresis.
Table 2Prevalence of equine piroplasms as revealed by PCR targeting 1.6 kb of 18S rRNA genefor both T. equi and B. caballi from horses and donkeys surveyed in the period fromOctober–November 2010 from different states in the Sudan.
State/Town Samples collected T. equi and B. caballicatch all(number, percent)
Horses Donkeys Total
North KurdofanEl Obied 4 19 23 (3, 13)Khowai 40 – 40 (8, 20)Khartoum StateOmdurman 39 – 39 (12, 30.85)Khartoum 17 – 17 NDKhartoum North 20 20 (5, 25)Gadaref StateGadaref 23 25 48 (35, 72.9)El Fao – 18 18 (13, 72.2)Showak – 16 16 (13, 81.3)Kassala StateKassala 53 14 67 (18, 26.9)Halfa 10 9 19 (14, 73.7)Total 206 102 307 (121, 39.4)
Table 3Prevalence of equine piroplasms in horses in South Darfur State.
Locality/Village Samples collected T. equi and B. caballi catchall (number, percent)
Shearia/Taisha 18 (1, 5.6)Nyala 17 (1, 5.9)Adayla/Tomat 51 (16, 31.4)Ed Al Fursan 24 (10, 41.7)Tulus 23 (10, 43.5)Jidad 25 (2, 8)Buram/Dimasoya 44 (22, 50)Total 202 (62, 30.75)
B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199 193
collected from five States in the Sudan, namely Kassala, Gadaref,Khartoum, North Kurdofan (Fig. 1) and South Darfur (Fig. 2). Sam-ples from the latter State were analyzed separately because of theirnumber and the many areas sampled within the State.
2.2. DNA extraction
Genomic DNA was extracted from six punches in each FTA cardto ensure correct estimation of prevalence as described elsewhere(Cox et al., 2010; Salim et al., 2011). Briefly, blood samples col-lected on FTA cards were dried thoroughly at room temperature.Using a sterile punch, each FTA card was punched out at 6 differentpositions; each was 3 mm in diameter. These were placed into asterile microcentrifuge tube and rinsed three times each in750 lL deionized water by vortexing for 5 s and discarding ofwater. DNA was eluted using a buffer that contained 90 lL deion-ized water plus 10 lL 10X ThermoPol Reaction Buffer (Biolabs, Inc.,England). Elution was performed by heating the sample at 95 �C for30 min using a heat block. Eluted DNA concentration ranged be-tween 100–250 ng/lL. DNA was stored at �20 �C until used.
2.3. PCR detection of equine piroplasms
The positive controls for both T. equi and B. caballi used in thisstudy were obtained from United State Department of Agriculture
(USDA) reference samples. Previously published PCR primers thatamplify the 18S rRNA gene of T. equi or B. caballi were used (Table 1).PCR reaction was performed in a total volume of 10 ll with 5 ll Go-Taq� Colorless Master Mix (Promega Co., USA), 0.5 ll of each10 mM primer, 2 ll RNase-free water and 2 ll extracted DNA of50 ng/ll concentration. Thermocycling profile started with initialhold for 2 min at 95 �C, followed by 35 cycles of 95 �C for 30 s, anannealing temperature according to the primer set (Table 1) for30 s and 72 �C for 1 min (except for T. equi 4F and 18SRev-TB prim-ers 1.5 min) and final extension step of 5 min at 72 �C.
To confirm the absence of B. caballi, another PCR targeting the B.caballi 48-kDa merozoite antigen (Table 1) was carried out. All PCRconditions were as before except the annealing temperature,which was 55 �C.
PCR products were electrophoresed in 2% agarose (agarose Ze-bra, BioTools Inc., Japan) in TAE buffer and stained using GelReddye (Biotium, Inc., Hayward, CA Biotium, Inc., USA) before beingvisualized under UV light.
2.4. Differentiation of Theileria equi and Babesia caballi by capillaryelectrophoresis
2.4.1. Primer designFifteen sequences of the 18S rRNA gene (nine from T. equi and
six from B. caballi) were retrieved from Genbank and aligned usingGenetyx software version 9 (http://www.sdc.co.jp/genetyx/up-dates/genetyx_9/) (Fig. 3). One set of primers was designed, itslocation within the gene and the fragment sizes generated areshown in Fig. 3 and Table 1.
2.4.2. Test samples and PCRDNA of all T. equi positive samples from Sudan was amplified
using the designed primers, which was labeled by 6-FAM™ dye
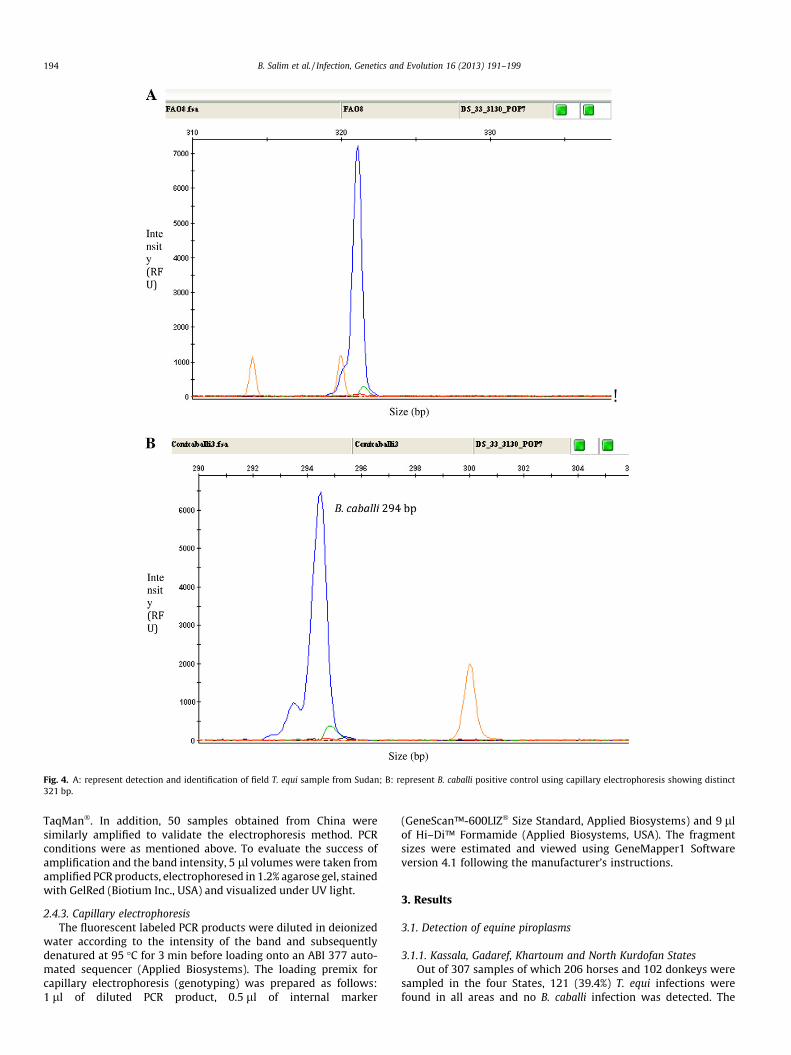
Fig. 4. A: represent detection and identification of field T. equi sample from Sudan; B: represent B. caballi positive control using capillary electrophoresis showing distinct321 bp.
194 B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199
TaqMan�. In addition, 50 samples obtained from China weresimilarly amplified to validate the electrophoresis method. PCRconditions were as mentioned above. To evaluate the success ofamplification and the band intensity, 5 ll volumes were taken fromamplified PCR products, electrophoresed in 1.2% agarose gel, stainedwith GelRed (Biotium Inc., USA) and visualized under UV light.
2.4.3. Capillary electrophoresisThe fluorescent labeled PCR products were diluted in deionized
water according to the intensity of the band and subsequentlydenatured at 95 �C for 3 min before loading onto an ABI 377 auto-mated sequencer (Applied Biosystems). The loading premix forcapillary electrophoresis (genotyping) was prepared as follows:1 ll of diluted PCR product, 0.5 ll of internal marker
(GeneScan™-600LIZ� Size Standard, Applied Biosystems) and 9 llof Hi–Di™ Formamide (Applied Biosystems, USA). The fragmentsizes were estimated and viewed using GeneMapper1 Softwareversion 4.1 following the manufacturer’s instructions.
3. Results
3.1. Detection of equine piroplasms
3.1.1. Kassala, Gadaref, Khartoum and North Kurdofan StatesOut of 307 samples of which 206 horses and 102 donkeys were
sampled in the four States, 121 (39.4%) T. equi infections werefound in all areas and no B. caballi infection was detected. The

Table 4Fragment analysis of samples from China.
No. Sample no. Base pairs
1 5 3472 7 3473 10 321–3474 11 3475 12 3476 14 3477 15 3478 16 3479 17 34710 18 34611 19 34712 20 34713 22 34714 23 34715 26 34716 27 33317 30 34718 32 35019 33 34720 36 33421 37 34722 42 34723 45 321–34524 48 35025 49 34726 51 34727 52 34728 53 347
B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199 195
highest prevalence was recorded in Gadaref State (35, 72.9%) fol-lowed by Kassala (18, 26.9%) (Table 2). T. equi infection was dem-onstrated in both horses and donkeys.
3.1.2. South Darfur StateThe prevalence of T. equi in this State was estimated to be
30.75% in horses since no donkeys were sampled in the State (Ta-ble 3). No samples were positive for B. caballi. The highest preva-lence was demonstrated in Buram (22, 50%) and the lowest wasin Nyala and Shearia (1, 5.9% and 1, 5.6%, respectively).
3.2. Capillary electrophoresis and fragment analysis of T. equi and B.caballi
The method gave a fragment size of 321 bp for T. equi in the po-sitive control and all T. equi field samples collected from Sudan(Fig. 4A), however B. caballi positive control showed a fragmentsize of 294 bp was obtained (Fig. 4B).
The method was further applied to 50 positive samples fromChina (T. equi and B. caballi) and the results revealed that someof the samples gave similar results to the Sudanese T. equi fragmentsize 321 bp (Fig. 5A) and that the others gave different fragmentsizes (Table 4). These differences in the fragment sizes may repre-sent different genotypes. B. caballi from China revealed the samefragment size to the positive control 294 bp (Fig. 5B).
None of the field samples were found to have mixed infection.Therefore, to mimic the case, positive controls of T. equi and B. cab-alli were first cloned then mixed as follows (0, 1, 2, 3, 4, 5 copy T.equi with their respective counterpart from B. caballi (5, 4, 3, 2, 1,0). In such mixtures, both parasites could be amplified and differ-entiated using capillary electrophoeresis (Fig. 6).
One sample from China (sample #45) revealed two bands inagarose gel and showed two-district fragment size when capillaryelectrophoresed 321 bp and 347 bp (Fig. 7). This sample was fur-ther sequenced and deletion of 26 bp was found to occur (Fig. 8).
4. Discussion
This study gave a comprehensive overview of equine piroplas-mosis in the Sudan focusing on areas with high population ofhorses in the Sudan, south Darfur. Additionally, we described anew and high throughput method for differentiating the causativeagents of the (EP) T. equi and B. caballi. Species differentiation willhelp to prescribe the drug of choice since the two parasites responddifferently to treatment and disease control.
The current study indicates that the disease is prevalent all overthe country where samples were collected. We suspect vector ticksresponsible for the transmission to be Hyalomma spp and Rhipi-cephalus spp, whose prevalence has been previously been reported(Salim et al., 2008).
The study showed that T. qui to be highly prevalent all in allareas sampled, with overall prevalence of 499 (35.95%). The high-est prevalence was scored in Showak and the lowest was in Shearialocality, South Darfur. This might be attributed to the type of ani-mals sampled and the tendency of T. equi to be highly prevalentin donkeys as in case of Showak than in horses as in case of Shearia.Donkeys usually show asymptomatic form of the disease, withlower T. equi parasitaemia when compared to horses (Kumaret al., 2009). The authors suggested that prevalence is of host spec-ificity rather than of place distribution. Although the vector distri-bution may play a role with this regard, but no data available onticks infesting equids in these areas.
There were no positive samples with B. caballi detected. Mostprobable reason would be that only healthy animals were sampledyet. It is well documented that B. caballi is known to be only found(positively identified) in acute cases. However, in our previousstudy (Salim et al., 2008), detected high levels of antibodies sug-gesting presence of B. caballi. Since there has been no cross-reaction reported between B. caballi and T. equi, then the possibilityof B. caballi occurring but being missed in the current study is prob-able, taking into consideration that two sets of primers not suffi-cient to conclude that none of the horses are infected. Definitelyinclusion of serological data would provided information aboutcumulative exposure horses to equine piroplasmosis and elucidateof the diseases have prevalence more compressively but that wasnot possible since sera were not collected.
To exclude any question regarding the gene target which mayhave played a role in limitation of the PCR since neither of thetwo PCRs targeting 18S rDNA and 48-kDa genes were capable ofdetecting a single positive case of B. caballi, we decided to employcapillary electrophoresis genotyping method to confirm.
Capillary electrophoresis genotyping method is a novel methodfor detection, differentiation of both T. equi and B. caballi in a singlereaction. The method has successfully been employed in differen-tiation of T. orientalis types Kamau et al. (2011). The authors brieflyrun a PCR amplification, Capillary gel electrophoresis and genotyp-ing based on ITS1 or ITS2 rRNA spacer regions, followed by scoringthe different fragment sizes as peaks representing different types.
This method can be used in addition to detection and identifica-tions of the species as quantifiable based on the capillary electro-phoresis peak height as we clearly illustrated in our work(Kamau et al., 2011).
When capillary electrophoresis genotyping method was appliedin detection and differentiation of equine piroplasmosis (EP) it wasfound to be highly sensitive and specific. Our choice of 18S rRNAgene as the target region was due to evolutionary stability of thegene, used worldwide for taxonomy and species differentiation.The polymorphic region used to design the primers used in thisstudy was found to be fixed and consistent throughout the T. equidetected from the Sudan. Interestingly this amplified region in thesamples collected from China gave almost consistent 347 bp but
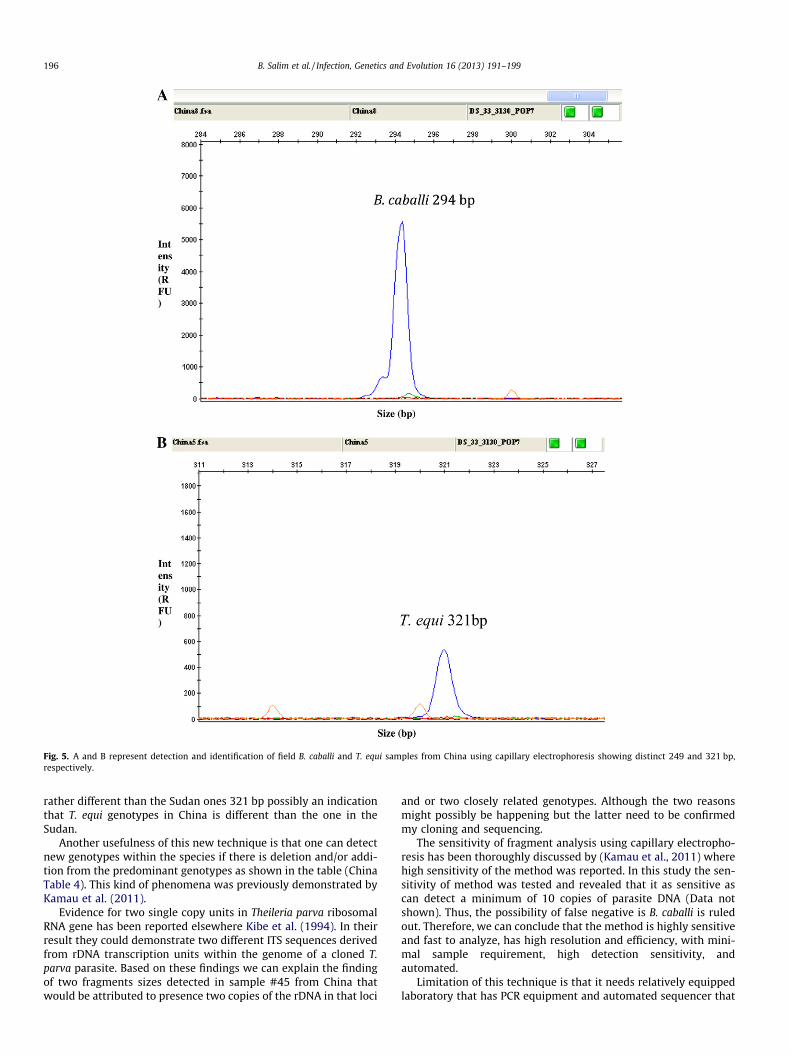
Fig. 5. A and B represent detection and identification of field B. caballi and T. equi samples from China using capillary electrophoresis showing distinct 249 and 321 bp,respectively.
196 B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199
rather different than the Sudan ones 321 bp possibly an indicationthat T. equi genotypes in China is different than the one in theSudan.
Another usefulness of this new technique is that one can detectnew genotypes within the species if there is deletion and/or addi-tion from the predominant genotypes as shown in the table (ChinaTable 4). This kind of phenomena was previously demonstrated byKamau et al. (2011).
Evidence for two single copy units in Theileria parva ribosomalRNA gene has been reported elsewhere Kibe et al. (1994). In theirresult they could demonstrate two different ITS sequences derivedfrom rDNA transcription units within the genome of a cloned T.parva parasite. Based on these findings we can explain the findingof two fragments sizes detected in sample #45 from China thatwould be attributed to presence two copies of the rDNA in that loci
and or two closely related genotypes. Although the two reasonsmight possibly be happening but the latter need to be confirmedmy cloning and sequencing.
The sensitivity of fragment analysis using capillary electropho-resis has been thoroughly discussed by (Kamau et al., 2011) wherehigh sensitivity of the method was reported. In this study the sen-sitivity of method was tested and revealed that it as sensitive ascan detect a minimum of 10 copies of parasite DNA (Data notshown). Thus, the possibility of false negative is B. caballi is ruledout. Therefore, we can conclude that the method is highly sensitiveand fast to analyze, has high resolution and efficiency, with mini-mal sample requirement, high detection sensitivity, andautomated.
Limitation of this technique is that it needs relatively equippedlaboratory that has PCR equipment and automated sequencer that
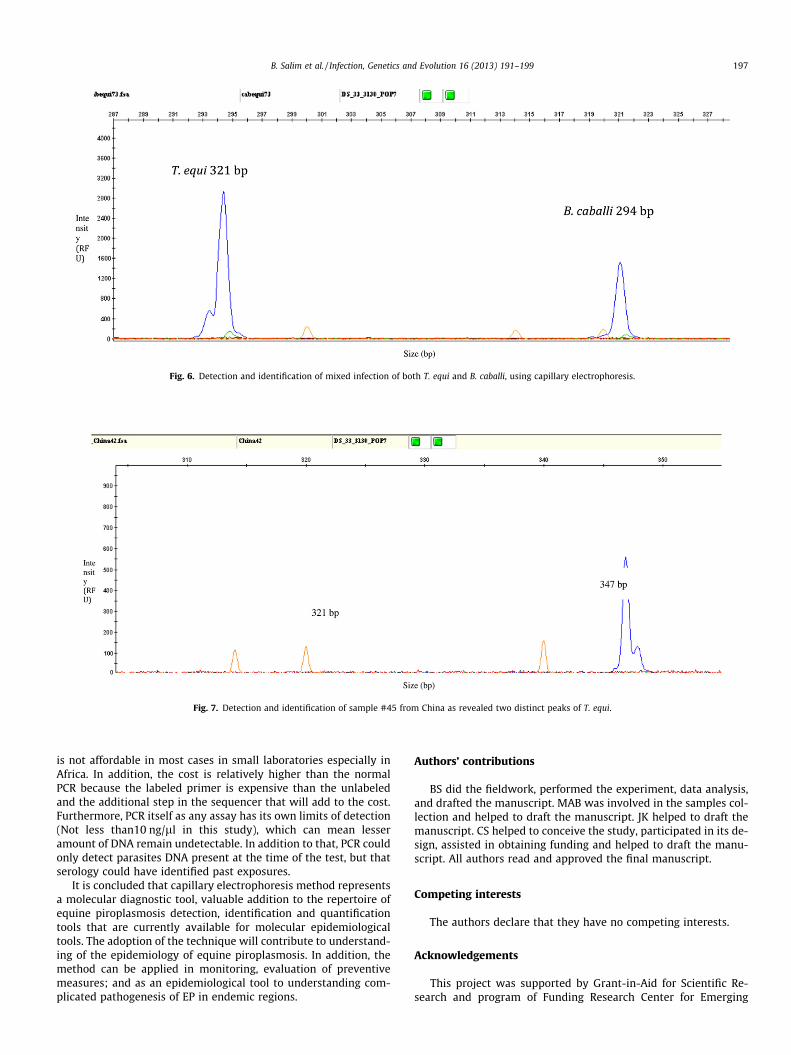
Fig. 6. Detection and identification of mixed infection of both T. equi and B. caballi, using capillary electrophoresis.
Fig. 7. Detection and identification of sample #45 from China as revealed two distinct peaks of T. equi.
B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199 197
is not affordable in most cases in small laboratories especially inAfrica. In addition, the cost is relatively higher than the normalPCR because the labeled primer is expensive than the unlabeledand the additional step in the sequencer that will add to the cost.Furthermore, PCR itself as any assay has its own limits of detection(Not less than10 ng/ll in this study), which can mean lesseramount of DNA remain undetectable. In addition to that, PCR couldonly detect parasites DNA present at the time of the test, but thatserology could have identified past exposures.
It is concluded that capillary electrophoresis method representsa molecular diagnostic tool, valuable addition to the repertoire ofequine piroplasmosis detection, identification and quantificationtools that are currently available for molecular epidemiologicaltools. The adoption of the technique will contribute to understand-ing of the epidemiology of equine piroplasmosis. In addition, themethod can be applied in monitoring, evaluation of preventivemeasures; and as an epidemiological tool to understanding com-plicated pathogenesis of EP in endemic regions.
Authors’ contributions
BS did the fieldwork, performed the experiment, data analysis,and drafted the manuscript. MAB was involved in the samples col-lection and helped to draft the manuscript. JK helped to draft themanuscript. CS helped to conceive the study, participated in its de-sign, assisted in obtaining funding and helped to draft the manu-script. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Acknowledgements
This project was supported by Grant-in-Aid for Scientific Re-search and program of Funding Research Center for Emerging

Fig. 8. Direct sequencing of sample #45 from China revealed two distinct sequences with 26 deletions.
198 B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199
and Re-emerging Infectious Disease, from Ministry of Education,Culture, Sports, Science and Technology of Japan (MEXT), Japan.The authors are grateful to Dr. Kyoko Hyashida and Dr. Ryo Nakaotheir valuable assistance.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.meegid.2013.02.008.
References
Alhassan, A., Pumidonming, W., Okamura, M., Hirata, H., Battsetseg, B., Fujisaki, K.,Yokoyama, N., Igarashi, I., 2005. Development of a single-round and multiplexPCR method for the simultaneous detection of Babesia caballi and Babesia equi inhorse blood. Vet. Parasitol. 129, 43–49.
Alhassan, A., Govind, Y., Tam, N.T., Thekisoe, O.M., Yokoyama, N., Inoue, N., Igarashi,I., 2007. Comparative evaluation of the sensitivity of LAMP, PCR and in vitroculture methods for the diagnosis of equine piroplasmosis. Parasitol. Res. 100,1165–1168.
Bashiruddin, J.B., Cammà, C., Rebêlo, E., 1999. Molecular detection of Babesia equiand Babesia caballi in horse blood by PCR amplification of part of the 16S rRNAgene. Vet. Parasitol. 84, 75–83.
Cox, A.P., Tosas, O., Tilley, A., Picozzi, K., Coleman, P., Hide, G., Welburn, S.C., 2010.Constraints to estimating the prevalence of trypanosome infections in EastAfrican zebu cattle. Parasit Vectors 3, 82.
Criado-Fornelio, A., Martinez-Marcos, A., Buling-Saraña, A., Barba-Carretero, J.C.,2003. Molecular studies on Babesia, Theileria and Hepatozoon in southernEurope: Part II. Phylogenetic analysis and evolutionary history. Vet. Parasitol.114, 173–194.
Kamau, J., Salim, B., Yokoyama, N., Kinyanjui, P., Sugimoto, C., 2011. Rapiddiscrimination and quantification of Theileria orientalis types using ribosomalDNA internal transcribed spacers. Infect. Genet. Evol. 11, 407–414.
Kibe, M.K., ole-MoiYoi, O.K., Nene, V., Khan, B., Allsopp, B.A., Collins, N.E., Morzaria,S.P., Gobright, E.I., Bishop, R.P., 1994. Evidence for two single copy units inTheileria parva ribosomal RNA genes. Mol. Biochem. Parasitol. 66, 249–259.
Kumar, S., Kumar, R., Sugimoto, C., 2009. A perspective on Theileria equi infections indonkeys. Jpn. J. Vet. Res. 56, 171–180.
Matjila, P.T., Leisewitz, A.L., Oosthuizen, M.C., Jongejan, F., Penzhorn, B.L., 2008.Detection of Theileria species in dogs in South Africa. Vet. Parasitol. 157, 34–40.
Nagore, D., García-Sanmartín, J., García-Pérez, A.L., Juste, R.A., Hurtado, A., 2004.Detection and identification of equine Theileria and Babesia species by reverseline blotting: epidemiological survey and phylogenetic analysis. Vet. Parasitol.123, 41–54.
OIE, 2010. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2010.<http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.05.08_EQUINE_PIROPLASMOSIS.pdf>.
Onnelly, J., Joyner, L.P., Graham-Jones, O., Ellis, C.P., 1980. A comparison of thecomplement fixation and immunofluorescent antibody titres in a survey of theprevalence of Babesia equi and Babesia caballi in horses in the Sultanate ofOman. Trop. Anim. Health Prod. 12, 50–60.

B. Salim et al. / Infection, Genetics and Evolution 16 (2013) 191–199 199
Oosthuizen, M.C., Zweygarth, E., Collins, N.E., Troskie, M., Penzhorn, B.L., 2008.Identification of novel Babesia sp. from a sable antelope (Hippotragus nigerHaris, 1838). J. Clin. Microbiol. 46, 2247–2251.
Rampersad, J., Cesar, E., Campbell, M.D., Samlal, M., Ammons, D., 2003. A fieldevaluation of PCR for the routine detection of Babesia equi in horses. Vet.Parasitol. 114, 81–87.
Salim, B.O., Hassan, S.M., Bakheit, M.A., Alhassan, A., Igarashi, I., Karanis, P.,Abdelrahman, M.B., 2008. Diagnosis of Babesia caballi and Theileria equiinfections in horses in Sudan using ELISA and PCR. Parasitol. Res. 103, 1145–1150.
Salim, B., Bakheit, M.A., Kamau, J., Nakamura, I., Sugimoto, C., 2010. Nucleotidesequence heterogeneity in the small subunit ribosomal RNA gene withinTheileria equi from horses in Sudan. Parasitol. Res. 106, 493–498.
Salim, B., Bakheit, M.A., Kamau, J., Nakamura, I., Sugimoto, C., 2011. Molecularepidemiology of camel trypanosomiasis based on ITS1 rDNA and RoTat 12 VSGgene in the Sudan. Parasit Vectors 4, 31.
Tenter, A.M., Freidhoff, K.T., 1986. Serodiagnosis of experimental and naturalBabesia equi and B. caballi infections. Vet. Parasitol. 20, 49–61.





























