Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Detection of microalgal resting cysts in European coastal sediments using aPCR-based assay
Antonella Penna a,�, Cecilia Battocchi a, Esther Garces b, Silvia Angl�es b, Emellina Cucchiari c,Cecilia Totti c, Anke Kremp d, Cecilia Satta e, Maria Grazia Giacobbe f, Isabel Bravo g, Mauro Bastianini h
a Department of Biomolecular Sciences, University of Urbino, Viale Trieste 296, 61100 Pesaro, Italyb Institut de Ci�encies del Mar, CSIC, Passeig Marıtim de la Barceloneta 37-43, 08003 Barcelona, Spainc Dipartimento di Scienze del Mare, Universit�a Politecnica delle Marche, Via Brecce Bianche, 60131 Ancona, Italyd Tvarminne Zoological Station, University of Helsinki, 10900 Hanko, Finlande Dip. Botanica ed Ecologia Vegetale, University of Sassari, 07100 Sassari, Italyf Istituto per l’Ambiente Marino Costiero, CNR, Spianata S. Raineri 86, 98122 Messina, Italyg Instituto Espanol de Oceanografıa, Apdo. 1552, 36200 Vigo, Spainh CNR-ISMAR, Istituto di Scienze Marine, Castello 1364/A, 30122 Venezia, Italy
a r t i c l e i n f o
Available online 23 September 2009
Keywords:
HAB species
Mediterranean Sea
PCR
Ribosomal genes
Resting stages
Sediments
a b s t r a c t
A PCR-based assay was developed and applied to sediment and sediment trap samples for the detection
of different cysts belonging to dinoflagellates and raphidophytes in European coastal areas.
Oligonucleotide primers were designed based on the ITS-5.8S and LSU ribosomal gene sequences.
The specificity and sensitivity of the PCR assay were assessed using genomic DNA from clonal cultures,
plasmid copy number of cloned target sequences, as well as from sediment samples. Qualitative PCR
determinations of different cysts in sediment and sediment trap samples were compared to taxonomic
examinations by light microscopy. This molecular methodology permitted a fast and specific detection
of target cysts in sediment samples. We also detected dinoflagellate and raphidophyte cysts at
concentrations not detectable by microscopic methods or that are difficult to identify. The results given
by molecular and microscopic methods were comparable. However, higher values of positive detection
for target cysts were obtained by PCR than with microscopy. Some taxa were detected in 100% of the
samples using PCR, while others were only found in 10% of the samples. The data obtained in this study
showed that the PCR-based method is a valid tool for cyst identification in marine sediments.
& 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Harmful Algal Blooms (HABs) are recurring events in Europeancoastal waters (Giacobbe et al., 2007; Smayda, 2007). Many HABspecies are responsible for these events and the majority aredinoflagellates (IOC Taxonomic reference list, http://www.bi.ku.dk/ioc/). Most HAB dinoflagellates display heteromorphic lifecycles, including motile planktonic stages, as well as immotilebenthic resting cysts. All stages of the life cycle, and especially thedormant cysts, have a large impact on bloom dynamics. Thebloom development of cyst-forming HAB species may also bedependent on the presence of cyst beds seeding local blooms(Garces et al., 2002; Steidinger and Garces, 2006). While greateffort has been dedicated to the study of the planktonic stages ofthese microorganisms, we still have limited knowledge of theirother life stages, especially benthic stages. In some dinoflagellates
and raphidophytes, the resting stages with their resistant wallspermits the species to tolerate environmental conditions andtherefore to expand their geographical distribution (Amorim et al.,2001; McGillicuddy et al., 2003; De Boer et al., 2004; Edvardsenand Imai, 2006).
Knowledge of the cyst bed composition and location in bottomsediments can provide information on the long-term presence of aspecies in an area, as well as enabling prediction of subsequentblooms. Mapping the cyst distribution of HAB species alsoprovides information on cyst transport to new areas, dependingon oceanic currents. Moreover, cyst distribution reflects thesedimentary dynamics and the location of blooms in the overlyingsurface waters (Angl�es et al., 2010). Further, cyst assemblage dataprovide an indication of the potential plankton diversity reservoirin a locality. All this information is necessary to understand andpredict potential HAB development and expansion. Therefore,monitoring the distribution of cyst densities in coastal areas priorto an outbreak is important for localizing hot spots for blooms andto minimize the damage caused by harmful blooms (Bravo et al.,2006).
ARTICLE IN PRESS
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/dsr2
Deep-Sea Research II
0967-0645/$ - see front matter & 2009 Elsevier Ltd. All rights reserved.
doi:10.1016/j.dsr2.2009.09.010
� Corresponding author.
E-mail address: [email protected] (A. Penna).
Deep-Sea Research II 57 (2010) 288–300

Author's personal copyARTICLE IN PRESS
Studies on resting cyst diversity and distribution have beenhampered by difficulties in the reliable identification of species-specific resting stages, since dinoflagellate and raphidophyte cystsdo not always have species-specific morphological features.Traditional microscopic methods, which are commonly employedfor monitoring of HAB species, often do not allow identification,unless the corresponding vegetative form can be providedthrough in vitro germination of the respective cysts. Moleculartechniques and in particular, PCR techniques based on theamplification of targeted ribosomal genes, have been developedfor the rapid and accurate detection and quantification ofvegetative cells of HAB species both in culture and field samples(Galluzzi et al., 2004; Coyne et al., 2005; Godhe et al., 2007; Pennaet al., 2007). The PCR detection technique can be applied tosamples from sediment cores and sediment traps containing avariety of cysts. Such molecular-based assays can accurately andrapidly identify a variety of specific taxa in the sediments,overcoming the problem of taxonomic identification by micro-scopy (Bolch, 2001; Bowers et al., 2006). However, to date, only afew dinoflagellates have been identified in sediment samplesusing the PCR approach (Godhe et al., 2002; Kamikawa et al.,2007). The success of the PCR-based approach in sediments isstrictly dependent on the inhibitory substances contained in thesediment material (Saito et al., 2002). Different strategies areapplied to overcome the problem of PCR inhibition caused bycontaminating substances: the use of commercial nucleic acidextraction kits that remove inhibitors and facilitate purification;the use of thermostable DNA polymerase; the dilution of templateDNA prior to PCR assay and the use of BSA (bovine serumalbumin) in the PCR assay.
In this study, a PCR-based assay was developed and applied tosediment and sediment trap samples for the detection of restingcysts from several taxa. The specificity and sensitivity of the PCRassay were assessed both in clonal microalgal culture and fieldsediment samples that were collected from several Europeancoastal areas. Oligonucleotide primers specific to several HABdinoflagellate species, namely Alexandrium spp., Lingulodinium
polyedrum, Protoceratium reticulatum, Gymnodinium catenatum,G. nolleri, and the raphidophyte Fibrocapsa japonica were applied inthe PCR-based assay to identify the corresponding cyst morpho-types in sediment and sediment trap samples from different areasof the Mediterranean, Baltic Sea, and the East Atlantic coast. Theassay was based on the LSU, 5.8S rRNA genes and ITS regions astarget regions for the taxa specific primers and was validated onthe genomic DNA of the clonal cultures and preserved cystsamples. A protocol for species-specific PCR detection of cystswas developed based on the use of a nucleic acid purification kit.The specificity and sensitivity of each assay was determined andcomparisons of the qualitative determinations of the PCR analysisand optical microscopy were made. Data on the detection of theresting cyst stage of the different HA species are shown.
2. Methods
2.1. Study sites
Surface sediment samples were collected during the years2006 and 2007 from 38 sampling stations located at 11 differentsites (Table 1) in the Mediterranean and Baltic Sea and the EastAtlantic (Fig. 1). In the Mediterranean Sea, the sampling sites weredistributed in four regional Seas: the North West Adriatic, Ionian,Tyrrhenian, and Catalan Sea. The sampling sites in the Baltic Seaand East Atlantic Ocean were located in the coastal NorthernBaltic Sea and the western coast of Spain, respectively. Stationswere located in areas where dinoflagellate or raphidophyte
blooms occur, and in sediment accumulation areas, as indicatedby deposition maps, e.g. for the Arenys harbor (Garces et al., 2004;Angl�es et al., 2010) and Ria de Vigo (Bravo and Anderson, 1994).
2.2. Cyst sample collection, purification, and microscopic
morphotype identification
Vertical sediment samples up to 8–10 cm in depth werecollected using a gravity corer (7.2 cm internal diameter). Threereplicate samples were taken per station. Sediment samples ofArenys de Mar (Catalan Sea) and Olbia (Tyrrhenian Sea) harborswere collected and processed as indicated in Satta et al. (2010).Sediment samples (7.5 ml) were extracted from the undisturbed3-cm surface layer in the sample core container using a syringeand were transferred into a 50-ml conical tube. In addition,sediment samples of 10 ml were collected from the bottom 3 cm ofthe sediment surface layer by a SCUBA diver using a plasticsyringe. Samples were preserved by adding Lugol’s solution(Andersen and Throndsen, 2003) in filtered seawater and storingthem in the dark at 4 1C until analysis. Three samples were takenfrom sediment traps, collected at the Arenys harbor (Catalan Sea)and Baiona site (East Atlantic coast); the sampling of sedimenttraps was carried out following the method of Garces et al. (2004).Settled material was collected from the traps every 4 days andtrap subsamples of 48 ml were fixed with Lugol’s solution andkept in the dark at 4 1C until analysis.
Subsamples of 2.5 ml of suspended sediments were sonicated(Bandelin, Germany) to disaggregate cysts from sediment parti-cles, sieved on steel membrane (Endecotts, UK) using sizefractionation of 100- and 10-mm membranes and transferred intofiltered seawater. The density gradient method was applied tosediment subsamples to separate cysts from detrital material(Amorim et al., 2001; Bravo et al., 2006). Samples were then usedboth for microscopic examination (2 ml subsamples) and mole-cular analysis (5-ml subsamples). The resting cysts were identifiedand counted under an inverted microscope at 200� and 400�magnification by scanning the entire Utermohl sedimentationchamber. Different morphotypes of dinoflagellate and raphido-phyte cysts were classified according to shape, color, wallthickness, and size. Cyst abundance was expressed as the cystnumber per volume of wet sediment sample and total cystnumber processed for molecular analysis.
2.3. Genomic DNA extraction of sediment and trap samples
Sediment samples (5 ml), obtained as above, were centrifugedat 4000 rpm for 10 min at room temperature; the supernatant wasgently discarded and 20 ml of filtered seawater were added;samples were again centrifuged at 4000 rpm for 10 min at roomtemperature. This washing step was repeated twice; then, the cystpellet was resuspended in 1 ml of sterile MilliQ water andcentrifuged at 10,000 rpm for 5 min at room temperature; thesupernatant was discarded and the pellet was frozen at �80 1Cuntil nucleic acid extraction. The pellet was thawed at +65 1C for15 min, and re-frozen at �80 1C for 15 min; this freeze–thaw stepwas repeated twice. After the last thawing, the pellet wassonicated for 20 min in an ultrasonic bath and then added to atube containing beads and lysis buffer of the UltraClean Soil DNaKit (MoBio Lab In., Solana Beach, CA). DNA extraction andpurification were carried out according to the manufacturer’sinstructions. Purified genomic DNA was quantified on an agarosegel using serially diluted Lambda DNA Marker (MBI Fermentas,Germany) and a gel-documentation apparatus (Bio-Rad, Hercules,CA, USA).
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300 289

Author's personal copyARTICLE IN PRESS
Ta
ble
1S
ed
ime
nt
sam
ple
sco
lle
cte
da
lon
gse
ve
ral
coa
sta
la
rea
sin
the
Me
dit
err
an
ea
n,
Ba
ltic
Se
aa
nd
Ea
ste
rnA
tla
nti
c,cy
sta
bu
nd
an
cep
er
sam
ple
an
dp
roce
sse
dcy
sta
bu
nd
an
cein
the
mo
lecu
lar
ba
sed
ass
ay.
Ge
og
rap
hic
al
are
aLo
cali
tie
sa
nd
sam
pli
ng
sta
tio
ns
Sta
tio
nco
ord
ina
tes
Sa
mp
lin
gp
eri
od
Sta
tio
n
nu
mb
er
Sa
mp
lin
gty
pe
Cy
sta
bu
nd
an
ce
(ml�
1�
we
tse
dim
en
t)
To
tal
pro
cess
ed
cyst
ab
un
da
nce
NW
Ad
ria
tic
Se
a,
Ita
lyV
en
ezi
a,
C1
04
51
150
0000
N,
121
450
6000
EJa
nu
ary
20
06
1S
ed
ime
nt
core
42
84
28
6
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,S
ou
thA
431
300
0900
N,
131
410
0000
EJu
ly2
00
62
Se
dim
en
tco
re1
26
18
90
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,S
ou
thC
431
300
0000
N,
131
410
0700
EJu
ly2
00
63
Se
dim
en
tco
re9
41
41
5
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,S
ou
thB
431
300
0500
N,
131
410
0300
EJu
ly2
00
64
Se
dim
en
tco
re7
71
10
5
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,S
ou
th-o
ut
CD
431
300
0000
N,
131
410
0700
ED
ece
mb
er
20
06
5S
ed
ime
nt
core
12
01
80
5
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
h-o
ut
AB
431
300
0900
N,
131
410
0000
ED
ece
mb
er
20
06
6S
ed
ime
nt
core
59
89
0
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,S
ou
th-i
nA
B4
31
300
0500
N,
131
410
0300
ED
ece
mb
er
20
06
7S
ed
ime
nt
core
78
11
75
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
hC
431
410
0400
N,
131
230
0000
ED
ece
mb
er
20
06
8S
ed
ime
nt
core
30
64
59
0
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
h-i
nA
BL
431
420
0100
N,
131
220
0000
ED
ece
mb
er
20
06
9S
ed
ime
nt
core
17
02
55
5
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
h-o
ut
AB
L4
31
430
0400
N,
131
190
0000
ED
ece
mb
er
20
06
10
Se
dim
en
tco
re2
47
37
00
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
hB
431
430
0400
N,
131
190
0000
EJu
ly2
00
61
1S
ed
ime
nt
core
51
27
68
0
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
hA
431
420
0100
N,
131
220
0000
EJu
ly,
20
06
12
Se
dim
en
tco
re4
70
70
50
NW
Ad
ria
tic
Se
a,
Ita
lyA
nco
na
,N
ort
h-o
ut
DE
R4
31
410
0400
N,
131
230
0000
EJu
ly2
00
61
3S
ed
ime
nt
core
38
65
79
0
NT
yrr
he
nia
nS
ea
,It
aly
Olb
iaA
401
550
4500
N,
91
300
2500
EM
ay
20
07
14
Se
dim
en
tco
re1
21
37
5
NT
yrr
he
nia
nS
ea
,It
aly
Olb
iaB
401
550
4500
N,
91
300
4500
EM
ay
20
07
15
Se
dim
en
tco
re5
91
77
NT
yrr
he
nia
nS
ea
,It
aly
Olb
iaG
401
550
1500
N,
91
300
1400
EM
ay
20
07
16
Se
dim
en
tco
re9
72
91
ST
yrr
he
nia
nS
ea
,It
aly
Vu
lca
no
381
250
0700
N,
141
570
1100
EM
ay
20
06
17
Se
dim
en
tco
re7
10
0
Ion
ian
Se
a,
Ita
lyS
t.1
C,
Sir
acu
sa3
71
030
5100
N,
1511
70
0100
EM
arc
h2
00
71
8S
ed
ime
nt
core
15
93
65
Ion
ian
Se
a,
Ita
lyS
t.2
C,
Sir
acu
sa3
71
030
2100
N,
1511
70
4000
EM
arc
h2
00
71
9S
ed
ime
nt
core
13
42
81
Ion
ian
Se
a,
Ita
lyS
t.E
,S
ira
cusa
371
030
5100
N,
151
170
01
EM
arc
h2
00
72
0S
ed
ime
nt
core
13
Ion
ian
Se
a,
Ita
lyA
qu
acu
lt.
B,
Sir
acu
sa3
71
020
0900
N,
151
170
20
EM
arc
h2
00
72
1S
ed
ime
nt
core
56
17
5
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,S
t.1
54
11
340
6900
N,
21
330
6800
EA
pri
l2
00
62
2T
rap
sam
ple
36
18
07
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,S
t.1
14
11
340
6900
N,
21
330
6800
EA
pri
l2
00
62
3T
rap
sam
ple
38
18
95
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,S
t.2
3-1
411
340
7300
N,
21
330
3400
EJu
ne
20
06
24
Se
dim
en
tco
re1
85
73
71
4
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,S
t.1
2+
St.
17
411
340
7200
N,
21
330
7000
E4
11
340
6000
N,
21
330
4400
E
Jun
e2
00
62
5S
ed
ime
nt
core
26
56
10
64
0
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,S
t.8
+S
t.1
34
11
340
6900
N,
21
330
4200
E4
11
340
6600
N,
21
330
7000
E
Jun
e2
00
62
6S
ed
ime
nt
core
25
91
10
36
4
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,2
411
340
7300
N,
21
330
3400
EO
cto
be
r2
00
62
7S
ed
ime
nt
core
24
15
48
30
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,1
/34
11
340
7300
N,
21
330
3400
EO
cto
be
r2
00
62
8S
ed
ime
nt
core
23
45
93
80
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,2
-4
411
340
7300
N,
21
330
3400
ED
ece
mb
er
20
06
29
Se
dim
en
tco
re2
83
71
13
48
NC
ata
lan
Se
a,
Sp
ain
Are
ny
s,1
/5/6
411
340
7300
N,
21
330
3400
ED
ece
mb
er
20
06
30
Se
dim
en
tco
re3
13
21
87
92
Ea
stA
tla
nti
c,S
pa
inB
aio
na
24
21
070
1200
N,
81
500
1500
EA
ug
ust
20
06
31
Tra
psa
mp
le1
14
05
70
0
Ea
stA
tla
nti
c,S
pa
inR
iad
eV
igo
a4
21
130
0800
N,
81
510
1400
EM
arc
h2
00
63
2S
ed
ime
nt
core
15
18
0
Ea
stA
tla
nti
c,S
pa
inR
iad
eV
igo
b4
21
130
0800
N,
81
510
1400
EM
arc
h2
00
63
3S
ed
ime
nt
core
11
41
36
8
Ea
stA
tla
nti
c,S
pa
inB
aio
na
28
421
070
1200
N,
81
500
1500
EJu
ly2
00
63
4T
rap
sam
ple
65
53
60
0
Ea
stA
tla
nti
c,S
pa
inB
aio
na
64
21
070
1200
N,
81
500
1500
ED
ece
mb
er
20
06
35
Se
dim
en
tco
re2
80
14
00
Ba
ltic
Se
a,
Sw
ed
en
Him
me
rsfj
ard
en
581
590
0700
N,
171
430
6000
EM
arc
h2
00
73
6S
ed
ime
nt
core
43
43
90
60
Ba
ltic
Se
a,
Fin
lan
dS
torf
jard
en
591
510
31
400
N,
2311
30
00
100
EM
ay
,2
00
73
7S
ed
ime
nt
core
88
01
79
21
35
Ba
ltic
Se
a,
Fin
lan
dLa
ng
ska
r5
91
470
33
400
N,
231;
200
28
400
EM
ay
20
07
38
Se
dim
en
tco
re6
34
22
85
39
0
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300290

Author's personal copyARTICLE IN PRESS
2.4. Microalgal cultures and genomic DNA extraction
Clonal cultures were used for taxon-specific PCR-based assays(Table 2). Cultures were kept in F/2 or L1 media (https://ccmp.bigelow.org/) at 2071 1C in a 12:12 h (light:dark) photoperiod.Illumination was provided by photon irradiance of 100mmolm–2 s–1. Culture subsamples containing 100,000 cells of eachtarget species were used for the genomic DNA extraction using aDNeasy Plant Kit (Qiagen, Valencia, CA, USA) according to Pennaet al. (2005).
2.5. PCR amplification detection assay
Genus (Alexandrium) and species-specific (Alexandrium minu-
tum, A. taylori, A. tamarense, A. catenella, L. polyedrum,P. reticulatum, F. japonica) primers were designed in the 5.8SrDNA-ITS regions and/or were derived from Penna et al. (2007).The primers for the genus Scrippsiella and species G. catenatum,G. nolleri and Scrippsiella hangoei were designed in the LSU and5.8S rDNA-ITS regions by using OLIGO software ver. 6.65 andBioEdit software ver. 7.0.5 (Table 3). The BLAST (Basic LocalAlignment Search Tool) analysis of alignment nucleotidesequences (Altschul et al., 1990) were as follows: primerG. catenatum: F0 ATTGCAGAATTCCGTGAA, total alignment length:18 nt, overall identity: 100% (18/18 nt). Primer G. nolleri
F0 TTTCAGCGATGGATGTCT, total alignment length: 18 nt, overallidentity: 100% (18/18 nt). Primer Scrippsiella sp. F0 CTGAAA
GGAAAGCGAATGGAG, total alignment length: 22 nt, overallidentity: 63.64% (14/22 nt). Primer S. hangoei F0 TTCGTTCGGAAGTGGTTT, total alignment length: 18 nt, overall identity:100% (18/18 nt). The LSU and 5.8S rDNA-ITS sequences forG. catenatum VGO-744 (AM998536), G. nolleri VGO-663(AM998534), G. nolleri 922I (AM998535), Scrippsiella rotunda
CBA-4 (AM998538), and S. trochoidea VGO-S3V (AM998537)were obtained in this study; the 5.8S rDNA-ITS sequences forS. hangoei were obtained from GenBank. Amplification fromsediment and sediment trap samples for the detection ofdifferent cyst taxa was performed in an Applied Biosystems DNAThermo Cycler 2720 (Foster City, CA, USA). PCR amplificationswere carried out directly using genus- or species-specific primersor by following two steps as follows: an initial PCR usingeukaryotic primers targeting the ITS-5.8S or LSU ribosomalgenes; then, a second (i.e. nested) PCR reaction, using genus-and species-specific primers on amplified products of ribosomalgenes (Fig. 2).
2.6. Direct PCR-based assay
Direct PCR using genus- or species-specific primers was asfollows: reaction tubes contained a 50mL mixture of 200–400mMof each dNTP; 0.2–0.4mM of each primer; 3.0–6.0 mM MgCl2; 1�Reaction Buffer (Eppendorf, Germany); 0.5–1� Enhancer Buffer(Eppendorf, Germany); 0.2–1.5mg/ul BSA (Table 4); 0.5 U of TaqPolymerase (Eppendorf, Germany) and 0.5–1.0 ng of sediment
Fig. 1. Locations of the sampling stations in the Mediterranean Sea, East Atlantic and Baltic Sea (see Table 1 for locality names).
Table 2Clonal cultures used together to test the specificity of genus and species specific PCR identification assay.
Species Strain Collection site Source
Alexandrium minutum VGO663 Tyrrhenian Sea, Olbia, Italy Fraga S.
Chaetoceros socialis CCMP 204 Catalan Sea, Marseille, France Berland B.
Fibrocapsa japonica CBA-1 Adriatic Sea, Ancona, Italy Totti C.
Gymnodinium catenatum GC12V East Atlantic, Vigo, Spain Figueroa R.I.
Gymnodinium nolleri 922I Kattegat, Denmark Ellegard M.
Scrippsiella hangoei SHTV-1 Baltic Sea, Tvarmine, Finland Kremp, A.
Scrippsiella rotunda CBA-4 Adriatic Sea, Pesaro, Italy Ingarao C.
Scrippsiella trochoidea VGOS3V East Atlantic, Vigo, Spain Bravo I.
Prymnesium parvum CCMP708 North Atlantic, Scotland, UK Droop M.
Skeletonema marinoi CBA-2 Adriatic Sea, Pesaro, Italy Penna A.
Ostreopsis ovata CNR-D1 Ligurian Sea, Genova, Italy Giacobbe M.G.
Pseudo-nitzschia spp. CBA-4 Adriatic Sea, Pesaro, Italy Capellacci S.
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300 291

Author's personal copyARTICLE IN PRESS
Table 3Oligonucleotide primers targeting the ITS-5.8S rDNA and LSU regions of different phytoplankton genera and species used in this study.
Target taxa Forward Primer Reverse primer Forward primer sequence (50- 30) -
Reverse primer sequence (50- 30) ’
Amplification
size (bp)
G + C % Primer positions
Alexandrium spp.a 5.8S-50 5.8S-30 F0-GCAADGAATGTCTTAGCTCAA 135 38.0 5.8S (50-30)
R0-GCAMACCTTCAAGMATATCCC 42.8 5.8S (30’50)
A. catenellaa ITS1c 5.8S-30 F0-AGCATGATTTGTTTTTCAAGC 226 33.3 ITS1 (50-30)
R0-GCAMACCTTCAAGMATATCCC 42.8 5.8S (30’50)
A. minutuma ITS1m 5.8S-30 F0-CATGCTGCTGTGTTGATGACC 212 52.3 ITS1 (50-30)
R0-GCAMACCTTCAAGMATATCCC 42.8 5.8S (30’50)
A. tamarensea 5.8S-50 ITS2t F0- TGTTACTTGTACCTTTGGGA 134 40.0 5.8S (50-30)
R0- ACAACACCCAGGTTCAAT 44.4 ITS2 (30’50)
A. tayloria ITS1t 5.8S-30 F0-TGGTGTTTGAATGCGGTTGT 297 45.0 ITS1 (50-30)
R0-GCAMACCTTCAAGMATATCCC 42.8 5.8S (30’50)
F. japonicaa Fibrocapsa F Fibrocapsa R F0-GCAGAGTCCAGCGAGTCATCA 180 57.1 5.8S (50-30)
R0-TAATATCCCAGACCACGCCAGA 50.0 ITS2 (30’50)
G. catenatumb Gymno-cat F Gymno-cat R F0-ATTGCAGAATTCCGTGAA 290 38.9 5.8S (50-30)
R0-GATCGATGCGAATGAAAC 44.4 ITS2 (30’50)
G. nollerib Gymno-nolleri F Gymno-nolleri R F0-TTTCAGCGATGGATGTCT 350 44.4 5.8S (50-30)
R0-TGAAGGCACGATTGACAC 50.0 ITS2 (30’50)
L. polyedruma L. Poly GF L. Poly GR F0-ATGTGTTCTCATCGGATGTTG 383 45.5 ITS1 (50-30)
R0-CACAGTACCGCTGCCACTTAAA 50.0 ITS2 (30’50)
P. reticulatuma P. ret F P. ret. R F0-TGCTGATTGCCATCTATCTT 382 40.0 ITS1 (50-30)
R0-CAGAAGCGCGTTAAACAG 50.0 ITS2 (30’50)
Scrippsiella sp.b Scripp F Scripp R F0-CTGAAAGGAAAGCGAATGGAG 221 47.6 LSU (D1-D2)
R0-CCGCAAATGAGTTCCAACAAG 47.6 LSU (D1-D2)
S. hangoeib Scripp-hangoei F Scripp-hangoei F F0-TTCGTTCGGAAGTGGTTT 274 44.4 ITS1 (50-30)
R0-CCCGAGAGCACCTTAACA 55.6 ITS2 (30’50)
Degenerate code D = A/G/T.
Degenerate code M = A/C.
Degenerate code Y = C/T.
Degenerate code R = A/G.
a From Penna et al. (2007).b designed in this study.
Fig. 2. Schematic representation of the PCR assays based on the use of genus and species-specific or eukaryotic primers. Direct or nested PCR assays were used for the PCR
amplification of target taxa cyst in sediment and trap samples.
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300292

Author's personal copyARTICLE IN PRESS
template DNA. PCR conditions were as follows: an initialdenaturation step of 10 min at 95 1C, 40 cycles of 30 s at 95 1C,30 s at 58 1C, and 30 s at 72 1C, and a final extension step of 7 minat 72 1C.
2.7. Nested PCR-based assay
For the nested PCR eukaryotic-specific primers targeting theITS-5.8S rDNA (Adachi et al., 1994) and LSU rDNA (Scholin et al.,1994) primers were used in the first PCR. Reaction tubes containeda 25mL mixture of: 200 mM of each dNTP; 2 pmol of each primer;4 mM MgCl2; 1�Reaction Buffer (Eppendorf, Germany); 0.5 U ofTaq Polymerase (Eppendorf, Germany) and 0.5–1.0 ng of sedimenttemplate DNA. PCR conditions were as follows: an initialdenaturation step of 10 min at 95 1C, 40 cycles of 30 s at 95 1C,30 s at 55 1C and 30 s at 72 1C, and a final extension step of 7 min at72 1C. The PCR-amplified products, which were derived from thePCR assay using universal eukaryotic primers and were not directlyvisualized on agarose gel, were used as template in nested PCRamplification with genus- or species-specific primers using 1mL ofthe first PCR-amplified product; the PCR conditions were asdescribed above for the direct PCR amplification with the exceptionof 35 cycles rather than 40.
2.8. Sequencing
The 5.8S gene and ITS regions of genomic DNA from microalgalspecies were amplified using ITSA/ITSB primers (Adachi et al.,1994). The PCR-amplified products were purified using the QiagenPurification Gel Extraction Kit (Qiagen, CA, California) according tothe manufacturer’s instructions and were then directly sequenced.PCR-amplified products were sequenced on an ABI PRISM 310Genetic Analyser (Applied Biosystems, USA) using the dyeterminator method according to the manufacturer’s instructions(ABI PRISM Big Dye Terminator Cycle Sequencing Ready ReactionKit, Applied Biosystems). Genus- and species-specific PCR-ampli-fied products obtained from sediment samples, such as those fromStation 23 (Arenys) for Alexandrium, Station 11 (Ancona, North B)for A. minutum, Station 8 (Ancona, North C) for A. tamarense,Station 27 (Arenys) for Scrippsiella, Station 18 (Siracusa) forL. polyedrum, Station 12 (Ancona, North A) for P. reticulatum,Station 9 (Ancona, North-in ABL) for F. japonica, Station 33 (Ria deVigo) for G. catenatum and Station 31 (Baiona) for G. nolleri, wereexcised from the gel, purified using QIAquick Gel extraction Kit(Qiagen) and directly sequenced.
2.9. Specificity and sensitivity of the PCR assay
The specificity and sensitivity of the PCR reactions had alreadybeen assessed for Alexandrium, A. minutum, A. tamarense,L. polyedrum, P. reticulatum and F. japonica in Penna et al. (2007).The specificity of the Scrippsiella spp., S. hangoei, G. catenatum andG. nolleri oligonucleotide primers were tested by amplifying targetDNA of the various microalgal genera and species from both clonalcultures of target species and sediment samples with designedspecies-specific primers. The Scrippsiella spp., S. hangoei, G. catena-
tum and G. nolleri genomic DNA was amplified with specific primersin the presence of non-target mixed genomic DNA obtained fromdifferent clonal strains; 1 ng of genomic DNA was used for eachbackground species. The 5.8S rDNA-ITS regions of Scrippsiella spp.,S. hangoei, G. catenatum and G. nolleri were amplified and cloned intothe Pcr 2.1 vector following the manufacturer’s instructions(Invitrogen, Carlsbad, CA, USA). Plasmids containing the target ITS-5.8S and LSU rDNA sequences and non-target mixed genomic DNAwere used as a positive and negative control, respectively.
To assess the sensitivity of the PCR assay, 100, 10 and 1 pg ofeach genomic DNA of Scrippsiella spp., S. hangoei, G. catenatum andG. nolleri were spiked into 10, 5 and 1 ng of the mixed cystpopulation DNA, providing background sediment DNA in themixture, which was then subjected to the PCR assays. Thesensitivity assays were done in triplicate for each amount ofgenomic DNA and taxon examined. The background sedimentsample was checked for the absence of the target taxa (Scrippsiella
spp., S. hangoei, G. catenatum and G. nolleri) by microscopy.Assessment of sensitivity was also performed on a retroviralcloned sequence in an external (non-microalgal) plasmid DNA(Casabianca et al., 1998); this non-microalgal plasmid DNA wasspiked as 104, 103, 102 and 101 copies into 10, 5 and 1 ng of mixedcyst population DNA. The PCR products were resolved on a 1.8%(w/v) agarose, 0.5X TBE buffer gel and were visualized by standardethidium bromide staining under UV light.
3. Results
3.1. Sample collection and cyst content identification by microscopy
A total of 38 sediment and sediment trap samples werecollected during the study in several coastal localities of theMediterranean and Baltic Sea and the north-western coast of Spain.A relatively high diversity of cyst taxa was found, with a total of 32cyst morphotypes identified by microscopy (Table 5). Dinoflagellate
Table 4Different concentrations of dNTPs, primers, MgCl2, Enhancer and BSA specific for each microalgal taxon used in the PCR based assay.
Taxa dNTPs (mM) Primers (mM) MgCl2 (mM) Enhancera BSA (mg/ml)
Alexandrium spp. 0.2 0.4 6.0 No added 0.2
Alexandrium catenella 0.2 0.4 6.0 0.5 0.2
Alexandrium minutum 0.2 0.4 6.0 1.0� 1.0
Alexandrium tamarense 0.4 0.4 6.0 No added 1.0
Alexandrium taylori 0.4 0.4 6.0 0.5 0.4
Gymnodinium catenatum 0.4 0.4 6.0 1x 1.5
Gymnodinium nolleri 0.4 0.4 6.0 No added 0.4
Fibrocapsa japonica 0.4 0.2 3.0 No added 0.2
Lingulodinium polyedrum 0.4 0.2 3.0 0.5 No added
Protoceratium reticulatum 0.4 0.4 3.0 No added 0.2
Scrippsiella spp. 0.1 0.4 6.0 0.5 0.2
Scrippsiella hangoei 0.4 0.4 3.0 No added 0.4
The Taq Master PCR enhancer is a buffer additive that improves thermostability (enzyme half-life) and processing of Taq DNA polymerase by stabilizing the enzyme during
PCR.
a The TaqMaster PCR enhancer is a buffer furnished with the Taq Polymerase (Eppendorf, Germany).
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300 293
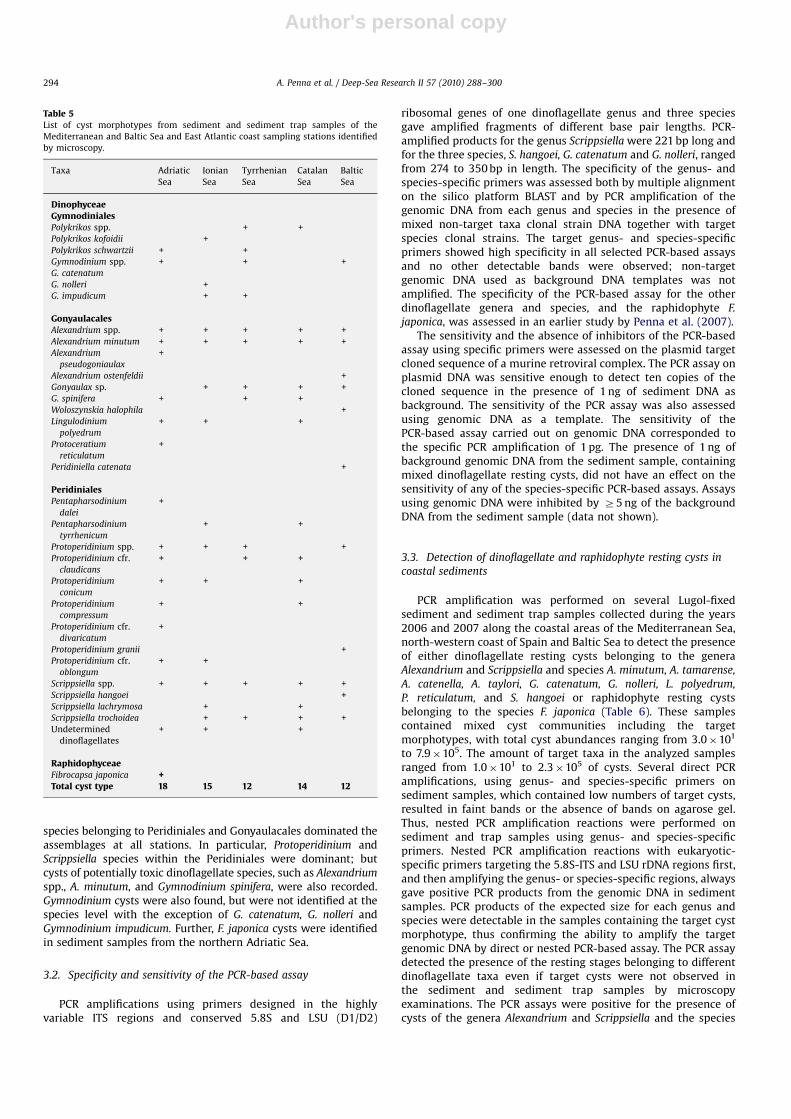
Author's personal copyARTICLE IN PRESS
species belonging to Peridiniales and Gonyaulacales dominated theassemblages at all stations. In particular, Protoperidinium andScrippsiella species within the Peridiniales were dominant; butcysts of potentially toxic dinoflagellate species, such as Alexandrium
spp., A. minutum, and Gymnodinium spinifera, were also recorded.Gymnodinium cysts were also found, but were not identified at thespecies level with the exception of G. catenatum, G. nolleri andGymnodinium impudicum. Further, F. japonica cysts were identifiedin sediment samples from the northern Adriatic Sea.
3.2. Specificity and sensitivity of the PCR-based assay
PCR amplifications using primers designed in the highlyvariable ITS regions and conserved 5.8S and LSU (D1/D2)
ribosomal genes of one dinoflagellate genus and three speciesgave amplified fragments of different base pair lengths. PCR-amplified products for the genus Scrippsiella were 221 bp long andfor the three species, S. hangoei, G. catenatum and G. nolleri, rangedfrom 274 to 350 bp in length. The specificity of the genus- andspecies-specific primers was assessed both by multiple alignmenton the silico platform BLAST and by PCR amplification of thegenomic DNA from each genus and species in the presence ofmixed non-target taxa clonal strain DNA together with targetspecies clonal strains. The target genus- and species-specificprimers showed high specificity in all selected PCR-based assaysand no other detectable bands were observed; non-targetgenomic DNA used as background DNA templates was notamplified. The specificity of the PCR-based assay for the otherdinoflagellate genera and species, and the raphidophyte F.
japonica, was assessed in an earlier study by Penna et al. (2007).The sensitivity and the absence of inhibitors of the PCR-based
assay using specific primers were assessed on the plasmid targetcloned sequence of a murine retroviral complex. The PCR assay onplasmid DNA was sensitive enough to detect ten copies of thecloned sequence in the presence of 1 ng of sediment DNA asbackground. The sensitivity of the PCR assay was also assessedusing genomic DNA as a template. The sensitivity of thePCR-based assay carried out on genomic DNA corresponded tothe specific PCR amplification of 1 pg. The presence of 1 ng ofbackground genomic DNA from the sediment sample, containingmixed dinoflagellate resting cysts, did not have an effect on thesensitivity of any of the species-specific PCR-based assays. Assaysusing genomic DNA were inhibited by Z5 ng of the backgroundDNA from the sediment sample (data not shown).
3.3. Detection of dinoflagellate and raphidophyte resting cysts in
coastal sediments
PCR amplification was performed on several Lugol-fixedsediment and sediment trap samples collected during the years2006 and 2007 along the coastal areas of the Mediterranean Sea,north-western coast of Spain and Baltic Sea to detect the presenceof either dinoflagellate resting cysts belonging to the generaAlexandrium and Scrippsiella and species A. minutum, A. tamarense,
A. catenella, A. taylori, G. catenatum, G. nolleri, L. polyedrum,
P. reticulatum, and S. hangoei or raphidophyte resting cystsbelonging to the species F. japonica (Table 6). These samplescontained mixed cyst communities including the targetmorphotypes, with total cyst abundances ranging from 3.0�101
to 7.9�105. The amount of target taxa in the analyzed samplesranged from 1.0�101 to 2.3�105 of cysts. Several direct PCRamplifications, using genus- and species-specific primers onsediment samples, which contained low numbers of target cysts,resulted in faint bands or the absence of bands on agarose gel.Thus, nested PCR amplification reactions were performed onsediment and trap samples using genus- and species-specificprimers. Nested PCR amplification reactions with eukaryotic-specific primers targeting the 5.8S-ITS and LSU rDNA regions first,and then amplifying the genus- or species-specific regions, alwaysgave positive PCR products from the genomic DNA in sedimentsamples. PCR products of the expected size for each genus andspecies were detectable in the samples containing the target cystmorphotype, thus confirming the ability to amplify the targetgenomic DNA by direct or nested PCR-based assay. The PCR assaydetected the presence of the resting stages belonging to differentdinoflagellate taxa even if target cysts were not observed inthe sediment and sediment trap samples by microscopyexaminations. The PCR assays were positive for the presence ofcysts of the genera Alexandrium and Scrippsiella and the species
Table 5List of cyst morphotypes from sediment and sediment trap samples of the
Mediterranean and Baltic Sea and East Atlantic coast sampling stations identified
by microscopy.
Taxa Adriatic
Sea
Ionian
Sea
Tyrrhenian
Sea
Catalan
Sea
Baltic
Sea
DinophyceaeGymnodinialesPolykrikos spp. + +
Polykrikos kofoidii +
Polykrikos schwartzii + +
Gymnodinium spp. + + +
G. catenatum
G. nolleri +
G. impudicum + +
GonyaulacalesAlexandrium spp. + + + + +
Alexandrium minutum + + + + +
Alexandrium
pseudogoniaulax
+
Alexandrium ostenfeldii +
Gonyaulax sp. + + + +
G. spinifera + + +
Woloszynskia halophila +
Lingulodinium
polyedrum
+ + +
Protoceratium
reticulatum
+
Peridiniella catenata +
PeridinialesPentapharsodinium
dalei
+
Pentapharsodinium
tyrrhenicum
+ +
Protoperidinium spp. + + + +
Protoperidinium cfr.
claudicans
+ + +
Protoperidinium
conicum
+ + +
Protoperidinium
compressum
+ +
Protoperidinium cfr.
divaricatum
+
Protoperidinium granii +
Protoperidinium cfr.
oblongum
+ +
Scrippsiella spp. + + + + +
Scrippsiella hangoei +
Scrippsiella lachrymosa + +
Scrippsiella trochoidea + + + +
Undetermined
dinoflagellates
+ + +
RaphidophyceaeFibrocapsa japonica +Total cyst type 18 15 12 14 12
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300294
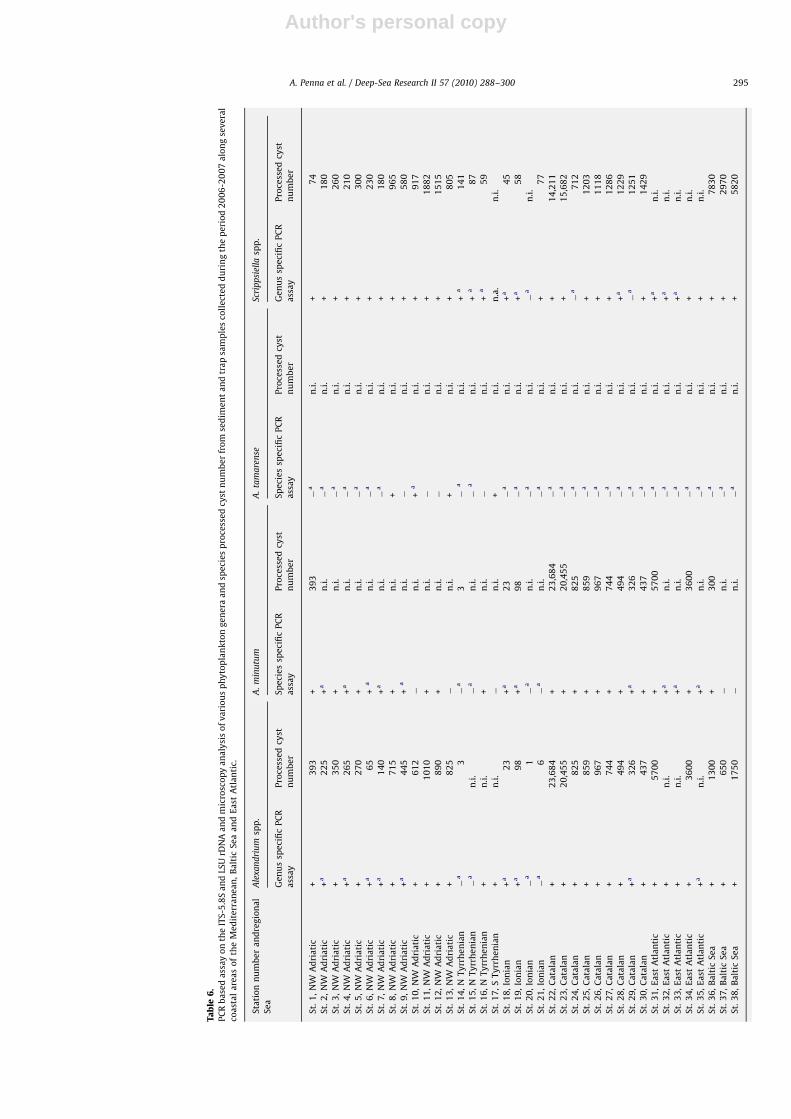
Author's personal copyARTICLE IN PRESS
Ta
ble
6.
PC
Rb
ase
da
ssay
on
the
ITS
-5.8
Sa
nd
LSU
rDN
Aa
nd
mic
rosc
op
ya
na
lysi
so
fv
ari
ou
sp
hy
top
lan
kto
ng
en
era
an
dsp
eci
es
pro
cess
ed
cyst
nu
mb
er
fro
mse
dim
en
ta
nd
tra
psa
mp
les
coll
ect
ed
du
rin
gth
ep
eri
od
20
06
-20
07
alo
ng
sev
era
l
coa
sta
la
rea
so
fth
eM
ed
ite
rra
ne
an
,B
alt
icS
ea
an
dE
ast
Atl
an
tic.
Sta
tio
nn
um
be
ra
nd
reg
ion
al
Se
a
Ale
xan
dri
um
spp
.A
.m
inu
tum
A.
tam
are
nse
Scri
pp
siel
lasp
p.
Ge
nu
ssp
eci
fic
PC
R
ass
ay
Pro
cess
ed
cyst
nu
mb
er
Sp
eci
es
spe
cifi
cP
CR
ass
ay
Pro
cess
ed
cyst
nu
mb
er
Sp
eci
es
spe
cifi
cP
CR
ass
ay
Pro
cess
ed
cyst
nu
mb
er
Ge
nu
ssp
eci
fic
PC
R
ass
ay
Pro
cess
ed
cyst
nu
mb
er
St.
1,
NW
Ad
ria
tic
+3
93
+3
93
�a
n.i
.+
74
St.
2,
NW
Ad
ria
tic
+a
22
5+
an
.i.
�a
n.i
.+
18
0
St.
3,
NW
Ad
ria
tic
+3
50
+n
.i.
�a
n.i
.+
26
0
St.
4,
NW
Ad
ria
tic
+a
26
5+
an
.i.
�a
n.i
.+
21
0
St.
5,
NW
Ad
ria
tic
+2
70
+n
.i.
�a
n.i
.+
30
0
St.
6,
NW
Ad
ria
tic
+a
65
+a
n.i
.�
an
.i.
+2
30
St.
7,
NW
Ad
ria
tic
+a
14
0+
an
.i.
�a
n.i
.+
18
0
St.
8,
NW
Ad
ria
tic
+7
15
+n
.i.
+n
.i.
+9
65
St.
9,
NW
Ad
ria
tic
+a
44
5+
an
.i.
�n
.i.
+5
80
St.
10
,N
WA
dri
ati
c+
61
2�
n.i
.+
an
.i.
+9
17
St.
11
,N
WA
dri
ati
c+
10
10
+n
.i.
�n
.i.
+1
88
2
St.
12
,N
WA
dri
ati
c+
89
0+
n.i
.�
n.i
.+
15
15
St.
13
,N
WA
dri
ati
c+
82
5�
n.i
.+
n.i
.+
80
5
St.
14
,N
Ty
rrh
en
ian
�a
3�
a3
�a
n.i
.+
a1
41
St.
15
,N
Ty
rrh
en
ian
�a
n.i
.�
an
.i.
�a
n.i
.+
a8
7
St.
16
,N
Ty
rrh
en
ian
+n
.i.
+n
.i.
�n
.i.
+a
59
St.
17
,S
Ty
rrh
en
ian
+n
.i.
�n
.i.
+n
.i.
n.a
.n
.i.
St.
18
,Io
nia
n+
a2
3+
a2
3�
an
.i.
+a
45
St.
19
,Io
nia
n+
a9
8+
a9
8�
an
.i.
+a
58
St.
20
,Io
nia
n�
a1
�a
n.i
.�
an
.i.
�a
n.i
.
St.
21
,Io
nia
n�
a6
�a
n.i
.�
an
.i.
+7
7
St.
22
,C
ata
lan
+2
3,6
84
+2
3,6
84
�a
n.i
.+
14
,21
1
St.
23
,C
ata
lan
+2
0,4
55
+2
0,4
55
�a
n.i
.+
15
,68
2
St.
24
,C
ata
lan
+8
25
+8
25
�a
n.i
.�
a7
12
St.
25
,C
ata
lan
+8
59
+8
59
�a
n.i
.+
12
03
St.
26
,C
ata
lan
+9
67
+9
67
�a
n.i
.+
11
18
St.
27
,C
ata
lan
+7
44
+7
44
�a
n.i
.+
12
86
St.
28
,C
ata
lan
+4
94
+4
94
�a
n.i
.+
a1
22
9
St.
29
,C
ata
lan
+a
32
6+
a3
26
�a
n.i
.�
a1
25
1
St.
30
,C
ata
lan
+4
37
+4
37
�a
n.i
.+
14
29
St.
31
,E
ast
Atl
an
tic
+5
70
0+
57
00
�a
n.i
.+
an
.i.
St.
32
,E
ast
Atl
an
tic
+n
.i.
+a
n.i
.�
an
.i.
+a
n.i
.
St.
33
,E
ast
Atl
an
tic
+n
.i.
+a
n.i
.�
an
.i.
+a
n.i
.
St.
34
,E
ast
Atl
an
tic
+3
60
0+
36
00
�a
n.i
.+
n.i
.
St.
35
,E
ast
Atl
an
tic
+a
n.i
.+
an
.i.
�a
n.i
.+
n.i
.
St.
36
,B
alt
icS
ea
+1
30
0+
30
0�
an
.i.
+7
83
0
St.
37
,B
alt
icS
ea
+6
50
�n
.i.
�a
n.i
.+
29
70
St.
38
,B
alt
icS
ea
+1
75
0�
n.i
.�
an
.i.
+5
82
0
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300 295
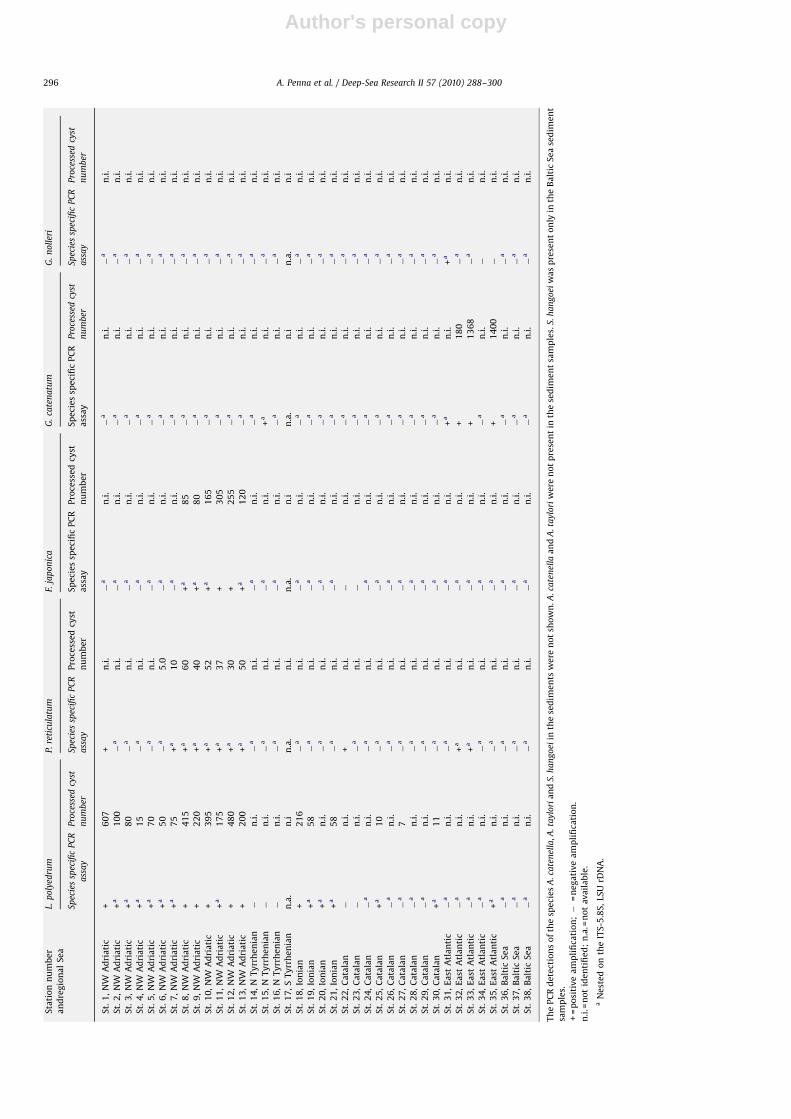
Author's personal copyARTICLE IN PRESS
Sta
tio
nn
um
be
r
an
dre
gio
na
lS
ea
L.p
oly
edru
mP.
reti
cula
tum
F.ja
po
nic
aG
.ca
ten
atu
mG
.n
oll
eri
Spec
ies
spec
ific
PC
R
ass
ay
Pro
cess
edcy
st
nu
mb
er
Spec
ies
spec
ific
PC
R
ass
ay
Pro
cess
ed
cyst
nu
mb
er
Sp
eci
es
spe
cifi
cP
CR
ass
ay
Pro
cess
ed
cyst
nu
mb
er
Sp
eci
es
spe
cifi
cP
CR
ass
ay
Pro
cess
edcy
st
nu
mb
er
Spec
ies
spec
ific
PC
R
ass
ay
Pro
cess
edcy
st
nu
mb
er
St.
1,
NW
Ad
ria
tic
+6
07
+n
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
2,
NW
Ad
ria
tic
+a
10
0�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
3,
NW
Ad
ria
tic
+a
80
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
4,
NW
Ad
ria
tic
+a
15
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
5,
NW
Ad
ria
tic
+a
70
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
6,
NW
Ad
ria
tic
+a
50
�a
5.0
�a
n.i
.�
an
.i.
�a
n.i
.
St.
7,
NW
Ad
ria
tic
+a
75
+a
10
�a
n.i
.�
an
.i.
�a
n.i
.
St.
8,
NW
Ad
ria
tic
+4
15
+a
60
+a
85
�a
n.i
.�
an
.i.
St.
9,
NW
Ad
ria
tic
+2
20
+a
40
+a
80
�a
n.i
.�
an
.i.
St.
10
,N
WA
dri
ati
c+
39
5+
a5
2+
a1
65
�a
n.i
.�
an
.i.
St.
11
,N
WA
dri
ati
c+
a1
75
+a
37
+3
05
�a
n.i
.�
an
.i.
St.
12
,N
WA
dri
ati
c+
48
0+
a3
0+
25
5�
an
.i.
�a
n.i
.
St.
13
,N
WA
dri
ati
c+
20
0+
a5
0+
a1
20
�a
n.i
.�
an
.i.
St.
14
,N
Ty
rrh
en
ian
�n
.i.
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
15
,N
Ty
rrh
en
ian
�n
.i.
�a
n.i
.�
an
.i.
+a
n.i
.�
an
.i.
St.
16
,N
Ty
rrh
en
ian
�n
.i.
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
17
,S
Ty
rrh
en
ian
n.a
.n
.in
.a.
n.i
.n
.a.
n.i
n.a
.n
.in
.a.
n.i
St.
18
,Io
nia
n+
21
6�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
19
,Io
nia
n+
a5
8�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
20
,Io
nia
n+
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
21
,Io
nia
n+
a5
8�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
22
,C
ata
lan
�n
.i.
+n
.i.
�n
.i.
�a
n.i
.�
an
.i.
St.
23
,C
ata
lan
�n
.i.
�a
n.i
.�
n.i
.�
an
.i.
�a
n.i
.
St.
24
,C
ata
lan
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
25
,C
ata
lan
+a
10
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
26
,C
ata
lan
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
27
,C
ata
lan
�a
7�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
28
,C
ata
lan
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
29
,C
ata
lan
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
30
,C
ata
lan
+a
11
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
St.
31
,E
ast
Atl
an
tic
�a
n.i
.�
an
.i.
�a
n.i
.+
an
.i.
+a
n.i
.
St.
32
,E
ast
Atl
an
tic
�a
n.i
.+
an
.i.
�a
n.i
.+
18
0�
an
.i.
St.
33
,E
ast
Atl
an
tic
�a
n.i
.+
an
.i.
�a
n.i
.+
13
68
�a
n.i
.
St.
34
,E
ast
Atl
an
tic
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�n
.i.
St.
35
,E
ast
Atl
an
tic
+a
n.i
.�
an
.i.
�a
n.i
.+
14
00
�n
.i.
St.
36
,B
alt
icS
ea
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
37
,B
alt
icS
ea
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
St.
38
,B
alt
icS
ea
�a
n.i
.�
an
.i.
�a
n.i
.�
an
.i.
�a
n.i
.
Th
eP
CR
de
tect
ion
so
fth
esp
eci
es
A.c
ate
nel
la,A
.ta
ylo
ria
nd
S.h
an
go
eiin
the
sed
ime
nts
we
ren
ot
sho
wn
.A.c
ate
nel
laa
nd
A.t
ayl
ori
wer
en
ot
pre
sen
tin
the
sed
ime
nt
sam
ple
s.S.
ha
ng
oei
wa
sp
rese
nt
on
lyin
the
Ba
ltic
Se
ase
dim
en
t
sam
ple
s.
+=
po
siti
ve
am
pli
fica
tio
n;�
=n
eg
ati
ve
am
pli
fica
tio
n.
n.i
.=n
ot
ide
nti
fie
d;
n.a
.=n
ot
ava
ila
ble
.
aN
est
ed
on
the
ITS
-5.8
S,
LS
UrD
NA
.
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300296
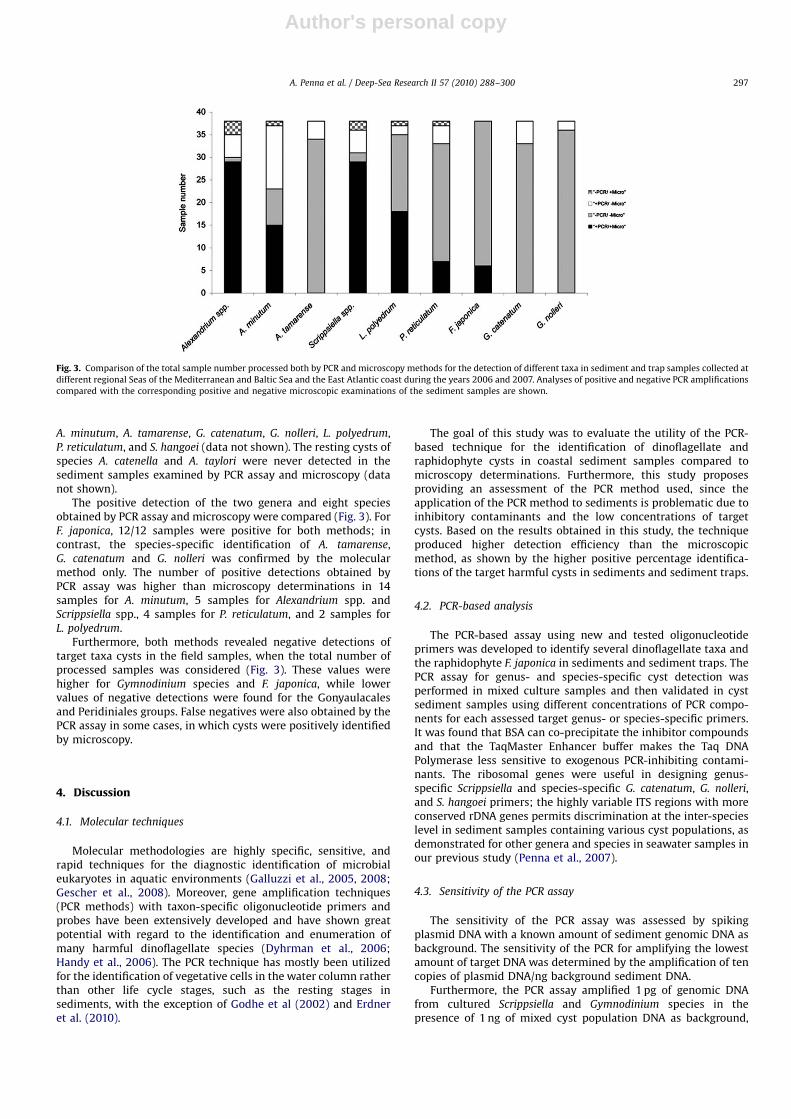
Author's personal copyARTICLE IN PRESS
A. minutum, A. tamarense, G. catenatum, G. nolleri, L. polyedrum,P. reticulatum, and S. hangoei (data not shown). The resting cysts ofspecies A. catenella and A. taylori were never detected in thesediment samples examined by PCR assay and microscopy (datanot shown).
The positive detection of the two genera and eight speciesobtained by PCR assay and microscopy were compared (Fig. 3). ForF. japonica, 12/12 samples were positive for both methods; incontrast, the species-specific identification of A. tamarense,G. catenatum and G. nolleri was confirmed by the molecularmethod only. The number of positive detections obtained byPCR assay was higher than microscopy determinations in 14samples for A. minutum, 5 samples for Alexandrium spp. andScrippsiella spp., 4 samples for P. reticulatum, and 2 samples forL. polyedrum.
Furthermore, both methods revealed negative detections oftarget taxa cysts in the field samples, when the total number ofprocessed samples was considered (Fig. 3). These values werehigher for Gymnodinium species and F. japonica, while lowervalues of negative detections were found for the Gonyaulacalesand Peridiniales groups. False negatives were also obtained by thePCR assay in some cases, in which cysts were positively identifiedby microscopy.
4. Discussion
4.1. Molecular techniques
Molecular methodologies are highly specific, sensitive, andrapid techniques for the diagnostic identification of microbialeukaryotes in aquatic environments (Galluzzi et al., 2005, 2008;Gescher et al., 2008). Moreover, gene amplification techniques(PCR methods) with taxon-specific oligonucleotide primers andprobes have been extensively developed and have shown greatpotential with regard to the identification and enumeration ofmany harmful dinoflagellate species (Dyhrman et al., 2006;Handy et al., 2006). The PCR technique has mostly been utilizedfor the identification of vegetative cells in the water column ratherthan other life cycle stages, such as the resting stages insediments, with the exception of Godhe et al (2002) and Erdneret al. (2010).
The goal of this study was to evaluate the utility of the PCR-based technique for the identification of dinoflagellate andraphidophyte cysts in coastal sediment samples compared tomicroscopy determinations. Furthermore, this study proposesproviding an assessment of the PCR method used, since theapplication of the PCR method to sediments is problematic due toinhibitory contaminants and the low concentrations of targetcysts. Based on the results obtained in this study, the techniqueproduced higher detection efficiency than the microscopicmethod, as shown by the higher positive percentage identifica-tions of the target harmful cysts in sediments and sediment traps.
4.2. PCR-based analysis
The PCR-based assay using new and tested oligonucleotideprimers was developed to identify several dinoflagellate taxa andthe raphidophyte F. japonica in sediments and sediment traps. ThePCR assay for genus- and species-specific cyst detection wasperformed in mixed culture samples and then validated in cystsediment samples using different concentrations of PCR compo-nents for each assessed target genus- or species-specific primers.It was found that BSA can co-precipitate the inhibitor compoundsand that the TaqMaster Enhancer buffer makes the Taq DNAPolymerase less sensitive to exogenous PCR-inhibiting contami-nants. The ribosomal genes were useful in designing genus-specific Scrippsiella and species-specific G. catenatum, G. nolleri,and S. hangoei primers; the highly variable ITS regions with moreconserved rDNA genes permits discrimination at the inter-specieslevel in sediment samples containing various cyst populations, asdemonstrated for other genera and species in seawater samples inour previous study (Penna et al., 2007).
4.3. Sensitivity of the PCR assay
The sensitivity of the PCR assay was assessed by spikingplasmid DNA with a known amount of sediment genomic DNA asbackground. The sensitivity of the PCR for amplifying the lowestamount of target DNA was determined by the amplification of tencopies of plasmid DNA/ng background sediment DNA.
Furthermore, the PCR assay amplified 1 pg of genomic DNAfrom cultured Scrippsiella and Gymnodinium species in thepresence of 1 ng of mixed cyst population DNA as background,
Fig. 3. Comparison of the total sample number processed both by PCR and microscopy methods for the detection of different taxa in sediment and trap samples collected at
different regional Seas of the Mediterranean and Baltic Sea and the East Atlantic coast during the years 2006 and 2007. Analyses of positive and negative PCR amplifications
compared with the corresponding positive and negative microscopic examinations of the sediment samples are shown.
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300 297
Author's personal copyARTICLE IN PRESS
while the taxon-specific PCR-based assay was inhibited in thepresence of 10 and 5 ng of sediment genomic DNA as background.It is likely that the presence of some inhibitory substances,which are not completely eliminated from sediments duringextraction and purification using the commercial kit, cannegatively affect the PCR reaction. Inhibitor substances, such ashumic acids, polyphenols, polysaccharides and metals, andnuclease activity, are the major concern when extracting genomicDNA from marine sediments (Stults et al., 2001). In fact, the co-precipitation of compounds that inhibit PCR confuses themolecular analyses of field samples by producing false nega-tive results (Tebbe and Vahjen, 1993). The most common strategyused to overcome PCR inhibition by contaminants is dilution to alower concentration of the template DNA. In this study, weapplied a total DNA extraction and purifying procedures using acommercial kit to eliminate the potential inhibitors of the PCRreaction.
4.4. Specificity of PCR assay
The PCR-based assay was effective for the qualitative detectionof the target cysts in sediment and sediment trap samples. TotalDNA extraction was accomplished with a commercial kit that waseffective in obtaining the total quantity of DNA visible on agarosegel, which was then quantifiable for subsequent PCR reactions.The commercial kit was also successful in removing majorinhibitors, since the direct PCR amplification using either taxon-specific or eukaryotic-specific primers targeting the LSU or 5.8SrDNA and ITS was positive. Nevertheless, absence of PCRamplifications were also obtained as direct PCR using genus-and species-specific primers or eukaryotic primers often resultedin very faint bands or no observed bands at all, thus yieldingapparent false negatives. A nested taxon-specific PCR amplifica-tion of the first amplified fragment using eukaryotic-specificprimers, always gave a positive PCR reaction. The lack ofPCR-amplified fragments was observed in those sediment samplescontaining a low fraction of target taxa cysts (n=7) (Fig. 3). Directand nested PCR-based amplifications with isolated single cystshave been achieved by minimizing cyst DNA loss by the Bolch(2001) and Ki and Han (2007) methods. However, a suitablemethod for the rapid and specific detection of dinoflagellate cystsin numerous sediment samples during monitoring activity isrequired. The single cyst PCR technique requires the manipulationand isolation of the single cysts from the sediment samples undera microscope, and a single isolated cyst may be sufficient to carryout only a few PCR amplification reactions.
4.5. Molecular PCR and microscopic determinations of sediment
samples
PCR detection of the target species was compared with themicroscopic analyses of the same sediment and sediment trapsamples. The molecular technique provided a higher positivedetection rate of target cysts than microscopy. Target cysts weredetected by PCR in sediment samples presenting mixed restingstages at low concentrations or concentrations below the detec-tion limit of the microscopy method, and when it was not possibleto recognize the target cyst taxonomically. Based on thePCR-based assay higher rates of positive detection were observedfor the genera Alexandrium and Scrippsiella, and speciesA. minutum, A. tamarense, G. catenatum, G. nolleri, L. polyedrum
and P. reticulatum due to a widespread distribution. Theraphidophyte F. japonica cysts were identified equally in sedimentsamples by the two methods. PCR analysis detected 100% ofA. tamarense, G. nolleri and G. catenatum cysts in sedimentsamples. This is a relevant finding as the different morphotypecysts of Gymnodinium spp. and Alexandrium spp. are quite difficultto recognize and distinguish (Bolch et al., 1999; Bravo et al., 2006).
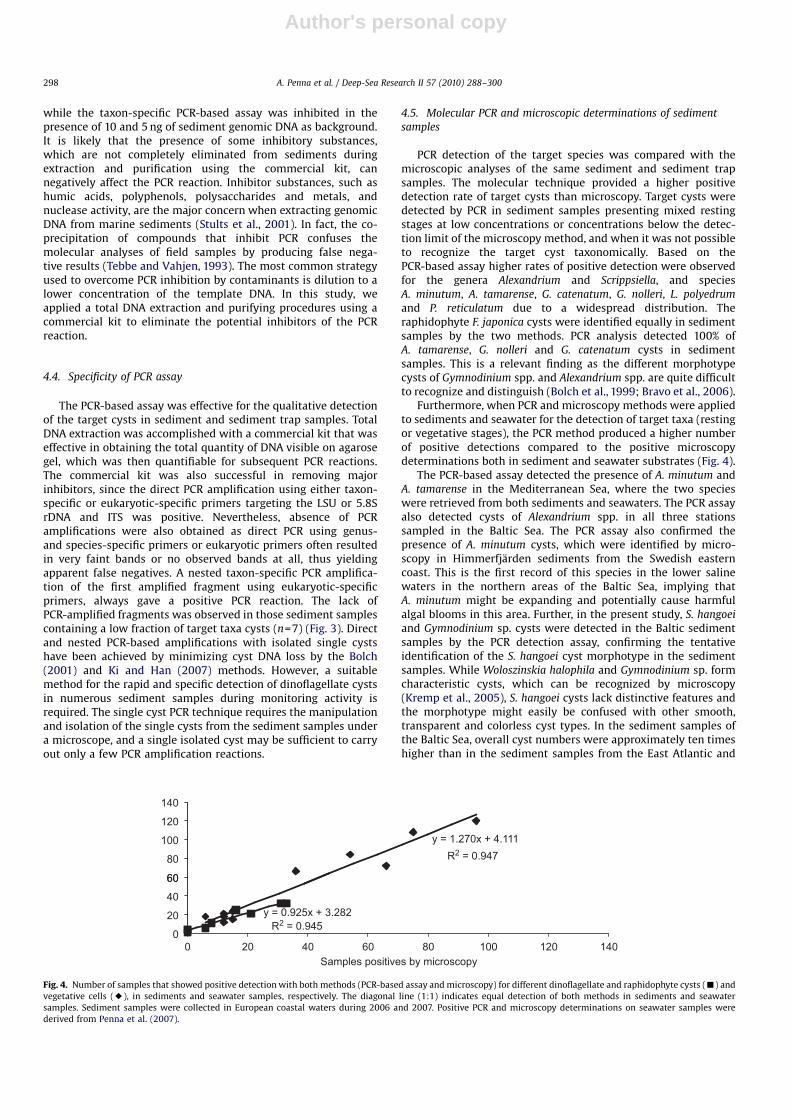
Furthermore, when PCR and microscopy methods were appliedto sediments and seawater for the detection of target taxa (restingor vegetative stages), the PCR method produced a higher numberof positive detections compared to the positive microscopydeterminations both in sediment and seawater substrates (Fig. 4).
The PCR-based assay detected the presence of A. minutum andA. tamarense in the Mediterranean Sea, where the two specieswere retrieved from both sediments and seawaters. The PCR assayalso detected cysts of Alexandrium spp. in all three stationssampled in the Baltic Sea. The PCR assay also confirmed thepresence of A. minutum cysts, which were identified by micro-scopy in Himmerfjarden sediments from the Swedish easterncoast. This is the first record of this species in the lower salinewaters in the northern areas of the Baltic Sea, implying thatA. minutum might be expanding and potentially cause harmfulalgal blooms in this area. Further, in the present study, S. hangoei
and Gymnodinium sp. cysts were detected in the Baltic sedimentsamples by the PCR detection assay, confirming the tentativeidentification of the S. hangoei cyst morphotype in the sedimentsamples. While Woloszinskia halophila and Gymnodinium sp. formcharacteristic cysts, which can be recognized by microscopy(Kremp et al., 2005), S. hangoei cysts lack distinctive features andthe morphotype might easily be confused with other smooth,transparent and colorless cyst types. In the sediment samples ofthe Baltic Sea, overall cyst numbers were approximately ten timeshigher than in the sediment samples from the East Atlantic and
y = 1.270x + 4.111
60
80
100
120
140
y = 0.925x + 3.282
0
20
40
60
0Samples positives by microscopy
20 40 60 80 100 120 140
R2 = 0.945
R2 = 0.947
Fig. 4. Number of samples that showed positive detection with both methods (PCR-based assay and microscopy) for different dinoflagellate and raphidophyte cysts (’) and
vegetative cells (E), in sediments and seawater samples, respectively. The diagonal line (1:1) indicates equal detection of both methods in sediments and seawater
samples. Sediment samples were collected in European coastal waters during 2006 and 2007. Positive PCR and microscopy determinations on seawater samples were
derived from Penna et al. (2007).
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300298

Author's personal copyARTICLE IN PRESS
the Mediterranean Sea. Such high total cyst concentrations aredue to the dominance of W. halophila, which encysts in largenumbers after the spring bloom, resulting in exceptionally highcyst fluxes in the sediments (Kremp and Heiskanen, 1999). Thegenus Scrippsiella was also highly abundant among the cyst-forming dinoflagellates in the examined areas; Scrippsiella cellsare commonly found in coastal waters of these regions (Montresoret al., 1994). The life strategy of this genus includes a shortdormancy period with rapid turnover rates in cyst/vegetative cells,which may explain the formation of the bloom and its abundancealong with its long-term presence in a widespread area.
The identification of Gymnodinium cysts was quite difficult dueto the many almost indistinguishable morphotypes, some ofwhich correspond to the same species (Matsuoka and Fukuyo,2003). Within the genus Gymnodinium, G. catenatum is a toxicspecies producing a reticulated cyst, which makes it easier todistinguish from other cysts, but it is difficult to differentiate fromG. nolleri cysts (Bolch et al., 1999). This species is reported to berecurrent on the western Atlantic coast of Spain (Bravo andAnderson, 1994) and in the Alboran Sea in the Mediterranean Sea(Bravo et al., 1990). Moreover, it has appeared sporadically in someregions of the western Mediterranean Sea and the cysts weredetected during occasional surveys in southern western Mediter-ranean sediments (Calbet et al., 2002). In this study, G. catenatum
cysts were found in the Spanish Atlantic sediments by microscopyand PCR assay, and also by the PCR method in one locality of theTyrrhenian Sea where the cysts had never been detected bymicroscopy.
With regards to the raphidophyte species, reports on F. japonica
cysts are found in the in vitro study of De Boer et al. (2004), andthe identification of the cyst morphotype is limited to marinesediments (Yoshimatsu, 1987). In the Mediterranean Sea,F. japonica blooms have only been registered in the Tyrrhenianand north-western Adriatic Seas (Cucchiari et al., 2008). In thislatter coastal area, F. japonica produces abundant blooms duringthe summer period (Totti C., pers. comm.). In the present study,cysts of F. japonica were detected in the north-western Adriaticsediment samples by both methods, proving for the first time thepresence of F. japonica resting stages in the area where this speciescauses blooms. Moreover, the taxonomic identification of cystmorphotype by microscopy was confirmed by molecular assay.The cysts of the toxic species L. polyedrum and P. reticulatum werefound at almost all sampling stations, since they are widespreadin European coastal seawaters.
5. Conclusions
In this study the specificity and sensitivity of the PCR-basedtechnique for the detection of target cysts in marine sediment andsediment trap samples was demonstrated. The PCR methodpermitted higher detection efficiency than the microscopic method,illustrated by the higher positive percentage identification of theharmful target cysts in sediments. Knowledge of species composi-tion is important to understand bloom events in the coastal areas;it is also crucial to have information on the presence of novel andpotentially introduced taxa and to confirm the recurrent events of aspecies. In the future, the PCR method could be used for mappingthe distribution of target species cysts in coastal sediments,particularly given its high specificity and sensitivity.
Acknowledgments
We thank S. Casabianca and A. Casabianca for molecularanalysis assistance and suggestions; S. Fraga for culture strains;
S. Capellacci for technical assistance. Thanks to the two anon-ymous reviewers who made an effort in improving the paper. Thisstudy was financed by the EU funded Research Project SEED(GOCE-CT-2005-003875). E. Garces was sustained by a Ramon yCajal contract from the Spanish Ministry of Science and Education.
References
Adachi, M., Sako, Y., Ispida, Y., 1994. Restriction fragment length polymorphism ofribosomal DNA internal transcribed spacer and 5.8S regions in JapaneseAlexandrium species (Dinophyceae). Journal of Phycology 30, 857–863.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic localalignment search tool. Journal of Molecular Biology 215, 403–410.
Amorim, A., Dale, B., Godinho, R., Brotas, V., 2001. Gymnodinium catenatum-likecysts (Dinophyceae) in recent sediments from the coast of Portugal. Phycologia40 (6), 572–582.
Andersen, P., Throndsen, J., 2003. Estimating cell number. In: Hallegraeff, G.M.,Anderson, D.M., Cembella, A.D. (Eds.), Manual on Harmful Marine Microalgae.UNESCO, Paris, pp. 99–129.
Angl�es, S., Garces, E., Jordi, A., Basterretxea, G., Palanques, A., 2010. Alexandriumminutum resting cyst distribution dynamics in a confined site. Deep-SeaResearch Part II 57 (3–4), 210–221.
Bolch, C.J., 2001. PCR protocol for genetic identification of dinoflagellates directlyfrom single cysts and plankton cells. Phycologia 40 (2), 162–167.
Bolch, C.J.S., Negri, A.P., Hallegraeff, G.M., 1999. Gymnodinium microreticulatum sp.nov. (Dinophyceae): a naked, microreticulate cyst-producing dinoflagellate,distinct from Gymnodinium catenatum and Gymnodinium nolleri. Phycologia 38(4), 301–313.
Bowers, H.A., Trice, M.T., Magnien, R.E., Goshorn, D.M., Michael, B., Schaefer, E.F.,Rublee, P.A., Oldach, D.W., 2006. Detection of Pfiesteria spp. by PCR in surfacesediments collected from Chesapeake Bay tributaries (Maryland). HarmfulAlgae 5 (4), 342–351.
Bravo, I., Garces, E., Diogene, J., Fraga, S., Sampedro, N., Figueroa, R.I., 2006.Resting cysts of the toxigenic dinoflagellate genus Alexandrium in recentsediments from the Western Mediterranean coast, including first descriptionof cysts of A. kutnerae and A. peruvianum. European Journal of Phycology 41 (4),293–302.
Bravo, I., Anderson, D.M., 1994. The effects of temperature, growth mediumand darkness on excystment and growth of the toxic dinoflagellate Gymno-dinium catenatum from northwest Spain. Journal of Plankton Research 16 (5),513–525.
Bravo, I., Reguera, B., Martınez, A., Fraga, S., 1990. First report of Gymnodiniumcatenatum in the Spanish Mediterranean coast. In: Graneli, E., Anderson, D.M.,Edler, L., Sundstrom, B. (Eds.), Toxic Marine Phytoplankton. Elsevier, New York,pp. 449–452.
Calbet, A., Broglio, E., Saiz, E., Alcaraz, M., 2002. Low grazing impact ofmesozooplankton on the microbial communities of the Alboran Sea: a possiblecase of inhibitory effects by the toxic dinoflagellate Gymnodinium catenatum.Aquatic Microbial Ecology 26 (3), 235–246.
Casabianca, A., Vallanti, G., Magnani, M., 1998. Competitive PCR for quantificationof BM5d proviral DNA in mice with AIDS. Journal of Clinical Microbiology 36,2371–2374.
Coyne, K.J., Handy, S.M., Demir, E., Whereat, E.B., Hutchins, D.A., Portune, K.J.,Doblin, M.A., Cary, S.C., 2005. Improved quantitative real-time PCR assays forenumeration of harmful algal species in field samples using an exogenous DNAreference standard. Limnology and Oceanography: Methods 3, 381–391.
Cucchiari, E., Guerrini, F., Penna, A., Totti, C., Pistocchi, R., 2008. Effect of salinity,temperature, organic and inorganic nutrients on growth of cultured Fibrocapsajaponica (Raphidophyceae) from the northern Adriatic Sea. Harmful Algae 7,405–414.
De Boer, M.K., Van Rijssel, M., Vrieling, E.G., 2004. Morphology of Fibrocapsajaponica cysts formed under laboratory conditions. In: Steidinger, K.A.,Landsberg, J.H., Tomas, C.R., Vargo, G.A. (Eds.), Harmful Algae 2002. UNESCO,Paris, pp. 455–457.
Dyhrman, S.T., Erdner, D., La Du, J., Galac, M., Anderson, D.M., 2006. Molecularquantification of toxic Alexandrium fundyense in the Gulf of Maine using real-time PCR. Harmful Algae 5 (3), 242–250.
Erdner, D.L., Percy, L., Lewis, J., Anderson, D.M., 2010. A quantitative real-timePCR assay for the identification and enumeration of Alexandrium cysts inmarine sediments. Deep-Sea Research Part II 57 (3–4), 279–287.
Edvardsen, B., Imai, I., 2006. The ecology of harmful flagellates within Prymne-siophyceae and Raphidophyceae. In: Graneli, E., Turner, J.T. (Eds.), Ecology ofHarmful Algae. Springer-Verlag, Berlin, pp. 67–79.
Galluzzi, L., Bertozzini, E., Penna, A., Perini, F., Pigalarga, A., Graneli, E., Magnani, M.,2008. Detection and quantification of Prymnesium parvum (Haptophyceae) byreal-time PCR. Letters in Applied Microbiology 46, 261–266.
Galluzzi, L., Penna, A., Bertozzini, E., Vila, M., Garces, E., Giacobbe, M.G., Prioli, S.,Magnani, M., 2005. Development of a qualitative PCR method for theAlexandrium (Dinophyceae) detection in contaminated mussels (Mytilusgalloprovincialis). Harmful Algae 4 (6), 973–983.
Galluzzi, L., Penna, A., Bertozzini, E., Vila, M., Garces, E., Magnani, M., 2004.Development of a real-time PCR assay for rapid detection and quantification of
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300 299

Author's personal copyARTICLE IN PRESS
Alexandrium minutum (a dinoflagellate). Applied and Environmental Micro-biology 70, 1199–1206.
Garces, E., Bravo, I., Vila, M., Figueroa, R.I., Maso, M., Sampedro, N., 2004.Relationship between vegetative cells and cyst production during Alexandriumminutum bloom in Arenys de Mar harbour (NW Mediterranean). Journal ofPlankton Research 26 (6), 637–645.
Garces, E., Zingone, A., Montresor, M., Reguera, B., Dale, B., 2002. Lifehab: LifeHistories of Microalgal Species Causing Harmful Blooms. Office for the OfficialPublications of the European Communities, Luxembourg, pp. 1-189.
Gescher, C., Metfies, K., Medlin, L.K., 2008. The ALEX CHIP-development of a DNAchip for identification and monitoring of Alexandrium. Harmful Algae 7 (4),485–494.
Giacobbe, M.G., Penna, A., Gangemi, E., Maso, M., Garces, E., Fraga, S., Bravo, I.,Azzaro, F., Decembrini, F., Penna, N., 2007. Recurrent high-biomass blooms ofAlexandrium taylorii (Dinophyceae), a HAB species expanding in the Mediter-ranean Sea. Hydrobiologia 580, 125–133.
Godhe, A., Cusack, C., Pedersen, J., Andersen, P., Anderson, D.M., Bresnan, E.,Cembella, A., Dahl, E., Diercks, S., Elbrachter, M., Edler, L., Galluzzi, L., Gescher,C., Gladstone, M., Karlson, B., Kulis, D., LeGresley, M., Lindahl, O., Marin, R.,McDermott, G., Medlin, L.K., Naustvoll, L.J., Penna, A., T +obe, K., 2007.Intercalibration of classical and molecular techniques for identification ofAlexandrium fundyense (Dinophyceae) and estimation of cell densities.Harmful Algae 6 (1), 56–72.
Godhe, A., Rehnstam-Holm, A-S., Karunasagar, I., Karunasagar, I., 2002. PCRdetection of dinoflagellate cysts in field sediment samples from tropic andtemperate environments. Harmful Algae 1 (4), 361–373.
Handy, S.M., Hutchins, D.A., Cary, S.C., Coyne, K., 2006. Simultaneous enume-ration of multiple raphidophyte species by quantitative real-time PCR:capabilities and limitations. Limnology and Oceanography: methods 4,193–204.
Kamikawa, R., Nagai, S., Hosoi-Tanabe, S., Itakura, S., Yamaguchi, M., Uchida, Y.,Baba, T., Sako, Y., 2007. Application of real-time PCR assay for detection andquantification of Alexandrium tamarense and Alexandrium catenella cysts frommarine sediments. Harmful Algae 6 (3), 413–420.
Ki, J.S., Han, M.S., 2007. Rapid molecular identification of the harmful freshwaterdinoflagellate Peridinium in various life stages using genus-specific single-cellPCR. Journal of Applied Phycology 19, 467–470.
Kremp, A., Elbrachter, M., Schweikert, M., Wolny, J., Gottschling, M., 2005.Woloszynskia halophila (Biecheler) comb. nov.—a bloom forming cold-waterdinoflagellate co-occurring with Scrippsiella hangoei (Dinophyceae) in theBaltic Sea. Journal of Phycology 41 (3), 629–642.
Kremp, A., Heiskanen, A.S., 1999. Sexuality and cyst formation of the spring-bloomdinoflagellate Scrippsiella hangoei in the coastal northern Baltic Sea. MarineBiology 134 (4), 771–777.
Matsuoka, K., Fukuyo, Y., 2003. Taxonomy of cysts. In: Hallegraeff, G.M., Anderson,D.M., Cembella, A.D. (Eds.), Manual on Harmful Marine Microalgae. UNESCO,Paris, pp. 563–592.
McGillicuddy, D.J., Signell, R.P., Stock, C.A., Keafer, B.A., Keller, M.D., Hetland, R.D.,Anderson, D.M., 2003. A mechanism for offshore initiation of harmfulalgal blooms in the coastal Gulf of Maine. Journal of Plankton Research 25,1131–1138.
Montresor, M., Montesarchio, E., Marino, D., Zingone, A., 1994. Calcareousdinoflagellate cysts in marine sediments of the Gulf of Naples (MediterraneanSea). Review of Paleobotany and Palynology 84, 45–56.
Penna, A., Bertozzini, E., Battocchi, C., Galluzzi, L., Giacobbe, M.G., Vila, M., Garces,E., Lugli�e, A., Magnani, M., 2007. Monitoring of HAB species in theMediterranean Sea through molecular methods. Journal of Plankton Research29 (1), 19–38.
Penna, A., Garces, E., Vila, M., Giacobbe, M.G., Fraga, S., Lugli�e, A., Bravo, I.,Bertozzini, E., Vernesi, C., 2005. Alexandrium catenella (Dinophyceae), a toxicribotype expanding in the NW Mediterranean Sea. Marine Biology 148 (1), 13–23.
Saito, K., Drgon, T., Robledo, J., Krupatkita, D., Vasta, G., 2002. Characterization ofthe rRNA locus of Pfiesteria piscicida and development of standard andquantitative PCR-based detection. Applied and Environmental Microbiology 68(11), 5394–5407.
Satta, C., Angles, S., Garces, E., Luglie, A., Padedda, B., Sechi, N., 2010. Dinoflagellatecysts in recent sediments from two semi-enclosed areas of the WesternMediterranean Sea subject to high human impact. Deep Sea Research II 57(3–4), 256–267.
Scholin, C.A., Herzog, M., Sogin, M., Anderson, D.M., 1994. Identification of group-and strain-specific genetic markers from globally distributed Alexandrium(Dinophyceae). II. Sequence analysis of fragments of the LSU rRNA gene.Journal of Phycology 30, 999–1011.
Smayda, T.J., 2007. Reflections on the ballast water dispersal—harmful algal bloomparadigm. Harmful Algae 6 (4), 601–622.
Steidinger, K.A., Garces, E., 2006. Importance of life cycles in the ecology of harmfulmicroalgae. In: Graneli, E., Turner, J.T. (Eds.), Ecology of Harmful Algae.Springer-Verlag, Berlin, pp. 37–49.
Stults, J.R., Snoeyenbos-West, O., Methe, B., Lovley, D.R., Chandler, D.P., 2001.Application of the 50 fluorogenic exonuclease assay (Taqman) for quantitativeribosomal DNA and rRNA analysis in sediments. Applied EnvironmentalMicrobiology 67, 2781–2789.
Tebbe, C.C., Vahjen, W., 1993. Interference of humic acids and DNA extracted directlyfrom soil in detection and transformation of recombinant DNA from bacteriaand a yeast. Applied and Environmental Microbiology 59, 2657–2665.
Yoshimatsu, S., 1987. The cysts of Fibrocapsa japonica (Raphidophyceae) found inbottom sediment in Harima Nada, eastern Inland Sea of Japan. Bulletin ofPlankton Society Japan 34, 25–31.
A. Penna et al. / Deep-Sea Research II 57 (2010) 288–300300





























