Embed Size (px)
Citation preview

1 23
Fish Physiology and Biochemistry ISSN 0920-1742Volume 38Number 2 Fish Physiol Biochem (2012) 38:441-454DOI 10.1007/s10695-011-9525-9
Digestive enzyme activities during earlyontogeny in Common snook (Centropomusundecimalis)
L. D. Jimenez-Martinez, C. A. Alvarez-González, D. Tovar-Ramírez, G. Gaxiola,A. Sanchez-Zamora, F. J. Moyano,F. J. Alarcón, G. Márquez-Couturier, etal.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media B.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Digestive enzyme activities during early ontogenyin Common snook (Centropomus undecimalis)
L. D. Jimenez-Martinez • C. A. Alvarez-Gonzalez • D. Tovar-Ramırez •
G. Gaxiola • A. Sanchez-Zamora • F. J. Moyano • F. J. Alarcon • G. Marquez-Couturier •
E. Gisbert • W. M. Contreras-Sanchez • N. Perales-Garcıa • L. Arias-Rodrıguez •
J. R. Indy • S. Paramo-Delgadillo • I. G. Palomino-Albarran
Received: 19 December 2009 / Accepted: 3 June 2011 / Published online: 14 June 2011
� Springer Science+Business Media B.V. 2011
Abstract Common snook (Centropomus undecimal-
is) is one of the most important marine species under
commercial exploitation in the Gulf of Mexico; for this
reason, interest in developing its culture is a priority.
However, larviculture remains as the main bottleneck
for massive production. In this sense, our objective was
to determine the changes of digestive enzymes activ-
ities using biochemical and electrophoretic techniques
during 36 days of Common snook larviculture fed with
live preys (microalgae, rotifers, and Artemia). During
larviculture, all digestive enzymatic activities were
detected with low values since yolk absorption, 2 days
after hatching (dah) onwards. However, the maximum
values for alkaline protease (6,500 U mg protein-1),
trypsin (0.053 mU 9 10-3 mg protein-1), and Leu-
cine aminopeptidase (1.4 9 10-3 mU mg protein-1)
were detected at 12 dah; for chymotrypsin at 25 dah
(3.8 9 10-3 mU mg protein-1), for carboxypeptidase
A (280 mU mg protein-1) and lipase at 36 dah
(480 U mg protein-1), for a-amylase at 7 dah
(1.5 U mg protein-1), for acid phosphatases at 34 dah
(5.5 U mg protein-1), and finally for alkaline phos-
phatase at 25 dah (70 U mg protein-1). The alkaline
protease zymogram showed two active bands, the first
(26.3 kDa) at 25 dah onwards, and the second
(51.6 kDa) at 36 dah. The acid protease zymogram
showed two bands (RF = 0.32 and 0.51, respectively)
at 34 dah. The digestive enzymatic ontogeny of
C. undecimalis is very similar to other strictly marine
carnivorous fish, and we suggest that weaning process
should be started at 34 dah.
Keywords a-Amylase � Centropomus undecimalis �Common snook � Lipase � PAGE � Phosphatase �Protease
L. D. Jimenez-Martinez � C. A. Alvarez-Gonzalez (&) �G. Marquez-Couturier � W. M. Contreras-Sanchez �N. Perales-Garcıa � L. Arias-Rodrıguez �J. R. Indy � S. Paramo-Delgadillo
DACBIOL Laboratorio de Acuacultura, Universidad
Juarez Autonoma de Tabasco, Carretera Villahermosa-
Cardenas km 0.5, 86139 Villahermosa, Tabasco, Mexico
e-mail: [email protected]
G. Gaxiola � A. Sanchez-Zamora �I. G. Palomino-Albarran
Unidad Multidisciplinaria de Docencia e Investigacion,
Facultad de Ciencias, UNAM, Puerto de abrigo s/n, Sisal,
Yucatan, Mexico
F. J. Moyano � F. J. Alarcon
Departamento de Biologıa Aplicada, Escuela Politecnica
Superior, Universidad de Almerıa, 04120 La Canada de
San Urbano, Almerıa, Spain
D. Tovar-Ramırez
Centro de Investigaciones Biologicas del Noroeste
(CIBNOR), Mar Bermejo 195, Col. Playa Palo de Santa
Rita, 23090 La Paz, B.C.S., Mexico
E. Gisbert
IRTA – Sant Carles de la Rapita, Crta. Poble Nou km 5.5,
43540 Sant Carles de la Rapita, Tarragona, Spain
123
Fish Physiol Biochem (2012) 38:441–454
DOI 10.1007/s10695-011-9525-9
Author's personal copy

Introduction
Marine fish culture in Mexico is a recent activity
mainly based on some species with high economic
value and adequate biological characteristics for their
culture, such as spotted sand bass (Paralabrax
maculatofasciatus), leopard grouper (Mycteroperca
rosacea), bullseye puffer (Sphoeroides annulatus),
rose snapper (Lutjanus guttatus), and more recently
cobia (Rachycentrum canadum) (Aviles-Quevedo
et al. 1995; Gracia-Lopez et al. 2004; Komar et al.
2004; Ibarra-Castro and Duncan 2007; Holt et al.
2007). The fat snook (Centropomus parallelus) and
the common snook (Centropomus undecimalis) of the
family Centropomidae, a group with highly important
commercial value in Mexico and the United States,
have been widely studied to understand their basic
biology and aiming for developing their culture
(Chavez 1961; Stephen and Shafland 1982; Sherwood
et al. 1993; Ramirez and Cerqueira 1994; Grier and
Taylor 1998; Grier 2000; Cequeira and Brugger
2001; Alvarez-Lajonchere et al. 2002; Tarcisio et al.
2005; Gracia-Lopez et al. 2006; Wainwright et al.
2006; Yanes-Roca et al. 2009). However, for
C. undecimalis, the bottleneck is still the massive
fry production due to feeding problems when micro-
algae, rotifers, and Artemia nauplii are used. These
live feeds are considered not adequate for fish larval
culture (Versichelle et al. 1989; Garcıa-Ortega et al.
1998), resulting in low growth and survival. For this
reason, many studies have been conducted to under-
stand the digestive physiology during early ontogeny
with many species, such as the seabream, Sparus
aurata (Moyano et al. 1996), Siberian sturgeon,
Acipenser baeri (Gisbert et al. 1999), white bream,
Diplodus sargus (Cara et al. 2003), yellowtail
amberjack, Seriola lalandi (Chen et al. 2006),
common seabream, Pagrus pagrus (Darias et al.
2006), P. maculatofasciatus (Alvarez-Gonzalez et al.
2008, 2010), and the orange-spotted grouper Epi-
nephelus coioides (Shaozhen et al. 2008). These
studies allow understanding the right moment to
conduct early weaning using artificial diets (Brock
et al. 1992; Zambonino-Infante and Cahu 1994;
Ribeiro et al. 1999; Cara et al. 2003; Fabillo et al.
2004). In consequence, the objective of this work was
to assess the development of digestive enzymes using
biochemical and electrophoretical techniques during
early ontogeny of C. undecimalis.
Materials and methods
Rearing and sampling of larvae
Twelve adult common snook C. undecimalis (4–5 kg
per fish) were maintained under controlled conditions
in four 13-m3 circular plastic tanks at the Unidad
Multidisciplinaria de Docencia e Investigacion
(UMDI) from UNAM in Sisal, Merida, Mexico for
2 years. Spawning induction was done in adults using
cholesterol implants with 150 lg of sGnRHa. fish-1
(Ovaplant, Syndel, Western Chemical, Fendale, WA,
USA). A total of 1,10,000 embryos were obtained after
32 h after injection. After the embryos hatched, the
yolk-sac larvae (3,250) were collected by siphoning
and placed in a 400-l cylinder-conical tank with
constant water exchange, and continuous aeration until
the larvae absorbed the yolk (24 h later). Salinity
(35.2 ± 1.1 ppt), dissolved oxygen (6.0 ± 0.3
mg l-1), and temperature (29.9 ± 1.1�C) were mon-
itored daily.
Larvae were fed four times per day (8:00, 12:00,
16:00, and 20:00 h), starting with green water culture,
using the microalgae Nannochloropsis sp.
(20 9 106 cells ml-1) and S-type rotifers Brachionus
rotundiormis (R, 2–10 preys ml-1) from mouth open-
ing (day 1 after hatching) until 10 dah. Rotifers were
mixed with newly hatched Artemia nauplii (AN, INVE
Aquaculture, Belgium, 2–10 preys ml-1) from day 10
until 25 dah and offered to the larvae. Finally, larvae
were fed exclusively with lipidic enriched (SELCO,
INVE Aquaculture, Belgium) Artemia meta-nauplii
(EAMN, 2–15 preys ml-1) from day 25 up to 36 dah
(end of the experiment). Nine samples of feed larvae
were taken from one culture tank using a 500-lm-
diameter mesh, in triplicate (the numbers in parenthe-
ses are the numbers of larvae sampled per replicate) on
day 0 (embryos, 80), 1 (600), 3 (600), 5 (600), 7 (600),
12 (400), 25 (100), 34 (50), and 36 (30) after hatching.
They were frozen with liquid nitrogen and stored at
-80�C until analysis was done. For growth analysis,
samples of 10 larvae were taken and fixed with
buffered borate formalin solution (4%) to measure
the total length for each larvae using a digital caliper
(Neiko-HKMUD473, Neiko-HKMUD473, CA, USA).
The individual wet weight (mg) of each larva was also
recorded with an analytic balance (OHAUS-Phoenix
GH-300, Pine Brook, NJ, USA; precision of 10-4 g)
after elimination of water excess with filter paper.
442 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy

Biochemical analyses
Sampled larvae were dissected individually to
remove the head and tail, and the visceral bulks
were homogenized as pool (30 mg ml-1) in cold
50 mmol l-1 Tris–HCl 20 mmol l-1 CaCl2 buffer,
pH 7.5. The supernatant obtained after centrifugation
(16,000g for 15 min at 5�C) was stored at -20�C
until enzyme analysis. The concentration of soluble
protein was determined by the Bradford (1976)
method using bovine serum albumin as a standard.
Total alkaline protease activity was measured
using casein (0.5%) in 50 mmol l-1 Tris–HCl buffer,
pH 9.0, following Kunitz’s (1947) method, modified
by Walter (1984). Acid protease activity was evalu-
ated according to Anson (1938) using 0.5% hemo-
globin in 0.1 mmol l-1 glycine–HCl, pH 2.0. One
unit of enzyme activity was defined as 1 lg tyrosine
released per minute using a coefficient of molar
extinction of 0.008 at 280 nm. Trypsin activity was
assayed using BAPNA (N-a-benzoyl-DL-arginine
4-nitroanilide hydrochloride) as substrate according
to Erlanger et al. (1961). Chymotrypsin activity in
extracts was determined using SAAPNA (N-succinyl-
ala–ala-pro-phe p-nitroanilide) according to DelMar
et al. (1979). Leucine aminopeptidase was deter-
mined using leucine p-nitroanilide (0.1 mmol l-1 in
DMSO) as substrate, according to Maraux et al.
(1973). For trypsin, chymotrypsin, and leucine ami-
nopeptidase activities, one unit of enzyme activity
was defined as 1 lmol p-nitroaniline released per
minute using coefficients of molar extinction of 8.2 at
410 nm. Carboxypeptidase A activity was measured
following the protocol of Folk and Schirmer (1963)
using HPA (hippuryl-L-phenyl-alanine) as substrate
dissolved in 25 mmol l-1 Tris–HCl, 10 mmol l-1
CaCl2 buffer, pH 7.8. One unit of enzyme activity
was defined as 1 lmol of hydrolyzed hippuryl per
minute using a coefficient of molar extinction of 0.36
at 254 nm.
Determination of a-amylase activity was carried out
following the Somoyi-Nelson procedure described by
Robyt and Whelan (1968). One unit of activity was
defined as the amount of enzyme able to produce 1 lg
of maltose per minute at 600 nm. Lipase activity was
quantified using b-naphthyl caprylate as substrate
according to Versaw et al. (1989). One unit of activity
was defined as 1 lg of naphthol released per minute
using a molar extinction coefficient of 0.02 at 540 nm.
Acid and alkaline phosphatases were assayed using
4-nitrophenyl phosphate in acid citrate buffer (pH 5.5)
or glycine–NaOH buffer (pH 10.1) according to
Bergmeyer (1974). One unit was defined as 1 lg of
nitrophenyl released per minute using a molar extinc-
tion coefficient of 18.5 at 405 nm. All assays were
performed by triplicate at 37�C.
Digestive enzyme activities were expressed as
U mg protein-1 and U larva-1 using the total number
of larvae in each homogenized pooled sample.
Electrophoretic analysis
The analysis of the alkaline protease isoforms was
done using sodium dodecyl sulfate–polyacrylamide
gel electrophoresis (SDS–PAGE; 10% polyacryl-
amide) for each larval enzyme preparation in a Mini
Protean II chamber (Bio-Rad) according to Laemmli
(1970) using 8 9 10 9 0.075-cm gels. Zymograms of
alkaline protease activities were obtained as described
by Garcıa-Carreno et al. (1993). Electrophoresis was
carried out during 60 min at a constant voltage of
100 V per gel at 5�C. After electrophoresis, the gels
were washed and incubated for 30 min at 5�C in a 0.5%
casein Hammerstein (Research Organics) solution at
pH 9.0. The gels were then incubated for 90 min in the
same solution at 25�C without agitation. Finally, the
gels were washed and fixed in 12% trichloroacetic acid
(Sigma–Aldrich) prior to staining with 0.1% Coomas-
sie brilliant blue R-250 (Research Organics) in a
solution of methanol-acetic acid (Sigma–Aldrich)-
water (50:20:50). Distaining was carried out in a
solution of methanol-acetic acid–water (35:10:55).
Clear zones, which indicated activity of alkaline
proteases, were visible after 24 h.
The acid protease activities in larval extracts were
analyzed by neutral native polyacrylamide electro-
phoresis (Williams and Reisfeld 1964). All electro-
phoresis procedures were performed at a constant
voltage and amperage (100 V and 64 mA). Acid
protease isoforms were revealed according to the
procedure of Dıaz-Lopez et al. (1998). Same quantity
of protein (30 lg per well) was applied to carry out
each electrophoresis. The gels were removed from
the cell and soaked in 100 mmol l-1 HCl to reach pH
2.0 where the enzymes become active. After 15 min,
the gel was soaked for 30 min at 4�C in a solution
Fish Physiol Biochem (2012) 38:441–454 443
123
Author's personal copy
containing 0.25% hemoglobin in 100 mmol l-1 Gly-
cine–HCl, pH 2.0, and then for 90 min in a fresh
hemoglobin solution at 37�C. The gels were washed
in distilled water and fixed for 15 min in a 12%
trichloroacetic acid solution. When the clear areas of
enzyme activity appeared, the gels were stained using
Coomassie brilliant blue R-250 solution. Destaining
was carried out as mentioned earlier. Clear zones
revealed the activity of acid proteases within a few
minutes although well-defined zones were obtained
only after 2–4 h of staining. A low-range molecular
weight marker (5 ll per well) containing phosphor-
ylase b (97 kDa), bovine serum albumin (66 kDa),
egg albumin (45 kDa), carbonic anhydrase (29 kDa),
trypsinogen (24 kDa), and soybean trypsin inhibitor
(20 kDa) was applied to each SDS–PAGE. The
relative electromobility (Rf) was calculated for all
zymograms (Igbokwe and Downe 1978), and the
molecular weight (MW) of each band in the SDS-
zymograms (alkaline protease) was calculated by a
linearly adjusted model between the Rf and the
decimal logarithm of MW proteins using Quality One
V. 4.6.5 (Hercules, CA) software program.
Statistical analysis
Larval growth was determined with an exponential
model Y = aebX, with logarithm base 10 transformed
data, and the model parameters were calculated by
using the least-squares technique. A Kruskal–Wallis
test was used to compare enzyme activity between
ages for each activity. A nonparametric Nemenyi test
was used when significant differences were detected.
All tests were carried out with Statistica v7.0
(StatSoft, Tulsa, OK, USA) software.
Results
Centropomus undecimalis larvae showed a typical
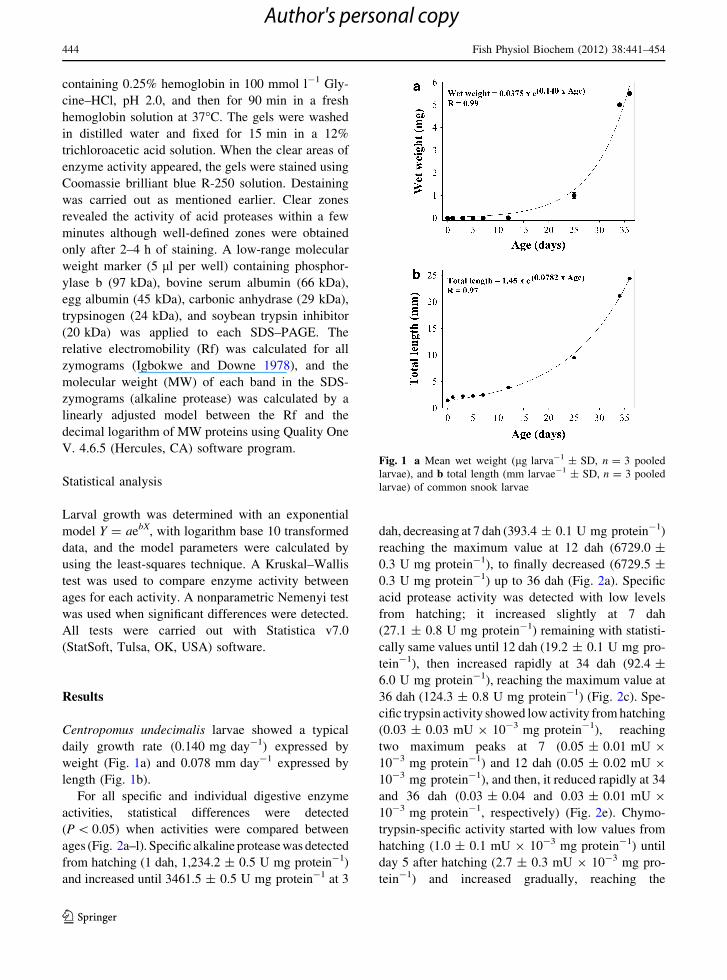
daily growth rate (0.140 mg day-1) expressed by
weight (Fig. 1a) and 0.078 mm day-1 expressed by
length (Fig. 1b).
For all specific and individual digestive enzyme
activities, statistical differences were detected
(P \ 0.05) when activities were compared between
ages (Fig. 2a–l). Specific alkaline protease was detected
from hatching (1 dah, 1,234.2 ± 0.5 U mg protein-1)
and increased until 3461.5 ± 0.5 U mg protein-1 at 3
dah, decreasing at 7 dah (393.4 ± 0.1 U mg protein-1)
reaching the maximum value at 12 dah (6729.0 ±
0.3 U mg protein-1), to finally decreased (6729.5 ±
0.3 U mg protein-1) up to 36 dah (Fig. 2a). Specific
acid protease activity was detected with low levels
from hatching; it increased slightly at 7 dah
(27.1 ± 0.8 U mg protein-1) remaining with statisti-
cally same values until 12 dah (19.2 ± 0.1 U mg pro-
tein-1), then increased rapidly at 34 dah (92.4 ±
6.0 U mg protein-1), reaching the maximum value at
36 dah (124.3 ± 0.8 U mg protein-1) (Fig. 2c). Spe-
cific trypsin activity showed low activity from hatching
(0.03 ± 0.03 mU 9 10-3 mg protein-1), reaching
two maximum peaks at 7 (0.05 ± 0.01 mU 9
10-3 mg protein-1) and 12 dah (0.05 ± 0.02 mU 9
10-3 mg protein-1), and then, it reduced rapidly at 34
and 36 dah (0.03 ± 0.04 and 0.03 ± 0.01 mU 9
10-3 mg protein-1, respectively) (Fig. 2e). Chymo-
trypsin-specific activity started with low values from
hatching (1.0 ± 0.1 mU 9 10-3 mg protein-1) until
day 5 after hatching (2.7 ± 0.3 mU 9 10-3 mg pro-
tein-1) and increased gradually, reaching the
Fig. 1 a Mean wet weight (lg larva-1 ± SD, n = 3 pooled
larvae), and b total length (mm larvae-1 ± SD, n = 3 pooled
larvae) of common snook larvae
444 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy
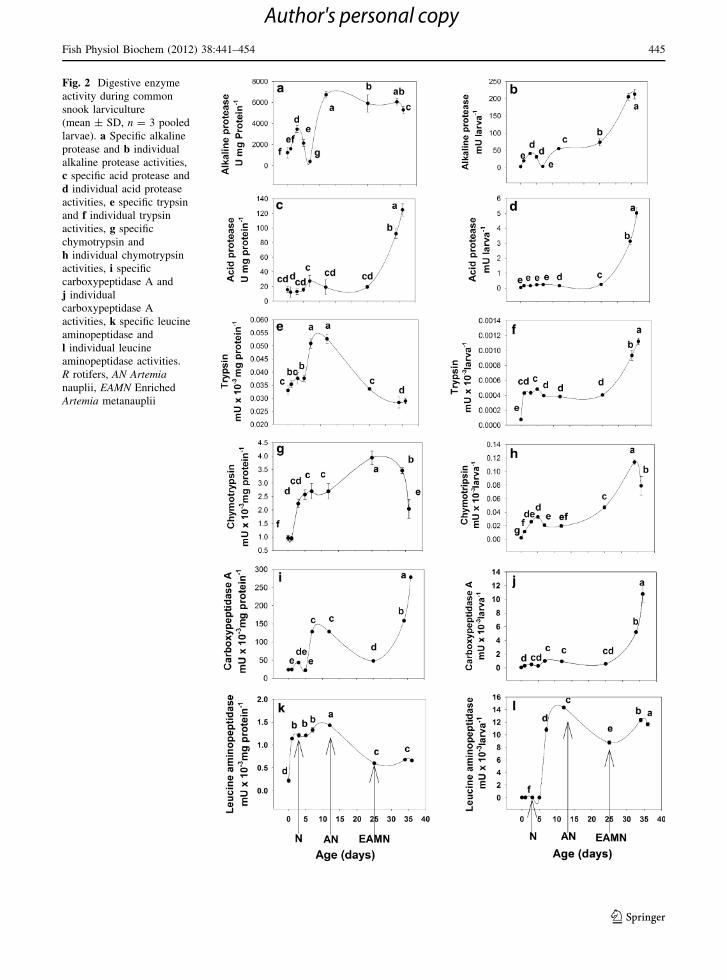
Fig. 2 Digestive enzyme
activity during common
snook larviculture
(mean ± SD, n = 3 pooled
larvae). a Specific alkaline
protease and b individual
alkaline protease activities,
c specific acid protease and
d individual acid protease
activities, e specific trypsin
and f individual trypsin
activities, g specific
chymotrypsin and
h individual chymotrypsin
activities, i specific
carboxypeptidase A and
j individual
carboxypeptidase A
activities, k specific leucine
aminopeptidase and
l individual leucine
aminopeptidase activities.
R rotifers, AN Artemianauplii, EAMN Enriched
Artemia metanauplii
Fish Physiol Biochem (2012) 38:441–454 445
123
Author's personal copy

maximum activity at 25 dah (3.9 ± 0.2 mU 9
10-3 mg protein-1) and then decreasing rapidly from
this day onwards (2.0 ± 1.0 mU 9 10-3 mg pro-
tein-1 at 36 dah) (Fig. 2g). Carboxypeptidase A-spe-
cific activity was low during the first days of
larviculture around 25.1 ± 0.0 mU mg protein-1,
and increased rapidly at days 7 (128.3 ± 0.3
mU mg protein-1) and 12 days after hatching
(128.1 ± 0.2 mU mg protein-1) to reduce at 25 dah
(47.8 ± 0.2 mU mg protein-1), and increased again at
34 and 36 dah (158.4 ± 0.1 and 278.2 ± 0.2 mU
mg protein-1, respectively) (Fig. 2i). Specific Leucine
aminopeptidase was first detected from hatching
(1 dah, 0.23 ± 0.02 mU mg protein-1); it increased
gradually until it reached the maximum peak of activity
at 12 dah (1.4 ± 0.2 mU mg protein-1) and then
decreased from this day up to the end of the larviculture
at 36 dah (0.60 ± 0.03 mU mg protein-1) (Fig. 2k).
For individual digestive proteases activities, a general
pattern was observed with a gradual increase from the
beginning of the larviculture to reach the maximum
peak at 36 dah. For alkaline protease (Fig. 2b), the
activity started from 20 to 50 mU larva-1, increasing
to reach their maximum values at 34 and 36 dah
(210 ± 3 and 225 ±
1 mU larva-1, respectively), for acid proteases
(Fig. 2d) the lowest values varied from 0.1 to
0.25 mU larva-1 from hatching until 12 dah, reaching
the highest values at 34 and 36 dah (3.5 ± 0.2 and
5.2 ± 0.8 mU larva-1, respectively), trypsin activity
(Fig. 2f) had the lowest values from hatching until 12
dah (0.001–0.004 mU 9 10-3 larva-1), and reaching
the highest values at 34 and 36 dah (0.1 ± 0.0
and 0.01 ± 0.00 mU 9 10-3 larva-1, respectively),
finally, carboxypeptidase A (Fig. 2j) showed the
lowest values from hatching until 25 dah (from 0.3
until 0.4 mU 9 10-3 larva-1), increasing the activity
rapidly until the maximum values at 34 and 36 dah
(5.9 ± 0.1 and 11.0 ± 0.04 mU 9 10-3 larva-1,
respectively). Chymotrypsin individual activity
showed a similar gradual increment as the specific
activity from hatching until 25 dah (0.01–0.05 mU 9
10-3 larvae-1), and reaching the maximum peak
at 34 dah (0.1 ± 0.0 mU 9 10-3 larva-1) and then
decreased rapidly at 36 dah (0.1 ± 0.0 mU 9
10-3 larva-1) (Fig. 2h). Finally, the leucine amino-
peptidase individual activity was different from the
other enzyme patterns; it showed null activity from
hatching up to day 5 after hatching, with a rapid
increase at 7 dah (11.0 ± 0.1 mU 9 10-3 larva-1),
reaching the maximum peak at 12 dah (14.5 ±
0.2 mU 9 10-3 larva-1); and then decreased at 25
dah (10.3 ± 0.2 mU 9 10-3 larva-1), and increasing
again at 34 (13.2 ± 0.1 mU 9 10-3 larva-1) and 36
dah (12.5 ± 0.1 mU 9 10-3 larva-1) (Fig. 2i).
Specific lipase activity showed slight increments at
3 (130.5 ± 0.3 U mg protein-1) and 7 dah (190.2 ±
0.3 U mg protein-1); afterward, the activity
remained low (90–110 U mg protein-1) until 36
dah (480.4 ± 0.3 U mg protein-1) when it increased
rapidly, being this day the peak with the maximum
activity (Fig. 3a). The individual lipase activity
gradually increased its activity from hatching (10 ±
1 mU 9 10-3 larva-1) until 12 dah (1.8 ± 0.1
mU larva-1), increasing its value (3.8 ± 0.9 mU
larva-1) at 12 dah, reaching the maximum value at
34 dah (10.3 ± 0.12 mU larva-1), and finally
decreasing rapidly at 36 dah (1.3 ± 0.1 mU larva-1)
(Fig. 3b). Specific a-amylase activity was present
from day 1 after hatching (0.2 ± 0.0 U mg pro-
tein-1); it increased from day 5 (0.9 ± 0.0
U mg protein-1) until day 12 after hatching (0.9 ±
0.0 U mg protein-1), reaching its maximum value at
7 dah (1.1 ± 0.0 U mg protein-1); the activity sud-
denly decreased from day 25 (0.1 ± 0.0 U mg pro-
tein- 1) until 34 dah (0.1 ± 0.0 U mg protein-1) and
to increase slightly 36 dah (0.5 ± 0.1 U mg pro-
tein-1) (Fig. 3c). For the individual activity of
a-amylase, two peaks were detected; the first one at 7
dah (8.0 ± 0.2 mU larva-1), with a gradual decrease
on the activity until day 34 after hatching (2.1 ±
0.4 mU larva-1) and a rapid increase at 36 dah
(18.2 ± 0.4 mU larva-1) being this the day of max-
imum activity (Fig. 3d). In the case of acid phospha-
tase-specific activity, low values were detected from
hatching until day 12 (0.5 ± 0.1 U mg protein-1);
however, from day 25 after hatching (3.8 ± 0.1
U mg protein-1) the activity increased, reaching the
maximum peak at 34 dah (5.1 ± 0.1 U mg pro-
tein-1) (Fig. 3e). The individual acid phosphatase
activity showed a similar tendency, being the max-
imum peaks obtained at 25 (50.5 ± 1.3 mU larva-1)
and 36 dah (Fig. 3f). Finally, the specific and
individual alkaline phosphatase activities (8–10
U mg protein-1 and 95–183 mU larva-1, respec-
tively) had the same pattern with low values from
446 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy
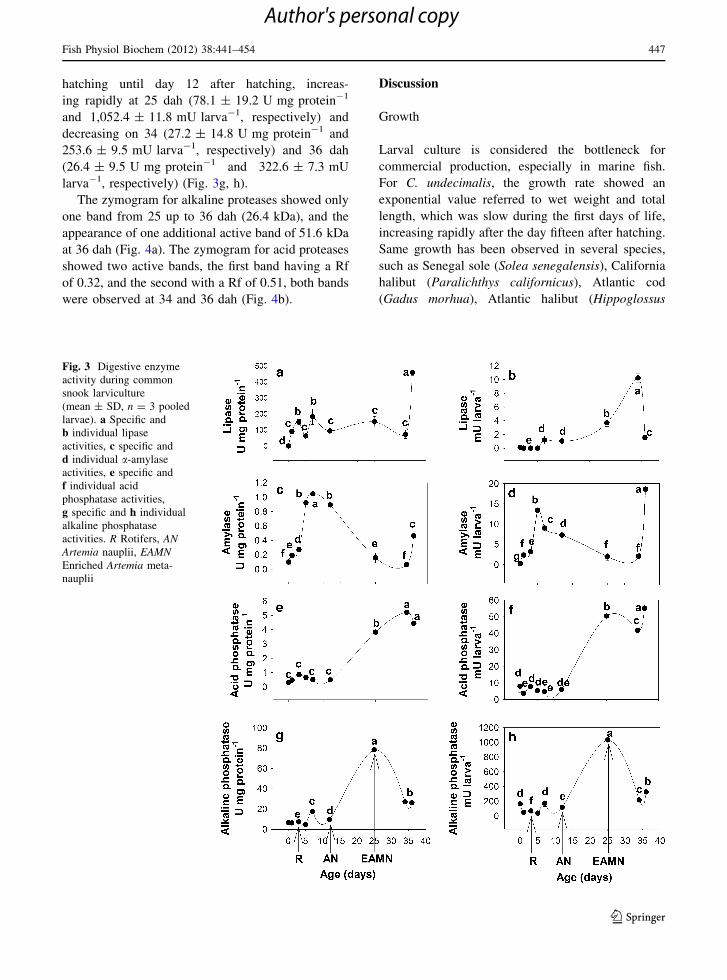
hatching until day 12 after hatching, increas-
ing rapidly at 25 dah (78.1 ± 19.2 U mg protein-1
and 1,052.4 ± 11.8 mU larva-1, respectively) and
decreasing on 34 (27.2 ± 14.8 U mg protein-1 and
253.6 ± 9.5 mU larva-1, respectively) and 36 dah
(26.4 ± 9.5 U mg protein-1 and 322.6 ± 7.3 mU
larva-1, respectively) (Fig. 3g, h).
The zymogram for alkaline proteases showed only
one band from 25 up to 36 dah (26.4 kDa), and the
appearance of one additional active band of 51.6 kDa
at 36 dah (Fig. 4a). The zymogram for acid proteases
showed two active bands, the first band having a Rf
of 0.32, and the second with a Rf of 0.51, both bands
were observed at 34 and 36 dah (Fig. 4b).
Discussion
Growth
Larval culture is considered the bottleneck for
commercial production, especially in marine fish.
For C. undecimalis, the growth rate showed an
exponential value referred to wet weight and total
length, which was slow during the first days of life,
increasing rapidly after the day fifteen after hatching.
Same growth has been observed in several species,
such as Senegal sole (Solea senegalensis), California
halibut (Paralichthys californicus), Atlantic cod
(Gadus morhua), Atlantic halibut (Hippoglossus
Fig. 3 Digestive enzyme
activity during common
snook larviculture
(mean ± SD, n = 3 pooled
larvae). a Specific and
b individual lipase
activities, c specific and
d individual a-amylase
activities, e specific and
f individual acid
phosphatase activities,
g specific and h individual
alkaline phosphatase
activities. R Rotifers, ANArtemia nauplii, EAMNEnriched Artemia meta-
nauplii
Fish Physiol Biochem (2012) 38:441–454 447
123
Author's personal copy

hippoglossus), and spotted sand bass (Paralabrax
maculatofasciatus) (Ribeiro et al. 1999; Alvarez-
Gonzalez et al. 2006, 2008; Kvale et al. 2007), which
has been related with the live food quality and
quantity (mainly rotifers and Artemia nauplii), feed-
ing frequency, weaning period, and morphophysio-
logical changes that are related with digestion
(enzymatic activity) and absorption of nutrients by
enterocytes (Moyano et al. 1996).
Alkaline proteases
The trend observed for alkaline protease-, trypsin-,
and chymotrypsin-specific activities during the first
days of larviculture is similar to that of other
carnivorous fishes like S. lalandi (Chen et al. 2006),
P. californicus (Alvarez-Gonzalez et al. 2006), red
drum, Sciaenops ocellatus (Lazo et al. 2007), and
P. maculatofasciatus (Alvarez-Gonzalez et al. 2008a),
which suggest that these enzymes play an important
role in larval digestion, previous to the stomach
functionality (Moyano et al. 1996). These changes in
the alkaline enzymatic activity agree with Cahu and
Zambonino-Infante (1994), who observed that pan-
creatic enzymes are responsible for most protein
hydrolysis in the gut, which is initiated in the luminal
zone, and then is continued in the parietal cell
epithelium (microvilli), before the absorption of
nutrients by enterocytes.
These variations in the alkaline protease activities
during C. undecimalis larviculture could be associ-
ated to morphophysiological changes and co-feeding
process when rotifers were replaced by Artemia
nauplii. These changes may increase the secretion of
pancreatic zymogens, possibly influenced by the type
and quantity of protein of the feed; also, the first
feeding after yolk absorption may induce the activa-
tion of alkaline proteases such as chymotrypsin that
occurred at 3 dah, which has been detected as the first
alkaline exoprotease that acts in the digestive tract
(Hjelmeland et al. 1983). In contrast, the level of
trypsin in our species was detected early at low level
and increased rapidly at 12 dah. This enzyme has also
been early detected in Senegal sole (Solea senegal-
ensis) and P. olivaceus larvae, even before the first
feeding. Indeed, this enzyme is detected before the
mouth opening, and it is related to the activation of
other pancreatic zymogens and the protein hydrolysis
in the lumen of S. senegalensis and P. olivaceus
(Kurokawa and Suzuki 1996; Saenz et al. 2005). The
differences between trypsin and chymotrypsin activ-
ities observed in C. undecimalis larvae could be
considered as an indicator of nutritional status; for
example, under normal conditions, the proportion of
trypsin increases as a response to protein hydrolysis
demand; however, if the larvae is reared with an
inadequate diet or suffers a feeding restriction, this
proportion decreases because relatively less trypsin is
produced against a constant secretion of chymotryp-
sin (Moyano et al. 1996; Cara et al. 2003).
For carboxipeptidase A in C. undecimalis larvae,
low activity was detected from the embryos until 5
dah, increasing from 7 to 12 dah, decreasing at 25
dah, and reaching its maximum values at 34 and 36
dah. This enzyme has been primarily studied on
adults of common carp, Cyprinus carpio (Cohen et al.
1981), amur catfish, Parasilurus asotus (Yoshinaka
et al. 1985), S. maximus (Munilla-Moran and Stark
1990), and small-spotted catshark, Scyliorhinus
Fig. 4 Zymograms of acid (a) and alkaline digestive proteases
(b) during larval development of common snook. Numbers at
the top of the gel indicate the mean of days after hatching. The
first well indicates LWMM (kDa): 97 Phosphorylase; 66Bovine serum albumin; 45 Ovoalbumin; 29 Carbonic anhy-
drase; 24 Trypsinogen, 20 Trypsin soybean inhibitor
448 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy

canicula (Hajjou et al. 1995), it is an important
digestive parietal enzyme that depends on Zn (me-
tallo-protease), and it is produced in the acinar
pancreatic cells to hydrolyze peptides from carboxyl
side (Vendrell et al. 2000). In P. olivaceus larvae has
only been reported procarboxypetidase A as the
major pancreatic enzyme, precursor for peptide
digestion in the intestinal lumen and is synthesized
at the first feeding (Srivastava et al. 2002).
Leucine aminopeptidase activity was detected with
low values in embryos, increasing between 2 and 12
dah, after that, decreasing was observed until the end
of the larviculture. This pattern coincides with turbot,
Scophthalmus maximus (Cousin et al. 1987) and
European sea bass, Dicentrarchus labrax (Cahu and
Zambonino-Infante 1997), where these parietal
enzymes were localized on microvilli of the entero-
cyte as proenzymes and then activated by alkaline
proteases (trypsin and chymotrypsin) to hydrolyze
peptides from the N-terminal amino acid.
Acid proteases (pepsin)
In C. undecimalis larvae, the maximum activity of
acidic protease was observed between 25 and 36 dah,
similarly to what is observed in other marine fish
larvae such as white bream (Diplodus sargus),
common dentex (Dentex dentex), and redbanded
seabream (Pagrus auriga) (Alarcon et al. 1998; Cara
et al. 2003; Moyano et al. 2005). It has been reported
that the appearance of pepsinogen like activity in the
bastard halibut Paralichthys olivaceus larvae at 45
dah (Kurokawa and Suzuki 1996) indicates the
starting of the juvenile stage and the settlement
period. Detection of high peaks of acid protease
activity (pepsin) indicates the presence of a func-
tional stomach (gastric cells and acid chloride
secretion) and could be taken, with other digestive
enzymes, as an indicator of maturation of the
digestive system (Ueberschaer 1993; Baglole et al.
1998; Kvale et al. 2007). In this sense, the change
from an undifferentiated straight tube in larvae to a
differentiated gastrointestinal tract, including the
presence on gastric cells, allows to start the weaning
period (Lazo et al. 2007). However, it is adequate to
complement the information of enzymatic activity
(biochemistry techniques) with the presence of gas-
tric cells through histology, as was done with the
D. dentex (Gisbert et al. 2009). On the other hand,
low activity pepsin activity was detected during the
first days of larviculture (from yolk-sac larvae until 12
dah) and is not necessarily from stomach origin, but it
could be another type of hydrolases (cathepsines),
which work at intracellular level under acidic condi-
tions and could be detected when whole-body
extracts are prepared (Moyano et al. 1996).
Lipase
Lipase activity was detected at 3 dah, increasing
rapidly at 36 dah, in accordance to Green and
McCormick (2001) who suggested that the presence
of these digestive enzymes before hatching is
strongly related with the absorption of nutritional
components of the yolk sac. Specifically for lipase
activity, diverse studies have been conducted through
larval development of marine fish, where the activity
shows two high peeks in a recurrent manner: the first
one occurs at early days of life related to lipid
hydrolysis of the yolk, and the second one when the
digestive system maturation was reached (Oozeki and
Baley 1995). However, these peeks could present
fluctuations, which are strictly related to either
changes in feed supplies and feed enrichment with
lipid emulsifiers (Hoehne-Reitan et al. 2001), as it
was observed in S. senegalensis larvae (Martınez
et al. 1999), in which two peeks of maximum activity
were detected at 7 and 36 dah. It is admitted that lipid
catabolism is performed primarily by esterase action
hydrolyzing fatty acids as energy source in the first
days of life, while true lipase is dependant on colipase
and bile salts, acting over phospholipids and triacyl-
glycerols (van Tilbeurgh et al. 1992); this enzyme is
responsible for releasing highly polyunsaturated fatty
acids and other more complex compounds, generally
observed when maturation of the digestive system
arrives (Ribeiro et al. 1999; Zambonino-Infante and
Cahu 1999; Gawlicka et al. 2000; Sidell and Hazel
2002; Murray et al. 2003; Morais et al. 2005; Gisbert
et al. 2009)
Amylase
Regarding a-amylase activity during ontogeny of
C. undecimalis, maximum values were detected between
5 and 12 dah, and later decreased, in agreement with
to earlier observations for other marine fish species,
where the highest activity was reported before
Fish Physiol Biochem (2012) 38:441–454 449
123
Author's personal copy

hatching and during the first days of life when
the absorption of yolk-sac occurred and was followed
by a subsequent reduction in activity (Cahu and
Zambonino-Infante 1994; Zambonino-Infante and
Cahu 2001). a-amylase activity and expression have
been detected in fish fertilized eggs by biochemical or
molecular approaches (Naz 2008; Darias et al. 2006)
though its function has not been understood at early
stages of fish development (Moyano et al. 1996;
Peres et al. 1996; Martınez et al. 1999; Buchet et al.
2000; Cuvier-Peres and Kestemont 2002). In larvae,
it has been considered as a digestive system matu-
ration indicator as it occurs in mammals with lactase
(Zambonino-Infante and Cahu 1994). Fish larvae
tend to maintain a-amylase at low levels of activity to
utilize carbohydrates in the feed in carnivorous
species (Munilla-Moran and Saborido-Rey 1996;
Alvarez-Gonzalez et al. 2008). Fange and Grove
(1979), Ugolev et al. (1983), and Hidalgo et al.
(1999) have proved that a-amylase activity tends to
be higher with a progressive activity increasing in
herbivorous and omnivorous fish when compared to
carnivorous fish.
Phosphatases
On the other hand, parietal digestive enzymes, such
as the brush border enzyme phosphatases, are very
important because they are responsible for conclud-
ing digestion at intestinal epithelium level besides
helping on hydrolyzed nutrient transport into the
enterocytes (Harpaz and Uni 1999; Smith et al. 2000;
Zambonino-Infante and Cahu 2001). Alvarez-Gon-
zalez et al. (2008) reported that phosphatases have
two main functions: the first one is to hydrolyze
inorganic phosphate used for energy production, and
the second one is to transport nutrients through cell
membranes (absorption process). In this sense,
Gawlicka et al. (2000) reported that when enterocytes
reach their maximum hydrolysis and absorption
capacity, phosphatase activity increases, which is
related to a drop in leucine-alanine peptidase activity
and maturation of enterocytes, as it has been detected
in D. labrax larvae (Peres et al. 1997) and conse-
quently determines the most adequate moment to
carry on the substitution of live prey to artificial feeds
(Ribeiro et al. 2002; Zambonino-Infante and Cahu
2007). In this manner, phosphatases act in the
digestion process facilitating other enzyme actions,
besides initiating the migration of nutrient processes
from the cryptic region to the microvilli border to
promote cell absorption (Copeland 1996).
Zymogram analysis
Zymogram of acid protease activity in C. undecimalis
larvae allowed the detection of two isoforms (0.32
and 0.51 Rf’s) up to 34 dah, which is similar in
juveniles as reported by Concha-Frıas (2008); this
author found that the total acid protease activity was
86% inhibited using pepstatin A in the stomach of
C. undecimalis juveniles. In this sense, pepsin activity
detection through this technique has been reported for
other species such as silk snapper, Lutjanus vivanus
(26.1 kDa active fraction); grunt, Haemulon plumi-
erii (24 kDa active fraction); spotted goatfish,
Pseudupeneus maculatus (24.0 kDa active fraction);
P. maculatofasciatus (one active fraction on 12 dah,
0.65 RF) (Rivera 2003; Rodrıguez 2004; Souza et al.
2007; Alvarez-Gonzalez et al. 2010) in agreement
that these detected enzymes are similar to the swine
pepsin A (35 kDa). However, the detection of a
second acid protease isoform in C. undecimalis larvae
could correspond not only to pepsin A, but also to
pepsin C that was reported for Pacific bluefin tuna
Thunnus orientalis juveniles using a molecular
approach (Tanji et al. 2009). For alkaline protease,
zymogram of C. undecimalis larvae showed only two
isoforms, which apparently correspond to trypsin-like
(51.6 kDa) detected at 36 dah, and chymotrypsin-like
(26.4 kDa) detected at 34 and 36 dah. In this sense,
an early detection using electrophoretical technique
of these isoforms was not possible because of the low
activity in whole-body homogenates until 25 dah,
which was corroborated using biochemical technique.
The molecular masses of active fractions found for
C. undecimalis larvae were similar to those detected
in wild juveniles of the same species (11 cm of total
length) by Concha-Frıas (2008); however, this
researcher found a third high molecular mass isoform
([75 kDa), which could correspond to an aminopep-
tidase. Our results are similar to those reported for
other strictly carnivorous marine fish species such as
the coho salmon (Oncorhynchus kisutch) and chinook
salmon (Oncorhynchus tschawytscha) with two
22 kDa isoforms, Northern bluefin tuna (Thunnus
thynnus) with three isoforms from 16.8 to 26.8 kDa
and spotted sand bass (P. maculatofasciatus) with
450 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy

two isoforms of 20.1–56.5 kDa, respectively. They
are different from omnivorous fishes such as S.
aurata (five active fractions ranging 24.5–90 kDa),
D. dentex (eight fractions ranging from 24.5 to
69.5 kDa), pirapita, Brycon orbignyanus (nine frac-
tions ranging from 7 to 70 kDa), blue disk, Symphys-
odon aequifasciata (eight fractions ranging from 19.2
to76.5 kDa), and rohu, Labeo rohita (five fractions
ranging from 20.9 to 69.4 kDa) (Dimes et al. 1994;
Alarcon et al. 1998; Garcıa-Carreno et al. 2002;
Chong et al. 2002; Essed et al. 2002; Chakrabarti
et al. 2006; Alvarez-Gonzalez et al. 2010).
We can conclude that the low activities of alkaline
protease, lipase, a-amylase, and phosphatase activity
detected in C. undecimalis larvae at hatching and
their increment throughout larviculture also the
presence of only two isoforms of alkaline protease
may be indicators of (1) genetic processes, specially
while observing a slight specific activity in starving
larvae during yolk absorption (for example, phos-
pholipids hydrolysis); (2) a response to ingestion
(feeding behavior), at the moment of adding live
feeds (rotifers, Artemia and enriched Artemia meta-
nauplii); and (3) progressive disappearance of the
metabolic function in which a specific enzyme is
involved (lactase in the case of mammals), or a
relative increase in the soluble protein pool in the
organism. Additionally, acidic protease activity,
which starts at 25 dah reaching its maximum activity
at 36 dah, allows us to consider C. undecimalis as a
juvenile focusing on a digestive physiology point of
view from 34 dah onwards, and let us situate this
species as a strictly carnivorous fish larvae, also the
weaning period for this species should be started after
this age.
Acknowledgments This work was made possible thanks to
the Project ‘‘Estudio sobre la fisiologıa digestiva del robalo
blanco Centropomus undecimalis’’ SEP-CONACyT (CB-2006-
1-58931). We thank Claudia Durruty Lagunes and Jaime
Suarez Bautista for their technical assistance. The Consejo
Nacional de Ciencia y Tecnologıa (CONACYT) of Mexico
provided a fellowship grant to the first author Luis Daniel
Jimenez-Martınez.
References
Alarcon FJ, Dıaz M, Moyano FJ, Abellan E (1998) Charac-
terization and functional properties of digestive proteases
in two sparids; gilthead seabream (Sparus aurata) and
common dentex (Dentex dentex). Fish Physiol Biochem
19:257–267
Alvarez-Gonzalez CA, Cervantes-Trujano M, Tovar-Ramırez
D, Conklin DE, Nolasco H, Gisbert E, Piedrahita R (2006)
Development of digestive enzymes in California halibut
Paralichthys californicus larvae. Fish Physiol Biochem
31:83–93
Alvarez-Gonzalez CA, Moyano-Lopez FJ, Civera-Cercedo R,
Carrasco-Chavez V, Ortiz-Galindo J, Dumas S (2008)
Development of digestive enzyme activity in larvae of
spotted sand bass (Palabrax maculatofasciatus). I: bio-
chemical analysis. Fish Physiol Biochem 34:373–384.
doi:10.1007/s10695-007-9197-7
Alvarez-Gonzalez CA, Moyano-Lopez FJ, Civera-Cercedo R,
Carrasco-Chavez V, Ortiz-Galindo J, Nolasco-Soria H,
Tovar-Ramırez D, Dumas S (2010) Development of diges-
tive enzyme activity in larvae of spotted sand bass (Palabraxmaculatofasciatus) II: electrophoretic analysis. Fish Physiol
Biochem 36:29–37. doi:10.1007/s10695-008-92976-4
Alvarez-Lajonchere L, Cequeira RV, Dos Reis M (2002) De-
sarrollo embrionario y primeros estadios larvales del robalo
chucumite, Centropomus parallelus Poey (Pices: Centrop-
omidae) con interes para su cultivo. Hidrobiologica 12(2):
89–100
Anson ML (1938) The estimation of pepsin, trypsin, papain
and cathepsin with hemoglobin. J Gen Physiol 22:79–89
Aviles-Quevedo A, McGregor-Pardo U, Rodrıguez-Ramos R,
Morales-Castro O, Huerta-Bello M, AIizawa M (1995)
Biologıa y cultivo de la cabrilla arenera Paralabrax ma-culatofasciatus (Steindachner, 1868). Secretarıa de Pesca.
Instituto Nacional de la Pesca. JICA. Mexico, p 85
Baglole CJ, Goff GP, Wright GM (1998) Distribution and
ontogeny of digestive enzymes in larval yellowtail and
winter flounder. J Fish Biol 53:767–784
Bergmeyer HV (1974) Phosphatases methods of enzymatic
analysis, vol 2. Academic Press, New York
Bradford MM (1976) A rapid and sensitive method for the
quantization of microgram quantities of protein utilizing the
principle of protein dye binding. Anal Biochem 72:248–254
Brock D, Robinette HR, Heinen J (1992) Culture system for
evaluating live and formulated diets for larval fish. Progr
Fish Cult 54:270–273
Buchet V, Zambonino-Infante JL, Cahu C (2000) Effect of lipid
level in a compound diet on the development of red drum
(Sciaenops ocellatus) larvae. Aquaculture 184:339–347
Cahu CL, Zambonino-Infante JL (1994) Early weaning of sea
bass (Dicentrarchus labrax) larvae with a compound diet:
effect on digestive enzymes. Comp Biochem Physiol 109A:
213–222
Cahu CL, Zambonino-Infante JL (1997) Is the digestive
capacity of marine fish larvae sufficient for compound diet
feeding? Aquacult Int 5:151–160
Cara JB, Moyano FJ, Cardenas S, Fernandez-Dıaz C, Yufera M
(2003) Assessment of digestive enzyme activities during
larval development of white bream. J Fish Biol 63:48–58
Cequeira RV, Brugger AM (2001) Effect of light intensity on
initial survival of fat snook (Centropomus parallelus, Pis-
ces: Centropomidae) larvae. Brazilian Arch Biol Technol
Int J 44(4):343–349
Chakrabarti R, Rathore RM, Kumar S (2006) Study of diges-
tive enzyme activities and partial characterization of
Fish Physiol Biochem (2012) 38:441–454 451
123
Author's personal copy

digestive proteases in a freshwater teleost, Labeo rohita,
during early ontogeny. Aquacult Nutr 12:35–43
Chavez H (1961) Estudio de una nueva especie de robalo del
Golfo de Mexico y redescripcion de Centropomus unde-cimalis (Bloch). Ciencia 2(2):75–86
Chen BN, Jian GQ, Martin SK, Wayne GH, Steven MC (2006)
Ontogenetic development of digestive enzymes in yel-
lowtail kingfish Seriola lalandi larvae. Aquaculture
256:489–501
Chong AS, Hashim R, Chow-Yang L, Ali AB (2002) Partial
characterization and activities of proteases from the
digestive tract of discus fish (Symphysodon aequifasciata).
Aquaculture 203:321–333
Cohen T, Gertler A, Birk Y (1981) Pancreatic proteolytic
enzymes from carp (Cyprinus carpio): 1. Purification and
physical properties of trypsin, chymotrypsin, elastase and
carboxypeptidase B. Comp Biochem Physiol Comp Bio-
chem 69B(3):639–646
Concha-Frıas B (2008) Evaluacion de la capacidad digestiva de
juveniles de Centropomus undecimalis (bloch, 1792) so-
bre diferentes ingredientes proteınicos. Master’s thesis,
Universidad Catolica del Norte de Chile, p 109
Copeland RA (ed) (1996) Structural components of enzymes.
In: Enzymes, a practical introduction to structure, mech-
anism and data analysis. Wiley, New York, pp 35–65
Cousin JCB, Baudin-Laurencin F, Gabaudan J (1987) Ontog-
eny of enzymatic activities in fed y fasting turbot,
Scophthalmus maximus L. J Fish Biol 30:15–33
Cuvier-Peres A, Kestemont P (2002) Development of some
digestive enzymes in Eurasian perch larvae Perca fluvia-tilis. Fish Physiol Biochem 24:279–285
Darias MJ, Murray HM, Gallant JW, Astola A, Douglas SE,
Yufera M, Martınez-Rodrıguez G (2006) Characterization
of a partial a-amylase clone from red porgy (Pagruspagrus): expression during larval development. Comp
Biochem Physiol 143:209–218
DelMar EG, Largman C, Broderick JW, Geokas MC (1979) A
sensitive new substrate for chymotrypsin. Anal Biochem
99:316–320
Dıaz-Lopez M, Moyano-lopez FJ, Alarcon-Lopez FJ, Garcıa-
Carreno FL, Navarrete del Toro MA (1998) Character-
ization of fish acid proteases by substrate-gel electro-
phoresis. Comp Biochem Physiol 121B:369–377
Dimes LE, Garcıa-Carreno FL, Haard NF (1994) Estimation of
protein digestibility. III. Studies on digestive enzyme from
the pyloric caeca of rainbow trout and salmon. Comp
Biochem Physiol 109:349–360
Erlanger B, Kokowsky N, Cohen W (1961) The preparation
and properties of two new chromogenic substrates of
trypsin. Arch Biochem Biophys 95:271–278
Essed Z, Fernandez I, Alarcon FJ, Moyano FJ (2002) Cara-
cterizacion de la actividad proteasa digestiva de atun rojo
Thunnus thynnus (Linnaeus, 1758). Bol Inst Esp Oceanogr
18(1–4):99–107
Fabillo MD, Herrera AA, Abucay JS (2004) Effects of delayed
first feeding on the development of the digestive tract y
skeletal muscles of Nile Tilapia, Oreochromis niloticus L.
In: Proceedings 6th international symposium on Tilapia in
aquaculture Philippine International Convention Center,
Roxas Boulevard, Manila, Philippines, pp 301–315
Fange R, Grove D (1979) Digestion. In: Hoar WS, Randall DJ,
Brett JR (eds) Fish physiology, vol 8. Academic Press,
NY, pp 161–260
Folk JE, Schirmer EW (1963) The porcine pancreatic car-
boxypeptidase a system. J Biol Chem 238:3884–3894
Garcıa-Carreno FL, Dimes LE, Haard NF (1993) Substrate-gel
electrophoresis for composition and molecular weight of
proteinases or proteinaceous proteinase inhibitors. Anal
Biochem 214:65–69
Garcıa-Carreno FL, Albuquerque-Cavalcanti C, Navarrete del
Toro MA, Zaniboni-Filho E (2002) Digestive proteinases
of Brycon orbignyanus (Characidae, Teleostei): charac-
teristics and effects of protein quality. Comp Biochem
Physiol 132B:343–352
Garcıa-Ortega A, Verreth JAJ, Coutteau P, Segner H, Huisman
EA, Sorgeloos P (1998) Biochemical and enzymatic
characterization of decapsulated cysts and nauplii of the
brine shrimp Artemia at different developmental stages.
Aquaculture 161:501–514
Gawlicka A, Parent B, Horn MH, Ross N, Opstad I, Torrinsen
OJ (2000) Activity of digestive enzymes in yolk-sac lar-
vae of Atlantic halibut (Hippoglossus hippoglossus):
indication of readiness for first feeding. Aquaculture
184:303–314
Gisbert E, Sarasquete MC, Willot P, Castello-Orvay F (1999)
Histochemistry of the development of the digestive sys-
tem of Siberian sturgeon during early ontogeny. J Fish
Biol 55:596–616
Gisbert E, Gimenez G, Fernandez I, Kotzamanis Y, Estevez A
(2009) Development of digestive enzymes in common
dentex Dentex dentex during early ontogeny. Aquaculture
287(3):381–387
Gracia-Lopez V, Kiewek-Martınez M, Maldonado-Garcıa M
(2004) Effects of temperature and salinity on artificially
reproduced eggs and larvae of the leopard grouper My-cteroperca rosacea. Aquaculture 237:485–498
Gracia-Lopez V, Rosas-Vazquez C, Brito-Perez R (2006)
Effects of salinity on physiological conditions in juvenile
common snook Centropomus undecimalis. Comp Bio-
chem Physiol 145A(3):340–345
Green BS, McCormick MI (2001) Ontogeny of the digestive
and feeding systems in the anemone fish Amphiprionmelanopus. Environ Biol Fishes 61:73–83
Grier H (2000) Ovarian germinal epithelium and folliculo-
genesis in the common snook, Centropomus undecimalis(Teleostei: centropomidae). J Morphol 243(3):265–281
Grier HJ, Taylor RG (1998) Testicular maturation and regression
in the common snook. J Fish Biol 53(3):521–542
Hajjou M, Smine A, Guerard F, Le Gal Y (1995) Purification
and some properties of a carboxypeptidase B from dogfish
Scyliorhinus canicula. Comp Biochem Physiol 110B(4):
791–798
Harpaz S, Uni Z (1999) Activity of intestinal mucosal brush
border membrane enzymes in relation to the feeding
habits of three aquaculture fish species. Comp Biochem
Physiol 124A:155–160
Hidalgo MC, Urea E, Sanz A (1999) Comparative study of
digestive enzymes in fish with different nutricional habits.
Proteolytic and amylase activities. Aquaculture 170:
267–283
452 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy

Hjelmeland K, Huse I, Jorgensen T, Molvik G, Raa J (1983)
Trypsin and trypsinogen as indices of growth and survival
potential of cod (Gadus morhua L.) larvae. Flodevigen
Rapp 3:1–17
Hoehne-Reitan K, Kjorsvik E, Reitan KI (2001) Bile saltde-
pendent lipase in larval turbot, as influenced by density
and lipid content of fed prey. J Fish Biol 58:746–754
Holt JG, Faulk CK, Schwarz MH (2007) A review of the lar-
viculture of cobia Rachycentron canadum, a warm water
marine fish. Aquaculture 268:181–187
Ibarra-Castro L, Duncan NJ (2007) GnRHa-induced spawning
of wild-caught spotted rose snapper Lutjanus guttatus.
Aquaculture 272:737–746
Igbokwe EC, Downe AER (1978) Electrophoretic and histo-
chemical comparison of three strains of Aedes aegypti.Comp Biochem Physiol 60B:131–136
Komar C, Turnbull JF, Roque A, Fajer E, Duncan NJ (2004)
Effect of water treatment and aeration on the percentage
hatch of demersal, adhesive eggs of the bullseye puffer
(Sphoeroides annulatus). Aquaculture 229:147–158
Kunitz M (1947) Crystalline soybean trypsin inhibitor II.
General properties. J Gen Physiol 30:291–310
Kurokawa T, Suzuki T (1996) Formation of the diffuse pan-
creas and the development of digestive enzyme synthesis
in larvae of the Japanese flounder Paralichthys olivaceus.
Aquaculture 141:267–276
Kvale A, Mangor-Jensen A, Moren M, Espe M, Hamre K
(2007) Development and characterization of some intes-
tinal enzymes in Atlantic cod (Gadus morhua L.) and
Atlantic halibut (Hippoglossus hippoglossus L.) larvae.
Aquaculture 264:457–468
Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature
227:680–685
Lazo JP, Mendoza R, Holt GJ, Aguilera C, Arnold CR (2007)
Characterization of digestive enzymes during larval
development of red drum (Sciaenops ocellatus). Aqua-
culture 265:194–205
Maraux S, Louvard D, Baratti J (1973) The aminopeptidase
from hog-intestinal brush border. Biochim Biophys Acta
321:282–295
Martınez MI, Moyano FJ, Fernandez-Dıaz C, Yufera M (1999)
Digestive enzyme activity during larval development of
the Senegal sole (Solea senegalensis). Fish Physiol Bio-
chem 21:317–323
Morais S, Rojas-Garcıa CR, Conceicao LEC, Rønnestad I
(2005) Digestion and absorption of a pure triacylglycerol
and a free fatty acid by Clupea harengus L. larvae. J Fish
Biol 67:223–238
Moyano FJ, Diaz M, Alarcon FJ, Sarasquete MC (1996)
Characterization of digestive enzyme activity during lar-
val development of gilthead sea bream (Sparus aurata).
Fish Physiol Biochem 15:121–130
Moyano FJ, Barros AM, Prieto A, Canavate JF, Cardenas S
(2005) Evaluacion de la ontogenia de enzimas digestivas
en larvas de hurta, Pagrus auriga (Pisces: Sparidae).
AquaNIC 22:39–47
Munilla-Moran R, Saborido-Rey F (1996) Digestive enzymes
in marine species. I. Proteinase activities in gut from red
fish (Sebastes mentella), seabream (Sparus aurata) and
turbot (Scophthalmus maximus). Comp Biochem Physiol
113B:395–402
Munilla-Moran R, Stark JR (1990) Metabolism in marine
flatfish—VI. Effect of nutritional state on digestion in
turbor, Scophthalmus maximus (L.). Comp Biochem
Physiol 95B(3):625–653
Murray HM, Gallant JW, Perez-Casanova JC, Johnson SC,
Douglas SE (2003) Ontogeny of lipase expression in
winter flounder. J Fish Biol 62:816–833
Naz M (2008) The changes in the biochemical compositions
and enzymatic activities of rotifer (Brachionus plicatilis,
Muller) and Artemia during the enrichment and starvation
periods. Fish Physiol Biochem 34:391–404
Oozeki Y, Baley M (1995) Ontogenetic development of
digestive enzyme activities in larval walleye pollock,
Theragra chalcogramma. Mar Biol 122:177–186
Peres A, Cahu CL, Zambonino-Infante JL, Legall MM,
Quazuguel P (1996) Amylase and trypsin responses to
intake of dietary carbohydrate and protein depend on the
developmental stage in sea bass (Dicentrarchus labrax)
larvae. Fish Physiol Biochem 15:237–242
Peres A, Cahu CL, Zambonino-Infante JL (1997) Dietary
spermine supplementation induces intestinal maturation in
sea bass (Dicentrarchus labrax) larvae. Fish Physiol
Biochem 16:479–485
Ramirez AB, Cerqueira VR (1994) Feeding behavior of young
robalo (Centropomus undecimalis Bloch, 1792). I. The
effect of chemical attractants. Aquaculture 124(1):289–290
Ribeiro L, Zambonino-Infante JL, Cahu C, Dinis MT (1999)
Development of digestive enzymes in larvae of Soleasenegalensis, Kaup 1858. Aquaculture 170:465–473
Ribeiro L, Zambonino-Infante JL, Cahu C, Dinis MT (2002)
Digestive enzymes profile of Solea senegalensis post
larvae fed Artemia and a compound diet. Fish Physiol
Biochem 27:61–69
Rivera SM (2003) Purification and characterization of trypsin
from intestinal and pyloric caecal tissues of the silk
snapper, Lutjanus vivanus (Cuvier 1828). Master’s thesis,
p 40
Robyt JF, Whelan WJ (1968) The a-amylase. In: Radley JA
(ed) Starch and its derivates. Chapman and Hall, London,
pp 430–497
Rodrıguez MAR (2004) Purification and kinetic characteriza-
tion of trypsin from the intestine and pyloric caeca of the
white grunt, Haemulon plumierii, (Lacepede, 1801).
Master’s thesis, p 29
Saenz RM, Alarcon FJ, Martınez MI, Ruiz F, Dıaz M, Moyano
FJ (2005) Caracterizacion de las proteasas digestivas del
lenguado senegales Solea senegalensis Kaup, 1858. Bol
Inst Esp Oceanogr 21(1–4):95–104
Shaozhen F, Wensheng L, Haoran L (2008) Characterization
and expression of the pepsinogen C gene and determina-
tion of pepsin-like enzyme activity from orange-spotted
grouper (Epinephelus coioides). Comp Biochem Physiol
149B:275–284
Sherwood NM, Grier HJ, Warby C, Peute J, Taylor RG (1993)
Gonadotropin-releasing hormones, including a novel
form, in snook Centropomus undecimalis, in comparison
with forms in black sea bass Centropristis striata. Regul
Pept 46(3):523–534
Fish Physiol Biochem (2012) 38:441–454 453
123
Author's personal copy

Sidell BD, Hazel JR (2002) Triacylglycerol lipase activities in
tissues of Antarctic fishes. Polar Biol 25:517–522
Smith TK, Tapia-Salazar M, Cruz-Suarez LE, Ricque-Marie D
(2000) Feed-borne biogenic amines: natural toxicants or
growth promoters? In: Cruz-Suarez LE, Ricque-Marie D,
Tapia-Salazar M, Olvera-Novoa MA, Civera-Cerecedo R
(eds) Avances en Nutricion Acuıcola V. Memorias del V
Simposium Internacional de Nutricion Acuıcola. Merida,
Yucatan, Mexico, pp 24–32, 19–22 Nov
Souza AAG, Amaral IPG, Alberico RES, Carvalho LB,
Bezerra RS (2007) Trypsin-like enzyme from intestine
and pyloric caeca of spotted goatfish (Pseudupeneusmaculatus). Food Chem 100:1429–1434
Srivastava AS, Kurokawa T, Suzuki T (2002) mRNA expres-
sion of pancreatic enzyme precursors and estimation of
protein digestibility in first feeding larvae of the Japanese
flounder, Paralichthys olivaceus. Comp Biochem Physiol
l32A:629–635
Stephen R, Shafland P (1982) Larval development of snook,
Centropomus undecimalis (Pisces: Centropomidae).
Copeia 3:618–627
Tanji M, Yakabe E, Kubota K, Kageyama T, Ichinose M, Miki
K, Ito H, Takahashi K (2009) Structural and phylogenetic
comparison of three pepsinogens from Pacific bluefin
tuna: molecular evolution of fish pepsinogens. Comp
Biochem Physiol 152B:9–19
Tarcisio T, Vinicius R, Brown J (2005) Early weaning of fat
snook (Centropomus parallelus Poey 1864) larvae.
Aquaculture 253:334–342
Ueberschaer B (1993) Measurement of proteolytic enzyme
activity: significance and application in larval fish
research. In: Walther BT, Fyhn HJ (eds) Physiological and
biochemical aspects of fish development, part III. Univ. of
Bergen, Norway, pp 233–239
Ugolev AM, Yegorova VV, Kuz’mina VV, Gruzdkov AA
(1983) Comparative molecular characterization of mem-
brane digestion on fish and mammals. Comp Biochem
Physiol 76B:627–635
van Tilbeurgh H, Sarda L, Verger R, Cambillau C (1992)
Structure of the pancreatic lipase-procolipase complex.
Nature 359:159–162
Vendrell J, Querol E, Aviles FX (2000) Metallocarboxypep-
tidases and their protein inhibitors. Structure, function and
biomedical properties. Biochim Biophys Acta 1477:
284–298
Versaw W, Cuppett SL, Winters DD, Williams LE (1989) An
improved colorimetric assay for bacterial lipase in nonfat
dry milk. J Food Sci 54:232–254
Versichelle D, Leger P, Lavens P, Sorgeloos P (1989) L’util-
isation d’artemia. In: Barnabe G (ed) Aquaculture. Tech-
nique et Documentation, Lavoisier, Paris, pp 241–259
Wainwright PC, Huskey SH, Turingan RG, Carroll AM (2006)
Ontogeny of suction feeding capacity in snook, Centrop-omus undecimalis. J Exp Zool 305(3):246–252
Walter HE (1984) Proteinases: methods with hemoglobin,
casein and azocoll as substrates. In: Bergmeyern HJ (ed)
Methods of enzymatic analysis, vol V. Verlag Chemie,
Weinham, pp 270–277
Williams DE, Reisfeld RA (1964) Disc electrophoresis in
polyacrylamide gels: extension to new conditions of pH
and buffers. Ann N Y Acad Sci 121:373–381
Yanes-Roca C, Rhody N, Nystrom M, Main KL (2009) Effects
of fatty acid composition and spawning season patterns on
egg quality and larval survival in common snook. Aqua-
culture 287(3):335–340
Yoshinaka R, Sato M, Tanaka H, Ikeda S (1985) Some enzy-
matic properties and digestive function of a pancreatic
metalloproteinase in the catfish (Parasilurus asotus).
Comp Biochem Physiol 80B(2):223–226
Zambonino-Infante JL, Cahu C (1994) Development and
response to a diet change of some digestive enzymes in
sea bass (Dicentrarchus labrax) larvae. Fish Physiol
Biochem 12(5):399–408
Zambonino-Infante JL, Cahu CL (1999) High dietary lipid levels
enhance digestive tract maturation and improve Dicen-trarchus labrax larval development. J Nutr 129:1195–1200
Zambonino-Infante JL, Cahu CL (2001) Ontogeny of the
gastrointestinal tract of marine fish larvae. Comp Biochem
Physiol 130C:477–487
Zambonino-Infante JL, Cahu CL (2007) Dietary modulation of
some digestive enzymes and metabolic processes in
developing marine fish: applications to diet formulation.
Aquaculture 268:98–105
454 Fish Physiol Biochem (2012) 38:441–454
123
Author's personal copy





























