Embed Size (px)
Citation preview

Breast Cancer Research and Treatment 31: 325-335, 1994. © 1994 Kluwer Academic Publishers. Printed in the Netherlands'.
Differentiation state and invasiveness of human breast cancer cell lines
Connie L. Sommers, Stephen W. Byers, Erik W. Thompson, Jeffrey A. Torri and Edward R Gelmann Department o f Cell Biology and the Vincent T. Lombardi Cancer Research Center, Georgetown University,
Washington, DC 20007, USA
Key words: breast cancer, vimentin, invasion, integrins, cadherins, epithelial
Summary
Eighteen breast cancer cell lines were examined for expression of markers of epithelial and fibroblastic differ- entiation: E-cadherin, desmoplakins, ZO-1, vimentin, keratin and 131 and ~4 integrins. The cell lines were distributed along a spectrum of differentiation from epithelial to fibroblastic phenotypes. The most well- differentiated, epithelioid cell lines contained proteins characteristic of desmosomal, adherens and tight j unc- tions, were adherent to one another on plastic and in the basement membrane matrix Matrigel and were keratin-positive and vimentin-negative. These cell lines were all weakly invasive in an in vitro chemoinvasion assay. The most poorly-differentiated, fibroblastic cell lines were E-cadherin-, desmoplakin- and ZO-l-nega- tive and formed branching structures in Matrigel. They were vimentin-positive, contained only low levels of keratins and were highly invasive in the in vitro chemoinvasion assay. Of all of the markers analyzed, vimentin expression correlated best with in vitro invasive ability and fibroblastic differentiation. In a cell line with unstable expression of vimentin, T47Dco, the cells that were invasive were of the fibroblastic type. The differ- entiation markers described here may be useful for analysis of clinical specimens and could potentially pro- vide a more precise measure of differentiation grade yielding more power for predicting prognosis.
Introduction
Breast tumors vary in their degree of differentia- tion with some tumors displaying relatively well-or- ganized glandular structures and some tumors con- taining sheets of pleiomorphic cells. Histologic grade, as defined by Bloom and Richardson, is based on irregularity in nuclear shape or size, num- ber of hyperchromatic nuclei and mitoses, and ex- tent of tubule formation and correlates with clinical prognosis [1]. Stenkvist et al. found that a combina- tion of mitotic frequency, size of primary tumor and differentiation of primary tumor (as determined by cluster size in fine needle biopsies) was highly pre-
dictive of recurrence within 2.5 years [2]. Part of the difficulty of using differentiation state as a prognos- tic indicator lies in the subjective nature of its eval- uation. Markers that could help define differentia- tion state might increase the power of this prognos- tic indicator.
We studied markers of epithelial differentiation in human breast cancer cell lines, immortal cultures established from the tumors or metastases of indi- vidual patients, to achieve several aims. First, we wanted to investigate markers that may be useful as prognostic indicators for future clinical studies. Sec- ondly, since the in vitro invasive abilities of these cell lines were known, we wanted to investigate if
Address for offprints: E.R Gelmann, Lombardi Cancer Research Center 'P' level, 3700 Reservoir Road, N.W. Washington, DC 20007, USA

326 CL Sommers et al.
any markers of epithelial differentiation correlated with in vitro invasiveness. We focused on epithelial cell adhesion and junction molecules and interme- diate filaments (tissue-specific cytoskeletal pro- teins) as markers since these molecules are likely to be involved in the organization of cell collectives. We have examined the expression and localization of members of two cell adhesion molecule families, a cadherin, involved in cell-cell adhesion, and two subunits of integrins, involved in cell-substrate ad- hesion. E-cadherin (epithelial cadherin, reviewed in [3]) is a homophilic cell adhesion molecule that functions in binding adjacent epithelial cells togeth- er. One could expect more invasive, metastatic cells to have less cell-cell adhesion and less E-cadherin. In fact, several recent clinical studies have shown a correlation between loss of E-cadherin expression and poorer differentiation grade and higher stage in breast cancer [4-6]. Integrin expression could be positively or negatively correlated with metastasis depending on whether the integrin functions to ad- here the carcinoma cell to basement membrane sur- rounding the primary tumor or whether it functions to aid in adhesion at a secondary metastatic site. Decreased expression of integrins ~[31 and c%[3 4 has been observed in some breast cancer specimens [7- 11].
Junctions involved in adhering normal epithelial cells together include adherens junctions (sites of attachment of actin filaments), desmosomes (sites of attachment of intermediate filaments), tight junctions (junctions that define the border between the apical and basolateral membranes) and gap junctions, which allow passage of small molecules between adjacent cells. We have analyzed adherens junctions in breast cancer cell lines by examining the presence and distribution of E-cadherin, a ma- jor component of adherens junctions. Loss of E- cadherin expression was correlated with poor dif- ferentiation grade in breast cancer and other can- cers [4-6, 12-16]. We examined desmosomal junc- tions using an antibody to desmoplakins I and II, components of desmosomes. Loss of immunoreac- tivity to desmosomal components has been observ- ed in poorly differentiated, invasive oral squamous cell carcinomas [17]. Tight junctions were examined using an antibody to ZO-1, a component of tight
junctions. We did not examine gap junctions, how- ever loss of eonnexins 26 and 43 (components of gap junctions) has already been described in breast can- cer cell lines [18].
Differentiation state is also reflected by expres- sion of tissue-specific intermediate filaments (see ref 19 for review). Keratins are normally expressed by epithelial cells and their derivative carcinomas. Vimentin is expressed by cells of mesenchymal ori- gin, such as fibroblasts and hematopoietic cells. We have described down regulation of keratin expres- sion and activation of vimentin expression in some breast cancer cell lines [20]. In addition there is a striking correlation between vimentin expression and in vitro invasiveness in these cell lines [21]. Vi- mentin expression has been observed in clinical breast cancer specimens, especially when alterna- tives to formalin fixation are employed [22-29]. Furthermore vimentin expression correlated with low levels of estrogen receptor and high Ki67 frac- tions, both poor prognostic indicators for breast cancer [23, 25, 26]. In this study, we have examined further the correlation between vimentin expres- sion and in vitro invasiveness in a cell line with vari- able expression of vimentin, T47Dco. Within this cell line, there was a significant correlation between cells that invaded a basement membrane-coated porous filter and vimentin expression.
Materials and methods
Cell culture
Cells were routinely cultured in IMEM (Biofluids, Rockville, MD) supplemented with 5 % heat inacti- vated fetal bovine serum (Biofluids) at 37 ° C in a humidified 5 % CO2 atmosphere. Cell lines were ob- tained from the sources described previously [30] or from the American Type Culture Collection, Rock- ville, MD (MDA-MB-435).
lmmunofluorescence microscopy
Cells grown on glass coverslips or 8-well chamber slides (Thomas Scientific, Swedesboro, N J) were
Differentiation state and invasiveness o f human breast cancer cell lines 327
MCF-7 SKBr3 MDA-MB-231
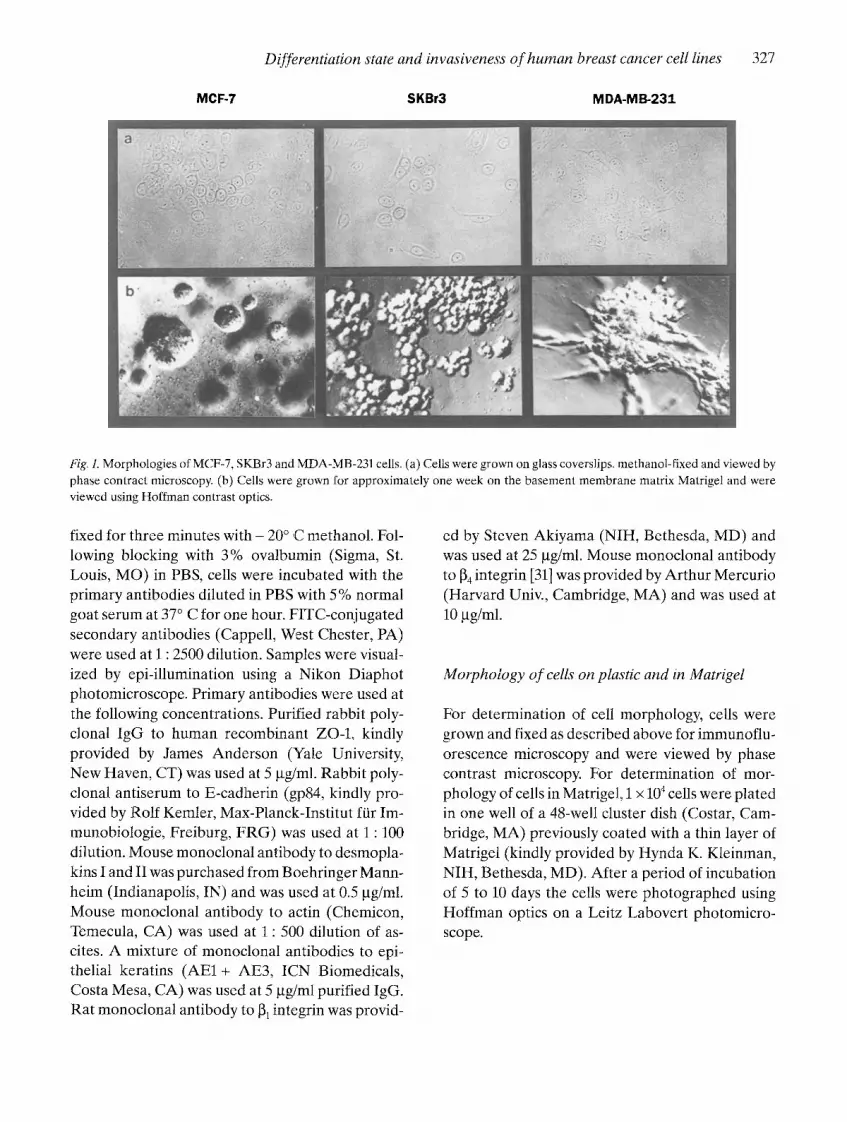
Fig. l. Morphologies of MCF-7, SKBr3 and MDA-MB-231 cells. (a) Cells were grown on glass coverslips, methanol-fixed and viewed by phase contract microscopy. (b) Cells were grown for approximately one week on the basement membrane matrix Matrigel and were viewed using Hoffman contrast optics.
fixed for three minutes with - 20 ° C methanol. Fol- lowing blocking with 3% ovalbumin (Sigma, St. Louis, MO) in PBS, cells were incubated with the primary antibodies diluted in PBS with 5% normal goat serum at 37 ° C for one hour. FfTC-conjugated secondary antibodies (Cappell, West Chester, PA) were used at l : 2500 dilution. Samples were visual- ized by epi-illumination using a Nikon Diaphot photomicroscope. Primary antibodies were used at the following concentrations. Purified rabbit poly- clonal IgG to human recombinant ZO-1, kindly provided by James Anderson (Yale University, New Haven, CT) was used at 5 gg/ml. Rabbit poly- clonal antiserum to E-cadherin (gp84, kindly pro- vided by Rolf Kemler, Max-Planck-Institut far Im- munobiologie, Freiburg, FRG) was used at 1 : 100 dilution. Mouse monoclonal antibody to desmopla- kins I and II was purchased from Boehringer Mann- heim (Indianapolis, IN) and was used at 0.5 gg/ml. Mouse monoclonal antibody to actin (Chemicon, Temecula, CA) was used at 1 : 500 dilution of as- cites. A mixture of monoclonal antibodies to epi- thelial keratins (AE1 + AE3, ICN Biomedicals, Costa Mesa, CA) was used at 5 gg/ml purified IgG. Rat monoclonal antibody to 131 integrin was provid-
ed by Steven Akiyama (NIH, Bethesda, MD) and was used at 25 gg/ml. Mouse monoclonal antibody to 134 integrin [31] was provided by Arthur Mercurio (Harvard Univ., Cambridge, MA) and was used at 10 gg/ml.
Morphology o f cells on plastic and in Matrigel
For determination of cell morphology, cells were grown and fixed as described above for immunoflu- orescence microscopy and were viewed by phase contrast microscopy. For determination of mor- phology of cells in Matrigel, 1 × 104 cells were plated in one well of a 48-well cluster dish (Costar, Cam- bridge, MA) previously coated with a thin layer of Matrigel (kindly provided by Hynda K. Kleinman, NIH, Bethesda, MD). After a period of incubation of 5 to 10 days the cells were photographed using Hoffman optics on a Leitz Labovert photomicro- scope.

328 CL Sommers et al.
In vitro invasion assays
Boyden chamber chemoinvasion assays have been described previously [30]. Briefly, cells were plated in the top chamber (3 x 105 cells/chamber) and a Matrigel-coated polycarbonate filter separated the top and bottom chambers. The bottom chamber contained conditioned medium for NIH3T3 cells as a chemoattractant. After 16 hours in a humidified, 5 % CO 2 37 ° C incubator, cells which had traversed to the bottom of the filters were stained with Diff- Quik (American Scientific Products, McGaw Park, IL) and quantitated electronically using the Zeiss IBAS 2000 image analysis system with Kontron Aiag processor interfaced with a Zeiss Axiophot microscope. For immunofluorescence microscopy of the top and bottom of the filters, instead of stain- ing with Diff-Quik, the filters were fixed with meth- anol for 10 minutes and stained as described for im-
munofluorescence microscopy as described above. Double staining for vimentin and actin was per- formed. For immunofluorescence microscopy of the tops of filters, ten-fold fewer cells (3 x 104 cells/ chamber) were plated at the onset of the experi- ment.
Results
Examination of the eighteen human breast cancer cell lines listed in Table i showed that the cell lines fell along a spectrum of epithelial differentiation that was related to in vitro invasiveness. The cell lines in Table 1 are listed with the most well-differ- entiated, epithelioid cell lines at the top of the table and the least well-differentiated, fibroblastic cell lines at the bottom of the table. Markers of differ- entiation and levels of estrogen receptor are shown
Table L S u m m a r y t a b l e o f m a r k e r e x p r e s s i o n in h u m a n b r e a s t c a n c e r cel l l i ne s
C e l l l i n e E R a E - c a d b D P - I / I I c Z O - I J V i m e [3t f [348 M a t r i g e l In vitro
m o r p h o l o g y h i n v a s i o n i
B T 4 8 3 + + + + + + - + + - f u s e d +
M D A - M B - 1 7 5 - 7 - + + + + - + - f u s e d +
M C F - 7 + + + + + + - + - f u s e d +
Z R - 7 5 - B + + + + + + - + ~ f u s e d +
T 4 7 D + + + + - + J f u s e d +
B T 4 7 4 + + + - - + - f u s e d +
M D A - M B - 3 6 1 + + + - - + _5 f u s e d +
M D A - M B - 4 6 8 - - + - - + + / - f u s e d +
S K - B r - 3 - - + / - + - + / - - s p h e r i c a l +
C A M A - 1 + - + + - + - s p h e r i c a l +
M D A - M B - 4 5 3 - - + / - - - + + s p h e r i c a l +
M D A - M B - 1 3 4 + - + / _ _ _ + .5 s p h e r i c a l +
M D A - M B - 4 3 6 - - - + + + - s t e l l a t e + +
M D A - M B - 4 3 5 . . . . + + - s t e l l a t e + +
M D A - M B - 2 3 1 . . . . + + - s t e l l a t e + + +
B T 5 4 9 . . . . + + - s t e l l a t e + + +
H S 5 7 8 t . . . . + + - s t e l l a t e + + +
M C F - 7 A o ~ . . . . + + - s t e l l a t ek + +
E s t r o g e n r e c e p t o r s t a t u s as p r e v i o u s l y p u b l i s h e d o r as d e t e r m i n e d u s i n g t h e A b b o t t E R - E I A m o n o c l o n a l a n t i b o d y k i t . b E - c a d h e r i n /
u v o m o r u l i n e x p r e s s i o n as d e t e c t e d b y i m m u n o f l u o r e s c e n c e m i c r o s c o p y u s i n g r a b b i t p o l y c l o n a l a n t i s e r u m gp84 . c D e s m o p l a k i n I + I I
e x p r e s s i o n as d e t e c t e d b y i m m u n o f l u o r e s c e n c e m i c r o s c o p y u s i n g m o u s e m o n o c l o n a l a n t i b o d y to D P I + I I . d Z O - 1 e x p r e s s i o n a s d e t e c t e d
b y i m m u n o f l u o r e s c e n c e m i c r o s c o p y u s i n g r a b b i t p o l y c l o n a l a n t i s e r u m to Z O - 1 . ° V i m e n t i n e x p r e s s i o n as d e t e c t e d b y i m m u n o f l u o r e s -
c e n c e m i c r o s c o p y u s i n g m o u s e m o n o c l o n a l a n t i b o d y V 9 to v i m e n t i n , f B e t a - 1 i n t e g r i n e x p r e s s i o n as d e t e c t e d b y i m m u n o f l u o r e s c e n c e
m i c r o s c o p y u s i n g r a t m o n o c l o n a l a n t i b o d y to b e t a - 1 i n t e g r i n , g B e t a - 4 i n t e g r i n e x p r e s s i o n as d e t e c t e d b y i m m u n o f l u o r e s c e n c e m i c r o s c o -
p y u s i n g m o u s e m o n o c l o n a l a n t i b o d y to b e t a - 4 i n t e g r i n , h M o r p h o l o g y o f ce l l s o r c o l o n i e s a f t e r g r o w t h in M a t r i g e l f o r o n e to t w o w e e k s .
A c t i v i t y in B o y d e n c h a m b e r c h e m o i n v a s i o n a s say , g r a d e d as p e r c e n t o f M D A - M B - 2 3 1 a c t i v i t y : + 0 - 4 0 % , + + 4 0 - 8 0 % , + + + > 8 0 % .
J O c c a s i o n a l s t a i n i n g a t f o c a l c o n t a c t s , k S p h e r i c a l f o r a b o u t 10 d a y s , t h e n s t e l l a t e .

Differentiation state and invasiveness of human breast cancer cell lines 329
Fig. 2. Immunofluorescence microscopy of markers of epithelial differentiation for MCF-7, SKBr3 and MDA-MB-231 cells. Immunoflu- orescence microscopy of tight junction protein ZO-1 (a-c), desmoplakins I and II (d-f), keratins (g-i) and vimentin (f-l) is shown for MCF-7 (a, d, g, j), SKBr3 (b, e, h, k) and MDA-MB-231 (c, f, i, 1).
with reference to invasive ability. The most well-dif- ferentiated, epithelioid cells grew as interconnect- ed colonies of polygonal cells on plastic and as fused colonies in the basement membrane matrix Matri- gel (see Fig. 1, MCF-7). The most poorly-differen- tiated, fibroblastic cells had various shapes includ-
ing spindle shapes when grown on plastic and tend- ed to grow over one another at high cell densities. In Matrigel, they formed colonies with large stellate projections (see Fig. 1, MDA-MB-231). Some cell lines with an intermediate degree of differentiation

330 C L Somrners et al.
Fig. 3. [~ integrin subunit expression in MCF-10A and breast cancer cell lines. Immunofluorescence microscopy of ~ integrin is shown for MCF-10A cells (a) and MDA-MB-435 cells (b). Immunofluorescence microscopy of [34 integrin is shown for MCF-10A cells, focused at the celbsubstrate interface (c), and MDA-MB-453 cells (d).
had varied shapes on plastic and grew as nonfused spheres in Matrigel (see Fig. 1, SKBr3).
We determined that some markers correlated with degree of differentiation in these breast cancer cell lines. As described previously [30], the most well-differentiated epithelioid breast cancer cell lines expressed E-cadherin (uvomorulin) and formed fused spherical colonies in Matrigel. Treat- ment of E-cadherin-positive cells with blocking an- tibody to E-cadherin resulted in a detached spher- ical morphology similar to that seen for SkBr3 in Fig. 1 [30]. Therefore, this morphological feature was indeed dependent on E-cadherin expression.
In this study, we also examined expression of des- moplakins I and II, components of desmosomes and ZO-1, a component of tight junctions. Desmo- somes, sites of attachment of keratin intermediate filaments, provide structural integrity to epithelial tissues. Loss of desmosomal structures could be necessary for an invasive phenotype. In the cell lines examined here, the most invasive cells did not express desmoplakins I and II (see Fig. 2f, MDA- MB-231). Well-differentiated cells such as MCF-7 did show bead-like staining at cell-cell borders us- ing the desmoplakin antibody (see Fig. 2d). Cells of
intermediate differentiation, such as SKBr3, showed an intermediate staining pattern with some cell-cell borders displaying desmosomal immuno- reactivity (Fig. 2e). A similar pattern of expression was seen for tight junctions, which form a seal around normal epithelial cells separating apical from basolateral components. Well-differentiated, epithelioid cells such as MCF-7 showed a line of staining at cell-cell borders (Fig. 2a), whereas fibro- blastic cells such as MDA-MB-231 showed no stain- ing (Fig. 2c) and intermediate cells such as SKBr3 showed an intermediate phenotype (Fig. 2b).
We had previously published the intermediate filament expression for most of the cell lines in Ta- ble 1 [20]. The most poorly-differentiated, fibro- blastic cell lines such as MDA-MB-231 expressed vimentin (Fig. 21) whereas cell lines such as MCF-7 and SKBr3 did not (Fig. 2j, k). The cell lines that expressed vimentin had reduced but detectable lev- els of keratins (see Fig. 2i) whereas cells lines such as MCF-7 and SKBr3 had easily detectable keratin intermediate filaments (Fig. 2g, h).
We also examined the integrin subunits [~ and ~4 since the integrin receptors ~213~ and %[34 are down- regulated in some breast cancer specimens [7-10]. ~

Differentiation state and invasiveness o f human breast cancer cell lines 331
integrin expression was detected in all of the cell
lines examined (see Table 1). The staining in
MCF-10A cells (established f rom normal m a m m a r y epithelium, ref 32) and in cell lines such as MCF-7 was concentrated at cell borders (see Fig. 3a). In poorly differentiated, fibroblastic cell lines such as MDA-MB-435 staining was observed all over the cell surface (see Fig. 3b). A different pat tern of
staining was observed for ~4 integrin. In MCF-10A cells, a scale-like pat tern was observed with ~34 in- tegrin concentrated at cell-substrate levels (Fig. 3c). However , the great majori ty of breast cancer cell lines did not have detectable expression of ~4 inte-
grin (see Table 1). The cell surface staining pat tern observed for MDA-MB-453 ( intermediate differ- entiation state) is shown in Fig. 3d. The integrin staining pat terns for MCF-10A were shown here since no cell lines in Table 1 showed a 'normal ' ex- pression pat tern for [34 integrin.
Matrigel morphology and in vitro invasive abil- ities correlated with expression of E-cadherin and vimentin (measures of differentiation state) in breast cancer cell lines [30]. In vitro invasion was measured by the ability of the cells to traverse a Ma- trigel-coated porous polycarbonate filter. As sum- marized in Table 1, the most well- differentiated cell lines (E-cad+/vim-) , which formed fused, spherical colonies in Matrigel, were only weakly invasive in
vitro. In termedia te cell lines (E-cad- /v im-) , which
formed detached spheres in Matrigel, were also only weakly invasive in vitro. The most poorly-dif-
ferentiated cells (E-cad-/vim+), which formed ag- gregates with stellate projections in Matrigel, were
highly invasive in vitro. Therefore, of all of the
markers examined, vimentin expression correlated
best with in vitro invasiveness. No strict correlation between invasiveness and estrogen receptor levels was found.
In general, the cell lines in Table 1had homogene-
ous expression of the markers examined, especially those cell lines at the extremes of differentiation.
However , the cell line T47Dco [33], was heteroge- nous in its expression of several of the markers de- scribed here, in particular, in its expression of vi- mentin. We hypothesized that if vimentin was a marker of invasiveness, that the invasive cells of
T47Dco would be vimentin-positive. To test this hy- pothesis, after the in vitro invasion assay described above, cells on the top or the bot tom of the filters were stained for vimentin expression. As shown in
Table 2, an average of 61.5% of cells on the top were vimentin-positive whereas nearly all of the cells
that had invaded to the bo t tom of the filter (94.5 %) were vimentin-positive. The number of vimentin- positive cells differ on the top versus the bo t tom of the filter as determined by Chi-square analysis (p < 0.0001). The overall percentage of vimentin-posi- tive cells did not differ on plastic and in the in vitro
invasion assay.
Discussion
Analysis of eighteen human breast cancer cell lines
has shown that these breast cancer cell lines fall along a spectrum of epithelial differentiation. Some
Table 2. Chemoinvasion of T47Dco cells.
# V + cells # Total cells % V + ceils
Expt 1 - top 226 357 63 Expt I - bottom 608 640 95 Expt 2 - top 284 473 60 Expt 2 - bottom 547 580 94 Both expts - top 61.5 _+ 2.1 Both expts - bottom 94.5 _+ 0.7
T47Dco cells were placed in the upper chamber of a modified Boyden chamber apparatus with a Matrigel-coated filter separating the top and bottom chambers. Chemoattractant was present in the bottom chamber. After 16 hours, the filters were fixed with methanol and cells on the top or the bottom of the filters were stained by immunofluorescence microscopy. Vimentin-positive cells (V+) were detected with monoclonal antibody vg. Total cells were detected with a monoclonal antibody to actin. Results from two experiments (expts) are shown. Means and standard deviations of % V+ cells from the two experiments are shown in the last two rows.

332 CL Sommers et aL
Well-differentiated Epithelioid
Poorly-differentiated Fibroblastic
e.g. BT483 e.g. MDA-MB-468 e.g. MDA-MB-436 MCF-7 CAMA-I MDA-MB-231 ZR-75-B SK-Br-3 BT549 T47D MDA-MB-453 HS578T
F/g. 4. Hypothetical progression of breast cancer. Breast cancer cell lines represent a spectrum of epithelial differentiation with well- differentiated lines represented at the left and poorly-differentiated lines represented at the right. Morphology in Matrigel is depicted in cartoon form. Wedges indicate amount of expression of E-cadherin (E-cad), desmoplakins I and II (DP I/II), tight junction protein ZO-1 and vimentin.
markers of epithelial differentiation (E-cadherin, desmoplakins, ZO-1) are present in normal breast epithelial cells and in well-differentiated cell lines whereas vimentin is expressed in poorly-differen- tiated cell lines (see Fig. 4). We hypothesize that breast cancer cells can progress along this spectrum during malignant progression. The MCF-7AD R cell line, selected for resistance to the chemotherapeut- ic drug adriamycin, and several other adriamycin- and vinblastine-resistant cell lines provide in vitro
evidence for this hypothesis [34]. The drug-resistant cell lines were more poorly-differentiated than the parental cell lines indicating a transition toward fi- broblastic differentiation.
Degree of differentiation was also compared to invasive ability in the cell lines as determined by analysis of Matrigel morphology and in vitro che- moinvasion ability. The most poorly-differentiated cells were the most invasive in vitro. Of the markers
that were examined in this study, vimentin expres- sion correlated best with in vitro invasive ability. Breast cancer cells that were selected for resistance to adriamycin (MCF-7ADR) were more invasive than parental cells in addition to being more fibroblastic [3@
Of the epithelial adhesion and junction mole- cules examined, E-cadherin was lost earliest in ma- lignant progression. Therefore E-cadherin may be a useful marker for defining a well-differentiated state. Indeed in the clinical studies reported thus far for several types of carcinomas, a strong correlation between E-cadherin and differentiation grade was found [4-6, 12-16]. In bladder cancer, E-cadherin expression also correlated with prognosis [16].
The desmoplakins and ZO-1, components of des- mosomes and tight junctions, were absent in poor- ly-differentiated, fibroblastic cell lines. Cell lines of intermediate degree of differentiation had variable

Differentiation state and invasiveness o f human breast cancer cell lines 333
expression of these markers. In some cases, immu- noreactive protein was detected but was not orga- nized into structures resembling those seen in well- differentiated cell lines. Loss of expression or func- tion of these junction molecules and the cell adhe- sion molecule E-cadherin may be required for an invasive phenotype [35-38]. This hypothesis could be tested by expressing these molecules in invasive breast cancer cells and testing the effect on inva- siveness. In the case of MDA-MB-435 cells, trans- fection of E-cadherin has in fact yielded a less in- vasive phenotype [36].
On the other hand, vimentin expression positive- ly correlated with invasive ability in vitro. However, we showed previously that transfection of MCF-7 cells with the vimentin cDNA did not result in an alteration of invasive ability [34]. Most likely, many factors, some of which are described here, are in- volved in the invasive phenotype. Other factors that are necessarily involved in the invasive phenotype include proteins that contribute to proteolytic ac- tivity. Future research which addresses what factors regulate differentiation state in breast cancer cells may allow exogenous control of differentiation state in these cells. In addition, these breast cancer cell lines could be treated with potential differen- tiating agents to screen for agents that may differ- entiate breast cancer cells in vivo.
We examined breast cancer cell lines for expres- sion of 13I and 134 integrin subunit expression to see if gain or loss of either of these subunits correlated with invasive ability. The patterns among the breast cancer cell lines were relatively uniform. 131 subunit was detected in all of the cell lines tested. This is in agreement with reports from clinical specimens [8]. However, integrins such as c~2[31 may still downreg- ulated in these cell lines as they are in breast cancer tissues [7, 8,10], which could be revealed by screen- ing with additional antibodies. On the other hand, most of the breast cancer cell lines did not express 134 integrin subunit whereas MCF-10A cells, establish- ed from normal mammary epithelium, did express this subunit. Therefore, integrins that contain [34 that are normally detected in breast tissue such as %134 may be downregulated early in malignancy. The loss of expression of [34 may provide an advan-
tage for cells to leave the primary tumor and be- come metastatic.
Markers that help to define differentiation state in breast cancer could provide useful prognostic in- formation. Instead of a subjective evaluation of de- gree of tubule formation or degree of differentia- tion, an index composed of expression of several markers, such as E-cadherin, desmoplakin, keratin and vimentin could be compared with other prog- nosticators in clinical studies. The cell lines de- scribed here provide a model system for screening such markers. In the future, analysis of markers chosen from in vitro studies and analysis of progno- sis can be applied to tumor specimens.
Acknowledgements
The authors would like to thank Miguel Saceda for providing information on estrogen receptor levels for some of the breast cancer cell lines used here. Study of markers of epithelial differentiation was made possible by the generosity of several col- leagues in providing antibodies: James Anderson (ZO-1 antibody), Roll Kemler (gp84), Steven Aki- yama ([31 integrin antibody) and Arthur Mercurio (134 integrin antibody). We would lastly like to thank Marc Lippman for encouragement and support throughout the course of these studies and Oscar Rodbell for expert assistance in photography.
References
1. Bloom HJG, Richardson WW: Histological grading and prognosis in breast cancer. Br J Cancer 11: 359-377, 1957
2. Stenkvist B, Bengtsson E, Dahlqvist B, Eklund G, Eriksson O, Jarkrans T, Nordin B: Predicting breast cancer recur- rence. Cancer 50: 2884-2893, 1982
3. Takeichi M: Cadherins: a molecular family important in se- lective cell-cell adhesion. Annu Rev Biochem 59: 237-252, 1990
4. Oka H, Shiozaki H, Kobayashi K, Inoue M, Tahara H, Ko- bayashi T, Takatsuka Y, Matsuyoshi N, Hirano S, Takeichi M, Mori T: Expression of E-cadherin cell adhesion mole- cules in human breast cancer tissues and its relationship to metastasis. Cancer Res 53: 1696-1701, 1993
5. Gamallo C, Palacios J, Suarez A, Pizarro A, Navarro R Quintanilla M, Cano A: Correlation of E-cadherin expres-

334 C L S o m m e r s et al.
sion with differentiation grade and histological type in breast carcinoma. Am J Patho1142: 987-993, 1993
6. Rasbridge SA, Gillett CE, Sampson SA, Walsh FS, Millis RR: Epithelial (E-) and placental (P-) cadherin cell adhe- sion molecule expression in breast carcinoma. J Patho1169: 245-250, 1993
7. Pignatelli M, Cardillo MR, Hanby A, Stamp GWH: Inte- grins and their accessory adhesion molecules in mammary carcinomas: loss of polarization in poorly differentiated tu- mors. Hum Pathol 23: 1159-1166, 1992
8. Zutter MM, Mazoujian G, Santoro SA: Decreased expres- sion of integrin adhesive protein receptors in adenocarcino- ma of the breast. Am J Patho1137: 863-870, 1990
9. Jones JL, Critchley DR, Walker RA: Alteration of stromal protein and integrin expression in breast-a marker of pre- malignant change? J Patho1167: 399406, 1992
10. Pignatelli M, Hanby AM, Stamp GWH: Low expression of beta-l, alpha-2 and alpha-3 subunits of VLA integrins in ma- lignant mammary tumours. J Patho1165: 25-32, 1991
11. Koukoulis GK, Virtanen I, Korhonen M, Laitinen L, Qua- ranta V, Gould VE: Immunohistochemical localization of integrins in the normal, hyperplastic, and neoplastic breast. Am J Pathol 4: 787-799, 1991
12. Mayer B, Johnson JR Leitl F, Jauch KW, Heiss MM, Schildb- erg FW, Birchmeier W, Funke I: E-cadherin expression in primary and metastatic gastric cancer: down-regulation cor- relates with cellular dedifferentiation and glandular disin- tegration. Cancer Res 53: 1690-1695,1993
13. Schipper JH, Frixen UH, Behrens J, Unger A, Jahnke K, Birchmeier W: E-cadherin expression in squamous cell car- cinomas of head and neck: inverse correlation with tumor dedifferentiation and lymph node metastasis. Cancer Res 51: 6328-6337,1991
14. Umbas R, Schalken JA, Aalders TW, Carter BS, Karthaus HF, Schaafsma HE, Debruyne FM, Isaacs WB: Expression of the cellular adhesion molecule E-cadherin is reduced or absent in high-grade prostate cancer. Cancer Res 52: 5104- 5109, 1992
15. Dorudi S, Sheffield JR Poulsom R, Northover JM, Hart IR: E-cadherin expression in colorectal cancer. An immunocy- tochemical and in situ hybridization study. Am J Patho1142: 981-986, 1993
16. Bringuier PR Umbas R, Schaafsma HE, Karthaus HFM, Debruyne FMJ, Schalken JA: Decreased E-cadherin immu- noreactivity correlates with poor survival in patients with bladder tumors. Cancer Res 53: 3241-3245,1993
17. Harada T, Shinohara M, Nakamura S, Shimada M, Oka M: Immunohistochemical detection of desmosomes in oral squamous cell carcinomas: correlation with dedifferentia- tion, mode of invasion, and metastatic potential. Int J Oral Maxillofac Surg 21: 346-349, 1992
18. Lee SW, Tomasetto C, Paul D, Keyomarsi K, Sager R: Tran- scriptional downregulation of gap-junction proteins blocks junctional communication in human mammary tumor cell lines. J Cell Bio1118: 1213-1221,1992
19. Bernal SD, Stahel RA: Cytoskeleton-associated proteins:
their role as cellular integrators in the neoplastic process. CRC Crit Rev Oncol Hematol 3: 191-204,1985
20. Sommers CL, Walker-Jones D, Heckford SE, Worland P, Valverius E, Clark R, McCormick F, Stampfer M, Abula- rach S, Gelmann EP: Vimentin rather than keratin expres- sion in some hormone-independent breast cancer cell lines and in oncogene-transformed mammary epithelial cells. Cancer Res 49: 42584263, 1989
21. Thompson EW, Paik S, Brunner N, Sommers CL, Zugmaier G, Clarke R, Shima TB, Torri J, Donohue S, Lippman ME, Martin GR, Dickson RB: Association of increased base- ment membrane-invasiveness with absence of estrogen re- ceptor and expression of vimentin in human breast cancer cell lines. J Cell Physio1150: 534-544, 1992
22. Heatley M, Whiteside C, Maxwell R Toner P: Vimentin ex- pression in benign and malignant breast epithelium. J Clin Pathol 46: 441-445, 1993
23. Domagala W, Lasota J, Bartowiak J, Weber K, Osborn M: Vimentin is preferentially expressed in human breast carci- nomas with low estrogen receptor and high Ki-67 growth fraction. Am J Patho1136: 219-227,1990
24. Domagala W, Leszek W, Lasota J, Weber K, Osborn M: Vi- mentin is preferentially expressed in high-grade ductal and medullary, but not in lobular breast carcinomas. Am J Path- o1137: 1059-1064, 1990
25. Raymond WA, Leong AS-Y: Vimentin - A new prognostic parameter in breast carcinoma? J Patho1158: 107-114, 1989
26. Cattoretti G, Andreola S, Clemente C, D'Amato L, Rilke F: Vimentin and p53 expression on epidermal growth factor receptor-positive, oestrogen receptor-negative breast carci- nomas. Br J Cancer 57: 353-357, 1988
27. Gould VE, Kouloulis GK, Jansson DS, Nagle RB, Franke WW, Moll R: Coexpression patterns of vimentin and glial filament protein with cytokeratins in the normal, hyperplas- tic, and neoplastic breast. Am J Patho1137: 1143-1155, 1990
28. Wada T, Yasutomi M, Hashmura K, Kunikata M, Tanaka T, Mori M: Vimentin expression in benign and malignant le- sions in the human mammary gland. Anticancer Res 122: 1973-1982, 1992
29. Azumi N, Battifora H: The distribution of vimentin and ker- atin in epithelial and nonepithelial neoplasms. Am J Clin Patho1188: 286-296, 1987
30. Sommers CL, Thompson EW, Torri JA, Kemler R, Gel- mann ER Byers SW: Cell adhesion molecule uvomorulin ex- pression in human breast cancer cell lines: relationship to morphology and invasive capacities. Cell Growth and Diff 2: 365-372, i991
31. Van Waes C, Kozarsky KF, Warren AB, Kidd L, Paugh D, Liebert M, Carey TE: The A9 antigen associated with ag- gressive human squamous carcinoma is structurally and functionally similar to the newly defined integrin alpha 6 be- ta 4. Cancer Res 51: 2395-2402, 1991
32. Tait L, Soule HD, Russo J: Ultrastructural and immunocy- tochemical characterization of an immortalized human breast epithelial cell line, MCF-10. Cancer Res 50: 6087- 6094,1990

Differentiation state and invasiveness of hurnan breast cancer cell lines 335
33. Graham ML, Krett NL, Miller LA, Leslie KK, Gordon DF, Wood WM, Wei LL, Horwitz KB: T47Dco cells, genetically unstable and containing estrogen receptor mutations, are a model for the progression of breast cancers to hormone re- sistance. Cancer Res 50: 6208-6217, 1990
34. Sommers CL, Heckford SE, Skerker JM, Worland R Torri J, Thompson EW, Byers SW, Gelmann EP: Loss of epithelial markers and acquisition of vimentin expression in adriamy- cin- and vinblastine-resistant human breast cancer cell lines. Cancer Research 52: 5190-5197, 1992
35. Behrens J, Mareel MM, Van Roy FM, Birchmeier W: Dis- secting tumor cell invasion: epithelial cells acquire invasive
properties after the loss of uvomorulin-mediated cell-cell adhesion. J Cell Bio1108: 2435-2447, 1989
36. Frixen UH, Behrens J, Sachs M, Eberle G, Voss B, Warda A, Lochner D, Birchmeier W: E-cadherin-mediated cell-cell adhesion prevents invasiveness of human carcinoma cells. J Cell Bio1113: 173-185,1991
37. Chen WC, Obrink B: Cell-cell contacts mediated by E-cad- herin (uvomorulin) restrict invasive behavior of L-cells. J Cell Bio1114: 319-327, 1991
38. Vleminckx K, Vakaet L, Mareel M, Fiers W, Van Roy F: Ge- netic manipulation of E-cadherin expression by epithelial tumor cells reveals an invasion suppressor role. Cell 66:10% 119,1991




























