Embed Size (px)
Citation preview

Cancer Genetics 205 (2012) 285e294
Detection of TET2 abnormalities by fluorescence insitu hybridization in 41 patients withmyelodysplastic syndromeIrene Dambruoso*, Marina Boni, Marianna Rossi, Patrizia Zappasodi,Celeste Calvello, Rita Zappatore, Paola Maria Cavigliano, Ilaria Giardini,Barbara Rocca, Marilena Caresana, Cesare Astori, Mario Cazzola,Carlo Castagnola, Paolo BernasconiDivision of Hematology, Fondazione IRCCS Policlinico San Matteo, University of Pavia, Pavia, Italy
Received Dece
29, 2012; accepted
* Corresponding
E-mail address:
2210-7762/$ - see
doi:10.1016/j.cance
TET2 haplo-insufficiency occurs through different molecular mechanisms and is promptly re-
vealed by array comparative genomic hybridization, single nucleotide polymorphism (SNP)
array, and next-generation sequencing (NGS). Fluorescence in situ hybridization (FISH) can
effectively demonstrate TET2 deletions and is often used to validate molecular results. In the
present study 41 MDS patients with and without 4q abnormalities were analyzed with a series
of bacterial artificial chromosome (BAC) probes spanning the 4q22.3-q25 region. On conven-
tional cytogenetic (CC) studies, a structural defect of the long arm of chromosome 4 (4q) was
observed in seven patients. In three, one each with a t(1;4)(p21;q24), an ins(5;4)(q23;q24qter),
and a t(4;17)(q31;p13) as the sole chromosomal abnormality, FISH with the RP11-356L5 and
RP11-16G16 probes, which cover the TET2 locus, produced one signal only. Unexpectedly, this
same result was achieved in 3 of the remaining 34 patients. Thus, a TET2 deletion was observed
in a total of six patients (14.6%). TET2 deletion was not correlated with any particular clinical
findings or outcome. These findings demonstrate that 1) FISH is an effective and economical
method to reveal cryptic abnormalities of band 4q22-q24 resulting in TET2 deletions; 2) in these
patients, TET2 deletion is the unifying genetic event; and 3) the different breakpoints within the
4q22-q25 region suggest that deletions are not mediated by repetitive sequences.
Keywords MDS, conventional cytogenetics, FISH, TET2
ª 2012 Elsevier Inc. All rights reserved.
Myeloproliferative disorder,myelodysplastic syndrome (MDS),and de novo/secondary acute myeloid leukemia (AML) aredistinct clonal hematopoietic stem cell disorders with over-lapping features (1). Recently, it has been observed that thesehematological disorders may share different alterations of thesame tumor suppressor gene (TSG), the TET2 (ten-eleventranlocation 2) gene, which maps at band 4q24 (2,3). In 2005,a fluorescence in situ hybridization (FISH) study provideda relevant contribution to the identification of this gene lesion(4). In fact, it disclosed that apparently balanced translocationsinvolving band 4q24 are always associated with the deletion of
mber 19, 2011; received in revised form March
March 29, 2012.
author.
front matter ª 2012 Elsevier Inc. All rights reserved.
rgen.2012.03.004
a common segment that is approximately 0.5 Mb in size.Subsequently, array comparative genomic hybridization(aCGH), single nucleotide polymorphism (SNP) arrays, andnext-generation sequencing (NGS) revealed that this same4q24 region is targeted by copy number imbalances (CNIs),uniparental disomies (UPDs), and mutations in varioushematological disorders with a frequency of 10e25% (5e19).Another recent study reported that TET2 deletions area recurrent and rare phenomenon in myeloid malignancieswith mostly complex cytogenetic abnormalities and arefrequently accompanied by TET2mutations on the remainingallele (20). Despite all these studies, the prognostic relevanceof TET2 lesions is still debated: Recent studies have reporteda favorable (10,15), poor (16), or inconclusive influence(11,21).
Based on these findings, we tested 41 MDS patients, withand without band 4q24 abnormalities, with 8 different

Table 1 BAC clones used to test band 4q24 deletions
Table 2 Patients’ clinical and hematological features at clinical
diagnosis
Total patients 41
Median age, years (range) 67 (29e85)
Sex
Male, n (%) 20 (48.7)
Female, n (%) 21 (51.2)
WHO classification
RARS 1
RA 4
MDS del(5q) 1
RCMDS 0
RCMD 3
RAEB-1 8
RAEB-2 24
MDS-U 0
IPSS
Low 5
Intermediate-1 6
Intermediate-2 16
High 14
Disease evolution
Advanced MDS 6
AML 15
Median follow-up, months (range) 6 (2e12)
Abbreviations: WHO, World Health Organization; MDS del(5q), 5q-
syndrome; RCMD, refractory cytopenia with multilineage displasia;
RCMDS, RCMD with ringed sideroblasts; RAEB-1, RAEB type 1;
RAEB-2, RAEB type 2; MDS-U, unclassifiable MDS; IPSS, Inter-
natonal Prognostic Scoring System.
286 I. Dambruoso et al.
bacterial artificial chromosome (BAC) probes (4). The goalsof our study were to evaluate whether FISH, a technique thatis less expensive than aCGH, SNP arrays, and NGS, is aneffective tool to reveal TET2 abnormalities and to definewhether the TET2 locus is always involved in structuraldefects of band 4q22-q24, even in patients with apparentlybalanced translocations/aneuploidies of chromosome 4.
Materials and methods
Patients
All the 41 consecutive MDS patients analyzed in the presentstudy were diagnosed at the Division of Haematology,Foundation IRCCS Policlinico San Matteo, University ofPavia (Pavia, Italy) within the period January throughDecember 2009. All these patients were classified accordingto the WHO classification (22). Our study does not includeany patient with either secondary MDS or with a white bloodcell (WBC) count above 12 � 109/L. The diagnostic proce-dures performed at clinical diagnosis were completed aspreviously reported (23).
Cytogenetic studies
Conventional cytogenetic (CC) studies were performed onbone marrow cells as previously reported (23). Chromosomeidentification and karyotype description were made accord-ing to the International System for Chromosome Nomencla-ture 2009 (24,25). In the presence of �3 chromosomalabnormalities, the karyotype was defined as complex.
FISH analysis
BAC clones (Table 1) were selected from the human libraryRPCI-11 according to the UCSC Human Genome Assembly(hg 19, February 2009). The RP11-356L5, and RP11-16G16probes were chosen because they cover the TET2 locus.Recombinant bacteria were obtained from BAC/PAC
Resources, of the Children’s Hospital and Research Centerat Oakland (Oakland, CACalifornia, USA) and cultured in LBBroth Medium with Cloramphenicol. BAC DNA extractionwas performed with the PhasePrep BAC DNA kit (Sigma-Aldrich, St. Louis, MO). Subsequently, probes were labeledwith the dig-dUTP Nick Translation Kit (Roche Diagnostics,Indianapolis, IN) and suspended in 50% formamide, 1%Tween-20, and 20% dextran sulfate along with salmon spermDNA and Cot-1 DNA (Roche Diagnostics).

Table 3 Clinical, cytogenetic, and FISH data of the 41 patients studied
Patient
number Sex Age Diagnosisa IPSS Karyotype FISH results
Disease
evolutionbFollow-up
(months)
1 M 22 RA Low 46,XY,t(1;4)(p32;q34)[20] Normal No 8/
2 F 22 RAEB-2 High 46,XX[22] Normal Yes, AML 12C
3 F 52 RAEB-2 High 46,XX[20] Normal Yes, AML 8C
4 F 48 RAEB-2 High 46,XX[20] Normal Yes, AML 3/
5 M 76 RCMD Int-2 46,XY,t(1;4)(p32;q24),t(2;14)(q25;q31),add(6)(p12)[18] Abnormal No 11/
6 M 64 RAEB-2 Int-2 46,XY,ins(5;4)(5q23;q24qter)[16] Abnormal Yes, AML 8/
7 F 48 RAEB-1 Int-2 45,XX,t(2;5)(q33;q31),del(13)(q14),del(17)(q21)[21] Normal No 6/
8 M 35 RAEB-2 Int-2 46,XY,del(4)(q31)[14] Normal Yes, AML 5/
9 M 43 RAEB-2 High 46,XY[22] Normal Yes, AML 9/
10 F 74 RAEB-2 Int-2 46,XX,del(5)(q14),add(11)(q14)[18] Normal No 6/
11 F 68 RAEB-2 High 51,XX,þ1,þ4,del(5)(q12),add(6)(q25),þ9,þ10,þ11,der(17)t(3;17)(q24;q12)[20] Trisomy 4 Yes, AML 7/
12 M 67 RAEB-2 High 46,XY[6]/ 47,XY,r(1)(p36q32),del(5)(q13),i(14)(q10),þmar [12]/
47,XY,t(1;3)(p36;p22),del(5)(q13),i(14)(q10),þmar[8]
Abnormal Yes, AML 11/
13 F 65 RAEB-1 Int-2 46,X,add(4)(q23),del(20)(q12),þmar [9]/idem,der(11)t(1;11)(q11;q25)[22] Normal Yes, AML 4/
14 F 79 RAEB-2 High 47,X,-4,del(5)(q11),add(12)(p11),-13,add(17)(p13),þ17,þmar,þmar[21] Monosomy 4 Yes, AML 6/
15 M 54 RAEB-2 Int-2 67,XY,-Y,-7,þder(7)t(7;?)(p22;?)x2,-10,þadd(12)(q24),-14,-18[12] Trisomy 4 Yes, AML 3/
16 F 66 RAEB-1 Int-1 46,XX[20] Normal Yes, RAEB-2 4/
17 F 81 RAEB-2 High 43,XX,-7,þ8,-9,-11,-11,i(11)(q10)x2,add(16)(p13),der(17)t(11;17)(q11;p12)[18] Normal Yes, AML 3/
18 M 62 RA Int-1 46,XY[16] Normal Yes, RAEB-2 12/
19 F 72 MDS del(5q) Low 46,XX,del(5)(q13q31)[18] Normal No 8/
20 M 70 RAEB-2 High 46,XY[21] Normal Yes, RAEB-2 6/
21 M 68 RAEB-2 Int-2 45,XY,-5,-7,-9,t(15;16)(p11;p11),add(16)(p13),add(17)(p12),add(19)(p13),-20,þr[12] Normal Yes, AML 5/
22 M 65 RAEB-2 Int-2 44,XY,del(5)(q13q33),-7,der(11)t(7;11)(p15;p15),-12,-13,-14,
-16,add(17)(p13),þmar,þmar,þmar[20]
Abnormal. Yes, AML 3/
23 F 37 RAEB-1 Int-1 44,XY,del(5)(q23q33),-7,-13,der(18)t(13;18)(q14;q23)[13] Normal Yes, RAEB-2 4/
24 M 65 RAEB-2 Int-2 49,XY,del(7)(q21),del(11)(p11),þr,þmar,þmar[12] Normal No 7/
25 M 68 RAEB-2 Int-2 47,XY,t(3;12)(p22;q24),del(5)(q23;q33),del(17)(q12),þmar[11] Normal No 3/
26 M 63 RAEB-2 Int-2 46,XY,t(3 ;3)(p22;q27);del(4)(q25),del(5)(q12q33),der(7)
t(7;13)(p11;q11),add(15)(p11),i(17)(q10),add(20)(q23)[18]
Abnormal No 4/
(Continued on the next page)
TET2deletio
nsin
MDS/AML
287

Table 3 (Continued )
Patient
number Sex Age Diagnosisa IPSS Karyotype FISH results
Disease
evolutionbFollow-up
(months)
27 F 60 RAEB-2 High 46,XX,del(5)(q13q33),der(7)t(7;17)(q11;q11),der(12)t(12;19)(q21;q13)[12] Abnormal Yes, AML 3/
28 M 53 RAEB-1 Int-1 46,XY[18] Normal No 2/
29 F 27 RAEB-2 High 46,XX,der(4)t(1;4)(p21;p16),del(11)(q14),þmar[13] Normal No 12/
30 M 88 RAEB-2 Low 46,XY [21] Normal No 6/
31 M 78 RAEB-1 Int-2 43,XY,del(5)(q11),der(7)t(7;14)(q11;p11),-10,add(12)(p12),-13,-14[14] Normal Yes, RAEB-2 8/
32 M 70 RA Low 46,XY[13] Normal No 9/
33 M 80 RAEB-2 High 45,XY,t(3;3)(q21;q26),-7[16] Normal No 4C
34 F 58 RARS Low 46,XX,del(20)(q12)[18] Normal Yes, RAEB-1 12/
35 F 73 RAEB-1 Int-1 47,XX,þ1,del(5)(q13q33),þ8,-16[13] Normal No 6C
36 F 82 RCMD Int-1 45,XX, del(1)(p11),del(6)(q13),-7,þ8,t(12;19)(q11;q11),-13[18] Normal No 3C
37 F 55 RCMD Int-2 44,XX,del(9)(q22),-17,-18,þmar[12] Normal No 11C
38 F 59 RA Int-2 49,XX,del(5)(q14),del(7)(q22),del(12)(p11),þmar,þmar,þmar[13] Normal No 10/
39 F 56 RAEB-2 High 49,XXadd(1)(p36),del(5)(q23q33),add(3)(q26),þmar,þmar,þmar[15] Normal No 7C
40 M 71 RAEB-1 Int-2 48,XY-5,del(7)(q21),add(11)(p11),þmar,þmar,þmar[13] Normal No 6/
41 F 65 RAEB-2 High 46,XX,t(2;3)(p23,q26),del(5)(q13q33),del(7)(q22)[12] Normal No 7/
Abbreviations: MDS del(5q), 5q-syndrome; RCMD, refractory cytopenia with multilineage displasia; RAEB-1, RAEB type 1; RAEB-2, RAEB type 2; IPSS, International Prognostic Scoring System;
Int, intermediate; / Z alive and well; C Z deceased.a According to World Health Organization (WHO) classification.b According to WHO guidelines (22).
288
I.Dambruosoetal.

TET2 deletions in MDS/AML 289
For FISH investigations, BAC probes and target DNAwere co-denaturated at 75�C for 5 minutes. Hybridizationwas performed in a humidified chamber at 37�C undera cover-slip. Post-hybridization washes were performed in0.4 � SSC with 0.3% Nonidet P40 (BDH Chemicals, Ltd.,UK) at 72�C for 2 minutes and then in 2 � SSC at roomtemperature for 2 minutes. The probes were labeled andrevealed by anti-digoxigenin-rhodamine FAB fragments(Sigma-Aldrich, St. Louis, MO). Slides were counterstainedwith DAPI (Abbott Molecular, Des Plaines, IL, USA). Meta-phases were analyzed with an Olympus BX60 epifluorencemicroscope (Olympus Corp., Milano, Italy), and images werecaptured by a Cytovision System (Olympus Corp.). After theanalysis of 300 bone marrow nuclei from 10 normal controls,in order to determine the normal reference range for each ofthe probe applied, a (one-sided) 95% confidence intervalusing a binomial distribution for the proportion of interphasecells with 1 and 2 red signals was calculated. Thus, the cut-off values were fixed at 10%.
Whole chromosome 4 and 5 painting probes (WCP4,WCP5), the centromeric probes for chromosomes 1 and 17(CEP1 and CEP17), the FIP1L1/CHIC/PDGRA probe(Kreatech Diagnostics, Amsterdam, the Netherlands), andthe Vysis LSI 5q31/5p15 (Abbott Molecular, Des Plaines, IL)were applied according to the manufacturers’ guidelines andused as internal controls.
Figure 1 Definition of the t(1;4)(p21,q24) translocation in patient 5
and G-banded chromosomes (normal chromosomes on the right, ch
with the RP11-356L5 and RP11-16G16 BAC probes (red spots) co
hybridization with the CEP4 probe (green spots) performed on the sam
the CEP1 probe (green spots) performed on another metaphase re
green arrow; (E) hybridization with the RP11-66J6 BAC probe (band
signals: one on the normal chromosome 4 and the other on der(1) (h
a red arrow.
Results
Patients
The clinical features of the 41 MDS patients are listed inTable 2. Their median age was 67 years (range 29e85).Twenty-one patients were females and 20 were males.According to the WHO classification, 4 patients were diag-nosed as refractory anemia (RA), 1 as 5q- syndrome, 3 asrefractory cytopenia with multilineage dysplasia (RCMD), 8as RA with excess of blasts type 1 (RAEB-1), 24 as RAEBtype 2 (RAEB-2), and 1 as RA with ring sideroblast (RARS).Median follow-up was 6 months (range 4e28). Within thisperiod, 7 patients (2 RCMD, 1 RAEB-1, and 4 RAEB-2) died,and 21 patients experienced disease progression (1 inRAEB-1, 5 in RAEB-2, and 15 in AML).
Conventional cytogenetics
CC analyses revealed a normal karyotype in 10 patients (2RA, 2 RAEB-1, and 6 RAEB-2), a single defect in 5 (1 RA, 15q- syndrome, 2 RAEB-2, and 1 RARS), 2 defects in 2 (2RAEB-2) and �3 defects (a complex karyotype) in 24 (3RCMD, 1 RA, 6 RAEB-1, and 14 RAEB-2) (Table 3).Considering the 31 patients with clonal defects, 7 presented
: (A) a schematic presentation of the chromosomal translocation
romosome 1 and 4 derivatives on the left); (B) on hybridization
vering the TET2 locus, one signal only is revealed (arrow); (C)
e cell reveals two normal chromosomes 4; (D) hybridization with
veals two chromosomes 1; the der(1) is indicated by a hatched
4q25) (red spot) performed on the same metaphase reveals two
atched green arrow). The chromosome 4 derivate is indicated by

Figure 2 Characterization of the ins(5;4)(q23;q24qter) in patient 6: (A) a schematic presentation of the chromosomal translocation
and G-banded chromosomes (normal chromosomes on the left, chromosome 4 and 5 derivatives on the right); (B) the hybridization
with the WCP4 probe (red signals) reveals a normal chromosome 4, a der(4) (red arrow), and a der(5) (hatched green arrow); (C)
hybridization with the WCP5 probe (red signals): the normal chromosome 5 is completely stained, and the der(5) is partially stained
(hatched green arrow).
290 I. Dambruoso et al.
with a structural defect affecting the long arm of chromosome4 as a single abnormality or as part of a complex karyotype,and 3 presented with chromosome 4 aneuploidies.
FISH results
Considering the seven patients with 4q structural abnormal-ities, FISH revealed a normal pattern in four patients: twowith a balanced translocation involving the long arm ofchromosome 4, one with a del(4)(q31), and one with anadd(4)(q23). Thus, imbalances involving bands 4q22-q24were identified in three patients (one RCMD and two RAEB-2) (Table 3). The RCMD patient (5), who carrieda t(1;4)(p36;q25), presented with the loss of the RP11-356L5, RP11-16G16, RP11-642P17, and RP11-788K3probes and the retention of the RP11-144B4, RP11-810D13, RP11-571L19, and RP11-752J12 probes in 88% ofthe interphase and mitotic cells examined (Figure 1). More-over, the RP11-66J6 probe, which maps at band 4q25, wastranslocated to chromosome 1 as revealed by the CEP1probe. Thus, these FISH results demonstrated that 4q
translocation breakpoints were localized at band 4q24 andnot at band 4q25 as suggested by CC.
AnRAEB-2 patient (patient 6)with an ins(5;4)(q23;q24qter)on CC analyses presented with some chromosomal materialderived from chromosome 4 inserted within the long arms ofchromosome 5 and no material derived from chromosome 5translocated to chromosome 4, after hybridization with theWCP4 andWCP5 probes (Figure 2). In addition, FISH studiesrevealed a cryptic deletion of the chromosomal segmentbetween bands 4q22.3 (RP11-144B4) and 4q24 (RP11-752J12) (Figure 3). Evenmore interestingwas the observationthat after hybridization with the RP11-144B4 probe, all of themitotic figures and 65%of the nuclei demonstrated two normalsignals, whereas 35% of the nuclei had one signal only. Thisdata suggests that an additional acquired rearrangementhad occurred within the clonal population displaying theins(5;4)(q23;q24qter). Furthermore, the RP11-66J6 probewas translocated onto the der(5) in a centromeric position asrevealed by the LSI 5q31/5p11 probes.
Another RAEB-2 patient (patient 26) with a del(4)(q31) onCC analysis presented with the deletion of the RP11-144B4,RP11-365L5, RP11-16G16, RP11-642P17, RP11-752J12,
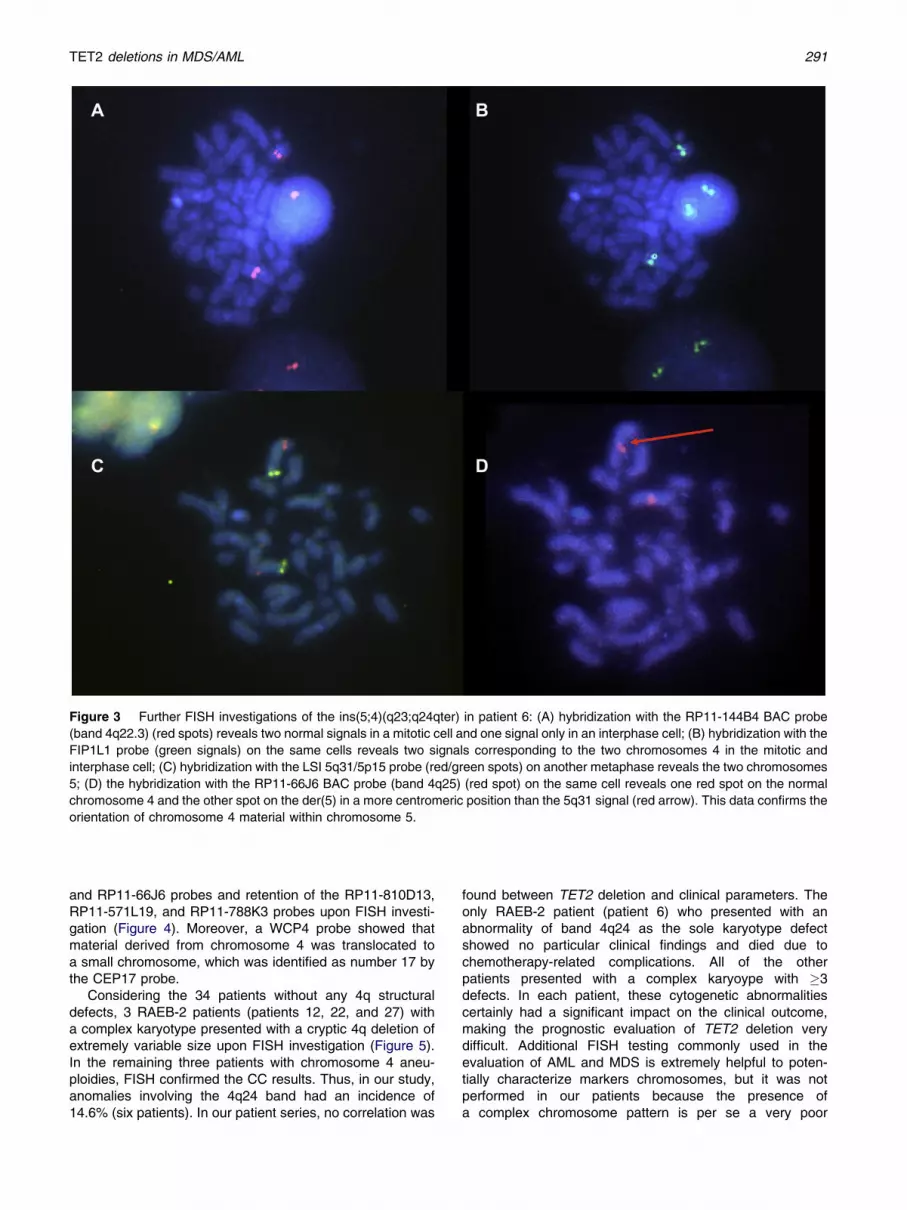
Figure 3 Further FISH investigations of the ins(5;4)(q23;q24qter) in patient 6: (A) hybridization with the RP11-144B4 BAC probe
(band 4q22.3) (red spots) reveals two normal signals in a mitotic cell and one signal only in an interphase cell; (B) hybridization with the
FIP1L1 probe (green signals) on the same cells reveals two signals corresponding to the two chromosomes 4 in the mitotic and
interphase cell; (C) hybridization with the LSI 5q31/5p15 probe (red/green spots) on another metaphase reveals the two chromosomes
5; (D) the hybridization with the RP11-66J6 BAC probe (band 4q25) (red spot) on the same cell reveals one red spot on the normal
chromosome 4 and the other spot on the der(5) in a more centromeric position than the 5q31 signal (red arrow). This data confirms the
orientation of chromosome 4 material within chromosome 5.
TET2 deletions in MDS/AML 291
and RP11-66J6 probes and retention of the RP11-810D13,RP11-571L19, and RP11-788K3 probes upon FISH investi-gation (Figure 4). Moreover, a WCP4 probe showed thatmaterial derived from chromosome 4 was translocated toa small chromosome, which was identified as number 17 bythe CEP17 probe.
Considering the 34 patients without any 4q structuraldefects, 3 RAEB-2 patients (patients 12, 22, and 27) witha complex karyotype presented with a cryptic 4q deletion ofextremely variable size upon FISH investigation (Figure 5).In the remaining three patients with chromosome 4 aneu-ploidies, FISH confirmed the CC results. Thus, in our study,anomalies involving the 4q24 band had an incidence of14.6% (six patients). In our patient series, no correlation was
found between TET2 deletion and clinical parameters. Theonly RAEB-2 patient (patient 6) who presented with anabnormality of band 4q24 as the sole karyotype defectshowed no particular clinical findings and died due tochemotherapy-related complications. All of the otherpatients presented with a complex karyoype with �3defects. In each patient, these cytogenetic abnormalitiescertainly had a significant impact on the clinical outcome,making the prognostic evaluation of TET2 deletion verydifficult. Additional FISH testing commonly used in theevaluation of AML and MDS is extremely helpful to poten-tially characterize markers chromosomes, but it was notperformed in our patients because the presence ofa complex chromosome pattern is per se a very poor

Figure 4 Characterization of the t(4;17)(q31;p13) chromosomal translocation in patient 26: (A) schematic presentation of the
chromosomal translocation and G-banded chromosomes (normal chromosomes on the left, chromosome 4 and 17 derivatives on the
right); (B) the hybridization with the WCP4 probe (red signals) reveals a normal chromosome 4, a completely stained der(4) (hatched
green arrow), and another partially stained chromosome; (C) the hybridization with the CEP17 probe (green signals) on the same
metaphase identifies the chromosome partially stained by the WCP4 as a number 17; (CeF) hybridization with the RP11-144B4,
RP11-752J12, and RP11-J6 BAC probes (red signals) on different mitotic figures reveals one signal only with each of the probes
applied. In each metaphase, the der(4) (red arrow) was identified with a CEP4 probe (data not shown).
Figure 5 The 4q deletion patterns of the six patients, together with the probes involved, are depicted in the figure.
292 I. Dambruoso et al.

TET2 deletions in MDS/AML 293
prognostic factor often resulting in patients needing alloge-neic bone marrow transplantation.
Discussion
The goal of the present study was to evaluate whether FISHis an effective method to precisely define band 4q22-q24structural defects and to reveal TET2 deletions in a series of41 MDS patients with and without 4q abnormalities upon CCanalysis. Currently, the most effective techniques used toidentify TET2 haplo-insufficiency are aCGH, SNP arrays,and molecular techniques (5e11). Usually, FISH is used tovalidate aCGH results, even if various studies have sug-gested that TET2 mutations and aberrations affecting the4q24 locus are independent genetic events (9e26). In 2005,a study applied FISH to four MDS/AML patients: three withan apparently balanced 4q24 translocation as the solekaryotype abnormality and the remaining patient witha del(4)(q23q24),þ4 (4). A set of BAC probes spanning the4q22-4q27 region was applied in order to precisely definethe 4q breakpoint. In the three patients with chromosome 4translocations, a common deletion region included the twocontiguous DNA probes, RP11-356L5 and RP11-16G16. Inthe remaining patient, this same deletion was carried by oneof the three homologs. Based on the size of the BAC probes,it was established that the extent of this common deletedsegment was approximately 0.5 Mb. In a more recentretrospective FISH study, 79 CMML patients and fourpatients diagnosed as AML post-CMMLwere analyzed (27);no patient carried the TET2 deletion. Other SNP array andNGS studies have revealed that TET2 deletions occur in2e14% of MDS/AML patients (3e11,19), and a more recentstudy has reported that loss of heterozygosity at the TET2locus occurs in 20% of MDS patients (26). In the presentseries, the loss of the RP11-356L5 and RP11-16G16probes, which hybridize to the centromeric and telomericregion of the TET2 locus, had an incidence of 14.6%. Froma cytogenetic point of view, two patients carried a 4qdefect as the sole karyotype abnormality, whereas theothers had a complex karyotype. In particular, one patienteach carried an apparently balanced chromosome 4qdefect, a t(1;4)(p21;q24), and an ins(5;4)(q23;q24qter); onepatient harbored a del(4)(q31); and three patients carriedtwo apparently normal chromosomes 4. Thus, upon FISHstudies, four patients presented with an absolutely unex-pected loss of the RP11-356L5 and RP11-16G16 probes. Inall patients, the size of the deletion was extremely variable(Figure 5), a finding suggesting that 4q22-q24 deletions arenot mediated by repetitive sequence (similar to segmentalduplications, Alu repeats, etc.). This conclusion is furtherstrengthened by the results achieved by FISH in patients 6and 26. Upon hybridization with the RP11-144B4 (4q22.3)probe, all of the metaphases and 65% of the nuclei frompatient 6 showed two normal signals, but 35% of his inter-phase cells showed one signal only. This result suggestsa clonal evolution within the population carrying theins(5;4)(q23;q24qter). The other patient (patient 26)demonstrated loss of the RP11-144B4, RP11-356L5, RP11-16G16, RP11-642P17, RP11-752J12, and RP11-66J6probes and the retention of the RP11-810D13, RP11-571L19, and RP11-788K3 probes (Figure 5). No patient
presented with a partial deletion (signal intensity approxi-mately reduced by one half) with any probe, a findingreported in a previous study (4).
The decision to perform FISH in our 34 patients withoutany structural defect at bands 4q22-q24, including 3 patientswith chromosome 4 aneuploidies, was dictated by an attemptto determine whether these sub-microscopic deletions mightbe a recurrent abnormality in MDS. As already reported, anabnormal FISH pattern was observed in three patients(8.8%), all with two apparently normal chromosomes 4. Inparticular, in one patient, the deletion was confined to theRP11-356L5 and RP11-16G16 probes, and in two patients, itinvolved additional probes (Figure 5). Moreover, in the threepatients with chromosome 4 aneuploidies, FISH investiga-tions were conducted based on literature data, which haveshown that TET2 behaves as a tumor suppressor gene (27).The RP11-356L5 and RP11-16G16 probes provided onesignal only in the �4 patient and three signals in the two þ4patients.
Importantly, a recent next-generation sequencing studyshowed that TET2 mutant clones were not only found inCD34þ and total bone marrow cells, but also in T cells(11). In our series, this evaluation could not be performed.In fact, the only patient who harbored the TET2 deletion asthe sole karyotype defect, a patient with RAEB-2, wasexamined at clinical diagnosis; on that occasion, CD34-positive and T cells were not submitted for immunologicselection.
In addition, in this patient, the prognostic significance ofthe TET2 deletion could not be assessed because he died ofchemotherapy-related complications. The other five patientswho presented with a TET2 deletion all had multiple alter-ations that hampered any potential correlation between TET2deletion and clinical outcome.
In conclusion, 1) FISH is an effective and economicmethod for revealing cryptic band 4q22-q24 abnormalitiesand TET2 deletions, which are recurrent defects in MDS; 2)in these patients, TET2 deletion is the unifying genetic event,and 3) deletions of band 4q22-q25 are probably not mediatedby repetitive sequences and do not occur through themechanism of non-allelic homologous recombination.
References
1. Levine RL, Carroll M. A common genetic mechanism in malignant
bone marrow diseases. N Engl J Med 2009;360:2355e2357.
2. Mohamedali AM, Smith AE, Gaken J, et al. Novel TET2 muta-
tions associated with UPD4q24 in myelodysplastic syndrome.
J Clin Oncol 2009;27:4002e4006.
3. Delhommeau F, Dupont S, Della Valle V, et al. Mutation in TET2
in myeloid cancer. N Engl J Med 2009;360:2289e2301.4. Vigui�e F, Aboura A, Bouscary D, et al. Common 4q24 deletion in
four cases of hematopoietic malignancy: early stem cell
involvement? Leukemia 2005;19:1411e1415.5. Mohamedali A, G€aken J, Twine NA, et al. Prevalence and
prognostic significance of allelic imbalance by single-nucleotide
polymorphism analysis in low-risk myelodysplastic syndromes.
Blood 2007;110:3365e3373.6. Gondek LP, Haddad AS, O’Keefe CL, et al. Detection of cryptic
chromosomal lesions including acquired segmental uniparental
disomy in advanced and low-risk myelodysplastic syndromes.
Exp Hematol 2007;35:1728e1738.

294 I. Dambruoso et al.
7. Gondek LP, Tiu R, O’Keefe CL, et al. Chromosomal lesions and
uniparental disomy detected by SNP arrays in MDS, MDS/MPD,
and MDS-derived AML. Blood 2008;111:1534e1542.
8. Starczynowski DT, Vercauteren S, Telenius A, et al. High-reso-
lution whole genome tiling path array CGH analysis of CD34þcells from patients with low-risk myelodysplastic syndromes
reveals cryptic copy number alterations and predicts overall and
leukemia-free survival. Blood 2008;112:3412e3424.9. Jankowska AM, Szpurka H, Tiu RV, et al. Loss of heterozygosity
4q24 and TET2 mutations associated with myelodysplas-
tic/myeloproliferative neoplasms. Blood 2009;113:6403e6410.
10. Kohlmann A, Grossmann V, Klein HU, et al. Next-generation
sequencing technology reveals a characteristic pattern of
molecular mutations in 72.8% of chronic myelomonocytic
leukemia by detecting frequent alterations in TET2, CBL, RAS
and RUNX1. J Clin Oncol 2010;28:3858e3865.11. Smith AE, Mohamedali AM, Kulasekararaj A, et al. Next-
generation sequencing of the TET2 gene in 355 MDS and
CMML patients reveals low-abundance mutant clones with early
origins, but indicates no define prognostic value. Blood 2010;
116:3923e3932.
12. Tefferi A, Lim KH, Levine R. Mutation in TET2 in myeloid
cancers. N Engl J Med 2009;361:1117.
13. Tefferi A, Pardanani A, Lim KH, et al. TET2 mutations and their
clinical correlates in polycythemia vera, essential thrombocy-
themia and myelofibrosis. Leukemia 2009;23:905e911.
14. Tefferi A, Levine RL, Lim KH, et al. Frequent TET2 mutations in
systemic mastocytosis: clinical, KITD816V and FIP1L1-
PDGFRA correlates. Leukemia 2009;23:900e904.
15. Kosmider O, Gelsi-Boyer V, Cheok M, et al. TET2 mutation is an
independent favorable prognostic factor in myelodysplastic
syndromes (MDSs). Blood 2009;114:3285e3291.
16. Kosmider O, Gelsi-Boyer V, Ciudad M, et al. TET2 mutation is
a frequent and adverse event in chronic myelomocytic
leukaemia. Haematologica 2009;94:1676e1681.
17. Langemeijer SMC, Kuiper RP, Berends M, et al. Acquired
mutations in TET2 are common in myelodysplastic syndromes.
Nat Genet 2009;41:838e842.
18. Hussein K, Abdel-Wahab O, Lasho TL, et al. Cytogenetic
correlates of TET2 mutations in 199 patients with myeloprolif-
erative neoplasms. Am J Hematol 2010;85:81e83.
19. Bacher U, Weissmann S, Kohlmann A, et al. TET2 deletions are
recurrent but rare phenomenon in myeloid malignancies and are
frequently accompanied by TET2 mutations on the remaining
allele. Br J Haematol 2002;156:67e75.
20. Couronn�e L, Lippert E, Andrieux J, et al. Analyses of TET2
mutations in post-myeloproliferative neoplasm acute myeloid
leukemias. Leukemia 2010;24:201e203.
21. Nibourel O, KosmiderO, CheokM, et al. Incidence and prognostic
value of TET2 alterations in de novo acute myeloid leukemia
achieving complete remission. Blood 2010;116:1132e1135
22. Swerdlow SH, Campo E, Harris NL, et al. WHO classification of
tumours, Volume 2. Geneva: IARC; 2008.
23. Bernasconi P, Klersy C, Boni M, et al. Incidence and prognostic
significance of karyotype abnormalities in de novo primary
myelodysplastic syndromes: a study on 331 patients from
a single institution. Leukemia 2005;19:1424e1431.
24. Shaffer LG, Slovak ML, Campbell LJ, editors. ISCN 2009: an
international system for human cytogenetic nomenclature.
Basel: Karger; 2009.
25. Mitelman F, Johansson B, Mertens F. Mitelman database of
chromosome aberrations in cancer. Available at:http://cgap.nci.
nih.gov/Chromosomes/Mitelman. accessed February 21, 2012.
26. Tefferi A, Lim KH, Abdel-Wahab O, et al. Detection of mutant
TET2 in myeloid malignancies other than myeloproliferative
neoplasms: CMML, MDS, MDS/MPN and AML. Leukemia 2009;
23:1343e1345.
27. Mallo M, Osca G, Sol�orzano J, et al. TET2 gene is not deleted in
chronic myelomonocytic leukemia: a FISH retrospective study.
Haematologica 2010;95:1798e1800.





























