Embed Size (px)
Citation preview

DEVELOPMENT OF CARBOHYDRATE ANALYSIS-BASED METHODS
FOR SUSTAINABLE WALNUT ORCHARD MANAGEMENT
Maciej Zwieniecki, Anna Davidson, Adele Amico Roxas
ABSTRACT
Tree growth and yield are dependent on a complex set of interactions involving genotype,
physiological and developmental processes, and the interaction of these processes with the
environment (abiotic and biotic stress). Non-structural carbohydrates (NSC; starch (ST) and
soluble carbohydrates (SC) like sucrose, glucose or fructose) are at the center of tree intrinsic and
extrinsic activities. As such, they are the key energy currency. NSC are produced during
photosynthesis and used to ‘pay’ for all biological services (growth, respiration, nutrient uptake,
defense against pathogens, reproduction, protection from abiotic stress). Evolved management of
NSC reserves is the key allowing the tree to survive adverse climatic conditions during vegetative
growth portion of the year (spring-summer-fall) and dormancy (winter). As the Central Valley
climate becomes more erratic and abiotic stresses more severe (summer like temperatures in the
fall and prolonged fall drought, loss of winter fog, large daily swings of temperature, and
increasing probability of winter frost), understanding the mechanisms responsible for tree energy
(NSC) management is crucial, yet still missing, is a piece of the puzzle to supplement existing
techniques (like nutrient analysis, water potential measurements) in mediating the impact of
current and future abiotic stresses and maintain high productivity.
Using a ‘citizen research’ approach, the Carbohydrate Observatory (CO), we are gaining a unique
insight to seasonal dynamics of NSC in walnut trees: (http://zlab-carb-
observatory.herokuapp.com). A large-scale approach and the ability to collect samples from a
diverse range of sites, tree ages, varieties and management practices provides opportunity to
determine the main factors linking productivity with NSC management. Currently, we have
determined that unlike almond and pistachio, walnut has a conservative strategy of NSC
management resulting in small swings of reserves between the summer productive phase and
dormancy. However, even these small variations seem important for yield capacity as indicated by
a significant positive correlation between NSC reserves in the winter and yield the following year.
Interestingly, we did not observe a significant negative correlation of NSC content in summer with
yield, as was observed in almond and pistachio. This suggests that trees with high NSC reserves
are always most productive.
In addition, analysis of the NSC seasonal pattern provides an opportunity to develop predictive
models of dormancy. The experimental approach to manipulate NSC availability in the fall by
defoliation and their redistribution by girdling revealed a complicated pattern that suggests a
significant impact of winter NSC storage on bloom success. In general, any disruption to natural
senescence and redistribution resulted in delayed bloom and reduced synchrony of the bloom.
However, the most pronounced effect was seen after early fall defoliation and girdling in the spring
– a situation that reduces NSC storage and does not allow for late spring retrieval of distally located
sugars. This insight provides prospect for the development of a model that uses climatic data and
trees’ NSC management to asses progression of dormancy and tree readiness for synchronous

bloom similar to an existing model for almond: http://zlab-chill-heat-model.herokuapp.com.
OBJECTIVES
Our main objective is to develop a carbohydrate analysis method as a tool to determine walnut’s
physiological status that would complement the currently used methods such as water potential
and nutrient analysis. The goal is to use carbohydrate analysis as a new option for sustainable
orchard management. The main objective represents a long-term goal with sub-yearly objectives
representing steps. Sub-objectives for the reported year were:
(1) Continue to maintain and possibly expand a network of walnut orchards to provide samples
to a large-scale, state-wide study of seasonal dynamics of carbohydrate using a ‘citizen
research approach’ that minimizes time and costs for research.
(2) Provide easy and informative access to NSC information for walnut growers by further
improvements to our online platform: http://zlab-carb-observatory.herokuapp.com
(3) Describe and publish seasonal patterns of NSC dynamics within the walnut crown that
links NSC management with tree phenology.
(4) Use data already collected to determine impact of NSC on yield.
(5) Experimentally test the role of NSC reserve levels and their redistribution on bud
development and bloom time.
SIGNIFICANT FINDINGS
(1) We retain a number of orchards that provide samples for analysis across a wide range of
climatic conditions although continuation of sampling from specific orchards was affected
by resent economic perturbances. We further improve and streamline our protocol for
sample analysis to determination content of soluble sugars and starch in walnut wood and
bark. Again, our methodology allows for cost effective large-scale analyses of
carbohydrate that provide means for a citizen research approach. In-house cost of analysis
is under $8 per sample compared to over $100 in commercial analytical labs.
(2) Online platform for sharing information on near real time of NSC content in walnut trees
(http://zlab-carb-observatory.herokuapp.com) has undergone significant improvement.
This included new capacity to compare orchard performance with state average and other
specific farms. In addition, it is now possible to compare NSC content in different
rootstocks, scions, age groups and geographical locations at the level of counties.
(3) A manuscript describing the impact of NSC management on tree phenology is under its
second round of revisions in Scientific Reports: Comparison of phenological traits, growth
patterns, and seasonal dynamics of non-structural carbohydrate in Mediterranean tree
crop species. 2019. Aude Tixier, Paula Guzmán-Delgado, Or Sperling, Adele Amico
Roxas, Emilio Laca, Maciej Zwieniecki. Scientific Reports (in revision).
(4) An analysis of NSC content in twigs was used to determine their impact on yield. Findings
suggest a significant positive correlation between NSC content in January and March on
following year yield. Surprisingly and unlike other nut species, no post-harvest NSC
exhaustion was found suggesting a highly conservative reproductive pattern.
(5) Detailed determination of temporal dynamics of NSCs content in walnut trees suggests that
early spring and late fall are the most significant periods affecting NSCs content and

redistribution. During late fall, walnut trees restore levels of NSCs required to survive
winter. During the spring, carbohydrates are mobilized for bud break. Experimentally
induced perturbances to these patterns significantly delayed or even halted flower and bud
development. Both early defoliation and girdling of phloem reduced bloom capacity and
delayed bloom.
PROCEDURES
The Carbohydrate Observatory is the research initiative providing analytical service to growers
interested in a better understanding of NSC management of their orchards. Growers provide three
twig samples (with xylem and phloem separated) per orchard that are subsequently analyzed for
soluble sugars and starch content in wood xylem – water conducting tissue) and bark (phloem –
sugar conducting tissue). Analytical results are being published on line at http://zlab-carb-
observatory.herokuapp.com.
Received samples are processed in the lab following the procedure described below:
• Each sample is ground into powder. A small amount (25 mg) is then washed in 1 mL of
pH buffer to dissolve all soluble carbohydrates.
• Using a colorimetric method (a spectrophotometer), the concentration of sugars is
measured (Anthrone method) in sample of buffer (50 uL) and recalculated to express SC
concertation per g of dry matter.
• The remaining material in the buffer is treated with two different enzymes that digest starch
to form the soluble sugars. These are again measured in a spectrophotometer using the
Anthrone method.
• For further details of the procedure please refer to published articles by the Zwieniecki lab,
on our website or request the procedure vial email ([email protected]).
Additional information related to each orchard is provided by growers on voluntary bases that
includes specific management practices, age, scion/rootstock combination and yield. This part
needs improvement due to a low participation from walnut growers. The quality of analysis
requires a high number of samples. Our lab is currently communicating with many growers to
explain the reasoning for the need of additional data. In addition, CIMIS and NOAA weather data
is used in the analysis. The additional site related information is crossed referenced with the NSC
analysis database and used in subsequent analyses of the role of NSCs in annual orchard
performance.
In winter of 2018/2019, a field experiment on the impact of NSC content and redistribution was
developed to study synchrony and timing of bloom in walnut. The study was performed during
2018-2019 in the experimental orchard owned by University of California, Davis (38°22’ N,
121°47’ W). For this study, we selected 20 homogenous mature walnut (Juglans regia L.) trees.
The experiment started in October and ended in May, to enable us to study the pre-dormancy,
dormancy and bud-break stages. Also, climatic data were collected from a data logger located in
the experimental site.
On the twenty selected trees, we applied experimental treatments. We defoliated the trees in
October and November to large limbs and girdled smaller branches three times (October,
December and April). Defoliation x girdling interaction treatments were also applied. Control
Branches were left intact. Girdling was done by removing ~1 cm of bark at the base of twigs using
a dull razorblade. The exposed xylem section was covered with Parafilm in order to avoid
desiccation. Every month we checked that the Parafilm was still in good condition. The sample
collection for carbohydrates measurement (NSC) was performed monthly starting October and
concluded in May. During every sample collection, twigs from the experimental orchard were
quickly transported to the laboratory. Bark was removed from ~3 cm of the twig and fresh weight
of the bark and wood was weighed. Samples were then dried at 75°C for 48 hours. Once dry,
samples were weighed; then ground into a fine and homogeneous powder using a ball mill for
NSC quantification. Soluble carbohydrates (SC) were extracted by incubating 25 mg of dry
material in 1 mL of sodium acetate buffer (0.2 M and 5.5 pH) for 15 minutes at 70 °C followed by
centrifugation (10 minutes at 21,000 g). The supernatant was diluted in deionized water (1:20,
v/v); the remaining pellet with the buffer was kept for starch analysis. The Pellet was incubated
for 10 min at 100°C to allow starch gelatinization and then digested with 0.7 U of amylase and 7
U of amyloglucosidase for 4 hours at 37 °C. Once the digestion was finished, samples were
centrifuged for 10 minutes at 21,000 g. The supernatant was diluted in deionized water (1:20, v/v).
SC and starch were quantified at the end of the extraction using Anthrone as a reagent (0.1 % (m/v)
in 98 % sulfuric acid) by reading absorbance at 620 nm (Leyva et al., 2008). In order to measure
the carbohydrate content in leaves, weekly we randomly selected five trees in October and
November. Leaf collection ended in the middle of November.
Phenological observation and bud measurement were performed on five randomly selected trees.
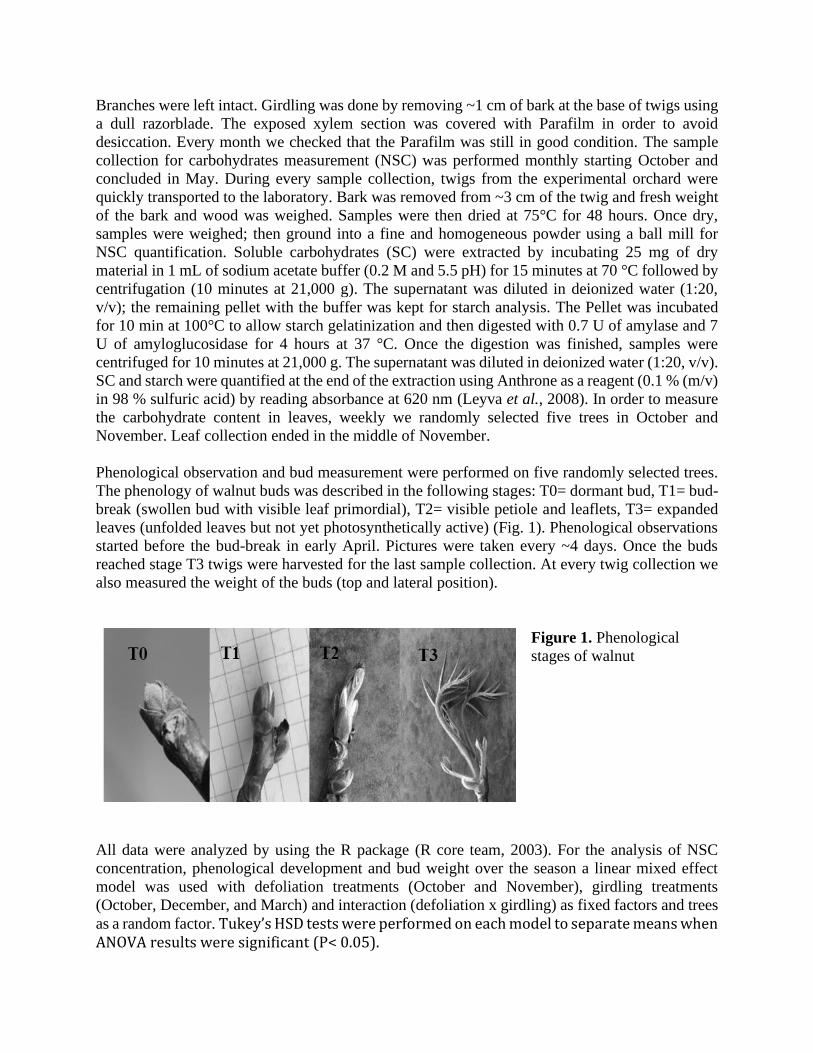
The phenology of walnut buds was described in the following stages: T0= dormant bud, T1= bud-
break (swollen bud with visible leaf primordial), T2= visible petiole and leaflets, T3= expanded
leaves (unfolded leaves but not yet photosynthetically active) (Fig. 1). Phenological observations
started before the bud-break in early April. Pictures were taken every ~4 days. Once the buds
reached stage T3 twigs were harvested for the last sample collection. At every twig collection we
also measured the weight of the buds (top and lateral position).
Figure 1. Phenological
stages of walnut
All data were analyzed by using the R package (R core team, 2003). For the analysis of NSC
concentration, phenological development and bud weight over the season a linear mixed effect
model was used with defoliation treatments (October and November), girdling treatments
(October, December, and March) and interaction (defoliation x girdling) as fixed factors and trees
as a random factor. Tukey’s HSD tests were performed on each model to separate means when ANOVA results were significant (P< 0.05).
RESULTS
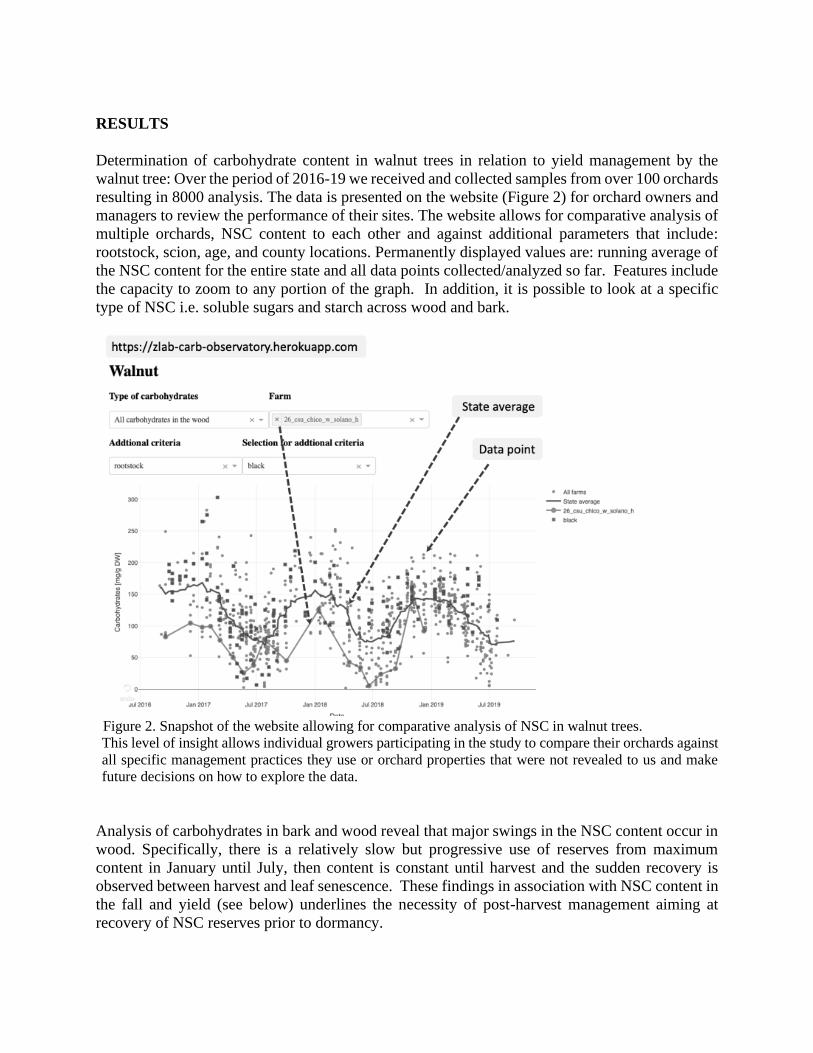
Determination of carbohydrate content in walnut trees in relation to yield management by the
walnut tree: Over the period of 2016-19 we received and collected samples from over 100 orchards
resulting in 8000 analysis. The data is presented on the website (Figure 2) for orchard owners and
managers to review the performance of their sites. The website allows for comparative analysis of
multiple orchards, NSC content to each other and against additional parameters that include:
rootstock, scion, age, and county locations. Permanently displayed values are: running average of
the NSC content for the entire state and all data points collected/analyzed so far. Features include
the capacity to zoom to any portion of the graph. In addition, it is possible to look at a specific
type of NSC i.e. soluble sugars and starch across wood and bark.
Figure 2. Snapshot of the website allowing for comparative analysis of NSC in walnut trees.
This level of insight allows individual growers participating in the study to compare their orchards against
all specific management practices they use or orchard properties that were not revealed to us and make
future decisions on how to explore the data.
Analysis of carbohydrates in bark and wood reveal that major swings in the NSC content occur in
wood. Specifically, there is a relatively slow but progressive use of reserves from maximum
content in January until July, then content is constant until harvest and the sudden recovery is
observed between harvest and leaf senescence. These findings in association with NSC content in
the fall and yield (see below) underlines the necessity of post-harvest management aiming at
recovery of NSC reserves prior to dormancy.
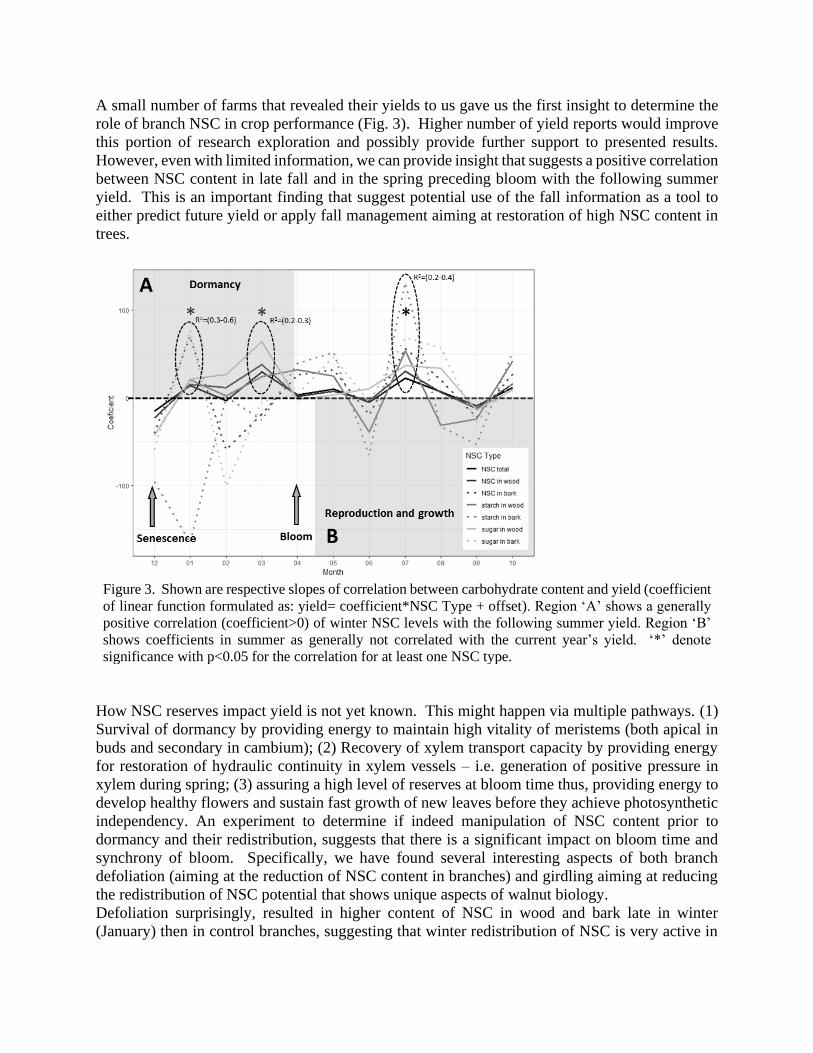
A small number of farms that revealed their yields to us gave us the first insight to determine the
role of branch NSC in crop performance (Fig. 3). Higher number of yield reports would improve
this portion of research exploration and possibly provide further support to presented results.
However, even with limited information, we can provide insight that suggests a positive correlation
between NSC content in late fall and in the spring preceding bloom with the following summer
yield. This is an important finding that suggest potential use of the fall information as a tool to
either predict future yield or apply fall management aiming at restoration of high NSC content in
trees.
Figure 3. Shown are respective slopes of correlation between carbohydrate content and yield (coefficient
of linear function formulated as: yield= coefficient*NSC Type + offset). Region ‘A’ shows a generally
positive correlation (coefficient>0) of winter NSC levels with the following summer yield. Region ‘B’
shows coefficients in summer as generally not correlated with the current year’s yield. ‘*’ denote
significance with p<0.05 for the correlation for at least one NSC type.
How NSC reserves impact yield is not yet known. This might happen via multiple pathways. (1)
Survival of dormancy by providing energy to maintain high vitality of meristems (both apical in
buds and secondary in cambium); (2) Recovery of xylem transport capacity by providing energy
for restoration of hydraulic continuity in xylem vessels – i.e. generation of positive pressure in
xylem during spring; (3) assuring a high level of reserves at bloom time thus, providing energy to
develop healthy flowers and sustain fast growth of new leaves before they achieve photosynthetic
independency. An experiment to determine if indeed manipulation of NSC content prior to
dormancy and their redistribution, suggests that there is a significant impact on bloom time and
synchrony of bloom. Specifically, we have found several interesting aspects of both branch
defoliation (aiming at the reduction of NSC content in branches) and girdling aiming at reducing
the redistribution of NSC potential that shows unique aspects of walnut biology.
Defoliation surprisingly, resulted in higher content of NSC in wood and bark late in winter
(January) then in control branches, suggesting that winter redistribution of NSC is very active in
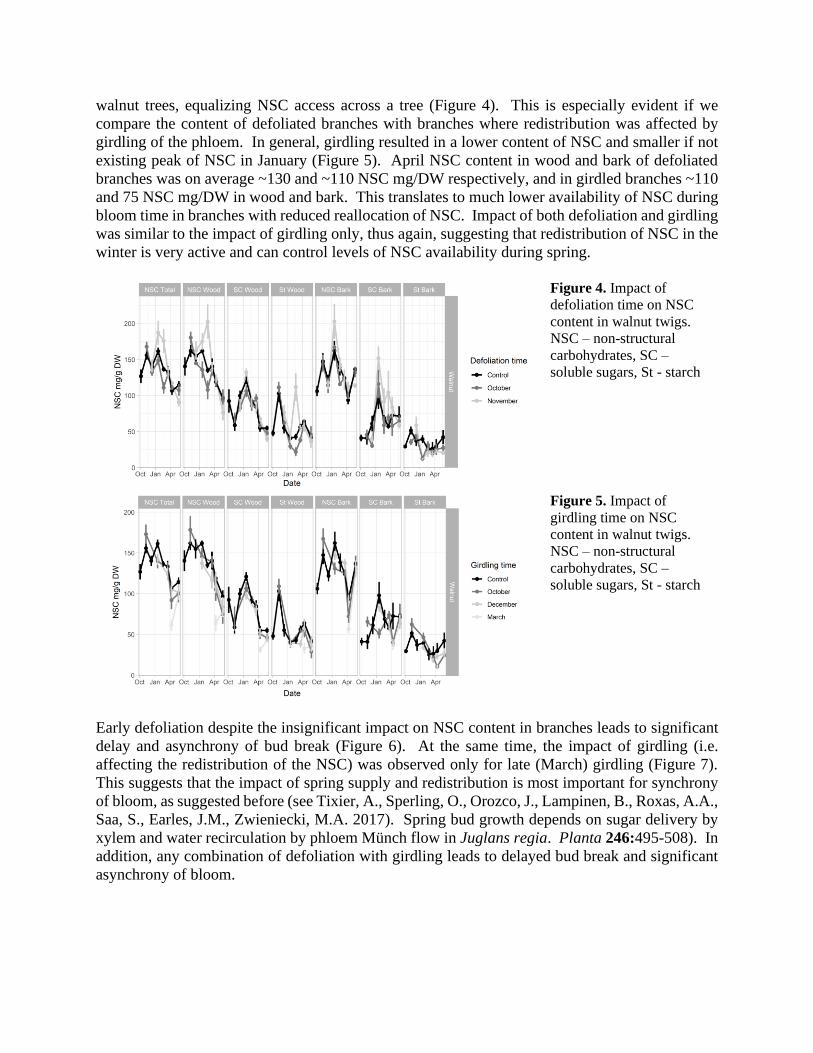
walnut trees, equalizing NSC access across a tree (Figure 4). This is especially evident if we
compare the content of defoliated branches with branches where redistribution was affected by
girdling of the phloem. In general, girdling resulted in a lower content of NSC and smaller if not
existing peak of NSC in January (Figure 5). April NSC content in wood and bark of defoliated
branches was on average ~130 and ~110 NSC mg/DW respectively, and in girdled branches ~110
and 75 NSC mg/DW in wood and bark. This translates to much lower availability of NSC during
bloom time in branches with reduced reallocation of NSC. Impact of both defoliation and girdling
was similar to the impact of girdling only, thus again, suggesting that redistribution of NSC in the
winter is very active and can control levels of NSC availability during spring.
Figure 4. Impact of
defoliation time on NSC
content in walnut twigs.
NSC – non-structural
carbohydrates, SC –
soluble sugars, St - starch
Figure 5. Impact of
girdling time on NSC
content in walnut twigs.
NSC – non-structural
carbohydrates, SC –
soluble sugars, St - starch
Early defoliation despite the insignificant impact on NSC content in branches leads to significant
delay and asynchrony of bud break (Figure 6). At the same time, the impact of girdling (i.e.
affecting the redistribution of the NSC) was observed only for late (March) girdling (Figure 7).
This suggests that the impact of spring supply and redistribution is most important for synchrony
of bloom, as suggested before (see Tixier, A., Sperling, O., Orozco, J., Lampinen, B., Roxas, A.A.,
Saa, S., Earles, J.M., Zwieniecki, M.A. 2017). Spring bud growth depends on sugar delivery by
xylem and water recirculation by phloem Münch flow in Juglans regia. Planta 246:495-508). In
addition, any combination of defoliation with girdling leads to delayed bud break and significant
asynchrony of bloom.
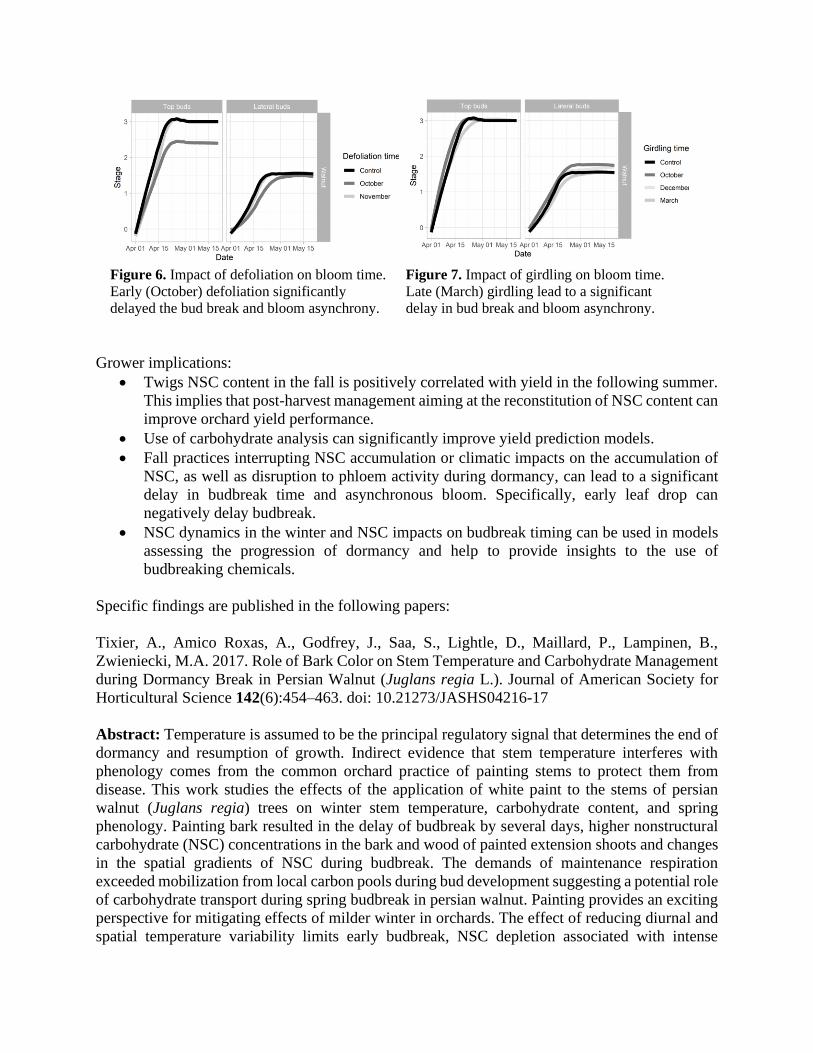
Figure 6. Impact of defoliation on bloom time.
Early (October) defoliation significantly
delayed the bud break and bloom asynchrony.
Figure 7. Impact of girdling on bloom time.
Late (March) girdling lead to a significant
delay in bud break and bloom asynchrony.
Grower implications:
• Twigs NSC content in the fall is positively correlated with yield in the following summer.
This implies that post-harvest management aiming at the reconstitution of NSC content can
improve orchard yield performance.
• Use of carbohydrate analysis can significantly improve yield prediction models.
• Fall practices interrupting NSC accumulation or climatic impacts on the accumulation of
NSC, as well as disruption to phloem activity during dormancy, can lead to a significant
delay in budbreak time and asynchronous bloom. Specifically, early leaf drop can
negatively delay budbreak.
• NSC dynamics in the winter and NSC impacts on budbreak timing can be used in models
assessing the progression of dormancy and help to provide insights to the use of
budbreaking chemicals.
Specific findings are published in the following papers:
Tixier, A., Amico Roxas, A., Godfrey, J., Saa, S., Lightle, D., Maillard, P., Lampinen, B.,
Zwieniecki, M.A. 2017. Role of Bark Color on Stem Temperature and Carbohydrate Management
during Dormancy Break in Persian Walnut (Juglans regia L.). Journal of American Society for
Horticultural Science 142(6):454–463. doi: 10.21273/JASHS04216-17
Abstract: Temperature is assumed to be the principal regulatory signal that determines the end of
dormancy and resumption of growth. Indirect evidence that stem temperature interferes with
phenology comes from the common orchard practice of painting stems to protect them from
disease. This work studies the effects of the application of white paint to the stems of persian
walnut (Juglans regia) trees on winter stem temperature, carbohydrate content, and spring
phenology. Painting bark resulted in the delay of budbreak by several days, higher nonstructural
carbohydrate (NSC) concentrations in the bark and wood of painted extension shoots and changes
in the spatial gradients of NSC during budbreak. The demands of maintenance respiration
exceeded mobilization from local carbon pools during bud development suggesting a potential role
of carbohydrate transport during spring budbreak in persian walnut. Painting provides an exciting
perspective for mitigating effects of milder winter in orchards. The effect of reducing diurnal and
spatial temperature variability limits early budbreak, NSC depletion associated with intense

maintenance respiration, freeze–thaw cycles and frost dehardening.
Tixier, A., Sperling, O., Orozco, J., Lampinen, B., Roxas, A.A., Saa, S., Earles, J.M., Zwieniecki,
M.A. 2017. Spring bud growth depends on sugar delivery by xylem and water recirculation by
phloem Munch flow in Juglans regia. Planta 246:495-508
Abstract: Main conclusion: During spring, bud growth relies on long-distance transport of
remotely stored carbohydrates. A new hypothesis suggests this transport is achieved by the
interplay of xylem and phloem. During the spring, carbohydrate demand of developing buds often
exceeds locally available storage, thus requiring the translocation of sugars from distant locations
like limbs, stems and roots. Both the phloem and xylem have the capacity for such long-distance
transport, but their functional contribution is unclear. To address this ambiguity, the spatial and
temporal dynamics of carbohydrate availability in extension shoots of Juglans regia L. were
analyzed. A significant loss of extension shoot carbohydrates in remote locations was observed
while carbohydrate availability near the buds remained unaffected. This pattern of depletion of
carbohydrate reserves supports the notion of long-distance translocation. Girdling and dye
perfusion experiments were performed to assess the role of phloem and xylem in the transport of
carbohydrate and water towards the buds. Girdling caused a decrease in nonstructural carbohydrate
concentration above the point of girdling and an unexpected concurrent increase in water content
associated with impeded xylem transport. Based on experimental observations and modeling, we
propose a novel mechanism for maintenance of spring carbohydrate translocation in trees where
xylem transports carbohydrates and this transport is maintained with the recirculation of water by
phloem Munch flow. Phloem Munch flow acts as a pump for generating water flux in xylem and
allows for transport and mobilization of sugars from distal locations prior to leaves photosynthetic
independence and in the absence of transpiration.
Predicting bloom dates by temperature mediated kinetics of carbohydrate metabolism in deciduous
trees. 2019. Or, S., Kamai, T., Tixier, A., Davidson, A., Jarvis-Shean, K., Raveh, E., DeJong, T.,
Zwieniecki, M.A. Agricultural and Forest Meteorology (in press):
https://doi.org/10.1016/j.agrformet.2019.107643
Abstract: Trees in seasonal climates gauge winter progression to assure vital and productive
blooming. However, how dormant plants asses environmental conditions remains obscure. We
postulated that it involves the energetic reserves required for bloom, and therefore studied winter
carbohydrate metabolism in deciduous trees. We quantified non-structural carbohydrates
throughout winter in almond, peach, and pistachio trees in California and Israel and characterized
winter metabolism. We constructed a carbohydrate-temperature (C–T) model that projects changes
in starch and soluble carbohydrate concentrations by temperature mediated kinetics. Then, we
tested the C–T model projections of bloom times by 20 years of temperature and phenology records
from California. The C–T model attributes winter carbohydrate regulation in dormant trees to
continuous updates of metabolic pathways. The model projects a surge in starch synthesis at the
end of winter, and critically low concentrations of soluble carbohydrates, that trigger bloom. This
is supported by field measurements of starch accumulation at the end of winter (˜50 mg g−1 DW
in almonds) that preceded bloom by ̃ 10 days. The C–T model provides a physiological framework
for bloom forecasts in deciduous orchards. It integrates contrasting notions of chill and heat and
elucidates why abnormal winter temperatures may compromise bloom in deciduous orchards.

Comparison of phenological traits, growth patterns, and seasonal dynamics of non-structural
carbohydrate in Mediterranean tree crop species. 2019. Aude Tixier, Paula Guzman Delgado, Or
Sperling, Adele Amico Roxas , Emilio Laca , Maciej Zwieniecki. Scientific Reports (in second
revision)
Abstract: Despite non-structural carbohydrate (NSC) importance for tree productivity and
resilience, little is known about their seasonal regulations and trade-off with growth and
reproduction. We characterize the seasonal dynamics of NSC in relation to the aboveground
phenology and temporal growth patterns of three deciduous Mediterranean species: almond
(Prunus dulcis (Mill.) D. A. Webb), walnut (Juglans regia L.) and pistachio (Pistacia vera L.).
Seasonal dynamics of NSC were synchronous between wood tissues from trunk, branches and
twigs. Almond had almost identical levels and patterns of NSC variation in twigs, branches and
trunks whereas pistachio and walnut exhibited clear concentration differences among plant parts
whereby twigs had the highest and most variable NSC concentration, followed by branches and
then trunk. While phenology had a significant influence on NSC seasonal trends, there was no
clear trade-off between NSC storage and growth suggesting that both were similarly strong sinks
for NSC. A temporal trade-off observed at the seasonal scale was influenced by the phenology of
the species. We propose that late senescing species experience C allocation trade-off at the end of
the growing season because of C-limiting thermal conditions and priority allocation to storage in
order to survive winter.





























